
Applied evolutionary ecology of insects of the subfamily Bruchinae (Coleoptera: Chrysomelidae)

The role of Chinese cabbage as a trap crop for
flea beetles (Coleoptera: Chrysomelidae)
in production of white cabbage
Stanislav Trdan *, Nevenka Valic, Dragan Znidarcic,Matej Vidrih, Klemen Bergant, Emil Zlatic, Lea Milevoj
University of Ljubljana, Biotechnical Faculty, Department of Agronomy, Chair of Entomology and
Phytopathology, Jamnikarjeva 101, SI-1111 Ljubljana, Slovenia
Received 7 April 2004; received in revised form 27 October 2004; accepted 7 March 2005
Abstract
During the years 2002 and 2003, preference of flea beetles, Phyllotreta spp., to white and Chinese
cabbage, grown in monoculture and in mixed crop, was tested. The aim of the research was to
determine if Chinese cabbage is an appropriate trap crop for this pest in the production of white
cabbage, an important vegetable in Europe and in North America. The number of beetles on Chinese
cabbage in monoculture and in mixed crop did not differ significantly. In both treatments the number
of adults of flea beetles on Chinese cabbage and the percentage of damaged leaf area they caused,
were significantly higher than that on white cabbage. Statistically significant and positive correlation
was established between leaf damage and number of flea beetles. It was stronger in 2003, which was
less favorable for the crop with regards to the weather conditions (drought and high air temperatures).
No significant differences were found in the number of adults and in most evaluations also in the
damage assessments on white cabbage when grown in monoculture and in mixed culture. Therefore,
we concluded that Chinese cabbage grown in mixed crop with white cabbage is not a suitable control
measure for reducing the damage caused by flea beetles.
# 2005 Elsevier B.V. All rights reserved.
Keywords: Phyllotreta spp.; Trap cropping; White cabbage; Chinese cabbage; IPM
www.elsevier.com/locate/scihorti
Scientia Horticulturae 106 (2005) 12–24
* Corresponding author. Tel.: +386 1 423 11 61; fax: +386 1 423 10 88.
E-mail address: [email protected] (S. Trdan).
0304-4238/$ – see front matter # 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.scienta.2005.03.005

1. Introduction
Flea beetles, Phyllotreta spp., are among the most important pests of cultivated
brassicas in Europe and North America (Stoner, 1992; Ester et al., 2003). Beetles can
cause substantial damage on host plants by feeding on the leaves, especially in early
stages of development (Palaniswamy and Lamb, 1992). In Central Europe and in
continental parts of Southern Europe, brassicas are planted in open fields in the
beginning of May (early white cabbage varieties), which usually coincides with high
population densities of flea beetles. Prevalent species of flea beetles are Phyllotreta
nemorum L., Phyllotreta undulata Kutschera, Phyllotreta cruciferae Goeze, and in some
districts also Phyllotreta vittula (Redtenbacher). In these areas flea beetles usually have
one generation per year (Vig, 2000) and prevail in the open until temperatures are too
low (end of October). Flea beetles cause less damage to older plants, because these
plants can ‘resist’ the pest with greater leaf surface. Beside causing a direct damage, flea
beetles can also be a vector for some plant pathogens (Dillard et al., 1998; Stobbs et al.,
1998).
Seedlings of late white cabbage (Brassica oleracea L. convar. capitata [L.] Alef. var.
alba DC.) varieties in Europe are planted in open fields from the beginning of July and
can also be important hosts to flea beetles. Beetles cause more damage in warm season,
when they feed more intensively, thus compensating a loss of water. Due to climatic
changes the hot season has been prolonging and average temperatures are higher than
they were few decades ago, which can also influence the pest bionomics (Yamaguchi
et al., 2001; Fuhrer, 2003). Brassicas seedlings can hardly recover from severe attack by
flea beetles, thus the use of insecticides is still the most common pest control strategy
applied in the early stages of plant development in white cabbage production.
Consequently, flea beetles have developed resistance to most of insecticides (Turnock
and Turnbull, 1994).
To reduce the use of pesticides, new environment friendly methods of pest control are
being developed. In the past 20 years, considerable research has been done in the field of
intercropping, and it has become an important part of integrated pest management (IPM) in
many places (Finch and Kienegger, 1997; Held et al., 2003). Compared with intercropping,
trap cropping is a less investigated method of an environment friendly pest control
(Boucher et al., 2003; Smyth et al., 2003). In this method trap plants (plants to which pests
are more attracted) are planted among the main crop. Pest populations on trap plants are
more numerous and so only these plants are treated with insecticides. The quantity of
pesticides is thus reduced, as well as the pest population. Beside direct influence (pests are
more susceptible to a trap plant), trap cropping and intercropping also have an indirect
impact: in mixed crops the level of parasitism is usually higher (Hokkanen, 1989), which
also reduces the damage.
In the literature some efficient models of trap cropping are reported (Asman, 2002; Rea
et al., 2002), though a combination of white cabbage and Chinese cabbage (Brassica
pekinensis [Lour.] Rupr.), both very popular vegetables in Europe, was not stated. Out of
practical experiences it is known that flea beetles cause more damage on Chinese cabbage
than on white cabbage (Kinoshita et al., 1979). As the growth and development of Chinese
cabbage and late white cabbage varieties coincides, the suitability of Chinese cabbage as a
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–24 13

trap plant for flea beetles in mixed crop with white cabbage was investigated in our study.
Our hypothesis was that the Chinese cabbage, as a trap plant, diverts the beetles from white
cabbage plants and thus reduces the damage caused by the beetles. To test the hypothesis,
differences between susceptibility of flea beetles to white cabbage and Chinese cabbage in
mixed crop and in monoculture crop were evaluated together with the differences in the
extent of damage on leaves.
2. Materials and methods
2.1. Plant material and insects
A 2-year experiment (2002–2003) was carried out in the field (Biotechnical Faculty in
Ljubljana, Slovenia, 468040 N latitude, 148310 E longitude, 300 m above sea level, UTM
33TVM50). Different brassicas, mostly white cabbage, have been grown on this location
for many years for research purposes, therefore population of flea beetles, Phyllotreta spp.,
there is permanent and numerous.
2.2. Agrotechnique
The site was a heavy clay soil with 2.6% organic matter content and 6.8 pH.
The climate at the site is a temperate continental. Seedlings of white and Chinese
cabbage were seeded in cell trays, one seed per cell (volume 33 cm3), filled with
commercial peat lite mix (1:1, v:v). The trays were placed in a greenhouse for 30 days,
watered each morning and fertilized once a week with Peter’s soluble 20-20-20
(N–P2O5–K2O).
Experimental plots were prepared with a rotary tiller cultivator. The seedlings were
transplanted on 1 August 2002 and on 4 September 2003 into raised beds mulched with
black polyethylene (15 mm thick). Beds were 1.3 m wide � 15 cm high, with 1.8 m
between the centers of contiguous beds. In both years, the previous crop was lettuce.
General agricultural procedures used in commercial brassicas production were followed
for both years, except the use of insecticides. Fertilizer was supplied in three split
applications of N, for a total of 240 kg ha�1, and a single application of P2O5 and K2O at
land preparation, for a total of 320 and 400 kg ha�1, respectively. The irrigation system
consisted of trickle-irrigation tubing laid on the bed. Irrigation for each treatment
was supplied by a combination of drip irrigation and natural rainfall, according to
evapotranspiration data. Water amounts were controlled for all treatments with
tensiometer.
2.3. Treatments
Experiment was designed as randomized complete block (3) with four treatments, each
replicated three times. Treatments were a monoculture of white cabbage, a monoculture of
Chinese cabbage and a mixed crop of the two host plants species (random distribution of
seedlings). Mixed crop were included in two different treatments, where both brassicas
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–2414

were planted identically, but each designated for separate crop to analyze. Forty seedlings
of white cabbage (variety ‘Ditmar’), 40 seedlings of Chinese cabbage (variety ‘Nagaoka’)
and 20 seedlings from both brassicas (mixed crops in two separate treatments) were
transplanted for each repetition. The spacing was 30 cm � 45 cm. The plot size
(experimental unit) was 4.0 m � 1.3 m, having four rows, each with 10 plants. Adjacent
plots were separated with non-planted buffer area (1.0 m � 1.3 m) with an aim to diminish
the impact of plants in neighboring plots. Only 15 randomly selected plants near the center
of each plot were sampled to eliminate confounding effect of adjacent plots as much as
possible. The extent of damaged leaf area caused by flea beetles was assessed four times in
2002 and three times in 2003. Additionally, flea beetles were counted ones in 2002 and
twice in 2003 (Table 1).
2.4. Field observations
EPPO directive (OEPP/EPPO, 2002) was used for evaluation of the extent (percentage)
of damaged leaf area. In the natural infestation trial the leaves were qualitatively assessed
on a scale from 1 (no damage) up to 5 (more than 25% leaf area eaten). The intermediate
values in the scale expressed the damage as follows: 2—up to 2% leaf area eaten,
3—between 3 and 10% leaf area eaten, and 4—between 11 and 25% leaf area eaten. The
growth stages of the plants were established using the BBCH scale for leaf vegetables
(forming heads) (BBA, 2001). The scale includes the principal growth stages as follows:
0—germination, 1—leaf development (main shot), 2—development of harvestable
vegetative plant parts, 5—inflorescence emergence, 6—flowering, development of fruit,
8—ripening of fruit and seed, and 9—senescence.
To minimize the migration of beetles between plants, all evaluations were carried out in
the morning, at about 8 a.m. with air temperature under 15 8C. Beetles were counted only
on the upper side of the leaf, as it is known that flea beetles mostly feed on this side (Vargas
and Kershaw, 1979). Moving the leaves or using some instruments (i.e. vacuum aspirator)
would disturb the flea beetles that would consequently aband the plants. As a result, the
data would be less accurate than in the case of our method. Insects were only counted as
long as leaves did not overlap (PGS 1 � 18).
2.5. Data analysis
An analysis of variance (ANOVA) was employed to assess the differences in the number
of flea beetles per plant, and in the percentage of damaged leaf area among the treatments.
Before analysis, each variable was tested for homogeneity of treatment variances. If
variances were not homogeneous, data were transformed to log(Y) before ANOVA.
Student–Newman–Keuls multiple range test (*P � 0.05) was used to separate mean
differences among parameters in all treatments. The relationship between mean damage
value, caused by feeding, and mean number of flea beetles was evaluated using linear and
polynomial regression analysis. All statistical analyses were performed with Statgraphics
Plus for Windows 4.0 (Statistical Graphics Corp., Manugistics, Inc.) and figures were
created with Sigmaplot 2002 for Windows 8.0 (Systat Software, Inc.). Data are presented
as untransformated mean � S.E.
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–24 15

S.
Trd
an
eta
l./Scien
tiaH
orticu
ltura
e1
06
(20
05
)1
2–
24
16
Table 1
Dates of flea beetles, Phyllotreta spp., countings and damage evaluations on leaves of white cabbage (C) and Chinese cabbage (CC) in 2002 and 2003
Year 2002 2003
No. of flea
beetles
Damaged leaf
area (%)
Growth stage (BBCH) No. of flea
beetles
Damaged leaf
area (%)
Growth stage (BBCH)
C CC C CC
First evaluation 14 August 14 August aPGS 1: 16–19 PGS 1: 18–19 18 September 18 September PGS 1: 14 PGS 1: 16
Second evaluation – 22 August PGS 1: 17–19 PGS 1: 19 30 September 30 September PGS 1: 17 PGS 1: 17–18
Third evaluation – 3 September bPGS 4: 41–42 PGS 4: 41 – 17 October PGS 1: 18–19 PGS 1: 19
Fourth evaluation – 20 September PGS 4: 44–47 PGS 4: 43–47 – – – –
a Principal growth stage 1: leaf development (main shot); 10—cotyledons completely unfolded, growing point or true leaf initial visible; 19—nine or more true leaves
unfolded.b Principal growth stage 4: development of harvestable vegetative plant parts; 41—heads begin to form, the two youngest leaves do not unfold; 49—typical size, form and
firmness of heads reached.

3. Results
For some of the treatments in both years, statistically significant differences were found
in the extent of damaged leaf area, caused by feeding of flea beetles, Phyllotreta spp., on
leaves of white cabbage and Chinese cabbage grown in monoculture and in mixed crop.
Differences were also found in some treatments in the occurrence of flea beetles on both
brassicas.
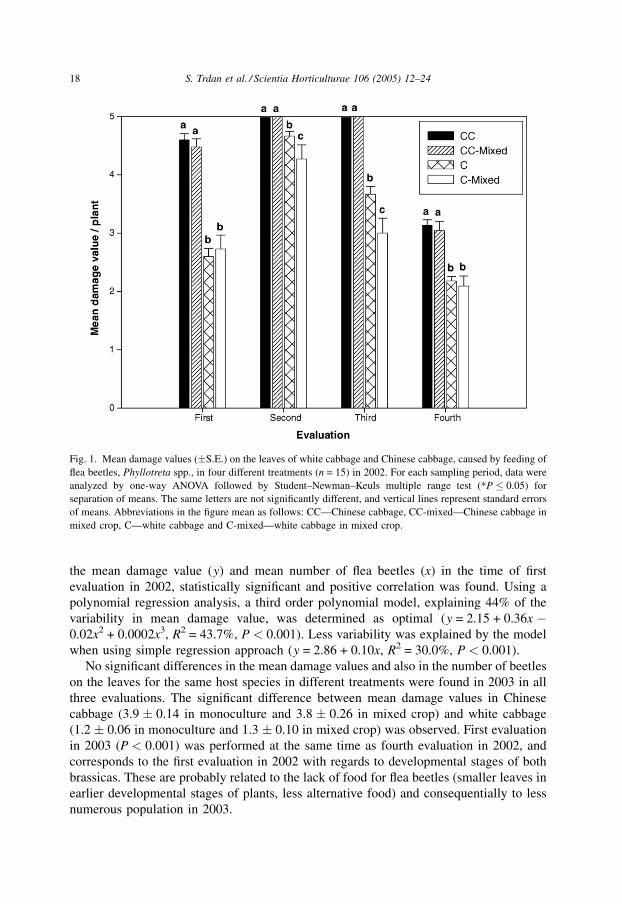
In the first evaluation in 2002 (P < 0.001), the most extended injuries were observed on
leaves of Chinese cabbage (from 8 up to 11 true leaves unfolded) grown in monoculture
(4.6 � 0.10), and in mixed crop with white cabbage (4.5 � 0.14). No statistically
significant differences were observed between these two treatments. In addition, no
statistically significant differences were found between the mean damage values for the
leaves of white cabbage (from 6 up to 10 true leaves unfolded) in monoculture (2.6 � 0.14)
and in mixed crop (2.7 � 0.24). On the other hand, mean damage values for white cabbage
were significantly lower than for Chinese cabbage for both treatments.
The results were slightly different in the second evaluation in 2002 (P < 0.001). The
same mean damage values were determined for Chinese cabbage (from 10 up to 17 true
leaves unfolded) grown in monoculture and in mixed crop (5.0 � 0.00). Therefore, no
statistically significant differences were found between these two treatments. On the
contrary, to the first evaluation, there were significant differences in mean damage values
for white cabbage (from 7 up to 10 true leaves unfolded) for different treatments. The value
was higher (4.7 � 0.08) when white cabbage was grown in monoculture than when grown
in mixed crop (4.3 � 0.24).
Statistically significant differences between some of the treatments were also found in
the third evaluation (P < 0.001), when both brassicas were in the stage BBCH 41–42
(beginning to form heads). Mean damage values for the white cabbage grown in
monoculture were significantly higher (3.7 � 0.14) than for the white cabbage grown in
mixed crop (3.0 � 0.25), but for both treatments lower than in second evaluation. Mean
damage values for Chinese cabbage were comparable for both treatments (5.0 � 0.00) and
significantly higher than for white cabbage. In the final (fourth) evaluation in 2002
(P < 0.001), when brassicas were in the stage BBCH 43–44 (30–70% of the expected head
size reached), mean damage values on the leaves of both brassicas were the lowest
compared with other three evaluations. The highest mean damage values (3.1 � 0.09) were
observed for Chinese cabbage grown in monoculture and in mixed crop with white cabbage
(3.0 � 0.16). The extent of damages on white cabbage was on average 1 grade lower
(2.2 � 0.08 in monoculture, and 2.1 � 0.17 in mixed crop). Generally, injuries on the
leaves of both brassica plants were on average assessed with higher grades in earlier
developmental stages (Fig. 1).
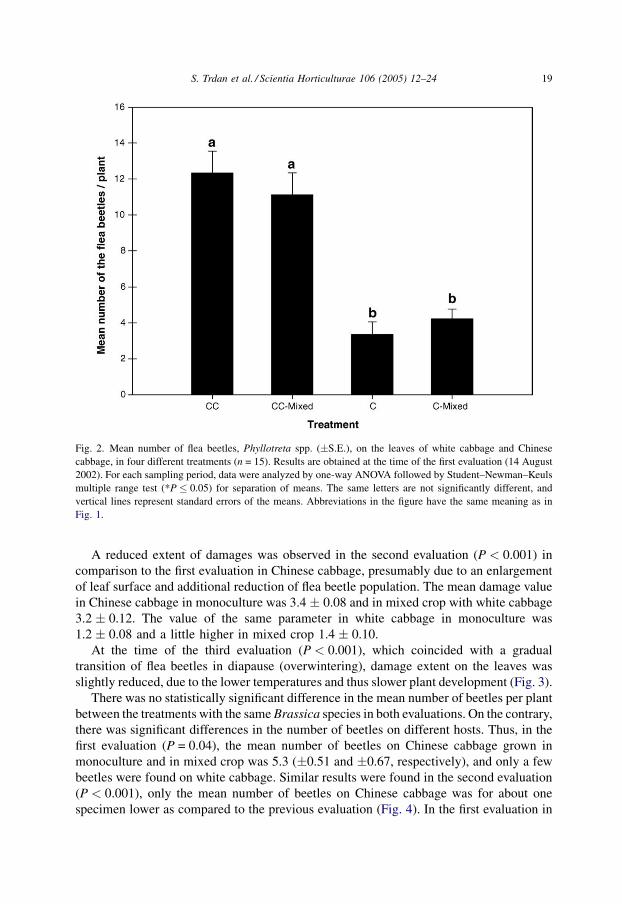
Significant differences were observed in the mean number of flea beetles, Phyllotreta
spp., on white cabbage and Chinese cabbage in 2002 (P < 0.001), irrespective of
treatment. The number of beetles on Chinese cabbage grown in monoculture (12.3 � 1.21)
and in mixed crop (11.1 � 1.21) was significantly higher in comparison to the number of
beetles on white cabbage in monoculture (3.4 � 0.70) and in mixed crop (4.2 � 0.54
beetles per plant). The number was not significantly different on the level of the same host
species but different treatment (CC versus CC-mixed, C versus C-mixed) (Fig. 2). Between
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–24 17
the mean damage value (y) and mean number of flea beetles (x) in the time of first
evaluation in 2002, statistically significant and positive correlation was found. Using a
polynomial regression analysis, a third order polynomial model, explaining 44% of the
variability in mean damage value, was determined as optimal (y = 2.15 + 0.36x �0.02x2 + 0.0002x3, R2 = 43.7%, P < 0.001). Less variability was explained by the model
when using simple regression approach (y = 2.86 + 0.10x, R2 = 30.0%, P < 0.001).
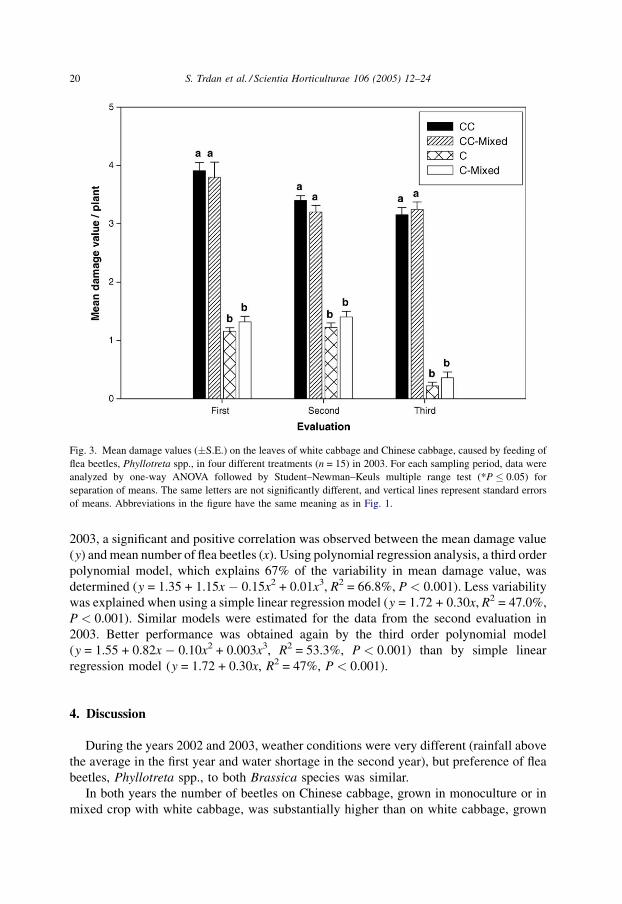
No significant differences in the mean damage values and also in the number of beetles
on the leaves for the same host species in different treatments were found in 2003 in all
three evaluations. The significant difference between mean damage values in Chinese
cabbage (3.9 � 0.14 in monoculture and 3.8 � 0.26 in mixed crop) and white cabbage
(1.2 � 0.06 in monoculture and 1.3 � 0.10 in mixed crop) was observed. First evaluation
in 2003 (P < 0.001) was performed at the same time as fourth evaluation in 2002, and
corresponds to the first evaluation in 2002 with regards to developmental stages of both
brassicas. These are probably related to the lack of food for flea beetles (smaller leaves in
earlier developmental stages of plants, less alternative food) and consequentially to less
numerous population in 2003.
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–2418
Fig. 1. Mean damage values (�S.E.) on the leaves of white cabbage and Chinese cabbage, caused by feeding of
flea beetles, Phyllotreta spp., in four different treatments (n = 15) in 2002. For each sampling period, data were
analyzed by one-way ANOVA followed by Student–Newman–Keuls multiple range test (*P � 0.05) for
separation of means. The same letters are not significantly different, and vertical lines represent standard errors
of means. Abbreviations in the figure mean as follows: CC—Chinese cabbage, CC-mixed—Chinese cabbage in
mixed crop, C—white cabbage and C-mixed—white cabbage in mixed crop.
A reduced extent of damages was observed in the second evaluation (P < 0.001) in
comparison to the first evaluation in Chinese cabbage, presumably due to an enlargement
of leaf surface and additional reduction of flea beetle population. The mean damage value
in Chinese cabbage in monoculture was 3.4 � 0.08 and in mixed crop with white cabbage
3.2 � 0.12. The value of the same parameter in white cabbage in monoculture was
1.2 � 0.08 and a little higher in mixed crop 1.4 � 0.10.
At the time of the third evaluation (P < 0.001), which coincided with a gradual
transition of flea beetles in diapause (overwintering), damage extent on the leaves was
slightly reduced, due to the lower temperatures and thus slower plant development (Fig. 3).
There was no statistically significant difference in the mean number of beetles per plant
between the treatments with the same Brassica species in both evaluations. On the contrary,
there was significant differences in the number of beetles on different hosts. Thus, in the
first evaluation (P = 0.04), the mean number of beetles on Chinese cabbage grown in
monoculture and in mixed crop was 5.3 (�0.51 and �0.67, respectively), and only a few
beetles were found on white cabbage. Similar results were found in the second evaluation
(P < 0.001), only the mean number of beetles on Chinese cabbage was for about one
specimen lower as compared to the previous evaluation (Fig. 4). In the first evaluation in
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–24 19
Fig. 2. Mean number of flea beetles, Phyllotreta spp. (�S.E.), on the leaves of white cabbage and Chinese
cabbage, in four different treatments (n = 15). Results are obtained at the time of the first evaluation (14 August
2002). For each sampling period, data were analyzed by one-way ANOVA followed by Student–Newman–Keuls
multiple range test (*P � 0.05) for separation of means. The same letters are not significantly different, and
vertical lines represent standard errors of the means. Abbreviations in the figure have the same meaning as in
Fig. 1.
2003, a significant and positive correlation was observed between the mean damage value
(y) and mean number of flea beetles (x). Using polynomial regression analysis, a third order
polynomial model, which explains 67% of the variability in mean damage value, was
determined (y = 1.35 + 1.15x � 0.15x2 + 0.01x3, R2 = 66.8%, P < 0.001). Less variability
was explained when using a simple linear regression model (y = 1.72 + 0.30x, R2 = 47.0%,
P < 0.001). Similar models were estimated for the data from the second evaluation in
2003. Better performance was obtained again by the third order polynomial model
(y = 1.55 + 0.82x � 0.10x2 + 0.003x3, R2 = 53.3%, P < 0.001) than by simple linear
regression model (y = 1.72 + 0.30x, R2 = 47%, P < 0.001).
4. Discussion
During the years 2002 and 2003, weather conditions were very different (rainfall above
the average in the first year and water shortage in the second year), but preference of flea
beetles, Phyllotreta spp., to both Brassica species was similar.
In both years the number of beetles on Chinese cabbage, grown in monoculture or in
mixed crop with white cabbage, was substantially higher than on white cabbage, grown
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–2420
Fig. 3. Mean damage values (�S.E.) on the leaves of white cabbage and Chinese cabbage, caused by feeding of
flea beetles, Phyllotreta spp., in four different treatments (n = 15) in 2003. For each sampling period, data were
analyzed by one-way ANOVA followed by Student–Newman–Keuls multiple range test (*P � 0.05) for
separation of means. The same letters are not significantly different, and vertical lines represent standard errors
of means. Abbreviations in the figure have the same meaning as in Fig. 1.

both ways. In 2002, the number of beetles in treatments CC and C was at ratio 3.67:1, and in
treatments CC-mixed and C-mixed at ratio 2.63:1. Considering trap cropping, the ratio C:C-
mixed 0.81:1 is the most relevant data (2002). This means that white cabbage in mixed crop
with Chinese cabbage is more exposed to flea beetle attack than if grown in monoculture. The
reason is that the number of beetles on Chinese cabbage is so high – for the reason of
intraspecific competition (Quiring and Timmins, 1990; Lucas et al., 1995) – that they migrate
to neighboring plants, in this case to white cabbage. In 2003 damage evaluation started
approximately 1 month later (beginning of September, due to very high temperatures which
enabled the seedlings to survive in August) than in 2002. The ratio between number of beetles
on both brassicas in 2003, grown in the same way (CC:C and CC -mixed:C-mixed) was over
10 times higher. Differences in susceptibility of the pest to both brassicas in both years are
mostly of abiotic nature, because moderate temperatures at the beginning of the experiment
in 2002 were more favorable for the pest and its hosts, as it is known for many phytophagous
insects and plants they attack (Raj et al., 1995; Tang et al., 1999).
The main reason for substantially greater values of CC:C and CC-mixed: C-mixed in
2003 than in 2002 is in the lower number of beetles on white cabbage in the second year.
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–24 21
Fig. 4. Mean number of flea beetles, Phyllotreta spp. (�S.E.), on the leaves of white cabbage and Chinese
cabbage, in four different treatments (n = 15). Results are obtained at the time of the first and second evaluation (18
September and 30 September 2003). For each sampling period, data were analyzed by one-way ANOVA followed
by Student–Newman–Keuls multiple range test (*P � 0.05) for separation of means. The same letters are not
significantly different, and vertical lines represent standard errors of the means. Abbreviations in the figure have
the same meaning as in Fig. 1.

The number of beetles on white cabbage at the time of first evaluation (when
developmental stages of both brassicas were comparable) was more than 26-times lower
(C-mixed2002:C-mixed2003 = 26.44:1). The exact ratio of number of beetles in monoculture
was not determined, since in first evaluation in 2003 not even a single beetle was found.
Ratio between C and C-mixed (0.00 beetles per plant/0.16 beetles per plant) in first
evaluation in 2003 is less relevant than the same ratio in second evaluation (0.42:1) in the
same year. Due to more favorable weather conditions in 2002, the ratio C:C-mixed for this
year was considered as more relevant. Results have thus confirmed previously known
statements of higher preference of flea beetles to Chinese cabbage, in comparison to white
cabbage (Kinoshita et al., 1979).
In 2002, which was more favorable for growing both brassicas, mean extent of damage
leaf area on Chinese cabbage in both treatments was very high (mean damage value about
4.5) already in the first evaluation. At the time of the second evaluation, all plants of
Chinese cabbage were assessed with the highest number on the scale. The situation was the
same in the third evaluation, while in the fourth evaluation about 10% of leaf area was
damaged on Chinese cabbage. This is probably the result of quick growth of leaf surface,
and natural decrease of population of the pest, bearing in mind that feeding of flea beetles
on damaged plants increases its natural mortality (Traw and Dawson, 2002).
Mean damage value on white cabbage, grown in monoculture or in mixed crop with
Chinese cabbage were substantially lower as compared with Chinese cabbage. Leaves of
white cabbage were most damaged at the time of the second evaluation, when the mean
damage value was more than 4. The population of flea beetles in 2003 was substantially
lower than in 2002 and so was the damage on the leaves of both brassicas. The extent of
damage on leaves of Chinese cabbage was the highest in first evaluation in both treatment.
The same parameter on leaves of white cabbage was surprisingly the highest in the second
evaluation. We assume that this is due to a slower growth of white cabbage and so the
leaves in second evaluation were already previously assessed. On the other hand, Chinese
cabbage grew more rapidly, thus compensated damage observed in first evaluation.
As the damage on leaves of both brassicas is not influenced only by the number of flea
beetles, but also by several biotic and abiotic factors, the direct impact of the number of flea
beetles on the damage was investigated. Polynomial regression was found as an appropriate
method for describing such relationship. Third order polynomial model explained 44–67%
of variability in damage as a function of the number of flea beetles. In more favorable years
for brassicas growing (in our case 2002), other factors beside the number of beetles
intensity the damage, caused primarily by the flea beetles feeding on leaves. Favorable
conditions for brassicas growing are also favorable for flea beetles, which migrate more
intensively between plants and between parcels. Less variability explained by the
polynomial model in the second evaluation in year 2003 (R2 = 0.53 versus 67%) can also be
explained as the holes on leaves were a result of flea beetles feeding during both – first and
the second – evaluations.
According to the results derived from the 2-year experiment we concluded, that Chinese
cabbage growing as a trap plant for flea beetles in mixed crop with white cabbage, is not a
sufficient control measure, because no statistically significant differences were found
between the number of beetles on white cabbage, grown in monoculture and in mixed
culture in a majority of evaluations. Obviously, Chinese cabbage is a very susceptible host
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–2422

for flea beetles, so the mass occurrence of beetles on this vegetable also causes higher
migration of beetles to white cabbage, thus causing a quite important extent of damage also
on white cabbage. On the other hand, no statistically significant differences were observed
in the number of beetles as well as in the damage extent on leaves of Chinese cabbage
grown in two different ways. Values of both parameters were in all evaluations statistically
and significantly higher from adequate values on white cabbage. We suggest that selective
chemical control of flea beetles on Chinese cabbage plants, grown in mixed crops where
white cabbage is the main crop, could diminish harmfulness of flea beetles, which will be in
Europe probably also in the future one of the most important pest of brassicas in early
developmental stages. In the area where this research was performed, early cabbage
varieties are harvested between the end of June and beginning of July, so spraying Chinese
cabbage with insecticides is recommended only in production of late white cabbage.
Acknowledgement
The authors would like to thank Irena Skapin Osborne for proof reading and editing the
draft article.
References
Asman, K., 2002. Trap cropping effect on oviposition behaviour of the leek moth Acrolepiopsis assectella and the
diamondback moth Plutella xylostella. Entomol. Exp. App. 105, 153–164.
BBA. 2001. Growth stages of mono- and dicotyledonous plants. http://www.bba.de/veroeff/bbch/bbcheng.pdf.
Boucher, T.J., Ashley, R., Durgy, R., Sciabarrasi, M., Calderwood, W., 2003. Managing the pepper maggot
(Diptera: Tephritidae) using perimeter trap cropping. J. Econ. Entomol. 96, 420–432.
Desh Raj, Nirmala Devi, Chandel, Y.S., 1995. Infestation of leaf miner, Chromatomyia horticola Goureau on
Brassica campestris in mid hill zone of Himachal Pradesh. J. Entomol. Res. 19, 107–110.
Dillard, H.R., Cobb, A.C., Lamboy, J.S., 1998. Transmission of Alternaria brassicicola to cabbage by flea beetles
(Phyllotreta cruciferae). Plant Dis. 82, 153–157.
Ester, A., de Putter, H., van Bilsen, J.G.P.M., 2003. Filmcoating the seed of cabbage (Brassica oleracea L. convar.
bapitata L.) and cauliflower (Brassica oleracea L. var. botrytis L.) with imidacloprid and spinosad to control
insect pests. Crop Prot. 22, 761–768.
Finch, S., Kienegger, M., 1997. A behavioural study to help clarify how undersowing with clover affects host-plant
selection by pest insects of brassica crops. Entomol. Exp. Appl. 84, 165–172.
Fuhrer, J., 2003. Agroecosystem responses to combinations of elevated CO2, ozone, and global climate change.
Agric. Ecosyst. Environ. 97, 1–20.
Held, D.W., Gonsiska, P., Potter, D.A., 2003. Evaluating companion planting and non-host masking odors for
protecting roses from the Japanese beetle (Coleoptera: Scarabaeidae). J. Econ. Entomol. 96, 81–87.
Hokkanen, H.M.T., 1989. Biological and agrotechnical control of the rape blossom beetle Meligethes aeneus
(Coleoptera Nitidulidae). Acta Entomol. Fenn. 53, 25–29.
Kinoshita, G.B., Svec, H.J., Harris, C.R., McEwen, F.L., 1979. Biology of the crucifer flea beetle, Phyllotreta
cruciferae (Coleoptera: Chrysomelidae), in southwestern Ontario. Can. Entomol. 111, 1395–1407.
Lucas, E., de Oliveira, D., Houle, M.J., 1995. Intraspecific competition by the Colorado potato beetle (Coleoptera:
Chrysomelidae) on potato plants, Solanum tuberosum. Environ. Entomol. 24, 576–580.
OEPP/EPPO, 2002. Guidelines for the efficacy evaluation of insecticides. Phyllotreta spp. on rape. Bull. OEPP/
EPPO Bull. 32, 361–365.
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–24 23

Palaniswamy, P., Lamb, R.J., 1992. Host preferences of the flea beetles Phyllotreta cruciferae and P. striolata
(Coleoptera: Chrysomelidae) for crucifer seedlings. J. Econ. Entomol. 85, 743–752.
Quiring, D.T., Timmins, P.R., 1990. Influence of reproductive ecology on feasibility of mass trapping Diabrotica
virgifera virgifera (Coleoptera: Chrysomelidae). J. Appl. Ecol. 27, 965–982.
Rea, J.H., Wratten, S.D., Sedcole, R., Cameron, P.J., Davis, S.I., Chapman, R.B., 2002. Trap cropping to manage
green vegetable bug Nezara viridula (L.) (Heteroptera: Pentatomidae) in sweet corn in New Zealand. Agric.
For. Entomol. 4, 101–107.
Smyth, R.R., Hoffmann, M.P., Shelton, A.M., 2003. Effects of host plant phenology on oviposition preference of
Crocidolomia pavonana (Lepidoptera: Pyralidae). Environ. Entomol. 32, 756–764.
Stobbs, L.W., Cerkauskas, R.F., Lowery, T., van Driel, L., 1998. Occurrence of turnip yellow mosaic virus on
Oriental cruciferous vegetables in southern Ontario, Canada. Plant Dis. 82, 351.
Stoner, K.A., 1992. Density of imported cabbageworms (Lepidoptera: Pieridae), cabbage aphids (Homoptera:
Aphididae), and flea beetles (Coleoptera: Chrysomelidae) on glossy and trichome-bearing lines of Brassica
oleracea. J. Econ. Entomol. 85, 1023–1030.
Tang, Y.Q., Lapointe, S.L., Brown, L.G., Hunter, W.B., 1999. Effects of host plant and temperature on the biology
of Toxoptera citricida (Homoptera: Aphididae). Environ. Entomol. 28, 895–900.
Theunissen, J., Booij, C.J.H., Lotz, L.A.P., 1995. Effects of intercropping white cabbage with clovers on pest
infestation and yield. Entomol. Exp. Appl. 74, 7–16.
Traw, M.B., Dawson, T.E., 2002. Reduced performance of two specialist herbivores (Lepidoptera: Pieridae,
Coleoptera: Chrysomelidae) on new leaves of damaged black mustard plants. Environ. Entomol. 31, 714–722.
Turnock, W.J., Turnbull, S.A., 1994. The development of resistance to insecticides by the crucifer flea beetle,
Phyllotreta cruciferae (Goeze). Can. Entomol. 126, 1369–1375.
Vargas, P., Kershaw, W.J., 1979. Host selection and choice of feeding site by the flea beetle Phyllotreta undulata
Kutsch. An. Inst. Nac. Investig. Agrar., Prot. Veg. 10, 81–93.
Vig, K., 2000. Data on the biology of the turnip flea beetle, Phyllotreta nemorum (Linnaeus, 1758) (Coleoptera,
Chrysomelidae Alticinae). Med. Fac. Landbouww. Univ. Gent 65, 201–212.
Yamaguchi, T., Kiritani, K., Matsuhira, K., Fukuda, K., 2001. The influence of unusual hot weather on the
occurrence of several arthropod crop pests. Jpn. J. Appl. Entomol. Zool. 45, 1–7.
S. Trdan et al. / Scientia Horticulturae 106 (2005) 12–2424
Copyright © 2022 FDOKUMEN




















