Influence of transonic inducer design on the performance of ...
Upload
independentCategory
view
3download
0
The FASEB Journal • Research Communication
Extracellular matrix metalloproteinase inducer/CD147promotes myofibroblast differentiation by inducing�-smooth muscle actin expression and collagen gelcontraction: implications in tissue remodeling
Eric Huet,* Benoit Vallee,* Dominika Szul,* Franck Verrecchia,† Samia Mourah,‡
James V. Jester,§ Thanh Hoang-Xuan,� Suzanne Menashi*,1 and Eric E. Gabison*,�
*CRRET Laboratory, CNRS UMR 7149, University of Paris XII, Creteil, France; †INSERM U697 and‡INSERM U716, Laboratoire de Pharmacologie, Hopital Saint-Louis, Paris, France; §Department ofOphthalmology, University of California at Irvine, Irvine, California, USA; and �Department ofOphthalmology at Fondation Ophtalmologique A. de Rothschild and Bichat Hospital, AP-HP,Paris, France
ABSTRACT Extracellular matrix metalloproteinase in-ducer (EMMPRIN) is a cell surface glycoprotein enrichedon tumor cells and normal epithelia. It is mainly knownfor its ability to induce matrix metalloproteinase produc-tion in fibroblasts following epithelial-stromal interaction.We sought to examine whether EMMPRIN has a broaderrole promoting fibroblast-to-myofibroblast differentia-tion. Because �-smooth muscle actin (�SMA) is consid-ered a marker of this differentiation process, we analyzedthe effect of EMMPRIN on its expression in corneal andskin fibroblasts by Western blots, immunocytochemistry,and a functional assay of collagen lattice contraction.Increasing EMMPRIN expression by cDNA transfectionor by treatment with exogenously added recombinantEMMPRIN resulted in an up-regulation of �SMA expres-sion. EMMPRIN also increased the contractile propertiesof the treated fibroblasts as demonstrated by the immu-nohistochemical appearance of stress fibers and by theaccelerated contraction of fibroblast-embedded collagenlattices. Blocking EMMPRIN expression by small interfer-ing RNA inhibited �SMA and collagen gel contractioninduced not only by EMMPRIN but also by transforminggrowth factor-�, a major mediator of myofibroblast dif-ferentiation that also regulated EMMPRIN expression.These findings, combined with the fact that EMMPRINand �SMA colocalized to the same cells in the stroma ofpathological corneas, expand on the mechanism by whichEMMPRIN remodels extracellular matrix during woundhealing and cancer.—Huet, E., Vallee, B., Szul, D., Ver-recchia, F., Mourah, S., Jester, J. V., Hoang-Xuan, T.,Menashi, S., Gabison, E. E. Extracellular matrix metal-loproteinase inducer/CD147 promotes myofibroblastdifferentiation by inducing �-smooth muscle actin ex-pression and collagen gel contraction: implications intissue remodeling. FASEB J. 22, 1144–1154 (2008)
Key Words: epithelial-stromal interactions � wound healing� MMPs � TGF�
In response to injury, resident cells in tissue adjacentto the damaged area initiate a cascade of events necessaryto restore tissue function. The wound repair process,regulated by a number of cytokines and growth factors, ischaracterized by the activation of quiescent fibroblastsand their differentiation into myofibroblasts, which thenleads to a wound contraction and extracellular matrix(ECM) remodeling. The differentiation of fibroblast tomyofibroblast can be identified by the expression of theactin isoform typical of vascular smooth muscle cells,�-smooth muscle actin (�SMA), which characterizes themyofibroblasts both in vitro and in vivo. The presence of�SMA stress fibers is generally related to the contractileactivities of the myofibroblasts, because a direct correla-tion was demonstrated between the level of �SMA andmyofibroblast contraction (1). The cytokine transforminggrowth factor-� (TGF�) is considered to be the majoractor in the differentiation of fibroblasts into myofibro-blasts because it is capable of up-regulating �SMA infibroblasts (2–4).
In addition to stimulating �SMA expression, TGF� isknown to be a potent stimulator of connective tissueformation as it controls both the expression of compo-nents of the ECM network, such as fibrillar collagensand fibronectin, and the expression of protease inhib-itors, including plasminogen activator inhibitor-1 ortissue inhibitors of metalloproteinases (TIMPs) (5). Bycontrast, the turnover of the laid down matrix isregulated by the activity of matrix metalloproteinases(MMPs), a family of enzymes capable of degradingalmost all ECM proteins. Therefore, the balance be-tween matrix formation and deposition by cytokineslike TGF� and their degradation by MMPs seeminglydictates the outcome of the healing process, whether
1 Correspondence: CRRET Laboratory, CNRS UMR 7149,University Paris XII, 61 Av du General de Gaulle, 94010Creteil Cedex, France. E-mail: [email protected]
doi: 10.1096/fj.07-8748com
1144 0892-6638/08/0022-1144 © FASEB

normal, hypertrophic/fibrotic or thinning/ulceration.In addition to its effect on the stimulation of ECMsynthesis and proteases inhibitors, TGF� can itselfregulate the expression of MMPs, as it was shown todown-regulate the interstitial collagenase (MMP-1) andstromelysin (MMP-3) but to increase MMP-2 andMMP-9 production in several cell types (6–9). CertainMMPs such as MMP-2, MMP-9, and MT1-MMP can inturn activate latent TGF� (10, 11), thus demonstratingthe complex interrelations that may exist in regulatingmatrix synthesis and degradation. One would expecttherefore that the myofibroblast, considered the keycell for the connective tissue remodeling that takesplace during wound healing, would be equipped withan elaborate regulatory system necessary for a finecontrol of matrix turnover.
Stroma cells with differentiated myofibroblast fea-tures also are found in different primary and metastaticepithelial tumors and are thought to play a central rolein tumor-associated tissue remodeling (12). These fi-broblastic cells adjacent to neoplastic cell nests, whichwere shown to express important amounts of �SMA,are normally referred to as peritumoral fibroblasts orreactive stroma. Their presence is considered to be aresponse of the host cells to inductive stimuli exertedby tumor cells, which in turn are able to participateactively in tumor progression by secretion of proteolyticenzymes, thus promoting tumor invasion and metasta-sis. Indeed, it is now generally admitted that an impor-tant proportion of such enzymes is produced by stromamyofibroblasts as a host response to tumor (13, 14).Therefore, tumor cells appear to signal neighboringfibroblasts on the one hand to differentiate to myofi-broblasts and, on the other, to secrete more MMPs.
The factor that induces the peritumoral fibroblasts toincrease MMP production has been identified as extracel-lular matrix metalloproteinase inducer (EMMPRIN)/CD147, a membrane-spanning molecule highly ex-pressed in tumor cells. Its most characterized role is theinduction of MMP production in neighboring cells (15,16) and its overexpression in vivo results in enhancedtumor growth (17). EMMPRIN also was shown tomodulate MMP expression during normal tissue re-modeling and differentiation as well as in nontumoralpathological conditions such as skin or corneal ulcer-ations (18, 19), rheumatoid arthritis (20), and lung andliver fibrosis (21, 22). As all these situations also areassociated with the presence of myofibroblasts, thepossibility that EMMPRIN may be involved in signalingfibroblasts not only to induce MMPs production butalso to change phenotype to that of myofibroblasts hasbeen addressed in this study using a corneal wound-healing model.
The cornea, with its simple cellular organizationproviding a particularly useful model for studyingwound-healing events, is a highly organized transparenttissue that needs to remain transparent to refract lightproperly. Penetrating incisions or ablation injury to thecorneal stroma stimulates a typical fibrotic repair re-sponse involving hypercellularity, expression of �SMA,
and deposition of disorganized ECM. This fibroticrepair response can be harmful to the cornea becauseit may cause opacity, and the contraction associatedwith remodeling alters corneal curvature (23). Chronicwound healing may be associated also with excessiveproteolysis and result in corneal thinning or ulcerationwith the risk of perforation (24), but the reason for thedifferent outcomes, fibrosis or thinning, followingchronic wound healing is yet to be determined.
We recently identified EMMPRIN in healthy anddiseased corneas and demonstrated its ability to induceMMP expression following epithelial-stromal interac-tion (19). In this study, we addressed the question ofwhether this MMP induction by EMMPRIN representsonly part of a broader process of fibroblast activationthat involves the differentiation of fibroblast to myofi-broblast. We therefore examined the effect ofEMMPRIN on �SMA in corneal and skin fibroblasts aswell as on the contractile potential of these cells. Therelationship between EMMPRIN and TGF� in thisdifferentiation process also was explored.
MATERIALS AND METHODS
Tissue, cells, and materials
Corneal tissues were obtained from the French Eye Bank(Banque Francaise des Yeux; Paris, France) and Belgian EyeBanque (Lieges, Belgium). Chronic ulcerated or perforatedcorneal specimens were obtained from patients undergoingpenetrating keratoplasty; use of specimen was performed inaccordance with protocols approved by the institutional ani-mal care and use committee. Tissues were flash frozen inoptimal cutting temperature compound and kept at �80°Cfor immunohistochemistry.
Telomerase-immortalized corneal fibroblasts (HTK cellline) (25) were cultured in Dulbecco modified Eagle medium(DMEM) (Invitrogen, Cergy Pontoise, France) supplementedwith 10% fetal calf serum (FCS; PAA Laboratories, LesMureaux, France).
Primary fibroblasts were obtained from skin biopsies of pa-tients undergoing breast plastic surgery, following informedconsent. Fibroblasts were propagated in DMEM supplementedwith 10% FCS. Fibroblasts were used at passages 4–6.
The antibodies: anti-EMMPRIN mAb used for immuno-blots and immunohistochemistry (anti-CD147, clone HIM6,Becton Dickinson France, Le Pont de Claix, France), anti-EMMPRIN pAb used for immunohistochemistry (anti-EMMPRIN, clone K-20, Santa Cruz Biotechnology, Tebu-bio,Le Perray en Yvelines, France), anti- �SMA (clone 1A4,Sigma-Aldrich, St. Quentin Fallavier, France), pan-TGF�blocking antibody (clone 1D11, R&D Systems, Lille, France),anti-MMP-1 mAb (clone 41–1E5, Calbiochem, VWR Interna-tional, Strasbourg, France), anti-GAPDH (clone 6C5, AppliedBiosystems, Courtabouef, France), horseradish-peroxidase[HRP]-conjugated anti-mouse and anti-rabbit antibodies(Jackson ImmunoResearch, Immunotech, Marseille, France),and anti-mouse IgG Alexa 488 and anti-goat IgG Alexa 594(Molecular Probes Inc., Eugene, OR, USA; Invitrogen).TIMP-1 pAb was kindly donated by Dr. R. Fridman (WayneState University, Detroit, MI, USA).
Other materials: TGF�1, epidermal growth factor (EGF),basic fibroblast growth factor (bFGF), and TNF-� were fromR&D Systems. (TGF�1 was used throughout this study and
1145EMMPRIN PROMOTES MYOFIBROBLAST DIFFERENTIATION

was referred to as TGF�.) Marimastat was obtained fromBritish Biotechnology (Oxford, UK). BM chemiluminescenceWestern blotting substrate (POD) and DAPI were from RocheDiagnostics (Meylan, France). Protease Inhibitor Cocktail SetV EDTA-free was from Calbiochem. Recombinant EMMPRIN(recEMMPRIN) used in the study was an affinity-purifiedFlag-tag protein consisting of the extracellular domain ofhuman EMMPRIN expressed in mammalian cells (19).EMMPRIN cDNA construct, human EMMPRIN full-lengthcDNA in pcDNA3 (pcDNA3 hEMMPRIN) (17), was kindlyprovided by Dr. Stanley Zucker (State University of New York,Stony Brook, NY, USA).
cDNA transfection
HTK corneal fibroblasts at 40–50% confluence were tran-siently transfected for 24 h with the pcDNA3 hEMMPRIN orwith pcDNA3 alone using FuGENE-6 (Roche Diagnostics) inDMEM/10% serum according to the manufacturer’s proto-col. Western blotting analysis confirmed increased EMMPRINexpression (mean 2.5�), which persisted over 3–4 days.
Small interfering RNA (siRNA) transfection
Two different EMMPRIN siRNA oligos, siEMMPRIN1 andsiEMMPRIN2 (Ambion, siRNA IDs: 10372 and 215973, re-spectively, Applied Biosystems) or scramble siRNA oligos(BLOCK-iT fluorescent oligo, Invitrogen), siScramble, (33nmol/L) were transfected into the cells using oligofectaminetransfection reagent in Opti-MEM I (Invitrogen) in theabsence of FCS and antibiotics. After 6 h incubation, an equalvolume of DMEM/20% FCS was added to the transfectionmixture, which was then cultured for a further 18 h. The cellsthen were washed and cultured in the presence of DMEM/10% FCS prior to analysis of EMMPRIN inhibition by Westernblot and further experiments. Density measurements of West-ern blot bands have shown a mean of 75% inhibition inEMMPRIN levels (4 different experiments), without signifi-cantly affecting TIMP-1 levels.
Three-dimensional collagen gel culture
Type I collagen was extracted from rat tail tendons as describedpreviously (26). Fibroblast-seeded collagen lattices were pre-pared by mixing on ice 5 ml collagen (2 mg/ml) in 0.1% aceticacid with 1 ml of 10� DMEM and 1 ml 0.1 N NaOH toneutralize the pH before adding 3 ml of cell suspension,harvested by trypsinization from subconfluent cultures, to give afinal cell density of 1.5 � 105/ml of lattice mixture. Collagengels were cast in 60 mm non-cell culture-treated dishes (4ml/gel). After polymerization (30 min at 37°C), gels weredetached and covered with 6 ml of DMEM with 1% FCS, with orwithout added TGF� (10 ng/ml) or EMMPRIN (2 �g/ml) asindicated. At each time point, the lattices were photographedand rates of gel contraction were calculated by determining theremaining surface area by computer-based analysis and ex-pressed as either percentage of initial area or percentage ofcontrol area. Data are presented graphically or in tabular formas mean � sd. Lattice area was statistically compared withStudent’s t test. Significant differences were determined atP � 0.001, P � 0.01, and P � 0.05.
For biochemical analysis on the embedded cells, gels werewashed by 30 min incubation with several changes of serum-free medium, centrifuged at 10,000 g for 2 min, and thepellets were homogenized using micropotters in radio-im-muno-precipitation assay (RIPA) buffer (50 mM Tris pH 7.5,150 mM NaCl, 5 mM EDTA, 0.1% SDS, 0.5% Nonidet P-40,0.5% sodium deoxycholate), supplemented with 1/100 Pro-
tease Inhibitor Cocktail Set V. After protein determinationusing the Pierce BCA Protein Assay kit (Perbio Science,Brebieres, France), extracts were analyzed by Western blots.
For collagen lattice populated with cDNA- or siRNA-trans-fected cells, 24 h after transfection, cells were recovered,counted, and embedded into collagen lattice at 1.5 � 105
cells/ml as described above.
Western blot analysis
Cells were lysed in RIPA buffer, incubated on ice for 5 min,and then scraped with a rubber policeman. Some The lysateswere clarified by centrifugation at 13,000 g for 5 min at 4°C,and 20 �g samples were analyzed by Laemmli SDS-PAGE(Bio-Rad, Marne la Coquette, France) on 10% gels and thentransferred to Immobilon-P PVDF membrane (Millipore,Guyancourt, France). Membranes were immunoblotted witheither anti-EMMPRIN mAb or anti-�SMA mAb overnight at4°C, or with GAPDH mAb for 1 h at room temperature,followed by 1 h incubation with HRP-conjugated anti-mouseantibody and visualized with BM chemiluminescence sub-strate. Protein loading was verified by ponceau red stainingand by comparing with the intensity of glyceraldehyde 3-phos-phate dehydrogenase (GAPDH) bands.
Northern blotting
Total RNA was obtained using RNeasy (Qiagen, Courtaboeuf,France) and analyzed by Northern hybridization (20 �g/lane) with 32P-labeled cDNA probes for EMMPRIN andGAPDH as described previously (5). The EMMPRIN probewas excised from pcDNA3 vector with EcoRI. Quantificationswere performed with a Storm 840 PhosphorImager (Amer-sham Biosciences, GE Healthcare, Orsay, France).
Confocal immunohistochemistry and cytochemistry
Cryostat sections (8 �m) were prepared from frozen corneaand were immunostained as described previously (27).Briefly, the sections were fixed in chilled acetone for 10 min,then rehydrated in PBS and incubated in blocking solution(3% BSA in PBS) for 30 min. Sections were incubated for 1 hwith either anti-EMMPRIN mAb, anti-EMMPRIN pAb, oranti-�SMA mAb, then for 30 min with conjugated affinity-purified donkey anti-mouse IgG and/or anti-goat IgG (Alexa488 and 594, respectively).
For immunocytochemistry, cells were seeded on type Icollagen (0.1 mg/ml)-coated glass slides, cultured for 2–3days in the presence of 10% FCS. The cells were then treatedin one of the following two ways: 1) the cells first were serumstarved for 48 h, after which they were treated with 2 �g/mlrecEMMPRIN for a further 48 h; 2) the cultured cells firstwere transfected with EMMPRIN siRNA for 6 h, after whichserum was added for a 18 h incubation as described above.The cells then were serum starved for 48 h and treated with 10ng/ml TGF� for a further 48 h in serum-free medium.
Slides then were washed and fixed in 4% paraformaldehyde/DMEM (v/v) and processed as above using either anti-EMMPRIN mAb, anti-EMMPRIN pAb, or anti- �SMA mAb,followed by conjugated affinity-purified donkey anti-mouseand/or anti-goat IgG (Alexa 488 and 594, respectively). Theslides were mounted and examined with a laser scanningconfocal microscope (Leica Lasertechnik, Heidelberg, Ger-many). Negative controls were prepared using the same proce-dure, but PBS was substituted for the primary antibody.
1146 Vol. 22 April 2008 HUET ET AL.The FASEB Journal

RESULTS
EMMPRIN expression is increased in thesubepithelial stroma of healing cornea
EMMPRIN is highly expressed in most cancer cells andin different normal epithelia. On the contrary, very lowlevels of EMMPRIN expression can usually be detectedin resting fibroblasts. Figure 1 confirms the low butdistinct staining of EMMPRIN in the stroma of normalA) human cornea, and B) human skin sections. How-ever, EMMPRIN expression was increased greatly in thestroma of injured human cornea with a developedulceration, as is shown by the much stronger andbroader staining of EMMPRIN in the affected areacompared to a distant nonulcerated region of the samecornea (Fig. 1C). This increase was more apparent atthe subepithelial stroma where interactions betweenepithelial and stromal cells are thought to take place.Examination of healed corneal lesions shows that thisup-regulation of EMMPRIN in the ulcerated stroma ismaintained even after complete epithelial closure (Fig.1D). Hence, EMMPRIN induction localizes with areasof stromal remodeling in the healing cornea.
TGF� regulates EMMPRIN expression in fibroblasts
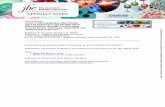
To investigate the possibility that cytokines regulateEMMPRIN expression, we examined the effect of TGF�,TNF-�, bFGF, and EGF on EMMPRIN expression incorneal fibroblast (HTK) cell culture. Figure 2 shows thatthe addition of TGF� had the greatest effect, increasingEMMPRIN expression at both the protein and RNAlevels. As illustrated (Fig. 2B), the increase of EMMPRINRNA expression is observed as early as 2 h after TGF�addition and reached a maximum at 6 h. Conversely,reducing endogenously produced TGF� by incubatingthe cells with TGF�-blocking antibodies inhibitedEMMPRIN levels (Fig. 2C), showing that EMMPRIN isconstitutively under TGF� regulation. This inhibition byTGF�-blocking antibodies was observed in both sparseand confluent cultures. Note that the EMMPRIN level wasmuch higher in sparse cultures, which are known toexpress more TGF� receptor (28) compared to confluent
cells, further supporting the notion that TGF� signalingregulates EMMPRIN production. The regulation ofEMMPRIN by TGF� also has been confirmed with pri-mary skin fibroblasts (not shown).
Differentiation of fibroblast to myofibroblast in vitrois associated with an increase in EMMPRIN levels
Because TGF� induces the differentiation of fibroblasts toa myofibroblast phenotype, we questioned the possibilitythat the up-regulation of EMMPRIN by TGF� may berelated to the differentiation process. The expression ofEMMPRIN therefore was measured in different experi-mental systems known to initiate myofibroblast differen-tiation, which were characterized by increased �SMAexpression. Treatment of fibroblasts with TGF� inducedthe expression of both EMMPRIN and �SMA (Fig. 3A) ina time-dependent manner. The coexpression ofEMMPRIN and �SMA also was observed in the presenceof serum (Fig. 3B) as well as when the cells were embed-ded in tridimensional collagen lattice (Fig. 3C), a culturesystem known also to induce myofibroblasts differentia-tion (29, 30). Time-course analysis of gel contractionreveals that both EMMPRIN and �SMA levels are highestat the point of maximal contraction. Hence, EMMPRINlevels are increased during myofibroblast differentiationinduced by either cytokines or by mechanical stress. Theseresults have been reproduced with primary skin fibro-blasts (not shown).
EMMPRIN induces �SMA production and acceleratescollagen gel contraction
To determine if EMMPRIN is an active player in thedifferentiation process, we investigated the direct effectof EMMPRIN on �SMA production and collagen gelcontraction.
Increased EMMPRIN level obtained by the transienttransfection of the corneal fibroblasts with EMMPRINcDNA caused an increase in �SMA expression and wasassociated with accelerated gel contraction, as shown inFig. 4A. Similarly, the treatment of corneal fibroblastswith exogenously added recEMMPRIN increased �SMAprotein expression and collagen gel contraction (Fig. 4B).
Figure 1. Confocal microscopy immunohistochemical staining of EMMPRIN in human normal and diseased corneas. A) Normalcornea, and B) normal skin for comparison. The staining of EMMPRIN is predominantly epithelial but some weakly stained stromalcells can be seen. C) Chronic corneal ulcers showing induction of EMMPRIN in the ulcerated region of the stroma, in particular atthe epithelio-stromal boundary. D) Chronic corneal wound after complete epithelial closure, showing persistent EMMPRINexpression just below the healed epithelium. Original: �20 (A, B); �10 (C, D). � � ulcerated area; � distance from ulceration.
1147EMMPRIN PROMOTES MYOFIBROBLAST DIFFERENTIATION
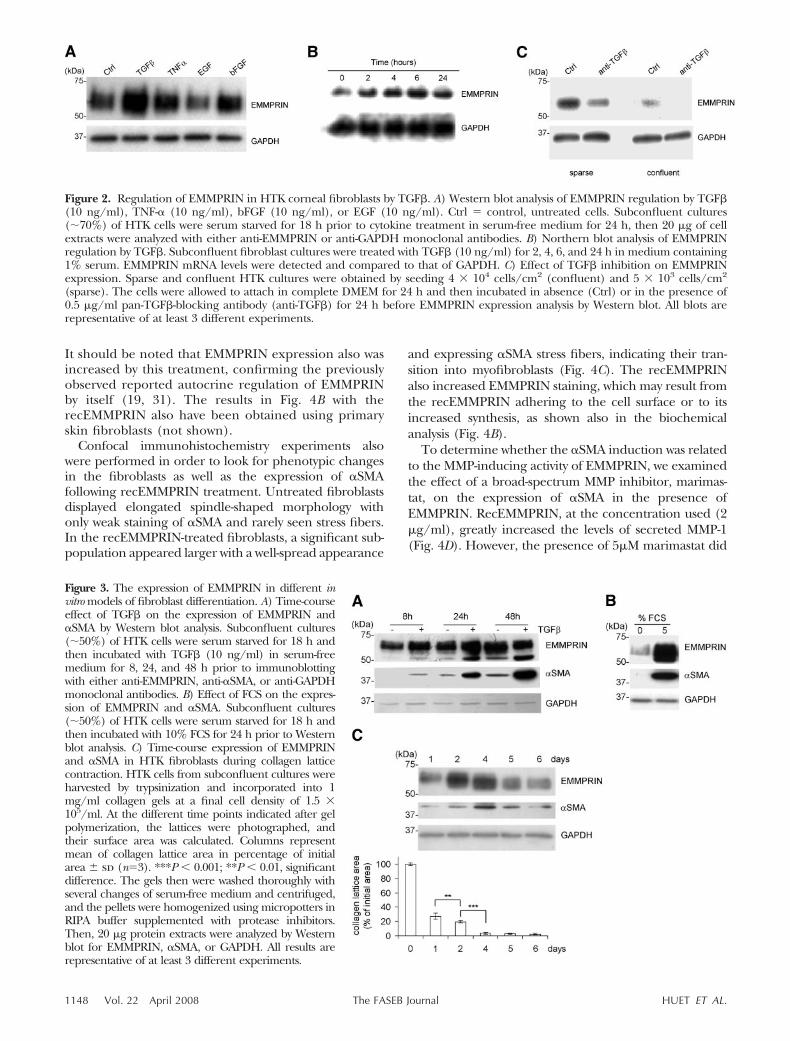
It should be noted that EMMPRIN expression also wasincreased by this treatment, confirming the previouslyobserved reported autocrine regulation of EMMPRINby itself (19, 31). The results in Fig. 4B with therecEMMPRIN also have been obtained using primaryskin fibroblasts (not shown).
Confocal immunohistochemistry experiments alsowere performed in order to look for phenotypic changesin the fibroblasts as well as the expression of �SMAfollowing recEMMPRIN treatment. Untreated fibroblastsdisplayed elongated spindle-shaped morphology withonly weak staining of �SMA and rarely seen stress fibers.In the recEMMPRIN-treated fibroblasts, a significant sub-population appeared larger with a well-spread appearance
and expressing �SMA stress fibers, indicating their tran-sition into myofibroblasts (Fig. 4C). The recEMMPRINalso increased EMMPRIN staining, which may result fromthe recEMMPRIN adhering to the cell surface or to itsincreased synthesis, as shown also in the biochemicalanalysis (Fig. 4B).
To determine whether the �SMA induction was relatedto the MMP-inducing activity of EMMPRIN, we examinedthe effect of a broad-spectrum MMP inhibitor, marimas-tat, on the expression of �SMA in the presence ofEMMPRIN. RecEMMPRIN, at the concentration used (2�g/ml), greatly increased the levels of secreted MMP-1(Fig. 4D). However, the presence of 5�M marimastat did
Figure 2. Regulation of EMMPRIN in HTK corneal fibroblasts by TGF�. A) Western blot analysis of EMMPRIN regulation by TGF�(10 ng/ml), TNF-� (10 ng/ml), bFGF (10 ng/ml), or EGF (10 ng/ml). Ctrl � control, untreated cells. Subconfluent cultures(70%) of HTK cells were serum starved for 18 h prior to cytokine treatment in serum-free medium for 24 h, then 20 �g of cellextracts were analyzed with either anti-EMMPRIN or anti-GAPDH monoclonal antibodies. B) Northern blot analysis of EMMPRINregulation by TGF�. Subconfluent fibroblast cultures were treated with TGF� (10 ng/ml) for 2, 4, 6, and 24 h in medium containing1% serum. EMMPRIN mRNA levels were detected and compared to that of GAPDH. C) Effect of TGF� inhibition on EMMPRINexpression. Sparse and confluent HTK cultures were obtained by seeding 4 � 104 cells/cm2 (confluent) and 5 � 103 cells/cm2
(sparse). The cells were allowed to attach in complete DMEM for 24 h and then incubated in absence (Ctrl) or in the presence of0.5 �g/ml pan-TGF�-blocking antibody (anti-TGF�) for 24 h before EMMPRIN expression analysis by Western blot. All blots arerepresentative of at least 3 different experiments.
Figure 3. The expression of EMMPRIN in different invitro models of fibroblast differentiation. A) Time-courseeffect of TGF� on the expression of EMMPRIN and�SMA by Western blot analysis. Subconfluent cultures(50%) of HTK cells were serum starved for 18 h andthen incubated with TGF� (10 ng/ml) in serum-freemedium for 8, 24, and 48 h prior to immunoblottingwith either anti-EMMPRIN, anti-�SMA, or anti-GAPDHmonoclonal antibodies. B) Effect of FCS on the expres-sion of EMMPRIN and �SMA. Subconfluent cultures(50%) of HTK cells were serum starved for 18 h andthen incubated with 10% FCS for 24 h prior to Westernblot analysis. C) Time-course expression of EMMPRINand �SMA in HTK fibroblasts during collagen latticecontraction. HTK cells from subconfluent cultures wereharvested by trypsinization and incorporated into 1mg/ml collagen gels at a final cell density of 1.5 �105/ml. At the different time points indicated after gelpolymerization, the lattices were photographed, andtheir surface area was calculated. Columns representmean of collagen lattice area in percentage of initialarea � sd (n�3). ***P � 0.001; **P � 0.01, significantdifference. The gels then were washed thoroughly withseveral changes of serum-free medium and centrifuged,and the pellets were homogenized using micropotters inRIPA buffer supplemented with protease inhibitors.Then, 20 �g protein extracts were analyzed by Westernblot for EMMPRIN, �SMA, or GAPDH. All results arerepresentative of at least 3 different experiments.
1148 Vol. 22 April 2008 HUET ET AL.The FASEB Journal
not inhibit �SMA induction by EMMPRIN (Fig. 4D),indicating an MMP-independent mechanism.
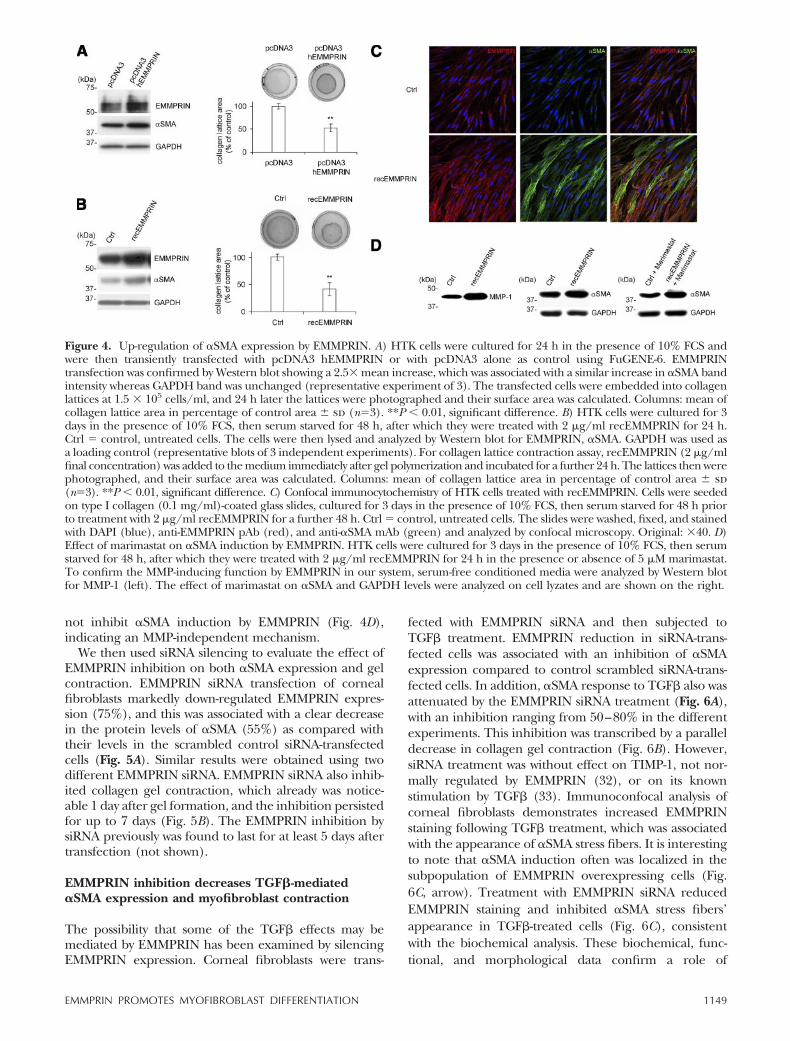
We then used siRNA silencing to evaluate the effect ofEMMPRIN inhibition on both �SMA expression and gelcontraction. EMMPRIN siRNA transfection of cornealfibroblasts markedly down-regulated EMMPRIN expres-sion (75%), and this was associated with a clear decreasein the protein levels of �SMA (55%) as compared withtheir levels in the scrambled control siRNA-transfectedcells (Fig. 5A). Similar results were obtained using twodifferent EMMPRIN siRNA. EMMPRIN siRNA also inhib-ited collagen gel contraction, which already was notice-able 1 day after gel formation, and the inhibition persistedfor up to 7 days (Fig. 5B). The EMMPRIN inhibition bysiRNA previously was found to last for at least 5 days aftertransfection (not shown).
EMMPRIN inhibition decreases TGF�-mediated�SMA expression and myofibroblast contraction
The possibility that some of the TGF� effects may bemediated by EMMPRIN has been examined by silencingEMMPRIN expression. Corneal fibroblasts were trans-
fected with EMMPRIN siRNA and then subjected toTGF� treatment. EMMPRIN reduction in siRNA-trans-fected cells was associated with an inhibition of �SMAexpression compared to control scrambled siRNA-trans-fected cells. In addition, �SMA response to TGF� also wasattenuated by the EMMPRIN siRNA treatment (Fig. 6A),with an inhibition ranging from 50–80% in the differentexperiments. This inhibition was transcribed by a paralleldecrease in collagen gel contraction (Fig. 6B). However,siRNA treatment was without effect on TIMP-1, not nor-mally regulated by EMMPRIN (32), or on its knownstimulation by TGF� (33). Immunoconfocal analysis ofcorneal fibroblasts demonstrates increased EMMPRINstaining following TGF� treatment, which was associatedwith the appearance of �SMA stress fibers. It is interestingto note that �SMA induction often was localized in thesubpopulation of EMMPRIN overexpressing cells (Fig.6C, arrow). Treatment with EMMPRIN siRNA reducedEMMPRIN staining and inhibited �SMA stress fibers’appearance in TGF�-treated cells (Fig. 6C), consistentwith the biochemical analysis. These biochemical, func-tional, and morphological data confirm a role of
Figure 4. Up-regulation of �SMA expression by EMMPRIN. A) HTK cells were cultured for 24 h in the presence of 10% FCS andwere then transiently transfected with pcDNA3 hEMMPRIN or with pcDNA3 alone as control using FuGENE-6. EMMPRINtransfection was confirmed by Western blot showing a 2.5� mean increase, which was associated with a similar increase in �SMA bandintensity whereas GAPDH band was unchanged (representative experiment of 3). The transfected cells were embedded into collagenlattices at 1.5 � 105 cells/ml, and 24 h later the lattices were photographed and their surface area was calculated. Columns: mean ofcollagen lattice area in percentage of control area � sd (n�3). **P � 0.01, significant difference. B) HTK cells were cultured for 3days in the presence of 10% FCS, then serum starved for 48 h, after which they were treated with 2 �g/ml recEMMPRIN for 24 h.Ctrl � control, untreated cells. The cells were then lysed and analyzed by Western blot for EMMPRIN, �SMA. GAPDH was used asa loading control (representative blots of 3 independent experiments). For collagen lattice contraction assay, recEMMPRIN (2 �g/mlfinal concentration) was added to the medium immediately after gel polymerization and incubated for a further 24 h. The lattices then werephotographed, and their surface area was calculated. Columns: mean of collagen lattice area in percentage of control area � sd(n�3). **P � 0.01, significant difference. C) Confocal immunocytochemistry of HTK cells treated with recEMMPRIN. Cells were seededon type I collagen (0.1 mg/ml)-coated glass slides, cultured for 3 days in the presence of 10% FCS, then serum starved for 48 h priorto treatment with 2 �g/ml recEMMPRIN for a further 48 h. Ctrl � control, untreated cells. The slides were washed, fixed, and stainedwith DAPI (blue), anti-EMMPRIN pAb (red), and anti-�SMA mAb (green) and analyzed by confocal microscopy. Original: �40. D)Effect of marimastat on �SMA induction by EMMPRIN. HTK cells were cultured for 3 days in the presence of 10% FCS, then serumstarved for 48 h, after which they were treated with 2 �g/ml recEMMPRIN for 24 h in the presence or absence of 5 �M marimastat.To confirm the MMP-inducing function by EMMPRIN in our system, serum-free conditioned media were analyzed by Western blotfor MMP-1 (left). The effect of marimastat on �SMA and GAPDH levels were analyzed on cell lyzates and are shown on the right.
1149EMMPRIN PROMOTES MYOFIBROBLAST DIFFERENTIATION
EMMPRIN in mediating, at least in part, TGF� effects onmyofibroblast differentiation.
The induction of EMMPRIN in the fibroblasts ofremodeling stroma in vivo is associated with �SMAexpression
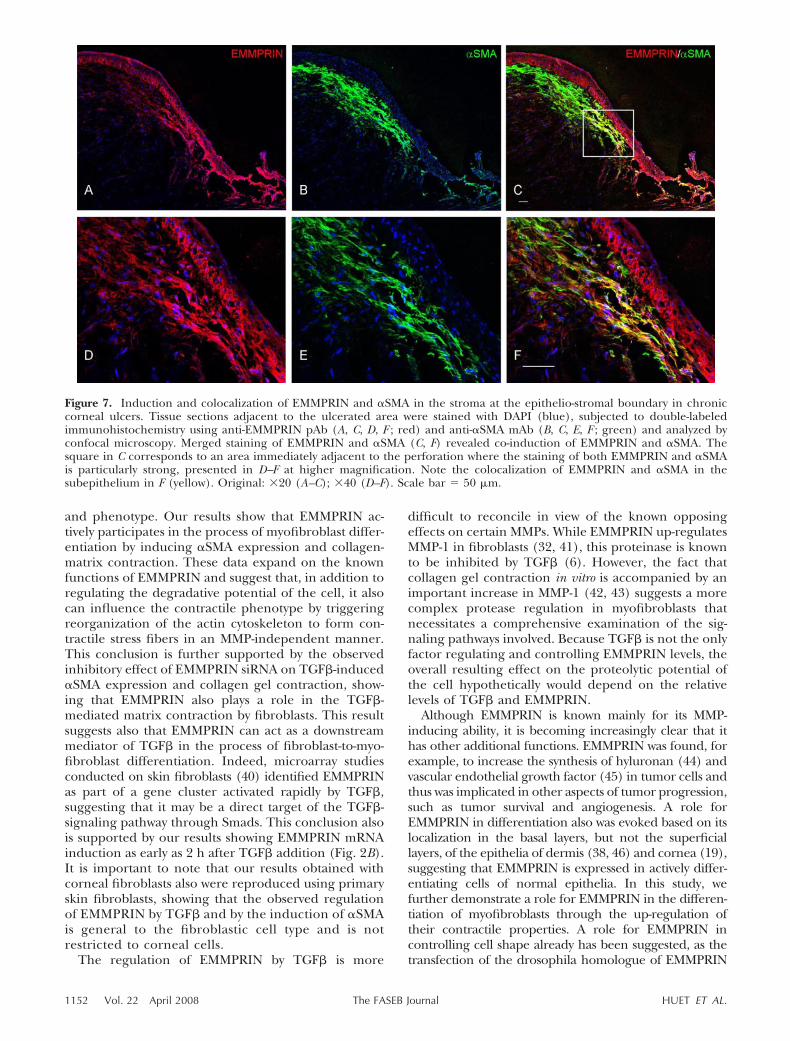
In this study we present evidence that EMMPRINinduces �SMA expression, whether EMMPRIN is exog-enously added, endogenously increased by transfection,or induced by TGF� treatment of fibroblasts. We soughtto examine whether the EMMPRIN overexpressing re-modeling stroma in vivo (Fig. 1) was associated with amyofibroblast phenotype. Double-labeled confocal mi-croscopy analysis of EMMPRIN (red) and �SMA (green)was performed on human cornea with chronic ulceration.
The staining in Fig. 7 shows that whereas most of theEMMPRIN-positive fibroblasts also expressed �SMA, amore detailed analysis reveals the presence of a subpopu-lation of fibroblasts localized in the subepithelial regionadjacent to the ulceration that express particularly highlevels of both EMMPRIN and �SMA. The presence ofthese heterogeneous populations in terms of the relativeexpression of EMMPRIN and �SMA may suggest theexistence of different induction pathways for �SMA in vivo.
DISCUSSION
Numerous studies have demonstrated a crucial role forEMMPRIN in regulating the expression of severalMMPs, mostly through the interaction of tumor cellswith neighboring fibroblasts. Our previous work hasshown that EMMPRIN also was responsible for theinduction of MMPs in corneal wound healing, follow-ing the direct interaction between corneal epithelial cellsand corneal fibroblasts (19), implicating EMMPRIN inthe MMP-mediated stromal thinning and ulceration.EMMPRIN itself is greatly increased in the stroma ofsuch pathological wounds, in particular at the level ofthe subepithelium, implicating direct interactions be-tween the epithelium and the stroma in its up-regula-tion. These epithelial-mesenchymal interactions play acritical role in development, cancer progression, andwound healing and were shown to modify the pheno-type of both epithelial cells and fibroblasts (34, 35).Known key mediators of these interactions includemembers of the TGF� family, released primarily fromthe epithelium to target stromal cells after disruption ofthe basement membrane (36). In this study, we dem-onstrate that TGF� up-regulates EMMPRIN expressionin fibroblasts and therefore is likely to contribute to theincreased EMMPRIN seen in the ulcerated stroma. Theinhibitory effect of TGF�-blocking antibody suggeststhat EMMPRIN is in fact constitutively under the regu-lation of TGF�, but its levels can be further increased inresponse to raised extracellular levels of this cytokine.This result concords with our findings that EMMPRINis up-regulated in low-density cultures, where TGF� andits receptors are known to be greatly increased (28).However, factors other than TGF� also may up-regulateEMMPRIN in this system. EGF, which was shown toregulate EMMPRIN in epithelial tumor cells (37), hadno such effect in fibroblasts, but the intervention ofother cytokines cannot be excluded. EMMPRIN itselfwas shown to be able to induce its own expression inneighboring cells (Fig. 4; 19, 31), and because it isparticularly enriched on the surface of epithelial cells(32, 38), it has the potential, following injury involvingbasement membrane rupture, to induce increased ex-pression in stromal cells. Because EMMPRIN can beshed from the cell surface (39), it also can act as adiffusible factor, reaching other cells in the vicinity.Therefore, likely both epithelial-derived EMMPRINand TGF� contribute to the increased EMMPRIN ex-pression observed in the injured stroma.
Figure 5. EMMPRIN siRNA inhibits the �SMA expression andcollagen lattice contraction. A) HTK cells were transfected usingoligofectamine transfection reagent with two different EMMPRINsiRNA, siEMMPRIN1 and siEMMPRIN2, or their correspondingscrambled control siRNA, siScramble, at 33 nmol/L concentra-tion, as described in Materials and Methods. Cells then werelysed and analyzed by Western blot for EMMPRIN, �SMA, andGAPDH (representative blots of several different experimentsare shown). B) Inhibition of collagen lattice contraction byEMMPRIN siRNA. The siRNA-transfected cells were embeddedinto collagen lattices at 1.5 � 105 cells/ml and, at different timepoints indicated after gel polymerization, the lattices werephotographed and their surface area was calculated. Represen-tative photographs of gels from 3 different experiments andtime-course quantification histogram are presented. Columns:mean of collagen lattice area in percentage of initial area � sd(n�3). **P � 0.01; *P � 0.05, significant difference.
1150 Vol. 22 April 2008 HUET ET AL.The FASEB Journal

In wound healing, TGF� is known to exert some itsactions through the differentiation of fibroblasts intomyofibroblasts, �SMA-containing contractile cells thatare essential for ECM remodeling during normal and
pathological wound healing. The fact that TGF� alsocan induce fibroblasts to up-regulate their EMMPRINexpression prompted us to examine the effect of theincreased EMMPRIN on myofibroblast differentiation
Figure 6. EMMPRIN siRNA inhibits the �SMA expression and collagen lattice contraction induced by TGF�. A) HTK cells weretransfected with EMMPRIN siRNA, siEMMPRIN1, or with the corresponding scrambled control siRNA, siScramble; serumstarved for 24 h; then subjected to TGF� treatment for 24 h in serum-free medium. EMMPRIN and �SMA protein levels wereevaluated by Western blot analysis of 20 �g cell lysates. GAPDH was used as a loading control. TIMP-1 was analyzed by Westernblots of nonconcentrated conditioned medium issue from an equal number of cells. Representative blots of several independentexperiments. B) Inhibition of TGF�-induced collagen lattice contraction by EMMPRIN silencing. The EMMPRIN siRNA,siEMMPRIN1, and scrambled control siRNA-, siScramble, transfected cells were embedded into collagen lattices at 1.5 � 105
cells/ml. Ctrl � control, untransfected cells. TGF� (10 ng/ml) was added to the medium after polymerization; 48 h later thelattices were photographed and their surface area was calculated. Columns: mean of collagen lattice area in percentage ofcontrol area � sd (n�3). **P � 0.01; *P � 0.05, significant difference. C) Confocal immunocytochemistry of EMMPRINsiRNA-transfected cells treated with TGF�. HTK cells were seeded on type I collagen (0.1 mg/ml)-coated glass slides. Thecultured cells first were transfected with EMMPRIN siRNA, siEMMPRIN1, or with the scrambled control siRNA, siScramble, asdescribed in Materials and Methods. The cells were then serum starved for 48 h and treated with 10 ng/ml TGF� for a further48 h in serum-free medium. The slides were washed; fixed; stained with DAPI (blue), anti-EMMPRIN pAb (red), and anti-�SMAmAb (green); and analyzed by confocal microscopy. Original: �40. Arrows represent cells overexpressing EMMPRIN and �SMA.
1151EMMPRIN PROMOTES MYOFIBROBLAST DIFFERENTIATION
and phenotype. Our results show that EMMPRIN ac-tively participates in the process of myofibroblast differ-entiation by inducing �SMA expression and collagen-matrix contraction. These data expand on the knownfunctions of EMMPRIN and suggest that, in addition toregulating the degradative potential of the cell, it alsocan influence the contractile phenotype by triggeringreorganization of the actin cytoskeleton to form con-tractile stress fibers in an MMP-independent manner.This conclusion is further supported by the observedinhibitory effect of EMMPRIN siRNA on TGF�-induced�SMA expression and collagen gel contraction, show-ing that EMMPRIN also plays a role in the TGF�-mediated matrix contraction by fibroblasts. This resultsuggests also that EMMPRIN can act as a downstreammediator of TGF� in the process of fibroblast-to-myo-fibroblast differentiation. Indeed, microarray studiesconducted on skin fibroblasts (40) identified EMMPRINas part of a gene cluster activated rapidly by TGF�,suggesting that it may be a direct target of the TGF�-signaling pathway through Smads. This conclusion alsois supported by our results showing EMMPRIN mRNAinduction as early as 2 h after TGF� addition (Fig. 2B).It is important to note that our results obtained withcorneal fibroblasts also were reproduced using primaryskin fibroblasts, showing that the observed regulationof EMMPRIN by TGF� and by the induction of �SMAis general to the fibroblastic cell type and is notrestricted to corneal cells.
The regulation of EMMPRIN by TGF� is more
difficult to reconcile in view of the known opposingeffects on certain MMPs. While EMMPRIN up-regulatesMMP-1 in fibroblasts (32, 41), this proteinase is knownto be inhibited by TGF� (6). However, the fact thatcollagen gel contraction in vitro is accompanied by animportant increase in MMP-1 (42, 43) suggests a morecomplex protease regulation in myofibroblasts thatnecessitates a comprehensive examination of the sig-naling pathways involved. Because TGF� is not the onlyfactor regulating and controlling EMMPRIN levels, theoverall resulting effect on the proteolytic potential ofthe cell hypothetically would depend on the relativelevels of TGF� and EMMPRIN.
Although EMMPRIN is known mainly for its MMP-inducing ability, it is becoming increasingly clear that ithas other additional functions. EMMPRIN was found, forexample, to increase the synthesis of hyluronan (44) andvascular endothelial growth factor (45) in tumor cells andthus was implicated in other aspects of tumor progression,such as tumor survival and angiogenesis. A role forEMMPRIN in differentiation also was evoked based on itslocalization in the basal layers, but not the superficiallayers, of the epithelia of dermis (38, 46) and cornea (19),suggesting that EMMPRIN is expressed in actively differ-entiating cells of normal epithelia. In this study, wefurther demonstrate a role for EMMPRIN in the differen-tiation of myofibroblasts through the up-regulation oftheir contractile properties. A role for EMMPRIN incontrolling cell shape already has been suggested, as thetransfection of the drosophila homologue of EMMPRIN
Figure 7. Induction and colocalization of EMMPRIN and �SMA in the stroma at the epithelio-stromal boundary in chroniccorneal ulcers. Tissue sections adjacent to the ulcerated area were stained with DAPI (blue), subjected to double-labeledimmunohistochemistry using anti-EMMPRIN pAb (A, C, D, F ; red) and anti-�SMA mAb (B, C, E, F ; green) and analyzed byconfocal microscopy. Merged staining of EMMPRIN and �SMA (C, F) revealed co-induction of EMMPRIN and �SMA. Thesquare in C corresponds to an area immediately adjacent to the perforation where the staining of both EMMPRIN and �SMAis particularly strong, presented in D–F at higher magnification. Note the colocalization of EMMPRIN and �SMA in thesubepithelium in F (yellow). Original: �20 (A–C); �40 (D–F). Scale bar � 50 �m.
1152 Vol. 22 April 2008 HUET ET AL.The FASEB Journal

was found to affect the intracellular architecture of differ-ent cell types in which it is expressed (47). Our resultsshow for the first time an effect of EMMPRIN on cytoskel-eton composition leading to a modification of cell shapeand tensile properties.
EMMPRIN is a transmembrane-spanning proteinthat can act as a receptor. It was identified as thesignaling receptor for extracellular cyclophilins in situ-ations related to chemotaxis and adhesion of immunecells (48). However, EMMPRIN�s receptor on fibroblastcells responsible for the stimulation of MMP produc-tion remains elusive (41, 49). It has been suggested thatEMMPRIN may serve as its own counter-receptor incancer cells via homophilic interactions, implying thatit can act also as a ligand (50). In this study, wecompared the effects of exogenously added solubleEMMPRIN lacking the transmembrane domain withthose of increased endogenous expression of the entiremolecule by cDNA transfection. In both cases, therewas an increase in �SMA expression. However, it isdifficult to speculate on the nature of EMMPRIN,whether receptor or ligand, involved in these twosituations because, when soluble EMMPRIN is added asa ligand, it induces endogenous EMMPRIN expressionin a positive autocrine feedback loop (19, 31; Fig. 4B).Thus, while the added recombinant molecule may actas a ligand, the overexpressed endogenous EMMPRINon the same cell potentially can act as a receptor.
�SMA generally has been used as a marker of fibrosisbecause �SMA-containing myofibroblasts have been ob-served in most fibrotic tissues, and TGF�, the cytokineprincipally associated with fibrosis, increases both theexpression of �SMA and that of ECM components. At thesame time, accumulated evidence indicates that the mainrole of the smooth-muscle-like features of myofibroblastsis in the force generation which results in wound contrac-tion (30). In view of our results showing that EMMPRIN,an MMP inducer, can itself induce �SMA, it is tempting tospeculate on the existence of several types of myofibro-blasts with different and even opposing effects on theECM: those with ECM synthetic phenotype present inmost fibrotic tissues and driven more by TGF�, and thosefound in ulcerated tissues having an increased degrada-tive potential due to a greater EMMPRIN effect. Indeed,�SMA-expressing myofibroblasts, expressing high levels ofmatrix-degrading proteinases such as MMP-2 and MT1-MMP, also were localized in the stroma surroundingmany cancer tissues, with the potential to promote tumorinvasion through tissue degradation (13, 14). Thus, thecellular pathways leading to the expression of �SMA maybe different from those of tissue remodeling, and theacquisition of myofibroblasts phenotype, characterized bythe expression of �SMA, may be more representative of thecontractile role of these cells. The effect of myofibroblastson matrix remodeling may then depend, among otherfactors, on the levels of TGF� and EMMPRIN, tilting thebalance toward synthesis or degradation, respectively.
The interplay and relative role of TGF� and EMMPRINin myofibroblast differentiation, �SMA expression, andmatrix remodeling in normal and pathological wound
healing is under investigation in our laboratory. Under-standing the underlying mechanisms controlling the myo-fibroblast phenotype has important implications for thecontrol of pathological wound-healing responses such asexcessive matrix contraction, scarring, or ulceration.
We thank N. Satterblad for confocal microscopy imaging.This work was supported by the Fondation de L’Avenir (studyET5-400). E.H. was supported by the Fondation des GueulesCassees and by Retina France.
REFERENCES
1. Hinz, B., Celetta, G., Tomasek, J. J., Gabbiani, G., and Chapon-nier, C. (2001) �-smooth muscle actin expression upregulatesfibroblast contractile activity. Mol. Biol. Cell 12, 2730–2741
2. Desmouliere, A., Geinoz, A., Gabbiani, F., and Gabbiani, G.(1993) Transforming growth factor-� 1 induces �-smooth mus-cle actin expression in granulation tissue myofibroblasts and inquiescent and growing cultured fibroblasts. J. Cell Biol. 122,103–111
3. Masur, S. K., Dewal, H. S., Dinh, T. T., Erenburg, I., andPetridou, S. (1996) Myofibroblasts differentiate from fibroblastswhen plated at low density. Proc. Natl. Acad. Sci. U. S. A. 93,4219–4223
4. Jester, J. V., Barry-Lane, P. A., Cavanagh, H. D., and Petroll,W. M. (1996) Induction of �-smooth muscle actin expressionand myofibroblast transformation in cultured corneal kerato-cytes. Cornea 15, 505–516
5. Verrecchia, F., and Mauviel, A. (2002) Transforming growthfactor-� signaling through the Smad pathway: role in extracel-lular matrix gene expression and regulation. J. Invest. Dermatol.118, 211–215
6. Yuan, W., and Varga, J. (2001) Transforming growth factor-�repression of matrix metalloproteinase-1 in dermal fibroblastsinvolves Smad3. J. Biol. Chem. 276, 38502–38510
7. Mauviel, A., Chung, K. Y., Agarwal, A., Tamai, K., and Uitto, J.(1996) Cell-specific induction of distinct oncogenes of the Junfamily is responsible for differential regulation of collagenasegene expression by transforming growth factor-� in fibroblastsand keratinocytes. J. Biol. Chem. 271, 10917–10923
8. Stuelten, C. H., DaCosta Byfield, S., Arany, P. R., Karpova, T. S.,Stetler-Stevenson, W. G., and Roberts, A. B. (2005) Breastcancer cells induce stromal fibroblasts to express MMP-9 viasecretion of TNF-� and TGF-�. J. Cell Sci. 118, 2143–2153
9. Philipp, K., Riedel, F., Germann, G., Hormann, K., and Sauer-bier, M. (2005) TGF-� antisense oligonucleotides reduce mRNAexpression of matrix metalloproteinases in cultured wound-healing-related cells. Int. J. Mol. Med. 15, 299–303
10. Karsdal, M. A., Larsen, L., Engsig, M. T., Lou, H., Ferreras, M.,Lochter, A., Delaisse, J. M., and Foged, N. T. (2002) Matrixmetalloproteinase-dependent activation of latent transforminggrowth factor-� controls the conversion of osteoblasts intoosteocytes by blocking osteoblast apoptosis. J. Biol. Chem. 277,44061–44067
11. Yu, Q., and Stamenkovic, I. (2000) Cell surface-localized matrixmetalloproteinase-9 proteolytically activates TGF-� and pro-motes tumor invasion and angiogenesis. Genes Dev. 14, 163–176
12. Desmouliere, A., Guyot, C., and Gabbiani, G. (2004) The stromareaction myofibroblast: a key player in the control of tumor cellbehavior. Int. J. Dev. Biol. 48, 509–517
13. Bisson, C., Blacher, S., Polette, M., Blanc, J. F., Kebers, F.,Desreux, J., Tetu, B., Rosenbaum, J., Foidart, J. M., Birembaut,P., and Noel, A. (2003) Restricted expression of membrane type1-matrix metalloproteinase by myofibroblasts adjacent to hu-man breast cancer cells. Int. J. Cancer 105, 7–13
14. Nielsen, B. S., Rank, F., Lopez, J. M., Balbin, M., Vizoso, F.,Lund, L. R., Dano, K., and Lopez-Otin, C. (2001) Collagenase-3expression in breast myofibroblasts as a molecular marker oftransition of ductal carcinoma in situ lesions to invasive ductalcarcinomas. Cancer Res. 61, 7091–7100
1153EMMPRIN PROMOTES MYOFIBROBLAST DIFFERENTIATION

15. Gabison, E. E., Hoang-Xuan, T., Mauviel, A., and Menashi, S.(2005) EMMPRIN/CD147, an MMP modulator in cancer, de-velopment and tissue repair. Biochimie 87, 361–368
16. Yan, L., Zucker, S., and Toole, B. P. (2005) Roles of themultifunctional glycoprotein, emmprin (basigin; CD147), intumour progression. Thromb. Haemost. 93, 199–204
17. Zucker, S., Hymowitz, M., Rollo, E. E., Mann, R., Conner, C. E.,Cao, J., Foda, H. D., Tompkins, D. C., and Toole, B. P. (2001)Tumorigenic potential of extracellular matrix metalloprotein-ase inducer. Am. J. Pathol. 158, 1921–1928
18. Norgauer, J., Hildenbrand, T., Idzko, M., Panther, E., Ban-demir, E., Hartmann, M., Vanscheidt, W., and Herouy, Y. (2002)Elevated expression of extracellular matrix metalloproteinaseinducer (CD147) and membrane-type matrix metalloprotein-ases in venous leg ulcers. Br. J. Dermatol. 147, 1180–1186
19. Gabison, E. E., Mourah, S., Steinfels, E., Yan, L., Hoang-Xuan,T., Watsky, M. A., De Wever, B., Calvo, F., Mauviel, A., andMenashi, S. (2005) Differential expression of extracellular ma-trix metalloproteinase inducer (CD147) in normal and ulcer-ated corneas: role in epithelio-stromal interactions and matrixmetalloproteinase induction. Am. J. Pathol. 166, 209–219
20. Konttinen, Y. T., Li, T. F., Mandelin, J., Liljestrom, M., Sorsa, T.,Santavirta, S., and Virtanen, I. (2000) Increased expression ofextracellular matrix metalloproteinase inducer in rheumatoidsynovium. Arthritis Rheum. 43, 275–280
21. Betsuyaku, T., Kadomatsu, K., Griffin, G. L., Muramatsu, T., andSenior, R. M. (2003) Increased basigin in bleomycin-inducedlung injury. Am. J. Respir. Cell Mol. Biol. 28, 600–606
22. Shackel, N. A., McGuinness, P. H., Abbott, C. A., Gorrell, M. D.,and McCaughan, G. W. (2002) Insights into the pathobiology ofhepatitis C virus-associated cirrhosis: analysis of intrahepaticdifferential gene expression. Am. J. Pathol. 160, 641–654
23. Fini, M. E., and Stramer, B. M. (2005) How the cornea heals:cornea-specific repair mechanisms affecting surgical outcomes.Cornea 24, S2–S11
24. Gabison, E. E., Chastang, P., Menashi, S., Mourah, S., Doan, S.,Oster, M., Mauviel, A., and Hoang-Xuan, T. (2003) Late cornealperforation after photorefractive keratectomy associated withtopical diclofenac: involvement of matrix metalloproteinases.Ophthalmology 110, 1626–1631
25. Jester, J. V., Huang, J., Fisher, S., Spiekerman, J., Chang, J. H.,Wright, W. E., and Shay, J. W. (2003) Myofibroblast differentiationof normal human keratocytes and hTERT, extended-life humancorneal fibroblasts. Invest. Ophthalmol. Vis. Sci. 44, 1850–1858
26. Mio, T., Adachi, Y., Romberger, D. J., Ertl, R. F., and Rennard,S. I. (1996) Regulation of fibroblast proliferation in three-dimensional collagen gel matrix. In Vitro Cell. Dev. Biol. Anim. 32,427–433
27. Lin, H. C., Chang, J. H., Jain, S., Gabison, E. E., Kure, T., Kato,T., Fukai, N., and Azar, D. T. (2001) Matrilysin cleavage ofcorneal collagen type XVIII NC1 domain and generation of a28-kDa fragment. Invest. Ophthalmol. Vis. Sci. 42, 2517–2524
28. Petridou, S., Maltseva, O., Spanakis, S., and Masur, S. K. (2000)TGF-� receptor expression and smad2 localization are cell densitydependent in fibroblasts. Invest. Ophthalmol. Vis. Sci. 41, 89–95
29. Hinz, B. (2006) Masters and servants of the force: the role ofmatrix adhesions in myofibroblast force perception and trans-mission. Eur. J. Cell Biol. 85, 175–181
30. Tomasek, J. J., Gabbiani, G., Hinz, B., Chaponnier, C., and Brown,R. A. (2002) Myofibroblasts and mechano-regulation of connectivetissue remodeling. Nat. Rev. Mol. Cell Biol. 3, 349–363
31. Tang, Y., Kesavan, P., Nakada, M. T., and Yan, L. (2004)Tumor-stroma interaction: positive feedback regulation of ex-tracellular matrix metalloproteinase inducer (EMMPRIN) ex-pression and matrix metalloproteinase-dependent generationof soluble EMMPRIN. Mol. Cancer Res. 2, 73–80
32. Toole, B. P. (2003) Emmprin (CD147), a cell surface regulatorof matrix metalloproteinase production and function. Curr. Top.Dev. Biol. 54, 371–389
33. Verrecchia, F., and Mauviel, A. (2002) Control of connective
tissue gene expression by TGF�: role of Smad proteins infibrosis. Curr. Rheumatol. Rep. 4, 143–149
34. Nakamura, T., Matsumoto, K., Kiritoshi, A., and Tano, Y. (1997)Induction of hepatocyte growth factor in fibroblasts by tumor-derived factors affects invasive growth of tumor cells: in vitroanalysis of tumor-stromal interactions. Cancer Res. 57, 3305–3313
35. Ronnov-Jessen, L., Petersen, O. W., and Bissell, M. J. (1996)Cellular changes involved in conversion of normal to malignantbreast: importance of the stromal reaction. Physiol. Rev. 76, 69–125
36. Kaartinen, V., Voncken, J. W., Shuler, C., Warburton, D., Bu, D.,Heisterkamp, N., and Groffen, J. (1995) Abnormal lung develop-ment and cleft palate in mice lacking TGF-� 3 indicates defects ofepithelial-mesenchymal interaction. Nat. Genet. 11, 415–421
37. Menashi, S., Serova, M., Ma, L., Vignot, S., Mourah, S., andCalvo, F. (2003) Regulation of extracellular matrix metallopro-teinase inducer and matrix metalloproteinase expression byamphiregulin in transformed human breast epithelial cells.Cancer Res. 63, 7575–7580
38. Chen, X., Kanekura, T., Tsuyama, S., Murata, F., and Kanzaki, T.(2001) Ultrastructural localization of basigin in normal humanepidermis. Histochem. Cell Biol. 115, 465–470
39. Sidhu, S. S., Mengistab, A. T., Tauscher, A. N., LaVail, J., andBasbaum, C. (2004) The microvesicle as a vehicle for EMMPRINin tumor-stromal interactions. Oncogene 23, 956–963
40. Verrecchia, F., Chu, M. L., and Mauviel, A. (2001) Identificationof novel TGF-�/Smad gene targets in dermal fibroblasts using acombined cDNA microarray/promoter transactivation ap-proach. J. Biol. Chem. 276, 17058–17062
41. Kataoka, H., DeCastro, R., Zucker, S., and Biswas, C. (1993)Tumor cell-derived collagenase-stimulatory factor increases ex-pression of interstitial collagenase, stromelysin, and 72-kDagelatinase. Cancer Res. 53, 3154–3158
42. Berton, A., Lorimier, S., Emonard, H., Laurent-Maquin, D.,Hornebeck, W., and Bellon, G. (2000) Contribution of theplasmin/matrix metalloproteinase cascade to the retraction ofhuman fibroblast populated collagen lattices. Mol. Cell Biol. Res.Commun. 3, 173–180
43. Langholz, O., Rockel, D., Mauch, C., Kozlowska, E., Bank, I., Krieg,T., and Eckes, B. (1995) Collagen and collagenase gene expressionin three-dimensional collagen lattices are differentially regulatedby �1 �1 and �2 �1 integrins. J. Cell Biol. 131, 1903–1915
44. Marieb, E. A., Zoltan-Jones, A., Li, R., Misra, S., Ghatak, S., Cao,J., Zucker, S., and Toole, B. P. (2004) Emmprin promotesanchorage-independent growth in human mammary carcinomacells by stimulating hyaluronan production. Cancer Res. 64,1229–1232
45. Tang, Y., Nakada, M. T., Kesavan, P., McCabe, F., Millar, H.,Rafferty, P., Bugelski, P., and Yan, L. (2005) Extracellular matrixmetalloproteinase inducer stimulates tumor angiogenesis byelevating vascular endothelial cell growth factor and matrixmetalloproteinases. Cancer Res. 65, 3193–3199
46. DeCastro, R., Zhang, Y., Guo, H., Kataoka, H., Gordon, M. K.,Toole, B., and Biswas, G. (1996) Human keratinocytes expressEMMPRIN, an extracellular matrix metalloproteinase inducer.J. Invest. Dermatol. 106, 1260–1265
47. Curtin, K. D., Meinertzhagen, I. A., and Wyman, R. J. (2005)Basigin (EMMPRIN/CD147) interacts with integrin to affectcellular architecture. J. Cell Sci. 118, 2649–2660
48. Yurchenko, V., Constant, S., and Bukrinsky, M. (2006) Dealingwith the family: CD147 interactions with cyclophilins. Immunol-ogy 117, 301–309
49. Guo, H., Zucker, S., Gordon, M. K., Toole, B. P., and Biswas, C.(1997) Stimulation of matrix metalloproteinase production byrecombinant extracellular matrix metalloproteinase inducer fromtransfected Chinese hamster ovary cells. J. Biol. Chem. 272, 24–27
50. Sun, J., and Hemler, M. E. (2001) Regulation of MMP-1 andMMP-2 production through CD147/extracellular matrix metal-loproteinase inducer interactions. Cancer Res. 61, 2276–2281
Received for publication April 30, 2007.Accepted for publication September 27, 2007.
1154 Vol. 22 April 2008 HUET ET AL.The FASEB Journal
Copyright © 2022 FDOKUMEN