
2 A Study of Chinese Cirrospilus Westwood (Hymenoptera Eulophidae)

Australian Journal of Entomology
(2004)
43
, 101–113
Blackwell Science, LtdOxford, UKAENAustralian Journal of Entomology1326-67562004 Australian Entomological SocietyMarch 2004432101113Original ArticleBiology of Leptocybe invasaZ Mendel
et al.
*Author to whom correspondence should be addressed (email:[email protected]).
Taxonomy and biology of
Leptocybe invasa
gen. & sp. n. (Hymenoptera: Eulophidae), an invasive gall inducer on
Eucalyptus
Zvi Mendel,
1
* Alexey Protasov,
1
Nicole Fisher
2
and John La Salle
2
1
Department of Entomology, Agriculture Research Organisation, The Volcani Center, Bet Dagan, 50250, Israel.
2
CSIRO Entomology, GPO Box 1700, Canberra, ACT 2601, Australia.
Abstract
Leptocybe invasa
Fisher & LaSalle, a new genus and species of Eulophidae, displays thelytokousreproduction, forming typical bump-shaped galls on the leaf midribs, petioles and stems of new growthof several
Eucalyptus
species. Presently, this wasp occurs in large areas in the Middle East, theMediterranean and Africa, and is a serious pest in young plantations. Heavy galling prevents furtherdevelopment of the infested growth.
Leptocybe invasa
is only known from females. Mean length of agall containing a single wasp is 2.1 mm, leaves of intensively growing trees may carry over 50 gallsper leaf. Mean development time from oviposition to emergence is 132.6 d in room temperature. InIsrael the wasp produces two or three overlapping generations annually. Mean survival time for waspsfed with honey and water is 6.5 days. Ten species were found to be suitable hosts in Israel:
E. botryoides
,
E. bridgesiana E. camaldulensis
,
E
.
globulus
.
E
.
gunii
.
E. grandis
,
E. robusta
,
E. saligna
,
E. tereticornis
, and
E. viminalis
.
Key words
biology, Eucalyptus, Eulophidae, invasive species, gall,
Leptocybe
, pest, Tetrastichinae.
INTRODUCTION
Eucalyptus
trees are the backbone of afforested areas in thelow-altitude arid and semiarid lands of the Middle East andNorth Africa.
Eucalyptus
plantations are a major source oftimber, firewood and honeybee foraging, and are used as rec-reation areas, as shelterbelts from drifting sands, and as wind-breaks surrounding cultivated and residential areas. A new pestof
Eucalyptus
was found in the Middle East and the Mediter-ranean region in 2000. It causes severe injury to young foliageof red river gum,
Eucalyptus camaldulensis
, by inducing galls,mainly on rapidly growing shoots. The wasps that emergedfrom isolated galls belong to the Eulophidae (Hymenoptera)and were recognised as an undescribed genus and species. Themassive presence of galls in young tree plantations and nurs-eries drew attention not only to the severe damage inflicted oninfested trees but also to the potential for causing seriouseconomic damage to the
Eucalyptus
forests in the area. Ourstudy has focused on
E. camaldulensis
. This species is one ofthe best known eucalyptus trees outside Australia and econom-ically is the most important hardwood species of the dry low-land areas in the entire Mediterranean and Middle East regions(FAO 1979). Planting of
E. camaldulensis
was stopped inIsrael because of extensive attacks by this wasp.
Until recent years, eucalypt trees were considered virtuallyfree of serious insect pests in the Middle East and the
Mediterranean. The eucalypt borer
Phoracantha semipunctata
(Fabricius) (Coleoptera: Cerambycidae), the sole introductionto those areas, is a minor pest attacking mainly drought-weakened trees (Mendel 1985). However, eucalypt trees intheir new habitats have been under assault from a constantstream of specific phytophagous insect pests originating fromtheir homeland (e.g., Withers 2001). Five species of gall-making wasps have established themselves on eucalypts out-side Australia:
Quadrastichodella nova
Girault (Flock 1957;Timberlake 1957; both as
Flockiella eucalypti
Timberlake),
Epichrysocharis burwelli
Schauff (Schauff & Garrison2000),
Ophelimus eucalypti
(Gahan) (Bain 1977; Viggiani &Nicotina 2001),
Aprostocetus
sp. (Beardsley & Perreira 2000)(Eulophidae), and
Nambouria xanthops
Berry & Withers(2002) (Pteromalidae). It is interesting that several of theseinvasions have occurred since 1990.
Leptocybe invasa
General& sp. n. is also believed to have originated in Australia, sim-ilarly to the above gall wasps.
Our study presents the taxonomy of
Leptocybe invasa
, aswell as basic aspects of its biology, with an emphasis ongall development, its seasonal history when attacking
E. camaldulensis,
and its plant-host specificity.
MATERIALS AND METHODS
Taxonomy
A combined diagnosis and description of this new taxon arepresented to validate both the genus and species namebecause, without additional species, it is not easy to assess the

102
Z Mendel
et al.
importance of the characters at the generic or at the specieslevel.
Terminology used in this paper is taken from Gibson(1997). Acronyms used in the text are as follows. ANIC,Australian National Insect Collection, CSIRO, Canberra,Australia; BMNH, The Natural History Museum, London,UK; QMB, Queensland Museum, Brisbane, Australia; USNM,United States National Museum of Natural History,Washington, DC, USA.
Wasp-rearing procedure
Biological studies were carried out in Israel between 2001 and2003, in the Volcani Center at Bet Dagan, The major field workwas conducted in Bet Dagan (Coastal Plain) and Nir David(Bet Shean Valley). Small branches with mature galls werecollected daily from
E. camaldulensis
stands and placed insmall, ventilated glass cages. The emerging wasps were exam-ined to determine the presence of both sexes, the gall-waspparasitoids and other arthropod associates. Soon after theiremergence, wasps were dissected and the developmental con-dition of the ovaries was determined. Observations were madeon wasps in the field, in a greenhouse and in Petri dishes inthe laboratory. Notes were taken on ovipositional behaviourand the timing of this activity.
Determination of gall growth stages as related to wasp development
The growth stages of the gall were determined by consideringthe steps of its development after wasps were introduced withintensively growing 2-year-old
E
.
camaldulensis
trees
.
Thewasp and trees were placed in plexiglas cages inside venti-lated greenhouse in Bet Dagan (Central Coastal Plain). Asimilar procedure was applied in 2- to 4-year-old-plantationsof this tree species at Nir David (Bet Shean Valley), and inprovenance trial plots with 4- to 10 years-old trees at BetDagan.
Descriptions and illustrations were prepared on the basisof microscopic examination of plant growth material and gallstaken from several different saplings and trees. We determinedfive stages of gall growth as well as the relationship betweenwasp development and gall growth stages. Mature galls fre-quently included several chambers, each containing a singlewasp. Galls were removed from the leaf surface at the surfacelevel of the leaf blade, from the petiole and from young shoots.Galls were weighed a few minutes after the removal, and theirlengths measured. They were dissected and the fresh mass oflarvae and pupae was determined.
Seasonal development
Seasonal development of wasps in
E. camaldulensis
standswas studied at Nir David and Bet Dagan, as well as in venti-lated greenhouses on the latter site. Development of galls wasmonitored on five artificially infested 1-year-old trees. Eachtree was exposed to 10 wasps and examined every third day.
The number of days between oviposition and the peak of eachgall stage was determined until the emergence of wasps wascomplete.
In June 2002, four cohorts of galls were selected, each ata different gall stage, from 1 to 4. Each cohort occurred on asingle branch with 8–10 twigs carrying between 50 and 70galls per cohort. Data on stage 1 galls refer to their mean ageper leaf. The same branches were used for each census. Thedevelopment of the galls was monitored at 10–30 d intervalsuntil the emergence of the wasps was complete.
The progress of gall stages through the winter was exam-ined in Nir David. We selected 4-year-old trees that carriedtwo distinct groups of gall stages in mid-October 2002. Onfour trees most of the galls observed were in stages 3–4, andon six trees they were in stage 1. Galls were monitored oncea month until May 2003, when emergence of the adult waspswas complete. On each tree, the same 10 small branches car-rying 10–15 leaves were marked and monitored. During thesemonthly samplings, the appearance of new foliage, and ofoviposition spots on new growth was recorded.
The information on the average maximum and minimumtemperatures came from the Israeli Meteorological Center atBet Dagan.
Adult-wasp longevity
Adult longevity was determined by means of six food treat-ments: (1) water; (2) young fresh foliage of
E. camaldulensis
;(3) honey solution (distilled water
+
honey, 1:1); (4) fresh
E. camaldulensis
flowers; (5) combination of treatments 2
+
3;and (6) no food. Wasps were placed in Petri dishes (95 mmdiameter) fitted with a filter-paper disk. The water or the honeysolutions were sprayed along a narrow strip on the cover ofeach Petri dish, and the food supply was renewed daily. Waspsbeing tested were collected on the day of emergence, removedwith a fine brush and placed in a Petri dish. Each treatmentcomprised three replicates, each of 10 wasps. The mortality ofthe wasps was determined daily. The test was conducted at25
∞
C and 80% relative humidity.Survival data on longevity of adult wasps were analysed
with the Life Test Procedure (SAS Institute Package 2002).Empirical survival distributions were fitted to the various feedtreatments. The homogeneity of the survival distributions wastested with the Wilcoxon test (log-rank statistics).
Host range
The host plant range was determined by means of severalsurveys carried out in 2001 and 2002. We focused mainly onpreviously established experimental plots and agroforestrystands planted with various species. We examined nurserystocks of 12- to 18-month-old trees at three main nurseries ofthe Forest Department of the Land Development Authoritiesin Israel. In all cases, shoots with typical mature galls weretaken to the laboratory and the emerged wasps identified.
Host suitability was tested in the laboratory by exposingseedlings of 36 species to
L. invasa
and observing whether gall

Biology of
Leptocybe invasa
103
formation occurred. In subsequent tests, three unsuitable spe-cies,
E. erythrocorys, E. gomphocephala,
and
E. occidentalis
,were tested more rigorously for host suitability and preference.Five saplings of each species were tested. Each was placed ina ventilated glass cage and was exposed to 10 adult wasps for24 h. The presence of eggs in the leaves, the development ofmature galls, and wasp emergence was examined.
Eucalyptuscamaldulensis
, the preferred host, was used as the control.
TAXONOMY
Leptocybe
Fisher & La Salle, gen. n.
Type species.
Leptocybe invasa
Fisher & La Salle, sp. n.
Etymology.
From the Greek
leptos
meaning fine, weak, thinand
kybe
meaning head; together signifying the weak area onthe head around the ocellar triangle. Gender feminine.
Leptocybe invasa
Fisher & La Salle, sp. n. (Figs 1–11)
Types.
Israel.
Holotype female, Bet Shean, 20.iv.2002, Z.Mendel, ex galls on
E. camaldulensis,
ANIC database no.32 014114 (ANIC). Paratypes: 118 females. 51 females, samedata as holotype (36 ANIC, 5 BMNH, 5 QMB, 5 USNM).
Syria.
67 females, Tartous, Sifsafa & Safita, 2003, W.Almatni,ex.
E. camaldulensis
, ANIC database no. 32 014060 (52ANIC, 5 BMNH, 5 QMB, 5 USNM).
Other material examined (not types).
Algeria
.
25 females,Pépinière CHLEF, x.2002, ex galls on
E. camaldulensis
, A.Guerroudj, ANIC database no. 32 014061 (ANIC).
Kenya.
57females, Nyanza Prov., Kisuma, G.Hailu, ex galls on leaves of
E. saligna
,
E. camaldulensis,
ANIC database no. 32 012679(ANIC).
Spain.
10 females, Cádiz, Estella del Marqués, 3.ix.2003, I. Sánchez, ex galls on
E. camaldulensis
(ANIC).
Diagnosis.
Head weak, with distinct groove and weakenedarea around ocellar triangle. Propodeum with a raised lobe ofthe callus that partially overhangs the outer rim of the spiracle;spiracular depression open to anterior margin of propodeum.Two longest cercal setae subequal in length, and straight oronly slightly curved. Postmarginal vein short, less than 0.25length of stigmal vein. Mesoscutum without median line, andwith 2–3 small adnotaular setae. Malar sulcus distinctlycurved. Dorsellum long, medially as long as propodeum.
Female.
Length 1.1–1.4 mm. Head and body brown withslight to distinct blue to green metallic shine; mouth marginlight brown to yellow. Fore coxa yellow, middle and hindcoxae the same colour with the body; legs and tarsi yellow,last tarsal segment brown apically. Antenna with scape yellow(darkened apically); funicle and club (brown to light brown).Wings hyaline or very faintly and evenly infumated, veinslight brown.
Head
(Figs 4–6). Generally strongly collapsing,particularly along weakened areas associated with a deep sul-cus around the ocellar triangle. POL distinctly longer thanOOL. Antennal torulus inserted about halfway between cly-peal margin and median ocellus, and above level of ventral eye
margin. Scrobal cavity with median line. Head weak, withdistinct groove and weakened area around ocellar triangle;frontofacial suture deep, forming anterior part of groovearound ocellar triangle. Malar sulcus distinctly curved. Genarelatively large and rounded behind malar sulcus. Clypeusweakly bilobed.
Antenna
(Fig. 3) with 4 anelli; 3 funicularsegments; 3 club segments. Scape slightly expanded ventrally.Pedicel long, over half the length of the scape. Funicularsegments all roughly quadrate.
Mesosoma
(Figs 6, 8, 9–11).Pronotum short. Midlobe of mesoscutum without median line,with 2–3 weak, short adnotaular setae at lateral margin. Scutel-lum with submedian and sublateral lines. Dorsellum somewhatelongate, medially as long as propodeum. Propodeum withoutmedian lines or lateral carinae or plicae. Propodeum with araised lobe of the callus that partially overhangs the outer rimof the spiracle; spiracular depression open to anterior marginof propodeum.
Forewing
(Fig. 2). Submarginal vein generallywith 3–4 dorsal setae, but may have 2 or 5. Postmarginal veinshort, less than 0.25 length of stigmal vein. Veins brown, witha faint hyaline area between parastigma and marginal vein.Basal cell without setae; basal vein usually with 1 seta. Specu-lum small; cubital row of setae not extending to basal vein(speculum open behind).
Gaster
(Fig. 7) short, ovate. Hypopy-gium extending just over half the length of the gaster. Twolongest cercal setae subequal in length and straight or onlyslightly curved. Ovipositor sheaths short, not reaching apex ofabdomen.
Male.
Unknown.
Discussion.
Bou ek (1988) provided a generic key toAustralasian Tetrastichinae. In this key
Leptocybe
would keyto
Aprostocetus
Westwood; either at couplet 112 (for thosespecimens with 3 or more setae on the dorsal surface of thesubmarginal vein), or at couplet 115 (for those specimens withtwo setae on the dorsal surface of the submarginal vein). Itdiffers from
Aprostocetus
by several characters, but these char-acters are in common with
Baryscapus
Förster (see below).
c
Fig. 1.
Leptocybe invasa
female, habitus.

104
Z Mendel
et al.
Figs 2–7.
Leptocybe invasa
female: (
2
) forewing; (
3
) antenna; (
4
) head, frontal view; (
5
) head, dorsal view; (
6
) head andmesosoma, lateral view; (
7
) gaster, lateral view.

Biology of
Leptocybe invasa
105
Leptocybe
is unique among the Tetrastichinae in having weak-ened areas on the vertex behind the ocelli. Other keys to generaof Tetrastichinae exist for Europe (Graham 1987; Graham1991) and North America (La Salle 1994; Schauff
et al
. 1997).
Leptocybe
is similar to
Baryscapus
. Characters which theyshare include: malar sulcus strongly curved; mesosternumconvex in front of trochantinal lobe, and without a distinctprecoxal suture; two longest cercal setae subequal in length,and straight or only slightly curved. However, Leptocybe dif-fers from Baryscapus as follows: propodeum with a raisedlobe of the callus that partially overhangs the outer rim of thespiracle; spiracular depression open to anterior margin of pro-podeum (in Baryscapus the propodeal spiracle has its entire
rim exposed); and the setae on the mesoscutum consist of asingle row of 2–3 adnotaular short, weak setae (Baryscapus isvariable in this character, but often has more than a single rowof adnotaular setae).
Leptocybe is also similar to Oncastichus La Salle. Oncas-tichus is another Australian gall inducer that has invadedCalifornia (Headrick et al. 1995). It does not attack Eucalyp-tus, but does attack another myrtaceous plant Chamelauciumuncinatum. Both these genera have weakened areas andgrooves on the vertex (although they are structurally different),and lack a median line on the mesoscutum. Leptocybe differsfrom Oncastichus in having the postmarginal vein less than0.25 the length of the stigmal vein (0.6–1.0 in Oncastichus);
Figs 8–11. Leptocybe invasa female: (8) mesosoma; (9) head and mesoscutum; (10) mesoscutum; (11) scutellum, dorsellumand propodeum.

106 Z Mendel et al.
and in Oncastichus the scape is long and slightly exceeds theupper margin of the vertex (scape normal in Leptocybe, andnot reaching upper margin of vertex).Included species. At present there is a single included species,L. invasa.Distribution. The Australian distribution for this species isstill unknown. This wasp is presently known from Algeria,Iran, Israel, Italy, Jordan, Kenya, Morocco, Spain, Syria,Turkey and Uganda.
BIOLOGY OF LEPTOCYBE INVASA
Adult behaviour and oviposition
Interestingly, among the several thousands of individual waspsthat emerged from E. camaldulensis, and smaller numbers col-lected from other Eucalyptus species, only female wasps werefound. Leptocybe invasa was the sole arthropod species thatemerged from these galls. Dissection of newly emergingwasps showed strict proovigenesis. When exposed to newgrowth, wasps insert their eggs in the epidermis of the upperside of newly developed leaves, 0.5–5 cm in length, on bothsides of the midrib, in the petiole of such leaves and in theparenchyma tissue of twigs. The attack takes place within the1–2 weeks of the bud break out. The eggs are laid at a distanceof 0.3–0.5 mm from each other and always in a lined group.Under laboratory conditions, oviposition may take place inPetri dishes or in small twigs covered by small polyethylenebags. Outdoors in the summer, wasps were observed on younggrowth during the morning and the late afternoon. Indoors,wasps were ready to attack the leaves at any time during theday. In Bet Shean Valley, where L. invasa has reached epi-demic levels, juvenile shoots were often killed due to eggoverloading. A day after oviposition a white substance, mostprobably gum coming out of the wound caused by the ovipos-itor, blocked the opening of the injury (Fig. 12).
Gall development
Five stages of gall development on E. camaldulensis treeswere identified. Stage 1 begins 1–2 weeks after oviposition,with the first symptoms of cork tissue appearing at the egginsertion spot. This stage is characterised by a small changein the morphology of the attacked tissue, the cork scarbecomes bigger and the section of the midrib that carries theeggs often changes its colour from green to pink (Fig. 13).Towards the end of the stage the galls are easily recognisedby their spherical appearance, their colour is glossy green, andeach of the galls can easily be separated from the others(Fig. 14). Stage 2 is characterised by development of thetypical bump shape and the galls reach their maximum size(width = 2.7 ± 0.5 mm). Stage 3 is characterised by the fadingof the green colour on the surface that tends to change to pinkwhile retaining its typical gloss. Stage 4 is characterised bythe loss of the glossiness of the gall surface, with colourchanges to light or dark red according to whether the galls arepresent on the leaves or on the stem. Stage 5 was recognised
as soon as emergence holes of the wasps were noticed, theircolour was light brown when displayed on the leaf and red-brown on the stem (Figs 15,16).
Determination of the five stages of gall growth enabled usto follow the development of galls as compared with the lifehistory of the wasp. The gall developed in parallel with thetissue of its producing plant. Stage 1, which probably startedwith the hatching of the larvae, covers the phases of hypoplasyand probably the early hyperplasy development of the gall(Rohfriotsch & Shorthouse 1982). During these phases, theleaves reached their full size, but their surface morphologyunderwent only small changes that could be regarded as exter-nal symptoms; the cork scars caused by oviposition increasedin width, and the section of the midrib that carried the eggschanged its colour from green to pink. At the end of Stage 2both the leaf and the gall reached their final dimensions. Therapid changes in the gall size suggest that this stage is charac-terised by the late hyperplasy and metaplasy phases of a typ-ical insect galls, i.e., a rapid overgrowth of the cells and thechange into typical gall tissue (see Abrahamson & Weis 1987).During this stage the gall, usually one ‘unit’ in a chain ofseveral, assumed its typical-bump shape and attained its max-imum size. The length of a gall of a single unit was about2.1 mm; that of one containing two ‘fused gall units’ (twowasps) was about 3.6 mm. There were no significant differ-ences between the length of a single gall or of a two-unit gall,whether they developed on the midrib, the petiole or the stem(Fig. 21). The number of wasps per leaf, contained in one orseveral multiunit galls, was highly variable, ranging from oneto 65; most frequently 3–6 for each leaf (blade and/or petiole).Leaves of intensively growing trees may carry much higherdensities (46.0 ± 14.3 wasps per leaf, n = 10). The mature gallson the stems were mostly one- or two-unit galls.
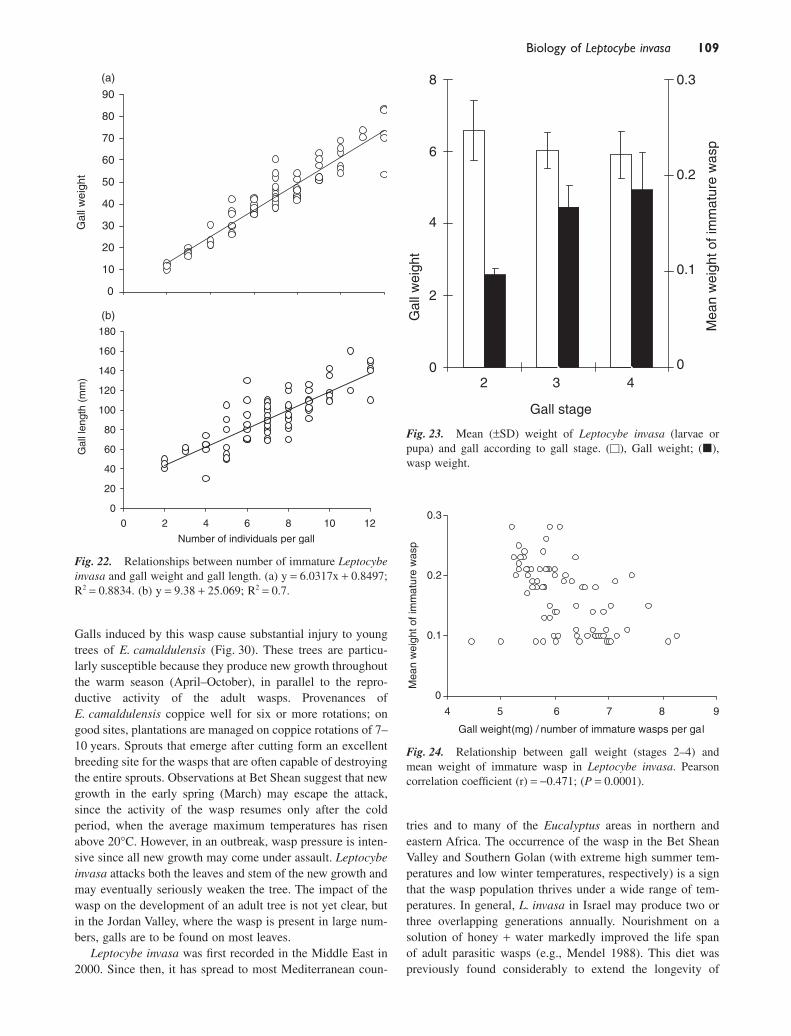
A high correlation was found between the number of waspsdeveloped in a gall, and its length and weight (Fig. 22).Whereas the gall weight did not change much during its devel-opment between stages 2 and 4, wasp larvae increased inweight between stages 2 and 3, and not at all between stages3 and 4 (Fig. 23). While mainly small larvae were found ingalls of stage 2 (Fig. 17), those in stage 3 galls were mainlymature larvae and prepupae (Figs 18,19). Individuals found instage 4 galls were mainly pupae and preadults; while adultsemerged from the stage 5 galls (Fig. 20). These results furtherconfirm our assumption that the fast development of the waspoccurs during the rapid growth of the gall. The inverse rela-tionship between the wasp density per gall and wasp weight(Fig. 24) suggests that, during stage 3, the larvae continue tofeed on the gall tissue.
Seasonal development
Oviposition was observed throughout the warm season (Aprilto October). On artificially infested trees in the greenhouse,development time from oviposition to wasp emergence was132.6 ± 8.1 d. Stage 1 lasted longest and accounted for about40% of the total development time. Stages 2 and 3 were theshortest (Fig. 25). During the warm months, all development

Biology of Leptocybe invasa 107
stages of the wasps were detected (Fig. 26). At Bet Dagan, thedevelopment time from oviposition to emergence was about4.5 months (cohort 1, Fig. 26) and the time between occur-rence of stage 4 and emergence was about 4 weeks for allcohorts. As observed at Nir David, development during thewinter was slow; the development of the later stage galls took4 months. Wasps that were only in stage 1 by early winter(October 2002) developed poorly, and all died before April2002 (Fig. 27). Wasps that inhabited galls at stages 3 or 4 inthe early winter (2002) started to emerge in April and theiremergence was complete by May (2003) (Fig. 27). Early inwinter, these individuals were already at the stage of maturelarvae or pupae and therefore could resume their developmentin the spring. They probably did not need to rely on thenutritive cell layers to complete their development. Leaves thatcarried mature galls in the autumn were shed during the win-ter. New spring growth of E. camaldulensis at Nir Davidappeared in February and developed on most branches exam-ined in March (Fig. 28). However, wasps resumed ovipositiononly in April, while in May the typical signs of ovipositionwere observed on all inspected twigs (Fig. 28). The timing of
these events closely matched that of the emergence of the over-wintering wasp population.
Adult longevity
The survival distribution of adult wasps differed significantlyamong treatments (Fig. 29). Wasps fed with honey and watersolution had the longest mean life span (±SE) (6.5 ± 0.2 d),which was significantly longer than those supplied with theflowers (4.7 ± 0.3 d), water (3.7 ± 0.2 d) or fresh young foli-age (3.5 ± 0.2 d). The life span of wasps on young foliage andwater treatments was significantly longer than that of those onthe ‘no food’ treatment (3.0 ± 0.2 d), but significantly shorterthan that obtained with flowers. Adding young foliage to thehoney plus water treatment did not significantly change thelongevity from that of the wasps fed on honey and water alone(6.0 ± 0.3 d).
Host specificity
We tested 36 species, belonging to 3 subgenera, 10 sections,and 21 series of Eucalyptus (classification following Brooker
Figs 12–16. Gall formation in Leptocybe invasa: (12) leaf 3 days after oviposition showing white secretions from the ovipositionwounds; (13) leaf after 3 weeks, showing early stages of gall formation; (14) gall after 6–7 weeks; (15–16) mature galls, showingexit holes.

108 Z Mendel et al.
2000). The following 10 species (in alphabetical order) werefound to be suitable hosts: Eucalyptus botryoides, E. bridge-siana E. camaldulensis, E. globulus. E. gunii. E. grandis,E. robusta, E. saligna, E. tereticornis, and E. viminalis.
These hosts belong to only three sections of the genus(Exsertaria, Latoangulata, Maidenaria) and no species of anyof these sections examined was found to be rejected for thedevelopment of L. invasa (Table 1). Saplings and adult trees(estimated at 1 years old) of two hybrids were examined,E. grandis ¥ E. camaldulensis and E. torquata ¥ E. woodwar-dii, known as E. torwood. Leptocybe invasa successfullydeveloped on the former and not on the latter, whose parentspecies are rejected host plants.
When L. invasa was offered five saplings of E. erythrocorys(as well as in those of E. camaldulensis, the control species),oviposition occurred. None of the five saplings of E. gompho-cephala or of E. occidentalis was attacked. Saplings ofE. erythrocorys developed similar injury symptoms to thoseexhibited by E. camaldulensis during the first days after ovi-position. However, no further development of the gall wasobserved, in contrast to gall maturation on E. camaldulensis.
DISCUSSION
Our findings suggest that L. invasa is a stenophagous feederdeveloping on eucalypt species in three sections of the genus.
Figs 17–20. Development of Leptocybe invasa: (17) gall cut open to show 10-week old L. invasa larvae; (18) gall cut open toshow a mature L. invasa larva; (19) gall cut open to show mature L. invasa pupae; (20) adult L. invasa emerging from gall.
Fig. 21. Average (±SD) gall length of Leptocybe invasa devel-oped on Eucalyptus camaldulensis as related to plant tissue andnumber of immature (fully grown larvae or pupae) wasps. (�), Asingle wasp; (�), two wasps.
0
1
3
2
4
5
Blade Petiole stem
Gall producing tissue
Gal
l le
ngth
Biology of Leptocybe invasa 109
Galls induced by this wasp cause substantial injury to youngtrees of E. camaldulensis (Fig. 30). These trees are particu-larly susceptible because they produce new growth throughoutthe warm season (April–October), in parallel to the repro-ductive activity of the adult wasps. Provenances ofE. camaldulensis coppice well for six or more rotations; ongood sites, plantations are managed on coppice rotations of 7–10 years. Sprouts that emerge after cutting form an excellentbreeding site for the wasps that are often capable of destroyingthe entire sprouts. Observations at Bet Shean suggest that newgrowth in the early spring (March) may escape the attack,since the activity of the wasp resumes only after the coldperiod, when the average maximum temperatures has risenabove 20∞C. However, in an outbreak, wasp pressure is inten-sive since all new growth may come under assault. Leptocybeinvasa attacks both the leaves and stem of the new growth andmay eventually seriously weaken the tree. The impact of thewasp on the development of an adult tree is not yet clear, butin the Jordan Valley, where the wasp is present in large num-bers, galls are to be found on most leaves.
Leptocybe invasa was first recorded in the Middle East in2000. Since then, it has spread to most Mediterranean coun-
tries and to many of the Eucalyptus areas in northern andeastern Africa. The occurrence of the wasp in the Bet SheanValley and Southern Golan (with extreme high summer tem-peratures and low winter temperatures, respectively) is a signthat the wasp population thrives under a wide range of tem-peratures. In general, L. invasa in Israel may produce two orthree overlapping generations annually. Nourishment on asolution of honey + water markedly improved the life spanof adult parasitic wasps (e.g., Mendel 1988). This diet waspreviously found considerably to extend the longevity of
Fig. 22. Relationships between number of immature Leptocybeinvasa and gall weight and gall length. (a) y = 6.0317x + 0.8497;R2 = 0.8834. (b) y = 9.38 + 25.069; R2 = 0.7.
90
(a)
(b)
80
70
60
50
40
30
20
10
0
Gal
l wei
ght
0
20
40
60
80
100
120
140
160
180
0 2 4 6 8 10 12
Number of individuals per gall
Gal
l len
gth
(mm
)
Fig. 23. Mean (±SD) weight of Leptocybe invasa (larvae orpupa) and gall according to gall stage. (�), Gall weight; (�),wasp weight.
0
2
2 3 4
6
4
8
Gall stageG
all w
eigh
t
0.3
0.2
0.1
0
Mea
n w
eigh
t of i
mm
atur
e w
asp
Fig. 24. Relationship between gall weight (stages 2–4) andmean weight of immature wasp in Leptocybe invasa. Pearsoncorrelation coefficient (r) = -0.471; (P = 0.0001).
0.3
0.2
0.1
0
4 5 6 7 8 9
Gall weight (mg) / number of immature wasps per gal
Mea
n w
eigh
t of i
mm
atur
e w
asp

110 Z Mendel et al.
Fig. 25. Development time, from oviposition to adult emergence, of galls of Leptocybe invasa on artificially infested leaves ofEucalyptus camaldulensis in a greenhouse. The plants were exposed to the wasps in July and adult emergence was observed betweenearly and mid November
0 10 20 30 40 50
Stage 4 - stage 5
Stage 3 - stage 4
Stage 2 - stage 3
Stage 1 - stage 2
Oviposition - stage 1
Gall development
Average (±SD) number of days
Table 1 Eucalyptus species examined in Israel in plantations and nurseries as related to theirsuitability for the development of Leptocybe invasa. Classification of Eucalyptus according toBrooker (2000)
Eucalyptus Subgenus Section Series Species Suitability
Corymbia Septentrionales Maculatae citriodora –Maculatae maculata –Torellianae torelliana –
Eudesmia Limbata Heteroptera erythrocorys –Symphyomyrtus Adnataria Siderophloiae crebra –
Buxeales hemiphloia = moluccana –Melliodorae leucoxylon –Siderophloiae melanophloia –Heterophloiae polyanthemos –Buxeales populnea –Melliodorae sideroxylon –
Bisecta Erectae astringens –Dundasianae dundasii –Elongatae erythronema –Loxophlebae loxophleba –Erectae occidentalis –Erectae platypus –Contortae salubris –Erectae sargentii –Erectae spathulata –Stricklandiae stricklandii –
Bolites gomphocephala –Dumaria Rufispermae kondininensis –
Torquatae torquata –Rufispermae woodwardii –
Exsertaria Rostratae camaldulensis +Erythroxyla tereticornis +
Latoangulata Annulares botryoides +Transversae grandis +Annulares robusta +Transversae saligna +
Maidenaria Bridgesianae bridgesiana +Globulares globulus +Orbiculares gunii +Viminales viminalis +
Sejuncta cladocalyx –

Biology of Leptocybe invasa 111
Fig. 26. Development of four cohorts of Leptocybe invasa galls on Eucalyptus camaldulensis at Bet Dagan, Israel. Each cohortconsisted of galls of a different gall development stage at the beginning of the survey (June 2002). N = number of galls per cohort.
Cohort 1, N = 60
Cohort 2, N = 70
Cohort 4, N = 50
Cohort 3, N = 50
Per
cent
age
of g
alls
of
each
sta
ge
100
80
60
40
20
0
Per
cent
age
of g
alls
of
each
sta
ge
100
80
60
40
20
0
Per
cent
age
of g
alls
of
each
sta
ge
100
80
60
40
20
0
Per
cent
age
of g
alls
of
each
sta
ge
100
80
60
40
20
0
35
30
25
20
15
VI VII VIII IXMonth
Examination time (2002)
June
6
June
20
June
27
July
4
July
10
July
24
Augus
t 7
Septe
mbe
r 5
Maximumminimum
(°C
)
1
2
3
4
5
Gall Stages
O. eucalypti (Withers et al. 2000). Our findings suggest thatL. invasa adults may prolong their activity by feeding on theflowers of their host plant.
The rapid spread and fast population growth are probablythe result of its thelytokous reproduction, mulivotinousdevelopment and, perhaps, also of the absence of its principalenemies.
The widespread and large populations of L. invasa in theMediterranean and the fact that the genus Leptocybe has notbeen detected as yet in Australia, suggests that in Australia,natural enemies play a significant role in reducing this waspspecies to below the observation threshold. These natural ene-mies may serve as appropriate candidates for classical biolog-ical control in the new areas colonised by L. invasa.

112 Z Mendel et al.
Fig. 27. Trend in gall stage of Leptocybe invasa through thewinter. Two distinct groups of gall stages were selected. The gallsdeveloped on Eucalyptus camaldulensis in the Bet Shean Valleyin Israel and were monitored each month between mid-October2002 and May 2003.
0
1
3
2
4
5
Octobe
r
Janu
ary
Decem
ber
Novem
ber
Mar
ch
Febru
ary
April
May
Mea
n ga
ll st
age
adult emergence
dead galls
4 trees
6 trees
Fig. 28. Occurrence of eggs of Leptocybe invasa as related todevelopment of young foliage of Eucalyptus camaldulensis at NirDavid and to the temperatures in the region during the time of thesurvey. (a) (�) Branches with new growth; (�) branches witheggs. (b) (�) Minimum; (�) maximum.
0
20
40
60
80
100
(a)
(b)
Num
ber
of b
ranc
hes
0
5
10
15
20
25
30
35
40
Octobe
r
Novem
ber
Decem
ber
Janu
ary
Febru
ary
Mar
chApr
ilM
ay
Month of observation (2002–2003)
Tem
pera
ture
°C
Fig. 29. Survival patterns of Leptocybe invasa at 25∞C as relatedto food possibilities. Each treatment involved 30 wasps. (�),Honey + water (-25.846 a); (�) honey + water + young leaves(-19.063 a); (¥) flowers (0.776 b); (�) water (11.810 c); (�)young leaves (13.976 cd); (�) no food (18.348 d). In parenthesisvalues of log-rank statistics with the same letter are not statisti-cally different (P < 0.0001).
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8
Days
Per
cent
age
surv
ivor
s
Fig. 30. Typical Leptocybe invasa galling deformation on abranch of Eucalyphis camaldulensis.

Biology of Leptocybe invasa 113
ACKNOWLEDGEMENTS
This study was partly supported by the Forests Department ofthe Jewish National Fund, Israel, and Jewish National Fund ofAustralia.
Our thanks to Fabienne Assael and Dani Blumberg(Volcani Center), Nitza Saphir, Ami Zehavi, Zion Madar andDavid Brand (Forests Department, Jewish National Fund, Esh-taol, Israel), and Ido Goffer (Nir David, DN Bet Shean, Israel)for field and laboratory assistance, to Abraham Weinstein (For-ests Department, Jewish National Fund) for the identificationof many Eucalyptus trees, and to John Connors (CSIROPlant Industry) who supplied information on Eucalyptusclassification.
REFERENCES
Abrahamson WG & Weis RE. 1987. Nutritional ecology of arthropod gallmakers. In: Nutritional Ecology of Insects, Mites and Spiders (edsF Slansky & JG Rodriguez), pp. 235–258. John Wiley, New York,USA.
Bain J. 1977. Megastigmus spermotrophus Wachtl (Hymenoptera:Chalcidoidea: Torymidae); Rhicnopeltella eucalypti Gahan(Hymenoptera: Chalcidoidea: Eulophidae), blue-gum chalcid. Forestand Timber Insects in New Zealand, New Zealand Forest Service no.14–15, 4pp.
Beardsley JW & Perreira WD. 2000. Aprostocetus sp. (Hymenoptera:Eulophidae: Tetrastichinae), a gall wasp new to Hawaii. Proceedingsof the Hawaiian Entomological Society 34, 203.
Berry JA & Withers TM. 2002. New gall-inducing species oformocerine pteromalid (Hymenoptera: Pteromalidae: Ormocerinae)described from New Zealand. Australian Journal of Entomology41, 18–22.
Bou ek Z. 1988. Australasian Chalcidoidea (Hymenoptera). CAB Inter-national, Wallingford, UK.
Brooker MIH. 2000. A new classification of the genus Eucalyptus.Australian Systematic Botany 13, 79–148.
FAO. 1979. Eucalyptus for Planting. FAO Forestry Series, 11. FAO,Rome, Italy.
Flock RA. 1957. Biological notes on a new chalcid-fly from seed-likeEucalyptus galls in California. Pan-Pacific Entomologist 33, 153–155.
Gibson GAP. 1997. Morphology and terminology. In: Annotated Keysto the Genera of Nearctic Chalcidoidea (Hymenoptera) (eds GAP
c
Gibson, JT Huber & JB Woolley), pp. 16–44. National ResearchCouncil Research Press, Ottawa, Ontario, Canada.
de Graham MWR & V. 1987. A reclassification of the European Tetras-tichinae (Hymenoptera: Eulophidae), with a revision of certaingenera. Bulletin of the British Museum (Natural History), EntomologySeries 55, 1–392.
de Graham MWR & V. 1991. A reclassification of the European Tetras-tichinae (Hymenoptera: Eulophidae): revision of the remaining gen-era. Memoirs of the American Entomological Institute 49, 1–322.
Headrick DH, La Salle J & Redak RA. 1995. A new genus of AustralianTetrastichinae (Hymenoptera: Eulophidae): an introduced pest ofChamelaucium uncinatum (Myrtaceae) in California. Journal ofNatural History 29, 1029–1036.
La Salle J. 1994. North American genera of Tetrastichinae (Hymenoptera:Eulophidae). Journal of Natural History 28, 109–236.
Mendel Z. 1985. Seasonal development of the eucalypt borer Phoracan-tha semipunctata (Coleoptera: Cerambycidae) in Israel. Phytopara-sitica 14, 85–93.
Mendel Z. 1988. Effect of food, temperature, and breeding conditions onthe life span of adults of three cohabitating bark beetles (Scolytidae)parasitoids (Hymenoptera). Environmental Entomology 17, 293–298.
Rohfriotsch O & Shorthouse JD. 1982. Insect galls. In: Molecular Biologyof Plant Tumors (eds G Kahl & J Schell), pp. 131–152. AcademicPress, New York, USA.
SAS Institute. 2002. SAS/STAT User’s guide, Version 6.11 Vol. 1+2., SASInstitute, Cary, NC, USA.
Schauff ME & Garrison RA. 2000. Introduced species of Epichrysocharis(Hymenoptera: Eulophidae) producing galls on Eucalyptus.California with notes on the described species and placement of thegenus. Journal of Hymenoptera Research 9, 176–181.
Schauff ME, La Salle J & Coote LD. 1997. Eulophidae. In: AnnotatedKeys to the Genera of Nearctic Chalcidoidea (Hymenoptera) (edsGAP Gibson, JT Huber & JB Woolley), pp. 327–429. NationalResearch Council Research Press, Ottawa, Canada.
Timberlake PH. 1957. A new entedontine chalcid-fly from seed capsulesof eucalyptus in California (Hymenoptera: Eulophidae). Pan-PacificEntomologist 33, 109–110.
Viggiani G & Nicotina M. 2001. The eulophid leaf gall-making of euca-lypts Ophelimus eucalypti (Gahan) (Hymenoptera: Eulophidae) inCampania. Bollettino Di Zoologia Agraria E Di Bachicoltura 33, 79–82.
Withers TM. 2001. Colonization of eucalypts in New Zealand byAustralian insects. Austral Ecology 26, 467–476.
Withers TM, Raman A & Berry JA. 2000. Host range and biology ofOphelimus eucalypti (Gahan) (Hym., Eulophidae), a pest of NewZealand eucalypts. New Zealand Plant Protection 53, 339–344.
Accepted for publication 3 November 2003.
Copyright © 2022 FDOKUMEN




















