
Expression Studies of Neogenin and Its Ligand Hemojuvelin in Mouse Tissues
13
http://jhc.sagepub.com/ Journal of Histochemistry & Cytochemistry http://jhc.sagepub.com/content/55/1/85 The online version of this article can be found at: DOI: 10.1369/jhc.6A7031.2006 2007 55: 85 J Histochem Cytochem Alejandra Rodriguez, Peiwen Pan and Seppo Parkkila Expression Studies of Neogenin and Its Ligand Hemojuvelin in Mouse Tissues Published by: http://www.sagepublications.com On behalf of: Official Journal of The Histochemical Society can be found at: Journal of Histochemistry & Cytochemistry Additional services and information for http://jhc.sagepub.com/cgi/alerts Email Alerts: http://jhc.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: What is This? - Jan 1, 2007 Version of Record >> by guest on September 21, 2014 jhc.sagepub.com Downloaded from by guest on September 21, 2014 jhc.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression Studies of Neogenin and Its Ligand Hemojuvelin in Mouse Tissues

http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/55/1/85The online version of this article can be found at:
DOI: 10.1369/jhc.6A7031.2006
2007 55: 85J Histochem CytochemAlejandra Rodriguez, Peiwen Pan and Seppo Parkkila
Expression Studies of Neogenin and Its Ligand Hemojuvelin in Mouse Tissues
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Jan 1, 2007Version of Record >>
by guest on September 21, 2014jhc.sagepub.comDownloaded from by guest on September 21, 2014jhc.sagepub.comDownloaded from

ARTICLE
Expression Studies of Neogenin and Its Ligand Hemojuvelin inMouse Tissues
Alejandra Rodriguez, Peiwen Pan, and Seppo Parkkila
Institute of Medical Technology, University of Tampere and Tampere University Hospital, Tampere, Finland (AR,PP,SP), andDepartment of Clinical Chemistry, University of Oulu, Oulu, Finland (SP)
SUMMARY Juvenile hemochromatosis is a severe hereditary iron overload disease causedby mutations in the HJV (hemojuvelin) and HAMP (hepcidin) genes. Hepcidin is an importantiron regulatory hormone, and hemojuvelin may regulate hepcidin synthesis via the multi-functional membrane receptor neogenin. We explored the expression ofmurine hemojuvelinand neogenin mRNAs and protein. Real-time RT-PCR analysis of 18 tissues from male andfemale mice was performed to examine the mRNA expression profiles. To further studyprotein expression and localization we used immunohistochemistry on several tissues fromthree mouse strains. Mouse Neo1 mRNA was detectable in the 18 tissues tested, the highestsignals being evident in the ovary, uterus, and testis. Neogenin protein was observed in thebrain, skeletal muscle, heart, liver, stomach, duodenum, ileum, colon, renal cortex, lung,testis, ovary, oviduct, and uterus. The spleen, thymus, and pancreas were negative for neo-genin. The highest signals for Hjv mRNA were detectable in the skeletal muscle, heart,esophagus, and liver. The results indicate that Neo1 mRNA is widely expressed in both maleand female mouse tissues with the highest signals detected in the reproductive system.Moreover, Hjv and Neo1 mRNAs are simultaneously expressed in skeletal muscle, heart,esophagus, and liver. (J Histochem Cytochem 55:85–96, 2007)
KEY WORDS
expression
hemochromatosis
hemojuvelin
iron
neogenin
HEREDITARY HEMOCHROMATOSIS (HH) comprises severalgenetic disorders of iron overload. In all cases, the nor-mal balance between iron absorption in the intestineand iron stores and needs is perturbed. HH mutationshave now been described in several different genes in-cluding SCL40A1 (encoding ferroportin1), HFE, TFR2(encoding the transferrin receptor-2), HJV (encodinghemojuvelin), and HAMP (encoding hepcidin) (Njajouet al. 2001; Fleming 2005; Le Gac and Ferec 2005).Disruption of the SCL40A1 gene causes an autosomaldominant disorder characterized by predominantly re-ticuloendothelial iron accumulation (Pietrangelo 2004).Disruption of any of the remaining four genes leadsto autosomal recessive disorders characterized by hightransferrin saturation, depletion of iron in macrophages,and deposition of iron in several organs including theliver, heart, and pancreas (Feder et al. 1996; Camaschella
et al. 2000; Roetto et al. 2003; Papanikolaou et al. 2004).These four HH variants also share an inappropriatelylow level of HAMP expression (Bridle et al. 2003; Roettoet al. 2003; Papanikolaou et al. 2004; Nemeth et al.2005). However, cases of HH with higher penetrance,faster iron loading, and earlier clinical complications areassociated with mutations in either HJV or HAMP. Theearly onset of this disorder is referred to as juvenile he-mochromatosis (JH) (Roetto et al. 2003; Papanikolaouet al. 2004).
Hepcidin is an antimicrobial peptide and iron regu-latory hormone. It binds to the iron exporter ferropor-tin and induces its internalization from the cell surfaceand subsequent degradation (Nemeth et al. 2004). Fer-roportin is expressed in hepatocytes, reticuloendo-thelial system cells, and in the basolateral membraneof duodenal enterocytes (Abboud and Haile 2000).Thus, hepcidin negatively regulates the rate of intestinaliron absorption and, in general, of iron efflux into theplasma. The low production of hepcidin observed inHH patients (see above) can therefore explain the highlevels of intestinal iron absorption and high transferrinsaturation; hence, the overall HH phenotype. More-
Correspondence to: Alejandra Rodriguez, Institute of MedicalTechnology, University of Tampere, Biokatu 6 FIN-33520, Tampere,Finland. E-mail: [email protected]
Received for publication June 13, 2006; accepted September 1,2006 [DOI: 10.1369/jhc.6A7031.2006].
TheJournal
ofHistoch
emistry&
Cytoch
emistry
C The Histochemical Society, Inc. 0022-1554/06/$3.30 85
Volume 55(1): 85–96, 2007
Journal of Histochemistry & Cytochemistry
http://www.jhc.org
by guest on September 21, 2014jhc.sagepub.comDownloaded from

over, HH-related HAMP downregulation indicates thatHFE protein, transferrin receptor-2, and hemojuvelinare upstream regulators of HAMP, although the un-derlying pathways remain obscure.
Hemojuvelin (HJV, HFE2, or RGMc) is a glycosylphosphatidylinositol-anchored protein for which fivespliced variants have been predicted, encoding proteinsof 200, 313, and 423 amino acids (Papanikolaou et al.2004; Zhang et al. 2005). To date, there is no clearevidence for the production of the two shorter peptides,and new findings suggest that under certain conditionsthe full-length HJV is posttranslationally processed toyield a shorter N-terminal and a larger C-terminal prod-uct (Zhang et al. 2005). Of additional importance, arecent study has revealed that hemojuvelin can be foundin the human body in two forms: membrane associatedand soluble hemojuvelin proteins. These two hemojuve-lin types appear to reciprocally regulate HAMP expres-sion, in response to extracellular iron concentrations inhuman primary hepatocytes, by physically competingfor a receptor-binding site (Lin et al. 2005). Interestingly,another recent report has shown evidence for a physicalassociation between hemojuvelin and the membrane re-ceptor neogenin [but not its homolog, Deleted in Colo-rectal Cancer (DCC)] in cultured embryonic kidney cells.This same study demonstrated that the hemojuvelin–neogenin interaction is required for iron accumula-tion is these cells (Zhang et al. 2005). Based on thesenew findings, a logical hypothesis would be that hemo-juvelin exerts its HAMP regulatory role through its re-ceptor, neogenin.
Neogenin is a type I transmembrane receptor that ishomologous to DCC, and both constitute a subfamilywithin the N-CAM family of cell adhesion molecules.The extracellular regions of neogenin and DCC containfour immunoglobulin domains with disulfide-bondedcysteines and six type-III fibronectin repetitions. Thecytoplasmic domains of these two receptors have nohomology to any other known proteins (Meyerhardtet al. 1997). Diverse functions of neogenin upon inter-action with different ligands have been reported, suchas the repulsive guidance of retinal axons and the reg-ulation of neuronal survival via the interaction withchick RGM (repulsive guidance molecule) (Monnier
et al. 2002; Matsunaga et al. 2004). Another functionof neogenin is its role in myotube formation, mediatedby its binding to netrin-3 (Kang et al. 2004).
It is significant that the binding ligands of neogeninare expressed in quite distinct locations (Schmidtmerand Engelkamp 2004; Barallobre et al. 2005), furthersuggesting that it may act as a multifunctional receptorwhose role is determined by the presence of specific in-teracting molecules. In our present study in the mouse,we elucidate sites of simultaneous expression of Neo1and Hjv mRNA. We also explore the spatial distribu-tion of the murine neogenin and hemojuvelin proteinsin different tissues.
Materials and Methods
RNA Extraction and First-strand cDNA Synthesis
Tissue samples for mRNA quantification were obtained from10-week-old NMRI mice (four males and four females) withthe approval of the Animal Care Committee of the Universityof Tampere, Tampere, Finland. The tissues extracted includedthe lung, brain, muscle, heart, spleen, thymus, pancreas, liver,esophagus, stomach, duodenum, jejunum, ileum, colon, kid-ney, testis, ovary, and uterus. All specimens used for quan-titative real-time PCR were snap frozen upon extraction andstored at �80C until use. Total RNA was isolated usingTRIZOL reagent (Invitrogen; Carlsbad, CA) according to themanufacturer’s instructions. After digestion with RNase-freeDNase I (Novagen; Madison, WI), the resulting RNA sam-ples were further purified using phenol/chloroform, followedby precipitation with ice-cold ethanol. RNA concentrationand purity were determined in each case by optical densitymeasurements at 260 and 280 nm. RNA extracts from malesand females were separately pooled to reduce the potential forindividual variation. Three Ag of each total RNA isolate wasconverted into first-strand cDNA with a First Strand cDNASynthesis kit (Fermentas; Burlington, Canada) and randomhexamer primers, according to the protocol recommended bythe manufacturer.
Quantitative Real-time PCR
The relative levels of mouse Neo1 and Hjv transcripts in dif-ferent tissues were assessed by real-time PCR using the RocheLightCycler detection system (Roche; Rotkreuz, Switzerland).The primer sets for the target genes used in these analyses weredesigned using Primer3 http://puma.fmvz.usp.br/primer3/
Table 1 Primer sequences for internal control genes
Gene Forward primer (59-39), reverse primer (59-39)
Actb (h-actin) AGAGGGAAATCGTGCGTGAC CAATAGTGATGACCTGGCCGThttp://web.ncifcrf.gov/rtp/gel/primerdb/ ID:634
Gapdh (glyceraldehyde-3-phosphate dehydrogenase) ATGGTGAAGGTCGGTGTG CATTCTCGGCCTTGACTG*Hprt1 (hypoxanthine phosphoribosyl-transferase I) AGCTACTGTAATGATCAGTCAACG AGAGGTCCTTTTCACCAGCA
http://web.ncifcrf.gov/rtp/gel/primerdb/ ID.10050Sdha (succinate dehydrogenase complex subunit A) GCTTGCGAGCTGCATTTGG CATCTCCAGTTGTCCTCTTCCA
http://pga.mgh.harvard.edu/primerbank/ ID: 15030102a2
*Designed using Primer3 (http://puma.fmvz.usp.br/primer3/primer3_www.cgi).
TheJournal
ofHistoch
emistry&
Cytoch
emistry
86 Rodriguez, Pan, Parkkila
by guest on September 21, 2014jhc.sagepub.comDownloaded from
primer3_www.cgi), based on the complete cDNA sequencesdeposited in GenBank [accession numbers: NM_008684 formouse neogenin (Neo1) and NM_027126 for mouse hemo-juvelin (Hjv)]. The specificity of the primers was verified usingNCBI Blast http://www.ncbi.nlm.nih.gov/BLAST/). To avoidamplification of contaminating genomic DNA, both primersfrom each set were specific to different exons. The Neo1forward 59-CCCTGGTCTCTACTCGCTTC-39 and reverse59-CCTGGCTGGCTGGTATTCTC-39 primers are specific toexons eight and nine, respectively. The Hjv forward 59-TC-TGACCTGAGTGAGACTGC-39 and reverse 59-GATGAT-GAGCCTCCTACCTA-39 primers are located in exons 1 and2, respectively. These primers amplify the mouse Hjv tran-scripts 1 and 3.
The Actb (h-actin), Gapdh (glyceraldehyde-3-phosphatedehydrogenase), Hprt1 (hypoxanthine phosphoribosyl-trans-ferase I), and Sdha (succinate dehydrogenase complex sub-unit A) genes were used as internal controls to normalize forpotential quality and quantity differences between samples.The primers for the internal controls are shown in Table 1.Every PCR reaction was performed in a total volume of 20 Alcontaining 1 Al of first-strand cDNA, 1X concentrated Quan-tiTect SYBR Green PCR Master Mix (Qiagen; Hilden, Ger-many), and 0.5 Amol/liter of each primer. Amplifications andsubsequent detection were carried out as described. After aninitial activation step of 15 min at 95C, amplification wasperformed in a three-step cycling procedure: denaturation at95C, 15 sec, ramp rate 20C/sec; annealing according to the
melting temperature of the primers, 20 sec, ramp rate 20C/sec; and elongation at 72C, 20 sec, ramp rate 20C/sec for45 cycles, and a final cooling step. The melting curve analysiswas always performed for each PCR amplicon to verify spe-cific amplification.
To quantify the concentration of the Actb, Gapdh, HprtI,Sdha, Neo1, and Hjv transcripts in the studied tissues, a stan-dard curve for each gene was established using 5-fold serialdilutions of known concentrations of purified PCR productsgenerated from the same primer sets. Every cDNA samplewas tested in duplicate and the obtained crossing point (Cp)value facilitated the determination of the levels of startingmessage using a specific standard curve. The geometric meanof the four internal control genes was used as an accurate nor-malization factor for gene expression levels (Vandesompeleet al. 2002). The level of expression of the target gene in eachtissue was divided by the corresponding normalization factorand subsequently multiplied by 102.
Immunohistochemistry
Tissue specimens were obtained from eight adult mice in-cluding four NMRI (two male and two female), two Balb/c(male and female), and two C57 (male and female) mice. Allavailable samples were used for neogenin studies, whereashemojuvelin staining was performed on samples from femaleC57 mice. After extraction, samples were fixed in 4% neutral-buffered formaldehyde at 4C for 24 hr and then dehydrated
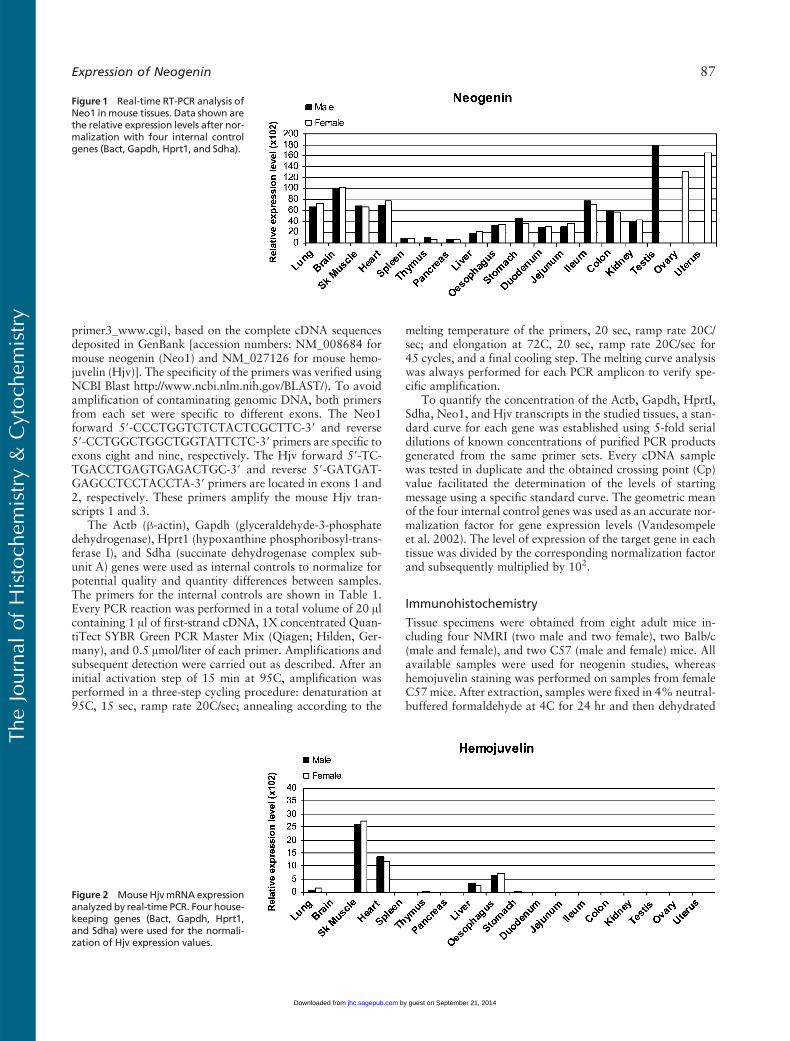
Figure 1 Real-time RT-PCR analysis ofNeo1 inmouse tissues. Data shown arethe relative expression levels after nor-malization with four internal controlgenes (Bact, Gapdh, Hprt1, and Sdha).
Figure 2 MouseHjvmRNAexpressionanalyzed by real-time PCR. Four house-keeping genes (Bact, Gapdh, Hprt1,and Sdha) were used for the normali-zation of Hjv expression values.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Expression of Neogenin 87
by guest on September 21, 2014jhc.sagepub.comDownloaded from
in an alcohol series, treated with xylene, and embeddedin paraffin wax. Four-Am sections were cut and placed onSuperFrost Plus (Menzel-Glaser; Braunschweig, Germany)mi-croscope slides.
After deparaffinization, immunostaining was performedby the biotin–streptavidin–peroxidase complex method.Briefly, antigen retrieval was performed in an autoclave at95C. The parameters for neogenin were 30 min in 10 mmol/liter citrate buffer, pH 6.0. For hemojuvelin experiments,slides were treated for 25 min in 10 mmol/liter citrate buffer,pH 9. After endogenous peroxidase activity was quenchedand nonspecific binding was blocked, the slides were incu-bated overnight at 4Cwith the rabbit anti-neogenin polyclonalantibody (1:50 dilution, sc-15337; Santa Cruz Biotechnology,Santa Cruz, CA) previously characterized (Lee et al. 2005) oralternatively with the 1:250 diluted rabbit anti-hemojuvelinpolyclonal antibody that has also been previously character-ized (Rodriguez Martinez et al. 2004). The slides were thenwashed three times with PBS for 10 min. Biotinylated goat
anti-rabbit secondary antibody (Zymed Laboratories; SouthSan Francisco, CA) was used at a dilution of 1:500 in a 60-minincubation step at room temperature. After washing, the sec-tions were incubated with streptavidin–horseradish per-oxidase conjugate (1:750 dilution; Zymed Laboratories) for30 min at room temperature. The slides were then washedagain four times for 5min in PBS and the staining reaction wascarried out using 3,39-diaminobenzidine tetrahydrochloride(DAB) as a chromogen. Finally, a 4-min incubation in he-matoxylin was performed to stain the nuclei of the cells andthus facilitate interpretation of the results. Neogenin experi-ments were accompanied by negative and positive controlstainings to detect for possible nonspecific signals. Negativecontrols were processed by replacing the primary antibodywith diluent. As a positive control parallel tissue sections wereimmunostained using a polyclonal rabbit anti-mouse PCNAantibody (Santa Cruz Biotechnology). Two kinds of negativecontrol were used to detect possible nonspecific signals in thehemojuvelin staining. The preimmune serum controls con-
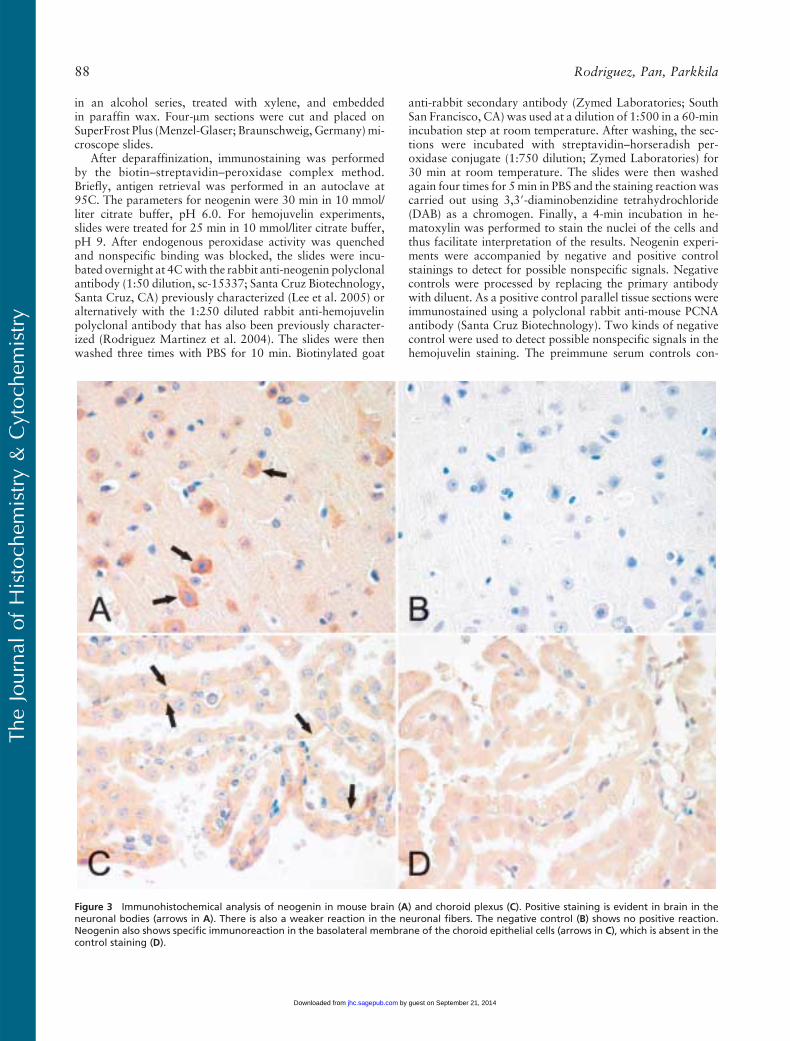
Figure 3 Immunohistochemical analysis of neogenin in mouse brain (A) and choroid plexus (C). Positive staining is evident in brain in theneuronal bodies (arrows in A). There is also a weaker reaction in the neuronal fibers. The negative control (B) shows no positive reaction.Neogenin also shows specific immunoreaction in the basolateral membrane of the choroid epithelial cells (arrows in C), which is absent in thecontrol staining (D).
TheJournal
ofHistoch
emistry&
Cytoch
emistry
88 Rodriguez, Pan, Parkkila
by guest on September 21, 2014jhc.sagepub.comDownloaded from
sisted of replacement of the primary antibodywith normal pre-immune rabbit serum. The diluent controls were performed asdescribed above.
Results
Neo1 and Hjv Gene Expression Profiles in Male andFemale Mice
Expression levels of Neo1 mRNA were examined in abroad range of mouse tissues by quantitative real timeRT-PCR. As shown in Figure 1, Neo1 transcripts weredetectable in each of the tissues tested for both gen-ders with the highest expression observed in the testis,ovary, and uterus. A relatively high signal was also ob-served in brain, followed by lung, skeletal muscle, andheart. In the digestive system, the highest expression forNeo1 was found in distal parts of the intestine (theileum and colon). However, moderate signal levels werealso observed in the esophagus, stomach, duodenum,
and jejunum, with a similar intensity as in the kidney,outside of the digestive system. The lowest Neo1 tran-script levels were observed in the liver, thymus, spleen,and pancreas. There were no marked differences in thelevels of expression observed in males vs females.
The expression of Hjv mRNA was also determinedby real-time RT-PCR (Figure 2). Positive tissues in-cluded skeletal muscle, heart, esophagus, and liver inwhich the signal intensities were at approximately thesame levels in both males and females. Very weak sig-nals were also evident in the lung and in the thymus andstomach of male mice.
Expression of Neogenin and HemojuvelinProteins in Mouse
Localization of neogenin protein in mouse tissues wasexamined by immunohistochemistry and revealed nomarked differences in the staining patterns between
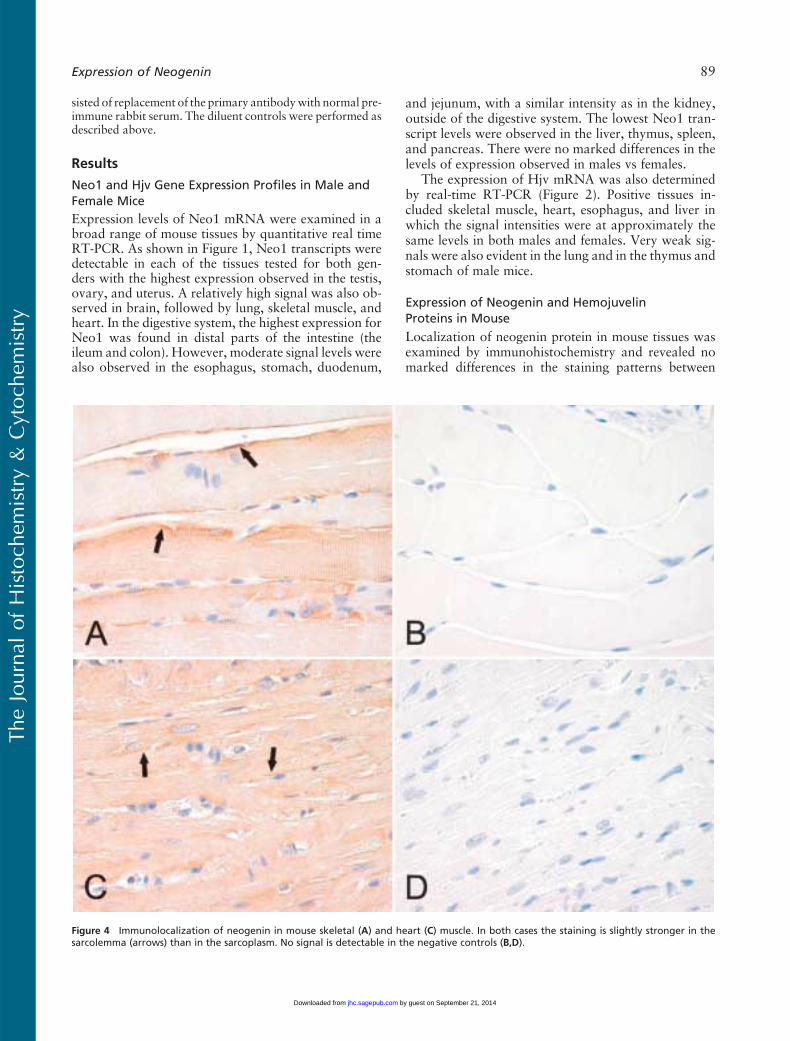
Figure 4 Immunolocalization of neogenin in mouse skeletal (A) and heart (C) muscle. In both cases the staining is slightly stronger in thesarcolemma (arrows) than in the sarcoplasm. No signal is detectable in the negative controls (B,D).
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Expression of Neogenin 89
by guest on September 21, 2014jhc.sagepub.comDownloaded from
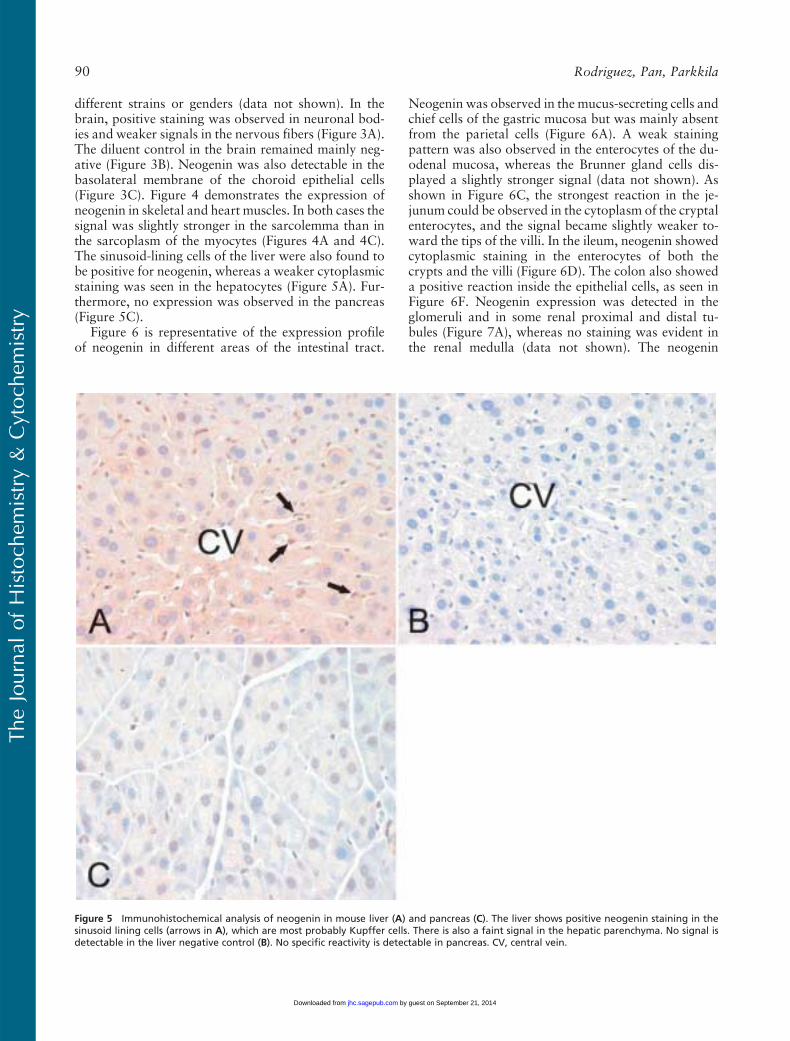
different strains or genders (data not shown). In thebrain, positive staining was observed in neuronal bod-ies and weaker signals in the nervous fibers (Figure 3A).The diluent control in the brain remained mainly neg-ative (Figure 3B). Neogenin was also detectable in thebasolateral membrane of the choroid epithelial cells(Figure 3C). Figure 4 demonstrates the expression ofneogenin in skeletal and heart muscles. In both cases thesignal was slightly stronger in the sarcolemma than inthe sarcoplasm of the myocytes (Figures 4A and 4C).The sinusoid-lining cells of the liver were also found tobe positive for neogenin, whereas a weaker cytoplasmicstaining was seen in the hepatocytes (Figure 5A). Fur-thermore, no expression was observed in the pancreas(Figure 5C).
Figure 6 is representative of the expression profileof neogenin in different areas of the intestinal tract.
Neogenin was observed in the mucus-secreting cells andchief cells of the gastric mucosa but was mainly absentfrom the parietal cells (Figure 6A). A weak stainingpattern was also observed in the enterocytes of the du-odenal mucosa, whereas the Brunner gland cells dis-played a slightly stronger signal (data not shown). Asshown in Figure 6C, the strongest reaction in the je-junum could be observed in the cytoplasm of the cryptalenterocytes, and the signal became slightly weaker to-ward the tips of the villi. In the ileum, neogenin showedcytoplasmic staining in the enterocytes of both thecrypts and the villi (Figure 6D). The colon also showeda positive reaction inside the epithelial cells, as seen inFigure 6F. Neogenin expression was detected in theglomeruli and in some renal proximal and distal tu-bules (Figure 7A), whereas no staining was evident inthe renal medulla (data not shown). The neogenin
Figure 5 Immunohistochemical analysis of neogenin in mouse liver (A) and pancreas (C). The liver shows positive neogenin staining in thesinusoid lining cells (arrows in A), which are most probably Kupffer cells. There is also a faint signal in the hepatic parenchyma. No signal isdetectable in the liver negative control (B). No specific reactivity is detectable in pancreas. CV, central vein.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
90 Rodriguez, Pan, Parkkila
by guest on September 21, 2014jhc.sagepub.comDownloaded from
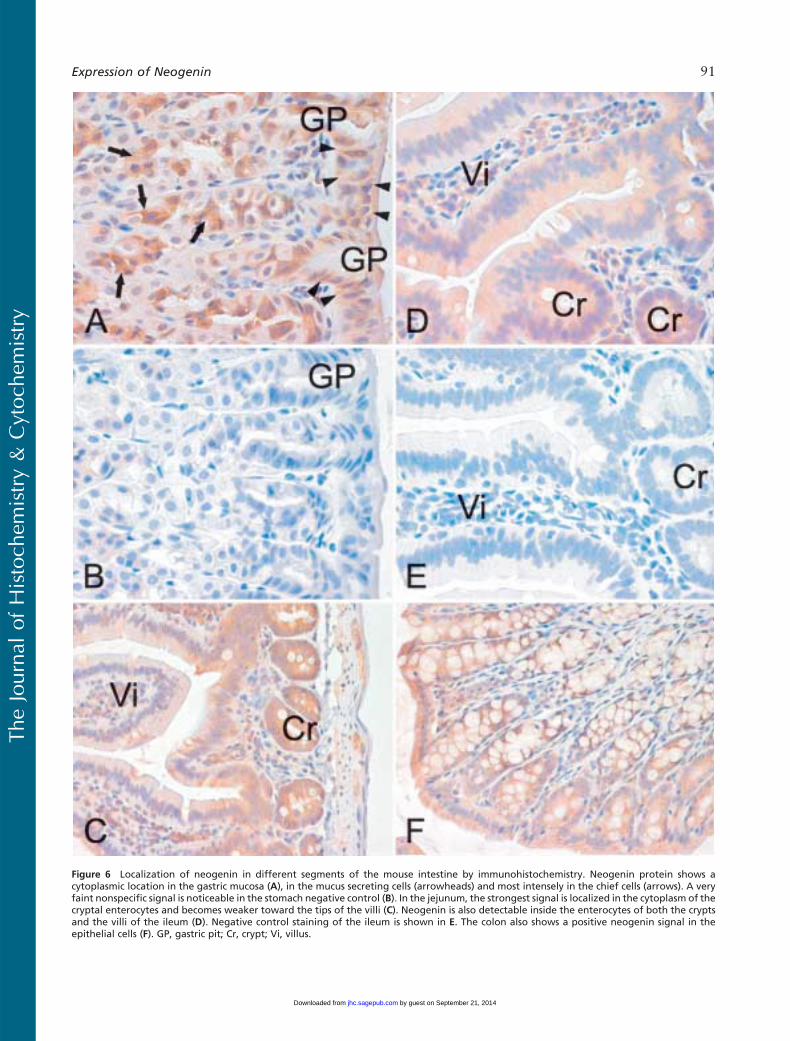
Figure 6 Localization of neogenin in different segments of the mouse intestine by immunohistochemistry. Neogenin protein shows acytoplasmic location in the gastric mucosa (A), in the mucus secreting cells (arrowheads) and most intensely in the chief cells (arrows). A veryfaint nonspecific signal is noticeable in the stomach negative control (B). In the jejunum, the strongest signal is localized in the cytoplasm of thecryptal enterocytes and becomes weaker toward the tips of the villi (C). Neogenin is also detectable inside the enterocytes of both the cryptsand the villi of the ileum (D). Negative control staining of the ileum is shown in E. The colon also shows a positive neogenin signal in theepithelial cells (F). GP, gastric pit; Cr, crypt; Vi, villus.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Expression of Neogenin 91
by guest on September 21, 2014jhc.sagepub.comDownloaded from
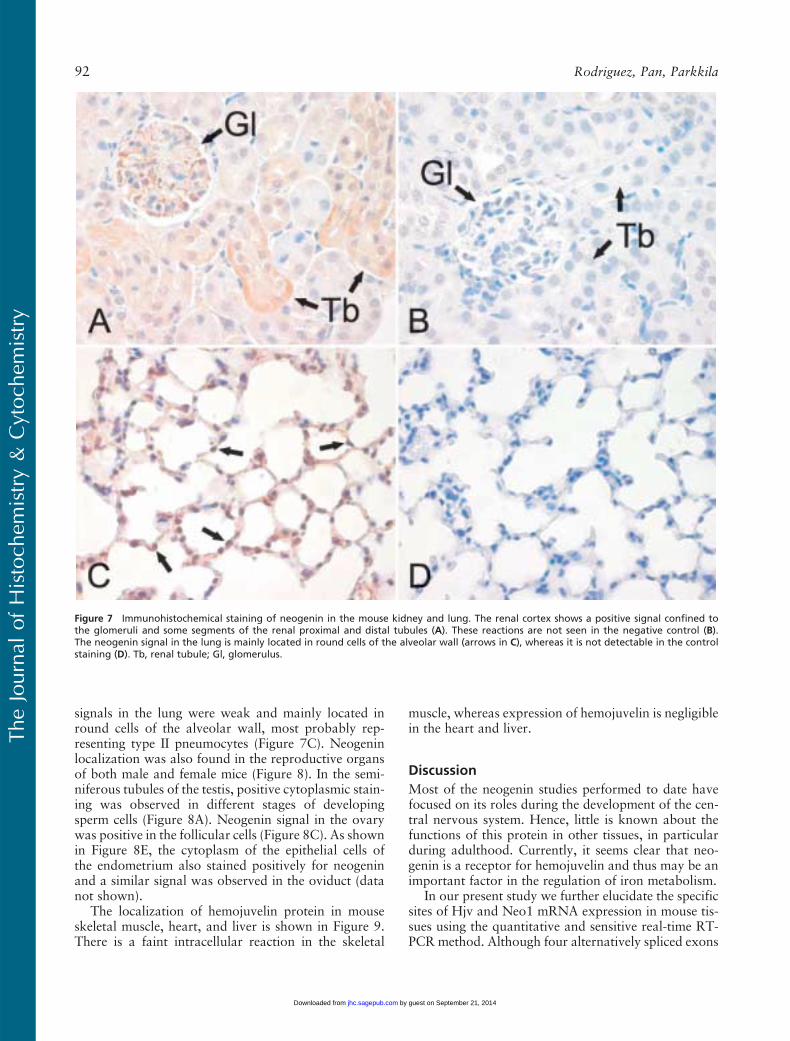
signals in the lung were weak and mainly located inround cells of the alveolar wall, most probably rep-resenting type II pneumocytes (Figure 7C). Neogeninlocalization was also found in the reproductive organsof both male and female mice (Figure 8). In the semi-niferous tubules of the testis, positive cytoplasmic stain-ing was observed in different stages of developingsperm cells (Figure 8A). Neogenin signal in the ovarywas positive in the follicular cells (Figure 8C). As shownin Figure 8E, the cytoplasm of the epithelial cells ofthe endometrium also stained positively for neogeninand a similar signal was observed in the oviduct (datanot shown).
The localization of hemojuvelin protein in mouseskeletal muscle, heart, and liver is shown in Figure 9.There is a faint intracellular reaction in the skeletal
muscle, whereas expression of hemojuvelin is negligiblein the heart and liver.
Discussion
Most of the neogenin studies performed to date havefocused on its roles during the development of the cen-tral nervous system. Hence, little is known about thefunctions of this protein in other tissues, in particularduring adulthood. Currently, it seems clear that neo-genin is a receptor for hemojuvelin and thus may be animportant factor in the regulation of iron metabolism.
In our present study we further elucidate the specificsites of Hjv and Neo1 mRNA expression in mouse tis-sues using the quantitative and sensitive real-time RT-PCR method. Although four alternatively spliced exons
Figure 7 Immunohistochemical staining of neogenin in the mouse kidney and lung. The renal cortex shows a positive signal confined tothe glomeruli and some segments of the renal proximal and distal tubules (A). These reactions are not seen in the negative control (B).The neogenin signal in the lung is mainly located in round cells of the alveolar wall (arrows in C), whereas it is not detectable in the controlstaining (D). Tb, renal tubule; Gl, glomerulus.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
92 Rodriguez, Pan, Parkkila
by guest on September 21, 2014jhc.sagepub.comDownloaded from
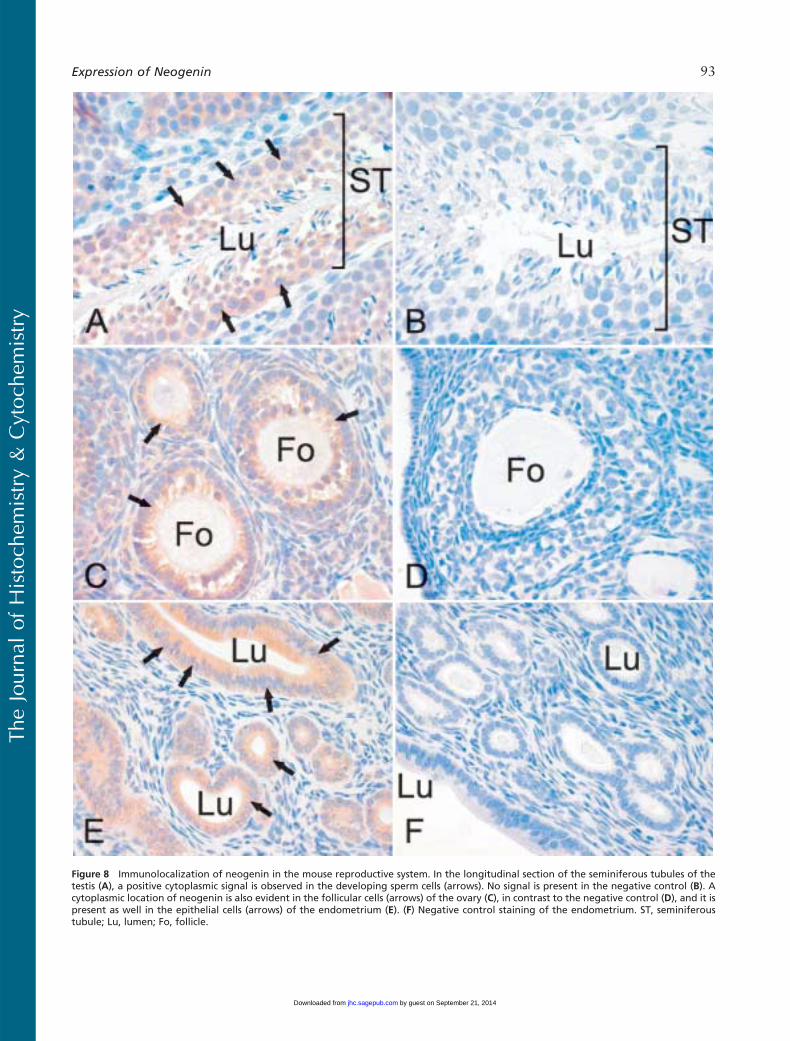
Figure 8 Immunolocalization of neogenin in the mouse reproductive system. In the longitudinal section of the seminiferous tubules of thetestis (A), a positive cytoplasmic signal is observed in the developing sperm cells (arrows). No signal is present in the negative control (B). Acytoplasmic location of neogenin is also evident in the follicular cells (arrows) of the ovary (C), in contrast to the negative control (D), and it ispresent as well in the epithelial cells (arrows) of the endometrium (E). (F) Negative control staining of the endometrium. ST, seminiferoustubule; Lu, lumen; Fo, follicle.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Expression of Neogenin 93
by guest on September 21, 2014jhc.sagepub.comDownloaded from
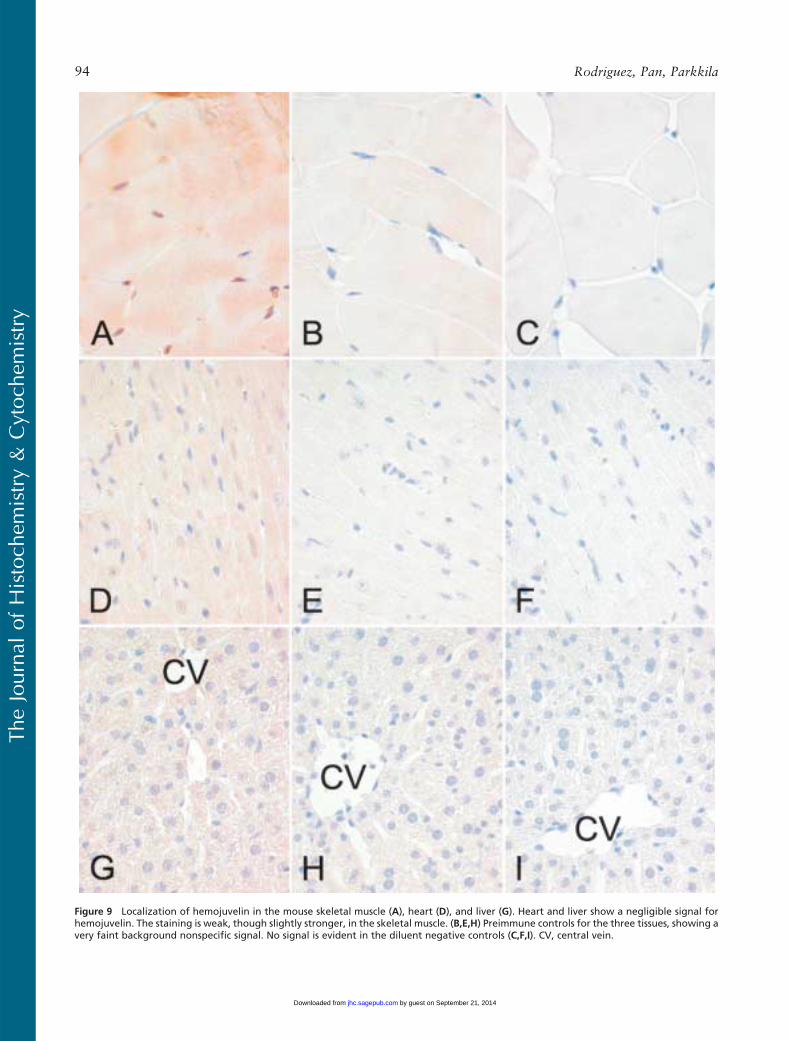
Figure 9 Localization of hemojuvelin in the mouse skeletal muscle (A), heart (D), and liver (G). Heart and liver show a negligible signal forhemojuvelin. The staining is weak, though slightly stronger, in the skeletal muscle. (B,E,H) Preimmune controls for the three tissues, showing avery faint background nonspecific signal. No signal is evident in the diluent negative controls (C,F,I). CV, central vein.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
94 Rodriguez, Pan, Parkkila
by guest on September 21, 2014jhc.sagepub.comDownloaded from

have been described for the mouse Neo1 gene (Keelinget al. 1997), our primer design ensured that there wouldbe no interference with any of these exons. Hence, ourassay detects all transcripts that are present in indi-vidual tissues. It is also noteworthy that our use of fourinternal control genes for the normalization of geneexpression levels enabled a higher degree of accuracy inour present study, compared with traditional normal-izations based on only one housekeeping gene. This isof special importance when treating heterogeneoussets of samples, in which case the use of a single internalcontrol may lead to erroneous conclusions.
Neo1 transcripts were detected in all tissue samplestested, with no notable variations observed betweenmale and female mice. Our results are in agreement witha previous report in which Neo1 expression was detect-able by Northern blotting in each of a smaller set ofsamples (Keeling et al. 1997). In addition, NEO1, thehuman ortholog of the mouse Neo1, was analyzedearlier by Northern blotting, with the strongest signalevident in skeletal muscle (Meyerhardt et al. 1997;Vielmetter et al. 1997). In our current experiments, thehighest levels of Neo1 mRNA expression were detect-able in the male and female reproductive systems (testis,ovary, and uterus). Accordingly, Neo1 had been shownpreviously to be present in the ovary by Northern blot-ting (Keeling et al. 1997). Northern analysis wasalso adopted in a study by Meyerhardt et al. (1997)that showed expression of NEO1 in testis and ovary.The role of neogenin and the identity of its ligand(s)in reproductive organs will certainly provide interestingnovel targets for future investigations.
We also explored the localization of the neogeninprotein in several mouse tissues using immunohisto-chemistry and found positive signals in the brain, skel-etal muscle, heart, liver, stomach, duodenum, ileum,colon, kidney, lung, testis, ovary, oviduct, and uterus.No neogenin protein was detectable in the spleen, thymus,or pancreas, and the absence of any protein product ismost likely due to a very low level of gene expression inthese tissues, as seen by quantitative PCR and to differ-ences in the sensitivities of the two methods.
Hjv mRNA expression profile was less extensivewhen compared with Neo1. As was observed for Neo1,differences between Hjv expression patterns in femaleand male tissues were small. Moreover, the expressionanalysis of murine Hjv mRNA reported herein is inagreement with previously reported Northern analysesof postnatal mice (Niederkofler et al. 2004). Previous insitu hybridization analyses in mouse embryos has alsoshown that Hjv is restricted to the skeletal and cardiacmuscle (Oldekamp et al. 2004) and, more recently, ithas been shown that mouse periportal hepatocytes ex-press hemojuvelin protein (Niederkofler et al. 2005).Earlier reports of human HJV expression profiles ob-tained by Northern blot and conventional RT-PCR are
also highly coincident with our present data (Papani-kolaou et al. 2004; Rodriguez Martinez et al. 2004).In hemojuvelin immunohistochemistry, the higheststaining reactions were detected in the skeletal muscle,which agrees well with the quantitative RT-PCR re-sults. It is notable that expression levels of HJV mRNAwere much lower as compared to neogenin. Thus, itis not surprising that hemojuvelin protein expressionseemed to be very low in murine tissues.
Neogenin has turned out to be a multifaceted re-ceptor with several ligands. The interaction betweenmembrane-associated hemojuvelin and neogenin canonly take place at sites where both molecules are ex-pressed, and we have determined these overlappingregions to be the skeletal muscle, heart, liver, andesophagus. Interestingly, the liver is also the major siteof hepcidin expression, which is considered to be adownstream target of hemojuvelin. Hepatocytes havebeen proposed to sense the iron status of the body andthen either release or downregulate hepcidin. Thus, theinteraction between hemojuvelin and neogenin mightplay a role in this sensing machinery. Although ourimmunohistochemical results indicated a very low ex-pression of both neogenin and hemojuvelin in hepa-tocytes, the tempting possibility remains that iron levelsor other factors could affect the expression and cellulardistribution of these iron regulatory proteins in theliver. On the other hand, in spite of the narrow he-mojuvelin expression profile that we identified in ourcurrent experiments, existence of a soluble form ofhemojuvelin implies that it can probably function inseveral organs and cell types, depending on the presenceof its receptor. The wide expression of neogenin alsosuggests that the hemojuvelin–neogenin pathway may,in fact, represent an important and extensive signalingcascade in the body.
Acknowledgments
This work was supported by grants from the SigridJuselius Foundation, Academy of Finland, and the FinnishCancer Foundation.
Literature Cited
Abboud S, Haile DJ (2000) A novel mammalian iron-regulatedprotein involved in intracellular iron metabolism. J Biol Chem275:19906–19912
Barallobre MJ, Pascual M, Del Rio JA, Soriano E (2005) The Netrinfamily of guidance factors: emphasis on Netrin-1 signalling. BrainRes Brain Res Rev 49:22–47
Bridle KR, Frazer DM, Wilkins SJ, Dixon JL, Purdie DM, CrawfordDH, Subramaniam VN, et al. (2003) Disrupted hepcidin regula-tion in HFE-associated haemochromatosis and the liver as a reg-ulator of body iron homoeostasis. Lancet 361:669–673
Camaschella C, Roetto A, Cali A, De Gobbi M, Garozzo G, CarellaM, Majorano N, et al. (2000) The gene TFR2 is mutated in anew type of haemochromatosis mapping to 7q22. Nat Genet 25:14–15
Feder JN, Gnirke A, Thomas W, Tsuchihashi Z, Ruddy DA, BasavaA, Dormishian F, et al. (1996) A novel MHC class I-like gene is
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Expression of Neogenin 95
by guest on September 21, 2014jhc.sagepub.comDownloaded from

mutated in patients with hereditary haemochromatosis. Nat Genet13:399–408
Fleming RE (2005) Advances in understanding the molecular basis forthe regulation of dietary iron absorption. Curr Opin Gastroenterol21:201–206
Kang JS, Yi MJ, Zhang W, Feinleib JL, Cole F, Krauss RS (2004)Netrins and neogenin promote myotube formation. J Cell Biol167:493–504
Keeling SL, Gad JM, Cooper HM (1997) Mouse neogenin, a DCC-like molecule, has four splice variants and is expressed widely inthe adult mouse and during embryogenesis. Oncogene 15:691–700
Le Gac G, Ferec C (2005) The molecular genetics of haemo-chromatosis. Eur J Hum Genet 13:1172–1185
Lee JE, Kim HJ, Bae JY, Kim SW, Park JS, Shin HJ, Han W, et al.(2005) Neogenin expression may be inversely correlated to thetumorigenicity of human breast cancer. BMC Cancer 5:154
Lin L, Goldberg YP, Ganz T (2005) Competitive regulation of hep-cidin mRNA by soluble and cell-associated hemojuvelin. Blood106:2884–2889
Matsunaga E, Tauszig-Delamasure S, Monnier PP, Mueller BK,Strittmatter SM, Mehlen P, Chedotal A (2004) RGM and its recep-tor neogenin regulate neuronal survival. Nat Cell Biol 6:749–755
Meyerhardt JA, Look AT, Bigner SH, Fearon ER (1997) Identi-fication and characterization of neogenin, a DCC-related gene.Oncogene 14:1129–1136
Monnier PP, Sierra A, Macchi P, Deitinghoff L, Andersen JS, MannM, Flad M, et al. (2002) RGM is a repulsive guidance moleculefor retinal axons. Nature 419:392–395
NemethE,RoettoA,GarozzoG,GanzT,CamaschellaC(2005)Hepci-din is decreased in TFR2 hemochromatosis. Blood 105:1803–1806
Nemeth E, Tuttle MS, Powelson J, Vaughn MB, Donovan A, WardDM, Ganz T, et al. (2004) Hepcidin regulates cellular iron ef-flux by binding to ferroportin and inducing its internalization.Science 306:2090–2093
Niederkofler V, Salie R, Arber S (2005) Hemojuvelin is essentialfor dietary iron sensing, and its mutation leads to severe iron over-load. J Clin Invest 115:2180–2186
Niederkofler V, Salie R, Sigrist M, Arber S (2004) Repulsive guidancemolecule (RGM) gene function is required for neural tube closure
but not retinal topography in the mouse visual system. J Neurosci24:808–818
Njajou OT, Vaessen N, Joosse M, Berghius B, van Dongen JW,Breuning MH, Snijders PJ, et al. (2001) A mutation in SLC11A3 isassociated with autosomal dominant hemochromatosis. Nat Genet28:213–214
Oldekamp J, Kramer N, Alvarez-Bolado G, Skutella T (2004)Expression pattern of the repulsive guidance molecules RGM A,B and C during mouse development. Gene Expr Patterns 4:283–288
Papanikolaou G, Samuels ME, Ludwig EH, MacDonald ML,Franchini PL, Dube MP, Andres L, et al. (2004) Mutations inHFE2 cause iron overload in chromosome 1q-linked juvenilehemochromatosis. Nat Genet 36:77–82
Pietrangelo A (2004) The ferroportin disease. Blood Cells Mol Dis32:131–138
Rodriguez Martinez A, Niemela O, Parkkila S (2004) Hepatic andextrahepatic expression of the new iron regulatory protein he-mojuvelin. Haematologica 89:1441–1445
Roetto A, Papanikolaou G, PolitouM, Alberti F, Girelli D, ChristakisJ, Loukopoulos D, et al. (2003) Mutant antimicrobial peptidehepcidin is associated with severe juvenile hemochromatosis. NatGenet 33:21–22
Schmidtmer J, Engelkamp D (2004) Isolation and expression patternof three mouse homologues of chick Rgm. Gene Expr Patterns 4:105–110
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, DePaepe A, Speleman F (2002) Accurate normalization of real-timequantitative RT-PCR data by geometric averaging of multipleinternal control genes. Genome Biol 3. Published online June 18,2002 (DOI: 10.1186/gb-2002-3-7-research0034)
Vielmetter J, Chen XN, Miskevich F, Lane RP, Yamakawa K,Korenberg JR, Dreyer WJ (1997) Molecular characterization ofhuman neogenin, a DCC-related protein, and the mapping of itsgene (NEO1) to chromosomal position 15q22.3-q23. Genomics41:414–421
Zhang AS, West AP Jr, Wyman AE, Bjorkman PJ, Enns CA (2005)Interaction of hemojuvelin with neogenin results in iron accumu-lation in human embryonic kidney 293 cells. J Biol Chem 280:33885–33894
TheJournal
ofHistoch
emistry&
Cytoch
emistry
96 Rodriguez, Pan, Parkkila
by guest on September 21, 2014jhc.sagepub.comDownloaded from




















