
Expression of Osteoclast Differentiation Signals by Stromal Elements of Giant Cell Tumors
10
Expression of Osteoclast Differentiation Signals by Stromal Elements of Giant Cell Tumors GERALD J. ATKINS, 1 DAVID R. HAYNES, 2 STEPHEN E. GRAVES, 1,3 ANDREAS EVDOKIOU, 1 SHELLEY HAY, 1 STELIOS BOURALEXIS, 1 and DAVID M. FINDLAY 1 ABSTRACT The mechanisms by which primary tumors of the bone cause bone destruction have not been elucidated. Unlike most other lytic bone tumors, osteoclastomas, otherwise known as giant cell tumors (GCT), contain osteoclast-like cells within the tumor stroma. A new member of the TNF-ligand superfamily member, osteoclast differentiation factor (ODF/OPGL/RANKL/TRANCE), was recently identified. ODF was shown to directly stimulate osteoclastogenesis, in the presence of M-CSF. In this study, the expression of ODF was examined in a number of tumor samples associated with bone lysis in vivo. In addition, we investigated expression of the ODF receptor on osteoclast precursors, RANK, as well as the ODF inhibitor osteoprotegerin (OPG), and another TNF-ligand superfamily member, TRAIL, previously shown to abrogate the inhibitory effects of OPG. We report here the novel finding that GCT stromal cells contain abundant ODF mRNA, whereas the giant cell population exclusively expresses RANK mRNA. These results are consistent with the osteoclast-mediated bone destruction by these tumors. We also report the expression of OPG and TRAIL mRNA in GCT samples. A comparison with other lytic and nonlytic tumors of bone showed that GCT express more ODF and TRAIL mRNA relative to OPG mRNA. In addition, GCT were found to express a number of cytokines previously reported to play central roles in osteoclastogenesis, namely, IL-1, -6, -11, -17, as well as TNF-a. Importantly, GCT were also found to express high levels of M-CSF mRNA, a cytokine shown to be an essential cofactor of ODF, and a survival factor for mature and developing osteoclasts. Furthermore, expression of these molecules by stromal cells isolated from GCT continued in vitro. Thus GCT constitutively express all of the signals that are currently understood to be necessary for the differentiation of osteoclasts from precursor cells. (J Bone Miner Res 2000;15:640 – 649) Key words: giant cell tumor, ODF, TRAIL, OPG, bone tumor INTRODUCTION G IANT CELL TUMORS (GCT), otherwise known as oste- oclastomas, are rare primary neoplasms of the skeleton, associated with extensive localized bone loss. GCT contain within the tumor mass variable numbers of large, multinu- cleated cells that are phenotypically indistinguishable from osteoclasts, (1) the cell type involved in bone resorption during normal homeostasis of the skeleton. It is believed that the stromal cells are the tumor cells that induce oste- oclastic bone resorption by recruiting osteoclast precursors and promoting their differentiation into functional oste- oclasts. (2) The mechanisms by which they do so are not understood. Recently, stromal cells of GCT have been 1 Department of Orthopaedics and Trauma, University of Adelaide, Adelaide, South Australia 5000, Australia. 2 Department of Pathology, University of Adelaide, Adelaide, South Australia 5000, Australia. 3 Department of Orthopaedic Surgery, Flinders Medical Centre and Repatriation General Hospital, Daw Park, South Australia 5041, Australia. JOURNAL OF BONE AND MINERAL RESEARCH Volume 15, Number 4, 2000 © 2000 American Society for Bone and Mineral Research 640
Transcript of Expression of Osteoclast Differentiation Signals by Stromal Elements of Giant Cell Tumors

Expression of Osteoclast Differentiation Signals byStromal Elements of Giant Cell Tumors
GERALD J. ATKINS,1 DAVID R. HAYNES,2 STEPHEN E. GRAVES,1,3 ANDREAS EVDOKIOU,1
SHELLEY HAY,1 STELIOS BOURALEXIS,1 and DAVID M. FINDLAY 1
ABSTRACT
The mechanisms by which primary tumors of the bone cause bone destruction have not been elucidated.Unlike most other lytic bone tumors, osteoclastomas, otherwise known as giant cell tumors (GCT), containosteoclast-like cells within the tumor stroma. A new member of the TNF-ligand superfamily member,osteoclast differentiation factor (ODF/OPGL/RANKL/TRANCE), was recently identified. ODF was shown todirectly stimulate osteoclastogenesis, in the presence of M-CSF. In this study, the expression of ODF wasexamined in a number of tumor samples associated with bone lysis in vivo. In addition, we investigatedexpression of the ODF receptor on osteoclast precursors, RANK, as well as the ODF inhibitor osteoprotegerin(OPG), and another TNF-ligand superfamily member, TRAIL, previously shown to abrogate the inhibitoryeffects of OPG. We report here the novel finding that GCT stromal cells contain abundant ODF mRNA,whereas the giant cell population exclusively expresses RANK mRNA. These results are consistent with theosteoclast-mediated bone destruction by these tumors. We also report the expression of OPG and TRAILmRNA in GCT samples. A comparison with other lytic and nonlytic tumors of bone showed that GCT expressmore ODF and TRAIL mRNA relative to OPG mRNA. In addition, GCT were found to express a number ofcytokines previously reported to play central roles in osteoclastogenesis, namely, IL-1, -6, -11, -17, as well asTNF-a. Importantly, GCT were also found to express high levels of M-CSF mRNA, a cytokine shown to bean essential cofactor of ODF, and a survival factor for mature and developing osteoclasts. Furthermore,expression of these molecules by stromal cells isolated from GCT continued in vitro. Thus GCT constitutivelyexpress all of the signals that are currently understood to be necessary for the differentiation of osteoclastsfrom precursor cells. (J Bone Miner Res 2000;15:640–649)
Key words: giant cell tumor, ODF, TRAIL, OPG, bone tumor
INTRODUCTION
GIANT CELL TUMORS (GCT), otherwise known as oste-oclastomas, are rare primary neoplasms of the skeleton,
associated with extensive localized bone loss. GCT containwithin the tumor mass variable numbers of large, multinu-cleated cells that are phenotypically indistinguishable from
osteoclasts,(1) the cell type involved in bone resorptionduring normal homeostasis of the skeleton. It is believedthat the stromal cells are the tumor cells that induce oste-oclastic bone resorption by recruiting osteoclast precursorsand promoting their differentiation into functional oste-oclasts.(2) The mechanisms by which they do so are notunderstood. Recently, stromal cells of GCT have been
1Department of Orthopaedics and Trauma, University of Adelaide, Adelaide, South Australia 5000, Australia.2Department of Pathology, University of Adelaide, Adelaide, South Australia 5000, Australia.3Department of Orthopaedic Surgery, Flinders Medical Centre and Repatriation General Hospital, Daw Park, South Australia 5041,
Australia.
JOURNAL OF BONE AND MINERAL RESEARCHVolume 15, Number 4, 2000© 2000 American Society for Bone and Mineral Research
640

shown to release chemokines such as macrophage chemoat-tractant protein 1(3) and interleukin-8,(4) which could recruitprecursors of the monocytic lineage into the stromal mass ofthe tumor, before their development into mature osteoclasts.
Considerable progress has been made toward an under-standing of the mechanisms responsible for physiologicalosteoclastogenesis. A large number of hormones and cyto-kines have been identified that can exert direct and indirectstimulatory and antagonisitic effects on the development ofosteoclasts from hematopoietic precursors.(5) Principalamong these are interleukin (IL) -1, -6, -11, -17, and -18;macrophage colony stimulating factor (M-CSF); PTHrP;and TNFa, as well as prostaglandin E2 (PGE2).
Most recently, a cell-surface TNF-ligand family member,termed osteoclast differentiation factor (ODF), previouslyidentified as TRANCE(6) and RANKL,(7) was shown to becentral to osteoclast development.(8) An engineered solubleform of ODF was shown to promote osteoclast developmentwhen added to peripheral blood mononuclear cells (PBMC)in stromal-free cultures, with the obligatory presence ofM-CSF.(8) ODF binds to a previously characterized TNFreceptor superfamily member, RANK,(9) on osteoclast pre-cursors, and ligation with ODF has recently been shown toactivate NF-kB by TNF-associated factors (TRAF) 2, 5, and6.(10) A soluble TNF-receptor family member termed osteo-protegerin (OPG), which can inhibit osteoclast formationand bone resorption, was shown to be an ODF antago-nist.(11) Overexpression of OPG in mice resulted in a mutedosteopetrotic phenotype, with reduced cancellous and in-creased cortical bone, but normal elongation and develop-ment of the growth plate.(11) Conversely, OPG-knockoutmice developed extensive osteoporosis associated with anincreased number of otherwise normal osteoclasts.(12) Like-wise, deletion of the ODF gene resulted in severe osteope-trosis and a complete lack of osteoclasts as a result of aninability of osteoblasts to support osteoclastogenesis.(13)
Another recently described member of the TNF-ligandsuperfamily, TRAIL/Apo2L,(14) has been shown to play arole in triggering apoptosis of cells bearing at least one oftwo receptors, DR4 and DR5. Two other receptors forTRAIL/Apo2L, DcR1 and DcR2, act as decoy receptors,protecting cells from apoptosis that also express one ormore of DcR1 and DcR2.(15) TRAIL has also been shown tobind to OPG, reversing its inhibitory effect on osteoclasto-genesis. Conversely, OPG is capable of inhibiting the apo-ptotic effect of TRAIL.(16) A role for TRAIL in physiolog-ical or pathological bone resorption has not been reported.
In light of these recently discovered molecules, it isplausible that their net local concentrations act to targetosteoclast formation in the microenvironment of the bone,and that their relative concentrations, in turn, are regulatedby circulating and local regulators of bone resorption, suchas PGE2 and 1a,25(OH)2 vitamin D3.
(17) In solid lytictumors of the bone, the cytokines and growth factors pro-duced by the tumor cells may disrupt the normal homeo-static balance of factors in the bone microenvironment,leading to increased local osteoclastogenesis and disregula-tion of osteoclastic activity. Alternatively, the GCT tumorcells themselves may express ODF and so directly supportosteoclast formation.
We hypothesized that the stromal element of GCT ex-presses these newly discovered osteoclastogenic molecules,which could account for the presence of osteoclasts withinthe tumor mass in vivo, and the levels of expression of thesepositive and negative mediators may account for the degreeof bone lysis observed in patients with GCT. We haveidentified within GCT samples expression of high levels ofmRNA encoding ODF, RANK, and TRAIL. In a smallseries of tumors, we found a relationship between expres-sion of the relative levels of ODF and TRAIL, in terms ofthe corresponding level of OPG, with the degree of bonelysis by these tumors in vivo. We provide evidence thatGCT tumor cells provide all of the necessary differentiationsignals required to support osteoclastogenesis consistentwith the pathological destruction of bone associated withthese tumors.
MATERIALS AND METHODS
Tumor classification and isolation
The diagnosis of all tumors was established by biopsybefore surgical management, and subsequently confirmedby repeat histology of the resected specimen. The clinicalclassification and bone lytic activity of all tumors used inthis study are shown in Table 1. The giant cell tumor (GCT)samples included tumors representative of highly aggres-sive, moderately aggressive, and mildly aggressive types,based on clinical assessment and radiological analysis (Ta-ble 1). The clinical assessment included time since onset ofsymptoms, tumor size, extent of bone involvement, and thedegree of extension into the soft tissue. The portion of eachtumor selected for analysis was isolated from within thetumor mass, but excluded the central necrotic tissue andnormal tissue surrounding the tumor.
Processing of tumor samples
Tumor samples were processed within 24 h of surgicalremoval. After being washed three times in sterile PBS,tissue was dissected in a petri dish using a scalpel blade,then either used directly for isolation of total RNA (seebelow) or enzymically digested for cell culture. For diges-tion, tissue was resuspended in 1 ml each of collagenase anddispase (10 mg/ml) and incubated for 1 h at37°C. After this,the cell suspension was diluted 10-fold inaMEM mediumcontaining 10% v/v FCS, filtered through a 40-mm cellstrainer (Becton Dickinson, Franklin Lakes, NJ, U.S.A.),and then washed twice in the same medium. Cells were theneither cryopreserved in a 20% solution of DMSO in mediumcontaining 20% FCS or cultured inaMEM medium con-taining 10% FCS.
Subfractionation of GCT populations
GCT samples were used freshly or cryopreserved GCTsamples were thawed and cultured for several days inDMEM medium containing 10% FCS in 75-cm2 flasks.Giant cells appeared in the stromal cell mass during thistime. Stromal cells were removed by brief trypsin digestion,
641OSTEOCLASTOGENESIS IN GIANT CELL TUMORS

achieved by rinsing the cells once in sterile PBS before theaddition of 2 ml trypsin solution (0.1% w/v in PBS), pre-heated to 37°C. After 30 s, most of the mononuclear stromalcells had detached, leaving giant cells and fan-shapedmononuclear cells attached. The trypsin was inactivated bythe addition of 8 ml DMEM medium containing 10% FCS,and the stromal cells were then removed and placed into afresh culture flask, maintaining a high-cell density. Theremaining giant cells were subsequently referred to asgiantcell–enrichedcultures and were processed immediately forisolation of total RNA (see below). After 2–4 days, furthergiant cells appeared in the replated stromal mass and thepreceding process was repeated. The subsequent stromalcell culture did not give rise to further giant cells.
Preparation of total RNA
Dissected tumor samples were dissolved in Trizol reagent(Life Technologies, Gaithersburg, MD, U.S.A.) at 2 ml/5mm2 of original tissue. Freshly digested GCT samples orgiant cell–enriched cultures and stromal cultures derivedfrom successive rounds of the brief trypsinization protocol,described earlier, were lysed in 1 ml/13 106 cells of Trizolreagent. Total RNA was prepared from the dissolved tissuesaccording to the manufacturer’s instructions.
RT-PCR analysis
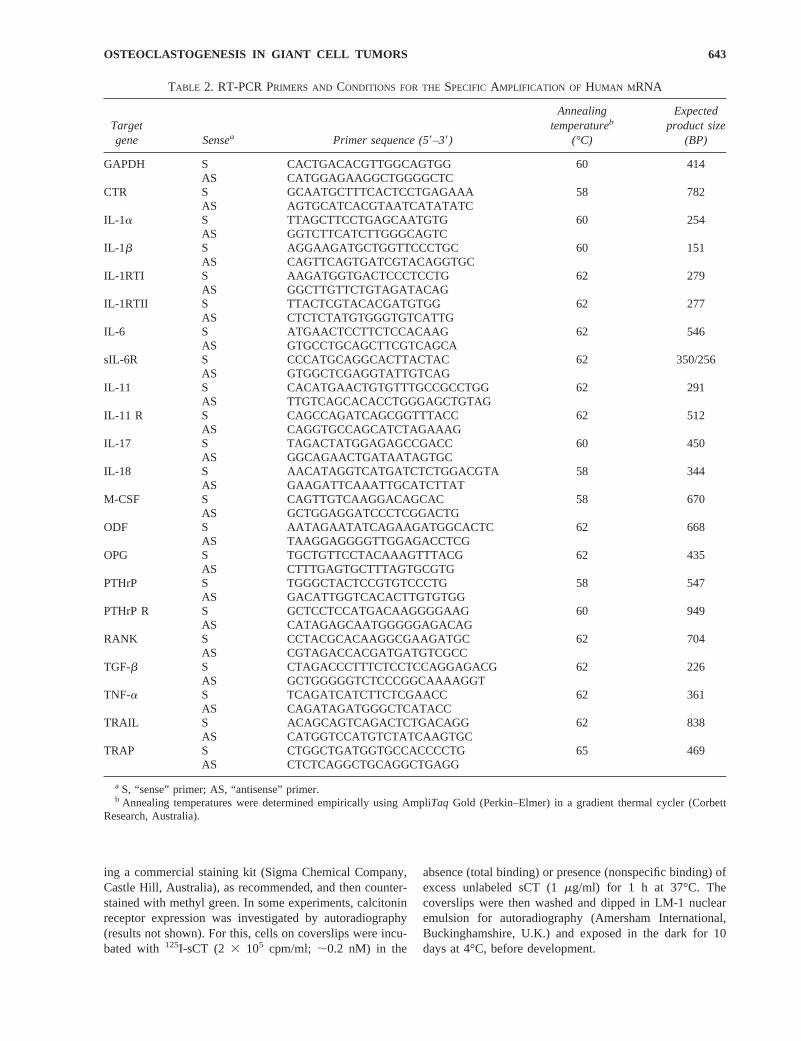
First-strand complementary DNA (cDNA) was synthe-sized from 1.6mg of total RNA from each sample, using acDNA synthesis kit, according to the manufacturer’s in-structions (Promega Corp., Madison, WI, U.S.A.). cDNAwas then amplified by PCR to generate products corre-sponding to mRNA encoding human gene products listed inTable 2. The 20-ml amplification mixture contained 1 U ofAmpliTaqGold DNA polymerase (Perkin–Elmer, Norwalk,CT, U.S.A.), 100 ng each of the 59 and 39 primers, 0.2 mMdNTPs (Pharmacia Biotech, Uppsala, Sweden), 1.5 mMMgCl2, 2 ml 103 reaction buffer, and sterile DEPC-H2O.
PCR was performed for 23 cycles for GAPDH and 30–35cycles for other primer pairs, such that all products could beassayed in the exponential phase of the amplification curve,in a thermal cycler (Corbett Research, Melbourne, Victoria,Australia). After an initial step at 95°C for 9 minutes toactivate the polymerase, each cycle consisted of 1 minute ofdenaturation at 94°C, 1 minute of annealing at the temper-atures indicated in Table 2, and 1 minute of extension at72°C. This was followed by an additional extension step at72°C for 1 minute. We designed human sequence-specificoligonucleotide primers on the basis of published sequences(Life Technologies). In all cases, the primers were mRNA-specific, in that the recognition sites of the upstream anddownstream primers resided in separate exons or at intron/exon boundaries in the genomic sequence. Primer se-quences and predicted PCR product sizes are shown inTable 2. Amplification products were resolved by electro-phoresis on a 2% w/v agarose gel and poststained withSYBR-1 Green (Molecular Probes, Eugene, OR, U.S.A.).The relative amounts of the PCR products were determinedby quantitating the intensity of bands using a FluorImagerand ImageQuant software (Molecular Dynamics, Sunny-vale, CA, U.S.A.). Amplified products corresponding toODF, OPG, RANK, and TRAIL mRNA are represented asa ratio of the respective PCR product/GAPDH PCR product.To show that there were no false-positive results, PCRreactions were carried out using non-reverse-transcribedRNA, and on reaction mixtures in which no RNA wasadded. The specificity of the PCR reaction was confirmedby Southern transfer onto a nylon membrane (Hybond-N;Amersham Life Science Inc., Arlington Heights, IL,U.S.A.) and hybridization with digoxigenin (DIG)-labeledinternal oligonucleotide probes (Table 1).
TRAP staining and calcitonin receptorautoradiography
Tumor cells cultured on glass coverslips were fixed andstained for tartrate-resistant acid phosphatase (TRAP), us-
TABLE 1. TUMOR SAMPLES USED IN THIS STUDY, SHOWING CLINICAL CLASSIFICATION, LYTIC PROPERTIES OFTUMOR IN THE
BONE, AND ASSESSMENT OFGCT AGGRESSIVENESS, AS DEFINED BY CRITERIA DESCRIBED IN THE TEXT
Code Classification Bone lytic activity Aggressiveness
GCT-A giant cell tumor lytic highGCT-B giant cell tumor lytic moderateGCT-C giant cell tumor lytic highGCT-D giant cell tumor lytic mildGCT-E giant cell tumor lytic moderateGCT-F giant cell tumor lytic highGCT-G giant cell tumor lytic highGCT-H giant cell tumor lytic moderateNGC-B plasmacytoma lytic N.A.a
NGC-C osteogenic sarcoma mixed lytic/osteogenic N.A.NGC-D extraosseus chondrosarcoma not involving bone N.A.NGC-F osteoblastoma nonlytic N.A.NGC-G osteogenic sarcoma nonlytic N.A.
a N.A. 5 not applicable.
642 ATKINS ET AL.
ing a commercial staining kit (Sigma Chemical Company,Castle Hill, Australia), as recommended, and then counter-stained with methyl green. In some experiments, calcitoninreceptor expression was investigated by autoradiography(results not shown). For this, cells on coverslips were incu-bated with125I-sCT (2 3 105 cpm/ml; ;0.2 nM) in the
absence (total binding) or presence (nonspecific binding) ofexcess unlabeled sCT (1mg/ml) for 1 h at 37°C. Thecoverslips were then washed and dipped in LM-1 nuclearemulsion for autoradiography (Amersham International,Buckinghamshire, U.K.) and exposed in the dark for 10days at 4°C, before development.
TABLE 2. RT-PCR PRIMERS AND CONDITIONS FOR THESPECIFIC AMPLIFICATION OF HUMAN M RNA
Targetgene Sensea Primer sequence (59–39)
Annealingtemperatureb
(°C)
Expectedproduct size
(BP)
GAPDH S CACTGACACGTTGGCAGTGG 60 414AS CATGGAGAAGGCTGGGGCTC
CTR S GCAATGCTTTCACTCCTGAGAAA 58 782AS AGTGCATCACGTAATCATATATC
IL-1a S TTAGCTTCCTGAGCAATGTG 60 254AS GGTCTTCATCTTGGGCAGTC
IL-1b S AGGAAGATGCTGGTTCCCTGC 60 151AS CAGTTCAGTGATCGTACAGGTGC
IL-1RTI S AAGATGGTGACTCCCTCCTG 62 279AS GGCTTGTTCTGTAGATACAG
IL-1RTII S TTACTCGTACACGATGTGG 62 277AS CTCTCTATGTGGGTGTCATTG
IL-6 S ATGAACTCCTTCTCCACAAG 62 546AS GTGCCTGCAGCTTCGTCAGCA
sIL-6R S CCCATGCAGGCACTTACTAC 62 350/256AS GTGGCTCGAGGTATTGTCAG
IL-11 S CACATGAACTGTGTTTGCCGCCTGG 62 291AS TTGTCAGCACACCTGGGAGCTGTAG
IL-11 R S CAGCCAGATCAGCGGTTTACC 62 512AS CAGGTGCCAGCATCTAGAAAG
IL-17 S TAGACTATGGAGAGCCGACC 60 450AS GGCAGAACTGATAATAGTGC
IL-18 S AACATAGGTCATGATCTCTGGACGTA 58 344AS GAAGATTCAAATTGCATCTTAT
M-CSF S CAGTTGTCAAGGACAGCAC 58 670AS GCTGGAGGATCCCTCGGACTG
ODF S AATAGAATATCAGAAGATGGCACTC 62 668AS TAAGGAGGGGTTGGAGACCTCG
OPG S TGCTGTTCCTACAAAGTTTACG 62 435AS CTTTGAGTGCTTTAGTGCGTG
PTHrP S TGGGCTACTCCGTGTCCCTG 58 547AS GACATTGGTCACACTTGTGTGG
PTHrP R S GCTCCTCCATGACAAGGGGAAG 60 949AS CATAGAGCAATGGGGGAGACAG
RANK S CCTACGCACAAGGCGAAGATGC 62 704AS CGTAGACCACGATGATGTCGCC
TGF-b S CTAGACCCTTTCTCCTCCAGGAGACG 62 226AS GCTGGGGGTCTCCCGGCAAAAGGT
TNF-a S TCAGATCATCTTCTCGAACC 62 361AS CAGATAGATGGGCTCATACC
TRAIL S ACAGCAGTCAGACTCTGACAGG 62 838AS CATGGTCCATGTCTATCAAGTGC
TRAP S CTGGCTGATGGTGCCACCCCTG 65 469AS CTCTCAGGCTGCAGGCTGAGG
a S, “sense” primer; AS, “antisense” primer.b Annealing temperatures were determined empirically using AmpliTaq Gold (Perkin–Elmer) in a gradient thermal cycler (Corbett
Research, Australia).
643OSTEOCLASTOGENESIS IN GIANT CELL TUMORS

Identification of resorption pit formation
To assess the extent of lacunar bone resorption by GCT,freshly isolated or freshly thawed cells were plated ontodiscs of whale dentine and cultured for 4 days inaMEMmedium containing 10% FCS. The dentine was then treatedwith 25% ammonia solution for 15 minutes, ultrasonicatedfor 5 minutes, followed by 15 minutes in trypsin at 37°Cand a final 5 minutes’ sonication to remove the adherentcells. The dentine was then dehydrated by passage throughgraded alcohol solutions from 70 to 100%, then dried undervacuum in a desiccator overnight before being mounted onstubs and carbon-coated for visualization using a PhilipsXL-20 scanning electron microscope (SEM).
RESULTS
Expression of osteoclast markers
Tumor samples (Table 1) were surgically removed andprocessed as described earlier. Cells isolated from giant celltumors were routinely plated onto whale dentine slices andin all cases formed resorption lacunae within 48–72 h ofplating. Cultured osteoclastoma cells were examined for theexpression of calcitonin receptor (CTR), which is expressedin bone exclusively by osteoclasts. Consistent with previousreports,(18) only multinucleated giant cells within the tumorbound125I-labeled sCT, whereas the stromal cells did not(not shown). RT-PCR also showed expression of the twopredominant isoforms of the human CTR(19) in the giantcell–enriched fraction of segregated tumor cells (Fig. 2).Cultures of dispersed GCT cells, both unfractionated andfractionated, were stained for TRAP. Both giant cells andsome mononuclear cells stained strongly positive for TRAP.In many of the stromal cells in unfractionated tumors,punctate staining for TRAP was observed, which may in-dicate internalization of exogenous TRAP by the stroma(Fig. 1). In addition, strong PCR signals for the osteoclastmarkers TRAP, carbonic anhydrase-II (CA-II) and cathep-sin K, were routinely derived from all GCT samples testedand were predominantly expressed by the giant cell–enriched fractions of segregated tumor samples (Fig. 2).
Expression of hematopoietic cytokine mRNA
Fractionated stromal and giant cell–enriched populationsof two GCT samples were tested for expression of a rangeof hematopoietic cytokines that have been implicated inosteoclastogenesis (Fig. 2). The tumorigenic stromal cells ofGCT-E were found to express mRNA encoding a number ofosteoclastogenic cytokines, including IL-6, IL-11, IL-17,PTHrP, and TGF-b. In addition, the stromal cells expressedthe functional IL-1 receptor type I (IL-1RTI). The stromalcells also expressed mRNA encoding M-CSF, an essentialcofactor in osteoclastogenesis.(8) The giant cell–enrichedfraction of GCT-E, however, expressed abundant IL-1a and-b, the nonfunctional decoy receptor for IL-1 (IL-1RTII),soluble IL-6 receptor (sIL-6R), TNF-a, GM-CSF, and theinhibitory IL-18. There were some differences in the ex-pression pattern observed between the tumors. For example,
the stromal cells of GCT-A expressed abundant mRNA forboth forms of IL-1, sIL-6R, IL-11R, IL-18, and TNF-a, inaddition to those described for GCT-E (data not shown). Wecannot rule out that the expression of the above-mentionedcytokines detected in the giant cell–enriched fractions ofGCT may represent minor contamination of this fractionwith stromal cells. Because the purpose of this study was toinvestigate expression of cytokines in the tumorigenic stro-mal element in particular, we did not attempt to removecompletely all of the stromal cells from the strongly adher-ent giant cells. The continued expression of the cytokineprofile, described here by GCT samples grown in vitro forup to 2 weeks, suggests that the stromal element of GCT iscapable of providing many of the recognized growth andsurvival factors for osteoclasts, in the absence of normalosteoblast and bone marrow stroma.
Expression of ODF, OPG, RANK, and TRAIL mRNA
ODF has been shown to be a central factor in osteoclastdevelopment. We were able to detect abundant ODF
FIG. 1. Photomicrographs showing giant cell tumor cells(GCT-A) cultured in vitro for 1 week and stained for TRAP.(A) Shows the unfractionated tumor and (B) shows anenriched population of giant cells after tryptic removal ofthe stromal cell layer, as described in Materials and Meth-ods.
644 ATKINS ET AL.

mRNA by RT-PCR in unfractionated GCT samples (Fig.3). To our knowledge, this is the first report of ODFexpression in any tumor type. To ascertain which celltype was likely to express which molecule, we fraction-ated GCT samples into preparations of stromal cellsalone and cultures highly enriched for giant cells. Asshown for GCT-E in Fig. 4, ODF mRNA was detected inthe stromal fraction. ODF mRNA expression was weaklydetectable in the giant cell– enriched fraction, althoughthis may be the result of contamination with stromalcells. Similar results were obtained in fractionated pop-ulations of GCT-A (data not shown).
The receptor for ODF on osteoclast precursor cells is anovel member of the TNF receptor superfamily,RANK.(9) RANK mRNA was found to be present in theprimary GCT samples (Fig. 3). Subfractionation of theGCT showed that RANK mRNA was exclusively presentin the giant cell– enriched fraction (Fig. 4), consistentwith a role for ODF/RANK ligation in osteoclast functionand survival.
We also examined the GCT for expression of anotherrecently described member of the TNF ligand superfam-ily, TRAIL/Apo2L. (14) This molecule has been found toplay a role in tumor apoptosis.(15) In addition, TRAIL canbind to OPG, reversing its inhibitory effect on osteoclas-togenesis. Conversely, OPG is capable of inhibiting theapoptotic effect of TRAIL.(16) We found high-level ex-pression of TRAIL mRNA in all GCT samples tested(Fig. 3), suggesting a functional role for TRAIL, perhaps
binding OPG and increasing the available ODF for liga-tion to RANK on the osteoclast precursors. TRAILmRNA was detected in both giant cell and stromal frac-tions of segregated GCT (Fig. 4).
Clinical correlation
It has been proposed that the ratio of ODF:OPG in thebone microenvironment may be an indicator of osteoclas-togenic potential. We were therefore interested to comparethis ratio between a number of GCT samples and also tocompare this parameter with the extent of observed osteol-ysis associated with particular tumors. In this small series oftumors, clear differences in the expression of ODF, OPG,TRAIL, and RANK mRNA between tumor types wereobservable. All giant cell tumors examined exhibited highlevels of ODF mRNA, which were generally greater thanobserved with the other tumors examined, including a lyticosteogenic sarcoma, an osteoblastoma, a sclerotic osteo-genic sarcoma, and an extraosseus chondrosarcoma (Fig.3A). A lytic plasmacytoma also expressed appreciable ODFmRNA (Fig. 3A). mRNA encoding TRAIL was also gen-erally abundant in the GCT samples and was less abundantin the non-GCT samples. The lowest levels of TRAILmRNA were recorded in the chondrosarcoma and the scle-rotic osteogenic sarcoma. High levels of both ODF andTRAIL are likely to provide an environment that stimulatesresorption, ODF increasing osteoclast differentiation and
FIG. 2. Hematopoietic cy-tokine expression by fraction-ated subpopulations of tumorstroma and giant cell–enriched (GC-enriched) pop-ulations of GCT-E. RT-PCRproducts were loaded onto2% agarose gels, electropho-resed, and stained withSYBR-1 Green. PCR prod-ucts were then visualized us-ing a FluorImager (MolecularDynamics), as described inMaterials and Methods.
645OSTEOCLASTOGENESIS IN GIANT CELL TUMORS

activation, and TRAIL, perhaps reducing the local effect ofOPG; therefore, high levels of either of these relative toOPG could potentially be a marker of tumor aggressiveness.We quantitated the levels of ODF and TRAIL mRNArelative to OPG mRNA and expressed these as their respec-tive ratios (Fig. 3D). As a group, the GCT exhibited higher
ratios of ODF:OPG than did the non-GCT tested. Generally,the ratios of TRAIL:OPG were higher for the GCT than thenon-GCT. Although larger numbers of tumors will need tobe examined to confirm these apparent clinical correlations,these data suggest that levels of these mRNA species mayprove useful in tumor identification.
FIG. 3. (A) RT-PCR anal-ysis of freshly resected tu-mor samples for expressionof mRNA species encodingGAPDH, ODF, OPG,RANK, and TRAIL. RT-PCR products were loadedonto 2% agarose gels, elec-trophoresed, and stainedwith SYBR-1 Green. PCRproducts were then visual-ized using a FluorImager(Molecular Dynamics), asdescribed in Materials andMethods. Quantitation ofRT-PCR products, shown in(A), expressed as (B) ratiosof ODF and OPG mRNAsnormalized to the level ofGAPDH mRNA, (C) ratiosof TRAIL and OPG mRNAnormalized to GAPDHmRNA, and (D) ratios ofODF and TRAIL mRNAsrelative to the level of OPGmRNA. * represents actualnumerical value of relativeexpression.
646 ATKINS ET AL.

DISCUSSION
ODF has been shown to be a central factor in osteoclastdevelopment, although to date, little information is availableregarding its role in pathological bone destruction by tu-mors. To our knowledge, this is the first report of theexpression of ODF in a pathological/oncological context.We examined a number of GCT, as well as other non-GCTof the bone associated with or without bone lysis, for theirexpression of ODF, RANK, OPG and TRAIL mRNA. Wewere able to detect abundant ODF mRNA by RT-PCR in allunfractionated GCT samples tested. Fractionation of oneGCT sample into stromal and giant cell fractions indicatedthat ODF mRNA expression is predominant in the stromalelement of the GCT (Fig. 4). The stromal elements of GCT,therefore, are equipped with a fundamental requirement forosteoclast development. In a physiological setting, the po-tency of ODF in its ability to stimulate osteoclast develop-ment is probably balanced by the local concentrations of its
inhibitor OPG, which is a potent inhibitor of osteoclasto-genesis.(11) It has been proposed that the ratio of ODF:OPGin the bone microenvironment may determine the localosteoclastogenic potential. Recently, the levels of ODFmRNA relative to OPG mRNA were shown to correlatewith the previously observed osteoclastogenic potential ofseveral osteoblastic cell lines.(20) We were interested tocompare this ratio, between a number of human bone tu-mors (including GCT and lytic and nonlytic non-GCT),with the extent of observed osteolysis in the correspondingindividuals. All GCT samples tested showed high ratios ofODF:OPG when compared with other nonlytic tumors (Fig.3B). The highest ratio of ODF:OPG was found in thestromal fraction of separated GCT, although our data do notrule out that giant cells themselves express ODF mRNA(Fig. 4).
The receptor for ODF on osteoclast precursor cells is anovel member of TNF receptor superfamily, RANK.(7,9)
Although mRNA for this molecule does not always corre-late with expression of protein at the cell surface,(7) abun-dant RANK mRNA was found to be expressed in most ofthe primary GCT samples tested. Subfractionation of theGCT showed that most RANK mRNA was present in thegiant cell–enriched fraction, consistent with its presence inthe hematopoietic element of the tumor, and with a role forODF/RANK ligation in osteoclast function and survival.(9)
In stroma passaged sufficiently to remove any further giantcells, we could no longer detect mRNA for RANK, sug-gesting the exclusive expression of RANK mRNA by giantcells in these tumors. Supportive of the above-mentionedconcepts, very little RANK mRNA was found in the non-lytic non-GCT examined here, whereas appreciable levelswere found in one of two lytic non-GCT.
Another recently cloned TNF-ligand family member,termed TRAIL, was characterized initially for its role inapoptosis.(15) TRAIL has been shown to be an antagonist ofOPG and can reverse the inhibitory effect of OPG onosteoclast formation.(16) The expression of TRAIL in thebone and its role in normal osteoclast physiology has notbeen established. We found high levels of expression ofTRAIL mRNA in all GCT samples tested (Fig. 3A), sug-gesting a functional role for TRAIL, perhaps binding localOPG and increasing the available ODF for ligation toRANK on the osteoclast precursors. In fractionated tumorsamples, TRAIL mRNA was clearly expressed by the stro-mal cells and equally strong signals for TRAIL were de-tected in the GC-enriched fraction (Fig. 4). The two lyticnon-GCT samples examined here expressed intermediatelevels of TRAIL mRNA, whereas the two nonlytic GCTexpressed the least. It will be of interest to examine theexpression of other TRAIL receptors(15) in these samples, toexplore a potential apoptotic/survival role for them in thesetumors.
In terms of ratios of ODF, TRAIL, and RANK mRNA toOPG mRNA, there was a clear distinction between giantcell tumors and other lytic tumors and tumors not associatedwith bone destruction. The nonlytic tumors did not expressabundant mRNA for ODF, RANK, or TRAIL, but didexpress abundant mRNA for the ODF inhibitor OPG. Thelytic non-GCT had increased levels of mRNA for ODF and
FIG. 4. (A) Expression of ODF, OPG, RANK, andTRAIL mRNA in the fractionated subpopulations of tumorGCT-E, and (B) quantitation of these with respect to thelevels of mRNA encoding the housekeeping gene GAPDH.
647OSTEOCLASTOGENESIS IN GIANT CELL TUMORS

TRAIL to OPG mRNA ratios but they remained markedlyless than what is observed for GCT. It is possible that theproduction of these factors in cases of non-GCT is a resultof stimulation of normal stroma, whereas in the giant celltumor it is thought that the stroma is tumorigenic, and islikely to be constitutively producing these mRNA species asa function of the transformed phenotype. It will be ofinterest to explore the molecular mechanisms responsiblefor this, particularly the role of potential autocrine factorsalso produced by the tumor cells, such as IL-1/IL-1RTI andIL-6/sIL-6R. Elevated PTHrP expression has been impli-cated in the molecular mechanism responsible for the es-tablishment and osteolysis associated with breast(21) andprostate(22) skeletal metastases, and the hypercalcemia ofhumoral malignancy such as multiple myeloma.(23) Interest-ingly, however, we observed low levels of PTHrP mRNA inthis study, suggesting that PTHrP may not play a major rolein GCT.
The expression of ODF by the stromal fraction of GCT isconsistent with the current body of evidence that suggeststhat these cells are mesenchymal in origin and osteoblast-like. Previously, it was shown that the stromal cells could beinduced to form mineralized nodules in vitro and formationof bone in SCID mice.(24) It was also shown that theosteoclast-like cells in GCT resorb bone in the presence, butnot the absence, of the stromal element of the tumor.(2) Thisis consistent with the recognized requirement for cell–cellcontact of the osteoclast with the osteoblast or stroma forfunction. We have shown evidence here that the mechanismby which GCT stromal cells induce the bone-resorbingfunction of giant cells is likely to be by ODF/RANK liga-tion, consistent with the normal situation of mature osteo-blasts and osteoclasts. To date, it has not been shown thatGCT stromal cells are capable of supporting osteoclasto-genesis from immature precursors, for example, CTR andvitronectin receptor-negative mononuclear cells. However,in addition to ODF, we have shown here the expression ofmRNA encoding a wide range of hematopoietic cytokinesimplicated in osteoclast formation and survival, includingIL-1a and -b, IL-1 receptor, IL-6, soluble IL-6 receptor,IL-17, TNF-a, and importantly, M-CSF. Although cytokinemRNA levels do not always correlate with secreted protein,we believe that these data are supportive of the overallosteoclastogenic nature of the GCT.
In summary, these data provide important new insightsinto the molecular biology of an osteolytic tumor type. Theexpression of molecules, thought to play a role in normalosteoclastogenesis, in GCT suggests the exciting possibilitythat they also play a role in pathological bone loss becauseof the stimulation of inappropriate osteoclastic activity. Itwill be important to determine the implications of thesefindings for other destructive primary bone and metastatictumors.
ACKNOWLEDGMENTS
This work was supported by grants from the Anti-CancerFoundation of Australia, The National Health and MedicalResearch Council of Australia, The Adelaide Bone and Joint
Research Foundation, Merck Sharp and Dohme (AustraliaPty Ltd Research Foundation), the Royal Adelaide HospitalResearch review committee, and Bristol-Myers Squibb/Zimmer. The authors thank Dr. Mark Clayer, M.D., Depart-ment of Orthopaedic Surgery, Queen Elizabeth Hospital,Woodville, South Australia, for providing one of the tumorsamples examined in this study.
REFERENCES
1. Goldring SR, Roelke MS, Petrison KK, Bhan A 1987 Humangiant cell tumors of the bone: Identification and characteriza-tion of cell types. J Clin Invest79:483–491.
2. James IE, Dodds RA, Lee-Rykaczewski E, Eichman CF, Con-nor JR, Hart TK, Maleeff BE, Lackman RD, Gowen M 1996aPurification and characterization of fully functional osteoclastprecursors. J Bone Miner Res11:1608–1618.
3. Zheng MH, Fan Y, Smith A, Wysocki S, Papadimitriou JM,Wood DJ 1998 Gene expression of monocyte chemoattractantprotein-1 in giant cell tumors of bone osteoclastoma: Possibleinvolvement in CD681 macrophage-like cell migration. J CellBiochem61:121–129.
4. Rothe L, Collin-Osdoby P, Chen Y, Sunyer T, Chaudhary L,Tsay A, Goldring S, Avioli L, Osdoby P 1998 Human oste-oclasts and osteoclast-like cells synthesize and release highbasal and inflammatory stimulated levels of the potent chemo-kine interleukin-8. Endocrinology139:4353–4363.
5. Martin TJ, Romas E, Gillespie MT 1998 Interleukins in thecontrol of osteoclast differentiation. Crit Rev Eukaryot GeneExpr 8:107–123.
6. Wong BR, Rho J, Arron J, Robinson E, Orlinick J, Chao M,Kalachikov S, Cayani E, Bartlett FS, Frankel WN, Lee SY,Choi Y 1997 TRANCE is a novel ligand of the tumor necrosisfactor receptor family that activates c-Jun N-terminal kinase inT cells. J Biol Chem272:25190–25194.
7. Anderson DM, Maraskovsky E, Billingsley WL, Dougall WC,Tometsko ME, Roux ER, Teepe MC, DuBose RF, Cosman D,Galibert L 1997 A homologue of the TNF receptor and itsligand enhance T-cell growth and dendritic-cell function. Na-ture 390:175–179.
8. Yasuda H, Shima N, Nakagawa N, Yamaguchi K, Kinosaki M,Mochizuki S, Tomoyasu A, Yano K, Goto M, Murakami A,Tsuda E, Morinaga T, Higashio K, Udagawa N, Takahashi N,Suda T 1998 Osteoclast differentiation factor is a ligand forosteoprotegerin/osteoclastogenesis inhibitory factor and isidentical to TRANCE/RANKL. Proc Natl Acad Sci USA95:3597–3602.
9. Nakagawa N, Kinosaki M, Yamaguchi K, Shima N, Yasuda H,Yano K, Morinaga T, Higashio K 1998 RANK is the essentialsignaling receptor for osteoclast differentiation factor in oste-oclastogenesis. Biochem Biophys Res Commun18:395–400.
10. Darnay BG, Haridas V, Ni J, Moore PA, Aggarwal BB 1998Characterization of the intracellular domain of receptor aciva-tor of NF-kB (RANK). J Biol Chem273:20551–20555.
11. Simonet WS, Lacey DL, Dunstan CR, Kelley M, Chang MS,Luthy R, Nguyen HQ, Wooden S, Bennett L, Boone T, Shi-mamoto G, DeRose M, Elliot R, Colombero A, Tan HL, TrailG, Sullivan J, Davy E, Bucay N, Renshaw-Gegg L, HughesTM, Hill D, Pattison W, Campbell P, Sander S, Van G,Tarpley J, Derby P, Lee R, Boyle WJ 1997 Osteoprotegerin: Anovel secreted protein involved in the regulation of bonedensity. Cell89:309–319.
12. Bucay N, Sarosi I, Dunstan CR, Morony S, Tarpley J, Cap-parelli C, Scully S, Tan HL, Xu W, Lacey DL, Boyle WJ,Simonet WS 1998 Osteoprotegerin-deficient mice developearly onset osteoporosis and arterial calcification. Genes Dev12:1260–1268.
648 ATKINS ET AL.

13. Kong Y-Y, Yoshida H, Sarosi I, Tan H-L, Timms E, Cappar-elli C, Morony S, Oliveira-dos-Santos, Van G, Itie A, Khoo W,Wakeham A, Dunstan CR, Lacey DL, Mak TW, Boyle WJ,Penninger JM 1999 OPGL is a key regulator of osteoclasto-genesis, lymphocyte development and lymph-node organogen-esis. Nature397:315–323.
14. Pitti RM, Marsters SA, Ruppert S, Donahue CJ, Moore A,Ashkenazi A 1996 Induction of apoptosis by Apo-2 ligand, anew member of the tumor necrosis factor cytokine family.J Biol Chem271:12687–12690.
15. Golstein P 1997 Cell death: TRAIL and its receptors. CurrBiol 7:R750–R753.
16. Emery JG, McDonnell P, Brigham Burke M, Deen KC, Lyn S,Silverman C, Dul E, Appelbaum ER, Eichman C, DiPrinzio R,Dodds RA, James IE, Rosenberg M, Lee JC, Young PR 1998Osteoprotegerin is a receptor for the cytotoxic ligand TRAIL.J Biol Chem273:14363–14367.
17. Tsukii K, Shima N, Mochizuki S-I, Yamaguchi K, KinosakiM, Yano K, Shibata O, Udagawa N, Yasuda H 1998 Osteoclastdifferentiation factor mediates an essential signal for boneresorption induced by 1a,25-dihydroxyvitamin D3, prosta-glandin E2, or parathyroid hormone in the microenvironmentof bone. Biochem Biophys Res Commun246:337–341.
18. Nicholson GC, Horton MA, Sexton PM, D’Santos CS, Mose-ley JM, Kemp BE, Pringle JA, Martin TJ 1987 Calcitoninreceptors of human osteoclastoma. Horm Metab Res19:585–589.
19. Moore EE, Kuestner RE, Stroop SD, Grant FJ, Matthews SL,Brady CL, Sexton PM, Findlay DM 1995 Functionally differ-ent isoforms of the human calcitonin receptor result fromalternative splicing of the gene transcript. Mol Endocrinol9:959–968.
20. Horwood NJ, Elliott J, Martin TJ, Gillespie MT 1998 Osteo-tropic agents regulate the expression of osteoclast differentia-tion factor and osteoprotegerin in osteoblastic stromal cells.Endocrinology139:4743–4746.
21. Rabbani SA, Gladu J, Harakidas P, Jamison B, Goltzman D1999 Over-production of parathyroid hormone-related peptideresults in increased osteolytic skeletal metastasis by prostatecancer cells in vivo. Int J Cancer80:257–264.
22. Guise TA, Yin JJ, Taylor SD, Kumagai Y, Dallas M, BoyceBF, Yoneda T, Mundy GR 1996 Evidence for a causal role ofparathyroid hormone-related protein in the pathogenesis ofhuman breast cancer-mediated osteolysis. J Clin Invest98:1544–1549.
23. Roodman GD 1997 Mechanisms of bone lesions in multiplemyeloma and lymphoma. Cancer80(Suppl 8):1557–1563.
24. James IE, Dodds RA, Olivera DL, Nuttall ME, Gowen M1996b Human osteoclastoma derived stromal cells: Correla-tion of the ability to form mineralized nodules in vitro withformation of bone in vivo. J Bone Miner Res11:1453–1460.
Address reprint requests to:David Findlay, Ph.D.
Department of Orthopaedics and TraumaUniversity of Adelaide
Royal Adelaide Hospital, North TerraceAdelaide, South Australia, 5000, Australia
Received in original form April 19, 1999; in revised form Septem-ber 17, 1999; accepted November 18, 1999.
649OSTEOCLASTOGENESIS IN GIANT CELL TUMORS




















