
Dynamic fragmentation and spheroidization of α phase grains ...
Upload
independentCategory
view
0download
0
Expression of natural antimicrobial human lysozyme in rice grains
Jianmin Huang1, Somen Nandi1, Liying Wu1, Dorice Yalda1, Glenn Bartley1,4, RaymondRodriguez2, Bo Lonnerdal3 and Ning Huang1,*1Ventria Bioscience, 4110 North Freeway Boulevard, Sacramento, CA 95834, USA; 2Section of Molecular andCellular Biology, University of California, Davis, CA 95616, USA; 3Department of Nutrition, University ofCalifornia, Davis, CA 95616, USA; 4Current address: United States Department of Agriculture, 800 BuchananStreet, Albany, CA 94710, USA; *Author for correspondence (e-mail: [email protected]; phone:916–921-6148; fax: 916–921-5611)
Received 8 January 2002; accepted in revised form 26 February 2002
Key words: Infant formula, natural antimicrobial, nutraceutical, recombinant human lysozyme, transgenic rice
Abstract
In the present study, we explored the expression of human lysozyme in maturing rice grains. Particle bombard-ment-mediated transformation was utilized to deliver the codon-optimized structural gene for human lysozymeto the callus of rice cultivar Taipei 309. Lysozyme expression is controlled by the promoter and signal peptidesequence for rice storage protein Glutelin 1. A total of 33 fertile plants were regenerated from independent trans-formation events and 12 of them with significant expression levels of lysozyme were advanced to further gen-erations. The transgenes were characterized by PCR and Southern blot analysis. Segregation analysis indicated atypical Mendelian 3: 1 inheritance, suggesting a single locus or closely linked loci of gene insertion. The ex-pression levels of lysozyme reached 0.6% of the brown rice weight or 45% of soluble proteins. Seven transgenicbreeding lines have been selected and followed over six generations. Lysozyme expression levels were main-tained in all generations. Biochemical, biophysical and functional comparisons of native and recombinant humanlysozyme revealed identical N-terminal sequence, molecular weight, pI and specific activity. Similar thermal andpH stability was observed for lysozyme from two sources. Furthermore, similar bactericidal activity was dis-played towards a laboratory strain of E. coli. The possibility of improving medical and nutritional quality ofinfant formulas and baby foods with rice flour or rice extract containing recombinant human lysozyme is dis-cussed.
Introduction
Human milk contains 50–400 �g/ml lysozyme (Hlys)and this enzyme is one of the most abundant proteinsin human milk (Lönnerdal 1985). In addition to hu-man milk, lysozyme is found also in other mamma-lian secretions, such as tears and saliva. Hlys consistsof unglycosylated polypeptide chain with 130 aminoacid residues, giving it a molecular weight of 15 kDa.Lysozyme (EC 3.2.1.17) kills bacterial cells by cleav-ing the linkage between N-acetylglucosamine and N-acetylmuramic acid in the peptidoglycan layer of thebacterial cell wall.
Pediatricians recommend newborns to be fed withbreast milk, a recommendation based on investiga-
tions like the comprehensive DARLING study (DavisArea on Lactation, Infant Nutrition and Growth). TheDARLING study documented for affluent middle-class families with well-educated mothers, thatbreast-fed infants have fewer incidences of diarrheaand upper respiratory infections than the formula-fedinfants. Upon infections, the duration of illness wassignificantly shorter in breast-fed infants than in for-mula-fed infants (Nommsen et al. 1991; Dewey et al.(1991, 1991, 1992); Peerson et al. 1993; Heinig et al.(1993, 1993); Dewey et al. 1993; Dewey and Heinig1995). Human milk contains a variety of unique pro-teins that actively protect newborns from diseases andthese contribute to the health of breast-fed infants(Lönnerdal 1985; Newman 1995). Unfortunately,
83Molecular Breeding 10: 83–94, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.

these important human milk proteins, such aslysozyme, lactoferrin and secretory IgA are notpresent in the commercially available infant formulasand baby foods (Lönnerdal 1985).
Various physiological malfunctions of motherand/or child as well as cultural habits can preventbreast feeding. In such cases, infant formulas have tobe used and their supplementation with biologicallyactive human milk proteins is desirable. Hlys is pres-ently purified from human milk, and therefore in lim-ited supply. Recombinant human lysozyme (rHlys)has been expressed in Saccharomyces cerevisiae, As-pergillus oryzae, mouse, Nicotiana tabacum and Dau-cus carota (Yoshimura et al. 1988; Tsuchiya et al.1992; Nakajima et al. 1997; Maga et al. 1998; Takai-chi and Oeda 2000), but expression levels were toolow to meet the requirement of high concentration ofHLys in human milk.
The production of recombinant human proteins inrice grains has several advantages: (1). Rice grainproteins can accumulate to 9–19% of grain weight(Lásztity 1996). (2). The endosperm proteins are syn-thesized during grain maturation and stored in proteinbodies for use in the germination and seedling growthof the next plant generation. (3). Grains can be storedfor years without loss of functionality, and thereforethe downstream processing can be conducted inde-pendently of growing seasons. (4). Specific promot-ers for expression of recombinant proteins are avail-able from the genes of the major rice storage proteins,the glutelin, Gt-1, Gt-2 and Gt-3 (Okita et al. 1989).
Rice grain, as one of the most important staplefoods for mankind, is consumed daily by more than40% of the world population. It is also recognized ashaving low allergenicity and has been recommendedas one of the first solid foods for infants and childrenwho have food allergies (Gastañaduy et al. 1990). Inthis study, we present the first effort to express hu-man lysozyme in rice grain. We have obtained an un-precedented high expression level of functional rHlys.Biochemical, biophysical and functional characteris-tics of the rHlys were identical to Hlys. Since ricegrain ingredients are well tolerated by infants, novelinfant formulas or baby foods containing rice extractwith Hlys are a realistic goal. By avoiding costly pro-tein purification steps for their preparation, the prod-ucts will become more viable and more readily avail-able.
Materials and methods
Cloning of the Gt-1 promoter and construction ofan expression plasmid
The promoter of the rice Glutelin 1 gene (Gt-1) andthe nucleotide sequence of the signal peptide wascloned with two primers based on the Gt1 gene se-quence (Okita et al. 1989). The forward primer withHindIII site was named MV-Gt-1-F1; 5�-ATC-GAAGCTTCATGAGTAATGTGTGAGCAT-TATGGGACCACG-3�. The reverse primer wasnamed Xba-Gt-1-R1; 5�-CTAGTCTAGACTC-GAGCCACGGCCATGGGGC CGGCTAGGGAGC-CATCGCACAAGAGGAA-3�. Crude DNA was iso-lated from leaves of rice variety M202 (Dellaporta etal. 1983). The PCR product amplified from the crudeDNA was cloned into pCR 2.1 (Invitrogen, Carlsbad,CA). The resulting plasmid was named pCRGt-1 orpAPI134.
To generate a Gt-1 expression plasmid, pAPI134was digested with HindIII and XbaI. The fragmentcontaining the Gt-1 promoter and Gt-1 signal peptidecode was cloned into a pUC19 based plasmid con-taining the nopaline synthase 3� (nos) terminator. Theresulting plasmid was named pAPI141 and containsthe rice Gt-1 promoter, the Gt-1 signal peptide code,a multiple cloning site and the nos terminator.
It has been shown that the codons for genes ex-pressed in rice are rich in guanine (G) or cytosine (C)in the third position (Huang et al. 1990). Changinglow G + C contents to high G + C contents has beenfound to increase the expression level of foreigngenes in barley grains (Horvath et al. 2000). The Hlysgene was synthesized by Operon (Operon Technolo-gies, Inc., Alameda, CA) based on the rice gene codonbias (also called codon-optimization) and named aslys-ger. Lys-ger was digested with DraI and XhoI andcloned into pAPI141 digested with NaeI and XhoIaccording to standard cloning techniques (Sambrooket al. 1989). The resulting plasmid was called pA-PI159 (Figure 1A).
Rice transformation and generation of R1 plantsexpressing human lysozyme
The basic procedures of particle bombardment-medi-ated rice transformation and plant regeneration werecarried out as described (Huang et al. (2001, 2001)).Individual R1 grains from R0 regenerants were dis-sected into embryos and endosperms. Expression lev-
84

els of rHlys in the endosperms were determined. Em-bryos from the individual R1 grains with high rHlysexpression were sterilized in 50% commercial bleachfor 25 min and washed with sterile H2O three timesfor 5 min each. Sterilized embryos were then placedin a tissue culture tube containing 1/2 strength of MSbasal salt (Sigma, St. Louis, MO) with addition of 1%sucrose and 0.05 mg/ml NAA (Sigma). Embryos weregerminated and plantlets having about 7 cm shootsand healthy root systems were obtained in about 2weeks. Mature R1 plants were generated as regener-ants.
Screening for plants expressing human lysozyme
Individual rice endosperms or grains were groundwith cold phosphate buffered-saline (PBS) with theaddition of 0.35 M NaCl. Grinding was conductedwith a pre-cooled mortar and pestle at 1 ml buffer/grain. Clear grain homogenate was obtained by sub-jecting the resulting grain extract to centrifugation at14,000 rpm for 10 min at 4 °C.
SDS-PAGE, electroblotting and Western blot anal-ysis were carried out with 18% precast gel (Invitro-gen, Carlsbad, CA) as described previously (Huanget al. 2001). The primary rabbit polyclonal antibodyagainst Hlys was purchased from Dako A/S (Den-mark) and used at 1:5000. Lysozyme was quantifiedby a turbidimetric activity assay with Micrococcusluteus (Sigma) on 96-well microtiter plate as de-scribed before (Humphrey et al. 2002). Briefly, 250�l of 0.015% M. luteus cell suspension was incubatedwith 10 �l of samples containing lysozyme with aconcentration less than 2.4 �g/ml. The reaction wasfollowed by the kinetic mode in Microplate Manager(Bio-Rad, CA) for 5 min at 450 nm. The concentra-tion of lysozyme was then determined in reference tothe standard curve.
Segregation was tested by Chi-square analysis inorder to determine the pattern of inheritance.
Cultivation of transgenic rice plants
Homozygous R1 plants were identified by analyzingpositive expressions of rHlys from a minimum of 20
Figure 1. A: Diagram of plasmid pAPI159 for expression of rHlys in transgenic rice grain. B & C: Identification of rHlys in rice grainextracts of transgenic plants by SDS-PAGE and Coomassie staining (B) or Western blot analysis (C). Amount of loaded protein was approxi-mately 10 �g per lane. TP-309 lane was loaded with soluble protein extract from non-transformed rice.
85

individual R2 grains. Homozygous lines derived fromthese plants were planted in a rice field in California.During growth, agronomic characteristics of bothtransgenic and non-transgenic plants, such as plantheight, percentage of fertility, number of effectivetillers, filled grains/panicle, non-filled grains/plant,time to maturity and 1000 grain weight were deter-mined and compared. Plants with satisfactory agro-nomic traits were selected and rHlys expression levelswere determined by lysozyme activity assay. Plantsthat met the criteria for satisfactory agronomic traitsand had more than 35 �g of rHlys/grain were ad-vanced to next generations.
Southern blot analysis of transgenic rice
Genomic DNA for Southern blot analysis was iso-lated from 3 g of young leaves following the proce-dure of Dellaporta (Dellaporta et al. 1983). About 5�g of DNA was then digested with HindIII andEcoRI, separated on a 1% agarose gel, blotted onto aHybond+ membrane (Amersham Pharmacia Biotech,Piscataway, NJ). The blot was probed with gel puri-fied human Hlys gene and developed by ECL directnucleic acid labeling and detection system (Amer-sham Pharmacia).
Tissue specific expression of lysozyme
The expression of the codon optimized lysozymegene was under control of the rice Glutelin 1 pro-moter, which is activated during rice endosperm de-velopment (Okita et al. 1989). To confirm that rHlysis expressed in the mature rice grain exclusively, tis-sues of mature and young leaf as well as mature andyoung root were collected and ground with 0.35 MNaCl in PBS. The resulting cell extract was centri-fuged and the clear supernatant was assayed by West-ern blot analysis for expression of rHlys.
Purification of recombinant human lysozyme
Transgenic rice grains expressing rHlys were dehu-sked and milled to flour conventionally. rHlys wasthen extracted by mixing the rice flour with 0.35 MNaCl in PBS at 100 g/l at room temperature for 1 h.The resulting mixture was first filtrated through 3 �mof a pleated capsule, then through 1.2 �m of a serumcapsule and finally through a Suporcap 50 capsulewith a 0.8 �m glass filter on top of a 0.45 �m filter(Gelman laboratory, Ann Arbor, MI).
The clear filtrate (1 L) was then dialyzed againstthe starting buffer (50 mM sodium phosphate, pH 8.5)at 4 °C overnight. Dialyzed sample was loaded ontoa cation exchange SP-Sepharose column (XK26/40,AmershamPharmacia Biotech) at 4 ml/min. The SP-Sepharose column had been equilibrated with thestarting buffer. After loading, the column was washedwith the same buffer until base-line of A280 readingwas achieved. Hlys was then eluted with 0.2 M NaClin 50 mM sodium phosphate, pH 8.5. Fractions con-taining rHlys were pooled and re-applied onto a gelfiltration Sephacryl S-100 column (AmershamPhar-macia Biotech). The column was equilibrated and runwith PBS. Pure rHlys fractions were identified by ac-tivity assay and total protein assay (Bradford 1976).Finally the purity of rHlys was confirmed by SDS-PAGE.
Bactericidal assay
E.coli strain JM 109 was used to investigate the bac-tericidal activity of lysozyme. An aliquot of an over-night JM 109 culture was grown in LB (Luria Broth)medium until mid-log phase. In order to increase thesensitivity of Gram-negative strain, like E. coli to thetreatment of lysozyme (Schütte and Kula 1990), 5mM EDTA was included in the incubation buffer.Buffer (20 mM sodium phosphate, pH 7.0 with 5 mMEDTA) alone, buffer containing Hlys, transgenic oruntransformed (TP309) grain extract were filter-ster-ilized. A standard inoculum of mid-log phase JM 109at 2 × 105 CFU (colony forming unit)/ml was incu-bated with sterilized samples at 37 °C for differentlengths of time. One fifth of the mixture volume wasplated onto LB agar plate and grown at 37 °C forovernight in order to determine the colony formingunits.
Determination of thermal stability, pH stability, pIand N-terminal sequences
Approximately 50 �l of Hlys or rHlys was dissolvedin PBS at 100 �g/ml and subjected to heat treatment.Four different temperatures of 65, 72, 85 and 100 °Cwere tested. With each temperature, 0 min, 0.33 min,1.5 min, 3 min, 5 min and 15 min were selected toanalyze the impact of incubation time on the stabilityof lysozyme.
For pH treatments at pH 2, 4 and 5, Hlys and rHlyswas dissolved in PBS adjusted to the correspondingpHs with HCl at 100 �g/ml. For pH 9 and 10,
86

lysozyme was dissolved in TBS and 150 mM sodiumcarbonate/bicarbonate at 100 �g/ml, respectively. Ap-proximately 100 �l of lysozyme solution was incu-bated at 37 °C for 30 min. The lysozyme activity wasassessed by activity assay.
Isoelectric points (pI) of Hlys and rHlys were de-termined by reverse IEF gel electrophoresis with aprecast Novex pH 3–10 IEF gel per instruction frommanufacturer (Invitrogen, Carlsbad, CA). PurifiedrHlys was resolved by 18% SDS-PAGE and blottedto a PVDF membrane (Bio-Rad). A rHlys band wasidentified by staining the membrane with CoomassieBrilliant Blue R-250 and sequenced at MolecularStructure Facility at University of California, Davis.
Results
Production of fertile transgenic rice plants
In order to increase the expression levels of Hlys inrice grains, 92 out of 130 codons for Hlys werechanged and the G + C content was raised from 46%to 68%. The synthetic Hlys gene was cloned into themultiple cloning site in an API base vector pAPI141that has the Gt-1 promoter, Gt-1 signal sequence codeand nos terminator in place (Figure 1A). A total of 77
transgenic rice R0 plants were generated and 33 setgrains sufficient for expression analysis. Individualgrains from the 33 R0 plants were dissected and theamount of rHlys in the endosperm estimated withCoomassie stained gels and Western blots. Expressionof rHlys in seven independently transformed riceplants is presented in Figures 1B & 1C. A dominatingband corresponding to the position of a protein withthe molecular mass of Hlys was detected in the saltsoluble fraction of crude extracts from the transgenicrice grains (Figure 1B). The identity of the proteinwas confirmed by its absence in crude grain extractfrom untransformed rice (Figure 1B, TP-309) and aspecific reaction with polyclonal antibody againstHlys (Figure 1C).
Segregation analysis and homozygous plantselection from R1 plants
Chi-square analyses from 15 representative R0 plantprogenies are shown in Table 1. The progenies from12 plants fitted a 3:1 segregation ratio, indicating thetransgene(s) were integrated into the same locus onthe rice chromosome in those plants. Progenies fromthree R0 plants (Plants 159–2-1, 159–21-1and 159–25-1) were not propagated because their segregation
Table 1. Segregation analysis of R0 plants.
Serial # R0 Plant # Grains X2 Value* Copy #
Screened Expression No Expression
1 159-1-1 17 12 5 0.14 4
2 159-2-1 39 23 16 4.94 ND**
3 159-5-1 20 12 8 2.07 2
4 159-12-1 39 27 12 0.57 1
5 159-18-1 44 37 7 2.21 4
6 159-20-1 44 32 12 0.09 2
7 159-21-1 45 22 23 15.7 ND**
8 159-25-1 20 20 0 7.4 ND**
9 159-26-1 50 37 13 0.03 2
10 159-29-1 20 18 2 2.9 2
11 159-37-1 20 18 2 2.9 2
12 159-38-1 10 7 3 0.13 6
13 159-41-1 6 3 3 1.6 3
14 159-43-1 20 12 8 2.06 5
15 159-53-1 20 12 8 2.06 5
Total 15 384 262 122
* X2 value: a value of less than 3.84 suggests that the chance of fitting 3:1 segregation ratio is over 95%.** ND: not determined.
87

failed to fit a 3:1 ratio, suggesting cases of instabilityof the transgenes in the rice chromosomes.
A total of 197 embryos from 12 selected R0 plantswere germinated and 157 R1 seedlings planted in thegreenhouse for generation of R2 grains. Individual R2
grains (n = 1502) from 109 R1 fertile plants werescreened for lysozyme expression by lysozyme activ-ity assay in order to identify 42 homozygous plants.
Expression levels of rHlys are maintained intransgenic plants
Grains from the 42 homozygous plants were propa-gated and the generated plants grown in a rice field inCalifornia. Several major agronomic characteristics(data not shown) and the expression levels of rHlys
from transgenic plants were monitored in every gen-eration. Expressions of rHlys of the seven lines overtwo to four generations determined by lysozyme ac-tivity assay are displayed in Figure 2. The amount ofrHlys synthesized in the individual lines was constantthroughout the period tested except line 159–41.Thehighest stable expression level of rHlys was achievedin line 159–53. It produced 0.6% rHlys per brown riceweight amounting to 45% of the total soluble proteinextract from rice grain. The different levels of expres-sion were evident from Coomassie stained gel (Fig-ure 1B). On average, the lines yielded 0.37% perbrown rice weight.
Figure 2. Stable rHlys expression in transgenic rice grains. Proteins from 1 g of brown rice flour were extracted with 40 ml of extractionbuffer containing 0.35 M NaCl in PBS. Extraction was conducted at room temperature for 1 h with shaking. Homogenate was centrifuged at14,000 rpm for 15 min at 4 °C. Protein supernatant was removed and diluted as needed for lysozyme turbidimetric activity assay (Humphreyet al. 2002). Extraction was repeated three times and standard deviation was shown as an error bar. Lysozyme yield was expressed as per-centage of total soluble protein (%TSP).
88

Southern blot analysis of transgenic lines
To determine the copy number of transgene integratedinto the rice genome, Southern blot analysis was per-formed on seven selected transgenic plants (Figure3A). By comparing to known amounts of the intact1470 bp Hlys gene, the intact copy number of the
transgenes, including promoter and Hlys gene, wasestimated to vary from 1 to 6 (Table 1). No positivecorrelation between copy number of the rHlys trans-gene and amount of rHlys synthesized was discern-ible (Table 1) & (Figure 2). The presence of larger orsmaller hybridizing fragments is probably due to theloss of one or several restriction enzyme recognition
Figure 3. Southern blot analysis of transgenic plants. Genomic DNA was isolated according to what was described in Materials and meth-ods. Genomic DNA was digested with HindIII and EcoRI and probed with the gel-purified human lysozyme gene. 1X = 1 copy genomeequivalent, 2X = 2 copies genome equivalent. A: Southern blot analysis of 7 transgenic lines; B: Southern blot analysis of a transgenic linethrough 5 generations.
89

sites in the course of integration or partial digestion.The different hybridization patterns of these lines fur-ther confirmed that they were from independent trans-formation events (Figure 3A). The integration pat-terns and copy numbers were conserved over 5generations as demonstrated for line 159–53-1 (Fig-ure 3B). Identical hybridization patterns of 159–53-1–13 and 159–53-1–16 also indicated that they werefrom the same mother plant.
Expression of rHlys is grain specific
Since the Gt-1 promoter is activated only in the de-veloping endosperm of the grain (Okita et al. 1989),rHlys is expected to be synthesized exclusively in thistissue. As shown in Figure 4, rHlys was only foundin mature and germinated grain, but not in any of theother tissues tested.
Purification of rHlys from rice grains
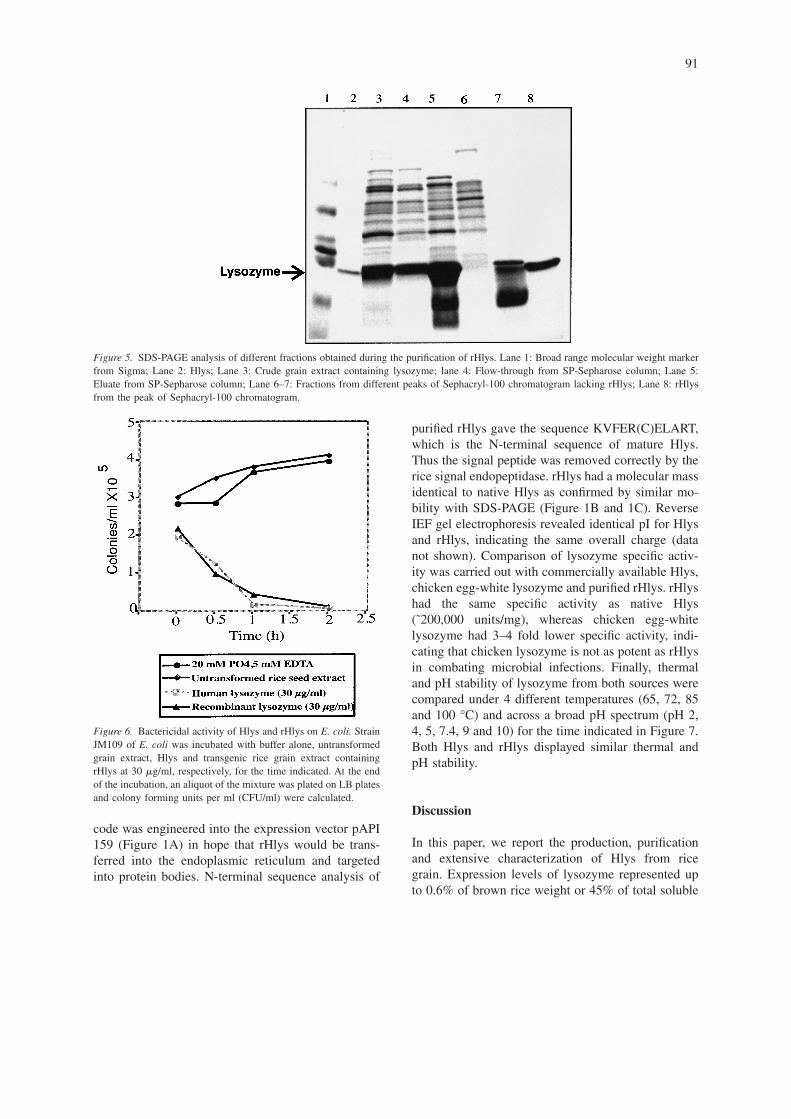
A 3-step filtration process was developed to clarifythe crude rice extract for column chromatographywithout the time consuming centrifugation step (cf.Materials and methods). More than 4 liters of cruderice extract could be clarified for column purification
within 30 min. Purification of rHlys was carried outby chromatography with a cation exchange SP-Sepharose column followed by gel filtration on aSephacryl S-100 column (cf. Materials and methods).rHlys bound to the first column and was eluted spe-cifically with 0.2 M NaCl (Figure 5, lane 5), while amajority of the rice proteins were removed by thischromatography step (Figure 5, lane 4). The gel fil-tration column permitted the isolation of a fractionwith electrophoretically homogenous rHlys (Figure 5,lane 8). The overall recovery of rHlys achieved bythis purification scheme was 60% (data not shown).
Characterizations of rHlys
The bactericidal activity of rHlys was compared withHlys by using an E. coli strain, JM109 (Figure 6). Ata concentration of 30 �g/ml, rHlys displayed a bac-tericidal time course activity similar to Hlys. In fact,after 1 h of incubation, almost all bacteria had beenkilled, while buffer (20 mM sodium phosphate, pH7.0 with 5 mM EDTA) alone and untransformed ricegrain extract had no killing effect (Figure 6).
After rHlys was purified to near homogeneity, sev-eral biochemical and biophysical characterizationswere carried out. The rice Glutelin 1 signal sequence
Figure 4. Analysis of rHlys in different rice tissues. Leaf, root and grain were harvested from both transformed (T) and un-transformed (U)plants. Proteins were extracted as described in Materials and methods. Crude extract was separated with an 18% precast SDS-PAGE.Lysozyme was identified by specific antibody.
90
code was engineered into the expression vector pAPI159 (Figure 1A) in hope that rHlys would be trans-ferred into the endoplasmic reticulum and targetedinto protein bodies. N-terminal sequence analysis of
purified rHlys gave the sequence KVFER(C)ELART,which is the N-terminal sequence of mature Hlys.Thus the signal peptide was removed correctly by therice signal endopeptidase. rHlys had a molecular massidentical to native Hlys as confirmed by similar mo-bility with SDS-PAGE (Figure 1B and 1C). ReverseIEF gel electrophoresis revealed identical pI for Hlysand rHlys, indicating the same overall charge (datanot shown). Comparison of lysozyme specific activ-ity was carried out with commercially available Hlys,chicken egg-white lysozyme and purified rHlys. rHlyshad the same specific activity as native Hlys(˜200,000 units/mg), whereas chicken egg-whitelysozyme had 3–4 fold lower specific activity, indi-cating that chicken lysozyme is not as potent as rHlysin combating microbial infections. Finally, thermaland pH stability of lysozyme from both sources werecompared under 4 different temperatures (65, 72, 85and 100 °C) and across a broad pH spectrum (pH 2,4, 5, 7.4, 9 and 10) for the time indicated in Figure 7.Both Hlys and rHlys displayed similar thermal andpH stability.
Discussion
In this paper, we report the production, purificationand extensive characterization of Hlys from ricegrain. Expression levels of lysozyme represented upto 0.6% of brown rice weight or 45% of total soluble
Figure 5. SDS-PAGE analysis of different fractions obtained during the purification of rHlys. Lane 1: Broad range molecular weight markerfrom Sigma; Lane 2: Hlys; Lane 3: Crude grain extract containing lysozyme; lane 4: Flow-through from SP-Sepharose column; Lane 5:Eluate from SP-Sepharose column; Lane 6–7: Fractions from different peaks of Sephacryl-100 chromatogram lacking rHlys; Lane 8: rHlysfrom the peak of Sephacryl-100 chromatogram.
Figure 6. Bactericidal activity of Hlys and rHlys on E. coli. StrainJM109 of E. coli was incubated with buffer alone, untransformedgrain extract, Hlys and transgenic rice grain extract containingrHlys at 30 �g/ml, respectively, for the time indicated. At the endof the incubation, an aliquot of the mixture was plated on LB platesand colony forming units per ml (CFU/ml) were calculated.
91

proteins from dehusked grain. To the best of ourknowledge, this represents the highest expressionlevel of human protein achieved in grain from trans-genic plants. The optimal production of human
lysozyme were achieved by a number of conditions:(A). Careful design of expression vector (selection ofpromoter and terminator, addition of genetic code forsignal sequence). The nucleotide sequence of the sig-
Figure 7. A: Thermal stability of Hlys and rHLys. Lysozyme was dissolved at 100 �g/ml in PBS. The mixtures were treated at differenttemperatures for different lengths of time as discussed in Materials and methods. At the end of each treatment, the remaining lysozymeactivity was assessed by activity assay. B: pH stability of Hlys and rHlys. Lysozyme was dissolved in different buffers as discussed in Ma-terials and methods at 100 �g/ml. The mixture was incubated at 37 °C for 30 min. The lysozyme activity was determined by activity assay.
92

nal sequence was included in the expression cassettewith the aim of directing the synthesized protein intothe lumen of the endoplasmic reticulum and accumu-lating in protein bodies. It was reported for barley thatin the absence of a signal sequence, recombinant pro-tein expression in the endosperm resulted in low yield(Horvath et al. 2000). This is probably due to the ex-tensive proteolytic activity in the cytosol during thefinal stages of grain maturation. (B). Codon optimi-zation of the transgene. (C) An efficient rice transfor-mation protocol allowed a sufficient number of trans-genic plants to be obtained and screened. Thus, highexpressing plants resulting from the transgene inte-gration at superactive loci (Shirsat et al. 1989) couldbe identified.
Although the potential application of rHlys as annutraceutical in the form of infant formula and babyfood has been pointed out in the introduction, thereare applications of lysozyme in other areas, such as afood preservative (Proctor and Cunningham 1988;Hughey et al. (1987, 1989)) or as a pharmaceutical(Proctor and Cunningham 1988) or as a cosmeticproduct (Sim et al. 2000). More significantly, rHlyshas also been shown to have a potential use as ananimal growth promotant in an effort to eliminate orreduce the antibiotics consumption in animal hus-bandry (Humphrey et al. 2002).
High level expression of human lysozyme in ricehas implications regarding the production of valuablehuman proteins in an economic and safe way. Rice isconsumed by a majority of the population in theworld and is being Generally Regarded As Safe(GRAS). This makes rice attractive as a “protein fac-tory” to produce biomedicals and nutraceuticals forhuman consumption. The cloning and expression ofhuman lysozyme in rice grains has opened a new av-enue for the bioproduction of other milk proteins. Itis the ultimate goal that by using rice grains as an ef-ficient, economic and safe “bio-reactor”, productionscale quantities of human milk proteins will be gen-erated. This study presented the first effort to engineerthe gene coding for one of the major human milkproteins, lysozyme, into the rice genome. Transforma-tion and characterization of other human milk pro-teins in transgenic rice grains are in progress. WithVentria rice varieties harboring different human milkprotein genes, we shall be able to develop a novelform of infant formulas and baby foods that mimicthe nutritional and medical values of human milk.
Acknowledgements
We thank Frank E. Hagie for encouragement and pro-fessor Diter von Wettstein from Washington StateUniversity for critical review. We thank Lifang Chen,Phuong Pham, Stephanie Spencer, and Kelli Bengefor technical assistance.
References
Bradford M.M. 1976. A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the princi-ple of protein-dye binding. Anal. Biochem. 72: 248–254.
Dellaporta S.L., Wood J. and Hick J.B. 1983. A plant DNA minipreparation; version II. Plant Mol. Biol. Report. 1: 19–21.
Dewey K.G., Heinig M.J., Nommsen L.A. and Lönnerdal B. 1991.Adequacy of energy intake among breast-fed infants in theDARLING study: relationships to growth velocity, morbidity,and activity levels. Davis Area Research on Lactation, InfantNutrition and Growth [see comments]. J. Pediatr. 119: 538–547.
Dewey K.G., Heinig M.J., Nommsen L.A. and Lonnerdal B. 1991.Maternal versus infant factors related to breast milk intake andresidual milk volume: the DARLING study. Pediatrics 87: 829–837.
Dewey K.G., Heinig M.J., Nommsen L.A., Peerson J.M. and Lön-nerdal B. 1992. Growth of breast-fed and formula-fed infantsfrom 0 to 18 months: the DARLING Study. Pediatrics 89:1035–1041.
Dewey K.G., Heinig M.J., Nommsen L.A., Peerson J.M. and Lön-nerdal B. 1993. Breast-fed infants are leaner than formula-fedinfants at 1 y of age: the DARLING study. Am. J. Clin. Nutr.57: 140–145.
Dewey K.G. and Heinig M.J. 1995. Nommsen-Rivers LA: Differ-ences in morbidity between breast-fed and formula-fed infants.J. Pediatr. 126: 696–702.
Gastañaduy A., Cordano A. and Graham G.G. 1990. Acceptability,tolerance, and nutritional value of a rice-based infant formula.J. Pediatr. Gastroenterol. Nutr. 11: 240–246.
Heinig M.J., Nommsen L.A., Peerson J.M., Lonnerdal B. andDewey K.G. 1993. Energy and protein intakes of breast-fed andformula-fed infants during the first year of life and their asso-ciation with growth velocity: the DARLING Study. Am. J. Clin.Nutr. 58: 152–161.
Heinig M.J., Nommsen L.A., Peerson J.M., Lonnerdal B. andDewey K.G. 1993. Intake and growth of breast-fed and formu-la-fed infants in relation to the timing of introduction of com-plementary foods: the DARLING study. Davis Area Researchon Lactation, Infant Nutrition and Growth. Acta. Paediatr. 82:999–1006.
Horvath H., Huang J., Wong O., Kohl E., Okita T., KannangaraC.G. et al. 2000. The production of recombinant proteins intransgenic barley grains. Proc. Natl. Acad. Sci. USA 97: 1914–1919.
Huang N., Simmons C.R. and Rodriguez R.L. 1990. Codon usagepatterns in plant genes. J. CAASS 1: 73–86.
93

Huang J., Sutliff T.D., Wu L., Nandi S., Benge K., Terashima M.et al. 2001. expression and purification of functional human al-pha-1-antitrypsin from cultured plant cells. Biotechnol. Prog.17: 126–133.
Huang N., Wu L., Nandi S., Bowman E., Huang J., Sutliff T. et al.2001. The tissue specific activity of a rice beta-glucanase 9promoter is used to selecte rice transformants. Plant Science161: 603–609.
Sim Y.C., Lee S.G., Lee D.C., Kang B.Y., Park K.M., Lee J.Y. etal. 2000. Stabilization of papain and lysozyme for applicationto cosmetic products. Biotechnol. Lett. 22: 137–140.
Proctor V.A. and Cunningham F.E. 1988. The chemistry oflysozyme and its use as a food preservative and a pharmaceu-tical. Crit. Rev. Food Sci. Nutr. 26: 359–395.
Hughey V.L., Wilger P.A. and Johnson E.A. 1987. Antimicrobialactivity of hen egg white lysozyme against Listeria monocyto-genes Scott A in foods. Appl. Environ. Microbiol. 55: 631–638.
Humphrey B.D, Huang N. and Klasing K.C. 2002. Rice Express-ing lactoferrin and lysozyme has antibiotic-like properties whenfed to chicks. J. Nutr. V132: 1214–1218.
Lásztity R. 1996. The Chemistry of Cereal Proteins. CRC Press,Boca Raton.
Lönnerdal B. 1985. Biochemistry and physiological function ofhuman milk proteins. Am. J. Clin. Nutr. 42: 1299–1317.
Maga E., Anderson G., Cullor J., Smith W. and Murray J. 1998.Antimicrobial properties of human lysozyme: Transgenicmouse milk. J. Food Protection 61: 52–56.
Nakajima H., Muranaka T., Ishige F., Akutsu K. and Oeda K. 1997.Fungal and bacterial disease resistance in transgenic plants ex-pressing human lysozyme. Plant Cell Rep. 16: 674–679.
Newman J. 1995. How breast milk protects newborns. ScientificAmerican 273: 76–79.
Nommsen L.A., Lovelady C.A., Heinig M.J., Lönnerdal B. andDewey K.G. 1991. Determinants of energy, protein, lipid, and
lactose concentrations in human milk during the first 12 mo oflactation: the DARLING Study. Am. J. Clin. Nutr. 53: 457–465.
Okita T.W., Hwang Y.S., Hnilo J., Kim W.T., Aryan A.P., LarsonR. et al. 1989. Structure and Expression of the Rice GlutelinMultigene Family. J. Biol. Chem. 264: 12573–12581.
Peerson J.M., Heinig M.J., Nommsen L.A., Lönnerdal B. andDewey K.G. 1993. Use of growth models to describe patternsof length, weight, and head circumference among breast-fedand formula-fed infants: the DARLING Study. Hum. Biol. 65:611–626.
Shirsat A., Wilford N. and Croy R. 1989. Gene copy number andlevels of expression in transgenic plants of a seed specific gene.Plant Sci. 61: 75–80.
Sambrook J., Maniatis T. and Fritsch E.F. 1989. Molecular Clon-ing: a Laboratory Manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, New York, USA.
Schütte H. and Kula M.R. 1990. Pilot- and process-scale tech-niques for cell disruption. Biotechnol. Appl. Biochem. 12: 599–620.
Hughey V.L., Wilger P.A. and Johnson E.A. 1989. Antibacterialactivity of hen egg white lysozyme against Listeria monocyto-genes Scott A in foods. Appl. Environ. Microbiol. 55: 631–638.
Takaichi M. and Oeda K. 2000. Transgenic carrots with enhancedresistance against two major pathogen, Erysiphe heraclei andAlternaria, dauci. Plant Sci. 25: 135–144.
Tsuchiya K., Tada S., Gomi K., Kitamoto K., Kumagai C., JigamiY. et al. 1992. High level expression of the synthetic humanlysozyme gene in Aspergillus oryzae. Appl. Microbiol. Biotech-nol. 38: 109–114.
Yoshimura K., Toibana A. and Nakahama K. 1988. Humanlysozyme: sequencing of a cDNA, and expression and secre-tion by Saccharomyces cerevisiae. Biochem. Biophys. Res.Commun. 150: 794–801.
94
Copyright © 2022 FDOKUMEN




















