
Truncated Navier-Stokes Equations with the Automatic Filtering
Upload
independentCategory
view
1download
0
Transgenic Research 11: 467–478, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
467
Expression of a truncated Brca1 protein delays lactational mammarydevelopment in transgenic mice
Melissa A. Brown1,2,3,4,∗, Hans Nicolai1,2, Kathy Howe1,2, Toyomasa Katagiri1, El-NasirLalani5, Kaylene J. Simpson3, Nathan W. Manning3, Andrew Deans3, Phil Chen6, Kum KumKhanna6, Mas Rina Wati4, Beatrice L. Griffiths2, Chun-Fang Xu1, Gordon W. H. Stamp5 &Ellen Solomon1
1Cancer Genetics Laboratory, Department of Medical and Molecular Genetics, King’s College London, Guy’sHospital, London, SE1 9RT, UK2Imperial Cancer Research Fund, Lincoln’s Inn Fields, London, WC2A 3PX, UK3Department of Biochemistry and Molecular Biology, University of Melbourne, Parkville, 3052, Australia4Department of Biochemistry and Molecular Biology, University of Queensland, St. Lucia, 4072, Australia5Department of Histopathology, Imperial College School of Medicine, The Hammersmith Hospital Campus,London, W12 0NN, UK6Queensland Institute of Medical Research, Herston, 4006, Australia
Received 14 November 2001; revised: 4 April 2002; accepted 5 April 2002
Key words: Brca1, dominant-negative, mammary gland, transgenic mice
Abstract
To address the hypothesis that certain disease-associated mutants of the breast-ovarian cancer susceptibility geneBRCA1 have biological activity in vivo, we have expressed a truncated Brca1 protein (trBrca1) in cell-lines andin the mammary gland of transgenic mice. Immunofluorescent analysis of transfected cell-lines indicates thattrBRCA1 is a stable protein and that it is localized in the cell cytoplasm. Functional analysis of these cell-linesindicates that expression of trBRCA1 confers an increased radiosensitivity phenotype on mammary epithelial cells,consistent with abrogation of the BRCA1 pathway. MMTV-trBrca1 transgenic mice from two independent linesdisplayed a delay in lactational mammary gland development, as demonstrated by altered histological profilesof lobuloalveolar structures. Cellular and molecular analyses indicate that this phenotype results from a defectin differentiation, rather than altered rates of proliferation or apoptosis. The results presented in this paper areconsistent with trBrca1 possessing dominant-negative activity and playing an important role in regulating normalmammary development. They may also have implications for germline carriers of BRCA1 mutations.
Introduction
The BRCA1 gene was isolated by positional clon-ing in 1994 as the first breast and ovarian cancersusceptibility gene. The product of BRCA1 is a nu-clear phosphoprotein that contains an N-terminal zincbinding RING domain, thought to mediate protein-protein interactions, and a C-terminal BRCT motif,which is found in a number of DNA repair proteins.
∗Author for correspondence: Department of Biochemistry andMolecular Biology, University of Queensland, St. Lucia, Queens-land, 4072, Australia: E-mail: [email protected]
A large number of BRCA1-associating proteins havebeen identified, which together implicate BRCA1 inthe regulation of DNA repair, transcription, the cellcycle and apoptosis (reviewed in Deng & Brodie,2000). Furthermore, BRCA1 has been classified as amember of the ‘caretaker’ family of tumour suppres-sors (reviewed in Brugarolas & Jacks, 1997; Kinzler& Vogelstein, 1997).
The products of germline tumour suppressor genemutations sometimes display biological activity. Forexample, they can block the activity of their nor-mal counterpart in a dominant-negative way. Some

468
versions of mutant p53 for instance are capable ofobstructing the function of wild-type p53, as demon-strated by inhibition of p53-mediated transcriptionalregulation in vitro and tumour suppressor activity invivo (Kern et al., 1992; Harvey et al., 1995). Mutantversions of WT1 (Haber et al., 1992; Englert et al.,1995), PMS2 (Nicolaides et al., 1998) and ATM(Morgan et al., 1997) have also been shown to limitthe function of their normal equivalent. Dominantnegative activity of tumour suppressor mutants has im-plications for germ-line carriers of these mutations andalso for the development of somatic models to studythe consequences of disrupting tumour suppressoractivity.
In this paper, we have addressed the hypothesisthat a truncated BRCA1 molecule (trBRCA1), con-taining approximately the first 300 amino acids andtherefore at least the first protein interaction domain,has biological activity in vivo. We have done thisby expressing trBRCA1 both in cell-lines and in themammary gland of transgenic mice. We show thattrBRCA1 is a stable cytoplasmic protein, that its ex-pression in mammary epithelial cells results in anincrease in radiosensitivity and that it is capable ofdisrupting lactational mammary gland development inmice.
Materials and methods
cDNA cloning, analysis, and transgene construction
Truncated BRCA1 cDNAs were generated by EcoRIdigestion of full-length BRCA1 cDNAs. This re-leases the 5′ end and cuts at nucleotide 1021 in hu-man BRCA1 (Accession no. HSU14680) and 1014in mouse Brca1 (Accession no. U32446). Transienttransfection experiments utilized a myc-tagged trun-cated human BRCA1 cDNA, generated by cloningthe EcoRI fragment of BRCA1 into pcDNA3.1-/Myc-His B (Invitrogen). Stable transfection constructscontained human full-length and truncated BRCA1cDNA cloned into the SV40 enhancer/promoter vectorpZeoSV (Invitrogen). Transgenic constructs containedtruncated mouse Brca1 cDNA cloned into the MMTVpromoter/SV40 polyA vector MMTV-Sv40-Bssk,originally described by Philip Leder (Harvard, MA)(Stewart et al., 1984) and provided by Clive Dickson(ICRF, London). All constructs were completely se-quenced prior to use in gene transfer experiments. Theprotein coding potential of these cDNAs was assessed
by in vitro translation using the TNT/T3 system (Pro-mega). The ability of anti-BRCA1 antibodies to recog-nize in vitro translated BRCA1 protein was assessedby western analysis of in vitro translated products us-ing the MS110 and MS13 anti N-term human BRCA1antibodies (from Ralph Scully, Dana Farber).
Mammalian cell culture and transfections
HBL100 cells were obtained from ICRF cell servicesor ATCC. Transient transfections were performed oncoverslips seeded with 5 × 104 cells 24 h prior to trans-fection, using 0.5 µg of DNA (or no DNA as a control)and FuGene reagent (Boehringer) and analysed 24 hpost-transfection. Stable transfections were performedusing Lipofectamine Plus reagent (Gibco-BRL), wereselected in 250 µg/ml Zeocin (Invitrogen) and expres-sion verified by either northern analysis, RT-PCR orwestern blotting.
Immunofluorescence and confocal microscopy
Transiently transfected cells grown on coverslips werefixed for 5 min at 4◦C in methanol/acetone (1:1).Endogenous BRCA1 expression was detected usingAP16 (hybridoma supernatant diluted 1:50), a mono-clonal antibody directed against a C-terminal epitope(1313-1863) (from Ralph Scully). The expression oftransfected trBRCA1 protein was detected using anantibody against the myc epitope tag (A-14, diluted1:200, Santa Cruz). Secondary antibody conjug-ates were FITC-conjugated goat anti-mouse im-munoglobulins (1:200) and TRITC-conjugated swineanti-rabbit immunoglobulins (1:100) (DAKO). Nu-clei were counterstained after the secondary antibodyincubations for about 5 min using Hoechst 33342(Sigma). Coverslips were mounted in fluorescentmounting medium (DAKO).
Radiosensitivity assay
Stably transfected HBL100 cells (6 × 102, 1.2 × 102,1.8 × 102, 2.4 × 103 and 3 × 103) were plated in du-plicate T25 flasks and allowed to attach overnight.Dishes were then exposed to 0, 1, 2, 3, 4 and 5 Gy ion-izing radiation (using a calibrated Caesium source atthe Royal Brisbane Hospital), respectively and incub-ated for between 10 and 14 days with regular mediumchanges. Cells were then fixed with methanol, stainedwith Geimsa and colonies of greater than 50 cellscounted using an inverted microscope at 40× totalmagnification.

469
Generation of transgenic mice
Linearized construct DNA (1–3 ng) was microinjectedinto fertilized CBA × C57BL/6 day 1 zygote pronuc-lei and introduced into pseudopregnant females. Ap-proximately 70 offspring were screened by Southernanalysis of EcoRI digested tail DNA using an 800 bpXbaI fragment of MMTV-Sv40-Bssk containing theSV40 polyA. Three founder lines were established andexamined for transgene expression by northern anal-ysis of RNA extracted from pregnant or post-partummammary gland tissue, using a 5′ EcoRI fragment ofBrca1 (nucleotides 25–1014) as a probe. Two linesof transgenic mice expressing the truncated Brca1construct were then expanded for further analysis.
Histological and mammary whole mount analysis oftransgenic mice
Transgenic mice and non-transgenic littermates wereanalyzed at different stages of pregnancy, deter-mined by timed mating plug date and confirmed bymicroscopic analysis and staging of embryos. Firstparity transgenic mice from the two independentfounder lines were examined, together with their non-transgenic littermates as controls. Between one andsix mice were analyzed at each time point. All ex-periments were performed ‘double-blind’ and in du-plicate (day 15, day 18 and post-partum). Mice werehumanely killed, the abdominal cavity opened andall tissues fixed by infusion of 10% formal salinefor 24–48 h. All tissues were processed to paraffinwax, sectioned at 4 µm and stained with haematoxylinand eosin. Mammary gland architecture was analyzedby whole mount processing exactly as described inJackson et al. (1997).
Expression analysis for markers of differentiation
Mammary tissue at days 14 and 18 of pregnancy wassnap frozen in liquid nitrogen and RNA extracted us-ing Trizol according to manufacturer’s instructions(Invitrogen). Total RNA (5 µg) was electrophoretic-ally separated in 1% agarose, 18% formaldehyde gelsand transferred to Hybond N membrane according tothe manufacturer’s instructions (Amersham Pharma-cia). Membranes were prehybridized for 3 h at 65◦C(in 0.5 M NaHPO4, 1% BSA, 1 mM EDTA and 7%SDS) and hybridized overnight with α-32P dATP ran-dom labelled partial cDNAs to mouse whey acidicprotein and β-casein genes (WAP: forward 5′TAG-CAGCAGATTGAAAGCATTATG3′, reverse 5′GA-CACCGGTACCATGCGTTG3′ (http://mammary.nih.
gov/tools/molecular/WAP/MMD.html) and β-casein:forward 5′ATGAAGGTCTTCATCCTCGCCTGCC3′,reverse 5′GCTGGACCAGAGACTGAGGAAGGTG-C3′ (Lindeman et al., 2001)). Membranes were wash-ed at 65◦C at high stringency (30 min at 0.2 × SSC/0.1% SDS, followed by 30 min at 0.1 × SSC/0.1 ×SDS) and exposed to Biomax MR film (Kodak) atroom temperature for 1–3 h.
Proliferation assay
Stably transfected mammary epithelial cell-lines weregenerated as described above. Proliferation rates ofthese cells were monitored by triplicate counting everyday for 7 days using a haemocytometer. Primary mam-mary epithelial cells were isolated from transgenic andnon-transgenic mice as described in Pullan and Streuli(1996). Briefly, the mammary tissue from 16 to 18 daypregnant MMTV-trBrca1 and non-transgenic controlmice was removed and digested to 50–200 cell aciniusing Collagenase C3 (Worthington – 12 mg/g tissue)in DMEM-F12 media containing insulin (5 µg/ml) andhydrocortisone (1 µg/ml) at 37◦C. The cell slurry wassubjected to a series of washes in the presence ofsoybean trypsin inhibitor (Sigma – 0.1 mg/ml) andDNase1 (Roche – 0.02 mg/ml) in Hanks Balanced SaltSolution and filtration through 150 and 53 µm meshbefore differential plating onto tissue culture plasticfor 1 h to remove fibroblasts. Proliferation of primarycells was monitored by plating similar volumes ofevenly sized organoids from comparative cell prepsand looking for any evidence of a growth advantage.
TUNEL assay
OCT-embedded cryopreserved mammary glands fromwild-type and transgenic animals at day 1 of lactationand day 2 involution (non-transgenic only) were sec-tioned at 10 µm. Sections were fixed, permeabilizedand stained using the Fluorescein In situ Cell DeathDetection kit (Roche Molecular Biochemicals) ac-cording to the manufacturer’s instructions. Propidiumiodide (0.2 µg/ml, Sigma) was used as a counterstain.Sections were analyzed using a Zeiss fluorescencemicroscope.
Results
trBRCA1 is a stable protein that is localized in thecytoplasm
To confirm that our cDNAs were capable of directingexpression of the correct protein, in vitro translation

470
Figure 1. trBRCA1 is a stable cytoplasmic protein. (A) In vitro translation (lane 1–6) and western analysis (lane 7–12) of transgene constructs.Lane 1 + 7: pZeoSV vector negative control, lane 2 + 8: pZeoSV plus 1 kb human trBRCA1 cDNA, lane 3 + 9: pZeoSV plus mouse 1 kbBrca1 cDNA, lane 4 + 10: pZeoSV plus 1kb antisense mouse Brca1 cDNA, lane 5 + 11: no DNA, lane 6 + 12: luciferase control cDNA.Western blot was probed with a combination of MS110 and MS13 monoclonal antibodies. (B) Immunofluorescent analysis of HBL100 cellstransiently transfected with myc tagged trBRCA1. TrBRCA1 is detected with rabbit-anti-myc antibody plus TRITC-labelled sheep-anti-rabbit(red). Endogenous BRCA1 is detected with AP16 anti BRCA1 C-term antibody and a FITC labelled goat anti-mouse secondary antibody(green).
and western analysis was performed. As shown in Fig-ure 1(A), human trBRCA1 translates a protein of theexpected size (lane 2) that is recognized by the anti-BRCA1 antibodies (lane 8). The mouse trBrca1 cDNA(lane 3) produced a major protein band slightly highercompared to the human BRCA1 cDNA (lane 2), pre-sumably due to slight differences in post-translationalmodification by rabbit reticulocyte component en-zymes, or illegitimate upstream initiation of transla-tion in the mouse, due to the lack of an in-frame stopupstream of the initiating methionine. This may alsoexplain the origin of a second smaller band in lane2. The mouse trBrca1 protein, as expected, was not
recognized by the anti-human BRCA1 antibody (lane9). Unfortunately, no suitable anti-mouse Brca1 anti-body is currently available for this analysis. No proteinproduct was detected in the antisense negative control(lane 4).
To determine if trBRCA1 could be expressed invivo, HBL100 human mammary epithelial cells weretransiently transfected with a myc-tagged trBRCA1construct and analyzed by immunofluoresence, us-ing an anti-Myc antibody to detect trBRCA1 (red)and AP16, an anti-C-term BRCA1 antibody to re-cognize endogenous BRCA1 (green). As shown inFigure 1(B), trBRCA1 can be expressed in vivo and

471
Figure 2. Increased radiosensitivity of cells expressing trBRCA1. Radiosensitivity analysis of HBL100 cells stably transfected with eithervector, trBRCA1 or full-length (fl) BRCA1. Results shown are an average of duplicate analyses.
synthesizes a protein that localizes in the cell cyto-plasm (red).
Expression of trBRCA1 in mammary epithelial cellsresults in an increase in radiosensitivity
To address the hypothesis that trBRCA1 disruptsBRCA1 function, one of the well established activ-ities of BRCA1 – response to DNA damage – wasassessed using a clonal radiosensitivity assay, whichmeasures the clonogenic capacity of cells after ex-posure to varying doses of ionizing radiation. Thisassay was performed on HBL100 cells stably trans-fected with either pZeoSV vector, pZeo trBRCA1or pZeo flBRCA1. Expression of these constructswas confirmed by RT-PCR utilizing transgene specificprimers, and western blotting using the MS110 anti-BRCA1 antibody (data not shown). Figure 2 showsthat cells transfected with trBRCA1 are much moresensitive to ionizing radiation than cells transfectedwith pZeoSV vector or full-length (fl) BRCA1. Thisresult is consistent with trBRCA1 limiting the func-tion of the DNA damage response pathway and thuswith BRCA1 function being compromised in thesecells.
Expression of trBrca1 in transgenic mice results in adelay in lactational mammary development
To study the physiological activity of trBRCA1, trans-genic mice were generated that expressed a truncated
mouse Brca1 cDNA, encoding amino acids 1-299, un-der the control of the MMTV promoter, which primar-ily directs expression to mammary epithelial cellsand is upregulated during pregnancy. This promoterhas been used extensively to study the role of vari-ous genes in mammary development and cancer (e.g.,Stewart et al., 1984 and reviewed in Amundadottiret al., 1996). The transgene is referred to as MMTV-trBrca1. Using standard procedures, two independentfounder lines that express MMTV-trBrca1 were gen-erated. Southern analysis of tail DNA and northernanalysis of mammary tissue confirms multiple integ-ration and expression of the transgene, respectively(Figures 3(B) and (C)). An approximately twofolddifference in transgene copy number relative to theloading control was noted between the two lines(Figure 3(B)). The expression levels were, however,comparable (data not shown).
Histological examination of all tissues was per-formed on mice through several stages of pregnancy(day 8, 12, 15, 18) and at 1 day post-partum. A delayin mammary gland development, reflecting an impair-ment of lobuloalveolar development, was observedduring pregnancies of both lines of transgenic mice(Figure 4(A)). At 8 days, controls (wild-type mice andnon-transgenic littermates) exhibited budding from theducts with early lobular formation accompanied bysome stromal induction (Figure 4(A)(b)), whereasin MMTV-trBrca1 mice, minimal budding from theductal system was present (Figure 4(A)(a)). At 12, 15and 18 days a normal progressive increase in lobular
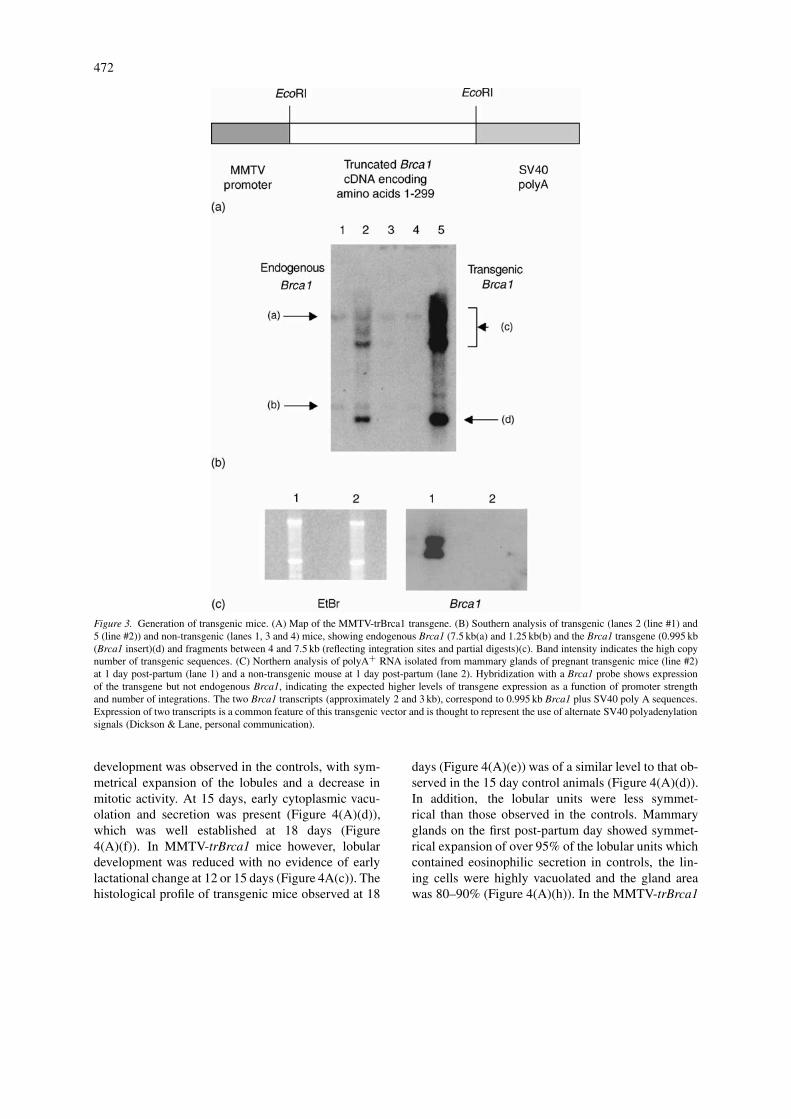
472
Figure 3. Generation of transgenic mice. (A) Map of the MMTV-trBrca1 transgene. (B) Southern analysis of transgenic (lanes 2 (line #1) and5 (line #2)) and non-transgenic (lanes 1, 3 and 4) mice, showing endogenous Brca1 (7.5 kb(a) and 1.25 kb(b) and the Brca1 transgene (0.995 kb(Brca1 insert)(d) and fragments between 4 and 7.5 kb (reflecting integration sites and partial digests)(c). Band intensity indicates the high copynumber of transgenic sequences. (C) Northern analysis of polyA+ RNA isolated from mammary glands of pregnant transgenic mice (line #2)at 1 day post-partum (lane 1) and a non-transgenic mouse at 1 day post-partum (lane 2). Hybridization with a Brca1 probe shows expressionof the transgene but not endogenous Brca1, indicating the expected higher levels of transgene expression as a function of promoter strengthand number of integrations. The two Brca1 transcripts (approximately 2 and 3 kb), correspond to 0.995 kb Brca1 plus SV40 poly A sequences.Expression of two transcripts is a common feature of this transgenic vector and is thought to represent the use of alternate SV40 polyadenylationsignals (Dickson & Lane, personal communication).
development was observed in the controls, with sym-metrical expansion of the lobules and a decrease inmitotic activity. At 15 days, early cytoplasmic vacu-olation and secretion was present (Figure 4(A)(d)),which was well established at 18 days (Figure4(A)(f)). In MMTV-trBrca1 mice however, lobulardevelopment was reduced with no evidence of earlylactational change at 12 or 15 days (Figure 4A(c)). Thehistological profile of transgenic mice observed at 18
days (Figure 4(A)(e)) was of a similar level to that ob-served in the 15 day control animals (Figure 4(A)(d)).In addition, the lobular units were less symmet-rical than those observed in the controls. Mammaryglands on the first post-partum day showed symmet-rical expansion of over 95% of the lobular units whichcontained eosinophilic secretion in controls, the lin-ing cells were highly vacuolated and the gland areawas 80–90% (Figure 4(A)(h)). In the MMTV-trBrca1

473
Figure 4.

474
Figure 4. Expression of truncated Brca1 in mouse mammary gland delays development during pregnancy. (A) Histological examination ofmammary glands from control mice at pregnancy day 8, 15, 18 and 1 day post-partum (b), (d), (f), (h) compared to corresponding areas intransgenic mice at the same time points (a), (c), (e), (g). Lactational activity is present in control mice at day 15 (d) but not in the mutants (c).Gland extent and development are considerably less at 18 days in transgenic mice (e) versus controls (f). In the first post-partum day, alveolardistension and secretion are less developed in the transgenic mice (g) whereas the control mice exhibit uniformly distended acini (h). (B) Wholemount preparations of thoracic mammary glands showing less volume and complexity of acini, with more asymmetric development, in theMMTV-trBrca1 mice at pregnancy day 15 (a) versus controls (b). In the 1st post-partum day, the alveolar units are less distended and subtleasymmetry is still present in the MMTV-trBrca1 mice (c) whereas the uniformity of the control lactating acini is striking (d).
mice, there was some irregularity in the distension ofthe lobular alveolar units and 10–15% did not containappreciable amounts of secretion, as judged by eosino-philic staining (Figure 4(A)(g)). More subtle changesincluded disorganization of the lobular alveolar devel-opment with asymmetry of the resulting units.
To substantiate and extend these observations,whole mount preparations of mammary glands wereanalyzed. Eight transgenic mice (one at pregnancy day15, five at pregnancy day 18 and two at 1 day post-partum) and five non-transgenic mice (one at day 15,three at day 18 and one at 1 day post-partum) wereanalyzed using this approach. A developmental delaywas clearly evident using this technique (Figure 4(B)),thus confirming the histological observations.
No mammary or other tumours were detected inany of the MMTV-trBrca1 mice, up to 18 monthsof age and through several pregnancies. No other
physiological changes were noted: adult mice ap-peared generally healthy, litter sizes were normal andthere was no obvious deficiency in the ability offemales to feed their young. No other histologicalchanges were noted in any of the other tissues ex-amined, including lung, heart, salivary gland, kidney,ureter, spleen, thyroid, adrenal gland, liver, pancreas,gall bladder, uterus, ovaries, oesophagus, stomach, in-testine, brain, bone, bone marrow and skeletal muscle.No changes of epithelial hyperplasia, epithelial atypiaor frank neoplasia were seen in mammary glands inany of the mice examined.
In summary, there was a clear delay in matura-tion of the lobular alveolar units and in lactationaldevelopment, maximally apparent at pregnancy day18 with a 2–3 day delay in development of the gland.Although subtle, these observations did not fall withinthe normal range of variation. Indeed, the differences

475
Figure 5. Expression of truncated Brca1 in mouse mammary glands results in downregulation of milk protein gene expression but no detectablechange in apoptosis. (A) Northern analysis of mammary gland RNA from day 14 (lanes 1 and 3) and day 18 (lanes 2 and 4) pregnantMMTV-trBrca1 transgenic mice (lanes 1 and 2) and littermate controls (lanes 3 and 4) for expression of β-casein (a) and Whey acidic protein(b). (B) TUNEL analysis of mammary glands from MMTV-trBrca1 transgenic mice (a) and littermate controls (b), (c) at day 1 of lactation (a),(b) or day 2 involution (c). TUNEL-positive cells are labelled with fluorescein (green), nuclei are counterstained with propidium iodide (red).Scale bar is 50 µm.
between our transgenic and control mice were clearlysignificant relative to those observed in a study oftransgenic MMTV-dnFGFR(111c) mice, in which we(G. W. H. S.) concluded no phenotypic effect (Jacksonet al., 1997). The phenotype of MMTV-trBrca1 micewas identical in the two independent founder linesexpressing comparable levels of trBrca1, indicatingthat the effect was specific to the transgene and nota position effect.
The observed delay in lactational mammarydevelopment is due to a defect in differentiation andnot altered proliferation or apoptosis
To elucidate the cellular nature of the phenotype in ourtransgenic mice, we examined cells and tissue sectionsfor evidence of altered proliferation, differentiationand apoptosis. Differentiation of mammary epithelialcells is accompanied by upregulation of a number ofgenes including the milk protein genes β-casein andwhey-acidic protein (WAP) and thus expression of
these genes is used as a marker of mammary differ-entiation (Robinson et al., 1995). Northern analysisof pregnant mammary glands indicated a downreg-ulation of β-casein at day 14 and WAP at day 18in MMTV-trBrca1 mice relative to littermate controls(Figure 5(A)). This result was reproducible in a totalof four independent sets of mice. This suggests thatexpression of trBrca1 results in defective differenti-ation of mammary epithelia. Proliferation rates weredetermined by triplicate cell counting using mam-mary epithelial cell-lines stably expressing trBrca1(relative to vector only control). Primary mammaryepithelial cells from pregnant transgenic mice werealso observed for any evidence of a proliferative ad-vantage relative to those from control animals. Nodifference in proliferation was observed in either cell-lines or primary cells (data not shown). To addressthe possibility that the phenotype reflected an increasein apoptosis, TUNEL studies were performed usingmammary gland sections taken from MMTV-trBrca1and control animals at day 1 of lactation. No TUNEL

476
positive cells were detected in either the transgenic orcontrol mice (Figure 5(B)). Gland sections from miceundergoing mammary involution were used as positivecontrols.
Discussion
We have shown that a truncated BRCA1 protein(trBRCA1) is stable, is localized in the cell cyto-plasm, and maintains the appropriate 3D structureto interact with anti-N-term BRCA1 antibodies. Inmammary cell-lines this protein confers an increase inradiosensitivity, consistent with abrogation of a DNAdamage response pathway and thus with BRCA1 func-tion. This finding, therefore, suggests that trBRCA1possesses dominant-negative activity. A dominant-negative hypothesis is supported by other in vitrostudies showing that certain mutants of BRCA1 (i)possess transforming activity (our unpublished data)and block BRCA1-mediated tumour suppressor activ-ity (Fan et al., 2001); (ii) can alter cell-cycle control(Larson et al., 1997); (iii) can interfere with BRCA1-mediated stimulation of p53 activity (Zhang et al.,1998) and inhibition of ER transcriptional activity(Fan et al., 2001); (iv) can block BRCA1-mediatedapoptosis (Thangaraju et al., 2000; Fan et al., 2001);and (v) that lymphocytes taken from carriers of asingle-allele germline BRCA1 mutation display an in-creased sensitivity to ionizing radiation (Foray et al.,1999).
With regard to the underlying molecular mech-anism of trBrca1 activity, a reasonable hypothesis isthat trBRCA1 sequesters BARD1 in the cytoplasm,limiting its previously described function in RNAprocessing (Kleiman & Manley, 2001), and proteinubiquitination (Hashizume et al., 2001). The findingthat trBRCA1 can interact with BARD1 (Wu et al.,1996, our unpublished data) supports this hypothesis.
Expression of trBrca1 in the mammary glandof transgenic mice results in a delay in lactationalmammary development. Interestingly a similar phen-otype is seen in mammary-specific conditional knock-outs of Brca1 (Xu et al., 1999), further supportinga dominant negative model. These findings indic-ate a critical role for Brca1 in the regulation ofmammary development. Such a role for Brca1 isfurther supported by the observations that Brca1 isexpressed in mouse mammary epithelial cells andupregulated during pregnancy (Lane et al., 1995;Marquis et al., 1995), is induced in ovariectomizedmice in response to exogenous ovarian hormones
(Marquis et al., 1995), and can be regulated in vitro, al-beit indirectly, by estrogen (Gudas et al., 1995; Markset al., 1997).
To address the cellular basis of this phenotype wehave analyzed stably transfected mammary epithelialcells and primary mammary epithelial cells from ourtransgenic mice. Expression levels of the genes encod-ing the mammary differentiation markers β-casein andWAP was reduced in MMTV-trBrca1 mice relative tocontrols (Figure 5(A)). This is consistent with the re-duced amount of secretion, as judged by eosinophilicstaining, observed in a proportion of alveoli (Fig-ure 4(A)) and suggests that the observed abnormalityin mammary development conferred by TrBrca1 re-flects a defect in differentiation. This conclusion isvindicated in our studies using an in vitro model forprolactin-induced mammary differentiation (Manninget al., manuscript in preparation). There was no changein the growth rate of either transfected cell-lines orprimary mammary epithelial cells from mice expres-sing TrBrca1 (data not shown). Consistent with this,is the finding that there is no difference in the levelsof expression of the cell cycle control proteins p21,cyclin D1 or cyclin E or amount of BrDU uptake intransgenic versus control mammary cells (Deans et al.,manuscript in preparation). Similar results have beenreported in independent studies of TrBRCA1 (Fanet al., 2001). TUNEL analysis indicates no detectableincrease in apoptosis in MMTV-trBrca1 mice at day1 of lactation (Figure 5(B)). Fan et al. also show noincrease in apoptosis in cells expressing TrBRCA1,indeed, they show an inhibition of drug-induced ap-optosis (Fan et al., 2001). This is in contrast to cellsin which BRCA1 function has been completely ab-lated (Xu et al., 1999) and may, therefore, reflect thefact that in cells expressing TrBRCA1 only a subsetof interacting proteins are sequestered and thus only asubset of functions limited.
None of our mice developed tumours. We believethat this indicates that introduction of trBrca1 alone,on the strain of mice used, is insufficient to inducetumorigenesis. This hypothesis is consistent with thecaretaker/gatekeeper model of tumour suppressor genefunction (Brugarolas & Jacks, 1997; Kinzler & Vogel-stein, 1997) and also with the paucity of tumours inthe Brca1 conditional knockout mice (Xu et al., 1999).The lack of tumours in our mice, in contrast to theconditional knockout mice, may reflect the differencein mouse strain used. Certainly mouse backgroundstrain has proved to be a contributing factor to tumori-genesis in mice expressing a truncated Brca2 protein

477
(Connor et al., 1997). To further address this we arecurrently backcrossing these mice onto other back-ground strains and studying the effect of concomitantgatekeeper mutation.
The observed effects of truncated BRCA1 haveimplications for germline carriers of BRCA1 muta-tions. This issue has been raised previously, with theobservation that lymphocytes from BRCA1 mutationcarriers are radiosensitive (Foray et al., 1999). Thefinding that BRCA1 mutations carriers also breast-feedtheir first infant for significantly shorter periods oftime (Jernstrom et al., 1998) further corroborates thesignificance of our findings and indicates a need toinvestigate this issue further.
Acknowledgements
The authors are very grateful to Vera Fantl and CliveDickson for advice on the generation and analysis oftransgenic mice, Ralph Scully for aliquots of anti-BRCA1 antibodies, Mary Seller for advice on stagingmouse embryos in order to confirm gestation dates,and Tracy Crafton, Sandra Peak, Karen Robinson andMax Walker for animal husbandry. Thanks also toDavid Grimwade and Aurélie Catteau for helpful dis-cussions and critical reading of the manuscript. Thiswork was supported by the Imperial Cancer ResearchFund, the Medical Research Council (UK), the Anti-Cancer Council of Victoria, Australia, the Queens-land Cancer Fund, Australia and the Selby ScientificFoundation.
References
Amundadottir LT, Merlino G and Dickson RB (1996) Transgenicmouse models of breast cancer. Breast Cancer Res Treat 39:119–135.
Brugarolas J and Jacks T (1997) Double indemnity: p53, BRCAand cancer. p53 mutation partially rescues developmental ar-rest in Brca1 and Brca2 null mice, suggesting a role for fa-milial breast cancer genes in DNA damage repair. Nat Med 3:721–722.
Connor F, Bertwistle D, Mee PJ, Ross GM, Swift S, Grigorieva Eet al. (1997) Tumorigenesis and a DNA repair defect in mice witha truncating Brca2 mutation. Nat Genet 17: 423–430.
Deng CX and Brodie SG (2000) Roles of BRCA1 and its interactingproteins. Bioessays 22: 728–737.
Englert C, Vidal M, Maheswaran S, Ge Y, Ezzell RM, IsselbacherKJ et al. (1995) Truncated WT1 mutants alter the subnuclearlocalization of the wild-type protein. Proc Natl Acad Sci USA92: 11960–11964.
Fan S, Yuan R-Q, Ma YX, Meng Q, Goldberg ID and Rosen EM(2001) Mutant BRCA1 genes antagonize phenotype of wild-typeBRCA1. Oncogene 20: 8215–8235.
Foray N, Randrianarison V, Marot D, Perricaudet M, Lenoir G andFeunteun J (1999) Gamma-rays-induced death of human cellscarrying mutations of BRCA1 or BRCA2. Oncogene 18: 7334–7342.
Gudas JM, Nguyen H, Li T and Cowan KH (1995) Hormone-dependent regulation of BRCA1 in human breast cancer cells.Cancer Res 55: 4561–4565.
Haber DA, Timmers HT, Pelletier J, Sharp PA and Housman DE(1992) A dominant mutation in the Wilms tumor gene WT1cooperates with the viral oncogene E1A in transformation ofprimary kidney cells. Proc Natl Acad Sci USA 89: 6010–6014.
Harvey M, Vogel H, Morris D, Bradley A, Bernstein A and Done-hower LA (1995) A mutant p53 transgene accelerates tumourdevelopment in heterozygous but not nullizygous p53-deficientmice. Nat Genet 9: 305–311.
Jackson D, Bresnick J, Rosewell I, Crafton T, Poulsom R, StampG et al. (1997) Fibroblast growth factor receptor signalling hasa role in lobuloalveolar development of the mammary gland. JCell Sci 110: 1261–1268.
Jernstrom H, Johannsson O, Borg A, Ivarsson H and Olsson H(1998) BRCA1-positive patients are small for gestational agecompared with their unaffected relatives. Eur J Cancer 34:368–371.
Kern SE, Pietenpol JA, Thiagalingam S, Seymour A, Kinzler KWand Vogelstein B (1992) Oncogenic forms of p53 inhibit p53-regulated gene expression. Science 256: 827–830.
Kinzler KW and Vogelstein B (1997) Cancer-susceptibility genes.Gatekeepers and caretakers. Nature 386: 761–763.
Lane TF, Deng C, Elson A, Lyu MS, Kozak CA and Leder P (1995)Expression of Brca1 is associated with terminal differentiation ofectodermally and mesodermally derived tissues in mice. GenesDev 9: 2712–2722.
Larson JS, Tonkinson JL and Lai MT (1997) A BRCA1 mutant al-ters G2-M cell cycle control in human mammary epithelial cells.Cancer Res 57: 3351–3355.
Lindeman GJ, Wittlin S, Lada H, Naylor MJ, Santamaria M,Zhang JG et al. (2001) SOCS1 deficiency results in acceleratedmammary gland development and rescues lactation in prolactinreceptor-deficient mice. Genes Dev 15: 1631–1636.
Marks JR, Huper G, Vaughn JP, Davis PL, Norris J, McDonnellDP et al. (1997) BRCA1 expression is not directly responsiveto estrogen. Oncogene 14: 115–121.
Marquis ST, Rajan JV, Wynshaw-Boris A, Xu J, Yin GY, Abel KJet al. (1995) The developmental pattern of Brca1 expression im-plies a role in differentiation of the breast and other tissues. NatGenet 11: 17–26.
Morgan SE, Lovly C, Pandita TK, Shiloh Y and Kastan MB(1997) Fragments of ATM which have dominant-negative orcomplementing activity. Mol Cell Biol 17: 2020–2029.
Nicolaides NC, Littman SJ, Modrich P, Kinzler KW and VogelsteinB (1998) A naturally occurring hPMS2 mutation can confer adominant negative mutator phenotype. Mol Cell Biol 18: 1635–1641.
Pullan SE and Streuli CH (1996) The mammary gland epithelialcells. In: Harris A (ed.), Epithelial Cell Culture. (pp 97–121)Cambridge University Press.
Robinson GW, McKnight RA, Smith GH and Hennighausen L(1995) Mammary epithelial cells undergo secretory differenti-ation in cycling virgins but require pregnancy for the establish-ment of terminal differentiation. Development 121: 2079–2090.
Stewart T, Pattengale PK and Leder P (1984) Spontaneous mam-mary adenocarcinomas in transgenic mice that carry and expressMTV/myc fusion genes. Cell 38: 627–637.

478
Thangaraju M, Kaufmann SH and Couch FJ (2000) BRCA1 facil-itates stress-induced apoptosis in breast and ovarian cancer celllines. J Biol Chem 275: 33487–33496.
Xu X, Wagner KU, Larson D, Weaver Z, Li C, Ried T et al. (1999)Conditional mutation of Brca1 in mammary epithelial cells
results in blunted ductal morphogenesis and tumour formation.Nat Genet 22: 37–43.
Zhang H, Somasundaram K, Peng Y, Tian H, Bi D, Weber BL et al.(1998) BRCA1 physically associates with p53 and stimulates itstranscriptional activity. Oncogene 16: 1713–1721.
Copyright © 2022 FDOKUMEN




















