
Expression of a dominant-negative mutant inhibitor-kappaBalpha of nuclear factor-kappaB in human...
8
[CANCER RESEARCH 59, 3468 –3474, July 15, 1999] Expression of a Dominant-Negative Mutant Inhibitor-kBa of Nuclear Factor-kB in Human Head and Neck Squamous Cell Carcinoma Inhibits Survival, Proinflammatory Cytokine Expression, and Tumor Growth in Vivo 1 Dianne C. Duffey, Zhong Chen, Gang Dong, Frank G. Ondrey, Jeffery S. Wolf, Keith Brown, Ulrich Siebenlist, and Carter Van Waes 2 Tumor Biology Section, Head and Neck Surgery Branch, National Institute on Deafness and Other Communication Disorders [D. D., Z. C., G. D., F. G. O., J. S. W., C. V. W.] and Laboratory of Immunoregulation, National Institute of Immunology, Allergy, and Infectious Diseases [K. B., U. S.], NIH, Bethesda, Maryland 20892 ABSTRACT We demonstrated recently that constitutive expression of proinflam- matory cytokines interleukin (IL)-1a, IL-6, IL-8, and granulocyte- macrophage colony-stimulating factor in head and neck squamous cell carcinoma is correlated with activation of transcription factor nuclear factor (NF)-kB/Rel A (p50/p65), which binds the promoter region within each of the genes encoding this repertoire of cytokines. NF-kB can be activated after signal-dependent phosphorylation and degradation of in- hibitor-kBa and has been reported to promote cell survival and growth. In the present study, we expressed a phosphorylation site mutant of inhibitor-kBa (IkBaM) in head and neck squamous cell carcinoma lines UM-SCC-9, -11B, and -38 to determine the effect of inhibition of NF-kB on cytokine expression, cell survival in vitro, and growth in vivo. After transfection with IkBaM, only a few UM-SCC-9 clones were obtained that stably expressed the mutant IkB, suggesting that expression of a mutant IkBa may affect survival of the transfected UM-SCC cell lines. After cotransfection of IkBaM with a Lac-Z reporter, we found that the num- ber of surviving b-galactosidase-positive cells in the three cell lines was reduced by 70 –90% when compared with controls transfected with vector lacking the insert. In UM-SCC-9 cells that stably expressed IkBaM, inhibition of constitutive and tumor necrosis factor-a induced NF-kB activation, and production of all four cytokines was observed. Although UM-SCC-9 IkBaM-transfected cells proliferated at the same rate as vector-transfected cells in vitro, a significant reduction in growth of tumor xenografts was observed in SCID mice in vivo. The decreased growth of UM-SCC-9 IkBaM-transfected tumor cells accompanied decreased im- munohistochemical detection of the activated form of NF-kB in situ. These results provide evidence that NF-kB and IkBa play an important role in survival, constitutive and inducible expression of proinflammatory cyto- kines, and growth of squamous cell carcinoma. NF-kB could serve as a potential target for therapeutic intervention against cytokine and other immediate-early gene responses that contribute to the survival, growth, and pathogenesis of these cancers. INTRODUCTION Autonomous production of proinflammatory cytokines has been detected in many different types of human and murine cancers, suggesting that their expression or the mechanism underlying their activation may provide a selective survival or growth advantage to cancer cells. We and others have observed previously that human head and neck SCCs 3 autonomously produce proinflammatory cytokines IL-1a, IL-6, IL-8, and GM-CSF in vitro and in vivo (1–5), and these cytokines have also been detected in tumor specimens and cell lines from patients with SCCs arising within the lungs, cervix, and skin (6 – 8). We reported that transformed murine keratinocytes that ex- press homologues of IL-1a, IL-6, IL-8, and GM-CSF at increased levels exhibit more aggressive growth and metastasis in vivo (9 –11). IL-1a has been shown to promote autocrine proliferation of trans- formed human keratinocytes and cervical SCC cell lines (12). Ele- vated expression of IL-8 by human lung SCC has been shown to promote angiogenesis and tumor growth in experimental models in vivo (8). Expression of GM-CSF has been associated with homing of CD341 granulocyte precursors that promote increased metastatic potential of human and murine SCC of the upper aerodigestive tract (13). Both IL-1 and IL-6 have been implicated in the increase in catabolism often observed in patients with these cancers (14). The molecular mechanisms that contribute to activation of this repertoire of cytokines in SCC have not been defined. The expression of proinflammatory cytokines can be induced in response to a variety of stimuli through activation of transcription factor NF-kB/Rel A (p50/p65), which is a heterodimer formed by members of the Rel family of proteins (15). We noted that the 59 promoter region of the IL-1a, IL-6, IL-8, and GM-CSF genes ex- pressed by HNSCC contain binding sites for the NF-kB/Rel family of transcription factors (16 –19), and we showed recently that the auton- omous expression of proinflammatory cytokines in HNSCC is corre- lated with constitutive activation of NF-kB/Rel A p50/p65 (20). Constitutive activation of NF-kB/Rel family members has also been demonstrated recently in other laboratories in several different neo- plasms, including lymphoma, melanoma, and carcinoma of the colon and breast (21–26). Members of the Rel family were originally iden- tified as viral and cellular oncogenes (27, 28). At present, the mech- anisms by which they can promote tumor growth have not been fully elucidated. Activation of NF-kB has been shown to involve signal-induced phosphorylation and degradation of cytoplasmic IkB proteins, which release NF-kB for nuclear translocation (29 –32), and for binding to the promoter sites of target genes. Studies in several laboratories have shown that mutations in the serine phosphoryl- ation sites at S32 and/or S36 of IkBa can inhibit the signal- dependent activation of NF-kB to a variety of stimuli (29 –32). Such phosphorylation mutants can therefore exert a dominant- negative effect, preventing the activation of NF-kB-dependent genes. By expression of a dominant-negative IkBa mutant, NF-kB has been shown to be important in activation of genes necessary for survival and protection of cells from injury by a variety of cyto- toxic stimuli, including cytokine TNF-a, chemotherapy, and radi- ation therapy (33–35). In the present study, we provide evidence that the expression of a phosphorylation mutant of IkBa (IkBaM) can reduce survival, constitutive and inducible activation of NF-kB, expression of proinflammatory cytokines, and tumor growth in vivo. Received 2/17/99; accepted 5/14/99. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 This work was supported by National Institute on Deafness and Other Communica- tion Disorders Intramural Research Project Z01-DC-00016. 2 To whom requests for reprints should be addressed, at National Institute on Deafness and Other Communication Disorders, Building 10, Room 5D55, MSC-1419, 10 Center Drive, Bethesda, MD 20892. E-mail: [email protected]. 3 The abbreviations used are: SCC, squamous cell carcinoma; HNSCC, head and neck SCC; IL, interleukin; GM-CSF, granulocyte-macrophage colony-stimulating factor; IkB, inhibitor kB; NF-kB, nuclear factor kB; TNF, tumor necrosis factor; X-gal, 5-bromo-4- chloro-3-indolyl-b-D-galactopyranoside; MTT, methyl thiazol tetrazolium salt; SCID, severe combined immunodeficient; b-gal, b-galactosidase. 3468 Research. on March 26, 2016. © 1999 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Expression of a dominant-negative mutant inhibitor-kappaBalpha of nuclear factor-kappaB in human...

[CANCER RESEARCH 59, 3468–3474, July 15, 1999]
Expression of a Dominant-Negative Mutant Inhibitor-kBa of Nuclear Factor-kB inHuman Head and Neck Squamous Cell Carcinoma Inhibits Survival,Proinflammatory Cytokine Expression, and Tumor Growth in Vivo1
Dianne C. Duffey, Zhong Chen, Gang Dong, Frank G. Ondrey, Jeffery S. Wolf, Keith Brown, Ulrich Siebenlist, andCarter Van Waes2
Tumor Biology Section, Head and Neck Surgery Branch, National Institute on Deafness and Other Communication Disorders [D. D., Z. C., G. D., F. G. O., J. S. W., C. V. W.] andLaboratory of Immunoregulation, National Institute of Immunology, Allergy, and Infectious Diseases [K. B., U. S.], NIH, Bethesda, Maryland 20892
ABSTRACT
We demonstrated recently that constitutive expression of proinflam-matory cytokines interleukin (IL)-1 a, IL-6, IL-8, and granulocyte-macrophage colony-stimulating factor in head and neck squamous cellcarcinoma is correlated with activation of transcription factor nuclearfactor (NF)-kB/Rel A (p50/p65), which binds the promoter region withineach of the genes encoding this repertoire of cytokines. NF-kB can beactivated after signal-dependent phosphorylation and degradation of in-hibitor- kBa and has been reported to promote cell survival and growth.In the present study, we expressed a phosphorylation site mutant ofinhibitor- kBa (IkBaM) in head and neck squamous cell carcinoma linesUM-SCC-9, -11B, and -38 to determine the effect of inhibition of NF-kBon cytokine expression, cell survivalin vitro, and growth in vivo. Aftertransfection with I kBaM, only a few UM-SCC-9 clones were obtained thatstably expressed the mutant IkB, suggesting that expression of a mutantIkBa may affect survival of the transfected UM-SCC cell lines. Aftercotransfection of IkBaM with a Lac-Z reporter, we found that the num-ber of surviving b-galactosidase-positive cells in the three cell lines wasreduced by 70–90% when compared with controls transfected with vectorlacking the insert. In UM-SCC-9 cells that stably expressed IkBaM,inhibition of constitutive and tumor necrosis factor-a induced NF-kBactivation, and production of all four cytokines was observed. AlthoughUM-SCC-9 IkBaM-transfected cells proliferated at the same rate asvector-transfected cellsin vitro, a significant reduction in growth of tumorxenografts was observed in SCID micein vivo. The decreased growth ofUM-SCC-9 IkBaM-transfected tumor cells accompanied decreased im-munohistochemical detection of the activated form of NF-kB in situ. Theseresults provide evidence that NF-kB and IkBa play an important role insurvival, constitutive and inducible expression of proinflammatory cyto-kines, and growth of squamous cell carcinoma. NF-kB could serve as apotential target for therapeutic intervention against cytokine and otherimmediate-early gene responses that contribute to the survival, growth,and pathogenesis of these cancers.
INTRODUCTION
Autonomous production of proinflammatory cytokines has beendetected in many different types of human and murine cancers,suggesting that their expression or the mechanism underlying theiractivation may provide a selective survival or growth advantage tocancer cells. We and others have observed previously that human headand neck SCCs3 autonomously produce proinflammatory cytokines
IL-1a, IL-6, IL-8, and GM-CSFin vitro and in vivo (1–5), and thesecytokines have also been detected in tumor specimens and cell linesfrom patients with SCCs arising within the lungs, cervix, and skin(6–8). We reported that transformed murine keratinocytes that ex-press homologues of IL-1a, IL-6, IL-8, and GM-CSF at increasedlevels exhibit more aggressive growth and metastasisin vivo (9–11).IL-1a has been shown to promote autocrine proliferation of trans-formed human keratinocytes and cervical SCC cell lines (12). Ele-vated expression of IL-8 by human lung SCC has been shown topromote angiogenesis and tumor growth in experimental modelsinvivo (8). Expression of GM-CSF has been associated with homing ofCD341 granulocyte precursors that promote increased metastaticpotential of human and murine SCC of the upper aerodigestive tract(13). Both IL-1 and IL-6 have been implicated in the increase incatabolism often observed in patients with these cancers (14). Themolecular mechanisms that contribute to activation of this repertoireof cytokines in SCC have not been defined.
The expression of proinflammatory cytokines can be induced inresponse to a variety of stimuli through activation of transcriptionfactor NF-kB/Rel A (p50/p65), which is a heterodimer formed bymembers of the Rel family of proteins (15). We noted that the 59promoter region of theIL-1a, IL-6, IL-8, and GM-CSF genes ex-pressed by HNSCC contain binding sites for the NF-kB/Rel family oftranscription factors (16–19), and we showed recently that the auton-omous expression of proinflammatory cytokines in HNSCC is corre-lated with constitutive activation of NF-kB/Rel A p50/p65 (20).Constitutive activation of NF-kB/Rel family members has also beendemonstrated recently in other laboratories in several different neo-plasms, including lymphoma, melanoma, and carcinoma of the colonand breast (21–26). Members of the Rel family were originally iden-tified as viral and cellular oncogenes (27, 28). At present, the mech-anisms by which they can promote tumor growth have not been fullyelucidated.
Activation of NF-kB has been shown to involve signal-inducedphosphorylation and degradation of cytoplasmic IkB proteins,which release NF-kB for nuclear translocation (29 –32), and forbinding to the promoter sites of target genes. Studies in severallaboratories have shown that mutations in the serine phosphoryl-ation sites at S32 and/or S36 of IkBa can inhibit the signal-dependent activation of NF-kB to a variety of stimuli (29 –32).Such phosphorylation mutants can therefore exert a dominant-negative effect, preventing the activation of NF-kB-dependentgenes. By expression of a dominant-negative IkBa mutant, NF-kBhas been shown to be important in activation of genes necessary forsurvival and protection of cells from injury by a variety of cyto-toxic stimuli, including cytokine TNF-a, chemotherapy, and radi-ation therapy (33–35). In the present study, we provide evidencethat the expression of a phosphorylation mutant of IkBa (IkBaM)can reduce survival, constitutive and inducible activation ofNF-kB, expression of proinflammatory cytokines, and tumorgrowth in vivo.
Received 2/17/99; accepted 5/14/99.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 This work was supported by National Institute on Deafness and Other Communica-tion Disorders Intramural Research Project Z01-DC-00016.
2 To whom requests for reprints should be addressed, at National Institute on Deafnessand Other Communication Disorders, Building 10, Room 5D55, MSC-1419, 10 CenterDrive, Bethesda, MD 20892. E-mail: [email protected].
3 The abbreviations used are: SCC, squamous cell carcinoma; HNSCC, head and neckSCC; IL, interleukin; GM-CSF, granulocyte-macrophage colony-stimulating factor; IkB,inhibitor kB; NF-kB, nuclear factorkB; TNF, tumor necrosis factor; X-gal, 5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside; MTT, methyl thiazol tetrazolium salt; SCID,severe combined immunodeficient;b-gal, b-galactosidase.
3468
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

MATERIALS AND METHODS
Cell Culture and Transfection of UM-SCC Cells with IkBaM andControl Vector. HNSCC cell lines UM-SCC-9, -11B, and -38 were obtainedfrom the well characterized UM-SCC series maintained by Dr. T. E. Carey(University of Michigan, Ann Arbor, MI; Ref. 36). Human SCC cell lineUM-SCC-9 was derived from a patient with a stage T2 SCC of the pharynx;UM-SCC11B was obtained from a patient, 65 years of age, with a T2N2a SCCof the hypopharynx; and UM-SCC-38 was cultured from a patient, 60 years ofage, with a T2N2a SCC of the oropharynx. The conditions for culture, karyo-type, and epithelial marker analysis of UM-SCC have been described previ-ously (36–38). The cell lines were tested and found to be free ofMycoplasmausing a Gibco MycoTect Kit (Life Technologies, Inc., Gaithersburg, MD).
The cDNA plasmid pCMX IkBaM contains a mutation at S36 of theNH2-terminus and a COOH-terminal PEST sequence mutation and was agenerous gift from Dr. Inder M.Verma (Salk Institute, La Jolla, CA; Ref. 35).The plasmid containing theneomycin(neo) resistance gene used is describedby Brown et al. (29). UM-SCC-9, -11B, and -38 cells were plated in 6-wellplates and transfected with 0.2mg of plasmid containingneomycinresistancegene, 2mg of plasmid DNA of either pCMVLacZ, pCMXIkBaM, or pCMXvector alone, and 20mg of LipofectAMINE (Life Technologies, Inc.) inserum-free medium. Expression of Lac-Z by X-gal staining and IkBaM byWestern blot was confirmed 72 h after transfection, and selection was carriedout in medium containing G418 sulfate (Life Technologies, Inc.) at a concen-tration of 1.2 mg/ml. After 2–3 weeks, clonal selection was performed bylimiting dilution, and individual clones were expanded and screened for IkBa
expression by Western blot analysis. Clones UM-SCC-9 I11 and I13, whichexpressed IkBaM most abundantly, and clones C11 and C13 transfected withvector control alone were expanded and used for additional studies. Weobserved that sustained expression of IkBaM in UM-SCC-9 cells was unstablein the absence of selective media, and we were unable to obtain stabletransformants of UM-SCC-11B or -38 expressing IkBaM from over 30 clonesplated from two independent transfections, despite evidence of transient ex-pression by Western blot at 72 h (data not shown).
b-Galactosidase Cell Viability Assay.UM-SCC cells growing in logphase were plated in six-well plates at 53 104 cells/well 1 day beforetransfection. Cultures were cotransfected with 0.5mg of pCMVLacZ and either2 mg of human IkBa mutant (S32GS36A, with NH2-terminal serine codon S32and S36 glycine and alanine substitutions) or the control vector PMT2T (39)with 16 ml of LipofectAMINE (Life Technologies, Inc.) in serum-free mediafor 5 h. Seventy-two h after transfection, the expression ofLacZgene by cellswas detected with X-gal staining procedure according to Saneset al. (40), andthe blue cells were counted after overnight staining under a microscope withmagnification of340. The average of theb-gal1 blue cells were calculatedfrom the counts in two representative fields in triplicate wells, and themeans6 SD from a representative experiment are presented.
Western Immunoblot Analysis of Murine and Endogenous HumanIkBa Expression. Expression of murine and human IkBa in transfectedUM-SCC-9 cells cultured in media or media containing 10,000 units of humanTNF-a (Knoll Pharmaceuticals, Parsippany, NJ) was compared by Westernimmunoblot analysis. Cell protein was extracted from cells grown to 80%confluency in T75 flasks by lysis in 50 mM Tris (pH 7.4), 100 mM NaCl, 50mM NaF, 30 mM sodium pyrophosphate, 0.5% NP40, and one Complete miniprotease inhibitor tablet (Boehringer Mannheim, Mannheim, Germany), fol-lowed by centrifugation at 14,0003 g for 15 min. The protein concentrationof extracts was determined using the BCA protein assay (Pierce, Rockford, IL),and proteins were separated by electrophoresis on 10% denaturing SDS-polyacrylamide gels (39) and transferred to nitrocellulose membrane (Bio-Rad,Hercules, CA) by electrophoresis (Hoeffer TE Transphor, San Francisco, CA).Immunostaining was performed as described (41) using polyclonal rabbitanti-mouse and anti-human IkBa (C-21; Santa Cruz Biotechnology, SantaCruz, CA), and goat anti-rabbit IgG-HRP conjugated antibody (Bio-Rad,Hercules, CA). Purified human IkBa (1–317; Santa Cruz Biotechnology),along with 20ml of MultiMark pigmented markers (Novex, San Diego, CA)were used as standards. Whole-cell extracts from the murine Pam LU-1 SCCcell line were used as a positive control for murine IkBa. Chemiluminescentdetection of horseradish peroxidase was obtained using the SuperSignal Sub-strate method (Pierce, Rockford, IL), followed by exposure on X-Omat ARfilm (Kodak, Rochester, NY).
Electromobility Shift Assay. EMSA were performed using standard meth-ods (42), and nuclear extracts were obtained from UM-SCC-9 in log phasegrowth using the methods of Dignamet al. (43) and Leeet al. (44). Double-stranded DNA oligonucleotide probes for NF-kB and OCT-1 were purchasedcommercially (Promega, Madison, WI and Santa Cruz Biotechnology). Theoligonucleotide sequences used were: NF-kB, 59-AGTTGAGGGGACTTTC-CCAGGC-39; and OCT-1, 59-TGTCGAATGCAA ATCACTAGAA-39. Themutant oligonucleotide double-strand probe used for a specificity control forNF-kB was 59- AGTTGAGGCGACTTTCCCAGGC-39. EMSA reactions in-cluded 5mg of nuclear extract protein incubated for 25 min at 20°C in buffercontaining 20 mM HEPES (pH 7.9), 4.6 mM MgCl2, 63 mM KCl, 11% glycerol,and 1 mM DTT, 1 mg poly(deoxyinosinic-deoxycytidylic acid) (Pharmacia,Piscataway, NJ), and 60,000 cpm of32P- labeled oligonucleotide probe. Forunlabeled and mutant probe competition, extracts were preincubated with a100-fold excess of each probe before the addition of labeled probe. Bindingcomplexes were resolved on 5%, 0.253 TBE polyacrylamide gel, and imagedwith a Packard Instant Imager (Packard Technologies, Downers Grove, IL).
NF-kB Luciferase Reporter Assay.The pIgkB Luc reporter constructcontaining two IgG-k chain NF-kB binding sites and the luciferase gene wasdescribed previously (45), and the pCMV Lac-Z reporter containing the CMVpromoter andLac-Z gene in pcDNA3 (Invitrogen, Carlsbad, CA) used fornormalization of the assay was prepared by Dr. G. Thomas in our laboratory.UM-SCC-9 cell cultures at 70% confluence were cotransfected with pIgkBLuc plasmid (2mg/ml) and pCMVLacZ (0.4mg/ml) in Opti-MEM mediacontaining 10mg/ml LipofectAMINE (Life Technologies, Inc., Gaithersburg,MD). After 4 h, the medium was removed, and cells were incubated incomplete media for 48 h. The relative luciferase activity was determined witha Dual Light Reporter Gene Assay kit (Tropix, Bedford, MA) with aMonolight 2010 luminometer (Analytical Luminescence Laboratories, SanDiego, CA).
Quantitation of Cytokine Secretion by ELISA. UM-SCC cells (33 106)were cultured in 12 ml of fresh medium in 75-cm2 flasks, and culture super-natants were collected after 5 days. Standardized ELISA assay kits were usedto quantify IL-1a, IL-6, IL-8, and GM-CSF (R&D Systems, Minneapolis,MN). The absorbance of the reaction product was quantitated with a mul-tichannel plate spectrophotometer at 450 nm (Biotek 311; Biotek Systems,Winooski, VT). All samples were assayed in triplicate. Results were standard-ized for amount of cytokine secreted as pg/106 cells/5 days6 SD.
MTT Assay. The cell proliferation rate of UM-SCC-9 and transfected cellsin vitro were compared by using the MTT reaction assay (Boehringer Mann-heim, Mannheim, Germany). Replicate cultures of each of the cell lines wereplated in 96-well, flat-bottomed microplates at an initial concentration of5 3 103 cells in 100-ml medium/well. On days 1–5, 10ml of MTT labelingagent were added, and cells were incubated for an additional 4 h. One hundredml of solubilization solution were added to lyse cells overnight. The absorb-ance of the colorimetric assay was determined at a wavelength of 570 nm usinga multichannel plate spectrophotometer (Biotek Instruments, Winooski, VT).
Growth of UM-SCC-9 Parental and Transfected Cell Xenografts inSCID Mice. UM-SCC-9 parental and transfected cell lines were inoculated inSCID mice at 4 weeks of age. UM-SCC-9, vector transfectants C11 and C13,and the IkBaM transfectants I11 and I13 cells in logarithmic growth phasewere harvested and injected s.c. into SCID mice, at a dose of 53 106 or1 3 107 cells. Three animals were injected for each cell line at each dose.Tumors were measured weekly. The data are presented as mean6 SE.Animals were sacrificed if the tumor exceeded 10% of the animal’s bodyweight or exceeded 2.5 cm2 in volume.
Immunohistochemical Staining of Activated NF-kB in SCC Tissue andCultured Cells. Immunohistochemical analysis of the activated form ofNF-kB p65 in tissue sections from UM-SCC-9 and transfectant tumor xe-nografts was performed using a modification of the protocol of Kaltschmidtetal. (46). Frozen tissue sections (10–12mm) from tumor xenograft specimenswere fixed and permeabilized in cold 1:1 methanol:acetone for 10 min, andendogenous peroxidase was quenched by incubation with 3% H2O2 in meth-anol for 30 min at room temperature. Sections were incubated with 10% horseserum in PBS (Vector Laboratories, Inc., Burlingame, CA) for 30 min, fol-lowed by incubation with 2mg/ml mouse monoclonal anti-human NF-kB p65diluted in a 1% BSA in PBS (Boeringer Mannheim, Mannheim, Germany).Onemg/ml of mouse anti-human pan cytokeratin K5, K6, K8, and K18 (IgG1;Novocastra Lab, Newcastle upon Tyne, United Kingdom) or isotype antibody
3469
NF-kB IN SQUAMOUS CELL CARCINOMA
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
(mouse IgG1; Dako, Carpenteria, CA) diluted in 1% BSA in PBS were used ascontrols. The samples were incubated with horse anti-mouse antibody (1:200in PBS containing 1.5% horse serum; Vector Lab), followed by a 30-minincubation with biotin/avidin horseradish peroxidase conjugates (VectastainElite ABC kit; Vector Lab), and an 8-min incubation with the chromogendiaminobenzidine tetrahydrochloride (Vector Lab). Stained sections were de-hydrated and mounted with Permount (Fisher Scientific, Fair Lawn, NewJersey).
RESULTS
Expression of a Phosphorylation Mutant of IkB in UM-SCCInhibits Survival of Transfected Cells. We transfected a phospho-rylation site-deficient mutant of murine IkBa (IkBaM, Ref. 35) or acontrol vector lacking an insert into head and neck squamous cellcarcinoma lines UM-SCC-9, -11B, and -38 to determine whether wecould obtain a UM-SCC line that stably expressed a dominant-nega-tive mutant of IkBa. After selection in G418 and cloning, we obtainedUM-SCC-9 cells expressing IkBaM, but we did not detect expressionof IkBaM in over 30 UM-SCC-11B or 38 clones plated from twoindependent transfections, despite similar transfection efficiencies andevidence of transient expression of mutant IkBa in transfected cul-tures by Western blot (data not shown). Because NF-kB/Rel transcrip-tion factors have been reported to regulate genes involved in cellularsurvival (21–25, 33–35), we examined the possibility that expressionof an IkBa mutant might reduce survival of transfected UM-SCCcells. UM-SCC-9, -11B, and -38 cell lines were cotransfected with aLac-Z reporter in the presence of a 4-fold excess of vector containinga human IkBa phosphorylation mutant (39) or control vector, and acomparison ofb-gal-positive cells in cultures was made 72 h aftertransfection.b-gal-positive blue staining cells were detected in allthree UM-SCC cell lines in the presence of control vector, at atransfection efficiency of 20–30%. However, Fig. 1 shows that aftercotransfection with the IkBa mutant, the number ofb-gal-positivecells was significantly reduced in the three cell lines by 70–90%.These data provide evidence that the difficulty obtaining stableIkBaM transfectants was due to decreased survival of UM-SCC cellstransfected with an inhibitor of NF-kB.
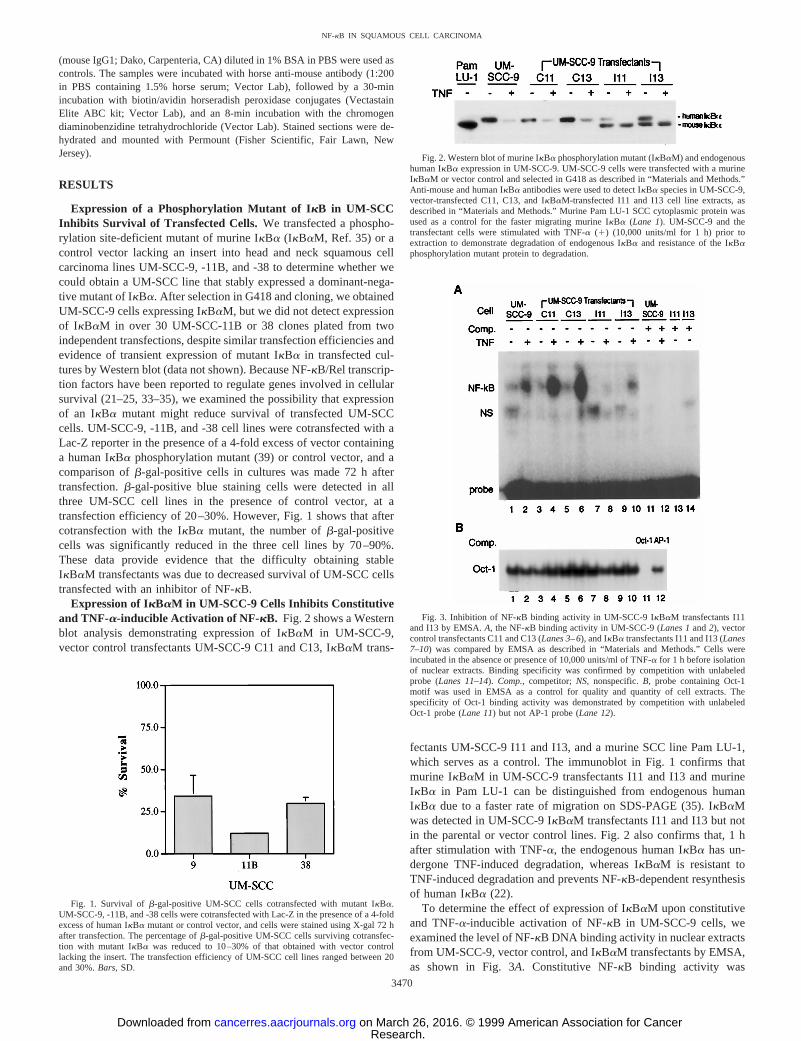
Expression of IkBaM in UM-SCC-9 Cells Inhibits Constitutiveand TNF-a-inducible Activation of NF-kB. Fig. 2 shows a Westernblot analysis demonstrating expression of IkBaM in UM-SCC-9,vector control transfectants UM-SCC-9 C11 and C13, IkBaM trans-
fectants UM-SCC-9 I11 and I13, and a murine SCC line Pam LU-1,which serves as a control. The immunoblot in Fig. 1 confirms thatmurine IkBaM in UM-SCC-9 transfectants I11 and I13 and murineIkBa in Pam LU-1 can be distinguished from endogenous humanIkBa due to a faster rate of migration on SDS-PAGE (35). IkBaMwas detected in UM-SCC-9 IkBaM transfectants I11 and I13 but notin the parental or vector control lines. Fig. 2 also confirms that, 1 hafter stimulation with TNF-a, the endogenous human IkBa has un-dergone TNF-induced degradation, whereas IkBaM is resistant toTNF-induced degradation and prevents NF-kB-dependent resynthesisof human IkBa (22).
To determine the effect of expression of IkBaM upon constitutiveand TNF-a-inducible activation of NF-kB in UM-SCC-9 cells, weexamined the level of NF-kB DNA binding activity in nuclear extractsfrom UM-SCC-9, vector control, and IkBaM transfectants by EMSA,as shown in Fig. 3A. Constitutive NF-kB binding activity was
Fig. 1. Survival ofb-gal-positive UM-SCC cells cotransfected with mutant IkBa.UM-SCC-9, -11B, and -38 cells were cotransfected with Lac-Z in the presence of a 4-foldexcess of human IkBa mutant or control vector, and cells were stained using X-gal 72 hafter transfection. The percentage ofb-gal-positive UM-SCC cells surviving cotransfec-tion with mutant IkBa was reduced to 10–30% of that obtained with vector controllacking the insert. The transfection efficiency of UM-SCC cell lines ranged between 20and 30%.Bars,SD.
Fig. 2. Western blot of murine IkBa phosphorylation mutant (IkBaM) and endogenoushuman IkBa expression in UM-SCC-9. UM-SCC-9 cells were transfected with a murineIkBaM or vector control and selected in G418 as described in “Materials and Methods.”Anti-mouse and human IkBa antibodies were used to detect IkBa species in UM-SCC-9,vector-transfected C11, C13, and IkBaM-transfected I11 and I13 cell line extracts, asdescribed in “Materials and Methods.” Murine Pam LU-1 SCC cytoplasmic protein wasused as a control for the faster migrating murine IkBa (Lane 1). UM-SCC-9 and thetransfectant cells were stimulated with TNF-a (1) (10,000 units/ml for 1 h) prior toextraction to demonstrate degradation of endogenous IkBa and resistance of the IkBaphosphorylation mutant protein to degradation.
Fig. 3. Inhibition of NF-kB binding activity in UM-SCC-9 IkBaM transfectants I11and I13 by EMSA.A, the NF-kB binding activity in UM-SCC-9 (Lanes 1and2), vectorcontrol transfectants C11 and C13 (Lanes 3–6), and IkBa transfectants I11 and I13 (Lanes7–10) was compared by EMSA as described in “Materials and Methods.” Cells wereincubated in the absence or presence of 10,000 units/ml of TNF-a for 1 h before isolationof nuclear extracts. Binding specificity was confirmed by competition with unlabeledprobe (Lanes 11–14). Comp.,competitor;NS, nonspecific.B, probe containing Oct-1motif was used in EMSA as a control for quality and quantity of cell extracts. Thespecificity of Oct-1 binding activity was demonstrated by competition with unlabeledOct-1 probe (Lane 11) but not AP-1 probe (Lane 12).
3470
NF-kB IN SQUAMOUS CELL CARCINOMA
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

observed in the UM-SCC-9 cell line and vector control transfectantsC11 and C13 and is further induced by addition of TNF-a (Fig. 3A,Lanes 1–6). The level of constitutive and TNF-a-induced NF-kBactivation is significantly reduced in UM-SCC-9 IkBaM transfectantsI11 and I13 (Fig. 3A, Lanes 7–10). The NF-kB DNA binding activityobserved in UM-SCC-9, C11, and C13 cells could be specificallyinhibited by competition with unlabeled NF-kB oligonucleotide (Fig.3A, Lanes 11–14). Fig. 3Bshows that expression of IkBaM did notinhibit DNA binding activity of the unrelated transcription factorOct-1 (Lanes 1–10), which could be blocked by unlabeled Oct-1 butnot AP-1 probe competition (Lanes 11and12).
To determine whether the reduction in NF-kB DNA binding activ-ity observed corresponded to a reduction in the functional activationof NF-kB, we examined NF-kB reporter activity in UM-SCC-9 vectorand IkBaM transfected clones. Fig. 4 confirms that functional acti-vation of NF-kB as determined by luciferase reporter assay is reduced;10-fold in the IkBaM transfectant I11 and I13 cells relative toUM-SCC-9 vector control transfectants C11 and C13. The reporteractivity was specific for NF-kB, because UM-SCC-9 cells transfectedwith a reporter containing a wild-type NF-kB sequence, but not amutant sequence, exhibited luciferase signal (data not shown).
Expression of Proinflammatory Cytokines IL-1a, IL-6, IL-8,and GM-CSF Is Inhibited in UM-SCC-9 Cells Transfected withIkBaM. We determined if expression of the IkBaM in UM-SCC-9resulted in reduced expression ofIL-1a, IL-6, IL-8, and GM-CSFcytokine genes that contain NF-kB sites in their 59 promoter regions(16–19). The quantity of each of these proinflammatory cytokinessecreted by vector control and IkBaM transfectants as determined byELISA is shown in Fig. 5. Secretion of all four of the proinflammatorycytokines was reduced in IkBaM transfectants I11 and I13 relative totwo vector control transfectants C11 and C13. Thus, expression byIkBaM in UM-SCC-9 inhibited activation of NF-kB and productionof proinflammatory cytokines IL-1a, IL-6, IL-8 and GM-CSF, indi-cating that NF-kB promotes constitutive expression of these proin-flammatory cytokines by UM-SCC-9.
Effect of Expression of IkBaM on Growth of UM-SCC-9 Cellsin Vitro and in Vivo. We compared the growth of UM-SCC-9IkBaM transfectants I11 and I13 and control vector transfectants C11and C13in vitro and as xenografts in SCID micein vivo. The growthof the cell linesin vitro was compared in a 5-day MTT assay asdescribed in “Materials and Methods,” and the results are shown inFig. 6A. There was no significant difference in growth betweenIkBaM transfectants I11 and I13 and control vector transfectants C11and C13in vitro. However, when SCID mice were inoculated s.c. with1 3 107 cells of IkBaM transfectants I11 and I13 or control vector
transfectants C11 and C13, the growth of the IkBaM transfectantswas significantly reduced when compared with vector-transfected celllines in two independent experiments (Fig. 6B). As shown in Fig. 6B,Experiment 1,we observed slower tumor growth of the IkBaM-transfected UM-SCC-9 I11 cell line when compared with vectortransfectant C11. We observed complete tumor regression of IkBaMtransfectant I13 at a tumor cell dose of 13 107 cells, whereas thecontrol line C13 grew progressively (Fig. 6B, Experiment 2). Similarbut smaller differences were obtained in both experiments in animalsinoculated with a lower dose of 53 106 cells (data not shown). Thus,expression of IkBaM in UM-SCC-9 cells inhibited tumor growthinvivo but did not inhibit tumor cell growthin vitro.
Inhibition of Activation of NF- kB in UM-SCC-9 IkBaM Trans-fectant Tumor Xenografts. To establish whether there is a relativedifference in constitutive activation of NF-kB in UM-SCC-9 andIkBaM transfectant tumor xenografts, we compared NF-kB activationin UM-SCC-9, C11, and I11 tumor specimensin situ by immunohis-tochemistry, using a monoclonal antibody specific for the nuclearlocalization sequence of the activated form of human NF-kB p65 (46).As shown in Fig. 7, staining for the activated form of NF-kB proteinwas observed in UM-SCC-9 (Fig. 7A) and vector transfectant C11cells (Fig. 7B) but was significantly decreased in IkBaM-transfectedI11 cells (Fig. 7C). All three tumor specimens stained positively withmonoclonal antibody against pan cytokeratin (Fig. 7,D–F) but notwith isotype-matched control antibody (Fig. 7,G–I). Thus, expressionof IkBaM inhibited constitutive activation of NF-kB in vivo as wellas in vitro.
DISCUSSION
In this study, we encountered difficulty in obtaining humanHNSCC lines that stably express a dominant-negative IkBa phos-phorylation site mutant, suggesting that expression of the mutantIkBa could affect survival of transfected UM-SCC cells. When thethree UM-SCC cell lines were cotransfected with a Lac-Z reporterin the presence of a 4-fold excess of vector containing a humanIkBa phosphorylation mutant or control vector, it was found thattransfection of mutant IkBa markedly reduced the survival of
Fig. 4. NF-kB reporter activity is reduced in the IkBaM transfectant relative toUM-SCC-9 vector control transfectants. Cells (23 105/well) were plated in six-wellplates the day before transfection. The cells were transfected by LipofectAMINE for 5 hwith IgkB-Luc and pCMVLacZ plasmids (5:1 ratio) in duplicate. Luciferase assay wasperformed within 48 h of transfection, and the relative light units were calculated.Bars,SD.p, significant difference between UM-SCC-9 C11 and C13versusIkBM transfectantsI11 and I13 (P , 0.05).
Fig. 5. Significant reduction of constitutive secretion of proinflammatory cytokinesIL-1a, IL-6, IL-8, and GM-CSF by IkBaM transfectantsI11 and I13 when compared withvector-transfected cell lines. ELISA was performed on supernatants of C11 and C13vector-transfected andI11 and I13 IkBaM-transfected cell lines, and the quantity ofcytokine was compared in pg/106 cells/5days.
3471
NF-kB IN SQUAMOUS CELL CARCINOMA
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

b-gal-positive cells by 70 –90% in cultures within 72 h (Fig. 1).These results are consistent with recent studies that show thatinhibition of activation or deletion of NF-kB/RelA inhibits sur-vival of a variety of normal and neoplastic cells of different tissueorigin (21–24, 33–35). The promotion of cell survival by NF-kBhas recently been attributed to several mediators. NF-kB has beenreported to induce TRAF1, TRAF2, c-IAP1, and c-IAP2, resultingin suppression of caspase-8 activation, thereby inhibiting apoptosis(47). A novel apoptosis inhibitor IEX-1 L has also been reported tobe involved in NF-kB-mediated cell survival (48). Recently, theprosurvival Bcl-2 homologue Bfl-1/A1 has been shown to be atranscriptional target of NF-kB, which can block TNF-a-inducedapoptosis (49). Although the pathways by which NF-kB promotessurvival in HNSCCs remain to be defined, our results and those ofothers suggest that NF-kB may be an important target for thera-peutic intervention against immediate-early responses that promotesurvival and resistance of cancer.
When we examined UM-SCC-9 cells that survive expression ofIkBaM, we found that IkBaM suppressed constitutive and TNF-a-induced activation of NF-kB, production of proinflammatorycytokines, as well as growthin vivo. We confirmed that expressionof IkBaM inhibited constitutive activation of NF-kB in UM-SCC-9 tumorsin situ. Whereas expression of IkBaM inhibited
growth of UM-SCC-9in vivo, there was no significant differencein growth between the IkBaM and vector control transfectantsinvitro. These results provide evidence that cytokines or other genesactivated by NF-kB in UM-SCC-9 promote interactions with thehost that contribute to malignant growthin vivo. These data areconsistent with observations made in our laboratory in an inde-pendent murine model of SCC, in which expression of the murinehomologues of these cytokines was found to be associated withmore aggressive growthin vivo but not in vitro (9 –11). We foundthat low cytokine-producing clones of the murine SCC line Pam212 grew slowly or regressed, whereas those that expressed thecytokine homologues IL-1a, IL-6, KC, and GM-CSF grew moreaggressivelyin vivo. However, no significant differences in pro-liferative rate were observedin vitro.
Among the NF-kB-induced cytokines expressed by human andmurine SCCs, there is evidence that the C-X-C family of chemokinesto which human IL-8 and murine KC belong can induce host angio-genesis responses and thereby provide a differential growth advantagein vivo. Smithet al. (8) have shown that expression of IL-8 by humanlung SCC can promote an increase in angiogenesis and growth oftumor xenografts in SCID mice. In murine SCC, we have obtainedpreliminary evidence that overexpression of murine cytokine KC inthe low KC expressing Pam 212 SCC line promotes an increase in
Fig. 6. Comparison of growth of UM-SCC-9cells expressing IkBaM in vitro and in vivo. A,UM-SCC-9 vector and IkBaM-transfected cloneswere plated at 53 103 cells/well in the 96-wellmicroplates, and MTT assay was performed on days1, 3, and 5 after initiation of cell culture. Data aremeans of quadruplicates of each condition;bars,SE.B, expression of IkBaM inhibits growth of UM-SCC-9 xenografts in SCID mice. Cells (107) ofUM-SCC-9 vector transfectants C11 and C13 andthe IkBaM transfectants I11 and I13 were s.c. in-jected into SCID mice (n5 3) in a volume of 0.1 ml.Tumors were measured weekly. The data were fromtwo representative experiments presented as meanarea (bars,SE), and similar results were observed inthe animal groups injected with 53 106 cells/mouse(not shown).
Fig. 7. Expression of IkBaM inhibits NF-kBactivation in tumor xenograftsin situ. Activation ofNF-kB in situ was determined by immunohisto-chemistry in frozen sections from the UM-SCC-9(A), vector transfected C11 (B), and IkBaM-trans-fected I11 (C) tumor xenografts using a murinemonoclonal antibody specific for activated humanNF-kB. Murine monoclonal antibody against hu-man pan cytokeratin was used as a positive controlfor human SCC cells (D–F), and isotype-matchedantibody was used as a negative control (G–I).Bars,SD.
3472
NF-kB IN SQUAMOUS CELL CARCINOMA
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

growth in vivo, whereas it has no effect on the rate of growthin vitro.4
The relative contribution of additional autocrine or paracrine mecha-nisms by which proinflammatory cytokines IL-1a, IL-6, GM-CSF,and others expressed by SCC may be found to promote growthremains to be determined. NF-kB and IkBa could serve as potentialtargets for therapy to inhibit expression of cytokine and other NF-kB-dependent genes that are ultimately found to contribute to the growthof SCCsin vivo.
Although NF-kB/Rel A and IkBa appear to be important in theregulation of expression of cytokines and other genes that promotegrowth in UM-SCC cell lines, it is possible that activation of otherpathways contribute to expression of these cytokines and other NF-kB-dependent genes. The inhibition of activation of cytokine expres-sion and growth by IkBa in UM-SCC-9 that we observed was lessthan complete. The promoter region of the cytokine genes expressedby SCC contains AP-1 and NF-IL6 as well as NF-kB binding se-quences (16–19), and coactivation of the promoter region of proin-flammatory cytokine genes by NF-kB, AP-1, and NF-IL6 has beenreported to have synergistic effects. Consistent with this possibility,both NF-kB and AP-1 have been reported to be activated and havetransforming activity in experimental murine tumors derived fromsquamous epithelia (50–52). Studies are under way to define theactivation, composition, and contribution of other immediate-earlytranscription factors that may contribute to the constitutive expressionof these cytokine genes in head and neck SCC. These studies shouldin turn facilitate the identification of additional genes that may pro-mote growth and malignant behavior of SCC.
The mechanism of constitutive activation of NF-kB/Rel A andIkBa in SCC and in other neoplasms remains to be determined. It willbe interesting to determine whether changes in expression or activa-tion of other signal transduction molecules upstream that lead toactivation of protective pathways, or inactivation of molecules thatinduce apoptosis, determine the requirement for activation of NF-kB.In addition to cytokine-induced activation, NF-kB and alternativepathways may be activated by alterations in epidermal growth factoror receptor expression, Ras activation, or through cell damage andchanges in oxidation-reduction state (26, 28, 53–55). Baldwin andcolleagues have shown previously that Bcr-Abl and Ras promoteactivation of NF-kB in leukemia, and an IkBa super-repressor caninhibit Bcr-Abl-mediated tumors in nude mice (56). Thus, F-kB couldserve as a common pathway for oncogenesis involving altered expres-sion or function of a variety of components of these pathways.
ACKNOWLEDGMENTS
The IkBaM construct provided by Dr. Inder Verma, the anti-NF-kB anti-body provided by Dr. Hubertus Stockinger of Boehringer Mannheim, andTNF-a provided by Dr. Melvin Spigelman of Knoll Pharmaceuticals areappreciated. The review of the original manuscript and helpful comments ofDrs. James B. Mitchell and James Mulshine of the National Cancer Instituteare gratefully acknowledged.
REFERENCES
1. Chen, Z., Colon, I., Ortiz, N., Callister, M., Dong, G., Pegram, M. Y., Arosarena, O.,Strome, S., Nicholson, J. C., and Van Waes, C. Effects of interleukin 1a, interleukin-1receptor antagonist and neutralizing antibody on proinflammatory cytokine expres-sion by human squamous cell carcinoma lines. Cancer Res.,58: 3668–3676, 1998.
2. Chen, Z., Malhotra, P. S., Thomas, G. R., Ondrey, F. G., Duffey, D. C., Smith, C. W.,Kroog, G. S., Enamorado, I., McCullagh, L., Rudy, S., Yeh, N. T., Mousa, S.,Quezado, M., Hersher, L., and Van Waes, C. Expression of proinflammatory and
proangiogenic cytokines in patients with head and neck cancer. Clin. Cancer Res., inpress, 1999.
3. Woods, K. V., El-Nagger, A., Clayman, G. L., and Grimm, E. A. Variable expressionof cytokines in human head and neck squamous cell carcinoma cell lines andconsistent expression in surgical specimens. Cancer Res.,58: 3132–3141, 1998.
4. Mann, E., Spiro, J., Chen, L., and Kreutzer, D. Cytokine expression by head and necksquamous cell carcinoma. Am. J. Surg.,164: 567–573, 1992.
5. Cohen, R., Contrino, J., Spiro, J., Mann, E., Chen, L., and Kreutzer, D. Interleukin-8expression by head and neck squamous cell carcinoma. Arch. Otolaryngol. HeadNeck Surg.,121: 202–209, 1995.
6. Yamamura, M., Modlin, R., Ohmen, J., and Moy, R. Local expression of anti-inflammatory cytokines in cancer. J. Clin. Invest.,91: 1005–1010, 1993.
7. Chopra, V., Dinh, T. V., and Hannigan, E. V. Circulating serum levels of cytokinesand angiogenic factors in patients with cervical cancer. Cancer Invest.,16: 152–159,1998.
8. Smith, D., Polverini, P., Kunkel, S., Orringer, M., Whyte, R., Burdick, M., Wilke, C.,and Strieter, R. Inhibition of interleukin 8 attenuates angiogenesis in bronchogeniccarcinoma. J. Exp. Med.,179: 1409–1415, 1994.
9. Chen, Z., Smith, C. W., Kiel, D., and Van Waes, C. Metastatic variants derivedfollowing in vivo tumor progression of anin vitro transformed squamous cellcarcinoma line acquire a differential growth advantage requiring tumor-host interac-tion. Clin. Exp. Metastasis,15: 527–537, 1997.
10. Dong, G., Loukinova, E., Smith, C. W., Chen, Z., and Van Waes, C. Genes differ-entially expressed with malignant transformation and metastatic tumor progress on ofmurine squamous cell carcinoma. J. Cell. Biochem. Suppl.,28/29: 90–100, 1997.
11. Smith, C. W., Chen, Z., Dong, G., Loukinova, E., and Van Waes, C. The hostenvironment promotes the development of primary and metastatic squamous cellcarcinomas that constitutively express proinflammatory cytokines IL-1a, IL-6, GM-CSF and KC. Clin. Exp. Metastasis,16: 655–664, 1998.
12. Woodworth, C., McMullin, E., Iglesias, M., and Plowman, G. Interleukin 1a andtumor necrosis factora stimulate autocrine amphiregulin expression and proliferationof human papillomavirus-immortalized and carcinoma-derived cervical epithelialcells. Proc. Natl. Acad. Sci. USA,92: 2840–2844, 1995.
13. Young, M. R. I., Wright, M. A., Lozano, Y., Prechel, M. M., Benefield, J., Leonetti,J. P., Collins, S. L., and Petruzelli, G. J. Increased recurrence and metastasis inpatients whose primary head and neck squamous cell carcinomas secreted granulo-cyte-macrophage colony stimulating factor and contained CD341 natural suppressorcells. Int. J. Cancer,74: 69–74, 1997.
14. Yoneda, T., Aufdemorte, T., Nishimura, R., Nishikawa, N., Sakuda, M., Alsina, M.,Chavez, J., and Mundy, G. Occurrence of hypercalcemia and leukocytosis withcachexia in a human squamous cell carcinoma of the maxilla in athymic nude mice.A novel experimental model of three concomitant paraneoplastic syndromes. J. Clin.Oncol.,9: 468–477, 1991.
15. Baeuerle, P. A. Pro-inflammatory signaling: last pieces in the NF-kB puzzle? Curr.Biol., 8: R19–R22, 1998.
16. Bailly, S., Fay, M., Israel, N., and Gougerot-Pocidalo, M. A. The transcription factorAP-1 binds to the human interleukin 1a promoter. Eur. Cytokine Netw.,7: 125–128,1996.
17. Dendorfer, U., Oettgen P., and Libermann, T. A. Multiple regulatory elements in theinterleukin-6 gene mediate induction by prostaglandins, cyclic AMP, and lipo-polysaccharide. Mol. Cell. Biol.,14: 4443–4454, 1994.
18. Mukaida, N., Okamoto, S., Ishikawa, Y., and Matsushima, K. Molecular mechanismof interleukin-8gene expression. J. Leukocyte Biol.,56: 554–558, 1994.
19. Thomas, R. S., Tymms, M. J., McKinlay, L. H., Shannon, M. F., Seth, A., and Kola,I. ETS1, NFkB and AP1 synergistically transactivate the human GM-CSF promoter.Oncogene,14: 2845–2855, 1997.
20. Ondrey, F. G., Dong, G., Sunwoo, J., Chen, Z., Wolf, J. S., Crowl-Bancroft, C. V.,Mukalda, N., and Van Waes, C. Constitutive activation of transcription factorsNF-kB, AP-1 and NF-IL6 in human head and neck squamous cell carcinomas thatexpress proinflammatory and proangiogenic cytokines. Mol. Carcinog., in press,1999.
21. Wu, M., Lee, H., Bellas, R. E., Schauer, S. L., Arsura, M., Katz, D., FitzGerald, M. J.,Rothstein T. L., Sherr D. H., and Sonenshein, G. E. Inhibition of NF-kB/Rel inducesapoptosis of murine B cells. EMBO J.,15: 4682–4690, 1996.
22. Bargou, R. C., Emmerich, F., Krappmann, D., Bommert, K., Mapara, M. Y., Arnold,W., Royer, H. D., Grinstein, E., Greiner, A., Scheidereit, C., and Dorken, B.Constitutive nuclear factor-kB-RelA activation is required for proliferation and sur-vival of Hodgkin’s disease tumor cells. J. Clin. Invest.,100: 2961–2969, 1997.
23. Shattuck-Brandt, R. L., and Richmond, A. Enhanced degradation of IkBa contributesto endogenous activation of NF-kB in Hs294T melanoma cells. Cancer Res.,57:3032–3039, 1997.
24. Nakshatri, H., Bhat-Nakshatri, P., Martin, D. A., Goulet, R. J., Jr., and Sledge, G. W.,Jr. Constitutive activation of NF-kB during progression of breast cancer to hormoneindependent growth. Mol. Cell. Biol.,17: 3629–3639, 1997.
25. Sovak, M. A., Bellas, R. E., Kim. D. W., Zanieski, G. J., Rogers, A. E., Traish, A. M.,and Sonenshein, G. E. Aberrant nuclear factor-kB/Rel expression and the pathogen-esis of breast cancer. J. Clin. Invest.,100: 2952–2960, 1997.
26. Cadoret, A., Bertrand, F., Baron-Delage, S., Levy, P., Courtois, G., Gespach, C.,Capeau, J., and Cherqui, G. Down-regulation of NF-kB activity and NF-kB p65subunit expression byrasand polyoma middle T oncogenes in human colonic Caco-2cells. Oncogene,14: 1589–1600, 1997.
27. Nehyba, J., Hrdlickova R., and Bose, H. R., Jr. Differences inkB DNA-bindingproperties of v-Rel and c-Rel are the result of oncogenic mutations in three distinctfunctional regions of the Rel protein. Oncogene,14: 2881–2897, 1997.
28. Siebenlist, U. G., Franzoso, G., and Brown, K. Structure, regulation and function ofNF-kB. Annu. Rev. Cell Biol.,10: 405–455, 1994.
4 E. Loukinova, G. Dong, Z. Chen, I. Enamorado, G. Thomas, and C. Van Waes. CXCchemokine KC/GRO-a promotes angiogenesis, growth and metastasis of murine squa-mous cell carcinoma by a CXCR2-dependent host mechanism, manuscript in preparation.
3473
NF-kB IN SQUAMOUS CELL CARCINOMA
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

29. Brown, K., Gerstberger, S., Carlson, L., Franzoso, G., and Siebenlist, U. Control ofIkB-a proteolysis by site-specific, signal-induced phosphorylation. Science (Wash-ington DC),267: 1485–1488, 1995.
30. Traenckner, E. B., Pahl, H. L., Henkel, T., Schmidt, K. N., Wilk, S., and Baeuerle,P. A. Phosphorylation of human IkB-a on serines 32 and 36 controls IkB-a prote-olysis and NF-kB activation in response to diverse stimuli. EMBO J.,14: 2876–2883,1995.
31. Brockman, J. A., Scherer, D. C., McKinsey, T. A., Hall, S. M., Qi, X., Lee, W. Y.,and Ballard, D. W. Coupling of a signal response domain in IkBa to multiplepathways for NF-kB activation Mol. Cell. Biol.,15: 2809-2818, 1995.
32. Verma, I. M., Stevenson, J. K., Schwarz, E. M., Van Antwerp, D., and Miyamoto, S.Rel/NF-kB/IkB family: intimate tales of association and dissociation. Genes Dev.,9:2723–2735, 1995.
33. Beg, A. A., and Baltimore, D. An essential role for NF-kB in preventing TNF-a-induced cell death. Science (Washington DC),274: 782–784, 1996.
34. Wang, C-Y., Mayo, M. W., and Baldwin, A. S. TNF and cancer therapy-inducedapoptosis: potentiation by inhibition of NF-kB. Science (Washington DC),274:784–787, 1996.
35. Van Antwerp, D. J., Martin, S. J., Kafri, T., Green, D. R., and Verma, I. Suppressionof TNF-a-induced apoptosis by NF-kB Science (Washington DC),274: 787–789,1996.
36. Krause, C., Carey, T., Ott, R., Hurbis, C., McClatchey, K., and Regezi, J. Humansquamous cell carcinoma. Establishment and characterization of new permanent celllines. Arch. Otolaryngol.,107: 703–710, 1981.
37. Bradford, C. R., Kimmel, K. A., Van Dyke, D. L., Worsham, M. J., Tilley, B. J.,Burk, D., del Rosario, F., Lutz, S., Tooley, R., Hayashida, D. J. S., and Carey,T. E. 11p deletions and breakpoints in squamous cell carcinoma: association withaltered reactivity with the UM-E7 antibody. Genes Chromosomes Cancer,3:272–282, 1991.
38. Van Waes, C., Mi Surh, D., Chen, Z., Kirby, M., Rhim, J. S., Brager, R., Sessions,R. B., Poore, J., Wolf, G. T., and Carey, T. E. Increase in suprabasilar integrinadhesion molecule expression in human epidermal neoplasms accompanies increasedproliferation occurring with immortalization and tumor progression. Cancer Res.,55:5434–5444, 1995.
39. Brown, K., Franzoso, G., Baldi, L., Carlson, L., Mills, L., Lin, Y. C., Gerstberger, S.,and Siebenlist, U. The signal response of IkBa is regulated by transferable N- andC-terminal domains. Mol. Cell. Biol.,17: 3021–3027, 1997.
40. Sanes, J. R., Rubenstein, J. R., and Nicolas, J. F. Use of a recombinant retrovirus tostudy post-implantation cell lineage in mouse embryos. EMBO J.,5: 3133–3142,1986.
41. Harlow, E., and Lane, D. In: Antibodies. A Laboratory Manual, pp. 473–510 and635–657. Cold Spring Harbor, NY: Cold Spring Harbor, 1988.
42. Garner, M. M., and Revzin, A. A gel electrophoresis method for quantifying thebinding of proteins to specific DNA regions: application to the components of the
Escherichia colilactose operon regulatory system. Nucleic Acids Res.,9: 3047–3060,1981.
43. Dignam, J. D., Lebovitz, R. M., and Roeder, R. G. Accurate transcription initiation byRNA polymerase II in a soluble extract from isolated mammalian nuclei. NucleicAcids Res.,11: 1475–1489, 1983.
44. Lee, K. A. W., Bindereif, A., and Green, M. R. A small scale procedure forpreparation of nuclear extracts that support efficient transcription and pre-mRNAsplicing. Gene Anal. Tech.,5: 22–31, 1988.
45. Kanno, T., Brown, K., and Siebenlist, U. Evidence in support of a role for human Tcell leukemia virus type I Tax in activating NF-kB via stimulation of signalingpathways. J. Biol. Chem.,270: 11745–11748, 1995.
46. Kaltschmidt, C., Kaltschmidt, B., Henkel, T., Stockinger, H., and Baeuerle, P. A.Selective recognition of the activated form of transcription factor NF-kB by amonoclonal antibody. Biol. Chem. Hoppe-Seyler,376: 9–16, 1995.
47. Wang, C-Y., Mayo, M. W., Korneluk, R. G., Goeddel, D. V., and Baldwin, A. S.,NF-kB antiapoptosis: induction of TRAF1 and TRAF2 and c-IAP1 and c-IAP2 tosuppress caspase-8 activation. Science (Washington DC),281: 1680–1683, 1998.
48. Wu, M. X., Ao, Z., Prasad, K. V. S., Wu, R., and Schlossman, S. F. IEX-1L, anapoptosis inhibitor involved in NF-kB-mediated cell survival. Science (WashingtonDC), 281: 998–1001, 1998.
49. Zong, W. X., Edelstein, L. C., Chen, C., Bash, J., and Gelinas, C. The prosurvivalBCl-2 homolog Bfl/A1 is a direct transcriptional target of NF-kB that blocks TNFa-induced apoptosis. Genes Dev.,13: 382–387, 1999.
50. Yuspa, S. H. The pathogenesis of squamous cell cancer: lessons learned from studiesof skin carcinogenesis. Cancer Res.,54: 1178–1189, 1994.
51. Li, J-J., Westergaard, C., Ghosh, P., and Colburn, N. H. Inhibitors of both nuclearfactor-kB and activator protein-1 activation block the neoplastic transformationresponse. Cancer Res.,57: 3569–3576, 1997.
52. Li, J-J., Rhim, J. S., Schlegel, R., Vousden, K. H., and Coburn, N. Expression ofdominant negative Jun inhibits elevated AP-1 and NF-kB transactivation and sup-presses anchorage independent growth of HPV immortalized human keratinocytes.Oncogene,16: 2711–2721, 1998.
53. Grilli, M., Chiu, J. J-S., and Lenardo, M. L. NFkB and Rel participants in a multiformtranscriptional regulatory system. Int. Rev. Cytol.,143: 1–62, 1993.
54. Sun, L., and Carpenter, G. Epidermal growth factor activation of NF-kB is mediatedthrough IkBa degradation and intracellular free calcium. Oncogene,16: 2095–2102,1998.
55. Mayo, M. W., Wang, C. Y., Cogswell, P. C., Rogers-Graham, K. S., Lowe, S. W.,Der, C. J., and Baldwin, A. S., Jr. Requirement of NF-kB activation to suppressp53-independent apoptosis induced by oncogenicRas. Science (Washington DC),278: 1812–1815, 1997.
56. Reuther, J. V., Reuther, G. W., Cortez, D., Pendergast, A. M., and Baldwin, A. S. Arequirement for NF-kB activation in Bcr-Abl-mediated transformation. Genes Dev.,12: 968–981, 1998.
3474
NF-kB IN SQUAMOUS CELL CARCINOMA
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

1999;59:3468-3474. Cancer Res Dianne C. Duffey, Zhong Chen, Gang Dong, et al.
in VivoExpression, and Tumor Growth Carcinoma Inhibits Survival, Proinflammatory Cytokine
B in Human Head and Neck Squamous CellκNuclear Factor- ofαBκExpression of a Dominant-Negative Mutant Inhibitor-
Updated version
http://cancerres.aacrjournals.org/content/59/14/3468
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/59/14/3468.full.html#ref-list-1
This article cites 50 articles, 26 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/59/14/3468.full.html#related-urls
This article has been cited by 56 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on March 26, 2016. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from




















