
Immunophenotype of hematopoietic stem cells from placental/umbilical cord blood after culture
Upload
independentCategory
view
1download
0
Ex vivo expansion of haematopoietic stem/progenitorcells from human umbilical cord blood on acellularscaffolds prepared from MS-5 stromal cell lineAbhilasha Tiwari1,5, Melinda L. Tursky2,3, Dolly Mushahary1, Samiksha Wasnik1,5, Fiona M. Collier2,Kantipudi Suma4, Mark A. Kirkland2,5 and Gopal Pande1*1CSIR Centre for Cellular and Molecular Biology (CCMB), Hyderabad, India2Barwon Biomedical Research (BBR), Geelong Hospital, VIC, Australia3Lowy Cancer Research Centre, University of New South Wales, NSW, Australia4Sridevi Nursing Home, 12-11-253, Warasiguda, Secunderabad, India5Deakin University, Waurn Ponds, Geelong, VIC, Australia
Abstract
Lineage-specific expansion of haematopoietic stem/progenitor cells (HSPCs) from human umbilical cordblood (UCB) is desirable because of their several applications in translational medicine, e.g. treatment ofcancer, bonemarrow failure and immunodeficiencies. The currentmethods forHSPC expansion use eithercellular feeder layers and/or soluble growth factors and selected matrix components coated on differentsurfaces. The use of cell-free extracellular matrices from bone marrow cells for this purpose has notpreviously been reported. We have prepared insoluble, cell-free matrices from a murine bone marrowstromal cell line (MS-5) grown under four different conditions, i.e. in presence or absence of osteogenicmedium, each incubated under 5% and 20%O2 tensions. These acellular matrices were used as biologicalscaffolds for the lineage-specific expansion of magnetically sorted CD34+ cells and the results wereevaluated by flow cytometry and colony-forming assays. We could get up to 80-fold expansion of someHSPCs on one of the matrices and our results indicated that oxygen tension played a significant role indetermining the expansion capacity of the matrices. A comparative proteomic analysis of the matricesindicated differential expression of proteins, such as aldehyde dehydrogenase and gelsolin, which havepreviously been identified as playing a role in HSPC maintenance and expansion. Our approach may beof value in identifying factors relevant to tissue engineering-based ex vivoHSPC expansion, and itmay alsoprovide insights into the constitution of the niche in which these cells reside in the bone marrow.Copyright © 2012 John Wiley & Sons, Ltd.
Received 23 August 2011; Revised 18 November 2011; Accepted 16 January 2012
Supporting information may be found in the online version of this article.
Keywords haematopoietic niche; acellular biological scaffold; extracellular matrix; umbilical cord blood;stem cell expansion; proteomics
1. Introduction
Haematopoietic stem and progenitor cells (HSPCs) areinherently migratory and they come in contact withseveral niches through the course of their development(Jones and Wagers, 2008). The interaction of HSPCs with
their niches regulates their differentiation into more com-mitted lineages, such as the common lymphoid progeni-tors (CLPs) and the common myeloid progenitors (CMPs)(Orkin and Zon, 2008). Four major components of stemcell niches are known to control the growth and differen-tiation of HSPCs: (a) extracellular signals emanating fromsoluble or insoluble growth factors; (b) specific cell–cellinteractions between HSPCs and other supporting cellswithin the niche; (c) signals generated by the diffusibleand non-diffusible proteins, glycoproteins and proteogly-cans present in the extracellular matrix (ECM) of the
*Correspondence to: G. Pande, CSIR Centre for Cellular andMolecular Biology, Uppal Road, Hyderabad 500007, India.E-mail: [email protected]
Copyright © 2012 John Wiley & Sons, Ltd.
JOURNAL OF TISSUE ENGINEERING AND REGENERATIVE MEDICINE RESEARCH ARTICLEJ Tissue Eng Regen Med (2012)Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/term.1479

niche; and (d) physicochemical factors, such as nutrientsand oxygen levels (Wilson and Trumpp, 2006). Thus,components of a stem cell niche not only provide struc-tural, trophic and mechanical support to the niche butcan also deliver specific three-dimensional (3D) topo-graphical information and physiological cues to the cells,regarding the availability of mobilized and immobilizedgrowth factors (GFs) that are vital to stem cell functions(Morrison and Spradling, 2008). The physiological environ-ment of haematopoietic niches in the bone marrow is oftenhypoxic (Dellatore et al., 2008; Mohyeldin et al., 2010), withthe vascular niche having comparatively higher O2
concentration than endosteal niche. This O2 concentrationgradient, along with other factors, directly affects therecruitment of multi-potent progenitors (MPPs) from theendosteal niche to the vascular niche, resulting in prolifera-tion and differentiation of HSPCs. Studies have demon-strated that CD34+CD38– cells show enhanced expansionin low (1.5%) O2, and SCID-repopulating cell frequency inNOD/SCID mice was also higher when cells were expandedin low or hypoxic O2 in comparison to standard (20%) O2
(Sullivan et al., 2006). While the direct effect of O2 levelson the expansion of HSPCs has been investigated, littleattention has been paid to the effect of O2 levels on thestromal cells that support HSPCs in the bone marrow niches.
The concept of stem cell niches has been used in thetissue-engineering field to prepare bioengineered 3Dscaffolds for the purpose of ex vivo stem and progenitorcell expansion (Adams, 2008; Guilak et al., 2009). Al-though many inorganic biomaterials have been used forthis application (Banu et al., 2001; Dellatore et al., 2008;Franke et al., 2007), the use of cell-free ECMs as biologicalscaffolds for tissue engineering has been less common(Hoshiba et al., 2010). Examples of cells that have beenexpanded on cell-free matrices include mesenchymalstem cells (MSCs) (Chen et al., 2007), neuronal cells(Aizman et al., 2009), osteogenic cells (Evans et al.,2010) and hepatic cells (Shiraki et al., 2011) derived frommurine embryonic stem cells.
A mouse stromal cell line, MS-5 (Itoh et al., 1989),derived from murine long-term marrow culture, supportscolony-forming unit–spleen (CFU-S) maintenance (Dexteret al., 1977), acts synergistically with human GFs tostimulate the formation of macroscopic colonies fromCD34+CD38– primitive progenitors in short-term methyl-cellulose assays (Issaad et al., 1993) and promotes a netincrease of long-term culture-initiating cells (LTC-IC)through self-renewal divisions (Kanai et al., 2000). Inthe present study we have used cell-free matrices fromMS-5 cells that mimic the endosteal and/or vascularniche-like conditions in biological scaffolds to supportthe ex vivo expansion of HSPCs. MS-5 cells were grownunder varying conditions of osteogenic induction andO2 tension and cell-free matrices were prepared by astandardized protocol. We analysed the growth and differ-entiation of human umbilical cord blood (UCB)-derivedCD34+ cells on these matrices for a period of 8 days.Our results showed optimal expansion of HSPCs onthe matrices of stromal cells that were grown at 20%
O2 in the presence of osteogenic medium (OGM). Com-parison of the protein profiles of the cell-free matricesof MS-5 cells, grown under different conditions, bytwo-dimensional (2D) gel electrophoresis and MALDI–TOF/MS analysis identified some candidate proteinsthat may be involved in the regulation of HSPC prolifer-ation and differentiation. Our results provide a newmethodology for the ex vivo expansion of humanCD34+ cells on heterologous cell-free biological scaf-folds. They also provide a model for investigating therole of O2 tension and osteogenic signals in regulatingHSPC differentiation through indirect effects on theniche stromal cells and ECM. Thus, our findings arehelpful in understanding the biochemical organizationof haematopoietic niches, and they also suggest thepossible design of bioactive and biomimetic scaffoldsthat could be used for haematopoeitic stem cell-basedtissue engineering.
2. Materials and methods
The sources of various reagents and cells, used in allexperiments, are provided in Supplementary materials(see Supporting information).
The overall experimental approach used in this study isshown schematically in Figure 1, where panels A, B andC, respectively, depict the steps for CD34+ cell isolation,biological scaffold (cell-free matrix) preparation and ex vivoHSPC expansion. Details of these steps are given below.
2.1. CD34+ isolation from UCB
Freshly collected UCB (40–55ml) was diluted four timeswith Dulbecco’s phosphate-buffered saline (PBS), pH7.2, containing 2mM EDTA, and mononuclear cells(MNCs) were collected from the buffy coat followingFicoll–Paque™ Plus density gradient centrifugation.CD34+ cells were isolated from MNCs by magnetic-assisted cell sorting (MACS) on an AutoMACS cell sorter,using the ‘double positive selection program’ provided bythe manufacturers, and the MACS CD34 MicroBead Kit,according to the manufacturer’s instructions. The percen-tages of CD34+ cells before and after MACS isolation, asassayed by flow cytometry, were 0.65� 0.24% and75.63�1.1%, respectively.
2.2. Stromal cell culture
MS-5 cells were maintained in conditions standardized inour laboratory for the expansion of UCB cells on MS-5cultures. The cells were grown in complete medium(CM) containing a-MEM, supplemented with 10% FCS,1� glutamax and 1� penicillin/streptomycin (P/S) at5% O2, 5% CO2 and 37�C. For matrix preparation, MS-5cells were seeded at 1�104 cells/cm2 in CM and grownunder low (5%) O2 and normal (20%) O2 conditions for
A. Tiwari et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

10days, with a change ofmedium every 72h. For osteogenicinduction CM was replaced with OGM, i.e. CM containing10mM b-glycerol phosphate, 100nM dexamethasone and100mM ascorbate 2-phosphate for 48h.
2.3. Preparation of decellularized biologicalscaffolds
Cell-free matrices which acted as biological scaffolds wereprepared from MS-5 cells using the following protocol.Confluent cells were rinsed in PBS and incubated withdeionized water (MQ) for 2–3min to loosen the cellattachment with the plastic surface. After removing MQ,0.02M NH4OH was applied to the cells for 1–2min atroom temperature (RT) for cell lysis, which was observedcarefully under an inverted microscope, and the NH4OHwas removed by gently inverting the culture plates. Thefragile matrices were allowed to dry overnight in thelaminar flow cabinet, washed three times with PBS andstored in PBS with P/S at 4�C. Thus, at the end of the pro-cedure, the following four types of decellularized matrices(MX1-4) were obtained from MS-5 cells (Figure 1B):
MX1 ¼ cells grown in 5% O2 þ OGMMX2 ¼ cells grown in 20% O2 þ OGMMX3 ¼ cells grown in 5% O2 � OGMMX4 ¼ cells grown in 20% O2 � OGM
2.4. Matrix characterization
In order to compare the physical and biochemical propertiesof the matrices, MS-5 cells and MX1-4 matrices were ana-lysed by phase-contrast and scanning electron microscopy,cytochemistry, immunostaining and proteomic analysis.Details of the methods used for this purpose are describedin Supplementary methods (see Supporting information).
2.5. HSPC expansion
Biological scaffolds (MX1-4) were prepared in 24-wellplates as described above. Negative control surface C1was prepared by incubating the wells (without cells) withCM only for 10 days followed by only OGM for next2 days. The negative control surface, C2, was preparedby incubating them with CM only for 12 days. The positive
Figure 1. Experimental flow chart. (A) Isolation of UCB-derived CD34+ HSPCs. MNCs were isolated by Ficoll gradient centrifugation andwere then labelled by CD34 microbeads and separated by AutoMACS. CD34+ HSPCs were characterized by flow and CFU assay. (B)Preparation of decellularized matrices. MS-5 cells were grown under two different oxygen conditions (5% and 20%) for 10days and wereincubated with or without osteogenic medium (OGM) for the last 2days. They were then treated with NH4OH for decellularization.(C) Co-culture of CD34+ HSPCs on acellular matrices and controls for 8days at 5% O2. C1, wells incubated with maintenancemedium for 10days and osteogenicmedium for the next 2days; C2, wells incubatedwithmaintenancemedium for all 12days, i.e. withoutosteogenic medium; C3, RetroNectin (RN)-coated wells, which were taken as positive controls
Haematopoietic progenitor cell expansion on acellular matrices
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

control surface, C3, was prepared by incubating the wellswith 10 mg/ml RetroNectin for 30min at RT; RetroNectin isused as a standard substrate for HSPC expansion, as reportedearlier (Uchida et al., 2011). Following these incubations, allcontrol surfaces were treatedwith the same decellularizationprotocol as has been described for the cell matrices.
Magnetically sorted UCB CD34+ cells, as a primary sourceof HSPCs, were resuspended in Stemline II serum-free me-dium supplemented with 1� P/S and GFs as indicated:thrombopoietin (TPO) 25ng/ml, stem cell factor (SCF)25ng/ml, interleukin-6 (IL-6) 50ng/ml; and Flt3 ligand(Flt3L) 40ng/ml; then seeded at 1�104 viable cells/ml inthe coated wells. For each category, MX1-4 and C1-3, tripli-cate cultures were prepared. All cultures were placed in thehypoxic incubator (5% O2, 5% CO2, 37�C, humidified) for8days. Cells were harvested by gentle flushing, followed bytwo washes with PBS. Viable cell counting was done with1% trypan blue and the cells were further analysed by flowcytometry and colony-forming assay (Figure 1C).
2.6. Flow cytometry
Flow-cytometric analysis for surface markers was done withthe following three antibody combinations: CD45 FITC+CD34 PE, CD34 FITC+CD38 PE and CD34 FITC+CD133PE; 7-amino-actinomycin D (7-AAD), at a concentration of0.25mg/1�106 cells, was added to each combination tomeasure cell viability. PE- and FITC-labelled anti-mouseisotype IgG stainings were used as compensation controls.Antibody-stained cells were analysed in a FACSCalibur flowcytometer, using CellQuest software. For determining themean and standard error values, data from three experi-ments, with triplicate samples in every experiment, wereused. Fold increase in total cell number and lineage-specificcells was calculated as follows:
(a) For total cells:
Count of total viable cells after expansionCount of total viable cells before expansion
(b) For lineage-specific cells:
Lineage � specific cell percentage� count of total viable cellsð Þ after expansionLineage � specific cell percentage� count of total viable cellsð Þ before expansion
(c) For percentage of HSPC subpopulations:
Fold expansion of HSPC subpopulation� 100Fold expansion of total viable cells
2.7. Colony-forming assays
Colony-forming assays were performed for all cell sam-ples, as triplicate cultures in Methocult media (Stem Cell
Technologies, cat no. H4434), to assess the numbers ofcolony-forming units (CFUs) for granulocyte, erythrocyte,macrophage and megakaryocyte common precursors(CFU-GEMM), granulocyte/macrophage common precur-sors (CFU-GM) and burst-forming units (BFU-E) for pureerythroid precursors, according to the manufacturer’sinstructions. Fold expansion of CFUs in each culture wascalculated as explained above for marker-based cellexpansion. The details of the experiment are describedin Supplementary methods (see Supporting information).
2.8. Protein analysis by MALDI–TOF/MS
The decellularized matrices of MX1-4 conditions werewashed with water and total matrix proteins were iso-lated in extraction buffer. 2-Dimensional electrophoresis(2-DE) analysis of proteins was performed as describedin Supplementary methods (see Supporting information)(O’Farrell, 1975). Stained gels were analysed by PDQuestSoftware v 7.2 and the differentially expressed spots inMX1, MX2 and MX4 gels were identified by keeping theMX3 gel spots as our standard culture condition. Anydifference which was more than two-fold (p≤ 0.05) wasconsidered as significant. MALDI–TOF/MS analysis of 52well-characterized spots from triplicate gels was doneafter in-gel digestion with trypsin, and the protein identi-ties of those spots were established using the IPIMousedatabase with significant MASCOT scores (> 62).
2.9. Statistical analysis
Statistical relevance of the mean change in cell percentageand colony numbers during each experiment was deter-mined via one-way ANOVA, using Minitab software.Pairwise comparison of interexperiment variation wasdone with a general linear model ANOVA to see theindividual and combined effect of the conditions on HSPCexpansion. Standard errors in the data points were deter-mined at the 95% confidence interval and p≤ 0.05 withineach experiment were considered as significant.
3. Results
3.1. Physicochemical characterization of thebiological scaffolds
Phase-contrast microscopy of the matrices before andafter decellularization showed the absence of intact cells(Figure 2Aa, b). This was confirmed by SEM analysis(Figure 2Ac–f), where matrix-bound cells were seen beforeNH4OH treatment (Figure 2Ac, e) but only an intact 3Dmatrix could be seen after cell removal (Figure 2Ad, f ).
The absence of cells in the matrices was also confirmedby DAPI staining for nuclei (Figure 2Ba, b). The presenceof collagen and GAGs in the cells and matrix was assessedby cytochemical staining with light green and alcian blue,respectively. Deep staining for both collagen and GAGs
A. Tiwari et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

could be seen in both conditions (Figure 2Bc–f), indicatingthat the decellularization process did not extract thesemolecules from the matrix. Specific immunostaining for
fibronectin, laminin and collagen type 1 was demonstratedunder both cellular and acellular conditions (see Support-ing information, Figure S1A, B and C, respectively). The
Figure 2. Characterization of cells andmatrices before and after decellularization. (A) Morphological properties of the cells and decellular-ized matrices generated under MX2 conditions (see Figure 1B). Phase-contrast microscopy of (a) the cells and (b) the matrices show theremoval of cells after NH4OH treatment (scale bar=100mm). SEM analysis of (c) the cells and (d) the matrix at lower magnification (scalebar=50mm) and (e, f) at higher magnification for the same (scale bar=10mm) shows the cells attached to the ECMproduced by them and,subsequently, the rough, 3D structure of thematrix after decellularization. (B) Cytochemical properties of theMX2 cells and their decellular-ized matrices. (a, c, e) Matrices with cells and (b, d, f) after decellularization. (a, b) Fluorescence microscopy for DAPI illustrates that thenucleus is removed byNH4OH treatment. (c, d) Light green staining for collagen shows that NH4OH removes cellswithout adversely affectingthe collagenous ECM on the culture surface. (e, f) Alcian blue staining for proteoglycans also proves the integrity of ECM-bound GAGs upondecellularization. Similar analysis was also done for cells grown under all other conditions (scale bar=100mm)
Haematopoietic progenitor cell expansion on acellular matrices
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term
absence of co-localized actin, after removal of cells (seeSupporting information; cf. Figure S1a and d, or c and f ),indicated that most of the cellular actin was removedduring the decellularization process. Similar results wereobtained for all matrices.
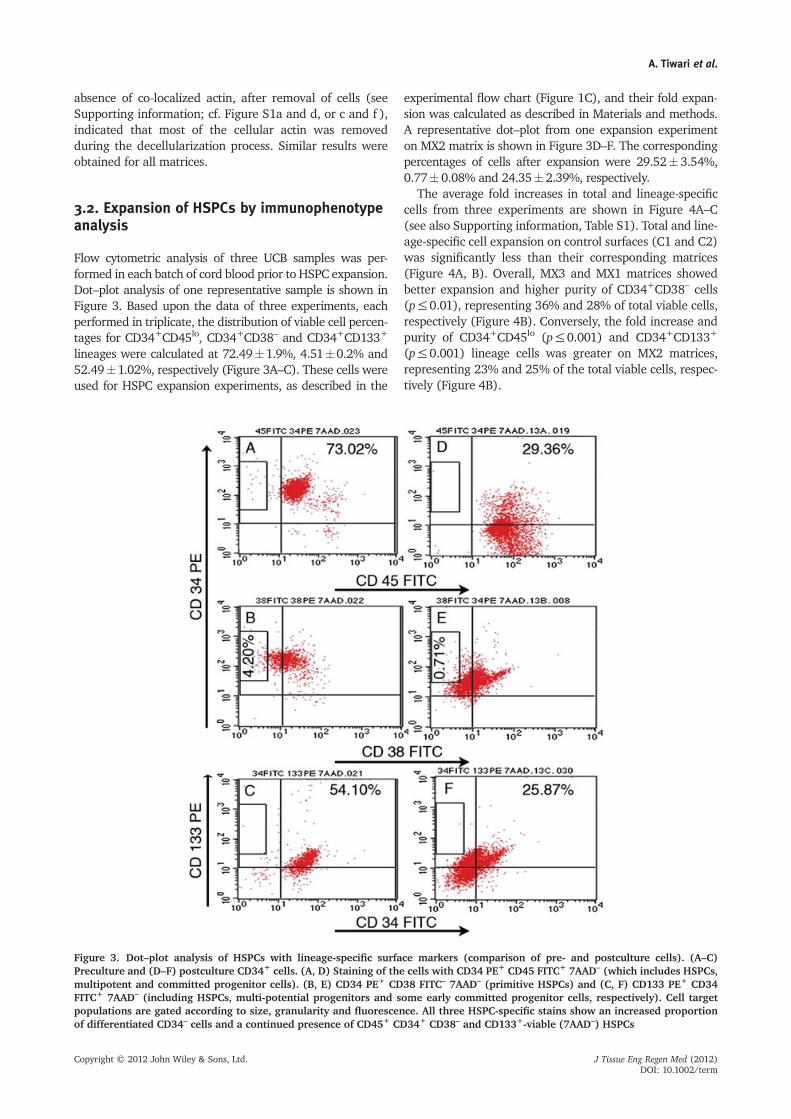
3.2. Expansion of HSPCs by immunophenotypeanalysis
Flow cytometric analysis of three UCB samples was per-formed in each batch of cord blood prior to HSPC expansion.Dot–plot analysis of one representative sample is shown inFigure 3. Based upon the data of three experiments, eachperformed in triplicate, the distribution of viable cell percen-tages for CD34+CD45lo, CD34+CD38– and CD34+CD133+
lineages were calculated at 72.49�1.9%, 4.51�0.2% and52.49�1.02%, respectively (Figure 3A–C). These cells wereused for HSPC expansion experiments, as described in the
experimental flow chart (Figure 1C), and their fold expan-sion was calculated as described in Materials and methods.A representative dot–plot from one expansion experimenton MX2 matrix is shown in Figure 3D–F. The correspondingpercentages of cells after expansion were 29.52� 3.54%,0.77� 0.08% and 24.35� 2.39%, respectively.
The average fold increases in total and lineage-specificcells from three experiments are shown in Figure 4A–C(see also Supporting information, Table S1). Total and line-age-specific cell expansion on control surfaces (C1 and C2)was significantly less than their corresponding matrices(Figure 4A, B). Overall, MX3 and MX1 matrices showedbetter expansion and higher purity of CD34+CD38– cells(p≤0.01), representing 36% and 28% of total viable cells,respectively (Figure 4B). Conversely, the fold increase andpurity of CD34+CD45lo (p≤0.001) and CD34+CD133+
(p≤0.001) lineage cells was greater on MX2 matrices,representing 23% and 25% of the total viable cells, respec-tively (Figure 4B).
Figure 3. Dot–plot analysis of HSPCs with lineage-specific surface markers (comparison of pre- and postculture cells). (A–C)Preculture and (D–F) postculture CD34+ cells. (A, D) Staining of the cells with CD34 PE+ CD45 FITC+ 7AAD– (which includes HSPCs,multipotent and committed progenitor cells). (B, E) CD34 PE+ CD38 FITC– 7AAD– (primitive HSPCs) and (C, F) CD133 PE+ CD34FITC+ 7AAD– (including HSPCs, multi-potential progenitors and some early committed progenitor cells, respectively). Cell targetpopulations are gated according to size, granularity and fluorescence. All three HSPC-specific stains show an increased proportionof differentiated CD34– cells and a continued presence of CD45+ CD34+ CD38– and CD133+-viable (7AAD–) HSPCs
A. Tiwari et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

The specific roles of O2 and OGM on the ability of thescaffold to support total and lineage-specific cell expan-sion are represented in Figure 4C. Matrices grown athigher O2 concentration (MX2 and MX4) expandedCD34+CD45lo and CD34+CD133+ cells more than matri-ces prepared at low O2 (MX1 and MX3), irrespective ofthe presence of OGM (Figure 4Cb, d). Matrices laid downby cells grown at low O2 appeared to support greaterexpansion of CD34+CD38– cells (Figure 4Cc), but thesedifferences were not statistically significant. The individ-ual role of OGM could not be determined in this analy-sis, as OGM alone did not show statistically significant
change in data. However, in combination with higherO2 concentration (MX2 matrix), cells grown in OGMsupported greater expansion of CD34+CD45lo (p≤ 0.05)and CD34+CD133+ cells (p≤0.05).
3.3. Fold expansion of HSPCs by CFU assay
Functional properties of the expanded cells were assessedby colony-forming assays in semi-solid media. This directlymeasured the in vitro potential of HSPC differentiation intovarious haematopoietic lineages. The fold expansion of
Figure 4. Fold expansion and purity of HSPCs, based on surfacemarkers. (A) Fold expansion of the HSPC subpopulation compared to thefold expansion for total cells. Statistical analysiswas doneusing one-wayANOVA (*p≤0.05; **p≤0.01; ***p≤0.005). (B) Percentage of thethree HSPCs subpopulations as compared to total cell expansion. (C) General linear model ANOVAwith 95% confidence and p≤0.05 wasdone to compare the O2 group, the OGM group and the combined effect of O2*OGM groups on HSPC expansion (*p≤0.05; **p≤0.01;***p≤0.005). (a) Total viable cells; there was no significant effect of either of two conditions on the expansion of total cell population.(b) Viable CD34+CD45low cells. There was a combined effect of both O2*OGM groups onMX2 condition (#p≤0.05) for expansion of thesecells. (c) Viable CD34+ CD38– cells. (d) Viable CD34+CD133+ cells, which also shows a combined effect of both O2*OGM groups on theMX2 condition (#p≤0.05). The individual role of O2 is quite prominent for CD34+CD45low and CD34+CD133+ cells, as the fold expansiononmatrices at 20%O2 (MX2 andMX4)was statistically better than that on 5%O2 (MX1 andMX3). The individual role ofOGMcould not bedetermined in this analysis, as OGM alone did not show a statistically significant change in data. Results are based on an average of threeindividual experiments with triplicates of each condition within an experiment
Haematopoietic progenitor cell expansion on acellular matrices
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

HSPCs based on CFU assays was higher than that seenin the flow-cytometry assays. The counts of BFU-E,CFU-GM and CFU-GEMM progenitors, before and afterthree expansion experiments and each performed in trip-licate, were used to calculate the fold expansion of therespective progenitors. Detailed analysis of these data isshown in Table S2 (see Supporting information).
The expansion of all colony-forming cell units wassignificantly higher using MX2 than in other experimentalmatrices and controls (Figure 5A). In particular, weobserved that on MX2 matrix the fold increase of
erythroid progenitors was equivalent to the fold increasein total viable cell number. The fold increase ofCFU-GMs and CFU-GEMMs was also significantly greateron MX2 than on other matrices. For all other matrices,the expansion of CFU was less than the level of totalviable cell expansion, but largely mirrored its pattern ofexpansion.
The individual and combined roles of O2 and OGMon colony-forming unit expansion are represented inFigure 5B, and multivariate analysis indicated a significantsynergistic effect of high O2 and OGM to enhance
Figure 5. Fold expansion of different cell populations by colony-forming assay. (A) Fold expansion of the CFUs compared to the foldexpansion for total cells. Statistical analysis was done using one-way ANOVA (*p ≤0.05; **p ≤0.01; ***p ≤0.005). (B) General linearmodel ANOVA with 95% confidence and p<0.05 was done to compare the O2 group, the OGM group and the combined effect ofO2*OGM groups (*p ≤0.05; **p ≤0.01; ***p ≤0.005). The presence of progenitor colonies were analysed by colony-forming assaysin semi-solid media after 14days and the colonies were manually counted for both pre- and postexpanded cells to calculate foldexpansion of those colonies. (a) BFU-E; (b) CFU-GM; and (c) CFU-GEMM; (d) total colony numbers, represented by the mean of allthe colonies. The individual role of O2 is seen for all the types of colony, as the fold expansion on matrices at 20% O2 (MX2 andMX4) was statistically better than that on 5% O2 (MX1 and MX3). We could also see the individual role of OGM on BFU-E, CFU-GEMMand total colonies, as the matrices with OGM (MX1 and MX2) showed significantly higher fold expansion compared to matriceswithout OGM (MX3 and MX4). All four types of colony had the combined effect of both O2*OGM groups on MX2 condition(#p ≤0.05 and ##p ≤0.01). Results are based on an average of three individual experiments, with triplicates of each condition withinan experiment. BFU-E, burst forming unit–erythroid; CFU-GM, colony-forming unit–granulocyte, macrophage; CFU-GEMM, colony-forming unit–granulocyte, erythroid, macrophage, megakaryocyte
A. Tiwari et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

expansion for all colony types – BFU-Es (p≤ 0.01), CFU-GMs (p≤ 0.05) and CFU-GEMMs (p≤ 0.05).
3.4. Proteomic analysis of the scaffold proteins
Proteins from all the scaffolds (MX1-4), were analysedusing standard 2D gel electrophoresis followed byMALDI–TOF/MS protocols, as described in Materials andmethods. Protein identities of approximately 300 spots,taken from all the four gels, with triplicates of each gel,were established using PDQuest software, and compari-son of protein expression in MX1, MX2 and MX4 scaffoldswas done using MX3 proteins as the reference scaffold. Atotal of 64 differential spots could be identified in MX1,MX2 and MX4, whose levels differed significantly (p< 0.05)from the MX3 gels. Positions of these spots have beenindicated in the gel pictures of the respective matrices(Figure 6). The MALDI–TOF/MS identities of 52 spots fromall matrices, along with their functional category, are shownin Table S3 (see Supporting information).
3.5. Identities of MX2-specific proteins
We studied the protein spots of MX2 scaffolds in moredetail, because MX2 had shown the most significantexpansion of clonogenic progenitors. These 29 spots repre-sented 23 unique protein identities whose comparativeexpression details are shown in Table 1. Of the 10 proteinsthat were uniquely regulated in the MX2 scaffold (see S.No. 1–10 in Table 1), only one protein (a-centractin) wasupregulated, whereas the rest were downregulated incomparison to other scaffolds. Among other proteins, fourwere commonly affected in MX2 and MX4 (S. Nos 11–14)and seven were commonly affected in MX2 and MX1 (S.Nos 15–21). It is noticeable that among 23 proteinsidentified in MX2, 18 were downregulated in comparisonto other matrices, indicating that their presence could beinhibitory for HSPC expansion.
The identities of the 23 MX2-related proteins (Table 1)indicate that several proteins linked with stem celldifferentiation functions are present in the stromalmatrices. For example, Aldh and Gelsolins are involvedin the regulation of stem cell signalling and they are alsoupregulated in the matrices of stromal cells.
4. Discussion
Ex vivo expansion of HSPCs on coated or uncoated inor-ganic substrates and ECM components has been reportedand reviewed in the literature. These scaffolds, in the pres-ence or absence of the specific growth factors and in thetime range 10days–9weeks expanded UCB-derivedCD34+ to different levels. They documented a three-fold in-crease on the Cytomatrix system (Ehring et al., 2003), 3.3–4.8-fold on biomimetic scaffolds (Tan et al., 2010), 6.8-foldon non-woven fibrous matrices (Li et al., 2001) and ~100
fold on FN-conjugated scaffold (Feng et al., 2006) forCD34+ cells. These reports have expanded our knowledgeabout the constitution of the haematopoietic niches andthey have also provided efficient and practical methodsfor obtaining large numbers of HSPCs for research and clin-ical use (Lund et al., 2009; Miyoshi et al., 2011). However,mimicking the interplay between cells and their cytokine-rich, tissue-specific environments remains relatively rare inthe field of tissue engineering (Discher et al., 2009). Ourresults have, for the first time, shown that it is possible to ex-pand human HSPCs on decellularized biological scaffoldsobtained from a bone marrow-derived stromal cell line.MS-5 cells with or without exogenous cytokines have been
Figure 6. Four representative 2-DE images of the decellular-ized matrices grown under different culture conditions(MX1-4). Proteins were separated in the first dimension onIPG strip (11 cm, pI range 4–7) and in the second dimensionby SDS–PAGE (4–15% gradient) on BioRad Criterion precastgels. Rectangles indicate upregulated/present and ovalsindicate downregulated/absent protein spots, as comparedto the standard condition (MX3)
Haematopoietic progenitor cell expansion on acellular matrices
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

shown earlier to support human HSPC expansion as feederlayers (Kanai et al., 2000; Keller et al., 2002), but expansionof human HSPCs has not been achieved on decellularizedmatrices of either mouse or human stromal cells. Wehave shown that stromal cells can significantly change theconstitution of their matrix and their ability to supportHSPC expansion in response to O2 levels and osteogenicinduction, and have undertaken preliminary analysis ofthe stroma to identify matrix components that maycontribute to this functional difference.
Stromal cell matrix proteins have been found to regulateHSPC expansion under laboratory conditions (Verfaillie etal., 1999). In our study, we aimed to mimic the functions ofthe endosteal and vascular niches by preparing cell-freematrices from MS-5 cells that were grown under differentO2 concentrations andwith/without osteogenic signals. Suchan approach is of significant advantage over the other tradi-tional method of using single ECM protein, as it capturesthe true complexity of ECM mix surfaces (Galan et al.,2007). As our matrices are intended for ex vivo HSPC expan-sion, our isolation procedure called for a balance betweencomplete removal of cells, and the preservation of ECM pro-tein activity (Grunert et al., 2007), and therefore differedsignificantly from other methods (Brown et al., 2011). Thematrices were characterized using different biochemicaland immunostaining methods and the results showed thatthe decellularization procedure removed almost all the cellsbut ECM components, such as collagen and ECM-boundGAGs, which are both important for haematopoiesis(Rodgers et al., 2008), were retained in the residualmatrices.
The optimized growth factor concentrations for humancord blood HSPC expansion ex vivo have previously been
established (Broxmeyer et al., 2009; Yao and Hwang,2007). However, in order to clearly identify the impact ofthe ECM on HSPC growth, the concentration of these GFswas reduced to half of the maximally stimulating levels.Although HSPC expansion experiments (in Stemline IImedium) on all matrices were performed at 5% O2,increase of total viable cell number was greater onmatrices prepared from MS-5 cells grown in 20% O2
(MX2 and MX4) compared to matrices prepared fromMS-5 cells grown in 5% O2 (MX1 and MX3). In addition,expansion of haematopoietic progenitors with pheno-type CD34+CD45lo and CD34+CD133+ was greater onmatrices obtained from MS-5 cells grown in 20% O2 inthe presence of osteogenic factors (MX2). This was alsoconfirmed by clonogenic assays, with MX2 matrixshowing the greatest expansion of all colony types. Incontrast, the expansion of cells with a more ‘primitive’phenotype (CD34+CD38–) was greatest using matricesgenerated at low O2 without osteogenic factors (MX3).
These data can be interpreted in several ways for therelative assessment of the individual and synergistic rolesof O2 and osteogenic induction in the haematopoieticniches for controlling HSPC expansion. The greaterexpansion of progenitors on matrices grown in higher O2
conditions is consistent with the concept that the vascularniche promotes expansion of committed progenitors, whilethe greater expansion of cells with a more primitive pheno-type on matrices grown under low O2 conditions is consis-tent with the concept that the hypoxic endosteal niche pro-motes self-renewal (Jones and Wagers, 2008; Mohyeldinet al., 2010). However, the synergistic effect of high O2
and OGM in promoting expansion of progenitors is less easy
Table 1. Differentially expressed proteins in the MX2 scaffold, relative to MX1 and MX4
Serial No. Protein name MX1 MX2 MX3 MX4 Functions
Uniquely affected in MX21 a-Centractin + " + + Involved in vesicle motility2 Peroxiredoxin-6 + - + + Anti-oxidant3 Isoform 3 of heterogeneous nuclear
ribonucleoprotein K (hnRNPs)+ - + + Downregulation helps in osteogenic differentiation
4 Annexin A5 + # + + Mediate vesicle/matrix interactions for bonemineralization. A5 also used as apoptotic marker5 Annexin A7 + # + +
6 78 kDa Glucose-regulated protein (HSPA5) + # + + Hypoxia-induced stress protein7 Actin, cytoplasmic 1 + - + + Cell motility8 Dynactin subunit 2 + # + + Motor activity, protein binding9 Actin, cytoplasmic 2 + # + + Cell motility10 T-complex protein 1 subunit e + # + + Chaperone, ATP bindingCommonly affected in MX2-MX1 and MX2-MX411 Aldehyde dehydrogenase (Aldh) + " + " Highly expressed in primitive HSCs12 ATP synthase subunit b, mitochondrial + " + " Cellular energy provision13 Nucleophosmin 1 + # + # Enhances HSC proliferation14 Isoform 1 of tropomyosin a1 chain + # + # IFP; involved in stroma-dependent hematopoiesis15 Annexin A1 " " + + MSC differentiation into osteoblasts16 L-Lactate dehydrogenase B chain (LDH) - + - + Related to hypoxia-induced factor. Levels reduced in
hypoxic condition17 Protein disulphide-isomerase A3 # # + + Calcium-binding antioxidant protein18 Calreticulin # # + + Regulates production of ECM19 Vimentin # # + + MSC marker20 Ferritin light chain 1 # # + + Ferric ion binding, oxidoreductase21 Guanine deaminase # # + + Guanine deaminase activity22 Isoform 2 of gelsolin " " + " Homing functions of MSCs23 Endoplasmin # # + # Stress protein
All scaffolds were compared by keeping MX3 protein levels as baseline.
A. Tiwari et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

to reconcile, considering the fact that such cells in vivo growin more hypoxic conditions and without osteogenicstimulus. The limitations of scaffolds prepared from MS-5matrices to support human HSPCs must be taken intoconsideration. MS-5 is an immortalized cell line habituatedto grow under non-physiological conditions, and the HSPCexpansion itself occurs in the presence of supraphysiologi-cal growth factor concentrations. Nevertheless, thesescaffolds allow the direct comparison of the effects ofECM generated from a single homogeneous cell type underdifferent conditions, which could enable us identify keyfactors involved in the control of HSPC growth and differen-tiation. We therefore undertook proteomics analysis of thematrices generated under different conditions.
Different levels of HSPC expansion on each matrixsuggests the role of differentially expressed matrixproteins prepared by culturing the cells in different condi-tions. Since MS-5 cells were originally maintained at 5%O2, as it represents the niche-like conditions and expan-sion of CD34+ cells was greater at lower oxygen concen-tration (Tursky et al., in press), MX3 (low O2, no OGM)was taken as the ‘baseline’ to identify the differentiallyexpressed proteins in ECM grown under other conditions.
The proteins differentially expressed under different O2
conditions (MX2/MX4 vs MX1/MX3) may indicate theirpossible role in O2 metabolism in stromal cells. Proteinssuch as b-chain of b-subunit of mitochondrial ATP synthase,L-lactate dehydrogenase-B (LDH-B) and aldehyde dehydro-genase (ALDH), that protect HSPCs from reactive O2
species (ROS) caused by higher O2 levels as well as otherprocesses relevant to stem cell regulation (Levi et al.,2009; Rossignol et al., 2003; Satyavani et al., 2009), areupregulated under high O2 conditions (MX2 and MX4).Gelsolin, which has a potential impact on the homingfunctions of stem cells (Forsberg et al., 2005), is also upre-gulated in MX2. Some hypoxia-induced stress proteins,such as HSPA5, were downregulated in MX2, as expected.Upregulation of annexin A1 in stroma grown in the pres-ence of OGM (MX1 and MX2) is consistent with its role inregulation of osteoblastic differentiation (Park et al., 2007;Zhang et al., 2007). Calreticulin, an ECM protein whichinteracts with thrombospondin and regulates the ECMproduction, and vimentin, which is a marker for MSCsand whose expression decreases with the differentiationof the MSCs into the osteogenic lineage, are downregulatedin both MX1 and MX2 (Kim et al., 2005; Satyavani et al.,2009). Ye et al. (2006) have shown that the expression ofPrx6, an oxidoreductase, and hnRNPs is downregulated indifferentiating MSCs, consistent with their absence in MX2matrices. It is also to be noted that 17 of 23 proteins weredownregulated in the MX2 matrix, indicating that osteo-genic induction and higher O2 levels reduce the expressionof proteins that could be inhibitory for HSPC expansion.
Our preliminary proteomic analysis of the matrices hasthus identified a number of proteins whose differentialexpression could be linked to the observed differences inthe ability of these matrices to support HSPC expansion.These results justify a more detailed and in-depth study ofthe protein constitution of the matrices and the gene
expression profile of MS-5 cells, to identify additionalfactors that may be involved. Our in vitro results on theexpansion of UCB-derived HSPCs on acellular MS-5matrices are of potential clinical value, since they may beindicative of the increased short-term repopulatingcapacity. However, we are unable to predict the capabilityof the expanded cells to support long-term repopulation ofall haematopoietic lineages. Further, since we haveperformed experiments only with the MS-5 cell matrices,we cannot be sure that experiments done on othermatrices would give similar results. Hence, future aspectsof this work would include investigating the role of thecandidate proteins as suggested by proteomics analysis, inHSPC expansion, conducting SCID mice repopulating cell(SRC) assays and testing the capability of matrices fromother cell lines for HSPC expansion.
In summary, we have prepared an acellular biologicalscaffold by modulating O2 tension and osteogenicinduction of mouse stromal cells that approaches mimick-ing the complexity of a stem cell niche and is conducive tothe efficient expansion of HSPCs, mainly of myeloidorigin, as shown by CFU assays. A limited analysis of theproteomic constitution of the matrices indicates the roleof proteins that could be involved in maintaining and/orexpanding HSPCs in vivo. Thus, this approach may be ofvalue for the identification of factors relevant to tissueengineering-based haematopoietic cell expansion, andmay provide insights into the constitution of the niche inwhich these cells reside in the bone marrow.
Acknowledgements
This study was supported by the Department of Science and Tech-nology, New Delhi, India (Grant Nos GAP0220 and GAP0258) andpartially by the Indo-Australian Biotechnology Fund (Grant No.GAP0311, to G.P.). A.T. and S.W. were supported by the DeakinUniversity Overseas Fellowship Programme at CCMB. We wouldlike to thank the staff of the CCMB Microscopy, FACS and Proteo-mics facilities for their help.
Supporting information on theinternet
The following supporting information may be foundin the online version of this article:
Supplementary materialsSupplementary methods
Figure S1. Immunostaining for actin and ECM pro-teins on MX2 matrices
Table S1. Fold expansion of HSPCs, based on surfacemarkers
Table S2. Fold expansion of different cell populationsby colony-forming assay
Table S3. Summary of proteins up and down-regulatedin MX1, MX2 and MX4 conditions as compared to thestandard (MX3) matrix
Haematopoietic progenitor cell expansion on acellular matrices
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

Author contributions
A.T., all experimental work, data analysis, statistical interpreta-tion and manuscript writing; M.L.T., flow-cytometry dataanalysis and statistics; D.M., proteomic data acquisition and
analysis; S.W., U.C.B. cell isolation; F.M.C., data analysis andstatistics; K.S., collection and provision of umbilical cord blood;M.A.K., study design, data monitoring and manuscript writing;G.P., study design, data monitoring, manuscript writing andcommunication and fund provisions.
References
Adams GB. 2008; Deconstructing the he-matopoietic stem cell niche: revealingthe therapeutic potential. Regen Med3: 523–530.
Aizman I, Tate CC, McGrogan M, et al.2009; Extracellular matrix produced bybone marrow stromal cells and bytheir derivative, SB623 cells, supportsneural cell growth. J Neurosci Res87: 3198–3206.
Banu N, Rosenzweig M, Kim H, et al. 2001;Cytokine-augmented culture of haemato-poietic progenitor cells in a novel three-dimensional cell growth matrix. Cytokine13: 349–358.
Brown BN, Freund JM, Han L, et al. 2011;Comparison of three methods for the der-ivation of a biologic scaffold composed ofadipose tissue extracellular matrix. TissueEng Part C Methods 17: 411–421.
Broxmeyer HE, Cooper S, Hass DM, et al.2009; Experimental basis of cord bloodtransplantation. Bone Marrow Transpl44: 627–633.
Chen XD, Dusevich V, Feng JQ, et al. 2007;Extracellular matrix made by bonemarrow cells facilitates expansion ofmarrow-derived mesenchymal progeni-tor cells and prevents their differentia-tion into osteoblasts. J Bone Miner Res22: 1943.
Dellatore SM, Garcia AS, Miller WM. 2008;Mimicking stem cell niches to increasestem cell expansion. Curr Opin Biotechnol19: 534–540.
Dexter TM, Allen TD, Lajtha LG. 1977; Condi-tions controlling the proliferation of hae-mopoietic stem cells in vitro. J Cell Physiol91: 335–344.
Discher DE, Mooney DJ, Zandstra PW. 2009;Growth factors, matrices, and forcescombine and control stem cells. Science324: 1673–1677.
Ehring B, Biber K, Upton TM, et al. 2003;Expansion of HPCs from cord bloodin a novel 3D matrix. Cytotherapy5: 490–499.
Evans ND, Gentleman E, Chen X, et al.2010; Extracellular matrix-mediatedosteogenic differentiation of murineembryonic stem cells. Biomaterials31: 3244–3252.
Feng Q, Chai C, Jiang XS, et al. 2006; Expan-sion of engrafting human hematopoieticstem/progenitor cells in three-dimensionalscaffolds with surface-immobilized fibro-nectin. J Biomed Mater Res A 78: 781–791.
Forsberg EC, Prohaska SS, Katzman S, et al.2005; Differential expression of novel po-tential regulators in hematopoietic stemcells. PLoS Genet 1: e28.
Franke K, Pompe T, Bornhauser M, et al.2007; Engineered matrix coatingsto modulate the adhesion of CD133+
human hematopoietic progenitor cells.Biomaterials 28: 836–843.
Galan I, DeLeon JA, Diaz L, et al. 2007;Effect of a bone marrow microenviron-ment on the ex vivo expansion of umbil-ical cord blood progenitor cells. Int J LabHematol 29: 58–63.
Grunert M, Dombrowski C, Sadasivam M,et al. 2007; Isolation of a native osteoblastmatrix with a specific affinity for BMP2. JMol Histol 38: 393–404.
Guilak F, Cohen DM, Estes BT, et al. 2009;Control of stem cell fate by physical inter-actions with the extracellular matrix. CellStem Cell 5: 17–26.
Hoshiba T, Lu H, Kawazoe N, et al. 2010;Decellularized matrices for tissueengineering. Expert Opin Biol Ther10: 1717–1728.
Issaad C, Croisille L, Katz A, et al. 1993; Amurine stromal cell line allows theproliferation of very primitive humanCD34++/CD38– progenitor cells inlong-term cultures and semi-solid assays.Blood 81: 2916–2924.
Itoh K, Tezuka H, Sakoda H, et al. 1989;Reproducible establishment of hemopoi-etic supportive stromal cell lines frommurine bone marrow. Exp Hematol17: 145–153.
Jones DL, Wagers AJ. 2008; No placelike home: anatomy and function of thestem cell niche. Nat Rev Mol Cell Biol 9:11–21.
Kanai M, Hirayama F, Yamaguchi M, et al.2000; Stromal cell-dependent ex vivo ex-pansion of human cord blood progeni-tors and augmentation of transplantablestem cell activity. Bone Marrow Transpl26: 837–844.
Keller U, Gotze KS, Duyster J, et al. 2002;Murine stromal cells producing hyper-interleukin-6 and Flt3 ligand supportexpansion of early human hematopoieticprogenitor cells without need of exoge-nous growth factors. Leukemia 16:2122–2128.
Kim SH, Jun S, Jang HS, et al. 2005; Identifi-cation of parathyroid hormone-regulatedproteins in mouse bone marrow cells byproteomics. Biochem Biophys Res Commun330: 423–429.
Levi BP, Yilmaz OH, Duester G, et al. 2009;Aldehyde dehydrogenase 1a1 is dispens-able for stem cell function in the mousehematopoietic and nervous systems. Blood113: 1670–1680.
Li Y, Ma T, Kniss DA, et al. 2001; Human cordcell hematopoiesis in three-dimensionalnonwoven fibrous matrices: in vitro sim-ulation of the marrow microenviron-ment. J Hematother Stem Cell Res 10:355–368.
Lund AW, Yener B, Stegemann JP, et al. 2009;The natural and engineered 3D microenvi-ronment as a regulatory cue during stemcell fate determination. Tissue Eng Part BRev 15: 371–380.
Miyoshi H, Murao M, Ohshima N, et al.2011; Three-dimensional culture ofmouse bone marrow cells within a po-rous polymer scaffold: effects of oxygenconcentration and stromal layer onexpansion of haematopoietic progenitorcells. J Tissue Eng Regen Med 5:112–118.
Mohyeldin A, Garzon-Muvdi T, Quinones-Hinojosa A. 2010; Oxygen in stemcell biology: a critical component ofthe stem cell niche. Cell Stem Cell7: 150–161.
Morrison SJ, Spradling AC. 2008; Stem cellsand niches: mechanisms that promotestem cell maintenance throughout life. Cell132: 598–611.
O’Farrell PH. 1975; High resolution two-di-mensional electrophoresis of proteins. JBiol Chem 250: 4007.
Orkin SH, Zon LI. 2008; Hematopoiesis: anevolving paradigm for stem cell biology.Cell 132: 631–644.
Park HW, Shin JS, Kim CW. 2007; Proteomeof mesenchymal stem cells. Proteomics7: 2881–2894.
Rodgers KD, San Antonio JD, Jacenko O.2008; Heparan sulfate proteoglycans: aGAGgle of skeletal–hematopoietic regula-tors. Dev Dyn 237: 2622–2642.
Rossignol F, Solares M, Balanza E, et al.2003; Expression of lactate dehydroge-nase A and B genes in different tissues ofrats adapted to chronic hypobaric hypoxia.J Cell Biochem 89: 67–79.
Satyavani RFA, Sundaram CS, Anabalagan C,et al. 2009; Proteomic analysis of the sidepopulation (SP) cells from murine bonemarrow. J Proteomics Bioinform 2: 398–407.
Shiraki N, Yamazoe T, Qin Z, et al. 2011; Effi-cient differentiation of embryonic stemcells into hepatic cells in vitro using afeeder-free basement membrane substra-tum. PLoS One 6: e24228.
Sullivan M, Galea P, Latif S. 2006; Whatis the appropriate oxygen tension forin vitro culture? Mol Hum Reprod12: 653.
Tan J, Liu T, Hou L, et al. 2010; Maintenanceand expansion of hematopoietic stem/progenitor cells in biomimetic osteoblastniche. Cytotechnology 62: 439–448.
Tursky ML, Collier FM, Ward AC, et al. Inpress; Systematic investigation of oxygenand growth factors in clinically valid exvivo expansion of cord blood CD34+ he-matopoietic progenitor cells. Cytotherapy(In press).
Uchida N, Hsieh MM, Hayakawa J, et al.2011; Optimal conditions for lentiviraltransduction of engrafting human CD34+
cells. Gene Ther 18: 1078–1086.Verfaillie CM, Gupta P, Prosper F, et al.1999; The hematopoietic microenviron-ment: stromal extracellular matrix com-ponents as growth regulators for human
A. Tiwari et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

hematopoietic progenitors. Hematology4: 321–333.
Wilson A, Trumpp A. 2006; Bone-marrowhaematopoietic–stem-cell niches. Nat RevImmunol 6: 93–106.
Yao CL, Hwang SM. 2007; Ex vivo ex-pansion of hematopoietic stem cells
from human cord blood in serum-freeconditions. Methods Mol Biol 407:165–175.
Ye NS, Chen J, Luo GA, et al. 2006; Proteomicprofiling of rat bone marrow mesenchymalstem cells induced by 5-azacytidine. StemCells Dev 15: 665–676.
Zhang A-X, Yu W-H, Ma B-F, et al. 2007;Proteomic identification of differentlyexpressed proteins responsible for 1os-teoblast differentiation from humanmesenchymal stem cells. Mol Cell Bio-chem 304: 167–179.
Haematopoietic progenitor cell expansion on acellular matrices
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term
Copyright © 2022 FDOKUMEN




















