
Evidence of Avian Leukosis Virus Subgroup E and Endogenous Avian Virus in Measles and Mumps Vaccines...
9
JOURNAL OF VIROLOGY, 0022-538X/99/$04.0010 July 1999, p. 5843–5851 Vol. 73, No. 7 Evidence of Avian Leukosis Virus Subgroup E and Endogenous Avian Virus in Measles and Mumps Vaccines Derived from Chicken Cells: Investigation of Transmission to Vaccine Recipients SHIRLEY X. TSANG, 1 WILLIAM M. SWITZER, 1 VEDAPURI SHANMUGAM, 1 JEFFREY A. JOHNSON, 1 CYNTHIA GOLDSMITH, 2 ANTHONY WRIGHT, 1 ALY FADLY, 3 DONALD THEA, 4 HAROLD JAFFE, 1 THOMAS M. FOLKS, 1 AND WALID HENEINE 1 * HIV and Retrovirology Branch, Division of AIDS, STD, and TB Laboratory Research, 1 and Infectious Disease Pathology Activity, National Center for Infectious Diseases, 2 Centers for Disease Control and Prevention, Atlanta, Georgia 30333; Avian Disease and Oncology Laboratory, U.S. Department of Agriculture, East Lansing, Michigan 48823 3 ; and Harvard Institute for International Development, Cambridge, Massachusetts 02138 4 Received 7 December 1998/Accepted 13 April 1999 Reverse transcriptase (RT) activity has been detected recently in all chicken cell-derived measles and mumps vaccines. A study of a vaccine manufactured in Europe indicated that the RT is associated with particles containing endogenous avian retrovirus (EAV-0) RNA and originates from the chicken embryonic fibroblasts (CEF) used as a substrate for propagation of the vaccine. We investigated the origin of RT in measles and mumps vaccines from a U.S. manufacturer and confirm the presence of RT and EAV RNA. Additionally, we provide new evidence for the presence of avian leukosis virus (ALV) in both CEF supernatants and vaccines. ALV pol sequences were first identified in particle-associated RNA by amplification with degenerate retroviral pol primers. ALV RNA sequences from both the gag and env regions were also detected. Analysis of hypervari- able region 2 of env revealed a subgroup E sequence, an endogenous-type ALV. Both CEF- and vaccine-derived RT activity could be blocked by antibodies to ALV RT. Release of ALV-like virus particles from uninoculated CEF was also documented by electron microscopy. Nonetheless, infectivity studies on susceptible 15B 1 chicken cells gave no evidence of infectious ALV, which is consistent with the phenotypes of the ev loci identified in the CEF. PCR analysis of ALV and EAV proviral sequences in peripheral blood mononuclear cells from 33 children after measles and mumps vaccination yielded negative results. Our data indicate that the sources of RT activity in all RT-positive measles and mumps vaccines may not be similar and depend on the particular endogenous retroviral loci present in the chicken cell substrate used. The present data do not support transmission of either ALV or EAV to recipients of the U.S.-made vaccine and provide reassurance for current immunization policies. Human vaccines made from live attenuated viruses have been used effectively worldwide to reduce and prevent mor- bidity and mortality from many viral infections. Most strains of the attenuated viruses were developed by adapting virus strains to certain culture conditions on primary animal-derived cells. The attenuated measles and mumps virus vaccines licensed in the United States are produced by a single manufacturer with primary chicken embryonic fibroblasts (CEF) (43). Measles and mumps vaccines are usually combined with attenuated rubella virus which is produced with human diploid cells, and the trivalent vaccine (MMR) is administered during childhood (14). MMR has been highly efficacious in preventing disease since the early 1970s (13, 30, 43). The general manufacturing regulations for these chicken-derived vaccines require that all avian embryo cell cultures used for propagation of vaccine strains originate from a closed, specific-pathogen-free, healthy flock which has been screened for absence of known chicken bacterial pathogens and viruses, including both avian retrovi- rus groups: the reticuloendotheliosis viruses (REV) and the avian leukosis-sarcoma viruses (ALV) (43). At least six sub- groups of ALV (A to E and J) have been identified in chickens based on differences in the envelope sequences. Only subgroup E viruses are expressed from endogenous sequences that are part of the chicken germ line; all other subgroup viruses are exogenous. The endogenous sequences are usually referred to as endogenous viral (ev) loci (32, 35). Reverse transcriptase (RT), an enzyme present in retrovi- ruses, was recently detected in live attenuated vaccines from several manufacturers (8). The RT-positive vaccines were all derived from chicken cells and included measles, mumps, and yellow fever vaccines (8, 34, 42). RT activity was not detected in the rubella vaccine produced in human cells (42). RT was identified in the vaccines by using a newly developed RT assay that uses PCR amplification as a detection system. This im- proved method has been found to be up to 1 million-fold more sensitive than conventional non-PCR-based RT assays (23, 33). The level of RT activity present in these vaccines was low and could not be detected by conventional RT assays (42). Both the evidence showing association of RT with particles and the observed sensitivity to specific RT inhibitors, such as zidovu- dine triphosphate, suggest a retroviral origin rather than non- specific polymerase activity (8, 34, 42). Weissmahr et al. recently examined the origin of RT in a CEF-derived measles vaccine from a European manufacturer * Corresponding author. Mailing address: HIV and Retrovirology Branch, Centers for Disease Control and Prevention, 1600 Clifton Rd., Mail Stop G-19, Atlanta, GA 30333. Phone: (404) 639-0218. Fax: (404) 639-1174. E-mail: [email protected]. 5843 on February 23, 2015 by guest http://jvi.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Evidence of Avian Leukosis Virus Subgroup E and Endogenous Avian Virus in Measles and Mumps Vaccines...

JOURNAL OF VIROLOGY,0022-538X/99/$04.0010
July 1999, p. 5843–5851 Vol. 73, No. 7
Evidence of Avian Leukosis Virus Subgroup E and EndogenousAvian Virus in Measles and Mumps Vaccines Derived from
Chicken Cells: Investigation of Transmission toVaccine Recipients
SHIRLEY X. TSANG,1 WILLIAM M. SWITZER,1 VEDAPURI SHANMUGAM,1 JEFFREY A. JOHNSON,1
CYNTHIA GOLDSMITH,2 ANTHONY WRIGHT,1 ALY FADLY,3 DONALD THEA,4 HAROLD JAFFE,1
THOMAS M. FOLKS,1 AND WALID HENEINE1*
HIV and Retrovirology Branch, Division of AIDS, STD, and TB Laboratory Research,1 and Infectious Disease PathologyActivity, National Center for Infectious Diseases,2 Centers for Disease Control and Prevention, Atlanta, Georgia 30333;
Avian Disease and Oncology Laboratory, U.S. Department of Agriculture, East Lansing, Michigan 488233;and Harvard Institute for International Development, Cambridge, Massachusetts 021384
Received 7 December 1998/Accepted 13 April 1999
Reverse transcriptase (RT) activity has been detected recently in all chicken cell-derived measles and mumpsvaccines. A study of a vaccine manufactured in Europe indicated that the RT is associated with particlescontaining endogenous avian retrovirus (EAV-0) RNA and originates from the chicken embryonic fibroblasts(CEF) used as a substrate for propagation of the vaccine. We investigated the origin of RT in measles andmumps vaccines from a U.S. manufacturer and confirm the presence of RT and EAV RNA. Additionally, weprovide new evidence for the presence of avian leukosis virus (ALV) in both CEF supernatants and vaccines.ALV pol sequences were first identified in particle-associated RNA by amplification with degenerate retroviralpol primers. ALV RNA sequences from both the gag and env regions were also detected. Analysis of hypervari-able region 2 of env revealed a subgroup E sequence, an endogenous-type ALV. Both CEF- and vaccine-derivedRT activity could be blocked by antibodies to ALV RT. Release of ALV-like virus particles from uninoculatedCEF was also documented by electron microscopy. Nonetheless, infectivity studies on susceptible 15B1 chickencells gave no evidence of infectious ALV, which is consistent with the phenotypes of the ev loci identified in theCEF. PCR analysis of ALV and EAV proviral sequences in peripheral blood mononuclear cells from 33 childrenafter measles and mumps vaccination yielded negative results. Our data indicate that the sources of RT activityin all RT-positive measles and mumps vaccines may not be similar and depend on the particular endogenousretroviral loci present in the chicken cell substrate used. The present data do not support transmission ofeither ALV or EAV to recipients of the U.S.-made vaccine and provide reassurance for current immunizationpolicies.
Human vaccines made from live attenuated viruses havebeen used effectively worldwide to reduce and prevent mor-bidity and mortality from many viral infections. Most strains ofthe attenuated viruses were developed by adapting virus strainsto certain culture conditions on primary animal-derived cells.The attenuated measles and mumps virus vaccines licensed inthe United States are produced by a single manufacturer withprimary chicken embryonic fibroblasts (CEF) (43). Measlesand mumps vaccines are usually combined with attenuatedrubella virus which is produced with human diploid cells, andthe trivalent vaccine (MMR) is administered during childhood(14). MMR has been highly efficacious in preventing diseasesince the early 1970s (13, 30, 43). The general manufacturingregulations for these chicken-derived vaccines require that allavian embryo cell cultures used for propagation of vaccinestrains originate from a closed, specific-pathogen-free, healthyflock which has been screened for absence of known chickenbacterial pathogens and viruses, including both avian retrovi-rus groups: the reticuloendotheliosis viruses (REV) and the
avian leukosis-sarcoma viruses (ALV) (43). At least six sub-groups of ALV (A to E and J) have been identified in chickensbased on differences in the envelope sequences. Only subgroupE viruses are expressed from endogenous sequences that arepart of the chicken germ line; all other subgroup viruses areexogenous. The endogenous sequences are usually referred toas endogenous viral (ev) loci (32, 35).
Reverse transcriptase (RT), an enzyme present in retrovi-ruses, was recently detected in live attenuated vaccines fromseveral manufacturers (8). The RT-positive vaccines were allderived from chicken cells and included measles, mumps, andyellow fever vaccines (8, 34, 42). RT activity was not detectedin the rubella vaccine produced in human cells (42). RT wasidentified in the vaccines by using a newly developed RT assaythat uses PCR amplification as a detection system. This im-proved method has been found to be up to 1 million-fold moresensitive than conventional non-PCR-based RT assays (23, 33).The level of RT activity present in these vaccines was low andcould not be detected by conventional RT assays (42). Both theevidence showing association of RT with particles and theobserved sensitivity to specific RT inhibitors, such as zidovu-dine triphosphate, suggest a retroviral origin rather than non-specific polymerase activity (8, 34, 42).
Weissmahr et al. recently examined the origin of RT in aCEF-derived measles vaccine from a European manufacturer
* Corresponding author. Mailing address: HIV and RetrovirologyBranch, Centers for Disease Control and Prevention, 1600 Clifton Rd.,Mail Stop G-19, Atlanta, GA 30333. Phone: (404) 639-0218. Fax: (404)639-1174. E-mail: [email protected].
5843
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from

(42). By using a novel PCR-based method designed to detectunknown particle-associated RNA sequences with conservedprimers in the tRNA primer binding site from the long termi-nal repeat (LTR), Weissmahr et al. identified RNA sequencesrelated to the endogenous avian virus (EAV-0). They alsodemonstrated that enzymatically active RT and EAV-0 RNAwere physically present together, which was interpreted as in-direct evidence for the presence of EAV virions. Additionalproof for the existence of EAV-0 retrovirus particles, includingvisualization of virions by electron microscopy and demonstra-tion of EAV RT activity that was distinct from other avianretroviral RT, was not shown (42).
EAV proviral sequences are present in the genome of allchickens, including line 0 chickens that have been bred toeliminate any endogenous viral (ev) loci related to ALV (20).EAV sequences are transcriptionally active during embryogen-esis but were not known to be associated with RT-positiveparticles (11). The findings of Weissmahr et al. of RT-associ-ated particles containing EAV RNA may explain the origin ofRT activity in ev2 chicken embryos reported 20 years ago(2–4). The implications of EAV-0 RNA for vaccine recipientsremain unknown.
Since all licensed vaccine producers follow similar manufac-turing regulations regarding screening of source chickens forexogenous ALV and REV infections, it is likely that any RTactivity found to be associated with these vaccines will be ofendogenous origin. However, it is not known whether RTactivity in vaccines from different manufacturers also origi-nates from particles with EAV-0 RNA. Expression of endog-enous retrovirus particles by chick cells and the characteristicsof such particles both are highly variable and depend largely onthe specific genetic profiles of the endogenous retroviral loci(e.g., EAV and ALV) of the cell substrate used. For instance,at least 30 ALV loci have been identified in different chickenstrains, and each chicken can carry multiple ev loci. ALV lociconfer a range of different phenotypes: expression of infectiousALV, expression of viral proteins, or no detectable expressionof any virus-related protein or RNA (32, 35). Therefore, RT inMMR vaccines propagated in chick cell substrates with differ-ent endogenous retroviral loci may be associated with endog-enous particles that have distinct biologic properties and thuspose different risks to vaccine recipients.
Therefore, it is important to study the origin of RT in alllicensed measles and mumps vaccines produced by variousmanufacturers. We focus in this study on the chicken-derivedMMR vaccine used in the United States. Identification of theavian retroviruses responsible for the RT in this vaccine andassessment of their risks of transmission to vaccine recipientsare necessary for a full understanding of the biological signif-icance of this RT activity. This information may be importantfor policy decisions regarding the use of RT-positive vaccines.We found evidence of both endogenous ALV and EAV in theU.S.-made MMR vaccine but were unable to propagate ALVin vitro or to demonstrate transmission of these avian retrovi-rus sequences to vaccine recipients.
MATERIALS AND METHODS
Vaccines, CEF, and CEF culture supernatant. CEF which had not been inoc-ulated with any vaccine virus strains (referred to hereafter as uninoculated orcontrol CEF) were provided by a U.S. MMR vaccine manufacturer. CEF werepooled from chicken embryos derived from a closed, specific-pathogen-free flockof White Leghorn chickens maintained by the manufacturer. Culture superna-tants of the cells were provided by the same manufacturer. Both CEF and CEFculture supernatants were shipped frozen and aliquoted after arrival. Aliquotswere stored at 270°C until use. Lyophilized U.S.-made MMR vaccines werepurchased and were stored at 4°C prior to use. The MMR vaccines were recon-stituted in diluent according to the instructions of the vaccine manufacturer.
Detection of RT activity. Detection of RT activity in cell-free culture super-natants or vaccines was performed by the PCR-based Amp-RT assay, as de-scribed previously (23). Testing was performed on 10 ml or less. Qualitativedetection of Amp-RT products was done by Southern blot hybridization, aspreviously described (23, 45).
Purification of particle-associated RNA. A volume of 250 ml of culture super-natant of uninoculated CEF was used for each CEF RNA extraction. Superna-tants were first clarified by centrifugation at 1,000 rpm for 10 min. Viral particleswere then pelleted by ultracentrifugation at 100,000 3 g (Beckman Ti 60 rotor)for 1 h at 4°C. For MMR vaccines, 50 doses were each dissolved in 0.5 ml ofdiluent and virus particles were ultracentrifuged as was done with the CEFsupernatant. Ultracentrifuged pellets from either preparation were resuspendedin 1 ml of phosphate-buffered saline, and free RNA and DNA were digested with250 ng of RNase (Boehringer Mannheim, Indianapolis, Ind.) per ml and 5 U ofDNase I (Boehringer Mannheim) for 1 h at 37°C in the presence of 10 mMMgCl2. Particles were pelleted by another round of ultracentrifugation, pelletswere lysed with 1 ml of RNA Stat60 buffer (TelTest B Inc., Friendswood, Tex.),and RNA was isolated and purified according to the instructions for use of RNAStat60 (TelTest B Inc.). Nucleic acids were washed with 70% ethanol, air dried,and dissolved in 50 ml of diethyl pyrocarbonate–double-distilled water. IsolatedRNA was treated with RNase-free DNase I for 1 h at 37°C, and DNase I wasinactivated by heating at 95°C for 10 min. To check for the absence of cellularmRNA, we performed an RT-PCR analysis with chicken b-actin primers for eachRNA preparation. Only isolated RNA with undetectable chicken b-actin se-quences was used for the study.
RT-PCR analysis. An RNA template-specific RT-PCR was used for initialdetection of the pol sequence in the particle-associated RNA (37). Primers mopFand mopR were used; they are derived from highly conserved retroviral polsequences corresponding to KVLPQG and YMDDLL, respectively. This assayformat is known to selectively amplify RNA target sequences in the presence oftarget DNA contamination. RNA was reverse transcribed at 37°C for 2 h by amopR primer containing a t30 tag (mopRt30), and RT was inactivated by heatingat 95°C for 10 min. The t30-tagged reverse-transcribed cDNA was then PCRamplified by t30 with forward primer mopF at a high annealing temperature of65°C.
RNA isolated from 5 to 10 ml of CEF supernatant or 1 dose of MMR wasapplied to a 50-ml RT reaction mixture containing 50 mM Tris-HCl (pH 8.3), 10mM MgCl2, 50 mM KCl, 10 mM dithioerythritol; 400 mM deoxynucleosidetriphosphate, 1 U of RNase inhibitor, and 10 U of murine leukemia virus RT(Promega, Madison, Wis.) with 100 ng of reverse primer (R) at 37°C for 2 h,followed by 95°C for 10 min. The cDNA product from the RT step was added toa PCR mixture (total volume of 100 ml) containing 2.5 U of Taq DNA polymer-ase (Perkin-Elmer, Foster City, Calif.) and 103 PCR buffer (Perkin-Elmer) with100 ng of forward primer (F). The cycling conditions were 95°C for 5 min; 35cycles of 1 min at 95°C, 1 min at 55°C, and 1 min at 72°C; and a final step of 7min at 72°C. For each RT-PCR, a control amplification in which no RT had beenadded was included to detect any residual target DNA contamination. Addi-tional negative controls containing water were also included.
Primers used in RT-PCR for ALV pol were as follows: mopF, 59-TGGAAGGTCTT(G/A)CC(C/A)CA(G/A)GG-39; mopR, 59-AGCAA(C/A)A(T/G)ATCATCCAT(G/A)TA-39; t30, 59-GAACATCGATGACAAGGTTAGGTATCGATA-39; mopRt30,59-GAACATCGATGACAAGCTTAGGTATCGATAAGCAA(C/A)A(T/G)ATCATCCAT(G/A)TA-39. The specific probe for HIV-1 pol was59-CAAAATCCAGACATAGTTATCTAT-39. Primers used in RT-PCR forALV gag and env sequences were alvgp85F (59-ACAGTGGTGACAGCGGAT-39) and alvgp85R (59-TTTGGTGAAACTACCTTG-39) for env and alvgagF and(59-TTCGTCCGCCTGTAAAACC-39) and alvgagR (59-CCCCTAATCCCAACCAAAAC-39) for gag. The internal oligonucleotide probe used for hybridizationof the ALV env product was 59-GGTGCATATGGCTACAG-39. The primers forchicken b-actin used in RT-PCR were CAF (59-CAGGCTGTGCTGTCC CTT-39) and CAR (59-GGAGGAG GATGAGGCAGC-39). The specific internalprobe for the chicken b-actin RT-PCR product was 59-AGGTTATAGCTTCACCAC-39. Primers used in RT-PCR for EAV-0 39 pol were EAVFI (59-CCCTTGGAATGTAGTCAC-39) and EAVRI (59-CATGAAGGGTAGCAACAA-39). The internal oligonucleotide probe used for hybridization of this EAV polfragment was 59-AAATCATTCAGACGTTCC-39. The EAV element primersequences were EAVFII (59-GATGTGAGGATGTCGAAGG-39) and EAVRII(59-AACGCAAATCCTAACTCTAT-39).
Cloning and sequencing. RT-PCR products of ALV pol and env were clonedinto pT7Blue vector (Novagen, Madison, Wis.). Plasmid DNA from recombinantclones was extracted with the Midi prep kit (Qiagen, Chatsworth, Calif.) andsequenced by either Sequenase 2.0 (U.S. Biochemical, Cleveland, Ohio) or anautomated sequencer (Perkin-Elmer Applied Biosystems Division). RT-PCRproducts of ALV gag and both EAV pol and EAV element were directly se-quenced by an automated sequencer (Perkin-Elmer Applied Biosystems Divi-sion). The Genetics Computer Group GCG (Madison, Wis.) sequence analysispackage was used for nucleic acid homology searches of the GenBank andEMBL sequence databases.
Electron microscopy. Specimens for ultrastructural studies were prepared asfollows. For CEF supernatant, 250 ml were ultracentrifuged at 43,000 rpm for 1 hat 4°C. For CEF, monolayer cells were scraped, pelleted at 1,000 rpm for 5 min,and then washed once in 0.2 M phosphate buffer. All pellets were subsequently
5844 TSANG ET AL. J. VIROL.
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from

fixed in buffered 2.5% glutaraldehyde, postfixed with 1% buffered osmium, anden bloc stained with 4% uranyl acetate. The pellets were dehydrated through agraded series of alcohol and propylene oxide and embedded in Epon substituteand Araldite (29). Sections were stained with uranyl acetate and lead citrate.
Neutralization of RT activity by anti-RT antisera. The ability of antisera toblock RT activity was measured by quantitative Amp-RT assays, which use anenzyme-linked immunosorbent assay (ELISA)-based, nonradioactive oligonucle-otide probing system to quantitate Amp-RT products (22). The RT neutraliza-tion reactions were set up with serial 10-fold dilutions of 1 ml of antiserum (i.e.,postimmune serum). Control reaction mixtures were made with similar dilutionsof the appropriate preimmune serum. Reactions included 9 ml of the sample and1 ml of serum. The level of RT tested in all samples was adjusted to be in thelinear range of the quantitative Amp-RT assay (22). The anti-avian myeloblas-tosis virus (AMV) RT antiserum and its preimmune serum were purchased fromQuality Biotech Incorporated-Resource Laboratory (Camden, N.J.).
In vitro infectivity assays. Samples of CEF culture supernatant were tested forinfectious exogenous and endogenous ALV, as described by Fadly and Witter(21). Briefly, either 1 ml of CEF supernatants concentrated from 250 ml, asdescribed above, or 1 ml of unconcentrated supernatant was inoculated on 15B1cells in 35-mm plates. Six inoculated 15B1 cultures, including three from theunconcentrated and three from the concentrated CEF supernatants, were mon-itored for production of ALV p27gag protein. At 7 to 9 days postinoculation, celllysates were tested for the presence of ALV p27 antigen by ELISA, as previouslydescribed (21). Control cultures were inoculated with 4,000 infectious units ofRAV-0, a prototypic infectious endogenous ALV isolate.
Proviral DNA PCR. Peripheral blood mononuclear cells (PBMC) or other celllysates were prepared at a concentration of 6 3 106 cells per ml of lysis buffer (50mM KCl, 10 mM Tris-HCl [pH 8.3], 1 mg of gelatin per ml, 0.45% Nonidet P-40,0.45% Tween 20), supplemented with 60 mg of proteinase K per ml at 56°C for1 h, followed by 94°C for 10 min. Aliquots of lysates from 150,000 PBMC fromMMR recipients or U.S. blood donors were PCR amplified for ALV env andEAV env-like sequences. The sensitivities of both PCR assays were measured byspiking known plasmid copies that contain either ALV or EAV target sequenceinto background DNA lysate from 150,000 PBMC. Both assays were found tohave a detection threshold of one copy (see Fig. 7). Standard amplificationconditions of 35 cycles were used, as described above. The PCR products weredetected by Southern blot hybridization of 32P end-labeled internal probes. Theprimers used for PCR detection of integrated proviral ALV sequence wereALVENVF2 (59-CGGTGCATATGGCTACAGATTTTG-39 and ALVENVR2(59-TTTCCACAACATCCGCTGACATTA-39); the specific probe wasALVENVP1 (59-AAGGAAATTAATGAGACAGAGCCG-39). The primersused for EAV were EAVF10 (59-ACAGAAGATCAAGATGCAGGCCGA-39)and EAVR10 (59-GCTCCTGAATGCGCTGATACACGT-39); the specificprobe was EAVP1 (59-CCGTGGCTAAAACAAATGCTT-39).
Genomic DNA from CEF was also isolated, by using the Easy-DNA kit(Invitrogen, San Diego, Calif.), and proviral ALV DNA was amplified accordingto the Expand long-template PCR system (Boehringer Mannheim). pRSA-2 is aRuppin-Schmidt A strain of ALV plasmid DNA used as a positive control. Theprimer pairs used for proviral DNA pol-env and env 39 LTR PCR wereALVPOLF (59-ATGACCTGTTCTCCCACTATCT-39) and RSEIIR1 (59-AACGGCTCTGTCTCATTAATTTCCT-39) for pol-env and ENVGp85F2 (59-ACAGTGGTGACAGCGGAT-39) and REVLTRSp (59-TGTGGTGAATGGTAAAATGGCG-39) for env 39 LTR. The specific probe used for env-E was 59-TGTGGAGATGTGCAGACAGTCAAATCCCCC-39.
Typing of ev loci in CEF by locus-specific PCR analysis. CEF DNA wasanalyzed for the presence of ev loci by using PCR-based methods described byBenkel (6). These validated methods are locus specific and use PCR primers thattarget ev LTRs and the unique flanking sequences of integration sites. Theamplifications produce diagnostic fragments of specific lengths which are indic-ative of the presence or absence of the locus in chicken cells. CEF can be typedas homozygous negative or positive for each locus. Heterozygosity is also indi-cated by the presence of both negative and positive diagnostic fragments. Wetested for the presence of six ev loci known to exist in White Leghorn chickens.The loci tested (and their corresponding phenotypes) are ev-1 (Gag2 Pol2 Env2
[very low level paticle producer]), ev-2 (infectious particles), ev-3 (Gag1 Env1),ev-6 (Gag2 Env1), ev-7 (RT1 [noninfectious particles]), ev-9 (Gag2 Env1), ev-12(infectious particles), and ev-21 (infectious particles).
Vaccine recipients and other test populations. Thirty-three children with doc-umented dates of MMR vaccination were included in the study. Pre-MMRsamples were obtained during the 6 months before the first MMR vaccination.Post-MMR samples were obtained 6 to 12 months after the first vaccination withMMR. Ninety-nine anonymous U.S. blood donors were also included and usedas controls.
Nucleotide sequence accession numbers. ALV sequences determined in thisstudy have been deposited in GenBank under accession no. AF087829,AF087830, and AF087831.
RESULTS
RT activity in CEF supernatants and CEF-derived vaccines.We determined the presence of RT activity by using Amp-RT,
which can detect RT activity from approximately 1 to 10 hu-man immunodeficiency virus type 1 (HIV-1) virions (22). RTactivity was detected in several lots of monovalent measles andmumps vaccines, as well as in trivalent MMR vaccines from aU.S. vaccine manufacturer. Testing of end-point 10-fold dilu-tions from a reconstituted 0.5-ml MMR vaccine showed thatthe detectable RT activity titer is equivalent to 0.01 ml ofvaccine (Fig. 1A, lanes 5 to 8). RT was also detected in super-natants of CEF that had not yet been inoculated with anyvaccine virus (Fig. 1A, lane 4). These results suggest that CEFis the source of RT in the vaccines. Amp-RT activity was alsofound on sucrose-banded CEF supernatants, with peak activityin a fraction with a density of 1.15 g/ml, suggesting that thisactivity is associated with retrovirus particles (data not shown).
Detection and identification of ALV pol RNA in CEF super-natants. To characterize the nucleotide sequence of any puta-tive retrovirus expressed in CEF culture, we analyzed the par-ticle-associated RNA that was extracted from uninoculatedCEF supernatants. The method used for isolating this RNAselects for pelletable particle-associated RNA and eliminatesfree DNA or RNA contaminants of cellular origin by pretreat-ment with RNases and DNases prior to extraction of particle-associated RNA. We have confirmed the absence of contam-ination of these RNA preparations with either cellular DNA orRNA. The negative PCR result (Fig. 1B) for chicken b-actinmRNA, a highly expressed transcript in chicken cells, reflectsthe lack of contamination with cellular mRNA. Also, the non-detectable RT-PCR product in the control reactions with noRT demonstrates the absence of contamination with any DNAin the CEF RNA (lane 5 of Fig. 1C).
To identify the presence of unknown RNA retroviral se-quences in the particle-associated RNA, we used an RT-PCRstrategy that employs primers derived from two sequences(KVLPQG and YMDDLL) that are conserved in all knownretroviral polymerase (pol) genes (36). These primers permitthe identification of a wide spectrum of pol-related sequencesto amplify virus-specific internal sequence (36, 44). A similarstrategy with primers from the YMDD motif of RT and an-other highly conserved motif in the protease gene has alsobeen successfully used to amplify pro-pol sequences from un-characterized retroviruses (31, 38). A positive PCR signal ofthe expected size was observed in RT-PCRs containing CEFRNA as well as in the HIV-1 positive control (Fig. 1C, lanes 1and 2, respectively). No PCR products were seen in the controlreactions with no added RT (Fig. 1C, lanes 5 and 6). This resultdemonstrates the absence of DNA that can be amplified bythese generic pol primers, thus confirming that the RT-PCRproduct in the CEF reaction originated from RNA only. Hy-bridization with an internal HIV-1-specific probe was positiveonly with the RT-PCR product of the HIV-1 control (Fig. 1D,lane 2), not with that from CEF RNA (Fig. 1D, lane 1). Thisfinding confirms the identity of the HIV-1 PCR product. Wehave cloned the RT-PCR pol fragment from CEF RNA andsequenced three representative clones. All internal 90-bp se-quences from the three clones were identical and were 97 to100% homologous to pol sequences of various ALV isolates (7,12).
Identification of endogenous ALV. To confirm the presenceof ALV RNA in CEF supernatants and to identify the sub-group of ALV, additional particle-associated gag and env ALVsequences were amplified from CEF RNA by ALV-specificRT-PCR analysis. Since ALV subgroups have distinct enve-lope glycoproteins which confer different tropisms to the virusfor both avian and nonavian cells, defining the env subgroup ofthe ALV in CEF RNA is important for predicting cellulartropism (10). Both env and gag sequences were detected (Fig.
VOL. 73, 1999 ALV AND EAV IN CEF-DERIVED MMR VACCINES 5845
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from
2A, lane 1), and both amplifications originated from RNA,since PCRs that were not reverse transcribed had no detect-able products (Fig. 2A, lane 4). The env sequence selected foramplification included hypervariable region 2 (hr2) of the en-
velope (gp85) gene of ALV, which is a subgroup-specific re-gion and thus can allow the differentiation among all fiveknown chicken ALV subgroups (A to E) (10, 16). The gag andenv RT-PCR products were cloned and sequenced. The gagsequence showed the highest homology (96 to 98%) to ALVgag sequences (12, 40). The hr2 sequence was highly related toendogenous subgroup E env sequences (99% homology at thenucleotide level) (19) but less related to hr2 sequences ofexogenous subgroups A to D (74 to 84% homology) (9, 10, 19).Two proviral regions encompassing an ;3-kb pol-env fragmentand a ;1.6-kb env 39 LTR sequence were also analyzed byPCR amplification of CEF DNA. Both products were ampli-fied, and both hybridized with an env-E-specific probe (datanot shown). These data confirm the presence of large endog-enous ALV sequences in these CEF.
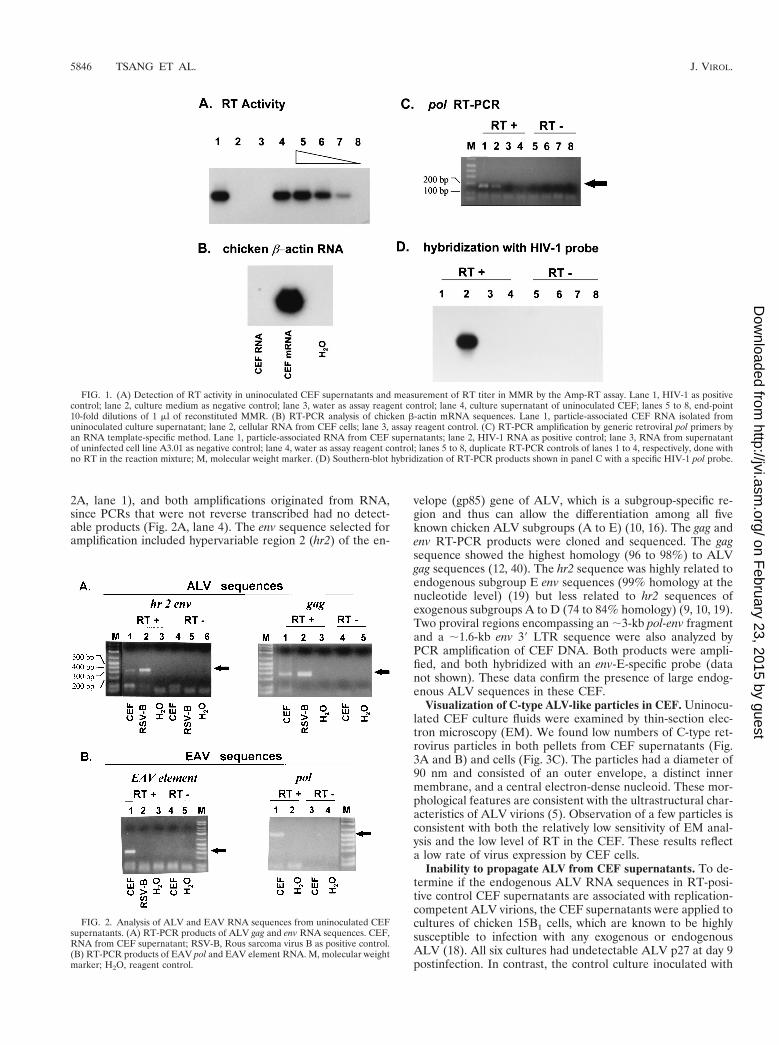
Visualization of C-type ALV-like particles in CEF. Uninocu-lated CEF culture fluids were examined by thin-section elec-tron microscopy (EM). We found low numbers of C-type ret-rovirus particles in both pellets from CEF supernatants (Fig.3A and B) and cells (Fig. 3C). The particles had a diameter of90 nm and consisted of an outer envelope, a distinct innermembrane, and a central electron-dense nucleoid. These mor-phological features are consistent with the ultrastructural char-acteristics of ALV virions (5). Observation of a few particles isconsistent with both the relatively low sensitivity of EM anal-ysis and the low level of RT in the CEF. These results reflecta low rate of virus expression by CEF cells.
Inability to propagate ALV from CEF supernatants. To de-termine if the endogenous ALV RNA sequences in RT-posi-tive control CEF supernatants are associated with replication-competent ALV virions, the CEF supernatants were applied tocultures of chicken 15B1 cells, which are known to be highlysusceptible to infection with any exogenous or endogenousALV (18). All six cultures had undetectable ALV p27 at day 9postinfection. In contrast, the control culture inoculated with
FIG. 1. (A) Detection of RT activity in uninoculated CEF supernatants and measurement of RT titer in MMR by the Amp-RT assay. Lane 1, HIV-1 as positivecontrol; lane 2, culture medium as negative control; lane 3, water as assay reagent control; lane 4, culture supernatant of uninoculated CEF; lanes 5 to 8, end-point10-fold dilutions of 1 ml of reconstituted MMR. (B) RT-PCR analysis of chicken b-actin mRNA sequences. Lane 1, particle-associated CEF RNA isolated fromuninoculated culture supernatant; lane 2, cellular RNA from CEF cells; lane 3, assay reagent control. (C) RT-PCR amplification by generic retroviral pol primers byan RNA template-specific method. Lane 1, particle-associated RNA from CEF supernatants; lane 2, HIV-1 RNA as positive control; lane 3, RNA from supernatantof uninfected cell line A3.01 as negative control; lane 4, water as assay reagent control; lanes 5 to 8, duplicate RT-PCR controls of lanes 1 to 4, respectively, done withno RT in the reaction mixture; M, molecular weight marker. (D) Southern-blot hybridization of RT-PCR products shown in panel C with a specific HIV-1 pol probe.
FIG. 2. Analysis of ALV and EAV RNA sequences from uninoculated CEFsupernatants. (A) RT-PCR products of ALV gag and env RNA sequences. CEF,RNA from CEF supernatant; RSV-B, Rous sarcoma virus B as positive control.(B) RT-PCR products of EAV pol and EAV element RNA. M, molecular weightmarker; H2O, reagent control.
5846 TSANG ET AL. J. VIROL.
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from
RAV-0 was positive for ALV p27 protein (data not shown).These findings do not support the presence of infectious ALVparticles in the RT-positive CEF fluids.
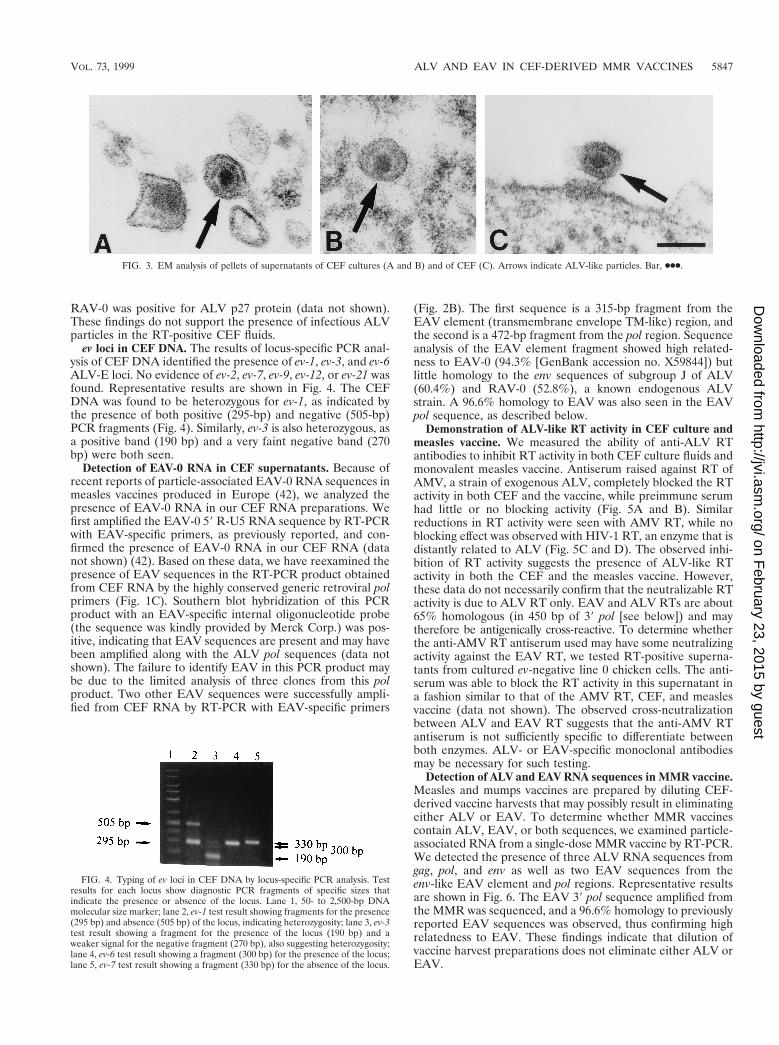
ev loci in CEF DNA. The results of locus-specific PCR anal-ysis of CEF DNA identified the presence of ev-1, ev-3, and ev-6ALV-E loci. No evidence of ev-2, ev-7, ev-9, ev-12, or ev-21 wasfound. Representative results are shown in Fig. 4. The CEFDNA was found to be heterozygous for ev-1, as indicated bythe presence of both positive (295-bp) and negative (505-bp)PCR fragments (Fig. 4). Similarly, ev-3 is also heterozygous, asa positive band (190 bp) and a very faint negative band (270bp) were both seen.
Detection of EAV-0 RNA in CEF supernatants. Because ofrecent reports of particle-associated EAV-0 RNA sequences inmeasles vaccines produced in Europe (42), we analyzed thepresence of EAV-0 RNA in our CEF RNA preparations. Wefirst amplified the EAV-0 59 R-U5 RNA sequence by RT-PCRwith EAV-specific primers, as previously reported, and con-firmed the presence of EAV-0 RNA in our CEF RNA (datanot shown) (42). Based on these data, we have reexamined thepresence of EAV sequences in the RT-PCR product obtainedfrom CEF RNA by the highly conserved generic retroviral polprimers (Fig. 1C). Southern blot hybridization of this PCRproduct with an EAV-specific internal oligonucleotide probe(the sequence was kindly provided by Merck Corp.) was pos-itive, indicating that EAV sequences are present and may havebeen amplified along with the ALV pol sequences (data notshown). The failure to identify EAV in this PCR product maybe due to the limited analysis of three clones from this polproduct. Two other EAV sequences were successfully ampli-fied from CEF RNA by RT-PCR with EAV-specific primers
(Fig. 2B). The first sequence is a 315-bp fragment from theEAV element (transmembrane envelope TM-like) region, andthe second is a 472-bp fragment from the pol region. Sequenceanalysis of the EAV element fragment showed high related-ness to EAV-0 (94.3% [GenBank accession no. X59844]) butlittle homology to the env sequences of subgroup J of ALV(60.4%) and RAV-0 (52.8%), a known endogenous ALVstrain. A 96.6% homology to EAV was also seen in the EAVpol sequence, as described below.
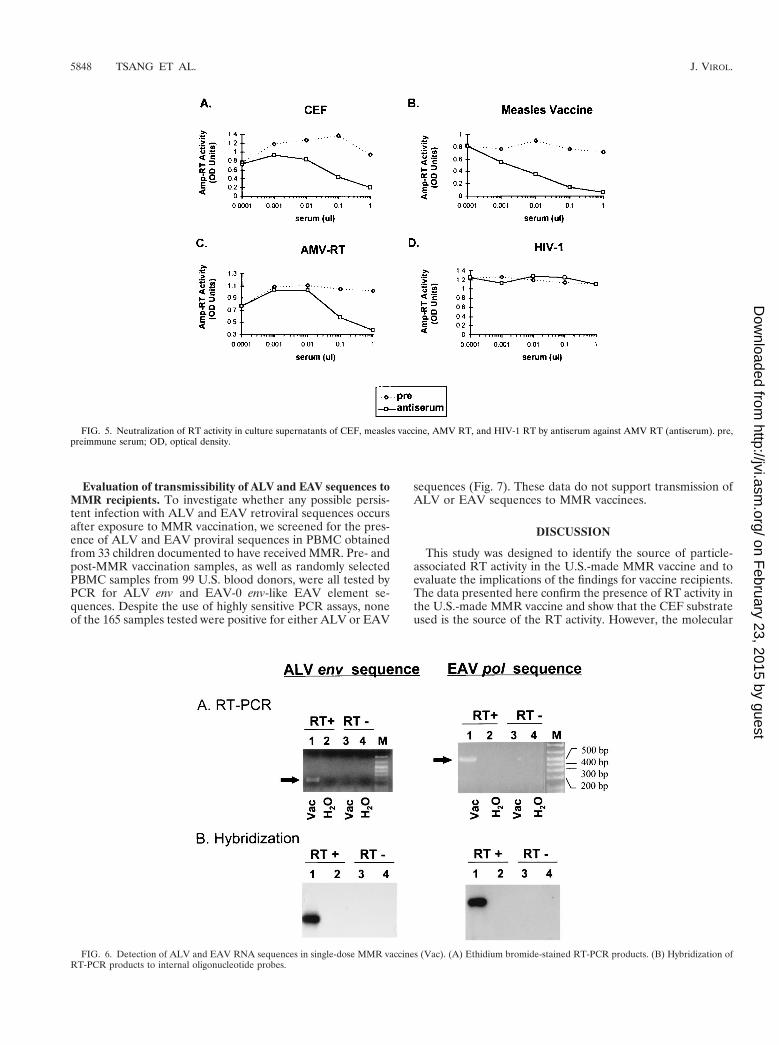
Demonstration of ALV-like RT activity in CEF culture andmeasles vaccine. We measured the ability of anti-ALV RTantibodies to inhibit RT activity in both CEF culture fluids andmonovalent measles vaccine. Antiserum raised against RT ofAMV, a strain of exogenous ALV, completely blocked the RTactivity in both CEF and the vaccine, while preimmune serumhad little or no blocking activity (Fig. 5A and B). Similarreductions in RT activity were seen with AMV RT, while noblocking effect was observed with HIV-1 RT, an enzyme that isdistantly related to ALV (Fig. 5C and D). The observed inhi-bition of RT activity suggests the presence of ALV-like RTactivity in both the CEF and the measles vaccine. However,these data do not necessarily confirm that the neutralizable RTactivity is due to ALV RT only. EAV and ALV RTs are about65% homologous (in 450 bp of 39 pol [see below]) and maytherefore be antigenically cross-reactive. To determine whetherthe anti-AMV RT antiserum used may have some neutralizingactivity against the EAV RT, we tested RT-positive superna-tants from cultured ev-negative line 0 chicken cells. The anti-serum was able to block the RT activity in this supernatant ina fashion similar to that of the AMV RT, CEF, and measlesvaccine (data not shown). The observed cross-neutralizationbetween ALV and EAV RT suggests that the anti-AMV RTantiserum is not sufficiently specific to differentiate betweenboth enzymes. ALV- or EAV-specific monoclonal antibodiesmay be necessary for such testing.
Detection of ALV and EAV RNA sequences in MMR vaccine.Measles and mumps vaccines are prepared by diluting CEF-derived vaccine harvests that may possibly result in eliminatingeither ALV or EAV. To determine whether MMR vaccinescontain ALV, EAV, or both sequences, we examined particle-associated RNA from a single-dose MMR vaccine by RT-PCR.We detected the presence of three ALV RNA sequences fromgag, pol, and env as well as two EAV sequences from theenv-like EAV element and pol regions. Representative resultsare shown in Fig. 6. The EAV 39 pol sequence amplified fromthe MMR was sequenced, and a 96.6% homology to previouslyreported EAV sequences was observed, thus confirming highrelatedness to EAV. These findings indicate that dilution ofvaccine harvest preparations does not eliminate either ALV orEAV.
FIG. 3. EM analysis of pellets of supernatants of CEF cultures (A and B) and of CEF (C). Arrows indicate ALV-like particles. Bar, ●●●.
FIG. 4. Typing of ev loci in CEF DNA by locus-specific PCR analysis. Testresults for each locus show diagnostic PCR fragments of specific sizes thatindicate the presence or absence of the locus. Lane 1, 50- to 2,500-bp DNAmolecular size marker; lane 2, ev-1 test result showing fragments for the presence(295 bp) and absence (505 bp) of the locus, indicating heterozygosity; lane 3, ev-3test result showing a fragment for the presence of the locus (190 bp) and aweaker signal for the negative fragment (270 bp), also suggesting heterozygosity;lane 4, ev-6 test result showing a fragment (300 bp) for the presence of the locus;lane 5, ev-7 test result showing a fragment (330 bp) for the absence of the locus.
VOL. 73, 1999 ALV AND EAV IN CEF-DERIVED MMR VACCINES 5847
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from
Evaluation of transmissibility of ALV and EAV sequences toMMR recipients. To investigate whether any possible persis-tent infection with ALV and EAV retroviral sequences occursafter exposure to MMR vaccination, we screened for the pres-ence of ALV and EAV proviral sequences in PBMC obtainedfrom 33 children documented to have received MMR. Pre- andpost-MMR vaccination samples, as well as randomly selectedPBMC samples from 99 U.S. blood donors, were all tested byPCR for ALV env and EAV-0 env-like EAV element se-quences. Despite the use of highly sensitive PCR assays, noneof the 165 samples tested were positive for either ALV or EAV
sequences (Fig. 7). These data do not support transmission ofALV or EAV sequences to MMR vaccinees.
DISCUSSION
This study was designed to identify the source of particle-associated RT activity in the U.S.-made MMR vaccine and toevaluate the implications of the findings for vaccine recipients.The data presented here confirm the presence of RT activity inthe U.S.-made MMR vaccine and show that the CEF substrateused is the source of the RT activity. However, the molecular
FIG. 5. Neutralization of RT activity in culture supernatants of CEF, measles vaccine, AMV RT, and HIV-1 RT by antiserum against AMV RT (antiserum). pre,preimmune serum; OD, optical density.
FIG. 6. Detection of ALV and EAV RNA sequences in single-dose MMR vaccines (Vac). (A) Ethidium bromide-stained RT-PCR products. (B) Hybridization ofRT-PCR products to internal oligonucleotide probes.
5848 TSANG ET AL. J. VIROL.
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from

data revealed the presence of endogenous ALV RNA in ad-dition to EAV RNA sequences not only in CEF supernatantsbut also in single-dose MMR vaccines. Neutralization of ALV-like RT activity with anti-AMV RT antibodies and visualiza-tion of particles with typical C-type ALV-like morphology pro-vide evidence that the detected ALV sequences are associatedwith ALV particles. Infectivity studies showed no evidence ofinfectious ALV, which is consistent with the predicted pheno-types of all three ev loci identified in CEF DNA. The negativeresults of PCR screening of MMR vaccine recipients does notsupport transmission of either ALV or EAV to these children.
Our findings of particle-associated RNA sequences ofEAV-0 support those of Weissmahr et al., who examined CEFfluids from a European vaccine manufacturer (42). The rea-sons for the discrepant result regarding the detection of en-dogenous ALV in our study but not in that of Weissmahr et al.are not clear but may be due to differences in the lines ofchickens used by the two manufacturers, as well as the effi-ciency of detection of ALV sequences by the methods used. Itis possible that the CEF analyzed by Weissmahr et al. eitherare free of ev loci or have different ev loci that are incapable ofexpressing ALV particles (e.g., ev-4, ev-5, and ev-8) (35). How-ever, this possibility cannot be ascertained until the ev types ofboth CEF substrates are determined.
The detection of endogenous ALV sequences in the U.S.-made vaccine demonstrates that different retrovirus particles,with potentially different properties and risks to vaccinees, mayaccount for the RT activity in the currently used RT-positiveMMR vaccines. The differences in the source of RT among thevarious RT-positive vaccines should not be unexpected, sincedifferent vaccine manufacturers use different chick cell sub-strates with distinct endogenous retroviral loci. Therefore, ourdata highlight the importance of studying independently theorigin and risks of each of the currently used RT-positivevaccines.
We found no evidence of infectious ALV that can replicatein indicator chicken 15B1 cells, despite the use of concentratedCEF inocula. These data suggest the presence of noninfectious
endogenous ALV particles in the CEF fluids. Nonchickencells, such as turkey embryo fibroblasts and Japanese quailprimary embryonic cells, have been used previously as suscep-tible cells for ALV subgroup E replication (27, 34). Furtherevaluation of infectious ALV from CEF fluids and MMR vac-cine on these nonchicken cells may be warranted to confirmthe present results seen on the chicken 15B1 cells.
The lack of evidence of infectious ALV suggests also theabsence of ev loci in CEF DNA that are associated with ex-pression of endogenous infectious ALV (e.g., ev-2, ev-10, ev-12,and ev-21) (35). These data are consistent with ev-1, ev-3, andev-6, which were identified in the CEF DNA. ev-3 and ev-6have large deletions in the gag-pol and 59 LTR gag regions,respectively, and therefore cannot independently generate ma-ture particles (35). ev-1 possesses a full-length genome butproduces low levels of noninfectious particles (17). Additionalanalysis of CEF DNA to identify the presence of other ev locimay be necessary.
Recently, an ALV subgroup J strain was identified in chick-ens and was found to possess a recombinant genome consistingpredominantly of an exogenous-type ALV backbone and anEAV-like env gene (1, 39). The evidence in our study for thepresence of both EAV and ALV RNA raise questions onwhether the detected EAV and ALV RNAs are copackaged inthe same particles or are parts of a recombinant genome sim-ilar to that seen in subgroup J ALV. While our present datacannot address the question of ALV-EAV copackaging, sev-eral observations argue against the presence of a subgroupJ-like virus in the CEF tested. First, we have not found evi-dence of ALV replication in 15B1 cells, which are known to besusceptible to infection with subgroup J virus (21). Second, theparticle-associated ALV env sequences identified in the CEFsupernatants were closest to the endogenous ALV sequencesand had little homology with that of subgroup J. Third, CEFfor vaccine production are obtained from a closely monitoredspecific-pathogen-free flock of chicken. Accidental introduc-tion of subgroup J ALV into this flock should be easily de-
FIG. 7. PCR analysis of PBMC of MMR recipients for the presence of ALV (A) or EAV (B) proviral DNA sequences. The detection thresholds of known copynumbers of target sequences are shown in the righthand panels. Cont, positive controls containing 1 or 1,000 copies; H2O, reagent control; pre and post, samples fromvaccine recipients obtained before and after MMR vaccination, respectively.
VOL. 73, 1999 ALV AND EAV IN CEF-DERIVED MMR VACCINES 5849
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from

tected by routine screening for ALV infection and by theability of this virus to quickly cause disease in chickens.
Our inability to detect either ALV or EAV sequences inPBMC samples from any vaccine recipient provides more di-rect evidence for the absence of infection with either ALV orEAV. These negative results were observed despite the use ofhighly sensitive PCR assays that employ specific primers andprobes derived from vaccine-associated EAV and ALV se-quences. Confirmation of these results by serologic screening,however, may be necessary. The relatively small number ofMMR recipients studied here and the single tissue (PBMC)analyzed from these subjects limit the strength of our conclu-sions regarding transmissibility. Therefore, additional labora-tory surveillance of EAV and ALV infection in recipients ofthis as well as other chicken-derived vaccines may be prudent.Nevertheless, the observed lack of evidence of either ALV orEAV infection in the vaccine recipients is consistent with thenoninfectious nature of ALV seen in this study and with otherin vitro data reported recently on the inability of other RT-positive uninoculated CEF fluids to infect a variety of humanand other mammalian cells, including PBMC (27, 34).
Little or no information is available from previous humanstudies on the prevalence of antibodies to subgroup E ALV. Afew studies have looked at risks of ALV infections in humansby serologic testing for antibodies to ALV and have reportedmostly negative or false-positive results (15, 25, 26). In manyprevious studies, the use of exogenous subgroup-specific neu-tralization assays limited the utility of the data in addressingrisks of human infection with subgroup E ALV. A more recentstudy reported positive Western blot results to ALV gag pro-teins in poultry workers and some persons with no occupa-tional exposure to ALV (24). However, additional evidence ofactive ALV infection in these persons, including virus isolationand detection of ALV sequences, was lacking. No serologictesting for antibodies to EAV in humans has been previouslyreported.
Documentation of ALV and EAV in MMR raises questionsabout the adequacy of using the present CEF as substrates forvaccine propagation and whether changing to a cell substratethat has no detectable RT activity is necessary. CEF originat-ing from line 0 chickens may provide a more suitable avian cellsubstrate that does not express any ALV. However, we foundthat culture fluids of CEF cells from line 0 chickens were alsoRT positive, most likely due to EAV expression. Therefore,obtaining an RT-negative substrate may require a change fromchicken cells to RT-negative cells from a different species.Since the cell substrate is also critical to the attenuation of thevaccine viruses, changing the cell type may have an unpredict-able effect on both the safety and the efficacy of the vaccine.Thus, any suggestion to change cell substrates must be ap-proached cautiously by balancing potential unproven and the-oretical risks associated with ALV-EAV against the risk ofpossibly compromising established vaccine safety or efficacy.
There is currently no evidence that exposure to ALVthrough vaccination is associated with adverse events. Ourinability to demonstrate infectious virus or to find any ALV-EAV sequences in recipients of the U.S.-made MMR vaccinesupports the safety of this vaccine product and suggests thatthese endogenous viruses are not xenotropic. The endogenoussubgroup E ALV virus is not known to be oncogenic in do-mestic fowl, and EAV-0 has not been associated with anyknown diseases in chickens (28). While exposure to exogenous-type ALV in preparations of yellow fever vaccine has beenpreviously demonstrated with vaccine recipients, no increase intumors has been found among the vaccinees (41). The proven
efficacy of MMR and the absence of evidence of any harm dueto MMR vaccination support current immunization policies.
ACKNOWLEDGMENTS
This work was supported in part by The National Vaccine ProgramOffice.
We thank Alison Mawle, Brian Mahy, William Bellini, Rima Khab-baz, Rafael Harpaz, Gina Terraciano, John Livengood, Arifa Khan,Andrew Lewis, Keith Peden, and Robert Breiman for critical reviewsof the manuscript.
REFERENCES
1. Bai, J., L. N. Payne, and M. A. Skinner. 1995. HPRS-103 (exogenous avianleukosis virus, subgroup J) has an env gene related to those of endogenouselements EAV-0 and E51 and an E element found previously only in sar-coma viruses. J. Virol. 69:779–784.
2. Bauer, G., R. R. Friis, G. Jilek, and P. H. Hofscneider. 1978. Purification andcharacterization of particles containing RNA-dependent DNA polymerase,in the allantoic fluid of uninfected leukosis virus-free chicken eggs. Biochim.Biophys. Acta 518:125–137.
3. Bauer, G., R. R. Friis, H. Mattersberger, and P. H. Hofscneider. 1978.Controlled release of particle-associated RNA-dependent DNA polymeraseby primary chick embryo cell cultures. Exp. Cell Res. 117:383–392.
4. Bauer, G., and P. H. Hofscneider. 1976. An RNA-dependent DNA polymer-ase, different from the known viral reverse transcriptase, in the chickensystem. Proc. Natl. Acad. Sci. USA 73:3025–3029.
5. Beard, J. W. 1973. Oncornaviruses. 1. The avian tumor virus, p. 261–281. InA. J. Dalton and F. Haguenau (ed.), Ultrastructure of animal viruses andbacteriophages: an atlas. Academic Press, Inc., New York, N.Y.
6. Benkel, B. F. 1998. Locus-specific diagnostic test for endogenous avian leu-kosis-type viral loci in chickens. Poult. Sci. 77:1027–1035.
7. Bieth, E., and J.-L. Darlix. 1991. Complete nucleotide sequence of a highlyinfectious avian leukosis virus. Nucleic Acids Res. 20:367.
8. Boni, J., J. Stalder, F. Reigel, and J. Schupbach. 1996. Detection of reversetranscriptase activity in live attenuated virus vaccines. Clin. Diagn. Virol.5:43–53.
9. Bova, C. A., J. P. Manfredi, and R. Swanstrom. 1986. env genes of avianretroviruses: nucleotide sequence and molecular recombinants define hostrange determinants. Virology 152:343–354.
10. Bova, C. A., J. C. Olson, and R. Swanstrom. 1988. The avian retrovirus envgene family: molecular analysis of host range and antigenic variants. J. Virol.62:75–83.
11. Boyce-Jacino, M. T., R. Resnick, and A. J. Faras. 1989. Structural andfunctional characterization of the unusually short long terminal repeats andtheir adjacent regions of a novel endogenous avian retrovirus. Virology173:157–166.
12. Broome, S., and W. B. Gilbert. 1985. Nucleotide sequence of Rous sarcomavirus. Cell 40:537–546.
13. Centers for Disease Control and Prevention. 1998. Advances in global mea-sles control and elimination: summary of the 1997 international meeting.Morbid. Mortal. Weekly Rep. 47:1–23.
14. Centers for Disease Control and Prevention. 1996. Immunization of adoles-cents: recommendations of the Advisory Committee on Immunization Prac-tices, the American Academy of Pediatrics, the American Academy of Fam-ily Physicians, and the American Medical Association. Morbid. Mortal.Weekly Rep. 47:1–16.
15. Choudat, D., G. Dambrine, B. Delemotte, and F. Coudert. 1996. Occupa-tional exposure to poultry and prevalence of antibodies against Marek’sdisease virus and avian leukosis retroviruses. Occup. Environ. Med. 53:403–410.
16. Coffin, J. M. 1989. Genetic variation in avian retroviruses. Dev. Biol. Stand.72:123–132.
17. Conklin, K. F., J. M. Coffin, H. L. Robinson, M. Groudine, and R. Eisenman.1982. Role of methylation in the induced and spontaneous expression of theavian endogenous virus ev-1: DNA structure and gene products. Mol. Cell.Biol. 2:638–652.
18. Crittenden, L. B., S. McMahon, M. S. Halpern, and A. M. Fadly. 1987.Embryonic infection with the endogenous avian leukosis virus Rous-associ-ated virus-0 alters responses to exogenous avian leukosis virus infection.J. Virol. 61:722–725.
19. Dorner, A. J., J. P. Stoye, and J. M. Coffin. 1985. Molecular basis of hostrange variation in avian retroviruses. J. Virol. 53:32–39.
20. Dunwiddie, C., and A. J. Faras. 1985. Presence of retrovirus reverse tran-scriptase-related gene sequences in avian cells lacking endogenous avianleukosis viruses. Proc. Natl. Acad. Sci. USA 82:5097–5101.
21. Fadly, A. M., and R. L. Witter. 1998. Oncornaviruses: leukosis/sarcoma andreticuloendotheliosis, p. 185–196. In D. E. Swayne, J. R. Glisson, M. W.Jackwood, J. E. Pearsonand, and W. Reed (ed.), A laboratory manual forisolation and identification of avian pathogens. American Association of
5850 TSANG ET AL. J. VIROL.
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from

Avian Pathologists, Kennett Square, Pa.22. Garcia-Lerma, G., S. Yamamoto, M. Gomez-Cano, V. Soriano, T. A. Green,
M. P. Busch, T. M. Folks, and W. Heneine. 1998. Measurement of humanimmunodeficiency virus type 1 plasma virus load based on reverse transcrip-tase (RT) activity: evidence of variabilities in levels of virion-associatedRT. J. Infect. Dis. 177:1221–1229.
23. Heneine, W., S. Yamamoto, W. M. Switzer, T. J. Spira, and T. M. Folks.1995. Detection of reverse transcriptase by a highly sensitive assay in serafrom persons infected with human immunodeficiency virus type 1. J. Infect.Dis. 171:1210–1216.
24. Johnson, E. S., L. G. Nicholson, and D. T. Durack. 1995. Detection ofantibodies to avian leukosis/sarcoma viruses (ALSV) and reticuloendotheli-osis viruses (REV) in humans by ELISA. Cancer Detect. Prev. 19:395–404.
25. Johnson, E. S. 1994. Poultry oncogenic retroviruses and humans. CancerDetect. Prev. 18:9–30.
26. Johnson, E. S., L. Overby, and R. Philpot. 1995. Detection of antibodies toavian leukosis/sarcoma viruses in humans by Western blot assay. CancerDetect. Prev. 19:472–486.
27. Khan, A. S., T. Maudru, A. Thompson, J. Muller, S. A. Aaronson, J. F. Sears,and K. W. C. Peden. 1998. The reverse transcriptase activity in cell-freemedium of chicken embryo fibroblast cultures is not associated with a rep-lication-competent retrovirus. J. Clin. Virol. 11:7–18.
28. Motta, J. V., L. B. Crittenden, H. G. Purchase, H. A. Stone, and R. L. Witter.1975. Low oncogenic potential of avian endogenous RNA tumor virus in-fection or expression. J. Natl. Cancer Inst. 55:687.
29. Mullenhauer, H. H. 1964. Plastic embedding mixtures for use in electronmicroscopy. Stain Technol. 39:11–114.
30. Orenstein, W. A., L. E. Markowitz, W. L. Atkinson, and A. R. Hinman. 1994.Worldwide measles prevention. Isr. J. Med. Sci. 30:469–481.
31. Patience, C., Y. Takeuchi, and R. A. Weiss. 1997. Infection of human cells byan endogenous retrovirus of pigs. Nat. Med. 3:282–276.
32. Payne, L. N. 1992. Biology of avian retroviruses, p. 299–376. In J. A. Levy(ed.), The Retroviridae. Plenum Press, New York, N.Y.
33. Pyra, H., J. Boni, and J. Schupbach. 1994. Ultrasensitive retrovirus detectionby a reverse transcriptase assay based on product enhancement. Proc. Natl.Acad. Sci. USA 91:1544–1548.
34. Robertson, J. S., C. Nicolson, A.-M. Riley, M. Bentley, G. Dunn, T. Corcoran,
G. C. Schild, and P. Minor. 1997. Assessing the significance of reversetranscriptase activity in chick cell-derived vaccines. Biologicals 25:403–414.
35. Rovigatti, U. G., and S. M. Austrin. 1983. Avian endogenous viral genes.Curr. Top. Microbiol. Immunol. 103:1–21.
36. Shih, A., R. Misra, and M. G. Rush. 1989. Detection of multiple, novelreverse transcriptase coding sequences in human nucleic acids: relation toprimate retroviruses. J. Virol. 63:64–75.
37. Shuldiner, A. R., A. Nirula, and J. Roth. 1990. RNA template-specific poly-merase chain reaction (RS-PCR): a novel strategy to reduce dramaticallyfalse positives. Gene 91:139–142.
38. Tristem, M., P. Kabat, L. Lieberman, S. Linde, A. Karpas, and F. Hill. 1996.Characterization of a novel murine leukemia virus-related subgroup withinmammals. J. Virol. 70:8241–8246.
39. Venugopal, K., L. M. Smith, K. Howes, and L. N. Payne. 1998. Antigenicvariants of J subgroup avian leukosis virus: sequence analysis reveals multi-ple changes in the env gene. J. Gen. Virol. 79:757–766.
40. Vogt, V. M., R. B. Pepinsky, and L. E. Southard. 1985. Primary structure ofp19 species of avian sarcoma and leukemia viruses. J. Virol. 56:31–39.
41. Waters, T. D., P. S. Anderson, Jr., G. W. Beebe, and R. W. Miller. 1972.Yellow fever vaccination, avian leukosis virus, and cancer risk in man. Sci-ence 177:76–77.
42. Weissmahr, R. N., J. Schupbach, and J. Boni. 1997. Reverse transcriptaseactivity in chicken embryo fibroblast culture supernatants is associated withparticles containing endogenous avian retrovirus EAV-0 RNA. J. Virol.71:3005–3012.
43. WHO Expert Committee on Biological Standardization. 1994. Requirementsfor measles, mumps and rubella vaccines and combined vaccines (live).Requirements for biological substances, no. 47. WHO Tech. Rep. Ser. 840:100–207.
44. Wolfgang, S., S. Heyko, K. Frank, R. Anja, H. Rudiger, and L. Christine.1995. Retrovirus-like particles released from the human breast cancer cellline T47-D display type B- and C-related endogenous retroviral sequences.J. Virol. 69:6408–6416.
45. Yamamoto, S., T. M. Folks, and W. Heneine. 1996. Highly sensitive qualita-tive and quantitative detection of reverse transcriptase activity: optimization,validation, and comparative analysis with other detection systems. J. Virol.Methods 61:135–143.
VOL. 73, 1999 ALV AND EAV IN CEF-DERIVED MMR VACCINES 5851
on February 23, 2015 by guest
http://jvi.asm.org/
Dow
nloaded from




















