
Spatial variations in zooplankton diversity in waters contaminated with composite effluents

August 2014⎪Vol. 24⎪No. 8
J. Microbiol. Biotechnol. (2014), 24(8), 1073–1080http://dx.doi.org/10.4014/jmb.1311.11051 Research Article jmbReview
Evaluation of β-1,4-Endoglucanases Produced by Bacilli Isolatedfrom Paper and Pulp Mill Effluents Irrigated SoilSangeeta Pandey, Rameshwar Tiwari, Surender Singh, Lata Nain, and Anil Kumar Saxena*
Division of Microbiology, Indian Agricultural Research Institute, New Delhi - 110012, India
Introduction
Earth’s most abundant renewable organic material, the
lignocellulosic biomass, is the credible source of bioenergy
and commodity chemicals, which can be made economically
viable through identification of cellulase-producing bio-
agents having capability to accelerate the conversion of
cellulose to glucose.
Cellulose is a crystalline structure having tightly packed
bundles of microfibrils, which prevents penetration of
hydrolytic enzymes and its enzymatic conversion to
glucose. Owing to the physical nature of the substrate, the
enzymatic conversion of cellulose to glucose is slow and
economically unviable. The β-1,4 endoglucanase acts
randomly along the cellulose chains. It produces small
cellulose fragments and generates new sites on which
exoglucanases act to produce cellobiose or oligosaccharides.
The synergistic action of the enzymes endo-1,4-glucanase
(E.C. 3.2.1.4), exo-1,4-glucanase (E.C. 3.2.1.91), and 1,4-β-D
glucosidases (E.C. 3.2.1.21) is required for complete
hydrolysis of cellulose [8].
β-1,4-Endoglucanase has many applications in same
industries. It is used to remove color from denim to impart
a good stonewashing effect [11]. In detergents, β-1,4-
endoglucanase is used as an additive for color brightening,
softening, and removal of soil particulate matter [21].
Furthermore, β-1,4-endoglucanase decreases the viscosity
of β-glucan solution by random cleaving of the glycosidic
bond of β-glucan. This process is useful in beer brewing
and improving the quality of animal feed [8]. To date,
numerous cellulases have been isolated and characterized
from bacteria, fungi, and animal species [8].
The microbial origin cellulase genes have been cloned
and the nucleotide sequences of some of the endoglucanse
genes have been identified [25]. The study of cellulolytic
enzymes at the molecular level revealed that some of the
modular features of these enzymes are attributable to their
catalytic activity [17]. The sequence comparison indicated
that the catalytic cores of cellulase belong to a restricted
number of families with significant diversity in their
activities [2]. Cellulases produced by two Bacillus strains
isolated from hot springs of Zimbabwe exhibited 100%
homology with the endoglucanase of Bacillus subtilis and
belonged to GH 5 [9]. The endoglucanase of B. licheniformis
Received: November 14, 2013
Revised: April 8, 2014
Accepted: April 13, 2014
First published online
April 18, 2014
*Corresponding author
Phone: +91-11-25847649;
Fax: +91-11-25846420;
E-mail: [email protected]
upplementary data for this
paper are available on-line only at
http://jmb.or.kr.
pISSN 1017-7825, eISSN 1738-8872
Copyright© 2014 by
The Korean Society for Microbiology
and Biotechnology
A total of 10 cellulase-producing bacteria were isolated from soil samples irrigated with paper
and pulp mill effluents. The sequencing of 16S rRNA gene revealed that all isolates belonged
to different species of genus Bacillus. Among the different isolates, B. subtilis IARI-SP-1
exhibited a high degree of β-1,4-endoglucanase (2.5 IU/ml), β-1,4-exoglucanase (0.8 IU/ml),
and β-glucosidase (0.084 IU/ml) activity, followed by B. amyloliquefaciens IARI-SP-2. CMC was
found to be the best carbon source for production of endo/exoglucanase and β-glucosidase.
The β-1,4-endoglucanase gene was amplified from all isolates and their deduced amino acid
sequences belonged to glycosyl hydrolase family 5. Among the domains of different isolates,
the catalytic domains exhibited the highest homology of 93.7%, whereas the regions of signal,
leader, linker, and carbohydrate-binding domain indicated low homology (73-74%). These
variations in sequence homology are significant and could contribute to the structure and
function of the enzyme.
Keywords: Cellulase, β-1,4-endoglucanase, Bacillus, Paper and pulp mill effluent
S
S

1074 Pandey et al.
J. Microbiol. Biotechnol.
of the same family has also been reported [3].
Soils irrigated with untreated pulp and paper mill
effluent over the years contain high alkalinity and organic
matter. Irrigation with such effluents facilitates growth of
lignocellulose degraders in these soils. Hence, these soils
can be explored for mining of novel microbes and genes
involved in the hydrolysis of cellulose. Studies conducted
in the past have also reported a dominance of culturable
lignocellulose degraders in such habitats [24, 27]. Nonetheless,
the diversity of cellulolytic bacteria and endoglucanase has
not been explored extensively. The aim of this study was to
explore the diversity of cellulolytic bacteria and the allelic
variation in their endoglucanase genes. The sequences of
these genes have been compared with known cellulase
genes and their modular structures have been defined.
Additionally, characterization of the enzyme was carried
out for prediction of its protein based on sequences of genes
amplified from different bacteria.
Materials and Methods
Sample Collection
The Century Pulp and Paper Mill, Lalkuan, Uttarakhand
(79°E10’E longitude and 29°3’N latitude) is one of the largest
paper and pulp mills of India. The untreated effluent of this mill is
utilized for irrigation of nearby agricultural lands. The soil samples
were collected from these agricultural lands and transported
immediately to the laboratory for further processing.
Isolation and Screening of Cellulase-Producing Microorganisms
Carboxymethyl cellulase (CMCase)-producing bacteria were
isolated by using carboxymethyl cellulose (CMC) agar containing
0.5% CMC; 0.1% NaNO3; 0.1% K2HPO4; 0.1%, KCl; 0.05%
MgSO4·7H2O; 0.05% yeast extract; and 1.5% agar, pH 7.0. All
colonies were picked and spot inoculated in Reese’s minimal
medium (RMM) containing 1% CMC to confirm their cellulase
production potential, as described by Pandey et al. [16]. The
substrate specificity of the crude enzyme was determined by
performing the assay with different substrates; Avicel, CMC, and
rice straw. All experiments were conducted in triplicates (n = 3).
Molecular Identification of Bacterial Isolates
For molecular characterization of the isolates, 16S rRNA gene
sequencing was done. Isolation of genomic DNA was carried out
by a standard DNA extraction protocol [20]. The extracted DNA
was used as the template for PCR amplification of the 16S rRNA
gene using universal 16S rRNA gene primers pA (5’-AGAGTT
TGATCCTGGCTCAG-3’) and pH (5’-AAGGAGGTGATCCAG
CCGCA -3’) to obtain a product of approximately 1,500 bp [4].
Aliquots of purified 16S rRNA PCR products were digested
separately with three different restriction endonucleases (AluI,
MspI, and HaeIII) in a 25 µl reaction volume by using the
manufacturer’s recommended buffer and temperature. Restricted
DNA was analyzed on 2% agarose gels. Strong and clear bands
were scored for similarity and clustering analysis using the
NTSYS-2.02e software package, Exeter software (http://www.
exetersoftware.com/). Similarity among the isolates was calculated
by Jaccard’s coefficient and a dendrogram was constructed using
the UPGMA method [12]. The 16S rRNA gene was sequenced by
Xcelris Labs (Ahmedabad, India) using Sanger’s dideoxynucleotide
sequencing method. A similarity search for the sequence was
carried out using the BLAST program of the National Center of
Biotechnology Information (http://www.ncbi.nlm.nih.gov/).
Hydrolytic Enzyme Production on Different Cellulosic Substrates
For enzyme production, cellulase-positive isolates were cultured
in RMM (KH2PO4 - 2 g, (NH4)2SO4 - 1.4 g, KNO3 - 1.4 g, MgSO4·7H2O
- 0.3 g, CaCl2·2H2O - 0.3 g, FeSO4·7H2O - 5 mg, MnSO4·2H2O - 1.6 mg,
ZnSO4·7H2O - 1.4 mg, CoCl2·6H2O - 2 mg, Agar - 20 g, Distilled
water - 1,000 ml) supplemented with CMC (1%), α-cellulose (1%),
Avicel PH101 (1%), Sigmacell 101 (1%), or rice straw (1%) as the
sole carbon source at 30°C and pH 7.0 ± 0.2. After 72h of
incubation, culture broths were centrifuged at 10,000 ×g for
15 min at 4ºC. The supernatant was collected and stored at 4ºC for
further enzyme assays. The substrate specificity of the crude
enzyme was determined by performing the assay with different
substrates; Avicel, CMC, and rice straw. All experiments were
conducted in triplicates (n = 3).
Enzyme Assays
The β-1,4-endoglucanase (CMCase) and β-1,4-exoglucanase
(FPase) activity was determined by using CMC and Whatman no
1 filter paper, respectively, as substrate [5] and measuring the
amount of reducing sugar [12]. One unit (IU) of filter paper
activity or CMCase corresponded to 1 µmole of glucose formed
per minute during hydrolysis. The β-glucosidase activity was
assayed by measuring the amount of p-nitrophenol released from
p-nitrophenyl-β-D-glucopyranoside [26]. One unit (IU) of β-glucosidase
activity was defined as the amount of enzyme releasing 1 µmole
of p-nitrophenol per milliliter of crude enzyme per minute.
Designing of Primers and Amplification of β-1,4-Endoglucanase
Thirty-five sequences of bacterial β-1,4-endoglucanase were
retrieved from NCBI GenBank. The amino acid sequences of these
genes were aligned using T-Coffee alignment (http://www.ebi.ac.uk/
Tools/msa/tcoffee/). The alignment of the Bacillus sequences was
further used in the primer design for amino acid positions 1 to 5
(MKRSI) and 505 to 509 (GTEPN) to obtain a complete coding
region of β-1,4-endoglucanases. The degenerate primers endo-F
(5’-ATGAARMGIWSIATH-3’) and endo-R (5’-RTTIGGYTCIGTNCCC-
3’) amplified approximately 1,500 bp of the β-1,4-endoglucanase
gene. PCR conditions were optimized by gradient PCR by keeping
the reactions at different annealing temperatures ranging from
51°C to 56°C. The final reaction mixture (25 µl) contained each
Evaluation of β-1,4-Endoglucanases Produced by Bacilli 1075
August 2014⎪Vol. 24⎪No. 8
primer at a concentration of 0.5 mM, each dNTP mix at a
concentration of 200 µM, 2.5 U of Taq DNA polymerase, 1.5 mM
MgCl2, 20 ng of template DNA, and 2.5 µl of 10× PCR buffer. The
following thermal profile was used for the PCR: 94ºC, 2 min; 35
cycles of 94ºC, 1 min; 53ºC, 1 min; 72ºC, 2 min; 1 cycle of 72ºC, 10
min. The PCR product was purified using the Qiaquick PCR
purification kit (Qiagen, Valencia, CA, USA) and sequenced by
Xcleris Lab (India).
Sequence and Phylogenetic Analysis
Sequence fragments were assembled with the codon code
aligner program (http://www.codoncode.com/aligner/). The sequenced
gene was compared with available sequences from GenBank using
the BLASTX program (http://blast.ncbi.nlm.-nih.gov/Blast/). The
deduced amino acid sequence was analyzed with the EXPASY tool
(http://expasy.org/). The signal peptide sequence of protein was
predicted by the SignalP 3.0 server (http://www.cbs.dtu.dk/services/
SignalP/) and conserved domain analysis was conducted with
Pfam (http://pfam.wustl.edu/hmmsearch.shtml). The phylogenetic
tree was drawn with Mega 5.0 [22]. In order to statistically
evaluate the confidence of branching, bootstrapping was carried
out with data resampled 1,000 times. Possible open reading frames
(ORFs) were identified with the ORF finder at the NCBI database
(http://www. ncbi.nlm.nih.gov). Closely related protein sequences
in the databases for the candidate cellulases were identified with
BLASTN and BLASTP of NCBI. Module structures of the enzymes
were predicted by the simple modular architecture research tool
(SMART; http://smart.embl-heidelberg.de).
Nucleotide Sequence Accession Numbers
The sequences generated in this study were deposited in NCBI
GenBank. The 16S rRNA gene sequences retrieved from the cellulase-
producing cultures were assigned accession numbers KF204578-
KF204587. The accession numbers of β-1,4-endoglucanase genes
obtained from these cultures were KF240847-KF240856.
Results
Isolation and Screening of Cellulase-Producing Bacteria
A total of 200 bacterial strains, isolated from effluent
irrigated soil, were screened for the presence of cellulase-
producing activity. Congo red test revealed that, out of 200
isolates, 10 isolates (IARI-SP-1 to IARI-SP-10) produced a
clear halo zone around the colony on CMC agar plate and
were taken as positive for cellulase activity.
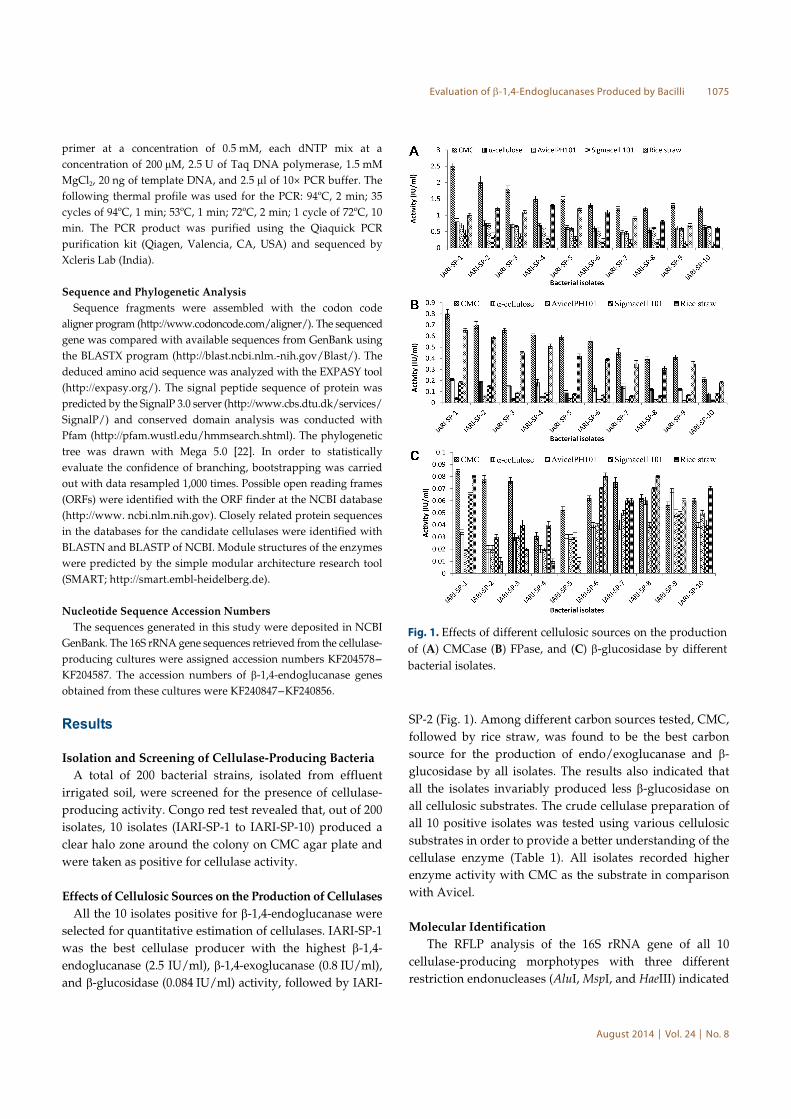
Effects of Cellulosic Sources on the Production of Cellulases
All the 10 isolates positive for β-1,4-endoglucanase were
selected for quantitative estimation of cellulases. IARI-SP-1
was the best cellulase producer with the highest β-1,4-
endoglucanase (2.5 IU/ml), β-1,4-exoglucanase (0.8 IU/ml),
and β-glucosidase (0.084 IU/ml) activity, followed by IARI-
SP-2 (Fig. 1). Among different carbon sources tested, CMC,
followed by rice straw, was found to be the best carbon
source for the production of endo/exoglucanase and β-
glucosidase by all isolates. The results also indicated that
all the isolates invariably produced less β-glucosidase on
all cellulosic substrates. The crude cellulase preparation of
all 10 positive isolates was tested using various cellulosic
substrates in order to provide a better understanding of the
cellulase enzyme (Table 1). All isolates recorded higher
enzyme activity with CMC as the substrate in comparison
with Avicel.
Molecular Identification
The RFLP analysis of the 16S rRNA gene of all 10
cellulase-producing morphotypes with three different
restriction endonucleases (AluI, MspI, and HaeIII) indicated
Fig. 1. Effects of different cellulosic sources on the production
of (A) CMCase (B) FPase, and (C) β-glucosidase by different
bacterial isolates.

1076 Pandey et al.
J. Microbiol. Biotechnol.
variations in the profile. A combined dendrogram constructed
based on similarity percentage among the isolates indicated
that all 10 cellulase-producing isolates belonged to different
clusters (Fig. 2). A partial 16S rRNA gene sequence of about
1,500 bp of all the endoglucanase-positive bacterial isolates
was analyzed by BLAST and the sequences were deposited
in NCBI GenBank. DNA sequencing and phylogenetic
analysis revealed that all positive isolates showed 95-100%
similarity with the known sequences in GenBank and
belonged to different species of genus Bacillus (Fig. 3).
Phylogenetic Analysis of β-1,4-Endoglucanase Gene
The nucleotide sequences of the cloned β-1,4-endoglucanase
genes were compared with those of the cellulase genes
registered in the NCBI database, using the BLASTN
program. The nucleotide sequences of IARI-SP-1 and IARI-
SP-2 indicated 99% and that of IARI-SP-3 showed 98%
similarity with β-1,4-endonuclease of B. subtilis strain BS-
02. The nucleotide sequences of IARI-SP-4, IARI-SP-8, and
IARI-SP-10 indicated 98%, 96%, and 97% similarity,
respectively, with B. subtilis strain shu-3. IARI-SP-5, IARI-
SP-6, IARI-SP-7, and IARI-SP-9 indicated 98%, 98%, 98%,
and 99% identity with B. subtilis strain AH18, B. subtilis
strain DR, Uncultured bacterium clone celWS14, and
B. megaterium strain AP25, respectively (Table 2). Based on
amino acid sequences of the 10 identified ORFs, it was
observed that all sequences are closely related to glycosyl
hydrolase family 5 (GH5). The comparison of deduced
amino acid sequences of the 10 bacterial isolates and
B. subtilis strain BS-02 (GenBank Accession No. JF965375)
reveals a close evolutionary relationship among the isolates
(Fig. 4). The analysis of the deduced amino acid sequences
of the cellulases from our 10 isolates using the pfam
program of Expasy revealed a modular enzyme composed
of two discrete domains in the following order: catalytic
domain (CD) (Q-48 through S-301) of the glucosyl hydrolase
family 5/A2 (endoglucanase, E.C. 3.2.1.4), and carbohydrate-
binding domain (CBD) (V-356 through G-437) of family
IIIa. Similar to the modular organization of many Bacillus
endoglucanases, the CDs of these enzymes were located in
the N-terminal region and the CBDs in the C-terminal
region. There were 7 to 8 amino acid residue substitutions
in the signal peptide region (1 through 29), and 22 amino
acid residues were conserved among all of the cellulases,
resulting in 79% homology. The leader region (30 through
47) consisted of 18 residues, and 16 amino acid residues
Table 1. Substrate specificity of the cellulases from different
Bacillus strains towards different substrates.
Bacterial
isolates
β-1,4-Endoglucanase (IU/ml)
CMC Avicel Rice straw
IARI-SP-1 9.5 ± 1.0 2.6 ± 0.9 7.5 ± 0.9
IARI-SP-2 8.0 ± 0.9 2.3 ± 0.8 6.8 ± 0.6
IARI-SP-3 4.5 ± 0.6 1.8 ± 0.7 4.5 ± 0.6
IARI-SP-4 6.8 ± 0.7 1.5 ± 0.6 5.3 ± 0.5
IARI-SP-5 6.3 ± 0.6 1.2 ± 0.8 4.5 ± 0.7
IARI-SP-6 5.5 ± 0.6 1.4 ± 0.6 4.0 ± 0.5
IARI-SP-7 4.3 ± 0.5 1.0 ± 0.7 2.8 ± 0.6
IARI-SP-8 4.5 ± 0.6 0.9 ± 0.8 2.5 ± 0.7
IARI-SP-9 2.5 ± 0.4 0.8 ± 0.7 1.5 ± 0.6
IARI-SP-10 1.1 ± 0.5 0.6 ± 0.8 0.9 ± 0.6
The results are presented as the mean ± SD (standard deviation), n = 3.
Fig. 2. Dendrogram showing clustering of 10 isolates generated from RFLP analysis of the 16S rRNA gene by three restriction
endonucleases (AluI, MspI, and HaeIII), using the UPGMA algorithim and Jaccard’s coefficient.

Evaluation of β-1,4-Endoglucanases Produced by Bacilli 1077
August 2014⎪Vol. 24⎪No. 8
among them were conserved, resulting in 89% homology.
The CD consisted of 254 residues (48 through 301), and 238
amino acid residues were conserved among the cellulases,
resulting in 93.7% homology. In the linker region (302
through 355) consisting of 54 residues, 40 amino acid
residues were conserved among the cellulases, resulting in
74% homology. In the CBD (356 through 437) consisting of
83 residues, 57 amino acid residues were conserved among
the cellulases, resulting in 68.6% homology. The CD
showed the highest 93.7% homology, whereas the regions
of signal, leader, linker, and CBD regions showed low
homology. The β-1,4-endoglucanase gene encoded a signal
peptide at the N-terminal end of the protein. The active site
131-H, 169-E, and 257-E are conserved among all isolates
except IARI-SP-4, in which Q is present at position 257.
IARI-SP-1 and IARI-SP-2, being the highest cellulase producers
found in this study, were found to be 100% homologous for
the β-1,4-endoglucanase gene.
Fig. 3. Phylogenetic tree of culturable bacteria based on the 16S rRNA gene sequence isolated from soil irrigated with effluents of
century paper mill.
Table 2. Phylogenetic affiliations of the β-1,4-endoglucanase gene, isolated from different Bacillus sp., with their nearest neighbor.
Bacterial Isolate Accession number Nearest neighbor of β-1,4-endoglucanase gene Homology (% )
B. subtilis IARI-SP-1 KF240847 B. subtilis strain BS-02 99
B. amyloliquefaciens IARI-SP-2 KF240848 B. subtilis strain BS-02 99
B. licheniformis IARI-SP-3 KF240849 B. subtilis strain BS-02 98
Bacillus sp. IARI-SP-4 KF240850 B. subtilis strain shu-3 98
B. pumilus IARI-SP-5 KF240851 B. subtilis strain AH18 98
B. thuringiensis IARI-SP-6 KF240852 B. subtilis strain DR 98
B. cereus IARI-SP-7 KF240853 Uncultured bacterium clone celWS14 98
B. mycoides IARI-SP-8 KF240854 B. subtilis strain shu-3 96
B. megaterium IARI-SP-9 KF240855 B. megaterium strain AP25 99
B. oceanisediminis IARI-SP-10 KF240856 B. subtilis strain shu-3 97

1078 Pandey et al.
J. Microbiol. Biotechnol.
Discussion
Soils perpetually irrigated with paper and pulp effluent
represents a unique niche with a predominance of cellulose-
degrading microorganisms, owing to their high organic
matter content. The predominance of Bacillus and Bacillus-
derived genera as dominant cellulase producers in the
present study confirms the earlier reports from such
environments [6, 18]. Genus Bacillus, being endospore
formers, are tolerant to the harsh conditions (high pH)
prevailing in such soils, which makes them the major
microflora of such environments [15].
Enzyme assays indicated that all isolates invariably
produced less β-glucosidase on different cellulosic substrates
tested. However, all isolates had greater catalytic activity
for CMC in comparison with other cellulosic substrates,
which revealed that enzymes produced by all isolates
could be categorized as endoglucanase with significant
Fig. 4. Alignment of deduced amino acid sequences of endoglucanases of the 10 isolates with B. subtilis strain shu-3.
Region 1-29, signal peptide; Region 48-301, cellulase catalytic domain (glucosyl hydrolase family 5/A2); Region 302-355, linker region; Region
356-437, cellulose-binding domain (CBD family IIIa); Sites 131-H, 169-E, and 257-E, active sites (proton donor or nucleophile).

Evaluation of β-1,4-Endoglucanases Produced by Bacilli 1079
August 2014⎪Vol. 24⎪No. 8
amount of exoglucanase activity. Many reports suggest
that the cellulase systems from bacilli are incomplete and
could not act on microcrystalline cellulose [1], but others
do confirm the exoglucanase activity in certain bacilli [9].
The exoglucanase activity of the crude enzymes might be
due to the presence of a CBD in the β-1,4-endoglucanase.
Earlier studies have indicated that the CBD plays crucial
roles in crystalline cellulose hydrolysis, by binding to the
cellulose surface and enhancing the sacchrification ability
of endoglucanses [14, 23]. In the present study, rice straw
was found to be a good carbon source for production of
cellulase and hence can be evaluated as a substrate for the
large-scale production of cellulase, replacing costly substrates
like CMC. There are several studies on cellulose degradation
by Bacillus sp., and large variations in enzyme activity
under both optimized and non-optimized conditions have
been reported [3, 7, 9, 18]. For example, Li et al. [7] have
reported that in a Bacillus sp., maximum cellulase activity
(0.26 U/ml) was detected when the culture was grown in
Luria broth supplemented with 1% CMC at 37°C for 24 h.
Our data on the β-1,4-exoglucanase, β-1,4-endoglucanase,
and β-glucosidase activities of all 10 isolates were generated
from crude culture supernatants, but were encouraging
as all 10 bacterial isolates exhibited considerably higher
endoglucanase activities (ranging from 1.1 to 2.5 IU/ml)
than earlier reported [18, 19, 28]. The maximum CMCase
activity in the alkaline range may be due to the alkaline
condition prevailing in the soil. The nucleotide sequencing
and BLAST search of β-1,4-endoglucanase revealed limited
variations, indicating a close evolutionary relationship
among the isolates. The high proportion of cellulase enzyme
belonging to the GH5 family among bacilli found in our
study is in agreement with earlier reports [3, 9]. The CDs of
these enzymes were located in the N-terminal region and
the CBDs in the C-terminal region, like for endoglucanases
of many other Bacillus species earlier reported. Currently, the
CDs of polysaccharides are grouped into at least 15 of the
more than 80 known glycosyl hydrolase families, whereas
CBDs fall into at least 13 families. Two basic residues at
positions 2 and 3 (lysine and arginine) in the hydrophilic
leader region were followed by a hydrophobic core of 18
amino acid residues rich in leucine and isoleucine, which is
consistent with the findings of Mezes et al. [10]. The β-1,4-
endoglucanase gene encoded a signal peptide at the N-
terminal end of the protein and was characterized by a
short, hydrophilic, basic region along with a subsequent
long, hydrophobic region. The presence of a signal peptide
suggests higher extracellular cellulase activity observed for
various strains screened during the present study.
The soils irrigated with pulp and paper mill effluent
specifically enrich cellulolytic bacilli that exhibit both endo-
and exoglucanase activities. The analysis of the β-1,4-
endoglucanase gene isolated from Bacillus sp. provides a
new insight into the potential capacity of the microbial
community to hydrolyze the lignocellulosic biomass.
Although all sequences found in the present study belonged
to the GH5 family, sequence analysis revealed significant
variations among them. These variations need to be further
exploited to correlate the efficiency of the enzyme, and its
ability to hydrolyze crystalline cellulose and also to be
secreted out of the cell in large amounts. Research is under
way to clone and overexpress the endoglucanase gene in a
suitable expression vector so that the product could be
utilized in the biofuel program being carried out at the
Institute.
Acknowledgments
The authors acknowledge funding from NAIP (70-24)
and ICAR National Fund for Basic, Strategic and Frontier
Application Research in Agriculture and sponsoring the
research project. The first author thanks the Indian Agricultural
Research Institute for providing fellowship during the
doctoral program.
References
1. Aa K, Flengsrud R, Lindahl V, Tronsmo A. 1994. Characterization
of production and enzyme properties of an endo-β-1,4-
glucanase from Bacillus subtilis CK-2 isolated from compost
soil. Antonie Van Leeuwenhoek 66: 319-326.
2. Béguin P, Aubert J-P. 1994. The biological degradation of
cellulose. FEMS Microbiol. Rev. 13: 25-58.
3. Bischoff KM, Rooney AP, Li X-L, Liu S, Hughes SR. 2006.
Purification and characterization of a family 5 endoglucanase
from a moderately thermophilic strain of Bacillus licheniformis.
Biotechnol. Lett. 28: 1761-1765.
4. Edwards U, Rogall T, Blöcker H, Emde M, Böttger EC. 1989.
Isolation and direct complete nucleotide determination of
entire genes. Characterization of a gene coding for 16S
ribosomal RNA. Nucleic Acids Res. 17: 7843-7853.
5. Ghose TK. 1987. Measurements of cellulase activities. Pure
Appl. Chem. 59: 257-268.
6. Kim Y-K, Oh H-J, Ko YH. 2012. Comparison of nucleotide
sequences of endo-B-1,4-glucanase genes from Bacillus
subtilis strains. Int. J. Biotechnol. Appl. 4: 130-133.
7. Li W, Zhang W-W, Yang M-M, Chen Y-L. 2008. Cloning of
the thermostable cellulase gene from newly isolated Bacillus
subtilis and its expression in Escherichia coli. Mol. Biotechnol.
40: 195-201.

1080 Pandey et al.
J. Microbiol. Biotechnol.
8. Lynd LR, Weimer PJ, Van Zyl WH, Pretorius IS. 2002.
Microbial cellulose utilization: fundamentals and biotechnology.
Microbiol. Mol. Biol. Rev. 66: 506-577.
9. Mawadza C, Hatti-Kaul R, Zvauya R, Mattiasson B. 2000.
Purification and characterization of cellulases produced by
two Bacillus strains. J. Biotechnol. 83: 177.
10. Mézes P, Blacher R, Lampen JO. 1985. Processing of Bacillus
cereus 569/H beta-lactamase I in Escherichia coli and Bacillus
subtilis. J. Biol. Chem. 260: 1218-1223.
11. Miettinen-Oinonen A, Suominen P. 2002. Enhanced
production of Trichoderma reesei endoglucanases and use of
the new cellulase preparations in producing the stonewashed
effect on denim fabric. Appl. Environ. Microbiol. 68: 3956-
3964.
12. Miller GL. 1959. Use of dinitrosalicylic acid reagent for
determination of reducing sugar. Anal. Chem. 31: 426-428.
13. Nei M, Li W. 1979. Mathematical model for studying
genetic variation in terms of restriction endonucleases. Proc.
Natl. Acad. Sci. USA 76: 5269.
14. Nguyen NH, Maruset L, Uengwetwanit T, Mhuantong W,
Harnpicharnchai P, Champreda V, et al. 2012. Identification
and characterization of a cellulase-encoding gene from the
buffalo rumen metagenomic library. Biosci. Biotechnol. Biochem.
76: 1075-1084.
15. Nicholson WL, Munakata N, Horneck G, Melosh HJ, Setlow
P. 2000. Resistance of Bacillus endospores to extreme terrestrial
and extraterrestrial environments. Microbiol. Mol. Biol. Rev.
64: 548-572.
16. Pandey S, Singh S, Yadav AN, Nain L, Saxena AK. 2013.
Phylogenetic diversity and characterization of novel and
efficient cellulase producing bacterial isolates from various
extreme environments. Biosci. Biotechnol. Biochem. 77: 1474-
1480.
17. Rabinovich M, Melnick M, Bolobova A. 2002. The structure
and mechanism of action of cellulolytic enzymes. Biochemistry
(Moscow) 67: 850-871.
18. Rastogi G, Bhalla A, Adhikari A, Bischoff KM, Hughes SR,
Christopher LP, Sani RK. 2010. Characterization of
thermostable cellulases produced by Bacillus and Geobacillus
strains. Bioresour. Technol. 101: 8798-8806.
19. Sadhu S, Saha P, Sen SK, Mayilraj S, Maiti TK. 2013.
Production, purification and characterization of a novel
thermotolerant endoglucanase (CMCase) from Bacillus strain
isolated from cow dung. SpringerPlus 2: 1-10.
20. Sambrook J, Russell DW. 2001. Molecular Cloning: A
Laboratory Manual. Cold Spring Harbor Laboratory Press,
Cold Spring Harbor, NY.
21. Shimonaka A, Koga J, Baba Y, Nishimura T, Murashima K,
Kubota H, and Kono T. 2006. Specific characteristics of family
45 endoglucanases from Mucorales in the use of textiles and
laundry. Biosci. Biotechnol. Biochem. 70: 1013-1016.
22. Tamura K, Peterson D, Peterson N, Stecher G, Nei M,
Kumar S. 2011. MEGA5: molecular evolutionary genetics
analysis using maximum likelihood, evolutionary distance,
and maximum parsimony methods. Mol. Biol. Evol. 28: 2731-
2739.
23. Tang Z-Z, Wu Z-F, Chen H, Lai X, Han X-Y, Wu Q. 2013.
Characterization of novel EGs reconstructed from Bacillus
subtilis endoglucanase. Appl. Biochem. Biotechol. 169: 1764-
1773.
24. Tripathi BM, Kaushik R, Kumari P, Saxena AK, Arora DK.
2011. Genetic and metabolic diversity of streptomycetes in
pulp and paper mill effluent treated crop fields. World J.
Microbiol. Biotechnol. 27: 1603-1613.
25. Wang F, Li F, Chen G, Liu W. 2009. Isolation and
characterization of novel cellulase genes from uncultured
microorganisms in different environmental niches. Microbiol.
Res. 164: 650-657.
26. Wood TM, Bhat KM. 1988. Methods for measuring cellulase
activities. Methods Enzymol. 160: 87-112.
27. Yadav S, Kaushik R, Saxena AK, Arora DK. 2011. Diversity
and phylogeny of plant growth-promoting bacilli from
moderately acidic soil. J. Basic Microbiol. 51: 98-106.
28. Yang D, Weng H, Wang M, Xu W, Li Y, Yang H. 2010.
Cloning and expression of a novel thermostable cellulase
from newly isolated Bacillus subtilis strain I15. Mol. Biol. Rep.
37: 1923-1929.
Copyright © 2022 FDOKUMEN




















