
DNA methylation in white blood cells: Association with risk factors in epidemiologic studies
Upload
independentCategory
view
3download
0
1
Epidemiologic and molecular characterization of human immunodeficiency virus
type 1 in Southern Brazil
Esmeralda A.J.M. Soares1, Rodrigo P. Santos2, José Augusto Pellegrini2, Eduardo
Sprinz2, Amilcar Tanuri1 and Marcelo A. Soares1*
aLaboratório de Virologia Molecular
Departamento de Genética Universidade Federal do Rio de Janeiro
CCS – Bloco A – Cidade Universitária – Ilha do Fundão 21944-970 Rio de Janeiro, RJ, Brazil
Telephone: 55-21-2562-6384 Fax: 55-21-2562-6397
E-mail: [email protected]
bHospital de Clínicas de Porto Alegre Universidade Federal do Rio Grande do Sul
Rua Ramiro Barcellos, 2350 - Bairro Rio Branco 90035-003 Porto Alegre, RS, Brazil
* To whom correspondence should be addressed
Work supported by the AIDS/STD National Program, Brazilian Ministry of Health, the
State Science Foundation of Rio de Janeiro Grant E-26/151.970/00, the Brazilian Council
for Scientific and Technologic Development Grant 462394/00-0.

2
ABSTRACT
Human immunodeficiency virus subtype C is the most prevalent subtype in the
world. Despite its recent expansion in Brazil, HIV-1C already prevails in the
southernmost state of Brazil, Rio Grande do Sul. This unique HIV epidemiology has
prompted us to characterize that population. Seventy-seven HIV-1-infected subjects
attending the largest HIV/AIDS clinic of the state had their viruses’ protease and reverse
transcriptase (RT) genes subtyped and genotyped. When subtype-specific infections were
plotted according to year of diagnosis, the prevalence of subtype C was shown to increase
over the last 18 years of the epidemic along with a concomitant decrease of subtype B.
Comparison of subtype C-infected treated and untreated subjects revealed amino acid
differences in protease and RT, in special the RT mutation D/G123S. The overall analysis
of drug resistance mutations in viruses from treated subjects has highlighted some
associations between subtypes and particular mutations, such as V82A/F/T/S in protease
and subtype F1, and M41L and L210W in RT and subtype B. The characterization of this
important population, which is one of a few in the developing world where a large
number of HIV-1C-infected subjects are under antiretroviral treatment, underscores its
potential usefulness in clinical, treatment and vaccine trials in Brazil.
Keywords: HIV-1, subtype C, AIDS epidemiology, drug resistance, Brazil

3
INTRODUCTION
Human immunodeficiency virus type 1 (HIV-1) subtype C is currently the most
prevalent subtype in the world, accounting for over half of the infected individuals
worldwide (1). It now dominates several areas of the world, such as southern Africa,
Israel, India and China, including countries previously dominated by other subtypes (2,3).
It is also increasing in incidence in some developed nations (4-7).
The dynamic nature of subtype C epidemic in the world highlights its importance
for HIV vaccine development (8,9). Subtype-specific immune responses have been
reported (10,11), and differences in the elicitation of responses by protective vaccination
might be important towards adequate vaccine strategies in different regions. Moreover,
differences in the kinetic parameters of non-B HIV-1 protease enzymes due to
polymorphic signatures of these viruses (12,13) may impact the long-term treatment of
non-B HIV-1 infection worldwide (14).
Differently from well-established countries where HIV-1 subtype C prevails,
there are regions where this variant is responsible for local endemic infections. Brazil can
be included in this profile. The prevailing subtype in the country is B, and subtypes F1
and C have been previously described on an isolated basis (15-17). However, a recent
nationwide analysis conducted by our group has shown that a large proportion of HIV-1
infections in southern Brazil, including the states of Rio Grande do Sul (RS) and Parana,
is represented by subtype C (prevalences of 45% and 30%, respectively; 18). That
subtype has also increased in Rio de Janeiro and Sao Paulo when compared to previous
surveys (19,20), suggesting a northward increase of subtype C in the country. In a more

4
detailed analysis of those variants, we have shown that the introduction of HIV-1 subtype
C in Brazil was likely to represent a single event (21).
To characterize the HIV-1-infected population from southern Brazil, which
accounts for over 20% of the country’s infections, we have surveilled the city of Porto
Alegre, capital of RS. This city has the highest reported rate of subtype C infections in
the country, and it is the city where subtype C sequences was initially described (17). It is
the main site in Brazil, and perhaps one of a few in the world, where a large number of
HIV-1 subtype C-infected individuals are treated at large. We describe here HIV-1
subtype prevalences and drug resistance mutation profiles from a major clinical site in
Porto Alegre. Hospital de Clínicas of Porto Alegre has the largest HIV/AIDS cohort and
provides care for HIV-infected people in that city as well as adjacent municipalities.
METHODS
Population and sample collection
Seventy-seven individuals regularly followed at the HIV/AIDS ambulatory care
unit in Hospital de Clinicas de Porto Alegre with confirmed HIV-1-positive serological
status were included in this study. Both treated and drug-naïve individuals for whom
extensive information, such as the date of diagnosis, HIV viral load and CD4 counts over
the period of months to years, and all histories of clinical manifestations and treatment
were included. Five of the patients had incomplete information and were discarded from
population-related analyses, but included in HIV-1 subtype prevalence. Subjects signed a
written consent for their inclusion in the study, and blood draw was part of their regular
follow up visits. The study was approved by the Brazilian IRB (project no. 526 –

5
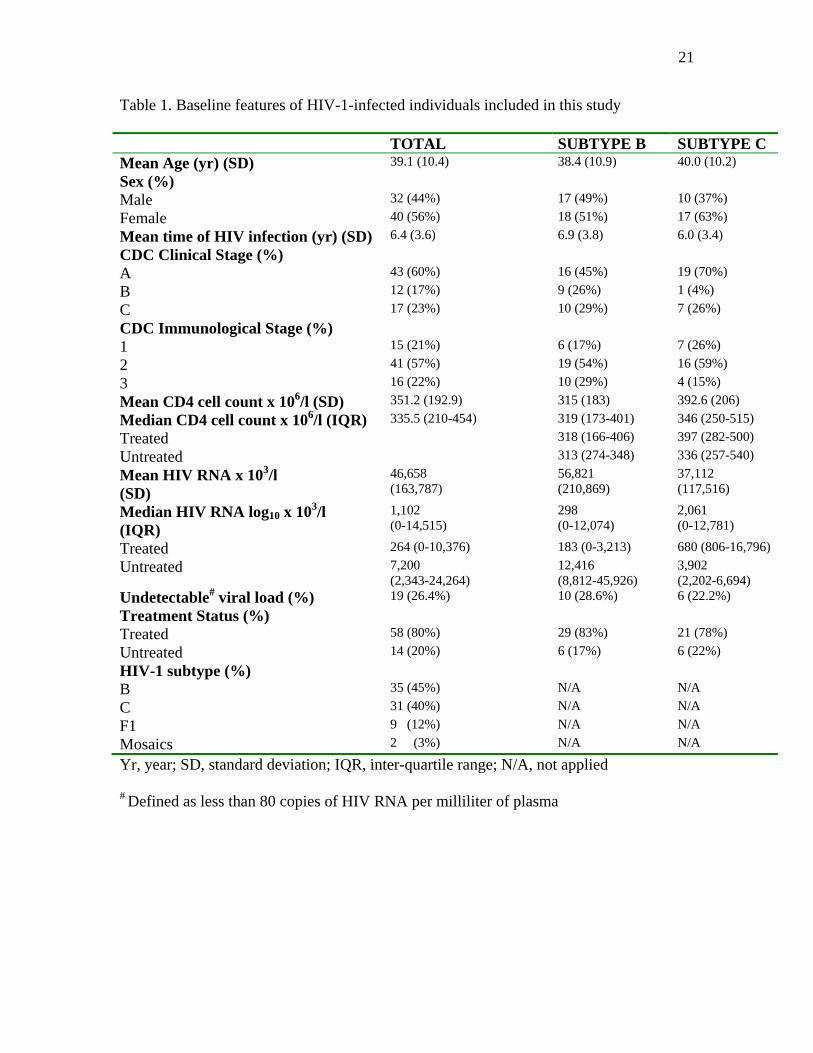
CONEP). Table 1 summarizes relevant baseline features and clinical data of the
individuals analyzed. All collections were done between July and September 2002, and
their plasma separated, aliquoted and stored at -80oC until processing.
RNA isolation, PCR and sequencing
Viral RNA was isolated as previously described (22). Following cDNA synthesis
using randon primers, nested PCR were conducted for the amplification of an HIV-1 pol
gene fragment spanning the entire protease (PR) coding region and the first 225 codons
of the reverse transcriptase (RT) region. In cases where no contiguous fragment was
obtained, separated PCR reactions were conducted for PR and RT. Primers and PCR
conditions used in the reactions have been previously described (22). PCR products were
purified in QIAGEN columns (QIAGEN, Inc., Valencia, CA) and sequenced in an ABI
3100 Genetic Analyzer (Applied Biosystems, Foster City, CA). Sequences were
assembled in PC/Windows computers using the software SeqMan (DNAStar, Inc.,
Madison, WI). All sequences obtained were subject to quality control procedures (23) to
ensure lack of sample mix-ups or contaminations. Sequences were reported to the
GenBank database under accession numbers AY275717 to AY275807.
Phylogenetic and Drug Resistance Mutation Analyses
Sequences were subjected to phylogenetic analysis for HIV-1 subtype
determination. Sequences in FASTA format (24) were aligned with an HIV-1 subtype
reference set from the Los Alamos database (http://hiv-web.lanl.gov) in ClustalW (25)
and manually edited for codon alignment in the Genetic Data Environment for Linux
(26). Phylogeny was performed using the neighbor-joining method with the F84 model of
substitution in PAUP* 4.0b2a (27). The HIV-1 group O sequence MVP5180 (28) was

6
used as outgroup. Sequences from which a contiguous PR/RT genomic region was
available for subtyping, but with unclear classification had the PR and RT regions further
uncoupled and separately analyzed. Sequences with discordant subtype assignment in PR
and RT regions were considered mosaics.
Subtype C PR and RT sequences were grouped according to treatment status
(treated or untreated) and subjected to VESPA analysis for consensi generation. Consensi
were then aligned in ClustalW to evidence potential differences in both sequence groups.
Primary and secondary drug resistance mutations (DRM) described for PR and
RT were analyzed using the Stanford Sequence Database (29, http://hivdb.stanford.edu).
Mutations were compiled according to the International AIDS Society USA consensus
(30).
Statistical Analyses
Differences in continuous characteristics of subtypes B and C-infected sub-
populations (HIV RNA viral load, CD4 T-cell counts, and mean ages and times of
infection) were evaluated by Kruskal-Willis ANOVA tests. Associations between HIV-1
subtypes and gender or treatment status, as well as the significance of differences in drug
resistance mutation frequencies among different HIV-1 subtypes, were evaluated by
Fisher’s exact tests. All statistical computations were performed in the package Analyze-
it® version 1.65 under Microsoft Excel for Windows XP.
RESULTS
Epidemiological Profile of the Population

7
Major baseline characteristics of the 77 individuals analyzed from Hospital das
Clinicas can be seen in Table 1. Overall values, as well as B and C subtype-specific
infected groups are shown in the second and third columns, respectively. Fifty-six percent
of the individuals were female (male to female ratio of 0.8). Mean age was 39.1 years
(SD 10.4). Median CD4 cell count was 335.5 x 106 cells/l, whereas median HIV-RNA
level was 1,101 copies/ml. A large range of viral load was observed, from undetectable
(<80 copies/ml) to over 106 copies/ml. Most of the patients (60%) were at CDC clinical
stage A at the time of sampling, and a similar number was at CDC immunological stage
2. Eighty percent of the patients were drug-experienced. All treated patients had been
subjected to HAART, with exception of one patient under mono- and one patient under
dual therapy. Of the remaining 54 patients, 7 had only experienced NNRTI-containing
regimens in addition to NRTI, 23 had IP-containing regimens, and 24 had experienced
both classes of drugs. The average number of drugs taken by patient among the treated
subjects was 5.5, whereas the average duration of therapy was 41.8 months (range 1-
109). There were no significant associations between CD4 cell counts, viral load, mean
age or time of infection from diagnosis to collection date and infecting subtype (p-values
of 0.5716, 0.1118, 0.4684 and 0.5320, respectively). Furthermore, no association between
gender or treatment status and infecting subtype was found (p-values of 0.5168 and
0.2222, respectively). However, subtype B-infected subjects seemed to have longer drug
exposure than subtype C-infected counterparts (46.6 versus 34.8 months, respectively).
PCR products from all patients were obtained (PR and RT, PR alone or RT
alone). Fifty-eight patients (75%) had both PR and RT sequenced, whereas for 6 (8%)
patients only PR was sequenced, and for 13 (17%) patients only RT was obtained.

8
Phylogenetic analysis of these fragments allowed HIV-1 subtype assignment for all
isolates (Table 1). A large number of subtype C viruses (40%) were observed, almost as
much as subtype B viruses (45%). Subtype F1 represented 12% of the samples. Two
mosaic viruses (with discordant phylogenies for PR and RT) was observed, accounting
for only 3% of the samples. These mosaic forms were not representative of the two most
prevalent subtypes in that area (B and C), but were rather similar to intersubtypic mosaics
previously characterized in Brazil, namely B/F and B/D (31,32).
HIV-1 Subtypes across the AIDS epidemic in Porto Alegre
Previous studies have suggested an increasing incidence of subtype C in the South
of Brazil (18,21), but we were able to analyze subtype prevalence over time in the city of
Porto Alegre. Samples were stratified by year of diagnosis and by infecting subtype (B,
C, F1 and mosaics), and then grouped in 5-year periods (1986-1990, 1991-1995, 1996-
2000, and 2001-2002). Figure 1 depicts that analysis. Our first subtype C virus appeared
as early as 1990, and to our knowledge this is the oldest subtype C infection described in
Brazil. Prior to that year, only subtype B-related viruses were detected. By 1991, the first
subtype F1 virus appeared in our dataset, again confirming the idea of a more recent
expansion of non-B subtypes in Brazil. A clear trend in the increase of subtype C
prevalence over the epidemic was observed: none prior to 1990, 39% from 1991 to 1995,
38% from 1996 to 2000, and 43% from 2001 and 2002. Direct comparisons between any
two consecutive time periods did not show statistical significance (data not shown).
Subtype C-Specific Mutational Analysis
To evaluate the impact of ARV treatment on PR and RT mutations in subtype C
viruses, consensi sequences from treated and untreated subjects were generated and

9
compared. In the PR region, only codon 15 differed between consensi (an isoleucine
(60%) among untreated sequences and a valine (84%) among treated sequences). In RT,
4 codons differed: 123 (an Asp/Gly (50%) in untreated and a Ser (54%) in treated); 135
(a Thr (75%) in untreated and an Ile (58%) in treated); 173 (an Ala (50%) in untreated
and a Lys (50%) in treated); and 177 (a Glu (50%) in untreated and an Asp (54%) in
treated). Of those differences, only the presence of a Ser at position 123 of RT from
treated subjects was statistically significant (p=0.021).
Drug Resistance Mutation Profile of the Population
The availability of treated subtype C-infected individuals in our study prompted
us to assess the prevalence of primary drug resistance mutations (DRM) in those
sequences. Patients already initiated on ARV treatment were subjected to DRM analysis.
Primary DRM associated with protease inhibitors (PI) and all mutations associated with
RT inhibitors (RTI) present in the International AIDS Society consensus (30) were
compiled. Although PI-associated secondary DRM are also important and contribute to
drug resistance, the high incidence of subtype C samples in this population would
artificially bring those numbers to high values, as several of these mutations are natural
polymorphisms in that subtype (21). For that reason, they were not analyzed in this study.
Tables 2 and 3 summarize all primary PR and RT DRM identified in each subtype group.
Among 58 treated patients, 25 (43.1%) of their viral isolates showed DRM to at least one
of the three classes of inhibitors (PI, NRTI or NNRTI). Six isolates (10.3%) had DRM for
2 different ARV classes, whereas 5 (8.6%) had DRM for all three classes. Among 43
isolates for which the PR sequence was available, 11 (25.6%) had one or more primary
PI-associated DRM. Twenty-one (44.7%) of those for which RT sequence was available

10
(n=47) had at least one NNRTI-associated mutation, whereas 9 (19.1%) had one or more
NNRTI-associated mutations.
The most prevalent PI-associated DRM was V82A/F/T/S (20.9%), followed by
M46I/L (11.6%) and L90M (9.3%). Mutations G48V, I50V and I84V were each
represented in one isolate. The mutation D30N was not found in any isolate analyzed in
this study, despite the use of the PI nelfinavir by 33.9% of the treated patients. Among
the NRTI mutations, the most prevalent was M184V (21.3%), followed by M41L and
T215Y/F (14.9%), V75I and K219Q/E (12.8%) and L210W (8.5%). Other mutations
appeared in lower frequencies, while mutations F77L and Y115F were not present in any
isolate. Finally, the main NNRTI-associated mutations found were K103N (14.9%),
L100I (8.5%) and V106A (4.2%). Mutations V108I and Y188C/H/L were not found
among our isolates. All observed mutations were compatible with their patient’s drug
treatment histories, with exception of three individuals (all subtype B-infected); one had
L74V but no use of ddI, ddC or ABC; one had T69D but no use of ddC; and one had
G190S but no use of NNRTI. When average numbers of DRM per genomic fragment
were calculated for each subtype, some differences were observed. In the RT, subtype B
(n=24) had a total of 54 DRM, and an average of 2.24 mutations per isolate, whereas
subtype C sequences had an average of 0.80 mutations per isolate (13 mutations in 17
isolates). Similarly, subtype B sequences had an average of 0.52 mutations per isolate in
the PR region, as compared to 0.07 for subtype C.
Fisher’s exact test was applied to evaluate the significance of selected DRM
comparing any two subtypes and/or B versus non-B subtypes. Of note, mutation
V82A/F/T/S of PR was significantly more frequent in subtype F1 than in subtype B

11
(p=0.0209). Similarly, mutation M41L of RT was significantly more frequent in subtype
B than in subtype C viruses (p=0.0308). This mutation was also more significantly found
in B subtype when compared to all non-B counterparts (p=0.011). We were not able to
find statistical significance in the difference between the occurrence of mutation L210W
in B and non-B subtypes, although a trend was observed towards a higher frequency in
subtype B (p=0.0554). No viruses isolated from drug-naïve individuals from our study had any primary
PI or RTI-associated mutations.
DISCUSSION
HIV/AIDS epidemic in Brazil is complex compared to other Latin American
countries regarding subtype distribution and prevalences. Early studies have shown a
predominance of subtype B, followed by subtype F1 and only sporadic instances of other
subtypes, such as C, D and A (15-17). More recently, however, we have described a
complex geographical pattern of HIV-1 subtype distribution in Brazil (18). We found that
subtype C is highly prevalent in the south of Brazil, ranging from 25% to 45% (18). In
the southeastern region, smaller frequencies of subtype C are found (5 to 10%), whereas
in the rest of the country it is virtually absent. In view of this complex distribution, and of
the importance of subtype C in the worldwide HIV/AIDS epidemic, we characterized the
HIV-infected population from the south of Brazil. We have chosen the largest AIDS
service in Porto Alegre, the capital of the state previously characterized with highest
proportion of subtype C in Brazil.

12
We found a high frequency of subtype C-harboring individuals (40%; Table 1), in
agreement with Brindeiro et al. (18). The slightly higher prevalence of 45% reported by
Brindeiro et al. is probably due to the fact that their analysis has been done on recently-
diagnosed (and likely recently-infected) individuals in 2001-2, and our current analysis is
based on total prevalence over 18 years (from 1986 to 2002). These data add up to the
growing body of evidence suggesting that subtype C is increasing in prevalence since its
introduction in the south of Brazil. Previous data on AIDS patients survival rates in the
city of Rio Grande, another city in RS state, have shown a prevalence of 22% for subtype
C viruses (33). When we stratify our dataset by year of diagnosis, we see a clear trend
towards a decrease of subtype B prevalence over time, accompanied by an increase of
non-B subtypes, in particular of subtype C, as well as of intersubtypic mosaics (Figure 1).
Although our data did not reach statistical significance, we are currently analyzing more
isolates, and soon we will be able to more rigorously test this hypothesis. All these data
together strongly suggest a shift in the HIV-1 subtype epidemiology over time in the
AIDS epidemic in RS. Noteworthy, we have also reported in this study the oldest isolate
of HIV-1 subtype C in Brazil, from an individual diagnosed in 1990.
A very small amount of mosaics (2/77, 3%) was present in our dataset, when
compared to what Brindeiro et al. (18) have described for RS in 2001-2 (25%). Despite
the similar subtype prevalence, the only two mosaics observed in our study were a B/F1
and a B/D virus, also differently from Brindeiro et al., which described most of the
mosaics being B/C recombinants. Although we cannot rule out the possibility that our
“pure” viruses are in fact mosaics with discordant subtypes in other genomic regions
(e.g., gag or env), the pol regions analyzed were the same as the ones studies by

13
Brindeiro et al. (18). This contrast argues in favor of a very recent start of intersubtypic
recombination events, with most of the B/C mosaic viruses being formed in the last few
years, as it has been previously suggested (21).
The universal free access to ARV drugs in Brazil has created a particular situation
in the south of the country where large numbers of subtype C-harboring individuals are
under treatment. We were able to compared subtype C sequences from treated and
untreated subjects. Of significance, we found that 54% of our subtype C sequences from
treated subjects had a serine residue at codon 123 of RT, whereas that residue was not
found in drug-naïve sequences of that subtype. A more careful analysis of ARV histories
from viruses with that substitution revealed that all had taken ZDV and/or 3TC. An
analysis of 30 subtype C-infected patients undergoing NRTI-based therapy from Stanford
database has also shown that 43% of them, but only 20% of drug-naïve isolates, harbored
that mutation. Those differences were also statistically significant (p=0.006). The
analysis of over 1,300 NRTI-treated and 665 untreated subtype B sequences from the
Stanford database revealed that both groups had very low and also similar (1-2%)
prevalences of 123S. It is possible that this mutation is only selected in the context of a
subtype C background under treatment, but a more extensive analysis with in vitro
phenotyping will be required to elucidate this issue.
We also assessed the patterns of known drug resistance mutations to currently
available ARV drugs among treated patients. Some significant subtype-specific
differences could be observed. Mutation V82A/F/T/S of PR was more frequent in
subtype F1-infected subjects than to subtype B counterparts (p=0.0209). This result
seems to corroborate previous observations by Caride et al. (34) in non-B-infected

14
individuals failing antiretroviral therapy. Their difference did not reach statistical
significance, probably due to the low number of isolates analyzed. In our study, however,
we were able to highlight that difference with statistical confidence for the first time.
Mutations in codon V82 do not seem to be significantly different between subtypes B and
C, corroborating previous observations (35), and may indeed be a specific feature of
subtype F1 and other subtypes not yet analyzed. However, it is important to note that our
number of subtype F1 sequences was still low and the analysis of larger numbers should
be done to draw definitive conclusions. In RT, mutation M41L was significantly more
frequent in subtype B than in subtype C (p=0.0308), and this difference was even more
significant when comparing subtypes B and non-B (p=0.011). Finally, mutation L210W
seemed to be more associated with non-B than with B subtype, although only a trend was
observed (p=0.0554). The importance of RT-associated drug resistance mutations in
different HIV-1 subtypes is less well known and understood than those of PR mutations
and only a few studies have been conducted to look at this issue (35-37). Of note, the
study by Grossman et al. has also noticed a statistically significant association between
mutations M41L and L210W with subtype B when compared to subtype C (35). It was
hypothesized that this difference could be due to differences in times of previous
exposure to NRTI mono- and/or dual therapy between both subtypes. Indeed, the average
times under therapy for each group in our study differed (46.7 months for subtype B and
34.8 months for subtype C), and this could explain higher rates of NRTI-associated
mutations in subtype B. The current inclusion of more isolates to this study and the
matching of NRTI exposure times among both groups will be needed to clarify this issue.

15
The idea that the efficacy of selected ARV drugs might be influenced by HIV-1
genetic polymorphisms and by specific baseline sequence patterns of different HIV-1
subtypes is of increasing concern (34-37). The similar prevalences of subtypes B and C in
our casuistic makes southern Brazil a perfect setting for clinical, treatment and vaccine
trials, where a control group of subtype B-infected individuals with similar socio-
demographical and ethnical characteristics is readily available.
ACKNOWLEDGEMENTS
We are in profound debt to Caroline Rech, Giancarlo Marafon and Mauricio Kunz
from Hospital de Clinicas de Porto Alegre for helping with subjects’ follow-up, study
enrollment and sample collection, and to Andre F.A. Santos and Thatiana M. Souza from
Universidade Federal do Rio de Janeiro for helping with processing and sequencing of
the samples.
REFERENCES
1. Osmanov S, C. Pattou, Walker N, et al. Estimated global distribution and regional
spread of HIV-1 genetic subtypes in the year 2000. J Acquir Immune Defic Syndr
2002; 29:184-90.
2. McCormack GP, Glynn JR, Crampin AC, et al. Early evolution of the human
immunodeficiency virus type 1 subtype C epidemic in rural Malawi. J Virol 2002;
76:12890-9.
3. Robbins KE, Kostrikis LG, Brown TM, et al. Genetic analysis of human
immunodeficiency virus type 1 strains in Kenya: a comparison using phylogenetic

16
analysis and a combinatorial melting assay. AIDS Res Hum Retroviruses 1999;
15:329-35.
4. Snoeck J, Van Dooren S, Van Laethem K, et al. Prevalence and origin of HIV-1
group M subtypes among patients attending a Belgian hospital in 1999. Virus Res
2002; 85:95-107.
5. Balotta C, Facchi G, Violin M, et al. Increasing prevalence of non-clade B HIV-1
strains in heterosexual men and women, as monitored by analysis of reverse
transcriptase and protease sequences. J Acquir Immune Defic Syndr 2001; 27:499-
505.
6. Puchhammer-Stockl E, Kunz C, Faatz E, et al. Introduction of HIV-1 subtypes C,
E and A into Austria. Clin Diagn Virol 1999; 9:25-8.
7. Sonnerborg A, Durdevic S, Giesecke J, et al. Dynamics of the HIV-1 subtype
distribution in the Swedish HIV-1 epidemic during the period 1980 to 1993. AIDS
Res Hum Retroviruses 1997; 13:343-5.
8. Novitsky V, Smith UR, Gilbert P, et al. Human immunodeficiency virus type 1
subtype C molecular phylogeny: consensus sequence for an AIDS vaccine
design? J Virol 2002; 76:5435-51.
9. Gaschen B, Taylor J, Yusim K, et al. Diversity considerations in HIV-1 vaccine
selection. Science 2002; 296:2354-60.
10. Cao H, Mani I, Vincent R, et al. Cellular immunity to human immunodeficiency
virus type 1 (HIV-1) clades: relevance to HIV-1 vaccine trials in Uganda. J Infect
Dis 2000; 182:1350-6.

17
11. Dorrell L, Dong T, Ogg GS, et al. Distinct recognition of non-clade B human
immunodeficiency virus type 1 epitopes by cytotoxic T lymphocytes generated
from donors infected in Africa. J Virol 1999; 73:1708-14.
12. Velazquez-Campoy A, Vega S, Freire E. Amplification of the effects of drug
resistance mutations by background polymorphisms in HIV-1 protease from
African subtypes. Biochemistry 2002; 41:8613-9.
13. Velazquez-Campoy A, Todd MJ, Vega S, et al. (2001). Catalytic efficiency and
vitality of HIV-1 proteases from African viral subtypes. Proc Natl Acad Sci USA
2001; 98:6062-7.
14. Holguín A, Álvarez A, Soriano V. High prevalence of HIV-1 subtype G and
natural polymorphisms at the protease gene among HIV-infected immigrants in
Madrid. AIDS 2002; 16:1163-70.
15. Bongertz V, Bou-Habib DC, Brigido LF, et al. HIV-1 diversity in Brazil: genetic,
biologic, and immunologic characterization of HIV-1 strains in three potential
HIV vaccine evaluation sites. Brazilian Network for HIV Isolation and
Characterization. J Acquir Immune Defic Syndr 2000; 23:184-93.
16. Couto-Fernandez JC, Morgado MG, Bongertz V, et al. HIV-1 subtyping in
Salvador, Bahia, Brazil: a city with African sociodemographic characteristics. J
Acquir Immune Defic Syndr 2000; 22:288-93.
17. Cornelissen M, Kampinga G, Zorgdrager F, et al. Human immunodeficiency virus
type 1 subtypes defined by env show high frequency of recombinant gag genes.
The UNAIDS Network for HIV Isolation and Characterization. J Virol 1996;
70:8209-12.

18
18. Brindeiro RM, Diaz RS, Sabino EC, et al. Brazilian Network for HIV Drug
Resistance Surveillance (HIV-BResNet): results from the first national survey.
AIDS 2003; 17:1063-1069.
19. Dumans AT, Soares MA, Pieniazek D, et al. Prevalence of protease and reverse
transcriptase drug resistance mutations over time in drug-naive human
immunodeficiency virus type 1-positive individuals in Rio de Janeiro, Brazil.
Antimicrob Agents Chemother 2002; 46:3075-9.
20. Turchi MD, Diaz RS, Martelli CM, et al. Genetic diversity and HIV-1 incidence
estimation among cocaine users in Sao Paulo, Brazil. J Acquir Immune Defic
Syndr 2002; 30:527-32.
21. Soares MA, Oliveira T, Brindeiro RM, et al. A specific subtype C of human
immunodeficiency virus type 1 circulates in Brazil. AIDS 2003; 17:1-11.
22. Stuyver L, Wyseur A, Rombout A, et al. Line probe assay for rapid detection of
drug-selected mutations in the human immunodeficiency virus type 1 reverse
transcriptase gene. Antimicrob Agents Chemother 1997; 41:284-91.
23. Korber BT, Learn G, Mullins JI, et al. Protecting HIV databases. Nature 1995;
378:242-4.
24. Durand P, Canard L, Mornon JP. Visual BLAST and visual FASTA: graphic
workbenches for interactive analysis of full BLAST and FASTA outputs under
Microsoft Windows 95/NT. Comput Appl Biosci 1997; 13:407-13.
25. Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity
of progressive multiple sequence alignment through sequence weighting,

19
position-specific gap penalties and weight matrix choice. Nucleic Acids Res 1994;
22:4673-80.
26. Smith SW, Overbeek CR, Woese W, et al. The Genetic Data Environment: An
expandable GUI for multiple sequence analysis. Comput Appl Biosci 1994;
10:671-5.
27. Swofford D. PAUP 4.0: Phylogenetic analysis using parsimony (and other
methods), 4.0b2a. Sunderland, MA, USA: Sinauer Associates, Inc. 1999.
28. Gurtler LG, Hauser PH, Eberle J, et al. A new subtype of human
immunodeficiency virus type 1 (MVP-5180) from Cameroon. J Virol 1994;
68:1581-5.
29. Kantor R, Machekano R, Gonzales MJ, et al. Human Immunodeficiency Virus
Reverse Transcriptase and Protease Sequence Database: an expanded data model
integrating natural language text and sequence analysis programs. Nucleic Acids
Res 2001; 29:296-9.
30. D’Aquila RT, Shapiro JM, Brun-Vézinet F, et al. Drug resistance mutations in
HIV-1. Topics HIV Med 2002; 10:21-5.
31. Ramos A, Tanuri A, Schechter M, et al. Dual and recombinant infections: an
integral part of the HIV-1 epidemic in Brazil. Emerg Infect Dis 1999; 5:65-74.
32. Tanuri A, Swanson P, Devare S, et al. HIV-1 subtypes among blood donors from
Rio de Janeiro, Brazil. J Acquir Immune Defic Syndr Hum Retrovirol 1999;
20:60-6.

20
33. Martinez AMB, Mendoza-Sassi G, Mendonça-Signorini V, et al. Prevalence of
subtypes B and C and correlation to survival in HIV-1-positive patients followed
at UH in the city of Rio Grande, RS. Braz J Infec Dis 2001; 5:S22.
34. Caride E, Hertogs K, Larder B, et al. Genotypic and phenotypic evidence of
different drug-resistance mutation patterns between B and non-B subtype isolates
of human immunodeficiency virus type 1 found in Brazilian patients failing
HAART. Virus Genes 2001; 23:193-202.
35. Grossman Z, Vardinon N, Chemtob D, et al. Genotypic variation of HIV-1
reverse transcriptase and protease: comparative analysis of clade C and clade B.
AIDS 2001; 15:1453-60.
36. Caride E, Brindeiro RM, Hertogs K, et al. Drug-resistant reverse transcriptase
genotyping and phenotyping of B and non-B subtypes (F and A) of human
immunodeficiency virus type I found in Brazilian patients failing HAART.
Virology 2000; 275:107-15.
37. Pérez-Álvarez L, Cuevas MT, Villahermosa ML, et al. Prevalence of drug
resistance mutations in B, non-B subtypes, and recombinant forms of human
immunodeficiency virus type 1 in infected individuals in Spain (Galicia). J Hum
Virol 2001; 4:35-8.
21
Table 1. Baseline features of HIV-1-infected individuals included in this study TOTAL SUBTYPE B SUBTYPE C Mean Age (yr) (SD) 39.1 (10.4) 38.4 (10.9) 40.0 (10.2)
Sex (%)
Male 32 (44%) 17 (49%) 10 (37%)
Female 40 (56%) 18 (51%) 17 (63%)
Mean time of HIV infection (yr) (SD) 6.4 (3.6) 6.9 (3.8) 6.0 (3.4)
CDC Clinical Stage (%)
A 43 (60%) 16 (45%) 19 (70%)
B 12 (17%) 9 (26%) 1 (4%)
C 17 (23%) 10 (29%) 7 (26%)
CDC Immunological Stage (%)
1 15 (21%) 6 (17%) 7 (26%)
2 41 (57%) 19 (54%) 16 (59%)
3 16 (22%) 10 (29%) 4 (15%)
Mean CD4 cell count x 106/l (SD) 351.2 (192.9) 315 (183) 392.6 (206)
Median CD4 cell count x 106/l (IQR) 335.5 (210-454) 319 (173-401) 346 (250-515)
Treated 318 (166-406) 397 (282-500)
Untreated 313 (274-348) 336 (257-540)
Mean HIV RNA x 103/l (SD)
46,658 (163,787)
56,821 (210,869)
37,112 (117,516)
Median HIV RNA log10 x 103/l (IQR)
1,102 (0-14,515)
298 (0-12,074)
2,061 (0-12,781)
Treated 264 (0-10,376) 183 (0-3,213) 680 (806-16,796)
Untreated 7,200 (2,343-24,264)
12,416 (8,812-45,926)
3,902 (2,202-6,694)
Undetectable# viral load (%) 19 (26.4%) 10 (28.6%) 6 (22.2%)
Treatment Status (%)
Treated 58 (80%) 29 (83%) 21 (78%)
Untreated 14 (20%) 6 (17%) 6 (22%)
HIV-1 subtype (%)
B 35 (45%) N/A N/A
C 31 (40%) N/A N/A
F1 9 (12%) N/A N/A
Mosaics 2 (3%) N/A N/A
Yr, year; SD, standard deviation; IQR, inter-quartile range; N/A, not applied
# Defined as less than 80 copies of HIV RNA per milliliter of plasma

22
Table 2. Prevalence of primary protease inhibitor-associated drug resistance mutations
among drug-experienced subjects analyzed in this study
MUTATION Sub B (n=21)
Sub C (n=15)
Sub F1 (n=5)
Mosaics (n=2)
D30N 0 0 0 0
M46I/L 3 0 1 1
G48V 0 0 1 0
I50V 0 0 1 0
V82A/F/T/S 3 1 4 1
I84V 1 0 0 0
L90M 4 0 0 0

23
Table 3. Prevalence of reverse transcriptase inhibitor-associated drug resistance
mutations among drug-experienced subjects analyzed in this study
MUTATION Sub B (n=24)
Sub C (n=17)
Sub F1 (n=4)
Mosaics (n=2)
NRTI M41L 7 0 0 0 E44D 1 0 0 0 A62V 0 1 0 0 K65R 1 0 0 0 D67N 1 1 1 0 T69D 1 1 0 0 69ins 0 0 0 0 K70R 2 0 1 0 L74V 2 0 0 0 V75I 5 1 0 0 F77L 0 0 0 0
Y115F 0 0 0 0 F116Y 0 1 0 0 V118I 1 1 0 0
Q151M 0 1 0 0 M184V 8 1 0 1 L210W 4 0 0 0
T215Y/F 5 1 1 0 K219Q/E 4 1 1 0 NNRTI L100I 3 1 0 0 K103N 5 2 0 0 V106A 2 0 0 0 V108I 0 0 0 0
Y181C/I 1 0 0 0 Y188C/L/H 0 0 0 0
G190S/A 1 0 0 0 NRTI, nucleoside reverse transcriptase inhibitor; NNRTI, non-nucleoside reverse transcriptase inhibitor

24
FIGURE LEGENDS
Figure 1. HIV-1 subtype prevalence according to year of diagnosis of infected individuals
followed at Hospital de Clínicas de Porto Alegre, Brazil

25
Subtype Prevalence by Year of Diagnosis
0102030405060708090
1986-1990 1991-1995 1996-2000 2001-2002
time (years)
% o
f sub
type
s
sub Bsub Csub F1mosaics
4
14
5
1
2
3
1 11
17
2
9
12
Copyright © 2022 FDOKUMEN




















