
Enhanced apoptotic action of trichosanthin in HIV1 infected cells
6
Enhanced apoptotic action of trichosanthin in HIV-1 infected cells Yuan-Yuan Wang a,c , Dong-Yun Ouyang a,c , Hai Huang b , Herman Chan b , Siu-Cheung Tam b , Yong-Tang Zheng a, * a Laboratory of Molecular Immunopharmacology, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming 650223, China b Department of Physiology, The Chinese University of Hong Kong, Hong Kong SAR, China c Graduate School of the Chinese Academy of Sciences, Beijing 100039, China Received 17 March 2005 Available online 13 April 2005 Abstract Trichosanthin (TCS) is a type 1 ribosome-inactivating protein (RIP) effective against HIV-1 replication. The mechanism is not clear. Present results suggested that the antiviral action may be partly mediated through enhanced apoptosis on infected cells. TCS induced apoptosis in normal H9 cells and this action was more potent in those infected with HIV-1. In flow cytometry study, TCS induced larger population of apoptotic H9 cells chronically infected with HIV-1 in a dose-dependent manner. At TCS concentration of 25 lg/ml, 8.4% of normal H9 cells were found to be apoptotic whereas the same concentration induced 24.5% in HIV-1 chron- ically infected cells. Such difference was not found in the control experiments without TCS treatment. Two other studies supported this action. Cytotoxic study showed that cell viability was always lower in HIV-1 infected cells after TCS treatment, and DNA frag- mentation study confirmed more laddering in infected cells. The mechanism of TCS induced apoptosis in normal or infected H9 cells is not clear. Results in this study demonstrated that TCS is more effective in inducing apoptosis in HIV-1 infected cells. This may explain in part the antiviral action of TCS. Ó 2005 Elsevier Inc. All rights reserved. Keywords: Trichosanthin; HIV-1; Apoptosis; Infected cells; Anti-HIV activity Trichosanthin (TCS) is a type 1 ribosome-inactivat- ing protein (RIP) isolated from root tuber of Trichosan- thes kirilowii. This single chain polypeptide depurinates A-4324 of the 28S rRNA and inhibits protein synthesis [1]. In its crude form, it was used in ancient China as an abortifacient agent. The active ingredient is now known as trichosanthin. This protein was found to inhi- bit HIV-1 replication in the late 1980s and generated some research interest in the past years [2–8]. Clinical trial of TCS on AIDS patients had been conducted, and one of the major drawbacks was anaphylaxis [6–8]. Recently, several TCS derivatives were designed with less antigenicity while retaining most of the activity [9–11]. These derivatives are therefore potential antiviral agent. TCS inhibits viral replication including herpes sim- plex virus (HSV-1) [12] and human immunodeficiency virus (HIV-1). Less work is done on its antiviral mecha- nism and it is generally presumed that this action is re- lated to ribosome inactivation. Almost all ribosome inactivating proteins are cytotoxic. Type 2 ribosome inactivating proteins like ricin are usually extremely toxic while type 1 like TCS is generally much milder [13]. Since most ribosome inactivating proteins are anti- viral in nature [14], it is logical to explore the role of cytotoxicity as a possible antiviral mechanism. When a virus infects a cell, it will try to ensure survival of the cell long enough for its own replication [15,16]. If cytotoxic effect of TCS is enhanced on infected cells, premature death of infected cells will limit massive viral replication 0006-291X/$ - see front matter Ó 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2005.03.230 * Corresponding author. Fax: +86 871 5191823. E-mail address: [email protected] (Y.-T. Zheng). www.elsevier.com/locate/ybbrc Biochemical and Biophysical Research Communications 331 (2005) 1075–1080 BBRC
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Enhanced apoptotic action of trichosanthin in HIV1 infected cells

www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 331 (2005) 1075–1080
BBRC
Enhanced apoptotic action of trichosanthin in HIV-1 infected cells
Yuan-Yuan Wang a,c, Dong-Yun Ouyang a,c, Hai Huang b, Herman Chan b,Siu-Cheung Tam b, Yong-Tang Zheng a,*
a Laboratory of Molecular Immunopharmacology, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming 650223, Chinab Department of Physiology, The Chinese University of Hong Kong, Hong Kong SAR, China
c Graduate School of the Chinese Academy of Sciences, Beijing 100039, China
Received 17 March 2005Available online 13 April 2005
Abstract
Trichosanthin (TCS) is a type 1 ribosome-inactivating protein (RIP) effective against HIV-1 replication. The mechanism is notclear. Present results suggested that the antiviral action may be partly mediated through enhanced apoptosis on infected cells. TCSinduced apoptosis in normal H9 cells and this action was more potent in those infected with HIV-1. In flow cytometry study, TCSinduced larger population of apoptotic H9 cells chronically infected with HIV-1 in a dose-dependent manner. At TCS concentrationof 25 lg/ml, 8.4% of normal H9 cells were found to be apoptotic whereas the same concentration induced 24.5% in HIV-1 chron-ically infected cells. Such difference was not found in the control experiments without TCS treatment. Two other studies supportedthis action. Cytotoxic study showed that cell viability was always lower in HIV-1 infected cells after TCS treatment, and DNA frag-mentation study confirmed more laddering in infected cells. The mechanism of TCS induced apoptosis in normal or infected H9 cellsis not clear. Results in this study demonstrated that TCS is more effective in inducing apoptosis in HIV-1 infected cells. This mayexplain in part the antiviral action of TCS.� 2005 Elsevier Inc. All rights reserved.
Keywords: Trichosanthin; HIV-1; Apoptosis; Infected cells; Anti-HIV activity
Trichosanthin (TCS) is a type 1 ribosome-inactivat-ing protein (RIP) isolated from root tuber of Trichosan-thes kirilowii. This single chain polypeptide depurinatesA-4324 of the 28S rRNA and inhibits protein synthesis[1]. In its crude form, it was used in ancient China asan abortifacient agent. The active ingredient is nowknown as trichosanthin. This protein was found to inhi-bit HIV-1 replication in the late 1980s and generatedsome research interest in the past years [2–8]. Clinicaltrial of TCS on AIDS patients had been conducted,and one of the major drawbacks was anaphylaxis[6–8]. Recently, several TCS derivatives were designedwith less antigenicity while retaining most of the activity
0006-291X/$ - see front matter � 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2005.03.230
* Corresponding author. Fax: +86 871 5191823.E-mail address: [email protected] (Y.-T. Zheng).
[9–11]. These derivatives are therefore potential antiviralagent.
TCS inhibits viral replication including herpes sim-plex virus (HSV-1) [12] and human immunodeficiencyvirus (HIV-1). Less work is done on its antiviral mecha-nism and it is generally presumed that this action is re-lated to ribosome inactivation. Almost all ribosomeinactivating proteins are cytotoxic. Type 2 ribosomeinactivating proteins like ricin are usually extremelytoxic while type 1 like TCS is generally much milder[13]. Since most ribosome inactivating proteins are anti-viral in nature [14], it is logical to explore the role ofcytotoxicity as a possible antiviral mechanism. When avirus infects a cell, it will try to ensure survival of the celllong enough for its own replication [15,16]. If cytotoxiceffect of TCS is enhanced on infected cells, prematuredeath of infected cells will limit massive viral replication
1076 Y.-Y. Wang et al. / Biochemical and Biophysical Research Communications 331 (2005) 1075–1080
and eventually save the population. The objective of thisstudy was to compare the cytotoxic effect of TCS onnormal and infected cells.
Methods
Trichosanthin. Natural TCS was purified from the root tuber ofT. kirilowii and obtained from Shanghai Jinshan PharmaceuticalLimited Company (Shanghai, PRC).
HIV-1 culture and treatment. Normal H9 cells and H9 cellschronically infected with HIV-1IIIB (H9/HIV-1IIIB) were obtained fromMedical Research Council, AIDS Regent Project, UK. They werecultured in RPMI-1640 medium (Gibco) supplemented with 10% fetalcalf serum together with 100 U/ml penicillin, 100 mg/L streptomycinand glutamine. Incubation was done at 37 �C in 5% CO2. H9/HIV-1IIIBcells were achieved by repeated exposure of the H9 cells to a highmultiplicity of infection (MOI). H9 cells were washed with PBS, re-suspended in 0.2–0.6 ml of HIV-1IIIB inocula, and incubated at 37 �Cfor 40–60 min. The cells were then washed again with PBS. Pelletedcells were re-suspended in 5–7 ml of complete RPMI-1640 medium and
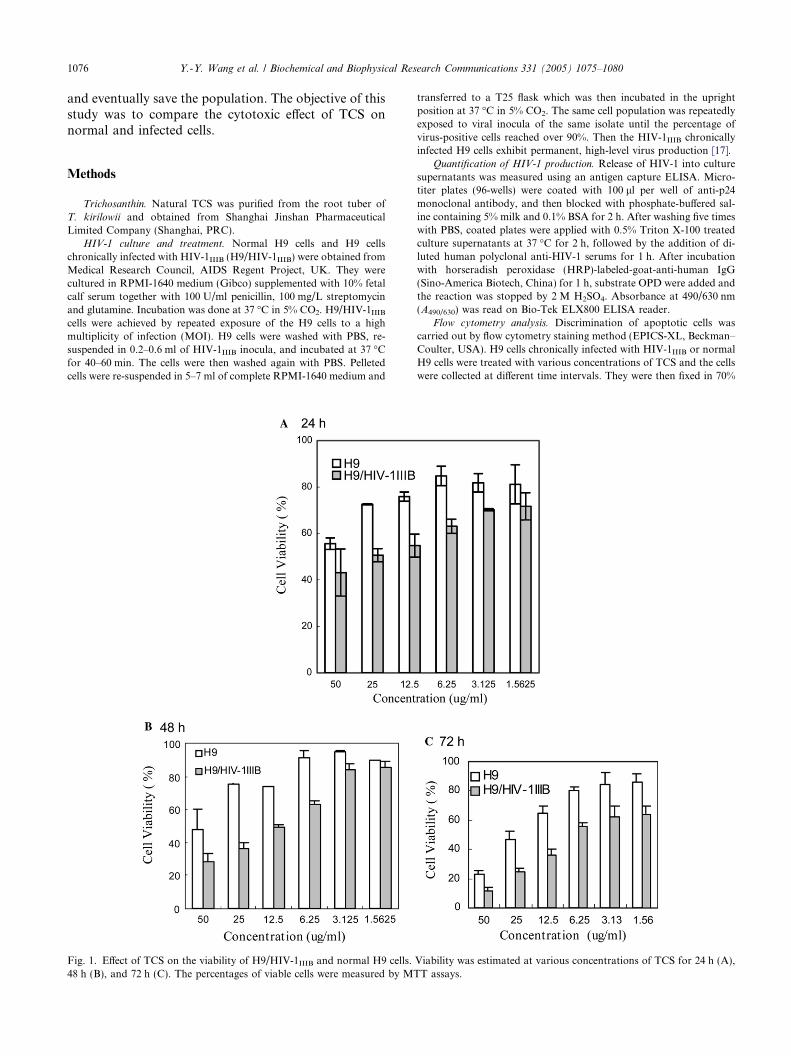
Fig. 1. Effect of TCS on the viability of H9/HIV-1IIIB and normal H9 cells.48 h (B), and 72 h (C). The percentages of viable cells were measured by MT
transferred to a T25 flask which was then incubated in the uprightposition at 37 �C in 5% CO2. The same cell population was repeatedlyexposed to viral inocula of the same isolate until the percentage ofvirus-positive cells reached over 90%. Then the HIV-1IIIB chronicallyinfected H9 cells exhibit permanent, high-level virus production [17].
Quantification of HIV-1 production. Release of HIV-1 into culturesupernatants was measured using an antigen capture ELISA. Micro-titer plates (96-wells) were coated with 100 ll per well of anti-p24monoclonal antibody, and then blocked with phosphate-buffered sal-ine containing 5% milk and 0.1% BSA for 2 h. After washing five timeswith PBS, coated plates were applied with 0.5% Triton X-100 treatedculture supernatants at 37 �C for 2 h, followed by the addition of di-luted human polyclonal anti-HIV-1 serums for 1 h. After incubationwith horseradish peroxidase (HRP)-labeled-goat-anti-human IgG(Sino-America Biotech, China) for 1 h, substrate OPD were added andthe reaction was stopped by 2 M H2SO4. Absorbance at 490/630 nm(A490/630) was read on Bio-Tek ELX800 ELISA reader.
Flow cytometry analysis. Discrimination of apoptotic cells wascarried out by flow cytometry staining method (EPICS-XL, Beckman–Coulter, USA). H9 cells chronically infected with HIV-1IIIB or normalH9 cells were treated with various concentrations of TCS and the cellswere collected at different time intervals. They were then fixed in 70%
Viability was estimated at various concentrations of TCS for 24 h (A),T assays.

Y.-Y. Wang et al. / Biochemical and Biophysical Research Communications 331 (2005) 1075–1080 1077
alcohol over 18 h at �20 �C. After washing with PBS, the cells weretreated with RNase A (10 mg/ml in 1 M Tris–Cl, pH 7.4) and stainedwith propidium iodide (PI) solution (100 lg/ml in phosphate-bufferedsaline, pH 7.4). PI fluorescence of individual cells was measured byflow cytometry. D-Sorbital is used as positive control.
DNA fragmentation assay. Cell treatment was similar to those de-scribed under flow cytometry. The cells were collected and suspendedin 20 ll of 50 mM Tris–Cl (pH 8.0), 10 mM EDTA, and 0.5 mg/mlproteinase K. After incubation at 50 �C for 2 h, 10 ll of 0.5 lg/mlRNase A solution was added, and the suspension was incubated for
Fig. 2. Programmed cell death induced by TCS. H9/HIV-1IIIB (A) and normwere washed in PBS and then fixed in 70% ethanol, stained with propidiumrepresent the percentages of apoptotic cells containing DNA at less than G1
another 2 h. DNA was analyzed by electrophoresis in 1.5% agarosegels, stained with ethidium bromide, and then photographed on a UVtransilluminator [18].
Results
TCS was incubated with H9 and H9/HIV-1IIIB cellsfor 24–72 h. The percentage of viable cells as determined
al H9 cells (B) were cultured with TCS at 25 lg/ml for 36 h. The cellsiodide, and analyzed by flow cytometry. Numbers above histograms(sub-diploid cells).

Fig. 4. Oligonucleosomal DNA fragmentation in TCS-treated cells.H9/HIV-1IIIB infected and normal H9 cells were treated with indicatedconcentrations of TCS for 36 h. After harvest of the cells, nuclearDNA fragmentation was analyzed by 1.5% agarose gel electrophoresis.1, H9/HIV-1IIIB TCS 25 lg/ml; 2, H9 TCS 25 lg/ml; 3, H9/HIV-1IIIBTCS 12.5 lg/ml; 4, H9 TCS 12.5 lg/ml; 5, H9/HIV-1IIIB TCS 1 lg/ml;6, H9 TCS 1 lg/ml; 7, H9/HIV-1IIIB D-sorbitol 300 mM; 8, H9D-sorbitol 300 mM; 9, H9/HIV-1IIIB cells; and 10, H9 cells.
1078 Y.-Y. Wang et al. / Biochemical and Biophysical Research Communications 331 (2005) 1075–1080
by MTT was always lower in the H9/HIV-1IIIB cells(Fig. 1). For incubation period of up to 48 h, viabilitybetween normal and infected cells was not significantlydifferent at low dosages of TCS (1.5625 lg/ml). Whenthe incubation period was extended to 72 h, there wassignificant difference in cell viability at all TCS dosages(p < 0.01). In the control experiment, viability of normaland infected cells was not different in the absence of TCSwithin the incubation period of up to 72 h. For example,proliferation of H9/HIV-1IIIB (2.8 ± 0.07-fold) and H9(2.9 ± 0.1-fold) is not significantly different after 48 hof incubation. To elaborate on the cause of death, apop-totic cells discriminated by the PI staining method wereestimated by flow cytometry. During apoptosis, DNAbreaks down into small fragments. Following appropri-ate fixation and elution, these fragments are lost fromthe nucleus. As these apoptotic cells now have a lowerDNA content, subsequent staining with a DNA-bindingdye (PI) will reveal these cells in the sub-G1 region (Fig.2). A summary of the fraction of apoptotic cells under
Fig. 3. TCS induced more apoptosis in H9/HIV-1IIIB cells thannormal H9 cells in a dose-dependent manner. Cells were treated withTCS at various concentrations for 24 h (A) to 36 h (B). Thepercentages of apoptotic cells were measured as sub-diploid cells byflow cytometric analysis. D-Sorbital is used as positive control. Dataare expressed as means ± SEM (n = 3).
various treatment conditions is shown in Fig. 3. TCSsignificantly induced more apoptosis in H9/HIV-1IIIBthan H9 (p < 0.01) in a dose-dependent manner. Largerdifferences were found with higher dosage of TCS.D-Sorbital was used as a positive control. This com-pound induced apoptosis but did not demonstrate selec-tivity between H9 and H9/HIV-1IIIB cells. In anothercontrol experiment done under similar condition andin the absence of TCS, the apoptotic ratio was lowand showed no difference between normal and infectedcells. To further substantiate the differential apoptoticeffect of TCS on H9 and H9/HIV-1IIIB cells, grossnucleosomal DNA ladder was monitored. Degradationof DNA into oligonucleosomal fragments was observedafter exposing H9/HIV-1IIIB cells to TCS at indicatedconcentrations. Even at the lowest concentration ofTCS 1 lg/ml, the ladder of H9/HIV-1IIIB cells was stillclear (Fig. 4).
Discussion
This study demonstrated that TCS is more toxic toHIV-1 infected cells than the normal host cells. The evi-dence showed consistency and is generated from differ-ent studies like flow cytometry study, cytotoxicitystudy, and DNA fragmentation study. The implicationof this enhanced toxic effect is that it may be in partresponsible for the antiviral mechanism of TCS. Anycompound that kills cells will inhibit viral replication,but these compounds cannot be qualified as an antiviralagent because this is simply a cellular massacre. An idealantiviral agent must be able to kill the infected cellswhile sparing the normal ones. TCS demonstrated selec-tivity in killing infected cells and we propose that thismay be in part responsible for the antiviral action ofTCS.

Y.-Y. Wang et al. / Biochemical and Biophysical Research Communications 331 (2005) 1075–1080 1079
When a virus infected its host, it will try to ensuresurvival of the cells for its replication. The signal tothe cells should be anti-apoptosis. It was previously re-ported that the MAPK pathway plays an important rolein HIV-1 production and virion infectivity [19–22]. If theantiviral action of TCS indeed works through cytotoxic-ity of infected cells, it will be most effective in the earlyphase of infection when there is active viral replicationand cells are gradually infected. In a previous study,early TCS or other RIP treatment to cells is much moreeffective than a late treatment [4,23].
HIV-1 integrated chronically into infected cells as thevirus reservoir. It is more resistant to the cytopathic ef-fect of the virus in vivo and continued to produce virion.This has been considered to play a central role in theprogression of HIV-1-associated diseases [24]. Currenttherapies used in the treatment of HIV-1 infected pa-tients include combinations of reverse transcriptaseinhibitors and HIV-1 protease inhibitors which canincrease CD4+ T cell counts through reducing the ‘‘by-stander’’ CD4+ T cell apoptosis [25]. Nevertheless,it cannot eradicate the chronically infected cells com-pletely. If TCS indeed works through killing of infectedcells, this enhanced killing may be an added advantageif it is included in the cocktail treatment regiment. The-oretically TCS can eradicate chronically infected cellsdue to enhanced killing.
In summary, this study provides some evidences inshowing that TCS induced apoptosis is enhanced inchronically HIV-1 infected cells.
Acknowledgments
We acknowledge MRC (UK), AIDS Research Pro-ject for providing cell lines and virus. We thank Dr.X.C. Wang and C.G. Jin (Tumor Institute of YunnanProvince) for flow cytometry technical assistance. Thework was supported by grants from the Natural ScienceFoundation of China (30471605) and Yunnan Province(2002C0066M; 2003C0022R); CAS Knowledge Innova-tion Projects (KSCX2-SW-216), Key Scientific andTechnological projects of China (2004BA719A14) andYunnan Province (2004NG12), and 863 Program(2003AA219142).
References
[1] J.S. Zhang, W.Y. Liu, The mechanism of action of trichosanthinon eukaryotic ribosomes-RNA N-glycosidase activity of thecytotoxin, Nucleic Acids Res. 20 (1992) 1271–1275.
[2] M.S. McGrath, S. Santulli, I. Gaston, Effects of GLQ223 on HIVreplication in human monocyte/macrophages chronically infectedin vitro with HIV, AIDS Res. Hum. Retroviruses 6 (1990) 1039–1043.
[3] M.S. McGrath, K.M. Hwang, S.E. Caldwell, I. Gaston, K.C.Luk, P. Wu, V.L. Ng, S. Crowe, J. Daniels, J. Marsh, T.
Deinhart, P.V. Lekas, J.C. Vennari, H.W. Yeung, J.D. Lifson,GLQ223: an inhibitor of human immunodeficiency virus replica-tion in acutely and chronically infected cells of lymphocyte andmononuclear phagocyte lineage, Proc. Natl. Acad. Sci. USA 86(1989) 2844–2848.
[4] J.H. Wang, H.L. Nie, S.C. Tam, H. Huang, Y.T. Zheng, Anti-HIV-1 property of trichosanthin correlates with its ribosomeinactivating activity, FEBS Lett. 531 (2002) 295–298.
[5] J.H. Wang, H.L. Nie, H. Huang, S.C. Tam, Y.T. Zheng,Independency of anti-HIV-1 activity from ribosome-inactivatingactivity of trichosanthin, Biochem. Biophys. Res. Commun. 302(2003) 89–94.
[6] J.O. Kahn, K.J. Gorelick, G. Gatti, C.J. Arri, J.D. Lifson, J.G.Gambertoglio, A. Bostrom, R. Williams, Safety, activity, andpharmacokinetics of GLQ223 in patients with AIDS and AIDS-related complex, Antimicrob. Agents Chemother. 38 (1994) 260–267.
[7] V.S. Byers, A.S. Levin, A. Malvino, L. Waites, R.A. Robins,R.W. Baldwin, A phase II study of effect of addition oftrichosanthin to zidovudine in patients with HIV disease andfailing antiretroviral agents, AIDS Res. Hum. Retroviruses 10(1994) 413–420.
[8] V.S. Byers, A.S. Levin, L.A. Waites, B.A. Starrett, R.A. Mayer,J.A. Clegg, M.R. Price, R.A. Robins, M. Delaney, R.W. Baldwin,Phase I/II study of trichosanthin treatment of HIV disease, AIDS4 (1990) 1189–1196.
[9] W.L. Chan, P.C. Shaw, X.B. Li, Q.F. Xu, X.H. He, S.C.Tam, Lowering of trichosanthin immunogenicity by site-specific coupling to dextran, Biochem. Pharmacol. 57 (1999)927–934.
[10] X.H. He, P.C. Shaw, L.H. Xu, S.C. Tam, Site-directed polyeth-ylene glycol modification of trichosanthin: effects on its biologicalactivities, pharmacokinetics, and antigenicity, Life Sci. 64 (1999)1163–1175.
[11] X.H. He, P.C. Shaw, S.C. Tam, Reducing the immunogenicityand improving the in vivo activity of trichosanthin by site-directedpegylation, Life Sci. 65 (1999) 355–368.
[12] Y.T. Zheng, W.L. Chan, P. Chan, H. Huang, S.C. Tam,Enhancement of the anti-herpetic effect of trichosanthin byacyclovir and interferon, FEBS Lett. 496 (2001) 139–142.
[13] F. Stirpe, L. Barbieri, Ribosome-inactivating proteins up to date,FEBS Lett. 195 (1986) 1–8.
[14] A.J. Vlietinck, T.D. Bruyne, S. Apers, L.A. Pieters, Plant-derivedleading compounds for chemotherapy of human immunodefi-ciency virus (HIV) infection, Planta Med. 64 (1998) 97–109.
[15] C. DeLuca, H. Kwon, N. Pelletier, M.A. Wainberg, J. Hiscott,NF-kB protects HIV-1-Infected myeloid cells from apoptosis,Virology 244 (1998) 27–38.
[16] R. Geleziunas, W. Xu, K. Takeda, H. Ichijo, W.C. Greene, HIV-1 Nef inhibits ASK1-dependent death signalling providing apotential mechanism for protecting the infected host cell, Nature410 (2001) 834–838.
[17] S. Gartner, M. Popovic, Virus isolation and production, in: A.Aldovini, B.D. Walker (Eds.), Techniques in HIV Research,Stockton, New York, 1990, pp. 53–69.
[18] K. Noguchi, M. Naito, S. Kataoka, S. Yonehara, T. Tsuruo, Arecessive mutant of the U937 cell line acquired resistance to anti-Fas and anti-p55 tumor necrosis factor receptor antibody-inducedapoptosis, Cell Growth Differ. 6 (1995) 1271–1277.
[19] C. Cartier, M. Deckert, C. Grangeasse, R. Trauger, F. Jensen, A.Bernard, A. Cozzone, C. Desgranges, V. Boyer, Association ofERK2 mitogen-activated protein kinase with human immunode-ficiency virus particles, J. Virol. 71 (1997) 4832–4837.
[20] J.M. Jacque, A. Mann, H. Enslen, N. Sharova, B. Brichacek,R.J. Davis, M. Stevenson, Modulation of HIV-1 infectivity byMAPK, a virion-associated kinase, EMBO J. 17 (1998) 2607–2618.

1080 Y.-Y. Wang et al. / Biochemical and Biophysical Research Communications 331 (2005) 1075–1080
[21] L. Shapiro, K.A. Heidenreich, M.K. Meintzer, C.A. Dinarello,Role of p38 mitogen-activated protein kinase in HIV type 1production in vitro, Proc. Natl. Acad. Sci. USA 95 (1998) 7422–7426.
[22] X. Yang, D. Gabuzda, Regulation of human immunodeficiencyvirus type 1 infectivity by the ERK mitogen-activated proteinkinase signalling pathway, J. Virol. 73 (1999) 3460–3466.
[23] G.J. Teltow, J.D. Irvin, G.M. Aron, Inhibition of herpes simplexvirus DNA synthesis by pokeweed antiviral protein, Antimicrob.Agents Chemother. 23 (1983) 390–396.
[24] S. Aquaro, P. Bagnarelli, T. Guenci, A. De Luca, M.Clementi, E. Balestra, R. Calio, C.F. Perno, Long-termsurvival and virus production in human primary macrophagesinfected by human immunodeficiency virus, J. Med. Virol. 68(2002) 479–488.
[25] F. Ensoli, V. Fiorelli, C. Alario, M. De Cristofaro, D. SantiniMuratori, A. Novi, M.G. Cunsolo, F. Mazzetta, A. Giovannetti,B. Mollicone, E. Pinter, F. Aiuti, Decreased T cell apoptosis andT cell recovery during highly active antiretroviral therapy(HAART), Clin. Immunol. 97 (2000) 9–20.




















