
Enhanced anti-inflammatory effects of a nitric oxide–releasing derivative of mesalamine in rats
10
Enhanced Anti-inflammatory Effects of a Nitric Oxide–Releasing Derivative of Mesalamine in Rats JOHN L. WALLACE,* NATHALIE VERGNOLLE,* MARCELO N. MUSCARA ´ ,* SAMUEL ASFAHA,* KEVIN CHAPMAN,* WEBB MCKNIGHT,* PIERO DEL SOLDATO, ‡ ANTONIO MORELLI, § and STEFANO FIORUCCI § *Department of Pharmacology and Therapeutics, University of Calgary, Calgary, Alberta, Canada; ‡ NicOx S.A., Nice, France; and § Clinica di Gastroenterologia ed Epatologia, Dipar timento di Medicina Clinica e Sperimentale, Universita ` degli Studi di Perugia, Perugia, Italy Background & Aims: Nitric oxide (NO)-releasing deriva- tives of cyclooxygenase inhibitors exhibit enhanced anti-inflammatory activity and greatly reduced gastroin- testinal toxicity. We evaluated whether a similar deriva- tization of mesalamine (5-aminosalicylic acid) would improve its anti-inflammatory activity. Methods: Ef- fects of an NO-releasing derivative of mesalamine (NCX-456; NO-mesalamine) were compared with those of mesalamine itself and 2 other NO donors in a rat model of colitis. These drugs were compared for their ability to inhibit leukocyte adherence to the vascular endothelium in vivo, interleukin (IL)-1b and interferon (IFN)-g release in vitro (splenocytes and colon), and messenger RNA expression in the inflamed colon. Results: NO-mesalamine was significantly more effec- tive than mesalamine in reducing the severity of colitis (damage and granulocyte infiltration). Unlike mesala- mine, NO-mesalamine significantly suppressed leuko- cyte adherence to the vascular endothelium in vivo. NO-mesalamine inhibited IL-1b and IFN-g release and caspase 1 activity in splenocytes; such effects were not found in the inflamed colon. Conclusions: These studies show that an NO-releasing derivative of mesala- mine has significantly enhanced anti-inflammatory ac- tivity, including improved efficacy in a rat model of colitis. The improved efficacy of this derivative is most likely caused by its enhanced ability to suppress leukocyte infiltration and possibly to scavenge peroxy- nitrite. M esalamine (5-aminosalicylic acid) is one of the most commonly used drugs for the treatment of active inflammatory bowel disease (IBD) and for maintenance of remission. Mesalamine is also believed to be the active moiety of sulfasalazine, another drug that is widely used in the treatment of IBD. Although mesalamine rarely produces significant adverse effects and has been shown to be an effective treatment for many patients, it fails to induce a marked improvement in a significant proportion of patients with IBD. 1 The mechanism of action of mesalamine is not clear, although it has been shown to exert many effects that could contribute to its anti- inflammatory activity. These include inhibition of leuko- triene synthesis, 2 scavenging of oxygen-derived free radi- cals, 3 scavenging of peroxynitrite, 4 and inhibition of interleukin (IL)-1b synthesis. 5 In recent years, we have shown that addition of a nitric oxide (NO)-releasing moiety to standard nonsteroidal anti-inflammatory drugs (NSAIDs) results in a substan- tial reduction in their toxicity in the gastrointestinal tract without a diminution of the anti-inflammatory, antipy- retic, or analgesic effects. 6–8 In some models, these NO-NSAIDs exhibited significantly enhanced anti- inflammatory and analgesic effects compared with the parent NSAIDs. 6,8,9 Moreover, an NO-releasing deriva- tive of diclofenac was shown to accelerate healing of gastric ulcers in the rat 10 and to be well tolerated by rats with colitis. 11 A significant acceleration of healing could be induced with another NO donor 10 or through admin- istration of a precursor for NO synthesis. 12 These findings suggested that the release of NO from the NO-NSAID accelerated healing of the ulcer and/or conferred protec- tion to the mucosa during an inflammatory event. Based on these findings, we hypothesized that adding an NO-releasing moiety to mesalamine may increase its effectiveness in reducing the severity of colitis. To test this hypothesis, we compared the effects of such a derivative (NCX-456; Figure 1; subsequently referred to as ‘‘NO-mesalamine’’) with those of mesalamine in a rat Abbreviations used in this paper: Ac-YVAD-CHO, acetyl Tyr-Val-Ala- Asp aldehyde; AFC, 7-amino-4-trifluoro-methylcoumarin; AMC, 7-amino-4-coumarin; DTT, diothiothreitol; ELISA, enzyme-linked im- munosorbent assay; FMLP, N-formyl-methionine-leucine-phenylala- nine; GAPDH, glyceraldehyde-3-phosphate-dehydrogenase; HPLC, high-performance liquid chromatography; IBD, inflammatory bowel disease; IFN-g, interferon gamma; IL-1b, interleukin 1b; iNOS, inducible nitric oxide; LPS, lipopolysaccharide; MPO, myeloperoxi- dase; NO, nitric oxide; RT-PCR, reverse-transcription polymerase chain reaction; TNBS, trinitrobenzene sulfonic acid. r 1999 by the American Gastroenterological Association 0016-5085/99/$10.00 GASTROENTEROLOGY 1999;117:557–566
Transcript of Enhanced anti-inflammatory effects of a nitric oxide–releasing derivative of mesalamine in rats

Enhanced Anti-inflammatory Effects of a NitricOxide–Releasing Derivative of Mesalamine in Rats
JOHN L. WALLACE,* NATHALIE VERGNOLLE,* MARCELO N. MUSCARA,* SAMUEL ASFAHA,*KEVIN CHAPMAN,* WEBB MCKNIGHT,* PIERO DEL SOLDATO,‡ ANTONIO MORELLI,§
and STEFANO FIORUCCI§*Department of Pharmacology and Therapeutics, University of Calgary, Calgary, Alberta, Canada; ‡NicOx S.A., Nice, France; and §Clinica diGastroenterologia ed Epatologia, Dipartimento di Medicina Clinica e Sperimentale, Universita degli Studi di Perugia, Perugia, Italy
Background & Aims: Nitric oxide (NO)-releasing deriva-tives of cyclooxygenase inhibitors exhibit enhancedanti-inflammatory activity and greatly reduced gastroin-testinal toxicity. We evaluated whether a similar deriva-tization of mesalamine (5-aminosalicylic acid) wouldimprove its anti-inflammatory activity. Methods: Ef-fects of an NO-releasing derivative of mesalamine(NCX-456; NO-mesalamine) were compared with thoseof mesalamine itself and 2 other NO donors in a ratmodel of colitis. These drugs were compared for theirability to inhibit leukocyte adherence to the vascularendothelium in vivo, interleukin (IL)-1b and interferon(IFN)-g release in vitro (splenocytes and colon), andmessenger RNA expression in the inflamed colon.Results: NO-mesalamine was significantly more effec-tive than mesalamine in reducing the severity of colitis(damage and granulocyte infiltration). Unlike mesala-mine, NO-mesalamine significantly suppressed leuko-cyte adherence to the vascular endothelium in vivo.NO-mesalamine inhibited IL-1b and IFN-g release andcaspase 1 activity in splenocytes; such effects werenot found in the inflamed colon. Conclusions: Thesestudies show that an NO-releasing derivative of mesala-mine has significantly enhanced anti-inflammatory ac-tivity, including improved efficacy in a rat model ofcolitis. The improved efficacy of this derivative is mostlikely caused by its enhanced ability to suppressleukocyte infiltration and possibly to scavenge peroxy-nitrite.
Mesalamine (5-aminosalicylic acid) is one of the mostcommonly used drugs for the treatment of active
inflammatory bowel disease (IBD) and for maintenance ofremission. Mesalamine is also believed to be the activemoiety of sulfasalazine, another drug that is widely usedin the treatment of IBD. Although mesalamine rarelyproduces significant adverse effects and has been shown tobe an effective treatment for many patients, it fails toinduce a marked improvement in a significant proportionof patients with IBD.1 The mechanism of action of
mesalamine is not clear, although it has been shown toexert many effects that could contribute to its anti-inflammatory activity. These include inhibition of leuko-triene synthesis,2 scavenging of oxygen-derived free radi-cals,3 scavenging of peroxynitrite,4 and inhibition ofinterleukin (IL)-1b synthesis.5
In recent years, we have shown that addition of a nitricoxide (NO)-releasing moiety to standard nonsteroidalanti-inflammatory drugs (NSAIDs) results in a substan-tial reduction in their toxicity in the gastrointestinal tractwithout a diminution of the anti-inflammatory, antipy-retic, or analgesic effects.6–8 In some models, theseNO-NSAIDs exhibited significantly enhanced anti-inflammatory and analgesic effects compared with theparent NSAIDs.6,8,9 Moreover, an NO-releasing deriva-tive of diclofenac was shown to accelerate healing ofgastric ulcers in the rat10 and to be well tolerated by ratswith colitis.11 A significant acceleration of healing couldbe induced with another NO donor10 or through admin-istration of a precursor for NO synthesis.12 These findingssuggested that the release of NO from the NO-NSAIDaccelerated healing of the ulcer and/or conferred protec-tion to the mucosa during an inflammatory event.
Based on these findings, we hypothesized that addingan NO-releasing moiety to mesalamine may increase itseffectiveness in reducing the severity of colitis. To testthis hypothesis, we compared the effects of such aderivative (NCX-456; Figure 1; subsequently referred toas ‘‘NO-mesalamine’’) with those of mesalamine in a rat
Abbreviations used in this paper: Ac-YVAD-CHO, acetyl Tyr-Val-Ala-Asp aldehyde; AFC, 7-amino-4-trifluoro-methylcoumarin; AMC,7-amino-4-coumarin; DTT, diothiothreitol; ELISA, enzyme-linked im-munosorbent assay; FMLP, N-formyl-methionine-leucine-phenylala-nine; GAPDH, glyceraldehyde-3-phosphate-dehydrogenase; HPLC,high-performance liquid chromatography; IBD, inflammatory boweldisease; IFN-g, interferon gamma; IL-1b, interleukin 1b; iNOS,inducible nitric oxide; LPS, lipopolysaccharide; MPO, myeloperoxi-dase; NO, nitric oxide; RT-PCR, reverse-transcription polymerasechain reaction; TNBS, trinitrobenzene sulfonic acid.
r 1999 by the American Gastroenterological Association0016-5085/99/$10.00
GASTROENTEROLOGY 1999;117:557–566

model of colitis. Having observed improved efficacy ofNO-mesalamine over mesalamine in this model, we thenproceeded to determine if there was a mechanistic basis tothe improved anti-inflammatory activity by comparingthe effects of the 2 drugs in terms of their ability to affectexpression of IL-1b and interferon (IFN)-g in the colonand their synthesis by isolated splenocytes. Also, effects ofNO-mesalamine and mesalamine on chemotaxin-inducedleukocyte adherence to the vascular endothelium wereexamined in vivo. These particular mechanistic studieswere selected because cytokines such as IL-1b and IFN-ghave been implicated in the pathogenesis of IBD13–15 andleukocyte adherence to the vascular endothelium is acritical event in inflammatory reactions.
Materials and Methods
AnimalsMale Wistar rats weighing 200–225 g were obtained
from Charles River Breeding Farms Ltd. (Montreal, Quebec,Canada; or Monza, Italy) and were housed in plastic cages. Therats were fed standard pellet chow and water ad libitum. Allexperimental protocols described were approved by the AnimalCare Committee of the University of Calgary and are inaccordance with the guidelines of the Canadian Council onAnimal Care.
Experimental Colitis
Colitis was induced by intracolonic instillation of thehapten trinitrobenzene sulfonic acid (TNBS; 60 mg/mL) in 0.5mL of 50% ethanol.16 The effects of mesalamine and NO-mesalamine on severity of colitis were examined using thefollowing treatment regimen. Groups of 12 rats each receivedNO-mesalamine (50 or 100 mg/kg), mesalamine (100 mg/kg),or vehicle intracolonically 1 hour after induction of colitis andat 12-hour intervals for 3 days thereafter. Longer treatmentperiods could not be evaluated because of a limited supply of
NO-mesalamine. A cannula similar to that used for instillationof TNBS into the colon was used for drug administration, withthe total volume delivered being 0.5 mL and the point ofdelivery approximately 9 cm proximal to the anus. All ratswere killed in a randomized manner on day 7 after TNBSadministration, and the severity of colitis was assessed (seebelow). Additional experiments were performed in which rats weretreated with sodium nitroprusside (an NO donor; n 5 6) orNCX-4016 (an NO-releasing aspirin derivative; n 5 10)7 at dosesequimolar to the 100-mg/kg dose of NO-mesalamine, to determineif other NO donors could produce beneficial effects in this model.
Assessment of Severity of Colitis
The colon was excised and pinned out, mucosal side up,on a wax platform. An observer unaware of the treatments therats had received assigned a score using criteria that have beenoutlined in detail previously.16 Briefly, the global score con-sisted of a colonic damage score for severity and extent ofulceration (from 0 to 10), summed with scores for the presenceor absence of diarrhea (0 or 1; diarrhea defined as loose orwatery stool) and adhesions (0, 1 for mild, or 2 for severe), andthe maximum thickness of the wall of the colon (in millime-ters). Tissue samples were then taken from a site of macroscopi-cally detectable inflammation (or a corresponding site in tissueswith no macroscopically detectable inflammation) for measure-ment of myeloperoxidase (MPO) activity, as an index of tissuegranulocyte numbers. The assay for MPO activity has beendescribed in detail previously.16 Other tissue samples weretaken from adjacent sites, fixed in neutral buffered formalin,and processed by routine techniques for histological evaluation.Slides were coded to prevent observer bias.
Nitrate and Nitrite Generation
To confirm that NO-mesalamine released NO in vivo,rats were given vehicle or NO-mesalamine at a dose of 50mg/kg orally (n 5 5/group). Three hours later the rats wereanesthetized with halothane. Blood was drawn from theinferior vena cava for measurement of serum nitrate and nitriteconcentrations, using a high-performance liquid chromatogra-phy (HPLC) assay described in detail previously.17 It was ourinitial intention to perform similar studies using sodiumnitroprusside at an equimolar dose, but rats receiving this drugorally did not survive for more than 1 hour because of profoundhypotension.
The rate of generation of NO from NO-mesalamine wascompared with that from sodium nitroprusside using an invitro system. Distal colon from 4 rats was excised, finelyminced with scissors, and suspended (167 mg/mL) in 10mmol/L sodium phosphate buffer (pH 7.4). The tissue was thenincubated at 37°C for 2 hours with either NO-mesalamine orsodium nitroprusside, both at concentrations of 10 mmol/L (anapproximation of the concentration to which the luminalsurface of the colon would have been exposed in the in vivostudies). Samples of the buffer were withdrawn at various timesof the incubation (5–120 minutes) for measurement of nitriteconcentration.17
Figure 1. Chemical structures of mesalamine (5-ASA) and NO-mesalamine (NCX-456).
558 WALLACE ET AL. GASTROENTEROLOGY Vol. 117, No. 3

Leukocyte Adherence In Vivo
The effects of mesalamine vs. NCX-456 on leukocyteadherence to the vascular endothelium were examined using anintravital microscopy preparation. Rats (n $ 6/group) werepretreated orally 1 hour before the start of these experimentswith vehicle, mesalamine, NO-mesalamine, or NCX-4016(each at 100 mg/kg). The rats were anesthetized with sodiumpentobarbital (60 mg/kg intraperitoneally). A midline abdomi-nal incision was made, and the rats were placed in a supineposition on an adjustable Plexiglas microscope stage. Asegment of the midjejunum was exteriorized through theabdominal incision, and the mesentery was prepared for in vivomicroscopic observation as previously described.18 Briefly, themesentery was draped over an optically clear viewing pedestalthat allows for transillumination of a 2-cm2 segment ofmesenteric tissue. The temperature of the pedestal was main-tained at 37°C with a constant temperature circulator. Theexposed bowel was covered with saline-soaked gauze to mini-mize tissue dehydration, and the mesentery was then super-fused with warm (37°C) bicarbonate-buffered saline, pH 7.4.The mesenteric microcirculation was observed using an intravi-tal microscope (Nikon optiphot-2; Melville, NY) with a 253objective lens (Leitz Wetzlar L25/0.35; Leipzig, Germany).Single unbranched mesenteric venules (25–40 µm in diameter)were selected for study. A videocamera mounted on themicroscope projected the image onto a monitor, and the imageswere recorded for playback analysis using a videocassetterecorder. A video time/date generator projected the time, date,and stopwatch function onto the monitor.
Images of the mesenteric microcirculation were recorded for5 minutes after a 15-minute equilibration period. This wasconsidered as time zero. The mesentery was then superfusedwith bicarbonate-buffered saline containing 5 µmol/L of thechemotactic peptide N-formyl-methionine-leucine-phenylala-nine (FMLP), and the images were recorded for 5 minutesbeginning 15, 30, 45, and 60 minutes after the beginning ofthe superfusion.
Venular diameter was measured on-line using a video caliper(model 908; IPM Ltd., San Diego, CA). Leukocyte adherencewas determined (on video playback) by an observer blind as tothe treatment the rats had received. A leukocyte was consideredadherent to the endothelium if it remained stationary for 30seconds or more.
Attempts were made to perform similar experiments usingrats with TNBS-induced colitis. However, the extensive adhe-sions between the inflamed colon and other abdominal tissuesprecluded the isolation of mesenteric vessels suitable for studyusing this apparatus.
Cytokine Release In Vitro: Splenocytes
The effects of mesalamine and NO-mesalamine onrelease of IL-1b and IFN-g were first examined using an assaythat involved the use of isolated rat splenocytes, as described indetail previously.19,20 Splenocytes (1 3 107/mL) were incubatedwith lipopolysaccharide (LPS) (100 ng/mL) alone or in combi-nation with mesalamine (0.1–10 µmol/L) or NO-mesalamine
(0.1–10 µmol/L) for 48 hours, and the release of IL-1b andIFN-g into the supernatant was measured using specificenzyme-linked immunosorbent assays (ELISA; Endogen, Mi-lan, Italy). Further experiments were performed in which theeffects of a selective inhibitor of caspase 1,20,21 acetyl Tyr-Val-Ala-Asp aldehyde (Ac-VAD.CHO; 10 µmol/L), were assessedby determining the effects of mesalamine and NO-mesalamine(each at 10 µmol/L) in the presence of this inhibitor.20–22
Cytokine Release In Vitro: Colon
The second assay used to determine the effects ofmesalamine and NO-mesalamine on release of IL-1b andIFN-g involved the use of inflamed colonic tissue from rat. Inthese studies, we also examined the effects of NCX-4016,another NO-releasing compound. Colitis was induced in 5 ratsby intracolonic administration of TNBS, as described above.On the third day after induction of colitis, the rats were killed,the inflamed region of the colon was excised and finely mincedwith scissors, and the tissue from each rat was divided into 4tubes. The tissue samples were weighed, then suspended (167mg/mL) in 10 mmol/L sodium phosphate buffer (pH 7.4). Thesamples were gently mixed and centrifuged (10,000g), and thesupernatant was discarded. The samples were then resuspendedin fresh 10 mmol/L phosphate buffer to which was addedvehicle, mesalamine, NO-mesalamine, or NCX-4016 (finalconcentration of each, 30 µmol/L). The samples were thenincubated in a shaking water bath (37°C) for 4 hours, afterwhich they were centrifuged (10,000g). The supernatants werefrozen at 220°C for subsequent measurement of IL-1b andIFN-g concentrations using specific ELISAs.
Caspase 1 Activity
Caspase 1, which is also known as interleukin-converting enzyme, is the enzyme responsible for cleaving theIL-1b precursor (pro–IL-1b) to its active form.23 To determineif NO-mesalamine produced inhibitory effects on cytokinerelease via inhibition of caspase 1 activity, its effects weremeasured using the rat splenocyte assay, as described previ-ously.21,22,24–26 This assay involved the measurement of theproteolytic cleavage of 7-amino-4-trifluoro-methylcoumarin(AFC)-conjugated YVAD-peptide as the substrate and AFC asthe standard. Inhibition of caspase 1 activity by NO-mesalamine could be due to the documented ability of NO tomodify protein function through S-nitrosation of thiolgroups.27–29 We tested this hypothesis by incubating spleno-cytes (in an ice-water bath) with NO-mesalamine (100 µmol/L)or sodium nitroprusside (100 µmol/L), alone or in combinationwith dithiothreitol (DTT; 10 µmol/L) for 1 hour. The DTTremoves any thiol-bound NO.20 Caspase 1 activity was thenassessed as described previously.22,22,24–26
Cytokine and Inducible NO SynthaseMessenger RNA Expression
Four groups of 5 rats each were given TNBS, asdescribed above, to induce colitis. A fifth group of 5 rats didnot receive TNBS and served as healthy controls. The rats that
September 1999 NO–RELEASING MESALAMINE 559

received TNBS were intracolonically treated, beginning 1 hourafter TNBS administration, with vehicle, mesalamine, NO-mesalamine, or NCX-4016 (each at 100 mg/kg). Drug orvehicle treatment was repeated at 12-hour intervals for 3 days,and the rats were killed 2 hours after the final (seventh) dose.Distal colon tissue was excised, messenger RNA (mRNA) wasextracted, and reverse-transcription polymerase chain reaction(RT-PCR) was performed, as described previously,30 to deter-mine the relative expression of mRNA for IL-1b, IFN-g, andinducible NO synthase (iNOS). Expression of mRNA forglyceraldehyde-3-phosphate dehydrogenase (GAPDH) was mea-sured as an internal control. The expression of mRNA for iNOSwas measured because this gene has previously been shown tobe markedly up-regulated in inflamed colon31 and because anNO-releasing derivative of flurbiprofen was previously shownto suppress iNOS expression.32
DNA amplification was conducted under the followingconditions: denaturation at 94°C for 1 minute, annealing at55°C for 30 seconds, and extension at 72°C for 1 minute. Toensure complete denaturation of the DNA with no backgroundpolymerase activity, Taq DNA polymerase was added to thePCR mixture during the hot start of cycle 1. Optimalcoamplification of each primer with GAPDH was determinedduring preliminary trials (data not shown). The IFN-g/GAPDH genes were coamplified for 35 and 20 cycles, theiNOS/GAPDH genes coamplified for 26 and 22 cycles, andIL-1b/GAPDH genes coamplified for 35 and 20 cycles,respectively.
After separation of the PCR products on a 1% agarose gelcontaining 10 µg of ethidium bromide, a picture was takenunder UV light using a Bio-Rad Gel Doc Video Documenta-tion System (Hercules, CA). Using Bio-Rad Quantity Onesoftware, densitometry analysis was completed and the geneproducts were normalized to control levels of GAPDH. TheiNOS and GAPDH products were amplified using primersdescribed previously.30 The IFN-g products were amplifiedusing the following primers: upstream, 58-ATG AGT GCTACA CGC CGC GTC TTG G-38; downstream, 58-GAG TTCATT GAC AGC TTT GTG CTG G-38.33 The IL-1b productswere amplified using the following primers: upstream, 58-GAA GCT GTG GCA GCT ACC TAT GTC T-38; down-stream, 58-CTC TGC TTG AGA GGT GGT GAT GTAC-38.34
Statistical Analysis
All data are expressed as the mean 6 SEM. Groups ofdata were compared using a one-way analysis of variancefollowed by a Student–Newman–Keuls test or with a Mann–Whitney U test (damage scores). The incidence of diarrhea wascompared across groups using the Fisher exact test. With allanalyses, an associated P value of ,5% was consideredsignificant.
Materials
TNBS was obtained from Fluka Chemica (Buchs,Switzerland). Mesalamine was generously provided by Axcan
Pharma Ltd. (St. Hilaire, Quebec, Canada). NO-mesalamine(NCX-456; 5-amino-2-hydroxybenzoic acid 4-(nitroxybutyl-ester) and NCX-4016 (2-acetoxybenzoate 2-[1-nitroxy-methyl]-phenyl ester) were synthesized by NicOx S.A. (Nice, France).ACF and Ac-YVAD-CHO were obtained from Alexis Corp.(Lausanne, Switzerland). The ELISA kits for measuring IL-1band IFN-g were obtained from Medicorp (Montreal, Quebec,Canada). All other chemicals, media, and sera were purchasedfrom Sigma Chemical Co. (St. Louis, MO), GIBCO (Paisley,England), or VWR Scientific (Edmonton, Alberta, Canada).
Results
Experimental Colitis
As described previously,16,35 intracolonic adminis-tration of TNBS resulted in extensive ulceration andinflammation of the distal colon of the rat. Macroscopi-cally visible damage typically extended 1–5 cm along thelongitudinal axis of the colon and was accompanied byextensive, transmural infiltration of granulocytes, macro-phages, and lymphocytes. As shown in Figure 2, themean colonic damage score in rats treated with vehicleover a 3-day period after induction of colitis was 7.7 60.7. Treatment over the same period with mesalamine(100 mg/kg) resulted in a significant decrease in thecolonic damage score (Figure 2). Treatment with NO-mesalamine (100 mg/kg) produced a highly significantdecrease in the colonic damage score (Figure 2). Reduc-tion of colonic damage with NO-mesalamine was signifi-cantly greater than that observed with mesalamine. Asshown in Table 1, a lower dose of NO-mesalamine (50mg/kg) was ineffective in this model.
Figure 2. Effects of mesalamine and NO-mesalamine (NCX-456) onseverity of colonic damage in rats killed 1 week after induction ofcolitis by intracolonic administration of TNBS. The rats were treatedintracolonically with the test drugs (each at 100 mg/kg) beginning 3hours after TNBS administration and at 12-hour intervals thereafterfor 3 days (total of 6 administrations). Each dot represents a rat; thehorizontal lines represent the mean damage scores for each group.
560 WALLACE ET AL. GASTROENTEROLOGY Vol. 117, No. 3
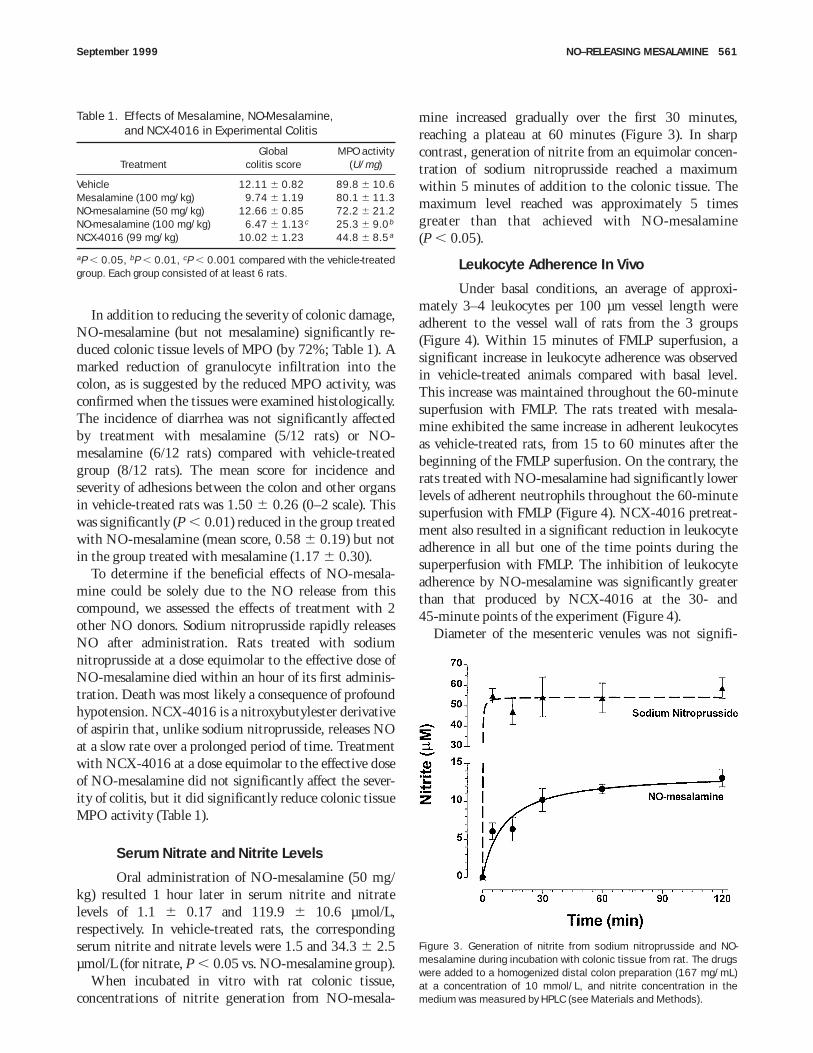
In addition to reducing the severity of colonic damage,NO-mesalamine (but not mesalamine) significantly re-duced colonic tissue levels of MPO (by 72%; Table 1). Amarked reduction of granulocyte infiltration into thecolon, as is suggested by the reduced MPO activity, wasconfirmed when the tissues were examined histologically.The incidence of diarrhea was not significantly affectedby treatment with mesalamine (5/12 rats) or NO-mesalamine (6/12 rats) compared with vehicle-treatedgroup (8/12 rats). The mean score for incidence andseverity of adhesions between the colon and other organsin vehicle-treated rats was 1.50 6 0.26 (0–2 scale). Thiswas significantly (P , 0.01) reduced in the group treatedwith NO-mesalamine (mean score, 0.58 6 0.19) but notin the group treated with mesalamine (1.17 6 0.30).
To determine if the beneficial effects of NO-mesala-mine could be solely due to the NO release from thiscompound, we assessed the effects of treatment with 2other NO donors. Sodium nitroprusside rapidly releasesNO after administration. Rats treated with sodiumnitroprusside at a dose equimolar to the effective dose ofNO-mesalamine died within an hour of its first adminis-tration. Death was most likely a consequence of profoundhypotension. NCX-4016 is a nitroxybutylester derivativeof aspirin that, unlike sodium nitroprusside, releases NOat a slow rate over a prolonged period of time. Treatmentwith NCX-4016 at a dose equimolar to the effective doseof NO-mesalamine did not significantly affect the sever-ity of colitis, but it did significantly reduce colonic tissueMPO activity (Table 1).
Serum Nitrate and Nitrite Levels
Oral administration of NO-mesalamine (50 mg/kg) resulted 1 hour later in serum nitrite and nitratelevels of 1.1 6 0.17 and 119.9 6 10.6 µmol/L,respectively. In vehicle-treated rats, the correspondingserum nitrite and nitrate levels were 1.5 and 34.3 6 2.5µmol/L (for nitrate, P , 0.05 vs. NO-mesalamine group).
When incubated in vitro with rat colonic tissue,concentrations of nitrite generation from NO-mesala-
mine increased gradually over the first 30 minutes,reaching a plateau at 60 minutes (Figure 3). In sharpcontrast, generation of nitrite from an equimolar concen-tration of sodium nitroprusside reached a maximumwithin 5 minutes of addition to the colonic tissue. Themaximum level reached was approximately 5 timesgreater than that achieved with NO-mesalamine(P , 0.05).
Leukocyte Adherence In Vivo
Under basal conditions, an average of approxi-mately 3–4 leukocytes per 100 µm vessel length wereadherent to the vessel wall of rats from the 3 groups(Figure 4). Within 15 minutes of FMLP superfusion, asignificant increase in leukocyte adherence was observedin vehicle-treated animals compared with basal level.This increase was maintained throughout the 60-minutesuperfusion with FMLP. The rats treated with mesala-mine exhibited the same increase in adherent leukocytesas vehicle-treated rats, from 15 to 60 minutes after thebeginning of the FMLP superfusion. On the contrary, therats treated with NO-mesalamine had significantly lowerlevels of adherent neutrophils throughout the 60-minutesuperfusion with FMLP (Figure 4). NCX-4016 pretreat-ment also resulted in a significant reduction in leukocyteadherence in all but one of the time points during thesuperperfusion with FMLP. The inhibition of leukocyteadherence by NO-mesalamine was significantly greaterthan that produced by NCX-4016 at the 30- and45-minute points of the experiment (Figure 4).
Diameter of the mesenteric venules was not signifi-
Table 1. Effects of Mesalamine, NO-Mesalamine,and NCX-4016 in Experimental Colitis
TreatmentGlobal
colitis scoreMPO activity
(U/mg)
Vehicle 12.11 6 0.82 89.8 6 10.6Mesalamine (100 mg/kg) 9.74 6 1.19 80.1 6 11.3NO-mesalamine (50 mg/kg) 12.66 6 0.85 72.2 6 21.2NO-mesalamine (100 mg/kg) 6.47 6 1.13c 25.3 6 9.0b
NCX-4016 (99 mg/kg) 10.02 6 1.23 44.8 6 8.5a
aP , 0.05, bP , 0.01, cP , 0.001 compared with the vehicle-treatedgroup. Each group consisted of at least 6 rats.
Figure 3. Generation of nitrite from sodium nitroprusside and NO-mesalamine during incubation with colonic tissue from rat. The drugswere added to a homogenized distal colon preparation (167 mg/mL)at a concentration of 10 mmol/L, and nitrite concentration in themedium was measured by HPLC (see Materials and Methods).
September 1999 NO–RELEASING MESALAMINE 561
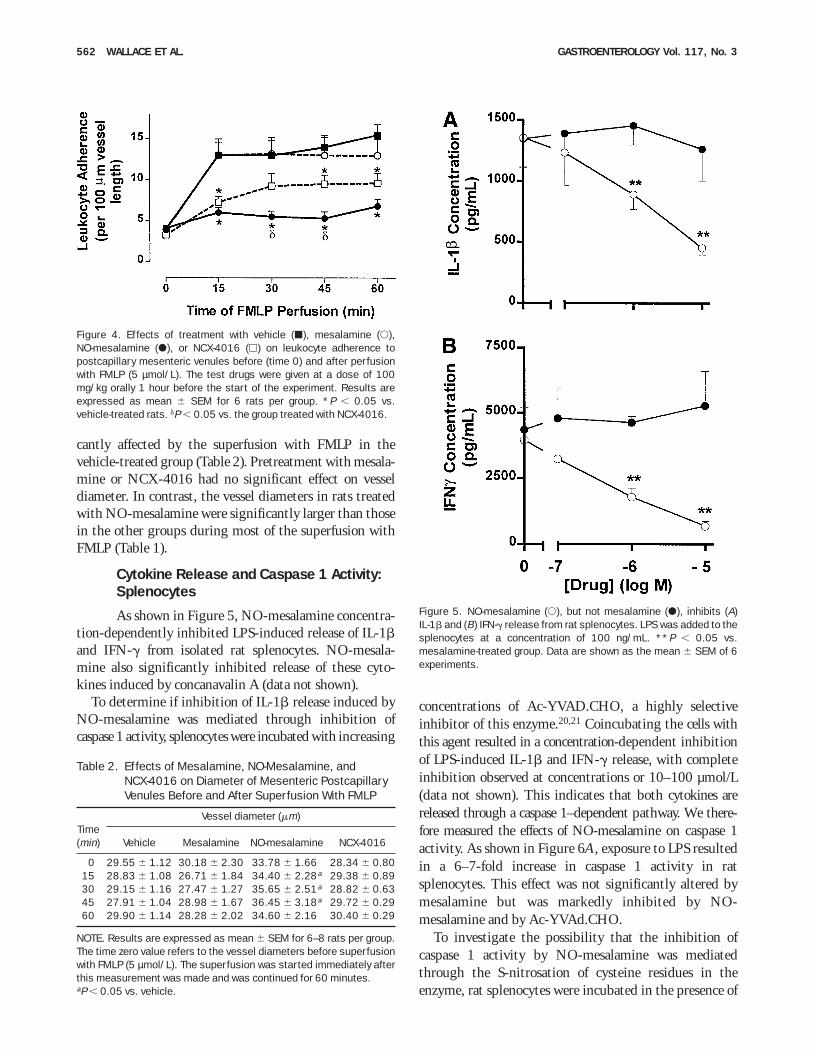
cantly affected by the superfusion with FMLP in thevehicle-treated group (Table 2). Pretreatment with mesala-mine or NCX-4016 had no significant effect on vesseldiameter. In contrast, the vessel diameters in rats treatedwith NO-mesalamine were significantly larger than thosein the other groups during most of the superfusion withFMLP (Table 1).
Cytokine Release and Caspase 1 Activity:Splenocytes
As shown in Figure 5, NO-mesalamine concentra-tion-dependently inhibited LPS-induced release of IL-1band IFN-g from isolated rat splenocytes. NO-mesala-mine also significantly inhibited release of these cyto-kines induced by concanavalin A (data not shown).
To determine if inhibition of IL-1b release induced byNO-mesalamine was mediated through inhibition ofcaspase 1 activity, splenocytes were incubated with increasing
concentrations of Ac-YVAD.CHO, a highly selectiveinhibitor of this enzyme.20,21 Coincubating the cells withthis agent resulted in a concentration-dependent inhibitionof LPS-induced IL-1b and IFN-g release, with completeinhibition observed at concentrations or 10–100 µmol/L(data not shown). This indicates that both cytokines arereleased through a caspase 1–dependent pathway. We there-fore measured the effects of NO-mesalamine on caspase 1activity. As shown in Figure 6A, exposure to LPS resultedin a 6–7-fold increase in caspase 1 activity in ratsplenocytes. This effect was not significantly altered bymesalamine but was markedly inhibited by NO-mesalamine and by Ac-YVAd.CHO.
To investigate the possibility that the inhibition ofcaspase 1 activity by NO-mesalamine was mediatedthrough the S-nitrosation of cysteine residues in theenzyme, rat splenocytes were incubated in the presence of
Figure 4. Effects of treatment with vehicle (j), mesalamine (s),NO-mesalamine (d), or NCX-4016 (h) on leukocyte adherence topostcapillary mesenteric venules before (time 0) and after perfusionwith FMLP (5 µmol/L). The test drugs were given at a dose of 100mg/kg orally 1 hour before the start of the experiment. Results areexpressed as mean 6 SEM for 6 rats per group. *P , 0.05 vs.vehicle-treated rats. dP , 0.05 vs. the group treated with NCX-4016.
Table 2. Effects of Mesalamine, NO-Mesalamine, andNCX-4016 on Diameter of Mesenteric PostcapillaryVenules Before and After Superfusion With FMLP
Time(min)
Vessel diameter (mm)
Vehicle Mesalamine NO-mesalamine NCX-4016
0 29.55 6 1.12 30.18 6 2.30 33.78 6 1.66 28.34 6 0.8015 28.83 6 1.08 26.71 6 1.84 34.40 6 2.28a 29.38 6 0.8930 29.15 6 1.16 27.47 6 1.27 35.65 6 2.51a 28.82 6 0.6345 27.91 6 1.04 28.98 6 1.67 36.45 6 3.18a 29.72 6 0.2960 29.90 6 1.14 28.28 6 2.02 34.60 6 2.16 30.40 6 0.29
NOTE. Results are expressed as mean 6 SEM for 6–8 rats per group.The time zero value refers to the vessel diameters before superfusionwith FMLP (5 µmol/L). The superfusion was started immediately afterthis measurement was made and was continued for 60 minutes.aP , 0.05 vs. vehicle.
Figure 5. NO-mesalamine (s), but not mesalamine (d), inhibits (A)IL-1b and (B) IFN-g release from rat splenocytes. LPS was added to thesplenocytes at a concentration of 100 ng/mL. **P , 0.05 vs.mesalamine-treated group. Data are shown as the mean 6 SEM of 6experiments.
562 WALLACE ET AL. GASTROENTEROLOGY Vol. 117, No. 3
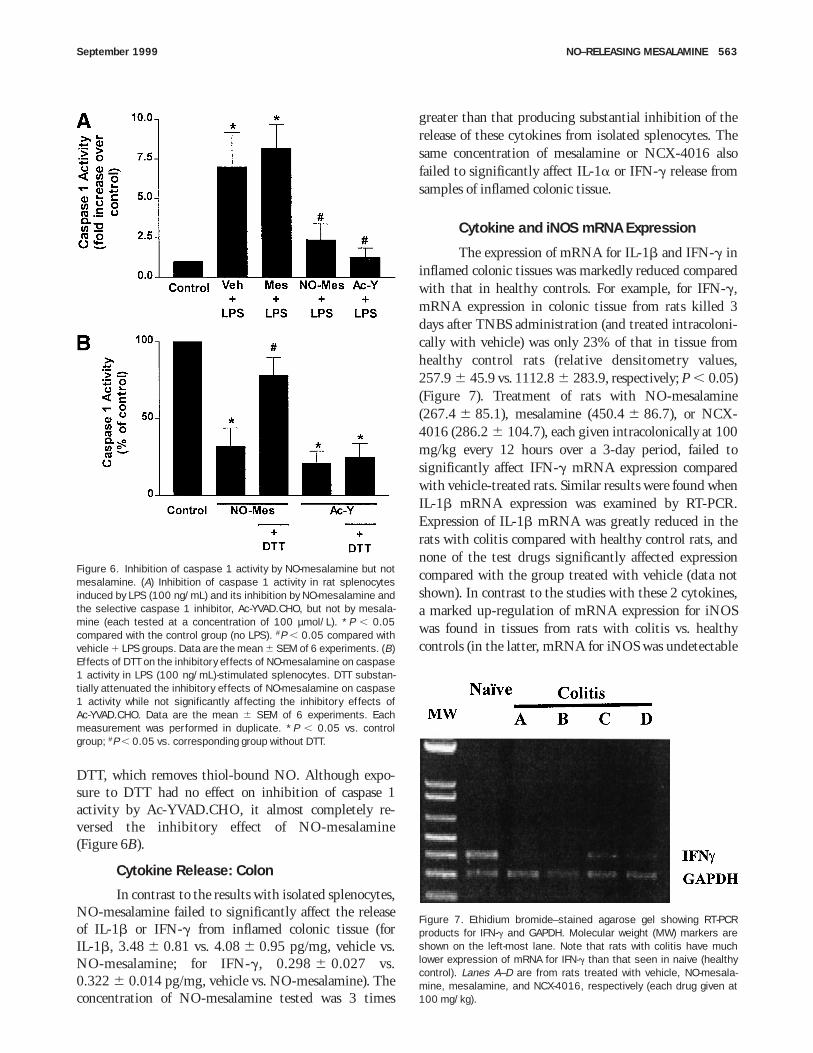
DTT, which removes thiol-bound NO. Although expo-sure to DTT had no effect on inhibition of caspase 1activity by Ac-YVAD.CHO, it almost completely re-versed the inhibitory effect of NO-mesalamine(Figure 6B).
Cytokine Release: Colon
In contrast to the results with isolated splenocytes,NO-mesalamine failed to significantly affect the releaseof IL-1b or IFN-g from inflamed colonic tissue (forIL-1b, 3.48 6 0.81 vs. 4.08 6 0.95 pg/mg, vehicle vs.NO-mesalamine; for IFN-g, 0.298 6 0.027 vs.0.322 6 0.014 pg/mg, vehicle vs. NO-mesalamine). Theconcentration of NO-mesalamine tested was 3 times
greater than that producing substantial inhibition of therelease of these cytokines from isolated splenocytes. Thesame concentration of mesalamine or NCX-4016 alsofailed to significantly affect IL-1a or IFN-g release fromsamples of inflamed colonic tissue.
Cytokine and iNOS mRNA Expression
The expression of mRNA for IL-1b and IFN-g ininflamed colonic tissues was markedly reduced comparedwith that in healthy controls. For example, for IFN-g,mRNA expression in colonic tissue from rats killed 3days after TNBS administration (and treated intracoloni-cally with vehicle) was only 23% of that in tissue fromhealthy control rats (relative densitometry values,257.9 6 45.9 vs. 1112.8 6 283.9, respectively; P , 0.05)(Figure 7). Treatment of rats with NO-mesalamine(267.4 6 85.1), mesalamine (450.4 6 86.7), or NCX-4016 (286.2 6 104.7), each given intracolonically at 100mg/kg every 12 hours over a 3-day period, failed tosignificantly affect IFN-g mRNA expression comparedwith vehicle-treated rats. Similar results were found whenIL-1b mRNA expression was examined by RT-PCR.Expression of IL-1b mRNA was greatly reduced in therats with colitis compared with healthy control rats, andnone of the test drugs significantly affected expressioncompared with the group treated with vehicle (data notshown). In contrast to the studies with these 2 cytokines,a marked up-regulation of mRNA expression for iNOSwas found in tissues from rats with colitis vs. healthycontrols (in the latter, mRNA for iNOS was undetectable
Figure 6. Inhibition of caspase 1 activity by NO-mesalamine but notmesalamine. (A) Inhibition of caspase 1 activity in rat splenocytesinduced by LPS (100 ng/mL) and its inhibition by NO-mesalamine andthe selective caspase 1 inhibitor, Ac-YVAD.CHO, but not by mesala-mine (each tested at a concentration of 100 µmol/L). *P , 0.05compared with the control group (no LPS). #P , 0.05 compared withvehicle 1 LPS groups. Data are the mean 6 SEM of 6 experiments. (B)Effects of DTT on the inhibitory effects of NO-mesalamine on caspase1 activity in LPS (100 ng/mL)-stimulated splenocytes. DTT substan-tially attenuated the inhibitory effects of NO-mesalamine on caspase1 activity while not significantly affecting the inhibitory effects ofAc-YVAD.CHO. Data are the mean 6 SEM of 6 experiments. Eachmeasurement was performed in duplicate. *P , 0.05 vs. controlgroup; #P , 0.05 vs. corresponding group without DTT.
Figure 7. Ethidium bromide–stained agarose gel showing RT-PCRproducts for IFN-g and GAPDH. Molecular weight (MW) markers areshown on the left-most lane. Note that rats with colitis have muchlower expression of mRNA for IFN-g than that seen in naive (healthycontrol). Lanes A–D are from rats treated with vehicle, NO-mesala-mine, mesalamine, and NCX-4016, respectively (each drug given at100 mg/kg).
September 1999 NO–RELEASING MESALAMINE 563

or only marginally detectable in the 5 rats studied).Treatment of rats with NO-mesalamine (relative density,728.4 6 84.6), mesalamine (859.3 6 39.3), or NCX-4016 (1035.9 6 144.6) failed to significantly affectiNOS mRNA expression compared with vehicle-treatedrats (715.2 6 133.2).
Discussion
Mesalamine is among the most commonly useddrugs to treat IBD, but it is only modestly effective orineffective in a significant proportion of patients. Theresults of this study show that addition of an NO-releasing moiety to mesalamine results in a markedincrease in its effectiveness in a rat model of colitis and ina substantial enhancement of its anti-inflammatory ac-tions. NO-mesalamine produced a significantly greaterreduction of colonic damage than mesalamine and pro-duced a significant reduction (72%) in granulocyteinfiltration to the colon (measured by MPO activity).Consistent with the latter, NO-mesalamine was able tosignificantly inhibit chemotaxin-induced leukocyte adher-ence, whereas the same dose of mesalamine had nosignificant effect. NO-mesalamine also exhibited supe-rior activity to mesalamine in terms of suppressingendotoxin-induced IL-1b and IFN-g release from iso-lated rat splenocytes.
The marked reduction of the severity of colitis in therats by NO-mesalamine was not solely attributable to theNO released by this compound. Administration of anequimolar dose of another NO-releasing drug (NCX-4016) had no significant effect in the colitis model. Thiscompound has been shown to release NO at rates similarto those reported in the present study for NO-mesalamine,7 to elevate cyclic guanosine monophosphatelevels in target cells,36 and to spare the gastric mucosa ofinjury normally associated with aspirin-like drugs.7,36 Inconsidering the mechanism through which NO-mesala-mine reduced the severity of colitis, it is important tobear in mind that a 100-mg/kg dose of this compounddelivers only about 50 mg/kg of mesalamine. Thus, theenhanced activity of NO-mesalamine vs. mesalamine inthe colitis model is even more significant when consid-ered on a molar basis. This also shows that NO-mesalamine did not act simply as a prodrug for mesala-mine.
The mechanism through which NO-mesalamine re-duced the severity of colonic injury may be related toseveral of its actions. As mentioned above, the ability ofNO-mesalamine, but not mesalamine, to inhibit leuko-cyte adherence to mesenteric postcapillary venules in vivosuggests that this could be the basis for enhanced
anti-inflammatory activity. The significant suppression ofcolonic MPO activity in the inflamed colon by NO-mesalamine, but not by mesalamine, is consistent withthis theory. Interestingly, NCX-4016 also significantlyinhibited leukocyte adherence induced by FMLP andreduced colonic MPO activity in the colitis model. Inboth cases, the effects of this drug were lesser inmagnitude to that of NO-mesalamine. The ability ofNO-mesalamine to increase blood flow, as was apparentby the significant vasodilation of mesenteric venules inrats treated with this drug but not in those treated withmesalamine or NCX-4016, may also contribute to re-duced tissue injury and/or accelerated repair.
The in vitro studies of rat splenocytes further showedthat NO-mesalamine has distinct biological propertiesfrom those of mesalamine. NO-mesalamine was found tomarkedly suppress cytokine release from splenocytes andto inhibit caspase 1 activity. However, it seems unlikelythat those actions contributed to the reduction of tissueinjury and granulocyte infiltration observed in the colitismodel. NO-mesalamine and mesalamine both failed tosignificantly affect the release of IL-1b or IFN-g frominflamed colonic tissue, and also did not affect theexpression of mRNA for these cytokines. Indeed, mRNAexpression for these cytokines was very low in theinflamed colon compared with healthy controls.
The ability of NO-mesalamine to inhibit leukocyteadherence more effectively than mesalamine or NCX-4016 is probably an important component of its mecha-nism of action, but there is at least one other attractivepossibility to explain the enhanced activity of thiscompound in experimental colitis. Grisham and Granger37
showed previously that mesalamine was able to scavengeoxygen- and nitrogen-derived free radicals at concentra-tions in the low micromolar range, whereas many of theother potential anti-inflammatory effects of mesalaminerequired much higher concentrations. A recent study bySandoval et al.4 showed that mesalamine was capable ofscavenging peroxynitrite, a potent oxidant that candirectly oxidize sulfhydryl groups as well as initiate lipidperoxidation.38,39 Peroxynitrite can also be degraded tozOH and NO2
z, which are highly toxic radicals.40 Peroxy-nitrite is produced in the inflamed colon,41 and intraco-lonic administration of peroxynitrite produces wide-spread colonic injury.42 Interestingly, delivery of exogenousNO could result in diminished peroxynitrite availabilitybecause of its ability to ‘‘scavenge’’ superoxide.43,44 Thatis, equimolar production of superoxide and NO can yieldperoxynitrite, but excess production of either of theseradicals can profoundly inhibit production of peroxyni-trite.45,46 In this regard, NO has been described to act asan antioxidant.41 Thus, NO-mesalamine can generate 2
564 WALLACE ET AL. GASTROENTEROLOGY Vol. 117, No. 3

agents (NO and mesalamine) capable of reducing theavailability of a substance, peroxynitrite, that may play akey role as a mediator of tissue injury in IBD.
In summary, the addition of an NO-releasing moietyto mesalamine results in a significant enhancement of itsbeneficial effects in experimental colitis. This compoundexhibits significantly greater activity than mesalamine interms of inhibiting chemotaxin-induced leukocyte adhe-sion to the vascular endothelium, suppressing granulo-cyte infiltration into a site of inflammation, and dilatingmesenteric venules. These actions could not be producedto the same extent by another, structurally related NOdonor (NCX-4016), suggesting that the 5-aminosalicylicacid moiety of NO-mesalamine contributed to its anti-inflammatory effects. Scavenging of peroxynitrite by theNO and mesalamine moieties of NO-mesalamine mayaccount for at least some of the beneficial effects of thiscompound in experimental colitis. Given the limitedefficacy of mesalamine in the treatment of IBD, assess-ment of the effects of NO-mesalamine in a clinical settingappears warranted.
References1. Sutherland LR, Martin F, Greer S, Robinson M, Greenberger N,
Saibil F, Martin T, Sparr J, Prokipchuk E, Borgen L. 5-Aminosali-cylic acid enema in the treatment of distal ulcerative colitis,proctosigmoiditis, and proctitis. Gastroenterology 1987;92:1894–1898.
2. Peskar BM, Dreyling KW, May B, Schaarschmidt K, Goebell H.Possible mode of action of 5-aminosalicylic acid. Dig Dis Sci1987;32:51S–56S.
3. Tamai H, Kachur JF, Grisham MB, Gaginella TS. Scavenging effectof 5-aminosalicylic acid on neutrophil-derived oxidants. Possiblecontribution to the mechanism of action in inflammatory boweldisease. Biochem Pharmacol 1991;41:1001–1006.
4. Sandoval M, Liu X, Mannick EE, Clark DA, Miller MJS. Peroxynitrite-induced apoptosis in human intestinal epithelial cells is attenu-ated by mesalamine. Gastroenterology 1997;113:1480–1488.
5. Mahida YR, Lamming CED, Gallagher A, Hawthorne AB, HawkeyCJ. 5-Aminosalicylic acid is a potent inhibitor of interleukin 1b
production in organ culture of colonic biopsy specimens frompatients with inflammatory bowel disease. Gut 1991;32:50–54.
6. Wallace JL, Reuter B, Cicala C, McKnight W, Grisham MB, Cirino G.Novel nonsteroidal anti-inflammatory drug derivatives with mark-edly reduced ulcerogenic properties in the rat. Gastroenterology1994;107:173–179.
7. Wallace JL, Muscara MN, McKnight W, Dicay M, Del Soldato P,Cirino G. In vivo anti-thrombotic effects of a nitric oxide–releasingaspirin derivative, NCX-4016. Thromb Res 1999;93:43–50.
8. Wallace JL, Reuter B, Cicala C, McKnight W, Grisham MB, Cirino G.A diclofenac derivative without ulcerogenic properties. Eur JPharmacol 1994;257:249–255.
9. Davies NM, Roseth AG, Appleyard CB, McKnight W, Del Soldato P,Calignano A, Cirino G, Wallace JL. NO-naproxen vs. naproxen:ulcerogenic, analgesic and anti-inflammatory effects. AlimentPharmacol Ther 1997;11:69–79.
10. Elliott SN, McKnight W, Cirino G, Wallace JL. A nitric oxide–releasing nonsteroidal anti-inflammatory drug accelerates gastriculcer healing in rats. Gastroenterology 1995;109:524–530.
11. Reuter BK, Cirino G, Wallace JL. Markedly reduced intestinaltoxicity of a diclofenac derivative. Life Sci 1994;55:PL1–PL8.
12. Konturek SJ, Brzozowski T, Majka J, Pytko-Polonczyk J, StachuraJ. Inhibition of nitric oxide synthase delays healing of chronicgastric ulcers. Eur J Pharmacol 1993;239:215–217.
13. Cominelli F, Nast CC, Clark BD, Schindler R, Llerena R, EysseleinVE, Thompson RC, Dinarello CA. Interleukin 1 (IL-1) gene expres-sion, synthesis, and effects of specific IL-1 receptor blockade inrabbit immune complex colitis. J Clin Invest 1990;86:972–980.
14. Cassini-Raggi V, Kam L, Chong YJT, Fiocchi C, Pizarro TT, CominelliF. Mucosal imbalance of IL-1 and IL-1 receptor antagonist ininflammatory bowel disease: a novel mechanism of chronicintestinal inflammation. J Immunol 1995;154:2434–2440.
15. Sartor RB. Pathogenetic and clinical relevance of cytokines ininflammatory bowel disease. Immunol Res 1991;10:456–471.
16. Wallace JL, Keenan CM. An orally active inhibitor of leukotrienesynthesis accelerates healing in a rat model of colitis. Am JPhysiol 1990;258:G527–G534.
17. Muscara MN, De Nucci G. Simultaneous determination of nitriteand nitrate anions in plasma, urine and cell culture supernatantsby high-performance liquid chromatography with post-columnreactions. J Chromatogr B Biomed Appl 1996;686:157–164.
18. McCafferty DM, Granger DN, Wallace JL. Indomethacin-inducedgastric injury and leukocyte adherence in arthritic versus healthyrats. Gastroenterology 1995;109:1173–1180.
19. Fiorucci S, Antonelli E, Migliorati G, Santucci L, Morelli O, FedericiB, Morelli A. TNFa-processing enzyme inhibitors prevent aspirin-induced TNFa release and protect against mucosal injury in rats.Aliment Pharmacol Ther 1998;12:1139–1154.
20. Fiorucci S, Santucci L, Federici B, Antonelli E, Distrutti E, MorelliO, Di Renzo G, Coata G, Cirino G, del Soldato P, Morelli A. Nitricoxide (NO)-releasing NSAIDs inhibit interleukin-1b convertingenzyme (ICE)-like cysteine proteases and protect endothelial cellsfrom apoptosis induced by TNFa. Aliment Pharmacol Ther 1999;13:421–435.
21. Thornberry NA, Bull HG, Calaycay JR, Chapman KT, Howard AD,Kostura MJ, Miller DK, Molineaux SM, Weidner JR, Aunins J,Elliston KO, Ayala JM, Casano FJ, Chin J, Ding GJF, Egger LA,Gaffney EP, Limjuco G, Palyha OC, Raju SM, Rolando AM, SalleyJP, Yamin TT, Lee TD, Shively JE, MacCross M, Mumford RA,Schmidt JA, Tocci MJ. A novel heterodimeric cysteine protease isrequired for interleukin-1b processing in monocytes. Nature1992;356:768–774.
22. Nicholson DW, Ali A, Thornberry NA, Vaillancourt JP, Ding CK,Gallant M, Gareau Y, Griffin PR, Labelle M, Lazebnik YA, MundayNA, Raju SM, Smulson ME, Yamin TT, Yu VL, Miller DK. Identifica-tion and inhibition of the ICE/CED-3 protease necessary formammalian apoptosis. Nature 1995;376:37–43.
23. Fletcher DS, Agarwal L, Chapman KT, Chin J, Egger LA, Luell S,MacIntyre DE, Peterson EP, Thornberry NA. A synthetic inhibitor ofinterleukin-1b converting enzyme prevents endotoxin-inducedinterleukin-1b production in vitro and in vivo. J Interferon CytokineRes 1995;15:243–248.
24. Thornberry NA. Interleukin-1 beta converting enzyme. MethodsEnzymol 1994;244:615–631.
25. Margolin N, Raybuck SA, Wilson KP, Chen W, Fox T, Gu Y,Livingston DJ. Substrate and inhibitor specificity of interleukin-1–converting enzyme and related caspases. J Biol Chem 1997;272:7223–7228.
26. Thornberry NA, Lazebnik Y. Caspases: enemies within. Science1998;281:1312–1316.
27. Dimmeler S, Haendeler J, Nehls M, Andreas M. Suppression ofapoptosis by nitric oxide via inhibition of interleukin-1 convertingenzyme (ICE)-like and cysteine protease protein (CPP)-32-likeproteases. J Exp Med 1997;185:601–607.
28. Young-Myeong K, Talanian RV, Billiar TR. Nitric oxide inhibitsapoptosis by preventing increases in caspase-3–like activity viatwo distinct mechanisms. J Biol Chem 1997;272:31138–31148.
29. Dimmeler S, Zeiher AM. Nitric oxide and apoptosis: another
September 1999 NO–RELEASING MESALAMINE 565

paradigm for the double-edged role of nitric oxide. Nitric Oxide:Biol Chem 1997;1:275–281.
30. Ferraz JG, Sharkey KA, Reuter BK, Asfaha S, Tigley AW, Brown ML,McKnight W, Wallace JL. Induction of cyclooxygenase 1 and 2 inthe rat stomach during endotoxemia: role in resistance todamage. Gastroenterology 1997;113:195–204.
31. Ribbons KA, Thompson JH, Liu XP, Pennline K, Clark DA, MillerMJS. Anti-inflammatory properties of interleukin-10 administra-tion in hapten-induced colitis. Eur J Pharmacol 1997;323:245–254.
32. Cirino G, Wheeler-Jones CPD, Wallace JL, Del Soldato P, BaydounAR. Inhibition of inducible nitric oxide synthase expression bynovel nonsteroidal anti-inflammatory derivatives with gastrointes-tinal-sparing properties. Br J Pharmacol 1996;117:1421–1426.
33. Morris GP, Beck PL, Herridge MS, Depew W, Szewczuk MR,Wallace JL. Hapten-induced model of chronic inflammation andulceration in the rat colon. Gastroenterology 1989;96:795–803.
34. Minami M, Kuraishi Y, Satoh M. Effects of kinic acid on messen-ger RNA levels of IL-1b, IL-6, TNFa, and LIF in the rat brain.Biochem Biophys Res Commun 1991;176:593–596.
35. Matsuda S, Uchikawa R, Yamada M, Arizono N. Cytokine mRNAexpression profiles in rats infected with intestinal nematodeNippostrongylus brasiliensis. Infect Immun 1995;63:4653–4660.
36. Wallace JL, McKnight W, Wilson TL, Del Soldato P, Cirino G.Reduction of shock-induced gastric damage by a nitric oxide–releasing aspirin derivative: role of neutrophils. Am J Physiol1997;273:G1246–G1251.
37. Grisham MB, Granger DN. 5-Aminosalicylic acid concentration inmucosal interstitium of cat small and large intestine. Dig Dis Sci1989;34:573–578.
38. Radi R, Beckman JS, Bush KM, Freeman BA. Peroxynitriteoxidation of sulfhydryls. The cytotoxic potential of superoxide andnitric oxide. J Biol Chem 1991;266:4244–4250.
39. Radi R, Beckman JS, Bush KM, Freeman BA. Peroxynitrite-induced membrane lipid peroxidation: the cytotoxic potential ofsuperoxide and nitric oxide. Arch Biochem Biophys 1991;288:481–487.
40. Pryor WA, Squadrito GL. The chemistry of peroxynitrite: a productfrom the reaction of nitric oxide with superoxide. Am J Physiol1995;26:L699–L722.
41. Miller MJS, Thompson JH, Zhang X-J, Sadowska-Krowicka H,Kakkis JL, Munshi UP, Sandoval M, Rossi JL, Eloby-Childress S,Beckman JS, Ye YZ, Rodi CP, Manning PM, Currie MG, Clark DA.Role of inducible nitric oxide synthase expression and peroxyni-trite formation in guinea pig ileitis. Gastroenterology 1995;109:1475–1483.
42. Rachmilewitz D, Stamler JS, Karmeli F, Mullins ME, Singel DJ,Loscalzo J, Xavier RJ, Podolsky DK. Peroxynitrite-induced ratcolitis—a new model of colonic inflammation. Gastroenterology1993;105:1681–1688.
43. Rubanyi GM, Ho EH, Cantor EH, Lumma WC, Botelho LHP.Cytoprotective function of nitric oxide: inactivation of superoxideradicals produced by human leukocytes. Biochem Biophys ResCommun 1991;181:1392–1397.
44. Rubbo H, Radi R, Trujillo M, Telleri R, Kalyanara B, Barnes S, KirkM, Freeman BA. Nitric oxide regulation of superoxide and peroxy-nitrite dependent lipid peroxidation: formation of novel nitrogencontaining oxidized lipid derivatives. J Biol Chem 1994;269:26066–26075.
45. Miles AM, Bohle DS, Glassbrenner PA, Hansert B, Wink DA,Grisham MB. Modulation of superoxide-dependent oxidation andhyroxylation reactions by nitric oxide. J Biol Chem 1996;271:40–47.
46. Wink DA, Cook JA, Kim SY, Vovovotz Y, Pacelli R, Krishna MC,Russo A, Mitchell JB, Jourd’heuil D, Miles AM, Grisham MB.Superoxide modulates the oxidation and nitrosation of thiols bynitric oxide–derived reactive intermediates. Chemical aspectsinvolved in the balance between oxidative and nitrosative stress.J Biol Chem 1997;272:11147–11151.
Received December 15, 1998. Accepted May 25, 1999.Address requests for reprints to: John L. Wallace, Ph.D., Depart-
ment of Pharmacology and Therapeutics, University of Calgary, 3330Hospital Drive Northwest, Calgary, Alberta, T2N 4N1, Canada.e-mail: [email protected]; fax: (403) 270-3353.
Supported by a grant from the Medical Research Council ofCanada (MRC). Dr. Wallace is an MRC Senior Scientist and anAlberta Heritage Foundation for Medical Research Senior Scientist.
The authors thank Barbara Federici for technical assistance.
566 WALLACE ET AL. GASTROENTEROLOGY Vol. 117, No. 3




















