
Embryonic Development of Caspian kutum, Rutilus frisii kutum
13
JOURNAL OF THE WORLD AQUACULTURE SOCIETY Vol. 41, No. 3 June, 2010 Embryonic Development of Caspian kutum, Rutilus frisii kutum Mahdieh Jafari, Mohd Salleh Kamarudin 1 , Che Roos Saad and Aziz Arshad Department of Aquaculture, Faculty of Agriculture, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia Shahrbanoo Oryan Department of Physiology, Faculty of Biology, Teacher Training University, Karaj, Tehran, PO Box 31979-37551, Iran Mohamad Hossein Tolouei Guilani Shahid Dr. Beheshti Sturgeon Propagation and Rearing Complex, Rasht, Guilan, PO Box 3117, Iran Abstract Rutilus frisii kutum is related to the Cyprinid family and an endemic fish of the Caspian Sea. This study was conducted to describe a complete embryonic development of kutum. Embryos were reared from zygote to larvae at 14–16 C. The embryonic period of this fish was divided into nine periods and 30 stages, from fertilization to hatching. Diagnostic structural characteristics were described for each stage. The main periods were zygote, cleavage, morula, blastula, gastrula, neurula, segmentation, pharyngula, and hatching. Hatching occurred at 8–9 d post-fertilization. Caspian kutum, Rutilus frisii kutum (Kamen- sky 1901), is a native, and commercial fish, of the Caspian Sea. This species is a migratory anadromous fish spawning in the rivers from March to April (Sharyati 1993; Razavi 1995), and on aquatic weeds and graveled and sandy substrates in rivers and lagoons (Abdoli 1999). It has three populations (one autumn and two spring) (Razavi 1997); a riverine/freshwater form exists in the South Caspian (Derzhavin 1934). Fish aged 5–6 yr compose the basic spawning stock. The absolute fecundity ranges from 27,000 to 280,000 eggs (Abdurakhmanov 1962). Decline in the stocks and catch of Caspian kutum was caused by overfishing, excessive catches of adults, increased pollu- tion, overexploitation of sands and sediments of the Caspian Sea, and the construction of 1 Corresponding author. bridges and dams that modified or blocked the natural spawning grounds (Azari Takami et al. 1979; Emadi 1979). For several years, start- ing from 1925, artificial breeding raised larvae for release in the 10 most important rivers. According to the Iranian Fisheries Organiza- tion’s report, more than 150 million juveniles are being released into the Caspian Sea (Ira- nian Fisheries Organization 2006). Spawning induction techniques using hormones provide high-quality gametes in good numbers for aqua- culture programs. Azari Takami et al. (1979), Woynarovich and Horv´ ath (1984), and Bart- ley and Rana (1998) detailed the technique for artificial spawning, incubation, and raising of fingerling Caspian kutum. At present, artificial propagation is the main method for maintaining and increasing the kutum population. Behzadi (1991) and Haghighi (2006) studied artificial production and embryonic stages of kutum, but there © Copyright by the World Aquaculture Society 2010 378
Transcript of Embryonic Development of Caspian kutum, Rutilus frisii kutum

JOURNAL OF THEWORLD AQUACULTURE SOCIETY
Vol. 41, No. 3June, 2010
Embryonic Development of Caspian kutum,Rutilus frisii kutum
Mahdieh Jafari, Mohd Salleh Kamarudin1, Che Roos Saad and Aziz Arshad
Department of Aquaculture, Faculty of Agriculture, University Putra Malaysia,43400 UPM Serdang, Selangor, Malaysia
Shahrbanoo Oryan
Department of Physiology, Faculty of Biology, Teacher Training University, Karaj, Tehran,PO Box 31979-37551, Iran
Mohamad Hossein Tolouei Guilani
Shahid Dr. Beheshti Sturgeon Propagation and Rearing Complex, Rasht, Guilan,PO Box 3117, Iran
AbstractRutilus frisii kutum is related to the Cyprinid family and an endemic fish of the Caspian Sea. This
study was conducted to describe a complete embryonic development of kutum. Embryos were rearedfrom zygote to larvae at 14–16 C. The embryonic period of this fish was divided into nine periodsand 30 stages, from fertilization to hatching. Diagnostic structural characteristics were described foreach stage. The main periods were zygote, cleavage, morula, blastula, gastrula, neurula, segmentation,pharyngula, and hatching. Hatching occurred at 8–9 d post-fertilization.
Caspian kutum, Rutilus frisii kutum (Kamen-sky 1901), is a native, and commercial fish, ofthe Caspian Sea. This species is a migratoryanadromous fish spawning in the rivers fromMarch to April (Sharyati 1993; Razavi 1995),and on aquatic weeds and graveled and sandysubstrates in rivers and lagoons (Abdoli 1999).It has three populations (one autumn and twospring) (Razavi 1997); a riverine/freshwaterform exists in the South Caspian (Derzhavin1934). Fish aged 5–6 yr compose the basicspawning stock. The absolute fecundity rangesfrom 27,000 to 280,000 eggs (Abdurakhmanov1962). Decline in the stocks and catch ofCaspian kutum was caused by overfishing,excessive catches of adults, increased pollu-tion, overexploitation of sands and sedimentsof the Caspian Sea, and the construction of
1 Corresponding author.
bridges and dams that modified or blocked thenatural spawning grounds (Azari Takami et al.1979; Emadi 1979). For several years, start-ing from 1925, artificial breeding raised larvaefor release in the 10 most important rivers.According to the Iranian Fisheries Organiza-tion’s report, more than 150 million juvenilesare being released into the Caspian Sea (Ira-nian Fisheries Organization 2006). Spawninginduction techniques using hormones providehigh-quality gametes in good numbers for aqua-culture programs. Azari Takami et al. (1979),Woynarovich and Horvath (1984), and Bart-ley and Rana (1998) detailed the technique forartificial spawning, incubation, and raising offingerling Caspian kutum.
At present, artificial propagation is themain method for maintaining and increasingthe kutum population. Behzadi (1991) andHaghighi (2006) studied artificial productionand embryonic stages of kutum, but there
© Copyright by the World Aquaculture Society 2010
378

EMBRYONIC DEVELOPMENT OF CASPIAN KUTUM 379
was no complete record of the developmentof embryonic substages. However, the above-mentioned studies did not provide a detaileddescription of the important characteristics ofthe kutum embryo. By knowing the commonembryonic development characteristics of thekutum, it was possible to distinguish habits,early life history, and probably embryo prob-lems in breeding and culture. The aim of thisstudy was to provide a detailed description ofkutum embryonic developmental stages.
Materials and Methods
Collection of Egg and Larvaeand Experimental Conditions
Eggs for this study were obtained fromone female and two male Caspian kutum,from broodstocks that were caught from HavyqRiver in Guilan province in the north of Iranand transferred to Shahid Dr. Beheshti Stur-geon Propagation and Rearing complex onMarch 4, 2007.
The broodstocks were kept for 2 d for adap-tation. Sperm were mixed with ovule obtainedafter pituitary extract injections; the first injec-tion 0.3 mg/kg and the second 3.5 mg/kg forfemale and one 2.0 mg/kg injection was admin-istered to the male, simultaneous to the secondinjection to the female. The gametes were gen-tly mixed for 5 min and washed with waterfor 45 min, resulting in the complete sep-aration of the eggs from each other. Theeggs were then incubated at 14–16 C inthree veis-type incubators (jar) (Bartley andRana 1998). The study period was 15 d. Thestudy was conducted based on two methods(Kimmel and Warger 1951): (1) using time ofdevelopment as a staging device and (2) usinganatomical features to determine and name eachstage.
Sampling
The eggs and embryos were studied atvarious developmental stages. A total of 30healthy eggs or larvae were sampled every hourfor 2 d post-fertilization and then six times perday. The chorion was removed, and the embryoand larvae were examined under a dissecting
microscope prior to fixation. The samples werefixed overnight in Bouin’s fixative and thentransferred to 10% buffered formalin. About 10of these samples were embedded in paraffin andsectioned at multiple levels or serially (5-μmthick). As the yolk was large, most of it wascut off as hatched larvae, so that they couldbe oriented more accurately. The sections werestained with hematoxylin and eosin.
Image Capture and Analysis
To obtain cell counts later in development,the embryo had to be separated from theyolk. The egg envelope was removed usingmicrodissecting scissors and forceps, and thecells were counted. A total of 30 embryos weremeasured for each stage. Total body length ofembryo or larvae was measured at 0.01 mmaccuracy; all measurements were performedusing the Nikon alphaphot YS microscope.Drawings were made from photographs takenusing camera (Nikon SMZ 800).
Statistical Analysis
The experiment was designed in three repli-cates and the results are given as mean ± SEM(n = 30 ) of each measured variable. The mor-phometric analysis employed independent t-testand chi-square test using SPSS 10.1. The levelof significant difference was set at P < 0.05.
Results
The characteristics of the embryonic stagesof R. frisii kutum are as follows, and aschematic model is presented in Figure 1. Also,there was no significant difference betweenincubation results.
Egg Characteristics
After matured ovules of Caspian kutum werereleased and fertilized, the germinal vesicle wasbroken and the fluid released. Water entered theegg membrane, expanding the membrane andcreating a space between the plasma and theegg membrane. The eggs were slightly demer-sal and drifted with the current. Before fertil-ization, the ovules were light yellow in color
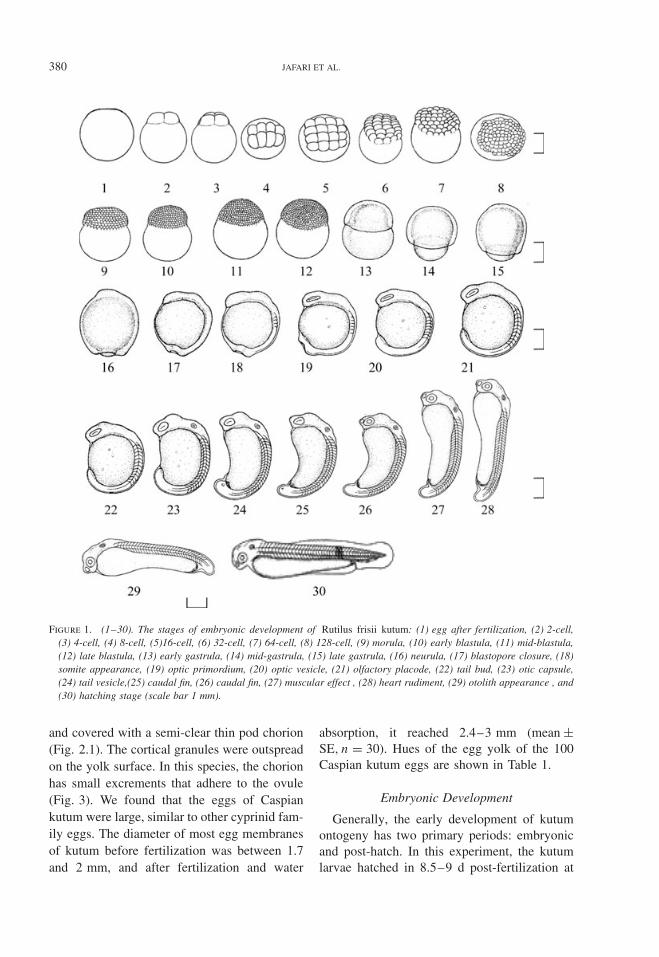
380 JAFARI ET AL.
Figure 1. (1–30). The stages of embryonic development of Rutilus frisii kutum: (1) egg after fertilization, (2) 2-cell,(3) 4-cell, (4) 8-cell, (5)16-cell, (6) 32-cell, (7) 64-cell, (8) 128-cell, (9) morula, (10) early blastula, (11) mid-blastula,(12) late blastula, (13) early gastrula, (14) mid-gastrula, (15) late gastrula, (16) neurula, (17) blastopore closure, (18)somite appearance, (19) optic primordium, (20) optic vesicle, (21) olfactory placode, (22) tail bud, (23) otic capsule,(24) tail vesicle,(25) caudal fin, (26) caudal fin, (27) muscular effect , (28) heart rudiment, (29) otolith appearance , and(30) hatching stage (scale bar 1 mm).
and covered with a semi-clear thin pod chorion(Fig. 2.1). The cortical granules were outspreadon the yolk surface. In this species, the chorionhas small excrements that adhere to the ovule(Fig. 3). We found that the eggs of Caspiankutum were large, similar to other cyprinid fam-ily eggs. The diameter of most egg membranesof kutum before fertilization was between 1.7and 2 mm, and after fertilization and water
absorption, it reached 2.4–3 mm (mean ±SE, n = 30). Hues of the egg yolk of the 100Caspian kutum eggs are shown in Table 1.
Embryonic Development
Generally, the early development of kutumontogeny has two primary periods: embryonicand post-hatch. In this experiment, the kutumlarvae hatched in 8.5–9 d post-fertilization at
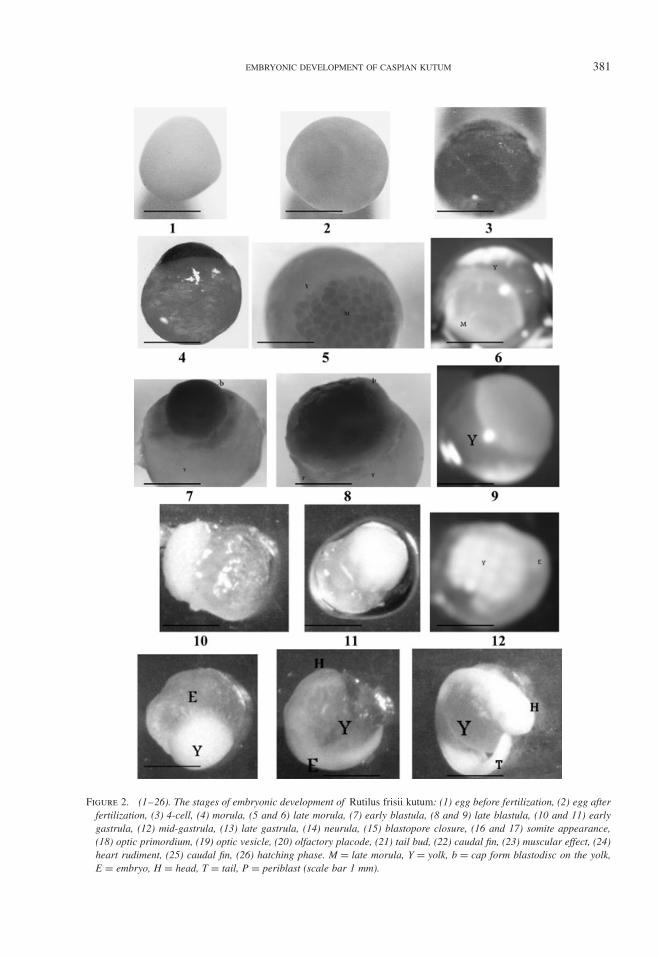
EMBRYONIC DEVELOPMENT OF CASPIAN KUTUM 381
Figure 2. (1–26). The stages of embryonic development of Rutilus frisii kutum: (1) egg before fertilization, (2) egg afterfertilization, (3) 4-cell, (4) morula, (5 and 6) late morula, (7) early blastula, (8 and 9) late blastula, (10 and 11) earlygastrula, (12) mid-gastrula, (13) late gastrula, (14) neurula, (15) blastopore closure, (16 and 17) somite appearance,(18) optic primordium, (19) optic vesicle, (20) olfactory placode, (21) tail bud, (22) caudal fin, (23) muscular effect, (24)heart rudiment, (25) caudal fin, (26) hatching phase. M = late morula, Y = yolk, b = cap form blastodisc on the yolk,E = embryo, H = head, T = tail, P = periblast (scale bar 1 mm).
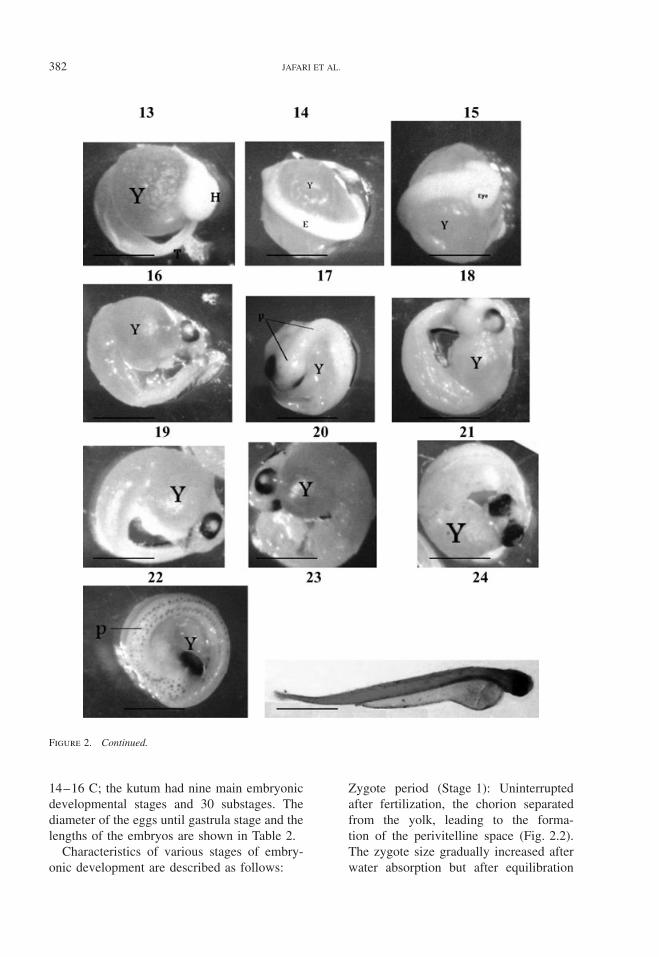
382 JAFARI ET AL.
Figure 2. Continued.
14–16 C; the kutum had nine main embryonicdevelopmental stages and 30 substages. Thediameter of the eggs until gastrula stage and thelengths of the embryos are shown in Table 2.
Characteristics of various stages of embry-onic development are described as follows:
Zygote period (Stage 1): Uninterruptedafter fertilization, the chorion separatedfrom the yolk, leading to the forma-tion of the perivitelline space (Fig. 2.2).The zygote size gradually increased afterwater absorption but after equilibration

EMBRYONIC DEVELOPMENT OF CASPIAN KUTUM 383
Figure 3. Photo micrograph of chorion in Rutilus frisii kutum egg. Y = yolk, C = chorion, PV = perivitelline, CE =chorion excrement.
Table 1. Hues of the egg yolk of Rutilus frisii kutum.
Color Number
Loquat yellow 24Apricot yellow 19Light green 40Butter yellow 10Cream colored 7Total 100
between perivitelline fluid and shell, waterabsorption stopped and no further changein zygote volume occurred.
Cleavage period (Stages 2–6): in thisperiod, five embryonic developmentalstages were observed.
Stage 2: the first cleavage occurred1 h after fertilization, and as inmedialecithal zygote, the cleavagewas of the meroblastic type andoccurred in the animal axis of thegerminal disc.Stage 3 (2 h 45 min–3 h 30 min):the eggs cleaved into four cells
(Fig. 2.3). Two blastomeres cleavedincompletely in a 2 × 2 array.Stage 4 (4 h 15 min–5 h 30 min):the eggs cleaved into eight cells andcut the blastodisc into a 2 × 4 arrayof blastomeres.Stage 5 (6 h 30 min–7 h 15 min):16 cells were arranged in a 4 ×4 array. The fourth set of cleavagealso occurred along two planes andproduced a 4 × 4 array of cells.Stage 6 (7 h 40 min–8 h 5 min):the cells were large and arrangedregularly; the cytoplasm diminishedand streamed toward the animalpole. Cleavages occurred along fourparallel planes. Frequently, 32 cellswere present in a 4 × 8 array.The cells became smaller and werearranged irregularly (Fig. 2.4).
Morula period (Stages 7, 8, and 9):
Early morula (Stage 7): this stageconsisted of 64 cells. Preblastincreased; cells of central blastodermtransformed into cylindrical forms,
384 JAFARI ET AL.
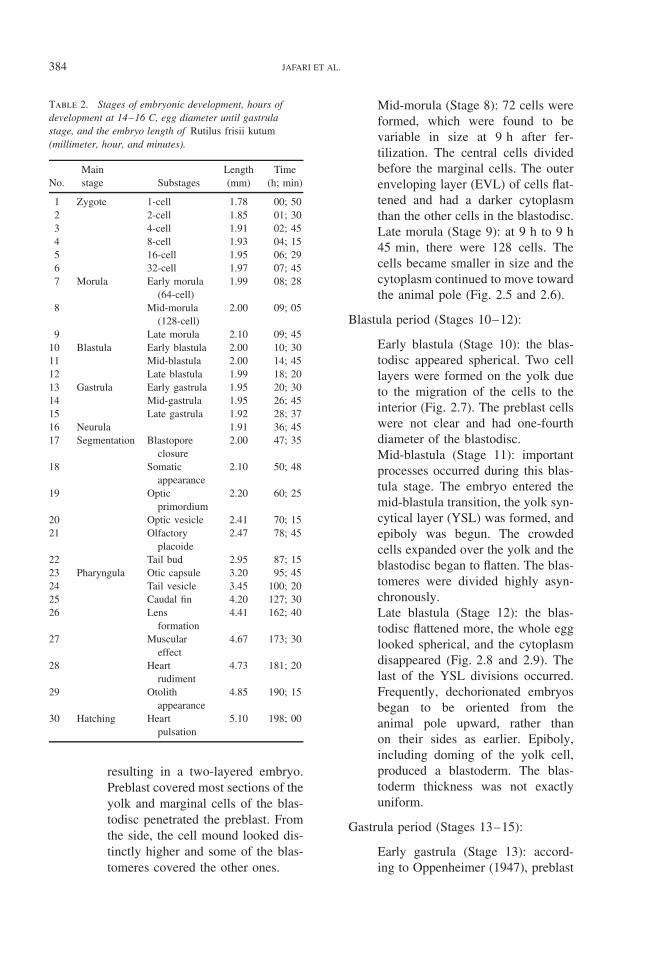
Table 2. Stages of embryonic development, hours ofdevelopment at 14–16 C, egg diameter until gastrulastage, and the embryo length of Rutilus frisii kutum(millimeter, hour, and minutes).
No.Mainstage Substages
Length(mm)
Time(h; min)
1 Zygote 1-cell 1.78 00; 502 2-cell 1.85 01; 303 4-cell 1.91 02; 454 8-cell 1.93 04; 155 16-cell 1.95 06; 296 32-cell 1.97 07; 457 Morula Early morula
(64-cell)1.99 08; 28
8 Mid-morula(128-cell)
2.00 09; 05
9 Late morula 2.10 09; 4510 Blastula Early blastula 2.00 10; 3011 Mid-blastula 2.00 14; 4512 Late blastula 1.99 18; 2013 Gastrula Early gastrula 1.95 20; 3014 Mid-gastrula 1.95 26; 4515 Late gastrula 1.92 28; 3716 Neurula 1.91 36; 4517 Segmentation Blastopore
closure2.00 47; 35
18 Somaticappearance
2.10 50; 48
19 Opticprimordium
2.20 60; 25
20 Optic vesicle 2.41 70; 1521 Olfactory
placoide2.47 78; 45
22 Tail bud 2.95 87; 1523 Pharyngula Otic capsule 3.20 95; 4524 Tail vesicle 3.45 100; 2025 Caudal fin 4.20 127; 3026 Lens
formation4.41 162; 40
27 Musculareffect
4.67 173; 30
28 Heartrudiment
4.73 181; 20
29 Otolithappearance
4.85 190; 15
30 Hatching Heartpulsation
5.10 198; 00
resulting in a two-layered embryo.Preblast covered most sections of theyolk and marginal cells of the blas-todisc penetrated the preblast. Fromthe side, the cell mound looked dis-tinctly higher and some of the blas-tomeres covered the other ones.
Mid-morula (Stage 8): 72 cells wereformed, which were found to bevariable in size at 9 h after fer-tilization. The central cells dividedbefore the marginal cells. The outerenveloping layer (EVL) of cells flat-tened and had a darker cytoplasmthan the other cells in the blastodisc.Late morula (Stage 9): at 9 h to 9 h45 min, there were 128 cells. Thecells became smaller in size and thecytoplasm continued to move towardthe animal pole (Fig. 2.5 and 2.6).
Blastula period (Stages 10–12):
Early blastula (Stage 10): the blas-todisc appeared spherical. Two celllayers were formed on the yolk dueto the migration of the cells to theinterior (Fig. 2.7). The preblast cellswere not clear and had one-fourthdiameter of the blastodisc.Mid-blastula (Stage 11): importantprocesses occurred during this blas-tula stage. The embryo entered themid-blastula transition, the yolk syn-cytical layer (YSL) was formed, andepiboly was begun. The crowdedcells expanded over the yolk and theblastodisc began to flatten. The blas-tomeres were divided highly asyn-chronously.Late blastula (Stage 12): the blas-todisc flattened more, the whole egglooked spherical, and the cytoplasmdisappeared (Fig. 2.8 and 2.9). Thelast of the YSL divisions occurred.Frequently, dechorionated embryosbegan to be oriented from theanimal pole upward, rather thanon their sides as earlier. Epiboly,including doming of the yolk cell,produced a blastoderm. The blas-toderm thickness was not exactlyuniform.
Gastrula period (Stages 13–15):
Early gastrula (Stage 13): accord-ing to Oppenheimer (1947), preblast

EMBRYONIC DEVELOPMENT OF CASPIAN KUTUM 385
increased and the blastodisc beganto invaginate, the germ ring formed,the dorsal lip appeared, and the blas-toderm covered about half of theyolk and was not in cap form. Ini-tial germ ring appeared dark andwide, and produced the germinaldisc (Fig. 2.10 and 2.11). This stagewas thoroughly different from theblastula stage. The dark section pro-duced an embryonic shield. Theperipheral blastomeres (the first-tierEVL cells) adhered to form the YSL(Kimmel et al. 1995). As in Caspiankutum, YSL formed a ring aroundthe blastodisc. The extra-embryonicectoderm at the center of the blas-toderm along with the preblast-covered yolk formed the yolk sac.The blastodisc covered about 10% ofthe yolk.Mid-gastrula (Stage 14): germ ringand margin of the blastoderm cov-ered about 10% of the yolk cell(Fig. 2.12). The germ ring beganto thicken and a thickened trian-gular region, the embryonic shield,extended from the germ ring towardthe animal pole. The YSL was nowcontinuous between the embryoniccells and the yolk. Epiboly was50–60% complete. Most of the blas-toderm was thin, except for theembryonic shield and the germ ring;therefore, distinguishing the differ-ent layers of the cells was difficult.Excluding the EVL and YSL, theembryonic shield consisted of aboutsix layers of cells separated into twolayers and the germ ring was abouttwo cell layers thick.Late gastrula (Stage 15): the embry-onic shield developed, the rudi-ment of the embryo became evident,and the blastoderm covered aboutfour-fifths of the yolk, which nowappeared pear-shaped. In three otherclutches, at about the same timeinterval, the blastoderm covered about
40% of the surface of the eggs andthe anterior–posterior axis of theembryo was visible as a thickeningin the center of the embryonic shield,extending from the animal pole tothe edge of the blastoderm.In the embryonic shield, Brachet’scleft separated the epiblast from thehypoblast and the central embryonicaxis extended into the yolk. TheEVL was so thin in the embryonicshield that it was difficult to see andthe YSL was continuous beneath theblastoderm. At the end of gastrula-tion, the embryonic shield was scro-biculate and announced the start ofthe neurulation stage. Epiboly was75% completed. The late gastrula-tion stage, nozzle of the blastoporewas still open (Fig. 2.13).
Neurula period (Stage 16):
The blastoderm was covered andthe embryo thickened substantially(Fig. 2.14). At this stage, the germi-nal axis was recognized clearly as asolid keel in the future skull section,and the opening of the nozzle of theblastopore gradually narrowed andclosed completely between 31 and37 h. Head section was absolutelydistinct and the cells were squeezedand appeared darker in the opticvesicle (Fig. 2.15).
Segmentation period (Stages 17–22):
Blastopore closure (Stage 17): thewhole embryo became thickened,especially at the end of the anteriorsection. In the posterior section, thetail bud extended ventrally from thegerm ring.Somite appearance (Stage 18): oneto three pairs of somites appeared atthe middle part of the embryo, thebrain rudiment thickened, and theblastopore was still visible (Fig. 2.16

386 JAFARI ET AL.
and 2.17). The first cleavage ofbrain occurred, distinguishing threemain sections of brain (front, middle,and back). Cells gathering aroundthe body axis was indicated by therise of the first pair of somite.Embryo was becoming lengthy andtail bud was sighted. Heart tubeswere formed and melanophoresincreased on the yolk. Epiboly wascompleted during this stage. Theeyes were visible but not pig-mented and there were a patch ofmelanophores on the egg surface oneach side of the embryo. At 48 h,Kupffer’s vesicle was still presentin embryos from one clutch. About15 somites were visible in sectionsand the formation of the first one tofour somites depended on whetherthe embryos were differentiated intomonomers with longitudinally ori-ented muscle fibers. Segmentationwas not yet complete and had notreached the posterior end of theembryo. The cells of the notochordstill had the “stack of pennies.”Optic primordium (Stage 19): theoptic primordium was long and ovalin shape. The lower edge appearedto have fine crenulations, the embryoembraced about five-sixths of theyolk, two to three oil dropletsappeared on the yolk cell, and thesomite number increased to four orfive pairs (Fig. 2.18).Optic vesicle (Stage 20): the opticprimordium enlarged slightly andbecame more visible, the embryoencircled most of the yolk, sothat the head and tail were inclose proximity, the somite num-ber was six or seven pairs, andthe notochords were clearly visi-ble (Fig. 2.19). In another clutch, at46 h myotomes had developed from17 to 18 somites, which extendedalong most of the embryo. The brainwas partitioned into the forebrain
(diencephalons and telencephalons),the dorsal (optic tectum) and ventralmidbrain, the cerebellum, and thehindbrain, which was divided intothe rhombomeres.Olfactory placode (Stage 21): theolfactory placode was barely vis-ible above the optic vesicle andthe somite number was nine pairs(Fig. 2.20). Besides pigmentation oneither side of the larva on the yolk,there were melanophores on the ven-tral side of the larva near the attach-ment to the yolk, as well as a fewnumber on the body dorsal to thisregion. The brain was enlarged andconsisted of three lobes, and therewas an ovoid otic vesicle with thick-ening of the walls. At this stage, theheart, which had been observed to becontracting rhythmically, was a sim-ple tube with a multilayered wall;lateral ventral was to the left sideof the pharynx. In kutum, the heartwas beating at 50 h and the tail wasfree of the yolk. The central part ofthe eyecup was more developed thanthe peripheral regions. In this centralregion, the layers of cells were con-nected by inner and outer plexiformlayers, and the photoreceptor nucleiwere aligned. Pigment granules werestarting to form in the outer layer ofthe optic cup. In this stage, brain’ssections were growing and expand-ing. A brain abdomen was going toshape up, which initially formed theoptical lobe, followed by the lensgrowth, and filled up the optic gob-let. Segmented nerves extended tothe myotomes, which were present atthe end of the tail. Two pronephrictubules were found to run from theglomeruli.Tail bud stage (Stage 22): the tailwas more prominent, the brain rudi-ment differentiated substantially, andthere were 10–14 pairs of somites(Fig. 2.21). The ectoderm was two

EMBRYONIC DEVELOPMENT OF CASPIAN KUTUM 387
cell layers thick, and unicellularhatching glands with prominentgranules that stained positively witheosin and orange G were present inthe ectoderm covering the head, tail,and surface of the yolk sac aroundthe embryo. Hatching glands werealso occasionally seen in the phar-ynx. Cement glands were presenton the head. There was a lumen inthe pharynx and esophagus, whichwere lined with cuboidal epithelium,and in the intestine, which was linedwith columnar epithelium. The neu-ral keel was enlarged anteriorly toform the brain rudiment, which wasnot subdivided yet, and optic primor-dial was present on either side of thisrudiment.
Pharyngula period (Stages 23–29):
Otic capsule (Stage 23): the otic cap-sule was clearly visible, the opticvesicle became enlarged and ellip-tical, and the somites numbered15–19 pairs. A cross-section of theposterior part of the brain revealedthe presence of an otic placode anda notochord ventral to the brain.The cells were beginning to elongatein the most anterior somites. Alongmost of the notochord, the cells hada typical “stack of pennies” appear-ance. The posterior part of the noto-chord, which could be seen in thetail bud dorsal to Kupffer’s vesicle,did not appearance yet. The Kupf-fer’s vesicle was lined ventrally bya depression in the YSL and theroof was lined with ciliated colum-nar cells.Tail vesicle (Stage 24): the tailextended outward and a vesicleappeared on the tail bud. Bloodislands that connected in yolk sacsection organized blood vessel lat-tice. The number of somites increasedto 19–20 pairs. The heart was a
thin-walled tube containing erythro-cytes. The liver was beginning toform between the yolk and gut. Theembryo had many melanophora onthe yolk. The first pigments formedon the retina; therefore, black coloreyes could be identified under astereomicroscope (after the removalof the chorion). Hearing cavitieswere completely formed. Formationof the brain abdomens, especially thefourth abdomen, was complete.Caudal fin (Stage 25): the embryoelongated further, the rudimentarycaudal fin formed, and the tail vesi-cle moved to the end of the tail.The yolk sac looked like a bean(Fig. 2.22). Pectoral fins formed onthe sides of the body. The rays ofdorsal fin and caudal fin were shapedup. The liver and intestine werespread. The liver covered the mainventral cavity.Lens formation (Stage 26): the eyelens formed. The eye consisted of anoptic cup with a ventral choroids fis-sure and a central lens primordium.Muscular effect (Stage 27): in thisstage, the yolk blood vessel latticecirculated in the back vessel andunder the tail vessel (Fig. 2.23). Theembryo could be seen in the eggwith chorion under a stereomicro-scope, because it had irregular andspasmodic movements and the eyepigments increased. Therefore, thisstage was termed eyed egg. Tailwas in the process of lengthening,passing above head slope form andcovered part of the brain (Fig. 3).Pectoral fins were very clear. Partof the spinal cord appeared distinctabove the notochord in the tailsection. Blood vessels gathered inthe caudal fin ray, and formation ofthe aorta was also complete. Smallglomerulus was observed under thebud caudal fin ray. Therefore, bloodcirculation was completed in this

388 JAFARI ET AL.
stage (Fig. 2.24). The lenses couldhardly be recognized because thelevel of pigmentation was increased.The head connected to the yolk.Embryo movements were increasedin the chorion, which was a veryclear cause of the presence of eyepigments in the chorion surface, andviewing of blood circulation in liversinus by unaided eye was made pos-sible by the removal of the chorion.The cells of the anterior part ofthe notochord were vacuolated andbeneath the notochord was the gut,which had a lumen. The posteriorgut did not appear to be ciliated andthe Kupffer’s vesicle was not seen.Heart rudiment (Stage 28): theembryo and yolk elongated further,the caudal fin extended, the embryowas of lash form, appearing occa-sionally side to side, and the heartappeared between the head and theyolk (Fig. 2.25). The heart was filledwith only a small amount of blood.Pectoral fins also had blood vessels.Gill net was not visible. Myotomes,notochord, and spinal cord appearedin the embryo tail.Otolith appearance (Stage 29): theembryo straightened but the headwas still curved around the ante-rior yolk sac. In ear otolith andsemicircular canals were grown andopercula were visible. Tail myomerswere visible in both kidneys. In thisstage, the eggs were about to behatched. Hatching time was depen-dent on water temperature. Some ofthe eggs that hatched had dark andpectoral fins that were growing. Inthis stage, embryo breathing withoxygen interchanged in yolk vessel.In our experiment, hatch occurred at14–16 C, 8–9 d after fertilization.The eyed egg was similar to freelarvae and the only difference wasthe presence of the egg shell.
Hatching period (Stage 30):
Hatching occurred in this stage andthe larvae were free of chorionshell (Fig. 2.26). The growth of finsand fin’s ray were recognized, andblood vessels penetrated them. Inmost kutum, hatching started 8–9d after fertilization and continuedfor about 24 h. Unhatched embryoswere coiled around a large yolk sac.There were patches of pigmentationon either side of the body posteriorto the pectoral fin, where the embryoattached to the yolk, and one or twopigments were spotted on the dorsalsurface of the embryo. The brain waswell developed, several gill archescould be distinguished around thepharynx, and the pronephric tubuleswere coiled anteriorly. There was asmall amount of lymphoid and ery-throid tissues around the pronephros.An oropharyngeal membrane cov-ered the mouth. The epithelial liningof the anterior part of the esopha-gus was beginning to stratify andposterior to this was a region ofcuboidal to columnar cells with palecytoplasm, which appeared to beciliated. A pneumatic duct with anarrow lumen attached the primor-dial swim bladder to the digestivetract. There were no goblet cells inthe esophagus or intestine, and therewas a small loop in the intestinearound the pancreas and the gall-bladder. A well-vascularized liverwas present and the ovoid gallblad-der was lined with cuboidal cells.Some exocrine pancreatic cells wereclosely associated with an insulin-positive endocrine islet. There werenumerous melanophores in the gutcavity. Primordial germ cells werefound near the pronephric tubules.Each gill arch contained a cartilagi-nous rod, which was present in theupper and lower jaws, and there was

EMBRYONIC DEVELOPMENT OF CASPIAN KUTUM 389
Figure 4. Photo micrograph of eye Rutilus frisii kutumembryo in hatching stage. The pigment layer (PL) of theeye extends to the lens and gill filaments (GF) form. Thebranchiostegal membrane (BM) covers the first pair ofgills, L, lens; PlL, plexiform layer, the retina (Re) of theeye is well developed (scale bar 0.1 mm).
a cartilaginous core in the pectoralfin. There were well-developed thy-mus glands on the dorsal wall of thepharyngeal cavity. There were tra-beculae in the ventricle of the heartand a thick-walled bulbus venosus.In the central part of the eyecup,the pigment cell layer was promi-nent and the nuclei of the rods andcones were aligned into two layers(Fig. 4).The otocyst was partially dividedinto the utriculus, sacculus, andlagena, each containing a singledense calcareous otolith. Olfactoryorgans were present. There wererudimentary cement glands on thehead, which were not as prominent.Neuromasts were also present onthe head. Numerous hatching glandswere present in the epidermis of thehead and tail, including the pectoralfins and the surface of the yolk sac
near the attachment of the embryo.A cross-section of the tail revealedthe presence of a hollow nerve cord,a vacuolated notochord, and caudalblood vessels. Posteriorly, the noto-chord turned upward, giving the taila heterocercal appearance. Usually,embryos first hatched from tail, leav-ing thin, ghost-like chorions behind.After hatching, the pectoral fins facedorsally. The egg membrane soft-ened, and the embryo was ready tohatch.
Discussion
A major goal of this study was to describethe acute embryonic stages in R. frisii kutum.Previous research on kutum larval organogene-sis was described by Behzadi (1991) and thenby Haghighi (2006), who investigated on arti-ficial breeding and embryo stages; however,they did not describe embryonic developmentexactly. This study describes the main embryostages according to the results of Behzadi andHaghighi. In this study, the staging system usedby Yi et al. (1966) for describing the embry-onic development in four species of cyprinidshas been utilized. We found that generally theembryonic development features presented inthis material were similar to that described ingrass carp by Yi et al. (1966). Although thereproductive habit of kutum was similar to thatof grass carp and black carp, and embryonicdevelopment was also similar, species-specificdifferences in morphology and time of organdifferentiation could be found by careful obser-vation of the various stages. Yolk-accumulatedcondition and oil drops were the main fac-tors differentiating the fish eggs. In kutum,yolk granules were very clear and perivitellinewas present around the yolk. The main differ-ences in the embryo characteristics had beendescribed in previous studies on grass, black,silver, and bighead carps; before the blastulastage, there was no obvious difference amongthe developing kutum eggs of other fish relatedto the cyprinid family. In the blastula stage,when the blastoderm margin covered half to

390 JAFARI ET AL.
two-thirds of the yolk surface, form of yolkwas different among other cyprinid species. Theentire yolk was spherical in shape. In the blasto-pore closure stage, when the embryo began toform, kutum head was smoothly curved. Theoptic rudiment did not appear until the olfactoryplacod stage. From the optic rudiment stage tothe otic capsule stage, oil droplets were foundon the yolk sac. From the otic capsule stage tothe heart pulsation stage, the head of the embryogradually developed a bump. At the otolithappearance stage, the yolk sac lengthened withthe development of the embryo, and the anteriorportion was wider than the posterior portion, butthe anterior one third of the yolk sac of grasscarp was spherical and the reminder was narrowand straight.
In black carp, the yolk sac gradually nar-rowed toward the vent. In silver carp, the fronttwo-thirds of the yolk sac was wide and theposterior one third was narrow (Yi et al. 1966).In Caspian kutum, the yolk sac was large andround similar to that of silver carp. Embry-onic development was similar to that of thesespecies, and specific differences in morphologyand time of organ differentiation could be madeby careful observation of the various stages.These result showed important changes in theembryo of Caspian kutum that could be veryimportant for achieving a successful hatch.
Acknowledgments
We are grateful to Mahmud Bahmani, deputyof International Sturgeon Research Institute, forhis assistance and to Soheil Shakouri for picturearrangement. Also, we are thankful to SafiehBehzadi as some of the original data werecollected by her as a part of her MSc thesis.
Literature CitedAbdoli, A. 1999. The Inland Water Fishes of Iran. Natural
and Wild Life Museum of Iran, Tehran, Iran. Pages198–200 (in Persian).
Abdurakhmanov, Yu. A. 1962. Fish of freshwater bodiesof Azerbaijan. Azerbaijan SSR AS. Pages 89–96.
Azari Takami, G. 1979. Fecundity of Rutilus frisii kutum.Journal of the Veterinary Faculty of the University ofTehran 35(1–2):66–78 (in Persian).
Bartley, M. D. and K.Rana. 1998. Iran promotes aqua-culture development. FAO Aquaculture Newsletter19:26–30.
Behzadi, S. 1991. Normal development of Rutilus frisiikutum. Master’s thesis. Islamic Azad University, Iran.
Derzhavin, A. N. 1934. Fresh water fishes of the southerncoast of the Caspian Sea. Trudy. Azerbaijankogo.Otdela zakavkazskogo filiala academia Nauk, SSSR,sector zoology 7:91–126 (in Russian).
Emadi, H. 1979. The state of the fishing and reproductionof the Kutum, Rutilus frisii kutum, in the Caspian seaof Iran. Journal of Ichthyology 19(4):151–154.
Haghighi, D. T. 2006. Embryonic development and nutri-tional requirements of kutum fry, Rutilus frisii kutum.PhD thesis. University Putra Malaysia.
Iranian Fisheries Organization. Accessed July 28, 2006at http.//www.Shilat.org.
Kimmel, C. B. and R. M. Warge. 1995. Tissue specificlinage of the zebra fish embryo in the gastrula. Science231:365–368.
Kimmel, C. B., W. W. Ballard, S. R. Kimmel, B. Ulman,and T. F. Schilling. 1995. Stages of embryonic devel-opment of the zebra fish. Developmental Dynamic203:253–310.
Oppenheimer, J. M. 1947. Organization of the teleostblastoderm. The Quarterly Review of Biology 22:105–118.
Razavi. S. B. 1995. Mahisefied, Rutilus frisii kutum. Ira-nian Fisheries Research Organization. 164 pp.
Razavi. S. B. 1997. Ecology of the Caspian Sea. IranianFishery Research Organization, Tehran, Iran. 90 pp.
Sharyati, A. 1993. Fishes of the Caspian Sea region (inPersian). Pages 77–79. Iranian Fisheries Company,Iran.
Woynarovich, E. and L. Horvath. 1980. The artificialpropagation of warm-water finfishes–a manual forextension. FAO Fish.Tech. Paper (201). 183 pp.
Yi, B. L., Z. T. Yu and Z. S. Liang. 1966. A comparativestudy of the embryonic development of grass carp,black carp, silver carp, big head and other fishes withdrifting eggs in the Yangtze River. Pages 37–53 in8th Symposium, Fisheries Research Committee of theWestern Pacific, Beijing. Science Press, Beijing.




















