
Effects of platelet‐derived growth factor‐AA and‐BB on embryonic cardiac development
10
Effects of Platelet-Derived Growth Factor-AA and -BB on Embryonic Cardiac Development ROBERT L. PRICE, 1 * STEPHEN T. HALEY, 2 TARA A. BULLARD, 1 EDIE C. GOLDSMITH, 1 DAVID G. SIMPSON, 3 THOMAS E. THIELEN, 4 MICHAEL J. YOST, 1 AND LOUIS TERRACIO 5 1 Department of Developmental Biology and Anatomy, School of Medicine, University of South Carolina, Columbia, South Carolina 2 Beckman Coulter Immunomics Operations, San Diego, California 3 Department of Anatomy, Medical College of Virginia, Virginia Commonwealth University, Richmond, Virginia 4 Medical University of South Carolina, Charleston, South Carolina 5 College of Dentistry, New York University, New York, New York ABSTRACT Several studies have shown that disruption of the normal expression patterns of platelet-derived growth factor (PDGF) ligands and receptors during development results in gross cardiac defects and embryonic or neonatal death. However, little is known about the specific role that PDGF plays in the differentiation of cardiac myocytes. In experiments complementing studies that utilized naturally-occurring Patch mice lacking the PDGFr , or knockout animals lacking a PDGF ligand or receptor, we used rat and mouse whole-embryo culture (WEC) techniques to increase the exposure of embryos to the PDGF-AA or -BB ligands. Following a 48-hr culture period, we analyzed heart growth and cardiac myocyte differentiation. Exposure of rat embryos to 50 ng/ml of PDGF-AA resulted in a 42% increase in total protein levels in the heart, but did not result in a significant increase in heart growth, as determined by measurements of the atrioventricular length and the left ventricular length and width. Exposure of embryos to 50 ng/ml of PDGF-BB resulted in a 77% increase in total protein levels and a significant (P 0.05) 8 –15% increase in the measured heart parameters. Although a comparison of control and PDGF-AA-treated embryos showed no increase in the overall size of the heart, confocal microscopy showed an increase in the size and number of myofibrillar bundles in the developing myocardium. In addition, transmission electron microscopy (TEM) revealed an increase in the presence of sarcomeres, indicating that myofibrils were more highly differentiated in these areas of the treated embryos. In PDGF-BB-treated embryos, the compact zone of the myocardium was thicker and, as shown by confocal microscopy and TEM, f-actin and well-developed sarcomeres were more prevalent, indicating that the myofibrils were more differen- tiated in the treated embryos than in the control embryos. These studies indicate that increased exposure of embryonic hearts to PDGF-AA or -BB increases the rate of myocardial development. Anat Rec Part A 272A:424 – 433, 2003. © 2003 Wiley-Liss, Inc. Key words: platelet-derived growth factor; cardiac development; myofibrillogenesis; confocal mi- croscopy; electron microscopy From several recent studies it has become evident that normal early embryonic heart development requires the coordinated temporal and spatial expression of a large number of growth factors and their specific receptors. The disruption of these specific patterns of expression by the use of transgenic animals, the addition of exogenous quan- tities of a growth factor, or the blocking of growth factor receptors frequently leads to developmental defects in the outflow tract, cardiac cushions, and myocardium that con- tribute to fetal or neonatal death (Price et al., 2001a). In studies utilizing the naturally-occurring Patch mutant (Morrison-Graham et al., 1992; Orr-Urtreger et al., 1992; Schatteman et al., 1992, 1995; Price et al., 2001b) and a series of knockout mice (Leve ´en et al., 1994; Kaminski et al., 2001; Soriano, 1994, 1997; Bostro ¨m et al., 1996; Frut- tiger et al., 1999; Kaminski et al., 2001) platelet-derived growth factor (PDGF) was identified as an essential sig- Grant sponsor: National Institutes of Health; Grant numbers: HD35253; HL58893; HL42249. *Correspondence to: Robert L. Price, Dept. of Developmental Biology and Anatomy, School of Medicine, University of South Carolina, Columbia, SC 29208. Fax: (803) 733-1533. E-mail: [email protected] Received 18 July 2002; Accepted 18 December 2002 DOI 10.1002/ar.a.10054 THE ANATOMICAL RECORD PART A 272A:424 – 433 (2003) © 2003 WILEY-LISS, INC.
Transcript of Effects of platelet‐derived growth factor‐AA and‐BB on embryonic cardiac development

Effects of Platelet-Derived GrowthFactor-AA and -BB on Embryonic
Cardiac DevelopmentROBERT L. PRICE,1* STEPHEN T. HALEY,2 TARA A. BULLARD,1
EDIE C. GOLDSMITH,1 DAVID G. SIMPSON,3 THOMAS E. THIELEN,4
MICHAEL J. YOST,1 AND LOUIS TERRACIO5
1Department of Developmental Biology and Anatomy, School of Medicine, Universityof South Carolina, Columbia, South Carolina
2Beckman Coulter Immunomics Operations, San Diego, California3Department of Anatomy, Medical College of Virginia, Virginia Commonwealth
University, Richmond, Virginia4Medical University of South Carolina, Charleston, South Carolina5College of Dentistry, New York University, New York, New York
ABSTRACTSeveral studies have shown that disruption of the normal expression patterns of platelet-derived growth factor
(PDGF) ligands and receptors during development results in gross cardiac defects and embryonic or neonatal death.However, little is known about the specific role that PDGF plays in the differentiation of cardiac myocytes. Inexperiments complementing studies that utilized naturally-occurring Patch mice lacking the PDGFr �, or knockoutanimals lacking a PDGF ligand or receptor, we used rat and mouse whole-embryo culture (WEC) techniques to increasethe exposure of embryos to the PDGF-AA or -BB ligands. Following a 48-hr culture period, we analyzed heart growthand cardiac myocyte differentiation. Exposure of rat embryos to 50 ng/ml of PDGF-AA resulted in a 42% increase intotal protein levels in the heart, but did not result in a significant increase in heart growth, as determined bymeasurements of the atrioventricular length and the left ventricular length and width. Exposure of embryos to 50 ng/mlof PDGF-BB resulted in a 77% increase in total protein levels and a significant (P � 0.05) 8–15% increase in themeasured heart parameters. Although a comparison of control and PDGF-AA-treated embryos showed no increase inthe overall size of the heart, confocal microscopy showed an increase in the size and number of myofibrillar bundles inthe developing myocardium. In addition, transmission electron microscopy (TEM) revealed an increase in the presenceof sarcomeres, indicating that myofibrils were more highly differentiated in these areas of the treated embryos. InPDGF-BB-treated embryos, the compact zone of the myocardium was thicker and, as shown by confocal microscopy andTEM, f-actin and well-developed sarcomeres were more prevalent, indicating that the myofibrils were more differen-tiated in the treated embryos than in the control embryos. These studies indicate that increased exposure of embryonichearts to PDGF-AA or -BB increases the rate of myocardial development. Anat Rec Part A 272A:424–433, 2003.© 2003 Wiley-Liss, Inc.
Key words: platelet-derived growth factor; cardiac development; myofibrillogenesis; confocal mi-croscopy; electron microscopy
From several recent studies it has become evident thatnormal early embryonic heart development requires thecoordinated temporal and spatial expression of a largenumber of growth factors and their specific receptors. Thedisruption of these specific patterns of expression by theuse of transgenic animals, the addition of exogenous quan-tities of a growth factor, or the blocking of growth factorreceptors frequently leads to developmental defects in theoutflow tract, cardiac cushions, and myocardium that con-tribute to fetal or neonatal death (Price et al., 2001a). Instudies utilizing the naturally-occurring Patch mutant(Morrison-Graham et al., 1992; Orr-Urtreger et al., 1992;Schatteman et al., 1992, 1995; Price et al., 2001b) and aseries of knockout mice (Leveen et al., 1994; Kaminski etal., 2001; Soriano, 1994, 1997; Bostrom et al., 1996; Frut-
tiger et al., 1999; Kaminski et al., 2001) platelet-derivedgrowth factor (PDGF) was identified as an essential sig-
Grant sponsor: National Institutes of Health; Grant numbers:HD35253; HL58893; HL42249.
*Correspondence to: Robert L. Price, Dept. of DevelopmentalBiology and Anatomy, School of Medicine, University of SouthCarolina, Columbia, SC 29208. Fax: (803) 733-1533.E-mail: [email protected]
Received 18 July 2002; Accepted 18 December 2002DOI 10.1002/ar.a.10054
THE ANATOMICAL RECORD PART A 272A:424–433 (2003)
© 2003 WILEY-LISS, INC.

naling molecule for several aspects of embryonic develop-ment.
The active forms of PDGF exist as dimers of the well-characterized PDGF-A and -B chains that can be ex-pressed as PDGF-AA and PDGF-BB homodimers, or asPDGF-AB heterodimers (Ataliotis and Mercola, 1997; Hel-din and Westermark, 1989), and the recently identifiedPDGF-CC (Ding et al., 2000; Kazlauskas, 2000; Li et al.,2000; Gilbertson et al., 2001) and PDGF-DD homodimers(Bergsten et al., 2001; LaRochelle et al., 2001). Two formsof receptors also exist. The PDGFr � is capable of bindingthe PDGF-A, -B, and -C isoforms (Gilbertson et al., 2001),while the PDGFr � binds the PDGF-B and -D isoforms(Bergsten et al., 2001). Both the PDGFr � and PDGFr �exhibit specific temporal and spatial expression patternsthat are related to cellular responses, including mitogen-esis, cell migration, chemotaxis, and differentiation (Atali-otis and Mercola, 1997; Betsholtz and Heldin, 1998).
The role of both the PDGF-A and -B isoforms and the �and � receptors in early cardiac development has been es-tablished through a series of studies with mice, in whichvarious components of the PDGF system were either missingor were blocked with specific antibodies. In these studies, theembryos usually died during fetal or early neonatal develop-ment, and exhibited severe cardiac dysmorphogenesis. InPatch mice, which lack PDGFr �, the hearts typically aresmall, with septal, valve, and outflow tract defects (Morri-son-Graham et al., 1992; Schatteman et al., 1995). Price etal. (2001b) used confocal and transmission electron micros-copy (TEM) to show that, in addition to the gross morpho-logical defects, the ventricular wall is thin and few sarco-meres are present in the myocytes. This indicates that themyocardium and myofibrils in Patch embryos are poorlydeveloped compared to littermate control embryos. In com-plementary studies, Schatteman et al. (1996) blocked thefunction of PDGF-A by the injection of blocking antibodiesinto the decidua of embryos at embryonic days (EDs) 8.5, 9.5,and 10.5. After an additional 48 hr of development, cardiacdefects similar to those observed in Patch mice were alsoseen in embryos injected with the blocking antibodies. How-ever, in recent studies of transgenic animals lacking thePDGFr-�, cardiac defects were limited to a dilated pericar-dium (Soriano, 1997).
Cardiac defects associated with deletion of the PDGF-Bchain differed significantly from those observed when ex-pression of the PDGF-A chain and PDGFr � were dis-rupted. The primary defects included enlarged hearts dueto ventricular and atrial dilation. Right ventricular de-fects included hypertrabeculation and areas where theventricular wall was a single cell layer thick. Left ventric-ular defects included dilated and congested pericardialblood vessels, indicating heart failure (Leveen et al.,1994). Cardiac development in mice lacking PDGFr �appeared normal, but this may have been a result ofcompensation by the PDGFr-� subunit (Soriano, 1994).
In the light of these observations on the fate of embryosin which specific components of the PDGF system wereblocked or absent, we used mouse and rat whole-embryoculture (WEC) systems to perform complementary studiesin which supplemental quantities of PDGF-AA and -BBwere added to the culture media to increase the exposureof the developing hearts to these growth factors. We thenanalyzed heart growth by two-dimensional measurementsof heart width and length, Western blot analysis for totalprotein content, and morphometric analysis of confocal
data sets. The organization of the myocardium and myo-fibrils in the developing hearts was also assessed by con-focal microscopy and TEM.
MATERIALS AND METHODSReagents
Unless otherwise noted, the reagents for WEC and im-munocytochemistry were purchased from Sigma ChemicalCo. (St. Louis, MO), and the reagents for electron micros-copy were obtained from Polysciences, Inc. (Warrington,PA).
AnimalsTimed-pregnant Sprague Dawley rats were obtained
from Harlan Laboratory (Indianapolis, IN). FounderC57BL/6 mice were also obtained from Harlan. A breedingcolony was subsequently established in the University ofSouth Carolina School of Medicine Animal Facility, andanimals were bred in-house. Day 0 of pregnancy was con-sidered to be midnight of the overnight breeding period.The animals were housed in a facility approved by theAmerican Association for Accreditation of Laboratory An-imal Care, and were provided food and water ad libitum.The University Institutional Animal Care and Use Com-mittee approved all of the experimental protocols.
WECThe methods for rat WEC were described in detail pre-
viously (Nakagawa et al., 1997; Price et al., 1997) and aresimilar to those originally described by New (1978) andmodified by Sadler et al. (1982). Rat WEC was performedwith embryos that were allowed to develop for 9.5 or 10.5days in vivo and an additional 48 hr in culture (9.5 � 48 hror 10.5 � 48 hr). Mouse WEC was performed with em-bryos that were allowed to develop for 8.5 or 9.5 days invivo and an additional 48 hr in culture. The 24-hr differ-ence in gestation times allowed comparison of similarlystaged animals, since mouse development occurs slightlyfaster than rat development during gestation. The meth-ods for mouse WEC were identical to our published meth-ods for rat WEC, with the exception that the O2 concen-tration was increased from 5% to 20% when ED 9.5animals were used.
For each experimental run, embryos were collected fromseveral mothers at the appropriate stage of gestation andpooled. They were then cultured for 2 hr to ensure thatthey had survived the isolation process and initial transferto the culture system. The number of mothers used foreach culture run varied depending on the size of individ-ual litters and the availability of culture bottles, but wastypically four to five dams per experiment. Following theinitial 2-hr culture period, the embryos were examinedand an assessment of age and developmental stage wasmade according to guidelines established by Brown andFabro (1981). This was necessary because in many casesembryos from a single litter are at significantly differentstages of development (Fujinaga and Baden, 1992). Fromthis group of pooled embryos, age- and developmentally-matched embryos were selected for culture. Those em-bryos that were considered either too young or too oldaccording to the Brown and Fabro system of scoring, or didnot appear healthy after this initial culture period, werediscarded. Embryos selected for further experimentationwere transferred with intact visceral yolk sacs into culture
425PLATELET-DERIVED GROWTH FACTOR-AA AND -BB
bottles (two to four embryos per bottle) containing 4.0 mlof rat serum (Harlan Bioproducts for Science, Cincinnati,OH) and 1.0 ml of Tyrode’s solution (0.137 M NaCl, 2.7mM KCl, 3.6 mM NaH2PO4H2O, 27 mM d-glucose, 3.6mM NaHCO3, 1.36 M CaCl2, 0.5 mM MgCl2, and 0.13 mMstreptomycin). For treated embryos, either PDGF-AA orPDGF-BB was added to the culture media, at a concen-tration of 50 ng/ml. Control cultures contained no exoge-nous PDGF. After the embryos were cultured for 24 hr, themedia were replaced with fresh media containing PDGFat the above concentrations. After 48 hr in culture, theembryos were removed from the culture bottles and pre-pared for microscopy and protein accumulation studies asdescribed below. Table 1 presents the number of cultureruns and embryos included in the analysis for each agegroup and experimental condition.
Assessment of Cardiac Growth FollowingExposure to PDGF in WEC
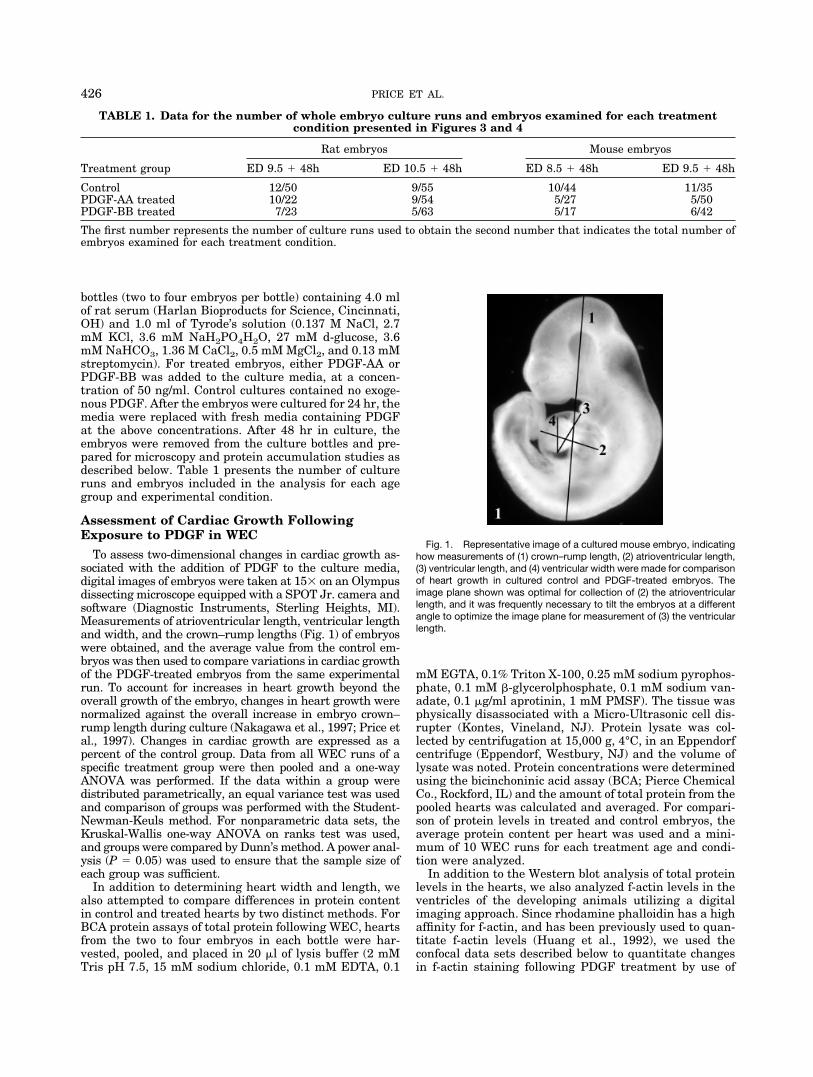
To assess two-dimensional changes in cardiac growth as-sociated with the addition of PDGF to the culture media,digital images of embryos were taken at 15� on an Olympusdissecting microscope equipped with a SPOT Jr. camera andsoftware (Diagnostic Instruments, Sterling Heights, MI).Measurements of atrioventricular length, ventricular lengthand width, and the crown–rump lengths (Fig. 1) of embryoswere obtained, and the average value from the control em-bryos was then used to compare variations in cardiac growthof the PDGF-treated embryos from the same experimentalrun. To account for increases in heart growth beyond theoverall growth of the embryo, changes in heart growth werenormalized against the overall increase in embryo crown–rump length during culture (Nakagawa et al., 1997; Price etal., 1997). Changes in cardiac growth are expressed as apercent of the control group. Data from all WEC runs of aspecific treatment group were then pooled and a one-wayANOVA was performed. If the data within a group weredistributed parametrically, an equal variance test was usedand comparison of groups was performed with the Student-Newman-Keuls method. For nonparametric data sets, theKruskal-Wallis one-way ANOVA on ranks test was used,and groups were compared by Dunn’s method. A power anal-ysis (P � 0.05) was used to ensure that the sample size ofeach group was sufficient.
In addition to determining heart width and length, wealso attempted to compare differences in protein contentin control and treated hearts by two distinct methods. ForBCA protein assays of total protein following WEC, heartsfrom the two to four embryos in each bottle were har-vested, pooled, and placed in 20 �l of lysis buffer (2 mMTris pH 7.5, 15 mM sodium chloride, 0.1 mM EDTA, 0.1
mM EGTA, 0.1% Triton X-100, 0.25 mM sodium pyrophos-phate, 0.1 mM �-glycerolphosphate, 0.1 mM sodium van-adate, 0.1 �g/ml aprotinin, 1 mM PMSF). The tissue wasphysically disassociated with a Micro-Ultrasonic cell dis-rupter (Kontes, Vineland, NJ). Protein lysate was col-lected by centrifugation at 15,000 g, 4°C, in an Eppendorfcentrifuge (Eppendorf, Westbury, NJ) and the volume oflysate was noted. Protein concentrations were determinedusing the bicinchoninic acid assay (BCA; Pierce ChemicalCo., Rockford, IL) and the amount of total protein from thepooled hearts was calculated and averaged. For compari-son of protein levels in treated and control embryos, theaverage protein content per heart was used and a mini-mum of 10 WEC runs for each treatment age and condi-tion were analyzed.
In addition to the Western blot analysis of total proteinlevels in the hearts, we also analyzed f-actin levels in theventricles of the developing animals utilizing a digitalimaging approach. Since rhodamine phalloidin has a highaffinity for f-actin, and has been previously used to quan-titate f-actin levels (Huang et al., 1992), we used theconfocal data sets described below to quantitate changesin f-actin staining following PDGF treatment by use of
Fig. 1. Representative image of a cultured mouse embryo, indicatinghow measurements of (1) crown–rump length, (2) atrioventricular length,(3) ventricular length, and (4) ventricular width were made for comparisonof heart growth in cultured control and PDGF-treated embryos. Theimage plane shown was optimal for collection of (2) the atrioventricularlength, and it was frequently necessary to tilt the embryos at a differentangle to optimize the image plane for measurement of (3) the ventricularlength.
TABLE 1. Data for the number of whole embryo culture runs and embryos examined for each treatmentcondition presented in Figures 3 and 4
Treatment group
Rat embryos Mouse embryos
ED 9.5 � 48h ED 10.5 � 48h ED 8.5 � 48h ED 9.5 � 48h
Control 12/50 9/55 10/44 11/35PDGF-AA treated 10/22 9/54 5/27 5/50PDGF-BB treated 7/23 5/63 5/17 6/42
The first number represents the number of culture runs used to obtain the second number that indicates the total number ofembryos examined for each treatment condition.
426 PRICE ET AL.

pixel intensity changes associated with rhodamine phal-loidin staining. Three-dimensional projections of data setscreated using VoxBlast were imported into Image ProPlus (version 4.1; Media Cybernetics, Silver Spring, MD)and a 5 � 5 line mask was applied to the image. Pixelintensities of each pixel along the lines in the mask werethen measured and exported to a Microscoft Excel spread-sheet for statistical analysis using ANOVA and a Tukeypost-hoc test. For the comparison of pixel intensity data,all specimens and images were treated identically.
Confocal and Electron Microscopy Analysis ofPDGF Receptor Localization and MyocardialOrganization
To enable visualization of the general heart morphol-ogy, the embryos were rinsed in Tyrode’s solution with 50mM KCl to relax the heart, and were then preserved in 2%paraformaldehyde (Price et al., 1996, 1997). To facilitatepenetration of rhodamine phalloidin and imaging of largerhearts, fixed embryos older than ED 10 were encased in13% polyacrylamide gel using a modification of the proce-dure described by Hansen and Dryer (1981). After fixa-tion, the embryos were rinsed in PBS plus 0.02% azide(PBS-A) and placed in polyacrylamide gel that was imme-diately polymerized with 2% ammonium persulfate. Thepolyacrylamide gel-encased embryos were sectioned at200 �m with an Oxford vibratome.
Whole embryos or acrylamide sections of embryos werethen rinsed in PBS-A and stained in a 1:20 dilution ofrhodamine phalloidin (Molecular Probes, Eugene, OR) inPBS-A overnight at 4°C. Rhodamine phalloidin is a spe-cific stain for f-actin, which is a primary component of thedeveloping cardiac contractile apparatus. At the concen-trations used, the fluorescence enhancement of rhodaminephalloidin is also linearly related to f-actin concentrations,and can therefore be used for f-actin quantitation (Huanget al., 1992). To determine the organization of the actin inthe myocardium of cultured control and PDGF-treatedembryos, we collected confocal data sets through 100-�-thick vibratome sections at 1-�m intervals with a Nikon60� Plan Apo lens (NA 1.4) and an iris setting of 1.1. Datasets were then reconstructed with Voxblast volume visu-alization software (version 1.44; VayTek, Inc., Fairfield,IA) on a Dell Precision Workstation 220. Volumes wererendered using the single volume mode, and the opacityvalues were adjusted to facilitate viewing of the myofi-brils. While several areas of the heart were examined bythis protocol, the data presented represent comparisons ofpixel intensities in the developing left ventricle of controland treated embryos, and all image sets used for compar-ison of the control and treated embryos were collected andtreated identically.
For labeling of the PDGF receptors, either whole em-bryos or acrylamide sections of embryos were incubated ina 1:20 dilution of a polyclonal antibody to the � or �receptor (Santa Cruz Biotechnology, CA) for 24 hr at 4°C,rinsed in PBS-A, and stained overnight at 4°C in a 1:50dilution of Cy2 (Jackson Immunochemical Research Lab-oratories, West Grove, PA). For double labeling, the em-bryos were first labeled for the PDGF receptors and thenby rhodamine phalloidin. Controls consisted of either non-immune IgG or PBS buffer substituted for the primaryantibody. All embryos and sections were examined using aBioRad MRC 1024ES confocal scanning laser microscope.
To determine whether increased organization of individ-ual myofibrils was associated with the observed changesin gross organization and f-actin levels in the myocar-dium, we also performed TEM on three hearts from eachgroup. To ensure that similar areas of control and treatedhearts were being examined, individual 2-�m-thick sec-tions were removed from epoxy-embedded hearts and im-aged with toluidine blue staining until similar regions ofthe embedded hearts were found. Thin sections were thencut on a diamond knife, stained with Reynold’s lead citrateand 2% aqueous uranyl acetate, and examined at 120kV ina JEOL 200CX TEM.
RESULTSLocalization of PDGF-� and -� Receptors
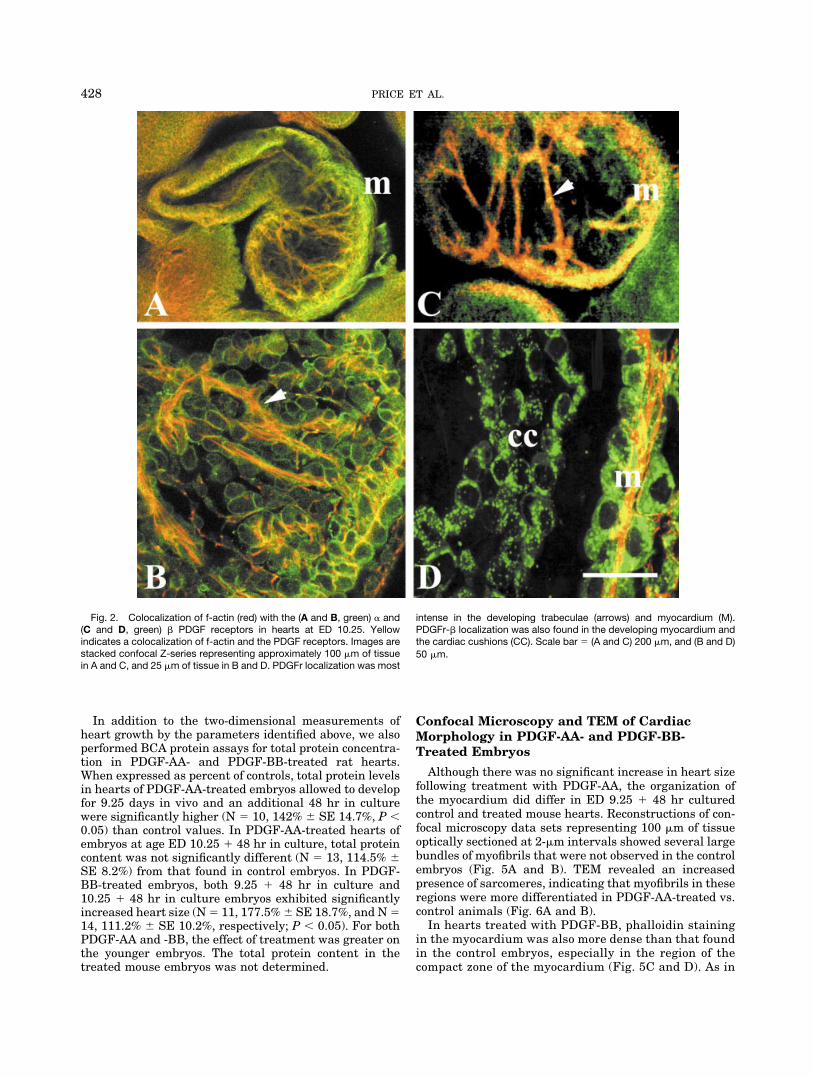
PDGF receptor localization has been well character-ized in the mouse system (Shinbrot et al., 1994; Price etal., 2001b). However, since this study utilized both ratand mouse embryos, and we could find no publisheddetails concerning PDGF receptor localization in the ratheart, confocal microscopy was used to determine thedistribution of both the PDGFr-� and -� receptors in theED 9.25–11.25 rat, which was the developmental win-dow used for the WEC studies. In animals at this stageof development, both types of receptors were localizedthroughout the developing heart (Fig. 2A and C). Local-ization of the PDGFr � was most intense on the devel-oping myocytes of the trabeculae and ventricular walls(Fig. 2A and B). Localization of the � receptor in thecardiac cushions was present, but was not as intense asthat found on myocytes. The PDGFr � was also foundthroughout the developing heart (Fig. 2C), including thecardiac cushions (Fig. 2D) and endothelial cells. Local-ization patterns of both receptor subtypes in the ratembryos were similar to those previously reported forthe mouse (Orr-Urtreger et al., 1992; Shinbrot et al.,1994; Takakura et al., 1997).
Effects of PDGF-AA and -BB on Cardiac Growthin WEC
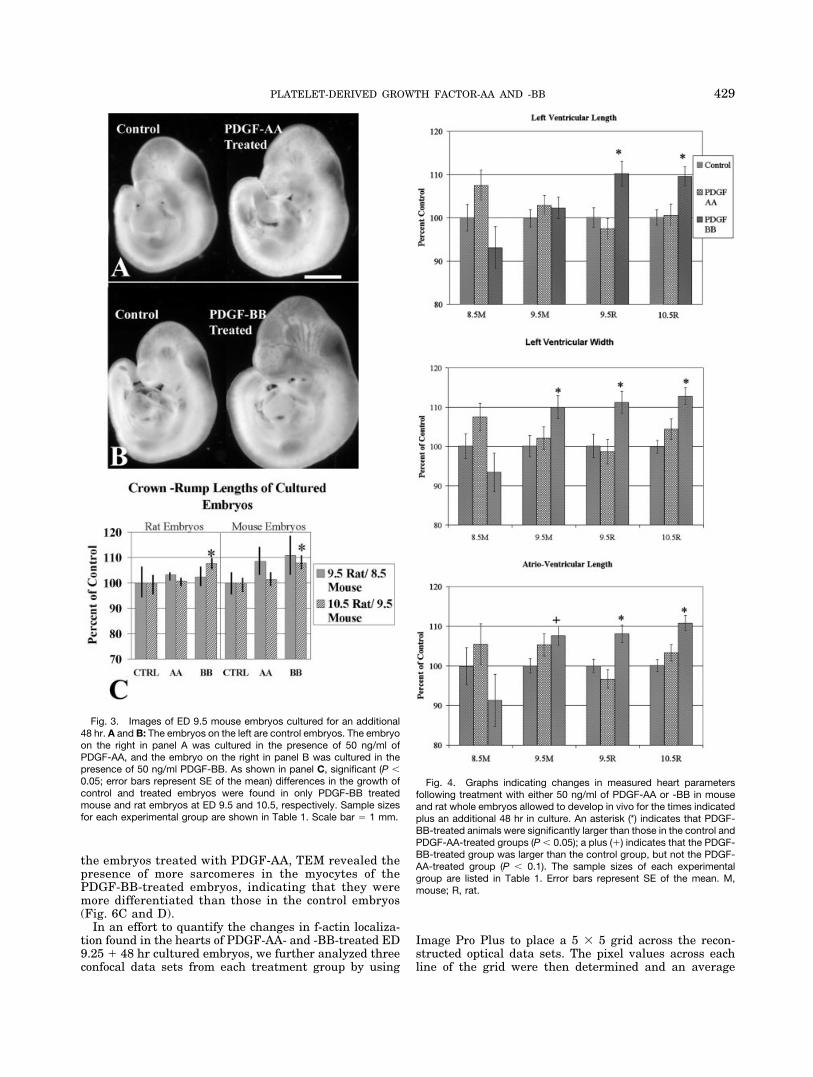
Figure 3 shows littermate mouse embryos following 9.5days of in vivo development and 48 hr in WEC in thepresence of 50 ng/ml of PDGF-AA (Fig. 3A) or 50 ng/ml ofPDGF-BB (Fig. 3B). Only PDGF-BB resulted in a signifi-cant increase in overall embryonic size at the later timepoints tested (Fig. 3C). It is to be expected that as overallbody size increases, a concomitant increase in heart sizeshould also occur. To account for this trend in increasedbody size, and to determine whether heart size increasedmore than expected, when we measured the heart param-eters we normalized the heart measurement to body sizeby dividing each measurement of the heart by the crown–rump length of the embryo.
The addition of 50 ng/ml of PDGF-AA to the culturemedia of both rat and mouse embryos did not result in asignificant increase in heart growth when compared to theoverall growth of the embryos. However, the addition of 50ng/ml of PDGF-BB to the culture media resulted in in-creased growth in both the in vivo ED 9.5 and 10.5 ratembryos and the ED 9.5 mouse embryos (Fig. 4) following48 hr of culture. These morphological measurements in-dicate that heart growth increased at a rate in excess ofoverall embryonic growth when exogenous quantities ofPDGF-BB were added to the culture media.
427PLATELET-DERIVED GROWTH FACTOR-AA AND -BB
In addition to the two-dimensional measurements ofheart growth by the parameters identified above, we alsoperformed BCA protein assays for total protein concentra-tion in PDGF-AA- and PDGF-BB-treated rat hearts.When expressed as percent of controls, total protein levelsin hearts of PDGF-AA-treated embryos allowed to developfor 9.25 days in vivo and an additional 48 hr in culturewere significantly higher (N � 10, 142% � SE 14.7%, P �0.05) than control values. In PDGF-AA-treated hearts ofembryos at age ED 10.25 � 48 hr in culture, total proteincontent was not significantly different (N � 13, 114.5% �SE 8.2%) from that found in control embryos. In PDGF-BB-treated embryos, both 9.25 � 48 hr in culture and10.25 � 48 hr in culture embryos exhibited significantlyincreased heart size (N � 11, 177.5% � SE 18.7%, and N �14, 111.2% � SE 10.2%, respectively; P � 0.05). For bothPDGF-AA and -BB, the effect of treatment was greater onthe younger embryos. The total protein content in thetreated mouse embryos was not determined.
Confocal Microscopy and TEM of CardiacMorphology in PDGF-AA- and PDGF-BB-Treated Embryos
Although there was no significant increase in heart sizefollowing treatment with PDGF-AA, the organization ofthe myocardium did differ in ED 9.25 � 48 hr culturedcontrol and treated mouse hearts. Reconstructions of con-focal microscopy data sets representing 100 �m of tissueoptically sectioned at 2-�m intervals showed several largebundles of myofibrils that were not observed in the controlembryos (Fig. 5A and B). TEM revealed an increasedpresence of sarcomeres, indicating that myofibrils in theseregions were more differentiated in PDGF-AA-treated vs.control animals (Fig. 6A and B).
In hearts treated with PDGF-BB, phalloidin stainingin the myocardium was also more dense than that foundin the control embryos, especially in the region of thecompact zone of the myocardium (Fig. 5C and D). As in
Fig. 2. Colocalization of f-actin (red) with the (A and B, green) � and(C and D, green) � PDGF receptors in hearts at ED 10.25. Yellowindicates a colocalization of f-actin and the PDGF receptors. Images arestacked confocal Z-series representing approximately 100 �m of tissuein A and C, and 25 �m of tissue in B and D. PDGFr localization was most
intense in the developing trabeculae (arrows) and myocardium (M).PDGFr-� localization was also found in the developing myocardium andthe cardiac cushions (CC). Scale bar � (A and C) 200 �m, and (B and D)50 �m.
428 PRICE ET AL.
the embryos treated with PDGF-AA, TEM revealed thepresence of more sarcomeres in the myocytes of thePDGF-BB-treated embryos, indicating that they weremore differentiated than those in the control embryos(Fig. 6C and D).
In an effort to quantify the changes in f-actin localiza-tion found in the hearts of PDGF-AA- and -BB-treated ED9.25 � 48 hr cultured embryos, we further analyzed threeconfocal data sets from each treatment group by using
Image Pro Plus to place a 5 � 5 grid across the recon-structed optical data sets. The pixel values across eachline of the grid were then determined and an average
Fig. 3. Images of ED 9.5 mouse embryos cultured for an additional48 hr. A and B: The embryos on the left are control embryos. The embryoon the right in panel A was cultured in the presence of 50 ng/ml ofPDGF-AA, and the embryo on the right in panel B was cultured in thepresence of 50 ng/ml PDGF-BB. As shown in panel C, significant (P �0.05; error bars represent SE of the mean) differences in the growth ofcontrol and treated embryos were found in only PDGF-BB treatedmouse and rat embryos at ED 9.5 and 10.5, respectively. Sample sizesfor each experimental group are shown in Table 1. Scale bar � 1 mm.
Fig. 4. Graphs indicating changes in measured heart parametersfollowing treatment with either 50 ng/ml of PDGF-AA or -BB in mouseand rat whole embryos allowed to develop in vivo for the times indicatedplus an additional 48 hr in culture. An asterisk (*) indicates that PDGF-BB-treated animals were significantly larger than those in the control andPDGF-AA-treated groups (P � 0.05); a plus (�) indicates that the PDGF-BB-treated group was larger than the control group, but not the PDGF-AA-treated group (P � 0.1). The sample sizes of each experimentalgroup are listed in Table 1. Error bars represent SE of the mean. M,mouse; R, rat.
429PLATELET-DERIVED GROWTH FACTOR-AA AND -BB

value was obtained. For high-affinity stains, such as phal-loidin, if all embryos and images are treated equally, ahigher average pixel value in the image will indicate anincreased presence of f-actin and increased differentiationof the myocytes. Pixel values from the control embryo datasets averaged 139.1 � SE 1.4, PDGF-AA-treated embryodata sets averaged 149.7 � SE 1.5, and PDGF-BB-treatedembryo data sets averaged 204.3 � SE 2.3. ANOVA testsshowed that pixel values in both PDGF-AA- and PDGF-BB-treated embryos were significantly higher than controlvalues, and that PDGF-BB values were significantlygreater than those found in the PDGF-AA-treated em-bryos (P � 0.005). Together with the confocal microscopyand TEM images, these data indicate that the myocytes inthe treated embryos were more differentiated than thosein the control embryos.
DISCUSSION
Both the PDGF-A and -B ligands and the � and �receptors are known to modulate a variety of cellularresponses during embryonic development. An animal inwhich any one of these components is knocked out willdie during embryonic or neonatal development (re-viewed in Ataliotis and Mercola, 1997; Betsholtz andHeldin, 1998). In the current experiments we usedmouse and rat WEC techniques to take a contrastingexperimental approach to previous knockout studies byincreasing the exposure of developing embryos to thePDGF ligands in WEC. The results for rat and mouseWEC were similar, with PDGF-BB stimulating agreater increase in total protein content and cardiacsize than PDGF-AA. This might be expected since
Fig. 5. Confocal microscopy data sets representing approximately100 �m of tissue from mouse hearts allowed to develop 9.25 days in vivoand an additional 48 hr in culture prior to fixation and optical sectioningat 2-�m intervals. Data sets were reconstructed using Voxblast. PanelsA and B are taken from control and PDGF-AA-treated embryos, respec-
tively, and illustrate the increased development of the trabeculae (ar-rows) in treated embryos. Panels C and D are taken from control andPDGF-BB-treated embryos, respectively, and illustrate the increaseddevelopment of the compact zone of the myocardium (delineated byarrows) in the PDGF-BB-treated embryos. Scale bar � 30 �m.
430 PRICE ET AL.
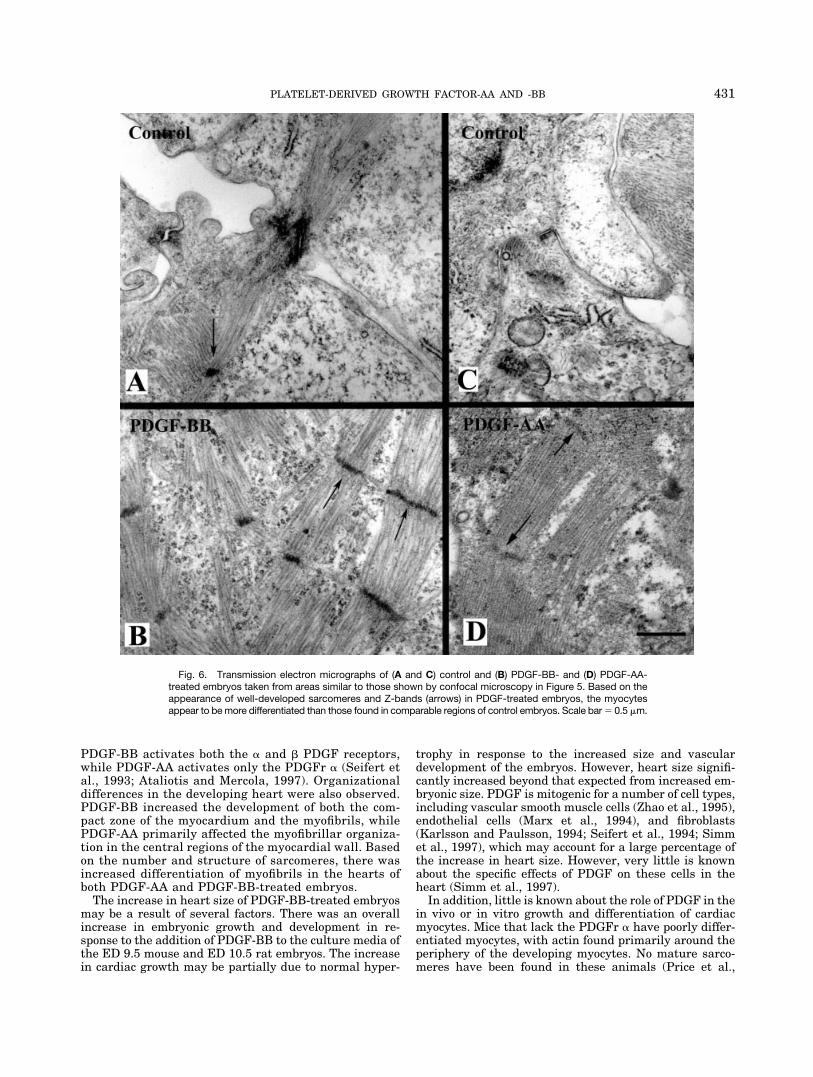
PDGF-BB activates both the � and � PDGF receptors,while PDGF-AA activates only the PDGFr � (Seifert etal., 1993; Ataliotis and Mercola, 1997). Organizationaldifferences in the developing heart were also observed.PDGF-BB increased the development of both the com-pact zone of the myocardium and the myofibrils, whilePDGF-AA primarily affected the myofibrillar organiza-tion in the central regions of the myocardial wall. Basedon the number and structure of sarcomeres, there wasincreased differentiation of myofibrils in the hearts ofboth PDGF-AA and PDGF-BB-treated embryos.
The increase in heart size of PDGF-BB-treated embryosmay be a result of several factors. There was an overallincrease in embryonic growth and development in re-sponse to the addition of PDGF-BB to the culture media ofthe ED 9.5 mouse and ED 10.5 rat embryos. The increasein cardiac growth may be partially due to normal hyper-
trophy in response to the increased size and vasculardevelopment of the embryos. However, heart size signifi-cantly increased beyond that expected from increased em-bryonic size. PDGF is mitogenic for a number of cell types,including vascular smooth muscle cells (Zhao et al., 1995),endothelial cells (Marx et al., 1994), and fibroblasts(Karlsson and Paulsson, 1994; Seifert et al., 1994; Simmet al., 1997), which may account for a large percentage ofthe increase in heart size. However, very little is knownabout the specific effects of PDGF on these cells in theheart (Simm et al., 1997).
In addition, little is known about the role of PDGF in thein vivo or in vitro growth and differentiation of cardiacmyocytes. Mice that lack the PDGFr � have poorly differ-entiated myocytes, with actin found primarily around theperiphery of the developing myocytes. No mature sarco-meres have been found in these animals (Price et al.,
Fig. 6. Transmission electron micrographs of (A and C) control and (B) PDGF-BB- and (D) PDGF-AA-treated embryos taken from areas similar to those shown by confocal microscopy in Figure 5. Based on theappearance of well-developed sarcomeres and Z-bands (arrows) in PDGF-treated embryos, the myocytesappear to be more differentiated than those found in comparable regions of control embryos. Scale bar � 0.5 �m.
431PLATELET-DERIVED GROWTH FACTOR-AA AND -BB

2001b). In isolated adult rat cardiac myocytes, PDGF-BB(50 ng/ml) stimulated protein synthesis by 87% (Neyses etal., 1991). However, Velez et al. (1994) reported only smallincreases of 6% in the cytoplasmic and 8% in the cytoskel-etal total protein levels in isolated chicken cardiac myo-cytes (Hamburger and Hamilton stage 19) stimulatedwith 3.64 ng/ml of PDGF. In older embryos (stages 29 and39), cytoplasmic protein levels dropped 4% and 8%, respec-tively, while cytoskeletal protein levels did not changewhen PDGF was added to the culture media. In subse-quent studies Velez et al. (1995, 2000) also found a slightdecrease in actin and �-actinin levels, and an increase invimentin levels in PDGF-treated chick cardiac myocytes,indicating that PDGF treatment may induce a reversion ofmyocytes to a less differentiated state. However, it may bedifficult to compare mammalian and chick myocytestreated with PDGF because there is no indication that thePDGFr � is present in the chick (Potts and Carrington,1993), and the PDGF ligand used in the Velez studies wasnot identified.
There are other differences between the current exper-iments and those of Velez et al. (1994, 1995, 2000). InWEC, cardiac myocytes essentially are allowed to developin vivo. This differs from most studies on cardiac myocytedevelopment, in which the cells are isolated during lateembryonic or neonatal stages of development and are sub-sequently established in culture. Following cell isolation,the contractile apparatus of cardiac myocytes breaks downand then reforms in an organizational pattern that fre-quently differs from that found in vivo (Simpson et al.,1996). This breakdown and reassembly does not occurduring WEC (Nakagawa et al., 1997). In addition, in WECthe myocytes are influenced by the surrounding intersti-tial cells, extracellular matrix components, and otherchemical and mechanical cell signaling mechanisms thatcontribute to myocyte differentiation. Although cell cul-ture studies are important in determining the roles ofspecific growth factors and signaling mechanisms, they donot take into account the influence of the surroundingcells and tissues.
In summary, the present WEC studies provide data thatcomplement the findings of previous knockout studies inwhich specific PDGF ligands or receptors were deleted, andfurther demonstrate the importance of PDGF in the earlyembryonic development of the heart. The addition of exoge-nous PDGF-AA and -BB to the culture media is analogous toan overexpression of the PDGF ligand, and results in thestimulation of the � and � receptors and an increase incardiac growth and myofibrillar differentiation.
ACKNOWLEDGMENTSThese studies were funded by National Institutes of
Health grants HD35253 (to R.L.P.), HL58893 (to L.T.),and HL42249.
LITERATURE CITEDAtaliotis P, Mercola M. 1997. Distribution and functions of platelet-
derived growth factors and their receptors during embryogenesis.Ann Rev Cytol 172:95–127.
Bergsten E, Uutela M, Li X, Pietras K, Ostman A, Heldin CH, AlitaloK, Eriksson U. 2001. PDGF-D is a specific, protease-activated li-gand for the PDGF �-receptor. Nat Cell Biol 3:512–516.
Betsholtz C, Heldin C-H. 1998. Platelet derived growth factor: aregulator of connective tissue development and reaction. In: ReedRK, Rubin K, editors. Connective tissue biology: integration andreductionism. London: Portland Press Ltd. p 179–193.
Bostrom H, Willetts K, Milos P, Leveen P, Lindahl P, Hedstrand H,Pekna M, Hellstrom M, Gebre-Medhin S, Schalling M, Nilsson M,Kurland S, Tornell J, Heath JK, Betsholtz C. 1996. PDGF-A sig-naling is a critical event in lung alveolar myofibroblast developmentand alveogenesis. Cell 85:863–873.
Brown NA, Fabro S. 1981. Quantitation of rat embryonic developmentin vitro: a morphological scoring system. Teratology 24:65–78.
Ding H, Wu X, Kim I, Tam PPL, Koh GY, Nagy A. 2000. The mousePdgfc gene: dynamic expression in embryonic tissues during orga-nogenesis. Mech Dev 96:209–213.
Fruttiger M, Karlsson L, Hall AC, Abramsson A, Calver AR,Bostrom H, Willetts K, Bertold CH, Heath JK, Betsholtz C,Richardson WD. 1999. Defective oligodendrocyte developmentand severe hypomyelination in PDGF-A knockout mice. Develop-ment 126:457– 467.
Fujinaga M, Baden JM. 1992. Variation in development of rat em-bryos at the presomite period. Teratology 45:661–670.
Gilbertson DG, Duff ME, West JW, Kelly JD, Sheppard PO, HofstrandPD, Gao Z, Shoemaker K, Bukowski TR, Moore M, Feldhaus AL,Humes JM, Palmer TE, Hart CE. 2001. Platelet-derived growthfactor C (PDG-C), a novel growth factor that binds to PDGF� and �receptor. J Biol Chem 276:27406–27414.
Hansen P, Dryer C. 1981. The use of polyacrylamide gel as an em-bedding medium for immunohistochemical studies of embryonictissue. Stain Technol 56:287–293.
Heldin CH, Westermark B. 1989. Platelet-derived growth factor:three isoforms and two receptor types. Trends Genet 5:108–111.
Huang ZJ, Haugland RP, You WM, Haugland RP. 1992. Phallotoxinand actin binding assay by fluorescence enhancement. Anal Bio-chem 1:199–204.
Kaminski WE, Lindahl P, Lin NL, Broudy VC, Crosby JR, HellstromM, Swolin B, Bowen-Pope D, Martin PJ, Ross R, Betsholtz C, RainesEW. 2001. Basis of hematopoietic defects in platelet-derived growthfactor (PDGF)-B and PDGF �-receptor null mice. Blood 97:1990–1998.
Karlsson C, Paulsson Y. 1994. Age related induction of platelet-de-rived growth factor A-chain mRNA in normal human fibroblasts.J Cell Physiol 158:256–262.
Kazlauskas A. 2000. A new member of an old family. Nat Cell Biol2:E78–E79.
LaRochelle WJ, Jeffers M, McDonald WF, Chillakuru RA, Giese NA,Lokker NA, Sullivan C, Boldog FL, Yang M, Vernet C, Burgess CE,Fernandes E, Deegler LL, Rittman B, Shimkets J, Shimkets RA,Rothberg JM, Lichenstein HS. 2001. PDGF-D, a new protease-activated growth factor. Nat Cell Biol 3:517–521.
Leveen P, Pekny M, Gebre-Medhin S, Swolin B, Larsson E, BetsholtzC. 1994. Mice deficient for PDGF B show renal, cardiovascular, andhematological abnormalities. Genes Dev 8:1875–1887.
Li X, Ponten A, Aase K, Karlsson L, Abramsson A, Uutela M, Back-strom H, Li H, Soriano P, Betsholtz C, Heldin CH, Alitalo K,Ostman A, Eriksson U. 2000. PDGF-C is a new protease-activatedligand for the PDGF� -receptor. Nat Cell Biol 2:302–309.
Marx M, Perlmutter RA, Madri JA. 1994. Modulation of platelet-derived growth factor receptor expression in microvascular endo-thelial cells during angiogenesis. J Clin Invest 93:131–139.
Morrison-Graham K, Schatteman GC, Bork T, Bowen-Pope DF, Wet-son JA. 1992. A PDGF receptor mutation in the mouse (Patch)perturbs the development of a non-neuronal subset of neural crest-derived cells. Development 115:133–142.
Nakagawa M, Price RL, Chintanowonges C, Simpson DG, Horacek M,Borg TK, Terracio L. 1997. Analysis of heart development in cul-tured rat embryos. J Mol Cell Cardiol 29:369–379.
New DAT. 1978. Whole embryo culture and the study of mammalianembryos during organogenesis. Biol Rev 53:81–122.
Neyses L, Nouskas J, Oberdorf S, Vetter H. 1991. Induction of theearly growth response gene-1 (Egr-1) and protein synthesis byplatelet derived growth factor BB(PDGF) and insulin in adult ratcardiomyocytes. Circulation 84(Suppl II):II396.
Orr-Urtreger A, Bedford MT, Do M, Eisenbach L, Lonai P. 1992.Developmental expression of the � receptor for platelet-derivedgrowth factor, which is depleted in the embryonic lethal Patchmutation. Development 115:289–303.
432 PRICE ET AL.

Potts JD, Carrington JL. 1993. Selective expression of the chickenplatelet-derived growth factor � (PDGF�) receptor during limb buddevelopment. Dev Dyn 198:14–21.
Price RL, Chintanowonges C, Shiraishi I, Borg TK, Terracio L. 1996.Local and regional variations in myofibrillar patterns in looping rathearts. Anat Rec 245:83–93.
Price RL, Carver W, Simpson DG, Fu L, Zhao J, Borg TK, Terracio L.1997. The effects of angiotensin II and specific angiotensin receptorblockers on embryonic cardiac development and looping patterns.Dev Biol 192:572–584
Price RL, Potts JD, Thielen TE, Borg TK, Terracio L. 2001a. Growthfactor regulation of embryonic, fetal, and neonatal cardiac develop-ment. In: Tomanek RJ, Runyan R, editors. Development of thecardiovascular system. New York: Springer-Verlag. p 171–199.
Price RL, Thielen TE, Borg TK, Terracio L. 2001b. Cardiac defectsassociated with the absence of the platelet-derived growth factoralpha receptor in the Patch mouse. Microsc Microanal 7:56–65.
Sadler TW, Horton WE, Warner CW. 1982. Whole embryo culture: ascreening technique for teratogens? Teratog Carcinog Mutagen2:243–253.
Schatteman GC, Morrison-Graham K, van Koppen A, Weston JA,Bowen-Pope DF. 1992. Regulation and role of PDGF receptor�-subunit expression during embryogenesis. Development 115:123–131.
Schatteman GC, Motley ST, Effman EL, Bowen-Pope DF. 1995. Plate-let-derived growth factor receptor alpha subunit deleted Patchmouse exhibits severe cardiovascular dysmorphogenesis. Teratol51:351–366.
Schatteman GC, Loushin C, Li T, Hart CE. 1996. PDGF-A is requiredfor normal murine cardiovascular development. Dev Biol 176:133–142.
Seifert RA, van Koppen A, Bowen-Pope DF. 1993. PDGF-AB requiresPDGF receptor �-subunits for high-affinity, but not for low-affinity,binding and signal transduction. J Biol Chem 268:4473–4480.
Seifert RA, Coats SA, Raines EW, Ross R, Bowem-Pope DF. 1994.Platelet-derived growth factor (PDGF) receptor �-subunit mutantand reconstituted cell lines demonstrate that transforming growthfactor-� can be mitogenic through PDGF A-chain-dependent and-independent pathways. J Biol Chem 269:13951–13955.
Shinbrot E, Peters KG, Williams LT. 1994. Expression of the platelet-derived growth factor � receptor during organogenesis and tissuedifferentiation in the mouse embryo. Dev Biol 199:169–175.
Simm A, Nestler M, Hoppe V. 1997. PDGF-AA, a potent mitogen forcardiac fibroblasts from adult rats. J Mol Cell Cardiol 29:357–368.
Simpson DG, Sharp WW, Borg TK, Price RL, Terracio L, SammarelAM. 1996. Mechanical regulation of cardiac myocyte protein turn-over and myofibrillar structure. Am J Physiol (Cell Physiol) 270:C1075–C1087.
Soriano P. 1994. Abnormal kidney development and hematologicaldisorders in PDGF �-receptor mutant mice. Genes Dev 8:1888–1896.
Soriano P. 1997. The PDGF� receptor is required for neural crest celldevelopment and for normal patterning of somites. Development124:2691–2700.
Takakura N, Yoshida H, Ogura Y, Kataoka H, Nishikawa S, Nih-sikawa SI. 1997. PDGFR� expression during mouse embryogenesis:immunolocalization analyzed by whole-mount immunohistostain-ing using the monoclonal anti-mouse antibody APA5. J HistochemCytochem 45:883–893.
Velez C, Aranega E, Melguizo C, Fernandez JE, Prados J, Aranega A.1994. Modulation of contractile protein troponin-T in chick myocar-dial cells by basic fibroblast growth factor and platelet-derivedgrowth factor during development. J Cardiovasc Pharmacol 24:906–913.
Velez C, Aranega E, Prados JC, Melguizo C, Alvarez L, Aranega A.1995. Basic fibroblast and platelet-derived growth factors as mod-ulators of actin and �-actinin in chick myocardiocytes during devel-opment. Proc Soc Exp Biol Med 210:57–63.
Velez C, Aranega E, Marchal JA, Melguizo C, Prados JC, Aranega A.2000. Development of chick cardiomyocytes: modulation of interme-diate filaments by basic fibroblast and platelet-derived growth fac-tors. Cells Tissues Organs 167:163–170.
Zhao XM, Frist WH, Yeoh TK, Miller GG. 1995. Confirmation ofalternatively spliced platelet-derived growth factor-A chain andcorrelation with expression of PDGF receptor-� in human cardiacallografts. Transplantation 59:605–611.
433PLATELET-DERIVED GROWTH FACTOR-AA AND -BB




















