



Electrostatic Association of Glutathione Transferase to the Nuclear Membrane: EVIDENCE OF AN ENZYME...
17
1 ELECTROSTATIC ASSOCIATION OF GLUTATHIONE TRANSFERASE TO THE NUCLEAR MEMBRANE Evidence of an enzyme defense barrier at the nuclear envelope Lorenzo Stella 1 , Valentina Pallottini 2 , Sandra Moreno 2 , Silvia Leoni 3 , Francesca De Maria 1 , Paola Turella 1 , Giorgio Federici 4 , Raffaele Fabrini 1 , Kutayba F. Dawood 1 , Mario Lo Bello 5 , Jens Z. Pedersen 5 and Giorgio Ricci 1 From the Departments of 1 Chemical Sciences and Technologies and 5 Biology, University of Rome “Tor Vergata”, 00133 Rome, 2 Department of Biology, University of Rome “Roma Tre”, 00146 Rome, and 3 Department of Cellular and Developmental Biology, University of Rome “La Sapienza” 00185 Rome, and 4 Children’s Hospital IRCCS “Bambin Gesù”, 00165 Rome, Italy Running Title: Nuclear association of glutathione transferases Address correspondence to: Prof. Giorgio Ricci, Department of Chemical Sciences and Technologies, University of Rome “Tor Vergata”, Via della Ricerca Scientifica, 00133 Rome, Italy. Tel: 39-0672594379 Fax: 39-0672594328; Email: [email protected] The possible nuclear compartmentalization of glutathione transferase isoenzymes has been the subject of contradictory reports. The discovery that the dinitrosyl-diglutathionyl-iron complex binds tightly to Alpha class GSTs in rat hepatocytes and that a significant part of the bound complex is also associated to the nuclear fraction (Pedersen et al., the accompanying paper), prompted us to reconsider the nuclear localization of GSTs in these cells. Surprisingly, we found that a considerable amount of GSTs corresponding to 10% of the cytosolic pool, is electrostatically associated to the outer nuclear membrane, and a similar quantity is compartmentalized inside the nucleus. Mainly Alpha class GSTs, in particular GSTA1-1, GSTA2-2 and GSTA3-3 are involved in this double modality of interaction. Confocal microscopy, immunofluorescence experiments and molecular modeling have been used to detail the electrostatic association in hepatocytes and liposomes. A quantitative analysis of the membrane bound Alpha GSTs suggests the existence of a multilayer assembly of these enzymes at the outer nuclear envelope that could represent an amazing novelty in cell physiology. The interception of potentially noxious compounds to prevent DNA damage could be the possible physiological role of the perinuclear and intranuclear localization of Alpha GSTs. Glutathione transferases (GSTs) are a superfamily of enzymes that protect the cell from toxic endogenous or xenobiotic compounds. Eight different, gene independent, isoenzymatic classes have been found in mammals, and two of these, Alpha and Mu classes, are abundantly expressed in rat liver, where they represent 43% and 56%, respectively, of the entire pool of cytosolic GSTs (1). Beside their catalytic competence to conjugate GSH to many toxic substances having an electrophilic centre, these enzymes also display peroxidase activity and ligandin properties (2, 3). In hepatocytes, Alpha GSTs efficiently trap a natural nitric oxide carrier, the dinitrosyl-diglutathionyl-iron complex (DNDGIC), thus preventing the irreversible inhibition of glutathione reductase caused by this compound (4). Preliminary evidence was also reported that GSTs could be associated to the nuclear membrane or compartmentalized in nuclei. The presence of GSTs on intracellular membranes is not a new finding. MGST1 is a peculiar trimeric microsomal integral membrane isoenzyme discovered and characterized many years ago (5). GSTA4-4, a specific isoenzyme able to detoxify hydroxyalkenals, displays a widespread mitochondrial, peroxisomal and cytosolic localization, but the plasma membrane also binds detectable amounts of this enzyme (6). Furthermore, the tight association of Alpha and Mu class GSTs with the microsomal membrane fraction of rat liver was demonstrated by Morgenstern et al. (7), and about 2% of the cytosolic GSTA1-1 has been found in the microsomal membrane of sheep liver cells (8). Immunohistochemical evidence suggested the presence of nuclear Alpha and Pi class GSTs in different human tissues, but these studies did not http://www.jbc.org/cgi/doi/10.1074/jbc.M609906200 The latest version is at JBC Papers in Press. Published on December 30, 2006 as Manuscript M609906200 Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc. by on January 12, 2007 www.jbc.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Electrostatic Association of Glutathione Transferase to the Nuclear Membrane: EVIDENCE OF AN ENZYME...

1
ELECTROSTATIC ASSOCIATION OF GLUTATHIONE TRANSFERASE TOTHE NUCLEAR MEMBRANE
Evidence of an enzyme defense barrier at the nuclear envelope
Lorenzo Stella1, Valentina Pallottini2, Sandra Moreno2, Silvia Leoni3, Francesca De Maria1, PaolaTurella1, Giorgio Federici4, Raffaele Fabrini1, Kutayba F. Dawood1, Mario Lo Bello5, Jens Z. Pedersen5
and Giorgio Ricci1
From the Departments of 1Chemical Sciences and Technologies and 5Biology, University of Rome“Tor Vergata”, 00133 Rome, 2Department of Biology, University of Rome “Roma Tre”, 00146 Rome, and
3Department of Cellular and Developmental Biology, University of Rome “La Sapienza” 00185 Rome,and 4Children’s Hospital IRCCS “Bambin Gesù”, 00165 Rome, Italy
Running Title: Nuclear association of glutathione transferasesAddress correspondence to: Prof. Giorgio Ricci, Department of Chemical Sciences and Technologies,University of Rome “Tor Vergata”, Via della Ricerca Scientifica, 00133 Rome, Italy. Tel: 39-0672594379Fax: 39-0672594328; Email: [email protected]
The possible nuclear compartmentalization ofglutathione transferase isoenzymes has been thesubject of contradictory reports. The discoverythat the dinitrosyl-diglutathionyl-iron complexbinds tightly to Alpha class GSTs in rathepatocytes and that a significant part of thebound complex is also associated to the nuclearfraction (Pedersen et al., the accompanyingpaper), prompted us to reconsider the nuclearlocalization of GSTs in these cells. Surprisingly,we found that a considerable amount of GSTscorresponding to 10% of the cytosolic pool, iselectrostatically associated to the outer nuclearmembrane, and a similar quantity iscompartmentalized inside the nucleus. MainlyAlpha class GSTs, in particular GSTA1-1,GSTA2-2 and GSTA3-3 are involved in thisdouble modality of interaction. Confocalmicroscopy, immunofluorescence experimentsand molecular modeling have been used to detailthe electrostatic association in hepatocytes andliposomes. A quantitative analysis of themembrane bound Alpha GSTs suggests theexistence of a multilayer assembly of theseenzymes at the outer nuclear envelope that couldrepresent an amazing novelty in cell physiology.The interception of potentially noxiouscompounds to prevent DNA damage could be thepossible physiological role of the perinuclear andintranuclear localization of Alpha GSTs.
Glutathione transferases (GSTs) are asuperfamily of enzymes that protect the cell from
toxic endogenous or xenobiotic compounds. Eightdifferent, gene independent, isoenzymatic classeshave been found in mammals, and two of these,Alpha and Mu classes, are abundantly expressed inrat liver, where they represent 43% and 56%,respectively, of the entire pool of cytosolic GSTs(1). Beside their catalytic competence to conjugateGSH to many toxic substances having anelectrophilic centre, these enzymes also displayperoxidase activity and ligandin properties (2, 3). Inhepatocytes, Alpha GSTs efficiently trap a naturalnitric oxide carrier, the dinitrosyl-diglutathionyl-ironcomplex (DNDGIC), thus preventing theirreversible inhibition of glutathione reductasecaused by this compound (4). Preliminary evidencewas also reported that GSTs could be associated tothe nuclear membrane or compartmentalized innuclei. The presence of GSTs on intracellularmembranes is not a new finding. MGST1 is apeculiar trimeric microsomal integral membraneisoenzyme discovered and characterized many yearsago (5). GSTA4-4, a specific isoenzyme able todetoxify hydroxyalkenals, displays a widespreadmitochondrial, peroxisomal and cytosoliclocalization, but the plasma membrane also bindsdetectable amounts of this enzyme (6). Furthermore,the tight association of Alpha and Mu class GSTswith the microsomal membrane fraction of rat liverwas demonstrated by Morgenstern et al. (7), andabout 2% of the cytosolic GSTA1-1 has been foundin the microsomal membrane of sheep liver cells (8).Immunohistochemical evidence suggested thepresence of nuclear Alpha and Pi class GSTs indifferent human tissues, but these studies did not
http://www.jbc.org/cgi/doi/10.1074/jbc.M609906200The latest version is at JBC Papers in Press. Published on December 30, 2006 as Manuscript M609906200
Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc.
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

2
quantify the specific isoenzymes involved (9, 10).Other investigations claimed either the presence orthe absence of nuclear compartmentalization ofGSTs. In particular, McCusker et al. did not reportany detectable nuclear GST activity (11), whileSoboll et al. (12), using a non-aqueous technique ofcell fractionation, found that both Alpha and MuGSTs are present in the nucleus. Other papersreported a nuclear localization of GSTs (13-15), butthe identification of the specific isoenzyme(s)involved in this association and their quantificationare uncertain.
The present study reveals for the first timethat beside a significant amount of Alpha GSTsinside the nucleus, an equivalent amount is found inelectrostatic association with the outer nuclearmembrane. This particular modality of interactionhas been detailed in cells and model systems usingconfocal microscopy, immunostaining experimentsand molecular modeling.
EXPERIMENTAL PROCEDURES
Materials – 6-(7-nitro-2,1,3-benzoxadiazol-4-y l t h i o ) h e x a n o l ( N B D H E X ) , dinitrosyl-diglutathionyl - iron complex (DNDGIC) and S-nitroso glutathione (GSNO) were synthesized asdescribed previously (16, 17). 1-chloro-2,4-dinitrobenzene (CDNB) and polyclonal antibodiesagainst Alpha and Mu GSTs were fromCalbiochem-Merck, Darmstadt, Germany.Fluorescein-conjugated goat anti-rabbit IgG wasfrom Vector, Burlingame, CA. Recombinant humanGSTA1-1, GSTM2-2 and GSTP1-1 were expressedin Escherichia coli and purified as describedpreviously (17-19). All other materials were ofreagent grade.Cells - CCRF-CEM cells (human T-lymphoblasticleukemia) were grown as described previously (20).Hepatocytes were prepared from Wistar male rats asdescribed in the accompanying paper (4).Experiments were carried out in accordance to theethical guidelines for animal research(Communication of the Italian Ministry of HealthNo. 246/H10, D.Lvo 116/92).Preparation of subcellular fractions – Subcellularfractions were prepared and characterized asreported in the accompanying paper (4) except thattwo parallel procedures were used; one using a
saline isotonic solution (0.05 M KCl, 0.04 MKH2PO4 and 0.1 M sucrose, pH 7.4), and one usingpure 0.25 M sucrose without salts.Purification of weakly and tightly bound GSTs –The nuclear fraction obtained from 10 g of liver waswashed three times with 20 ml of 0.25 M sucroseand resuspended in 20 ml of 0.25 M sucrosecontaining 10 mM potassium phosphate buffer, pH7.4. The suspension was rapidly centrifuged at 1000× g and the procedure was repeated. Further rapidextractions did not increase the amount of GSTs inthe supernatant. The collected supernatants wereconcentrated and represent the “weakly boundGSTs”. The pellet was again resuspended in thesame buffer solution and incubated under gentleagitation for one hour. This procedure was repeatedthree times. The collected supernatants wereconcentrated and represent the “tightly boundGSTs”. Both weakly and tightly bound GSTs werepurified by affinity chromatography through acolumn (1 × 4 cm) of glutathione-sepharose matrix(21). The first eluate was again passed through thecolumn to retain quantitatively the Alpha GSTs.GST activity – The standard test for glutathionetransferase activity contained 1 mM GSH and 1 mMCDNB in 1 ml of 0.1 M potassium phosphate buffer,pH 6.5. An isotonic enzymatic test for GST was alsoperformed using a standard PBS buffer at pH 6.5containing 1 mM GSH and 1 mM CDNB. Theactivity was followed spectrophotometrically at 340nm. Activity of nuclear GST was measured bydiluting aliquots of a nuclear suspension in thestandard assay mixture or using the isotonic testdescribed above. After two min of preincubation,the time course of the reaction was linear. A similarprocedure was used for activity determination in themitochondria, microsomes and lysosomes fractions.With these activity measurements it is not possibleto distinguish whether the activity is due to GSTsinside the intact structures or to GSTs bound to theoutside of the membranes.SDS-PAGE and immunoblot analysis – Proteinsamples were analyzed by SDS-PAGE andvisualized with Coomassie Brilliant Blue R-250 andtransferred to Hybond-ECL nitrocellulosemembranes (GE Healthcare). The proteins wereimmunoblotted with either a polyclonal anti-AlphaGST antibody, or a polyclonal anti-Mu GSTantibody.
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

3
EPR spectroscopy – EPR measurements were madeas described previously (4).HPLC analysis of GSTs – The GSTs extracted fromnuclei were resolved on a reverse-phase (C18, 4.6mm × 250 mm) column, essentially as reported byYeh et al. (1).Immunofluorescence studies – The nuclei pellet wasf ixed in 4% freshly depolymer izedparaformaldehyde in 0.25 M sucrose for 4 h at 4 °C.After extensive washings in 0.25 M sucrose, floatingpellets were incubated with a polyclonal anti-AlphaGST diluted 1:100 in 0.25 M sucrose overnight at 4°C, and washed again in 0.25 M sucrose for 20 min.Pellets were then incubated with fluoresceinconjugated goat anti-rabbit IgG diluted 1:200 in0.25 M sucrose, for 1 h at room temperature,washed in 0.25 M sucrose, and finally mounted onslides with Vectashield (Vector). Slides wereobserved in a laser scanning confocal microscope(Nikon) and micrographs were digitally captured.Fluorescence labelling of GSTA1-1, GSTM2-2 – 1mM NBD-Cl was reacted with 10 µM purifiedhuman GSTA1-1 or GSTM2-2 in 0.1 M potassiumphosphate buffer, pH 6.0. After two hours the excessof reagent was removed by G-25 Sephadexchromatography and the modified enzymes wereanalyzed spectrophotometrically. The selectivealkylation of enzyme cysteines was confirmed onthe basis of the diagnostic absorption peak centeredat about 430-440 nm.Liposome preparation – Phospholipids werepurchased from Avanti Polar Lipids (Alabaster,AL). Large unilamellar vesicles (LUVs) wereprepared according to a procedure reportedpreviously (22). Briefly, lipids were dissolved in achloroform/methanol solution (1:1 v/v), and thesolvents were evaporated under reduced argonatmosphere, until a thin film formed. Completeevaporation was ensured by applying a rotaryvacuum pump for at least 2 h, and the film washydrated with a 0.25 M sucrose solution. Aftervigorous stirring and 10 freeze and thaw cycles, theliposome suspension was extruded for 31 timesthrough two stacked polycarbonate membranes with100 nm pores (Avestin, Ottawa, ON, Canada). Finalphospholipid concentration was determined by theStewart method (23). Anionic liposomes wereformed by egg phosphatidylcholine, eggphosphatidylglycerol and cholesterol (8:1:1 molarratio), to reproduce the charged lipid and cholesterol
content of the nuclear membrane (24). Neutralliposomes were formed by phosphatidylcholine andcholesterol (9:1 molar ratio).
Giant unilamellar vesicles (GUVs) wereprepared by the electroformation method (25).Lipids were dissolved in a choloroform/acetonitrilesolution (95:5 v/v) at a 5 mM total concentration;0.25 ml of this solution was deposited on a 25 mm ×30 mm indium tin oxide (ITO) covered glass slide,and spin coated at 600 rpm for 5 min. Completesolvent evaporation was ensured by applying arotary vacuum pump for at least 2 h. A 15 mm × 15mm × 1.5 mm electroformation chamber wasformed by separating the lipid covered ITOelectrode by another bare ITO glass slide with apoly(dimethylsiloxane) spacer. The chamber wasfilled with a 0.25 M sucrose solution, and a 1.1 V(peak to peak), 10 Hz ac potential was applied to thetwo ITO electrodes, using a Wavetek 182 Afunction generator. After 1.5 h the potential wasswitched to 3 V, 4 Hz for 10 minutes, to favordetachment of GUVs from the electrode. Thesolution contained in the electroformation chamberwas gently removed, and aliquots were deposited onsilanized microscope slides for observation in theconfocal microscope. The lipid compositions ofGUVs were the same employed for LUVs, exceptt h a t 1 % r h o d a m i n e - l a b e l e dphosphatidylethanolamine was included to allowvisualization of the liposomes by confocalmicroscopy.Electrostatic calculations – Molecular structures ofhuman and rat GSTs were derived from thefollowing Protein Data Bank entries: 2GSD (humanA1-1), 2GTU (human M2-2), 6GSS (human P1-1),1EV4 (rat A1-1), 1B4P (rat M2-2). Protein chargeswere calculated by using the PDB2PQR software(26), and Poisson-Boltzmann calculations ofelectrostatic potential were performed with theAPBS program (27), with 161 × 161 × 161 gridpoints, a 110 Å coarse grid and a 83 Å fine griddimension, dielectric constants equal to 2 and 78.54for protein and water, respectively, and Debye-Hückel boundary conditions. Protein dipoles wereestimated by employing the Dipole server(bioportal.weizmann.ac.il/dipol/), and moleculargraphics were realized with the MOLMOL (28) andUCSF Chimera softwares (29).Confocal imaging – Confocal images were acquiredwith a confocal laser scanning microscope, Nikon
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

4
PCM 2000 (Nikon Instruments) equipped withSpectra Physics Ar-ion laser (488 nm, 514 nm) andHe-Ne laser (543.5 nm) sources. A 60×/1.4 oilimmersion objective was used for the observations.
RESULTS
GST activity in subcellular fractions of rathepatocytes – Many peripheral membrane proteinscan easily be detached with mild treatments likeincreasing the ionic strength. As most standardprocedures for the isolation of subcellularcomponents include the use of buffered solutions,some of the electrostatic protein-membraneinteractions occurring in intact cells may be lost.Thus, in our study on the cellularcompartmentalization of GSTs, we used only anisotonic solution of sucrose (0.25 M) forhomogenization of the liver, as well as for isolationof the subcellular components. In addition, weadopted a particular GST activity determination onintact organelles (see Experimental Procedures) thatavoids inactivation caused by sonication steps or bydetergent extractions. As shown in Table 1, crudenuclear, lysosomal, mitochondrial and microsomalfractions all contain GST activity. The highestactivity is recovered in the nuclear pellet, thatcontains an amount corresponding to about 15% ofthe overall cytosolic GST activity. Interestingly,about 50 % of the GST activity of the fractions islost when the subcellular components are isolated inisotonic saline solution (Table 1), suggesting thatabout half of the GST could be electrostaticallybound. Due to the considerable amount of GSTactivity associated to the nuclear pellet, only thisspecific subcellular fraction was further studied andcharacterized.
GST activity in hepatocyte nuclear pellet – Thenuclear pellet from rat liver, isolated afterhomogenization in 0.25 M sucrose (1:10), waswashed three times with ten volumes of sucrose 0.25M to remove any contamination of the cytosolicGSTs. Optical microscopy shows that the nuclei areintact. The nuclear pellet resuspended in 0.25 Msucrose, contains a total of 16 GST units per gtissue. After treatment of nuclei with ten volumes of10 mM potassium phosphate buffer or 20 mM NaClin the presence of 0.25 M sucrose, about 8 units are
released into the surrounding solution, confirmingthe presence of GSTs electrostatically bound to thenuclei (weakly bound GST). No additional release isobserved after increasing the concentration of thebuffer up to 0.1 M. The remaining activityassociated to the nuclear pellet, termed “tightlybound GST” (about 8 units) can be only partiallyextracted by multiple and prolonged incubations (1hour each) with ten volumes of 0.25 M sucrose in 10mM potassium phosphate buffer. Sonicationtreatment of the nuclear pellet or the use ofdetergents like Triton X-100 even at lowconcentrations (1%) causes partial and irreversibleinactivation of the enzyme. The use of concentratedcarbonate does not increase the rate and extent ofthe extraction, indicating that the tightly bound GSTdoes not behave like a peripheral protein (8). Themost likely localization of the tightly bound GST isthe nuclear interior, as indicated by confocalmicroscopy (see below).
Further evidence of the presence ofequivalent levels of GSTs associated to the nucleusis provided by EPR spectroscopy. It has beendemonstrated that the paramagnetic speciesDNDGIC binds tightly to Alpha and Mu GST with1:1 stoichiometry, giving an EPR spectrum differentfrom that of the free complex (30). Titration ofGSTs with DNDGIC therefore allows quantificationof GSTs. Experiments performed on nuclei isolatedin 0.25 M sucrose or in saline solution are shown inFig. 1 where the loss of activity due to the bindingof DNDGIC is also reported. Starting from a ratliver of 10 grams (51 mg total GSTs), the weaklybound GST is about 5 mg and a similar amount isrecovered as tightly bound GST.
GSTA1-1 and GSTA2-2 are involved in the nuclearassociation – The extracts containing the solubilizedweakly and tightly bound GSTs contain otherproteins as well. In fact, they display specificactivities of approximately 3 U/mg which rise to 16U/mg after glutathione-sepharose affinitychromatography. SDS-PAGE electrophoresis,performed after the affinity step, indicates that thesesamples have similar protein composition (Fig. 2A)with a main component at about 25 kDa and a minorcomponent of 24 kDa (about 20%). The immunoblotin Fig. 2B shows that only the major component at25 kDa cross-reacts with the anti-Alpha GSTantibody, whereas neither the 25 kDa nor the 24
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

5
kDa component react with the anti-Mu GSTantibody. The minor component, which neitherbelongs to the Alpha nor to the Mu class, has not yetbeen analyzed further. To identify the specificAlpha isoenzyme(s) involved in the weak and tightassociation, a simple reversed phase HPLC analysiswas performed, according to the proceduredescribed by Yeh et al. (1). The weakly bound GSTsare mainly represented by GSTA1-1, GSTA2-2 andGSTA3-3, approximately with the same relativeabundance as found in the cytosol. Conversely, thetightly bound GSTs are mainly represented byGSTA1-1 and GSTA3-3 (Fig. 3).
Direct observation of the association of Alpha GSTsto the nuclear membrane – Direct evidence ofintranuclear and perinuclear association of AlphaGSTs is provided by confocal fluorescencemicroscopy using an anti-Alpha GST antibody and asecondary fluorescent antibody. All nuclei displayintense staining at the periphery but also inside thenuclei (Fig. 4A). NBDHEX, a specific fluorescentprobe for Alpha GST, gives further confirmation. Ithas recently been demonstrated that this fluorescentcompound, able to trigger apoptosis in human tumorcells, acts as a strong inhibitor of Alpha, Pi and MuGSTs by binding tightly to the active site of theseenzymes (31). Interaction of this compound withMu and Pi GSTs causes a dramatic loss of itsintrinsic fluorescence, while the fluorescencespectrum is almost unchanged when NBDHEXbinds to Alpha GST (KD = 5.3 × 10-6 M) (31).Incubation of intact nuclei with 10 µM NBDHEXcauses a distinct accumulation of fluorescence at thenuclear membrane, and also faint fluorescenceinside the nucleus (Fig. 4B). When the sameexperiment is performed with nuclei extensivelywashed with 10 mM potassium phosphate buffer,only faint intranuclear fluorescence is observed (notshown).
Electrostatic binding of Alpha GST to the nuclearmembrane: reality or artifact? – The data reportedabove prove that a part of the intracellular GST poolis associated to the nucleus. However, they do notexclude the possibility that this association mightnot be present in the intact hepatocyte, but isinduced artificially by the homogenization insucrose in the absence of buffer or other inorganicsalts. Convincing evidence that this interaction
really occurs in intact cells is achieved usingNBDHEX as a fluorescent intracellular marker forAlpha GSTs. NBDHEX not only specifically labelsGSTs but also accumulates in the cell within a fewminutes (32). After exposure of rat hepatocytes to0.1 mM NBDHEX, faint fluorescence is visible inthe cytosol, but also a strong staining of the nuclei,mainly localized on the nuclear envelope (Fig. 4C).To verify if NBDHEX labels selectively GSTs andto prove the absence of non-specific interactionswith the nuclear membrane, rat hepatocytes wereincubated with 1 mM GSNO after treatment withNBDHEX. As shown in the accompanying paper(4), this compound causes the formation of about0.15 mM of DNDGIC inside the cell. As DNDGICbinds to Alpha GSTs with an affinity a thousandtimes higher than NBDHEX (KD = 10-10 M), the ironcomplex will displace this fluorescent label. Thefluorescence observed after exposure to NBDHEXand localized near the nuclear envelope fades almostcompletely after 1 hour of incubation with 1 mMGSNO (Fig. 4D). The absence of non-specificbinding of NBDHEX to the membrane is alsodemonstrated by exposing a human tumor cell line(CEM) to NBDHEX. These tumor cells do notexpress Alpha or Mu GSTs but exclusively the Piclass GSTP1-1. After incubation with 50 µMNBDHEX, the nuclei appear like black holes, whilea detectable fluorescence is visible in the cytosol,indicating that NBDHEX enters the cells (Fig. 4E).
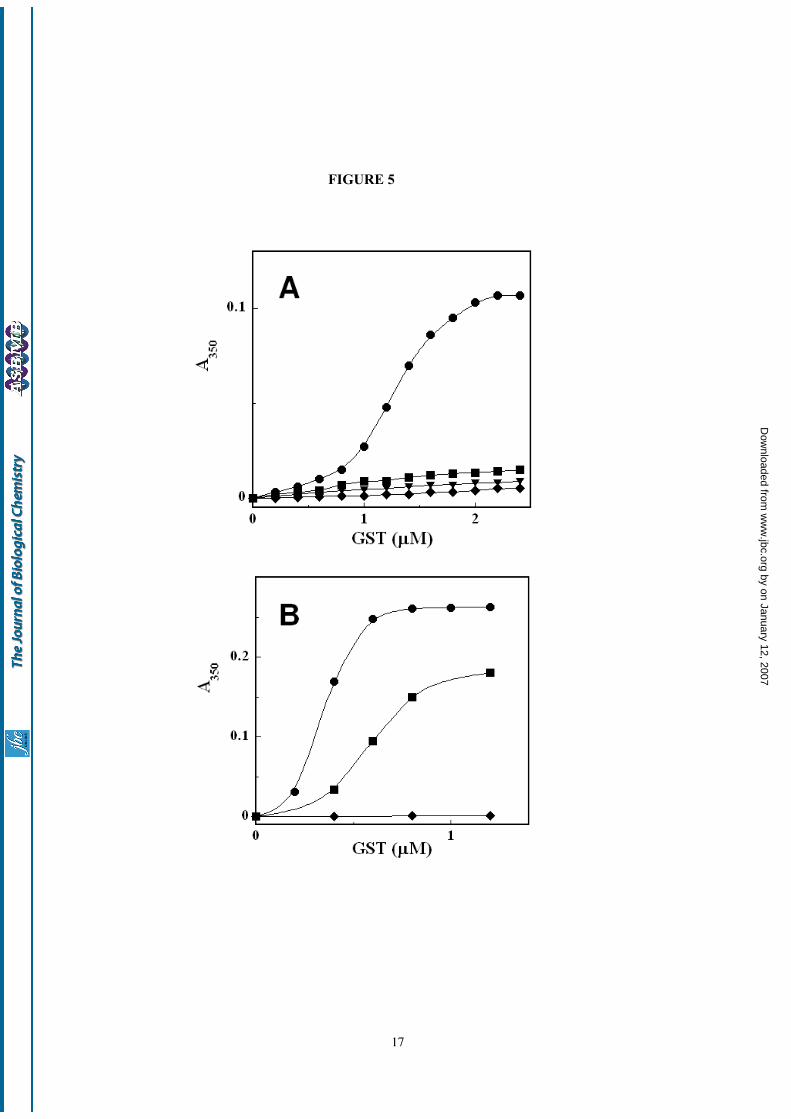
Interaction of GSTs with model membranes –Liposomes were used to confirm the electrostaticassociation of Alpha GSTs to the nuclearmembrane. Addition of increasing amounts ofGSTA1-1 to a liposome suspension (lipidconcentration 40 µM) in the absence of salts (pH isset to 7.4 by suitable addition of KOH) causesvisible turbidity of the solution, clearly indicating aprotein-membrane interaction (Fig. 5). A similarbehavior is commonly observed when polycationsassociate electrostatically to vesicles containingacidic lipids, because this interaction perturbs thestability of the colloidal liposome suspension byshielding the electrostatic repulsion betweenliposomes (33). Very likely this is also the case withGSTA1-1, which has an isoelectric point of 8.9 (34),and which at pH 7.4 has a total charge of +10 (perdimer), as calculated with the PDB2PQR software(26). The hypothesis that vesicle aggregation is
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

6
induced by protein binding to the membrane andthat the origin of this interaction is essentiallyelectrostatic is confirmed by the absence ofaggregation with neutral liposomes, lacking anioniclipids (data not shown). Interestingly, at the samepH value this phenomenon is completely absentwith GSTM2-2 and GSTP1-1 (Fig. 5A), indicating aspecific interaction with Alpha GST, in agreementwith the isoenzyme composition of the nuclear GSTfractions. The lack of effect seen for Mu and Pi classenzymes is consistent with an electrostaticinteraction, since the isoelectric points of GSTM2-2and GSTP1-1 are 5.4 and 4.5, respectively (totalcalculated charges at pH 7.4 are –4 and –6, forGSTM2-2 and GSTP1-1, respectively).
This conclusion is further supported by thepH and ionic strength dependence of the liposomeaggregation process (Fig. 5B). By lowering the pHto 5.4, a lower concentration of GSTA1-1 is neededto induce vesicle aggregation, in agreement with thehigher total charge of the enzyme under theseconditions (+12). Interestingly, at pH 5.4 liposomeaggregation is caused also by GSTM2-2, which isclose to its isoelectric point at this pH. Finally, theaddition of 10 mM NaCl completely inhibits vesicleaggregation at pH 7.4 in the presence of GSTA1-1(Fig. 5A). Since a relatively high ionic strengthreduces the intensity of electrostatic interactions,this result cannot be due to a direct effect of theionic strength on the aggregation process (whichwould be favored by the reduced intervesiclerepulsion in the presence of salts), but only to thelack of protein-membrane association under theseconditions. This finding nicely parallels theobservation that dissociation of nuclear GSTs can beinduced by the addition of 10 mM potassiumphosphate.
Direct visualization of GSTA1-1 associationto anionic lipid membranes is obtained byemploying the so-called giant unilamellar vesicles(GUVs), i.e. liposomes with a diameter of severalmicrometers, which can be easily observed byoptical microscopy. In order to allow directvisualization by confocal fluorescence microscopy,GSTA1-1 and GSTM2-2 are labeled with NBD-Cl,a fluorescent probe (λem = 520 nm) that binds toproteins by reacting with lysines and cysteines. Welabel the proteins at pH 6.0 to achieve selectivealkylation of cysteines (35), in order to minimize theperturbation of the electrostatic potential of the
protein surface. As shown in Fig. 4F accumulationof GSTA1-1 on the surface of anionic GUVs isdemonstrated by strong NBD fluorescence. Thisphenomenon is absent with GSTM2-2 (Fig. 4G) orin the presence of salts (not shown).
Molecular modeling of GST electrostatics – A moredetailed analysis of the electrostatic properties of theGST isoenzymes provides further informationregarding the mode of association. In addition to anet positive charge, GSTA1-1 also exhibits anasymmetric distribution of electric charges, whichendows this protein with a strong electric dipole(1000 Debye, as calculated by the Dipole server), asshown in Fig. 4. Interestingly, this is not the case forthe other isoenzymes (the calculated electric dipolesof GSTM2-2 and P1-1 are 240 and 40 Debye,respectively). A further illustration of this point isprovided by a calculation of the electrostaticpotential generated by GSTs, performed by solvingthe Poisson-Boltzmann equation with the APBSsoftware (27). Fig. 4 shows the isopotential surfacescorresponding to +3 kT/e (in blue) and –3 kT/e (inred), demonstrating a significant positive potentialon the surface of the dimer located on the proteinside opposite to the active sites. Very likely this isthe protein region interacting electrostatically withthe membrane. In agreement with this conclusion, apositive potential region is not present in the otherisoenzymes, which do not interact with the nuclearmembrane or with liposomes. It is worth noting thatvery similar results were obtained for both humanand rat enzymes (data not shown).
DISCUSSION
Data reported in this paper clearly show thatspecific GSTs are involved in subcellularcompartmentalization in rat hepatocytes. About 20%of cytosolic Alpha class GSTs are electrostaticallyassociated to the outer nuclear membrane and anequivalent amount is present in the nuclear interior.The use of 0.25 M sucrose without exogenous saltsfor nuclei isolation, and the particular assay for GSTactivity of intact nuclei, made it possible to discoverthis double modality of association that, to a lesserextent, may also be present in other subcellularcomponents (see Table 1). Previous observationscontrary to the nuclear compartmentalization ofGST were probably due to the use of standard
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

7
subcellular fractionation procedures that caused theloss of the electrostatically bound GST, and to theextensive inactivation that occurs during sonicationof nuclei or extraction with detergents.Immunostaining and chromatographic data indicatethat GSTA1-1. GSTA2-2 and GSTA3-3 are theenzymes mainly involved in the nuclear association.Confocal microscopy and the use of NBDHEX, aspecific fluorescent probe for Alpha GSTs, give adirect visualization of the presence of theseisoenzymes both at the outer nuclear membrane andin the nuclear interior. The electrostatic binding ofAlpha GSTs observed in isolated nuclei is wellreproduced with liposomes, and the results alsoconfirm that Mu and Pi class GSTs do not interact atphysiological pH values. Although our experimentswith liposomes have been done with GSTA1-1, asimilar behavior is expected for GSTA2-2 andGSTA3-3 that display very similar electrostaticproperties. Finally, electrostatic potentialcalculations, performed on Alpha, Mu and Pi GSTsindicate that only Alpha GSTs display a properelectrostatic potential at the protein surface tointeract with the negatively charged membrane. Inthis context, another peculiar feature of the Alphadimer is the strong dipolar character that is notfound in the other GST isoenzymes.
An examination of the present data disclosesa surprising scenario. The amount of the cytosolicAlpha GSTs is about 2.2 mg/g of liver (reaching aconcentration of 0.3 mM in the cytosol, 43% of thetotal GSTs) while about 0.5 mg is electrostaticallyassociated to the nuclear membrane and anequivalent amount is probably compartmentalizedinside the nucleus. As the nuclear volume is about10% of the cytosolic volume, it results that theintranuclear GST concentration is approximately 0.7mM, a value comparable to that found in cytosol.This concentration may reflect a free diffusionalin/out traffic of Alpha GSTs between the cytosolicand nuclear compartments. Notably, the nuclearaccess seems to be denied for Mu GSTs. In contrast,the local concentration of Alpha GSTs at the outernuclear membrane will be much higher. Anestimation of the minimum surface occupied by allelectrostatically bound GSTA1-1 assembled in alayer (calculated on the basis of an area of 19.6 nm2
for each GSTA1-1 dimer, based on the X-raystructure) paradoxically results about five timeslarger than the surface of the entire nuclear
membrane (Table 2). In addition, the specificactivity of the Alpha GSTs, extracted from thenuclear membrane after salt treatment, is about 3U/mg, five times lower than the one of the purifiedenzymes (16 U/mg). Thus, other proteins must beelectrostatically associated to the nuclear envelopetogether with GSTs; this appears to be an additionalparadox, given the absence of free membrane areafor further electrostatic interactions. One possibleexplanation for this excessive amount of boundproteins is that Alpha GSTs could be assembled in amultilayer disposition near the nuclear membrane inan alternate sequence with negatively chargedproteins. This peculiar onion layer-like assembly ofproteins is not a complete novelty. It has beendemonstrated in vitro that positively chargedproteins easily aggregate in such a way to negativelycharged colloidal particles in the presence of anionicpolyelectrolytes (36, 37) and our preliminary resultsalso indicate a prevalent anionic nature of theaccompanying proteins. To our knowledge, thepresent data represent the first indication for theexistence in vivo of a similar protein assembly, thatobviously needs further confirmation. Thepossibility that a few cytosolic enzymes are not in acompletely disordered distr ibution butelectrostatically ordered near specific intracellularmembranes opens a new and fascinating scenario incell physiology. Is it possible that this hypotheticalprotein shell escaped visualization by advancedspectroscopy and microscopy techniques? Theanswer may be affirmative. We must remember thatthe protein concentration in the cytosol is above 300mg/ml, a border-line value for proteincrystallization. Thus all cytosolic proteins areextremely crowded but no spectroscopic ormicroscopic evidence has been reported for thisalmost crystal-like condition. Obviously in theabsence of a detectable selective marker no tracecan be expected if this crowding is formed byselected proteins near specific subcellularcomponents. Whatever the true assembly of AlphaGSTs at the nuclear envelope, the presence of largeamounts of this specific GSTs near the nucleusdemonstrated here is a novelty and probably has aphysiological finality. In fact all GSTs represent avery efficient defense system against many toxicalkylating compounds but Alpha class GSTs displayan additional and peculiar peroxidase activity, notshown by Mu and Pi GSTs, and eliminates
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

8
efficiently dangerous organic peroxides (2). Alphaclass GSTs also have a ten times higher affinity thanPi and Mu GSTs for DNDGIC, a natural carrier ofNO that displays strong oxidizing properties andinactivates irreversibly key enzymes like glutathionereductase (38, 39). Thus, the present results seem toindicate that the nucleus and its precious geneticcontent have a further mechanism of protection, notconsidered until now. Quite a few questions remainopen. For example, given the similar lipidcomposition of most intracellular membranes, it isnot clear why the Alpha GST displays a soimpressive accumulation on the nuclear membrane(see Table 2). Interestingly, it was shown many
years ago by Virtanen that the outside of the nucleusis strongly negatively charged while the innermembrane is practically neutral (40). However, it isalso possible that specific nuclear membraneproteins recognize Alpha GSTs and favor theiraccumulation. We did not explore in this paper thestatus of the tightly bound GSTs, that probablyreside in the nuclear interior. The curiously similaramount of the “external” and “internal” Alpha GSTscould be merely a coincidence, or it could reflect aspecular modality of interaction of Alpha GSTs withthe outer and inner membrane. Work is in progressto answer these questions.
REFERENCES
1. Yeh, H.-I., Hsieh, C.-H., Wang, L.-Y., Tsai, S.-P., Hsu, H.-Y., and Tam, M. F. (1995) Biochem. J. 308,69-75
2. Hurst, R., Bao, Y., Jemth, P., Mannervik, B., and Williamson, G. (1998) Biochem. J. 332, 97-100
3. Mannervik, B. (1986) Chem. Scripta 26, 281-284
4. Pedersen, J. Z., De Maria, F., Turella, P., Federici, G., Mattei, M., Fabrini, R., Dawood, K. F., Massimi,M., Caccuri, A. M., and Ricci, G. (2006) J. Biol. Chem. The accompanying paper
5. Morgenstern, R., DePierre, J. W., and Jornvall, H. (1985) J. Biol. Chem. 260, 13976-13983
6. Singh, S. P., Janecki, A. J., Srivastava, S. K., Awasthi, S., Awasthi, Y. C., Xia, S. J., and Zimniak, P.(2002) J. Biol. Chem. 277, 4232-4239
7 Morgenstern, R., Guthenberg, C., Mannervik, B., and DePierre, J. W. (1983) FEBS Lett. 160, 264-268
8. Sandeep, K., Prabou, K. S., Reddy, P. V., Gumpricht, E., Hildenbrandt, G. R., Scholz, R. W., Sordillo,L. M., and Reddy, C. C. (2001) Biochem. J. 360, 345-354
9. Campbell, J. A. H., Corrigal, A. V., Guy, A., and Kirsch, R. E. (1991) Cancer 67, 1608-1613
10. Abei, M., Harada, S., Tanaka, N., McNeil, M., and Osuga, T. (1989) Biochim. Biophys. Acta 995, 279-284
11. McCusker, F. M., Phillips, M. F., Boyce, S. J., and Mantle, T. J. (1990) Glutathione S-Transferases andDrug Resistance (Hayes, J. D., Pickett, C. B., and Mantle, T. J., eds.), Taylor and Francis, London
12. Soboll, S., Grundel, S., Harris, J., Kolb-Bachofen, V., Ketterer, B., and Sies, H. (1995) Biochem. J.311, 889-894
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

9
13. Rogers, L. K., Gupta, S., Welty, S. E., Hansen, T. N., and Smith, C. V. (2002) Toxicol. Sci. 69, 279-285
14. Bennett, C. F., Spector, D. L., and Yeoman, L. C. (1986) J. Cell. Biol. 102, 600-609
15. Rozell, B., Hansson, H. A., Guthenberg, C., Tahir, M. K., and Mannervik, B. (1993) Xenobiotica 23,835-849
16. Turella, P., Cerella, C., Filomeni, G., Bullo, A., De Maria, F., Ghibelli, L., Ciriolo, M. R., Cianfriglia,M., Mattei, M., Federici, G., Ricci, G., and Caccuri, A. M. (2005) Cancer Res. 65, 3751-3761
17. Lo Bello, M., Nuccetelli, M., Caccuri, A. M., Stella, L., Parker, M. W., Rossjohn, J., McKinstry, W. J.,Mozzi, A. F., Federici, G., Polizio, F., Pedersen, J. Z., and Ricci, G. (2001) J. Biol. Chem. 276, 42138-42145
18. Board, P. G., and Pierce, K. (1987) Biochem. J. 248, 937-941
19. Ross, V. L., and Board, P. G. (1993) Biochem. J. 294, 373-380
20. Turella P., Cervella C., Filomeni G., Bullo A., De Maria F., Ghibelli L., Circolo M.R., Cianfriglia M.,Mattei, M., Federici G., Ricci G., and Caccuri, A. M. (2005) Cancer Res. 65, 65, 3751-3761
21. Simmons, P. C., and Vander Jagt, D. L. (1977) Anal. Biochem. 82, 334-341
22. Stella, L., Mazzuca, C., Venanzi, M., Palleschi, A., Didone, M., Formaggio, M., Toniolo, C., andPispisa, B. (2004) Biophys. J. 86, 936-945
23. Stewart, J. C. M. (1980) Anal. Biochem. 104,10-14
24. Tandford, C. (1973) The Hydrophobic effect: formation of Micelles and Biological Membranes, Wileyand sons, eds. New York
25. Estes, D. J., and Mayer, M. (2005) Colloids Surf. B 42, 115-123
26. Dolinsky, T. J., Nielsen, J. E., McCammon, J. A., and Baker, N. A. (2004) Nucleic Acids Res. 32,W665-W667
27. Baker, N. A., Sept, D., Joseph, S., Holst, M. J., and McCammon, J. A. (2001) Proc. Natl. Acad. Sci.USA 98, 10037-10041
28. Koradi, R., Billeter, M., and Wüthrich, K. (1996) J. Mol. Graphics 14, 51-55
29. Pettersen, E. F., Goddard, T. D., Huang, C. C., Couch, G. S., Greenblatt, D. M., Meng, E. C., andFerrin, T. E. (2004) J. Comput. Chem. 25, 1605-1612
30. De Maria, F., Pedersen, J. Z., Caccuri, A. M., Antonini, G., Turella, P., Stella, L., Lo Bello, M.,Federici, G., and Ricci, G. (2003) J. Biol. Chem. 278, 42283 - 42293
31. Ricci, G., De Maria, F., Antonini, G., Turella, P., Bullo, A., Stella, L., Filomeni, G., Federici, G., andCaccuri, A. M. (2005) J. Biol. Chem. 280, 26397-26405
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

10
32. Turella, P., Filomeni, G., Dupuis, M. L., Ciriolo, M., Molinari, A., De Maria, F., Tombesi, M.,Cianfriglia, M., Federici, G., Ricci, G., and Caccuri, A. M. (2006) J. Biol. Chem. 281, 23725-23732
33. Arnold, K., and Krumbigel, O. M. (1990) Chem. Phys. Lipids 55, 301-307
34. Stockman, P. K., Mclellan, L. I., and Hayes, J. D. (1987) Biochem. J. 244, 55-61
35. Del Boccio, G., Pennelli, A., Whitehead, E. P., Lo Bello, M., Petruzzelli, R., Federici, G., and Ricci, G.(1991) J. Biol. Chem. 266, 13777-13782
36. Caruso, F. (2004) Colloids and Colloid Assemblies, Wiley-VCH, Mannheim
37. Ai, H., Jones, S. A., and Lvov, Y. M. (2003) Cell. Biochem. Biophys. 39, 23-43
38. Biese, M., Keese, M. A., Becker, K., Büsse, R., and Mülsch, A. (1997) J. Biol. Chem. 272, 21767-21773
39. Becker, K., Savvides, S. N., Keese, M., Schimer, R. H., and Karplus, P. A. (1998) Nature Struct. Biol.5, 267-271
40. Virtanen, I. (1978) Cell Biol. Int. Rep. 2, 33-39
41. Alberts, B., Bary, D., Lewis, J., Raff, M., Roberts, K., and Watson, J. D. (1984) Molecular Biology ofthe cell, Garland Pubs., New York
42. Mannervik, B., Board, P. G., Hayes, J. D., Listowsky, I., and Pearson, W. R. (2005) Methods Enymol.401, 1-8
FOOTNOTES
Acknowledgements: We thank F. Cavalieri, G. Paradossi and M. Ranalli for their kind assistance with theconfocal microscope and prof. A. Trentalance for her helpful discussion of the manuscript.
1Abbreviations used are: CDNB, 1-chloro-2,4-dinitrobenzene; DNDGIC, dinitrosyl-diglutathionyl-ironcomplex; DNGIC, dinitrosyl-glutathionyl-iron complex; DNIC: dinitrosyl-iron complex; EPR, electronparamagnetic resonance; GSNO, S-nitrosoglutathione; GST, glutathione transferase; GUV, giant unilamellarvesicle; LUV, large unilamellar vesicle; NBD-Cl, 4-chloro-7-nitro-2,1,3-benzoxadiazole; NBDHEX, 6-(7-nitro-2,1,3-benzoxadiazol-4-ylthio)hexanol.
FIGURE LEGENDS
FIGURE 1. Titration of the nuclear GST with DNDGIC – The nuclear pellet isolated in 0.25 M sucrose isincubated with ten volumes of 10 µM DNDGIC at pH 7.4. After removal of the free complex by G-25Sephadex chromatography, GST activity under isotonic conditions and EPR quantifications are performed asdescribed under “Experimental Procedures”. The same experiment is performed with the nuclear pellet isolated
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

11
in 10 mM potassium phosphate buffer, pH 7.4 and 0.25 M sucrose. Data are the mean ± SD of threeexperiments.
FIGURE 2. SDS-PAGE and Immunoblotting – Panel A: SDS-PAGE. Lane 1: protein markers; lane 2: purifiedhuman GSTA1-1; lanes 3 and 7: purified pool of cytosolic GSTs isoenzymes; lanes 4 and 8: weakly boundGST; lanes 5 and 9: tightly bound GST; lane 6: purified human GSTM2-2; lane 10: GSTP1-1 used as negativecontrol. Panel B: Immunoblotting using anti-Alpha GST (left side) and anti-Mu GST (right side).
FIGURE 3. HPLC analysis – Tightly and weakly bound GSTs, extracted as reported under “ExperimentalProcedures” were purified through affinity chromatography and analyzed by reverse phase HPLC as describedpreviously (1). Identification of GST isoenzymes were made according to (1). Upper panel: representativechromatogram of the cytosolic GSTs, previously purified through two subsequent affinity chromatographysteps on glutathione-sepharose matrix (21)(recovery ≥ 93%). Panel A: Relative amounts of the five cytosolicisoenzymes present in the GST pool. Isoenzymes below 2% are not reported. The total amount of cytosolicGSTs is 5.1 mg per gram of liver. Panel B: relative amounts of isoenzymes of the weakly bound GSTs. Thetotal amount of the weakly bound GSTs is 0.5 mg per gram of liver, corresponding to 10% of the cytosolicGSTs. Panel C: relative amounts of isoenzymes of the tightly bound GSTs. The amount of the tightly boundGSTs after partial extraction (60%) is 0.3 mg per gram of liver, corresponding to 6% of the cytosolic GSTs.The nomenclature used for rat GST isoenzymes is in accordance with the one proposed recently (42).Percentages represent the mean of three experiments.
FIGURE 4. Immunofluorescence, fluorescence label experiments and electrostatic calculations – Upper panel:A: high-power oil-immersion confocal micrograph of nuclei pellets immunoreacted with anti-Alpha GST,diluted 1:100, followed by fluorescein conjugated secondary antibody (for details see ExperimentalProcedures). Bar, 5 µm; B: nuclei pellets incubated with NBDHEX. Bar, 10 µm; C: intact rat hepatocytesincubated with NBDHEX. Bar, 12 µm. The inset shows a two fold magnification; D: rat hepatocytes, treated asin C and exposed to 1 mM GSNO for 1 hour. Bar, 20 µm; E: low-power confocal micrograph of CEM cellsincubated with NBDHEX. Bar, 20 µm; F: GUVs after interaction with GSTA1-1 labelled with NBD-Cl; G: asin F after interaction with GSTM2-2 labelled with NBD-Cl; H: GUVs control showing only rhodaminefluorescence. Lower panel: Molecular models represent the crystal structures of GSTA1-1, GSTM2-2 andGSTP1-1 and their isopotential surfaces corresponding to +3 kT/e (in blue) and –3 kT/e (in red). Only GSTA1-1 shows a significant positive potential on the surface of the dimer located on the protein side opposite to theactive sites. In the model of hGSTA1-1 a dipolar moment of 1000 Debye was calculated and represented as ared arrow inside the dimer.
FIGURE 5. Electrostatic association of GSTA1-1, GSTM2-2 and GSTP1-1 to liposomes as followed byturbidimetric measurements – Panel A: Anionic liposomes (40 µM phospholipids) were incubated with variableamounts of purified human GSTs at pH 7.4. Turbidity changes was monitored at 350 nm. (●): GSTA1-1; (■):GSTM2-2; (◆): GSTP1-1. (▼): GSTA1-1 in the presence of 10 mM NaCl; Panel B: the same experiment as inA, performed at pH 5.4.
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

12
TABLE 1
Subcellular localization of GST activitya
GST(U/g tissue)
Cell components
SucroseSucrose + potassium
phosphate buffer
CytosolNucleiMitocondriaLysosomesMicrosomes
101 ± 8 16 ± 2
1.6 ± 0.4 2.9 ± 0.7
4.2 ± 1
110 ± 10 8 ± 2
0.7 ± 0.3 1.6 ± 0.2 2.1 ± 0.4
aActivity measurements were done on intact organelles, isolated in pure sucrose (0.25 M) or in 0.1 M sucrose,0.05 M KCl and 0.04 M KH2PO4, pH 7.4, using the isotonic assay medium described under “ExperimentalProcedures” section.
TABLE 2
Quantitative analysis of GSTs bound electrostatically to subcellular membranes
Cellcomponents
Membrane areaa
(m2 /g )GST /membrane area
(mg/m2)Area of GST/
membrane area
NucleiMitocondriaLysosomesMicrosomes
0.0220.770.044
5.5
22.70.062.4
0.02
50.010.5
0.01
aData derived from ref. 41.
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from

13
FIGURE 1
0
20
40
60
80
100
Nuclei in sucrose
Nuclei in sucrose + DNDGIC
Nuclei in sucrose + DNDGIC
Nuclei in sucrose
plus buffer
Nuclei in sucroseplus buffer+ DNDGIC
■ = GST activity
■ = DNGIC-GST complex
%
by on January 12, 2007 w
ww
.jbc.orgD
ownloaded from




















