
Electrophysiology of the Right Anterior Approach to the Atrioventricular Node:.: Studies In Vivo and...
16
47 Electrophysiology of the Right Anterior Approach to the Atrioventricular Node: Studies In Vivo and in the Isolated Perfused Dog Heart MATTHIAS ANTZ, M.D., BENJAMIN J. SCHERLAG, PH.D., EUGENE PATTERSON, PH.D.. KENICHIRO OTOMO. M.D., CLAUDIO TONDO. M.D., JAN PITHA, M.D., PH.D..I= MARIO D. GONZALEZ. M.D. WARREN M. JACKMAN, M.D., and RALPH LAZZARA, M.D. From the Cardiovascular Section. Departments of Medicine and *Pathology. University of Oklahoma Health Sciences Center, and Department of Veterans Affairs Medical Center. Oklahoma City, Oklahoma Electrophysiology of Anterior AVN Input. Introduction: Previous reports have described electrophysiologic properties and rate-dependent responses in the transitional cell zone of the pos- terior AV nodal input (slow pathway). The purpose of this study was to investigate the electro- physiology of the anterior transitional cell zone (fast pathway) in vivo and in a Langendorff prepa- ration perfused with a nonhlood solution containing hutanedionemonoxime to inhihit contraction. Methods and Results: In five anesthetized dogs, the His-bundle electrogram recorded from the aortic root included atrial activity in close proximity to the anterior limbus of the fossa ovalis. Dur- ing decrementai atrial pacing, the atrial potential exhihited amplitude alternans at a pacing cycle length (CL) of 135 ± 14 msec. In ten isolated pertused canine hearts, a bipctlar electrode catheter was positioned with its tip against the right anterior interatrial septum just superior to the tendon of Todaro. The AV Wenckebach CL (WCL) averaged 262 ± 21 msec. During further decreases in pacing CL, the bipolar atrial potential developed a 2:1 amplitude alternans (9/10 dogs) at CL = 168 ± 15 msec and then split into two components with subsequent 2:1 block between these compo- nents (10/10 dogs) at CL = 152 ± 19 msec. Radiofrequency ablation at this site in six dogs pro- longed the stimulus to HB interval from 113 ± 19 to 151 ± 30 msec (P < 0.01) without changing the WCL, consistent with ablation of the fast AV nodal pathway. In six other isolated perfused canine hearts, an octapolar catheter (2-mm spacing) was positioned along the anterior limbus of the fossa ovalLs with the tip electrode located over the anterior portion (apex) of the triangle of Koch. The aforementioned 2:1 amplitude alternans occurred at a longer CL in the distal electrodes ltKated at the tendon of Todaro than in the proximal electrodes at farther distances from the tendon of Todaro (185 ± 25 vs 171 ± 20 msec, P < 0.05). as did the 2:1 block between tbe two components (161 ± 18 vs 150 ± 18 msec, P < 0.05). Microelectrode recordings obtained adjacent to tbe catheter demonstrated 2:1 alternans and block patterns in the action potentials of transitional cells but not in atrial cells, which exhibited 1:1 conduction at all CL. Conclusions: The transitional cell zone in the anterior Interatrial septum exhibits a specific rate-dependent, spatial gradient of conduction block, which can be recorded in bipolar electro- grams as well as microelectrode recordings. Electrophysiologic changes induced by radiofre- quency ablation of this anterior atrial/transitional cell zone (corrohoraled by histology) pro- vide strong presumptive evidence tbat this area constitutes all or a major part of the fast AV nodal pathway. (J Cardiovasc Electrophysiol Vol. 8. pp. 47-61, January 1997) atrioventricular conduction, atrioventricular node, fast pathway, radiofrequency ablation, canine Langendorff preparation, microelectrode recordings, transitional cells Dr. Antz is supported by a fellowship gram from the •'Deutsche OU Health Sciences Center. VA Medical Cenler (I51-F). 921 Forschungsgemein.schafi." Bonn. Germany. Dr. Tondo is the re- N.E. !3th Street. Okhthoma City, OK 7.M04. Fax: 405-270- cipient of a Bristol-Myers Squibb Cardiovascular International 5132. Fellowship Award. Manuscript received 17 May 19%; Accepted for publtcation u Address for correspondence: Benjamin J. Scherlag, Ph.D.. August
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Electrophysiology of the Right Anterior Approach to the Atrioventricular Node:.: Studies In Vivo and...

47
Electrophysiology of the Right Anterior Approach to theAtrioventricular Node: Studies In Vivo and in the Isolated
Perfused Dog Heart
MATTHIAS ANTZ, M.D., BENJAMIN J. SCHERLAG, PH.D.,
EUGENE PATTERSON, PH.D.. KENICHIRO OTOMO. M.D.,CLAUDIO TONDO. M.D., JAN PITHA, M.D., PH.D..I= MARIO D. GONZALEZ. M.D.
WARREN M. JACKMAN, M.D., and RALPH LAZZARA, M.D.
From the Cardiovascular Section. Departments of Medicine and *Pathology. University of OklahomaHealth Sciences Center, and Department of Veterans Affairs Medical Center. Oklahoma City, Oklahoma
Electrophysiology of Anterior AVN Input. Introduction: Previous reports have describedelectrophysiologic properties and rate-dependent responses in the transitional cell zone of the pos-terior AV nodal input (slow pathway). The purpose of this study was to investigate the electro-physiology of the anterior transitional cell zone (fast pathway) in vivo and in a Langendorff prepa-ration perfused with a nonhlood solution containing hutanedionemonoxime to inhihit contraction.
Methods and Results: In five anesthetized dogs, the His-bundle electrogram recorded from theaortic root included atrial activity in close proximity to the anterior limbus of the fossa ovalis. Dur-ing decrementai atrial pacing, the atrial potential exhihited amplitude alternans at a pacing cyclelength (CL) of 135 ± 14 msec. In ten isolated pertused canine hearts, a bipctlar electrode catheterwas positioned with its tip against the right anterior interatrial septum just superior to the tendonof Todaro. The AV Wenckebach CL (WCL) averaged 262 ± 21 msec. During further decreases inpacing CL, the bipolar atrial potential developed a 2:1 amplitude alternans (9/10 dogs) at CL =168 ± 15 msec and then split into two components with subsequent 2:1 block between these compo-nents (10/10 dogs) at CL = 152 ± 19 msec. Radiofrequency ablation at this site in six dogs pro-longed the stimulus to HB interval from 113 ± 19 to 151 ± 30 msec (P < 0.01) without changing theWCL, consistent with ablation of the fast AV nodal pathway. In six other isolated perfused caninehearts, an octapolar catheter (2-mm spacing) was positioned along the anterior limbus of the fossaovalLs with the tip electrode located over the anterior portion (apex) of the triangle of Koch. Theaforementioned 2:1 amplitude alternans occurred at a longer CL in the distal electrodes ltKated atthe tendon of Todaro than in the proximal electrodes at farther distances from the tendon ofTodaro (185 ± 25 vs 171 ± 20 msec, P < 0.05). as did the 2:1 block between tbe two components(161 ± 18 vs 150 ± 18 msec, P < 0.05). Microelectrode recordings obtained adjacent to tbe catheterdemonstrated 2:1 alternans and block patterns in the action potentials of transitional cells but notin atrial cells, which exhibited 1:1 conduction at all CL.
Conclusions: The transitional cell zone in the anterior Interatrial septum exhibits a specificrate-dependent, spatial gradient of conduction block, which can be recorded in bipolar electro-grams as well as microelectrode recordings. Electrophysiologic changes induced by radiofre-quency ablation of this anterior atrial/transitional cell zone (corrohoraled by histology) pro-vide strong presumptive evidence tbat this area constitutes all or a major part of the fast AVnodal pathway. (J Cardiovasc Electrophysiol Vol. 8. pp. 47-61, January 1997)
atrioventricular conduction, atrioventricular node, fast pathway, radiofrequency ablation, canineLangendorff preparation, microelectrode recordings, transitional cells
Dr. Antz is supported by a fellowship gram from the •'Deutsche OU Health Sciences Center. VA Medical Cenler (I51-F). 921Forschungsgemein.schafi." Bonn. Germany. Dr. Tondo is the re- N.E. !3th Street. Okhthoma City, OK 7.M04. Fax: 405-270-cipient of a Bristol-Myers Squibb Cardiovascular International 5132.Fellowship Award.
Manuscript received 17 May 19%; Accepted for publtcation uAddress for correspondence: Benjamin J. Scherlag, Ph.D.. August

48 Journal of Cardiovascular Electrophysiology Vol. 8. No. I. January 1997
Introduction
The cure of AV nodal reentrant tachycardia(AVNRT) by radiofrequency catheter ablationhas renewed interest in the anatomy and physiol-ogy of the AV node and its atrial inputs. Potentialsarising from the atrial input to the slow pathwaywere described in 1992.'-
Less attention has been directed to the anteriorapproaches to the AV node.'"^ The purpose of thepresent study was lo examine the electrophysio-logie properties and histologic characteristics ofthe right anterior approach to the AV node in thein vivo and ex vivo dog heart. We attempted toelueidate the types of cells (i.e., atrial or transi-tional) found in this zone and to determine theirrate-dependent responses. The possible relation-ship of this transitional zone to the fast pathwayinput to the AV node was further explored by theeffects of radiofrequency ablation.
Methods
In Vivo Studies
Twenty-four mongrel dogs weighing between15 and 25 kg were anesthetized with sodiumpentobarbital (30 mg/kg. followed by 50 to 100mg as needed to maintain anesthesia) and me-chanically ventilated with room air. A right tho-racotomy was performed in the fifth intereostalspaee. and the heart was exposed by excision ofthe upper pulmonary lobe and periciirdiotomy. Epi-eardial plaque electrodes were sutured to the highright atrium (HRA) at its confluence with the lat-eral margin of the superior vena cava and to thecrux of the heart near the os of the coronary si-nus (CS). A pair of plunge wire electrodes was in-serted in the right ventricular outflow tract (RVOT)for ventricular pacing. The sinus node was crushedwith a Satinsky clamp, and atiial pacing was per-formed from the HRA electrode. The followingvessels were surgically exposed and cannulated:the left femoral arteiy for recording arterial pres-sure, the left external juguhir vein for drug deliv-ery, and the left common carotid artery for ad-vancing a hexapolar electrode catlieter into the non-coronary cusp of the aortic root to record theHis-bundle eleetrogram.^
HRA pacing was perfonned at a constant cy-cle length (CL) of 400 msec to determine the stim-ulus to His-bundle (S-H) interval. During pro-grammed stimulation of the HRA and RVOT(basic drive CL 4(K) m.sec), the effective refractory
period of the atrium (AERP) and the ventricle(VERP) were evaluated. Dect^mental atrial pacingwas used to deteniiine the Wenckebach cyele length(WCL). Simultaneous i-ecordings of ECG leads IIand aVR, and the His-bundle. CS, and HRA elec-trognuns were displayed on a monitor and storedon an optical disk (Bard Electrophysiology, Tewks-bury, MA. USA). The endwardial and epicai'dialrecordings were filtered at 30 to 5CX) Hz.
Catheter Recordings In Vivo
Fibers ofthe anterior limbus ofthe fossa ovalisfonning the right anterior approach to tlic AV nodeare located adjacent to the aortic wall just aboveits noneoronary cusp. Due to tbis proximity, elee-trogranis from the aortic root record potentials thatarise from the anterior limbus of the fossa ovalis.In six dogs, the His catheter was placed in the cor-responding area by manipulating the calheier inthe aortic root until a large atrial potential and asmall His-bundle potential were recorded. Simi-lar electrograms are obtained from the right airialendocardium from sites at whieh fast pathway ab-lation is successfully performed in patients."
Deereinental HRA pacing was performed be-ginning at a CL of 400 msec and decreasing theCL in 10-msec decrements to 100 msec or untilatrial fibrillation was induced. For better identifi-cation of the atrial potentials at the shorter CLs,transient stimtilation of the isolated vagosympa-thetic trunk was performed (20 Hz. 1 to 10 V. 0.5msec) to transiently prcxluee high-grade AV block.Electrograms were evaluated only if the catheterwas in a stable position as defined by the ab-sence of any detectable change in morphology ofthe His bundle and ventricular potentials.
Ex Vivo Studies
After intravenous administration of heparin(3,(KX) units), the hearts were excised and imme-diately submerged in iced oxygenated (95% 0,-5%CO;) modified Tyrode's solution with the follow-ing compt)sition: 117 mM NaCI, 4.1 mM KCl, 0.5niM MgCK. 1.35 mM CaCl^. 24 mM NaHCO,,1.8 mM NaHiPOj. 5.5 mM dextrose, 2 mM stxliumpyruvate, 0.6 ;j,M albumin, and 10 to 15 mM bu-tanedionemonoxime (DAM). This small amountof albumin has been shown to significantly increasethe stability of a Langendorff preparation.^ DAMwas added because of its marked negative inotropicproperties.*^ The concentration of DAM used {10to 15 mM) reduces myocardial contraetion and al-
Antz. etal. Electrophysiology of Anterior AVN Input 49
lows stable catheter positions and microelectrodeimpalements without causing major electrophysi-ologic changes.-^-^
The a.scending aorta of the excised canine heartwas cannulated and perfused with oxygenated,wanned (37° ± 0.5°C) modified Tyrode's solutionof the same composition as described previously (pH7.40 ± 0.05). Perfusion was pertormed at a con-stant pressure of 90 to 1(X) mmHg. The flow waskept stable at a rate of 2(X) to 250 mLVmin to avoidllow-dependent electrophysiologic changes.'" Aftertlie initial washout of blood, the heart was transferredinto a tissue bath that kept the myocanJium moistand wium. The incxiified Tyrode's solution was re-circulated using a peristaltic pump. Pin electrodeswere inserted into the plastic surface of the tissuebath to construct Einthoven's triangle for recordingECGs that were similar to those obtained in vivo.
Stability of the Langendorff Preparation
In eight canine hearts, the AERP and VERP,WCL, and S-H interval during HRA stimulation(basic pacing CL 400 msec) as well as the pres-ence of ventriculoatrial (VA) conduction duringventricuUu- pacing at a CL of 400 msec were as-sessed in vivo and at 30-minute intervals duringperfusion of the isolated heart. After a median du-ration of 3.5 hours of perfusion (range 2.5 to 6hours), epinephrine (0.33 /iM) was added to theperfusate. and the WCL, S-H interval, and thepresence of VA conduction during ventricular pac-ing at a CL of 400 msec were again assessed.
Catheter Recordings and Radiofrequency AblationEx Vivo
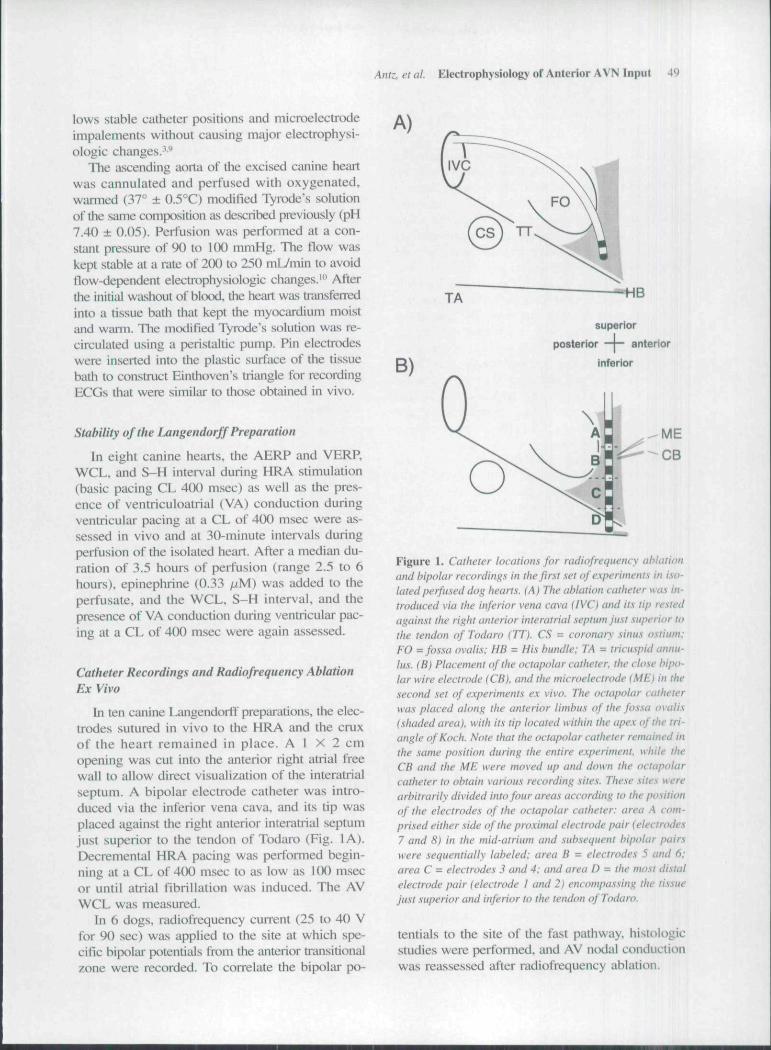
In ten canine LangendorfF preparations, the elec-trodes sutured in vivo to the HRA and the cruxof the heart remained in place. A 1 X 2 cmopening was cut into the anterior right atrial freewall to allow direct visualization of the interatrialseptum. A bipolar electrode catheter was intro-duced via the inferior vena cava, and its tip wasplaced against the right anterior interatrial septumjust superior to the tendon of Todaro (Eig. I A).Decremental HRA pacing was performed begin-ning at a CL of 400 msec to as low as lCX) msecor until atrial fibrillation was induced. The AVWCL was measured.
In 6 dogs, radiofrequency current (25 to 40 Vfor 90 sec) was applied to the site at which spe-cific bipolar potentials from the anterior transitionalzone were recorded. To correlate the bipolar po-
A)
B)
•iHB
superior
posterior — j — anterior
inferior
MECB
Figure 1. Catheter locations for radiofrequency ahlationand hipolar recordings in the first set ofe.Kperinwnts in iso-lilted perfused dog hearts. {A) The ablation catheter was in-troduced via the inferior vena cava (IVC) and its tip re.'itedagainst the right anterior interatrial septum just superior tathe tendon of Todaro (TT). CS = coronary sinus ostiurn:FO = fossa ovalis: HB = His bundle: TA = tricuspid annu-lus. (B) Placement of the octapolar catheter, the clo.se hipo-lar wire electrode (CB). and the microelectrode (ME) in the.second .set of experiments ex vivo. The octapolar eatheterwas placed along the anterior limbus of the fossa ovalis(shaded area), with its tip located within the apex of the tri-angle of Koch. Note that the octapolar catheter remained inthe .same position during the entire experiment, while theCB and the ME were moved up and down the octapolarcatheter to obtain various recording sites. These sites werearbitrarily divided imo four areas according to the positionof the electrodes of the octapolar catheter: area A com-prised either side of the proximal electrode pair (electrodes7 and 8) in the mid-atrium and subsequent bipolar pairswere sequentially labeled: area B = electrodes 5 and 6:area C = electrodes 3 and 4: and area D = the most distalelectrode pair (electrode I and 2) encompassing the ti.s.suejust superior and inferior to the tendon of Todaro.
tentials to the site of the fast pathway, histologicstudies were p>erformed, and AV mxlal conductionwas reassessed after radiofrequency ablation.

50 Journal of Cardiovascular Glectrophysiology Vol. 8. ^o. I. January 1997
Closely Spaced Bipolar and MicroelectrodeRecordings
In 6 other isolated and perfused canine hearts,a 7-French octapolar catheter (i-mm electrodewidth and 2-mm intereleclrode distaiice center toeenter) was visually placed along the anterior lim-bus of the fossa ovalis, with its tip located withinthe apex of the triangle of Koch. According to theposition of the electrode pairs, four aieas were de-fined £iround the oetapolar calhetcr (Fig. IB). AreaA encompassed electrodes 7 and 8 and was usu-ally located along the luiterior linibus adjacent tothe superior half of the fossa ovalis; area B en-compassed electrodes 5 and 6 and was usuallylocated anterior to the inferior half of the fossaovalis; area C encompassed electrodes 3 and 4 andwas usually located just anterior to the region be-tween the fossa ovalis and the tendon of Todaro;and area D encompassed electrodes 1 and 2 andwas usually located over the anterior portion (apex)of the triangle of Koch.
Bipolar recordings were obtained from adjacentelectrode pairs. To correlate these recordings withmore localized activation, a single bipolar electrodeconsisting of two Teflon-coated silver wires (each0.010 in inch diameter; 0.4-mm interelectrode dis-tance) and a single intracelluhir micn:)electr(xle wereplaced adjacent to every electrode and interelectrodespace ofthe octapolar catheter (ai'eas A, B, C. andD). Trans me mbnuie action pt tentials were obtainedwith the standard glass microelectrode filled with 3M KCI and connected to a high-impedance ampli-fier (mtxlel FD 223, World Precision Instruments,Inc., Sarasota, FL, USA). The signals wens displayedon an oscilloscope {model 2214. Tektronix Inc..Beaverton, OR, USA) and recorded simultaneouslywith the bipolar eleetrograms using the Bard sys-tem (see above). Filter settings for microelectroderecordings were set to 0.01 to 2()(K) Hz. For as-sessment ofthe resting potential, amplitude, and up-stroke velocity, the action potentials were alsorecorded on a second calibrated recording system(Gould TA 2000, Gould, Inc., Valley View, OH,USA). The upstroke velocity of the action poten-tials (dV/dt) was detennined elecu^onically with hn-earity up to 6(X) V/sec.
Differentiation of Cell Types
For the AV nodal area, electrophysiologie dif-ferentiation between cell types is conventionallymade on the basis of their transmembrane actionpotential chaiacteristics. For this purpose, mea-surements of activation times, upstroke velocity.
and various other electrophysiologic parameters aswell as the assessment of action potential mor-phology during ba.seline or extrastimulus deliveryhave been commonly used. '""
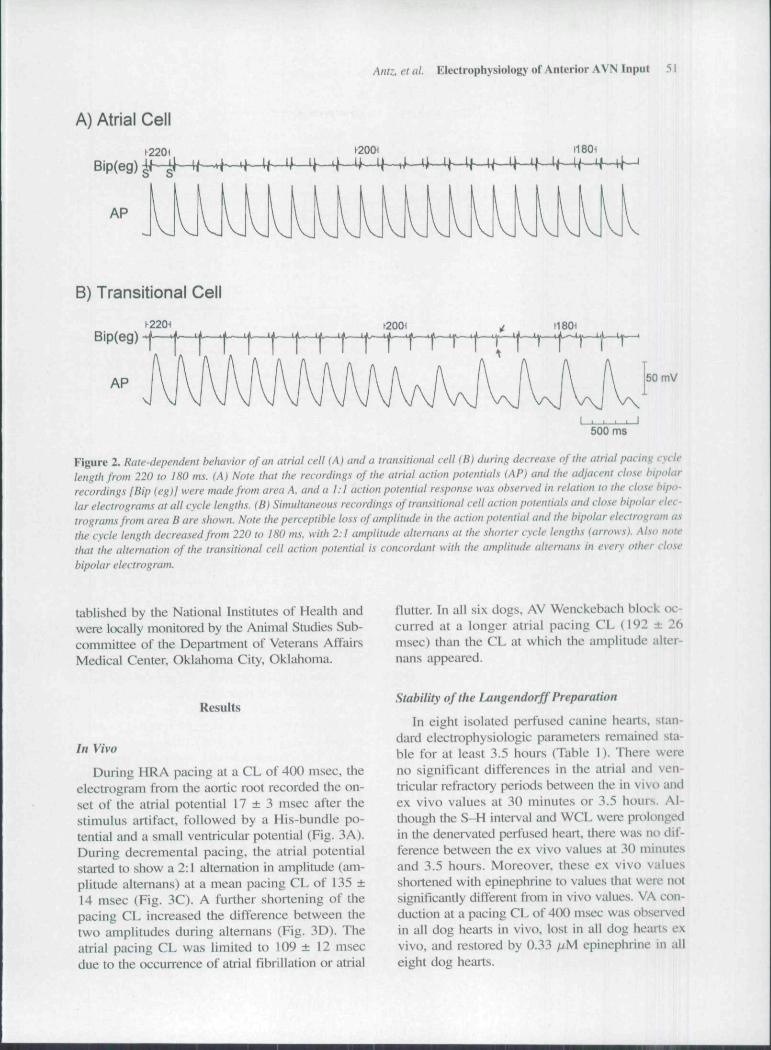
For this study, differentiation between atrial (A)cell and transitional (T) cell action potentials lo-cated within the interatrial septum was made us-ing the aforementioned parameters in addition totheir rate dependent electrophysiologic properties.Atrial ceils (Fig. 2A) were identified by their tri-angular shape, little decrease in amplitude at shortCLs, and their I: I action potential response lo stim-ulation down to the shortest CL (UXJ msec). Tran-sitional ceils (Fig. 2B) were identified by their morerounded peaks, slower upstrokes, decremental re-sponse, and viirious forms of conduction block overa wide range of pacing CLs down to KH) msec.
Histologic Studies
Histologic studies of the anterior limbus of thefossa ovalis were peiiomied in 4 canine hearts.The tissue was cut parallel to the endocardial sur-face at 40-pm intervals at 14 levels. Sections werestained with hematoxylin and eosin and examinedby light microscopy.
Transitional cells were distinguished from atrialcells according to previously published criteria,'-'''*i.e., by their smaller diameters, lesser degree ofeosinophilia, and separation into smaller fasciclesby connective tissue. The compact AV ntxle wasidentified using the criteria of Anderson et al.,'''i.e., as a structure that contained small cells thatfornied a half-oval, were aggregated into tightlyknit fascicles, and were not separated by muchconnective tissue.
Statistical Analysis
Data are expressed as mean ± I SD. In case ofan asymmetrical, non-Gaussian distribution of mea-sured parameters, the median is given instead ofthe mean. Comparisons between continuous vari-ables were analyzed by Student's two-tailed /-testfor paired and unpaired samples. Factorial or re-peated measures analysis of v;mance was used forcomparison between data in the same measure-ments groups (ANOVA), followed by multiplecomparisons (Scheffe F-test). Differences wereconsidered significant at P < 0.05.
Study Guidelines
Studies were pertbrmed according to the guide-lines for humane care and treatment of animals es-
Antz. et al. Electrophysiology of Anterior AVN Input
i180i
B) Transitional Cell
^220^Bip(eg) ^^^~Y~^^-~YTTT^rT^^
AP 50 mV
500 ms
Figure 2. Rate-dependent behavior of an atrial celt lA) and a transitional cell (B) during decrease of the atrial pacing cyclelength from 220 to 180 ms. (A) Note that the recordings of the atrial action potentials {AP) ami the adjacent close bipolarrecordings {Bip {eg)} were made from area A, and a 1:1 action potential response was observed in relation to the close bipo-lar electrograms at all cycle lengths. (Bi Simultaneous recordings of transitional cell action potentials and close bipolar elec-trograms from area B are shown. Note the perceptible loss of amplitude in the action potential and the bipolar electrogram asthe cycle length decreased from 220 to ISO ms, with 2:1 amplitude alternatis at the shorter cycle lengths (arrows). Al.^o notethat the alternation of the transitional celt action potential is concordant with the amplitude altemans in every other closebipolar electrogram.
tablished by the National Institutes of Health andwere locally monitored by the Animal Studies Sub-committee of the Department of Veterans AffairsMedical Center, Oklahoma City. Oklahoma.
flutter. In all six dogs, AV Wenckebacb block oc-curred at a longer atrial pacing CL (192 ± 26msec) than the CL at which the amplitude alter-nans appeiired.
Results
In Vivo
During HRA pacing at a CL of 400 msec, theelectrogram from the aortic root recorded tbe on-set of tbe atrial potential 17 ± 3 msec after tbestimulus artifact, followed by a His-bundle po-tential and a small ventricular potential (Fig. 3A).During decrementa! pacing, the atrial potentialstarted to show a 2:1 alternation in amplitude (am-plitude altemans) at a mean pacing CL of 135 ±14 msec (Fig. 3C). A further shortening of thepacing CL increased the difference between thetwo amplitudes during altemans (Fig. 3D). Theatrial pacing CL was litnited to 109 ± 12 msecdue to the occurrence of atrial fibrillation or atrial
Stability of the Langendorff Preparation
In eight isolated perfused catiine hearts, stan-dard electrophysiologic paraineters remained sta-ble for at least 3.5 hours (Table 1). There wereno significant differences in the attial and ven-tricular refractory pericxis between the in vivo andex vivo values at 30 minutes or 3.5 hours. Al-though the S-H interval and WCL wet^ prol{)ngedin the denervatcd perfused heart, there was no dif-ference between tbe ex vivo values at 30 minutesand 3.5 hours. Moreover, these ex vivo valuesshortened with epitiephrine to values that were notsignificantly different from in vivo values. VA cc«i-duction at a pacing CL of 4(X) msec was observedin all dog hearts in vivo, lost in all dog hearts exvivo, and restored by 0.33 fuM epinephrine in alleight dog hearts.

52 Journal of Cardiovascular Electrophysiology Vol. 8. No. /. Januan-1997
A)
B)
H R A ^ ^ - ^ , ^ i ^HBE
C)II
HRA^ T 140 T T
HBE- ' r—^h^
D)
100 ms
Figure 3. Effect of rapid atrial pacing in vivo. No ampli-tude altemans was seen in the atrial potential recordedfrom the electrode catheter (HBE) located in the aortic rootat cycle lengths of 400 to 150 m.\. Amplitude altemans wasinitially obser\-ed at a cycle length of 140 ms and becamemore marked at a cycle length of 110 ms. There were nocorre.sponding changes in the atrial potential recordedfrom the high right atrium {HRA) over the same range ofcycle lengths. Note that recording.\ from the electrode cath-eter show a very large atrial potential and a very .small Hisbundle and ventricular potential.
Bipolar Potentials and Radiofrequency Ablation ExVivo
When tbe mapping catheter was placed with itstip against tbe anterior interatrial septum just su-perior to the tendon of Todart> in 10 dogs, a largeatrial potential was recorded, whicb was cau.sed bytbe almost simultaneous activation of atrial (A)
TABLE 1Slabilily of the Electrophysiologic Parameters DuringLuiigeiidorff Preparation and Comparison to In Vivo
V.\ \'ivii K\ Vivo V.\ Vivo I'ostIn Vivit ut 30 mill ut ..1.5 li Kpint'phriiic
WCL 188 ± 2(. 266 ± 38* 270 ± 23* 187 ± 77**(msec)
S-Hinlerval 77 ± 9 132 ± 29* 133 + 30* SK) ± 26**(msec)
AERP [|3 ± 23 lOi i 4 107 ± 15(msec)
VERP 143 ± 24 126 ± 14 139 ± 18(nxsec)
VA 8/8 hearts 0/8 tieans* 0/8 hearts* 8/8 hearts**conduction
No siynificanl dilTerence was found between ex vivo al 30min and ex vivo at 3.5 h,*P < 0.05 vs in vivo by ANOVA; **P < 0.05 vs ex vivoat 30 min and ex vivo at 3.5 h hy ANOVA.AERP = airial effeclivc refractory period: S-H interval =atrial stimulus-His bundle interval: VA conduction =ventriculoiuriiti conduction; VERP = ventricular effectiverefractory period: WCL = Wenckbach cycle length.
as well as transitional (T) cells (see t^elow). ThisA/T potential (Fig. 4) was typically followed bya His-bundle potential and a small ventricular ptvtential. During decremental HRA pacing, the AVWCL averaged 262 ± 21 msec. During further re-duction of tbe stimulation CL, the extracelluhtr A/Tpotential showed a 2:1 amplitude alternans be-ginning at a mean CL of 168 ± 15 msec in 9 of10 dogs (Fig. 4B). This response was usually as-sociated with a splitting of the A/T potential intotwo components. At very short CLs. a 2:1 blockbetween the two comp()nents was seen beginningat a mean CL of 152 ± 19 msec in 10 of 10dogs. These findings suggest that the extracellu-lar A/T potential recorded at baseline representsthe superimposed activation of two componentswith different rate-dependent characteristics.
It was hypothesized that tbe specialized transi-tional cells in the imterior approacb to the AV nodewould block at a longer CL tban nottiial uttialtissue, i.e.. the transitional cells would block 2:1at a CL at wbich local atrial activation would stillpropagate in a 1:1 fashion. As a result, unblockedactivity would represent atrial activity, and theblocked component would represent potentials thatare specific for activation of tbe anterior transi-tional zone. To verify the hypothesis that the siteat which this phenomenon occurred was correlatedto the U^ation of the fast pathway, radiofreqtiencycurrent was applied between the two mapping elec-trodes that recorded the 2:1 block in the A/T po-tential in six dogs (compare Figs. 4A and 4D).
In all dogs, ablation caused a significant pro-longation of the S-H interval (from 113 ± 19 to

Antz. et al. Electrophysiology of Anterior AVN Input 53
A
B
D After Ablation/v.
HRA 400
154
CS
Figure 4. Effect of rapid atrial pacing ex vivo. The elec-trode catheter is located in the anterior interatriat septum(AS) superior to the tendon of Todaro {see Fig. I A). Notethe large atrial activity {A/T: caused by the activation ofatrial [A] as welt as transitional [T] cells), small His bun-dle {H). and tow-amplitude ventricular (V) potential. (A) Ata pacing cycle length of 400 ms. no change in the A/T etec-trogram is observed in two consecutive beats. {B) At a .s-tim-ulation cycle length of 160 ms, the A/T potential shows a2:1 amptitude altemans. (C) At a cycte length of 140 m.s.the large part of the A/T potential blocks in a 2:1 fashion.Note that the atrial activity as recorded in the high rightatrium (HRA) and the coronary sinus (CS) electrodesshowed no such rate-dependent changes. (D) After ra-diofrequency delivery to the ablation catheter, the S-H in-terval is markedly prolonged, and the A/T potential exhibitssimilar low-amplitude deflections as those observed during2:1 A/T potential block (C).
151 ± 30 msec: P < 0.01) without affecting theAV WCL (253 ± 15 vs 253 + 27 ms; P = NS).suggestitig damage to ibe fast pathway with pt eser-vation of slow pathway and AV nodal function.Verification of the atriaJ end of tbe fast pathwayby assessment of earliest retrograde atrial activa-tion was not possible, because VA conduction wasnot present before or after radiofrequency ablation
in any of the catecholaniine-depleted, isolated, per-fused canine hearts.
To investigate tbe origin of the extracelluhir A/Tpotential, closely spaced bipolar (extracellular) andmicioelectnxle (intracellular) recordings were ob-tained along the length of the tx-taptiUu- catheter onthe anterior limbus of the fossa ovalis (Fig. IB). Tliedistal pair of the octapolar catheter (electrodes 1-2)recorded a large ventricuUir potential, a His-bundlepotential, and a small atrial potential (Fig. 5). wliilethe electrogi-am frotn the most proximal pair of elec-trodes (7-8) showed only a large attial potential.
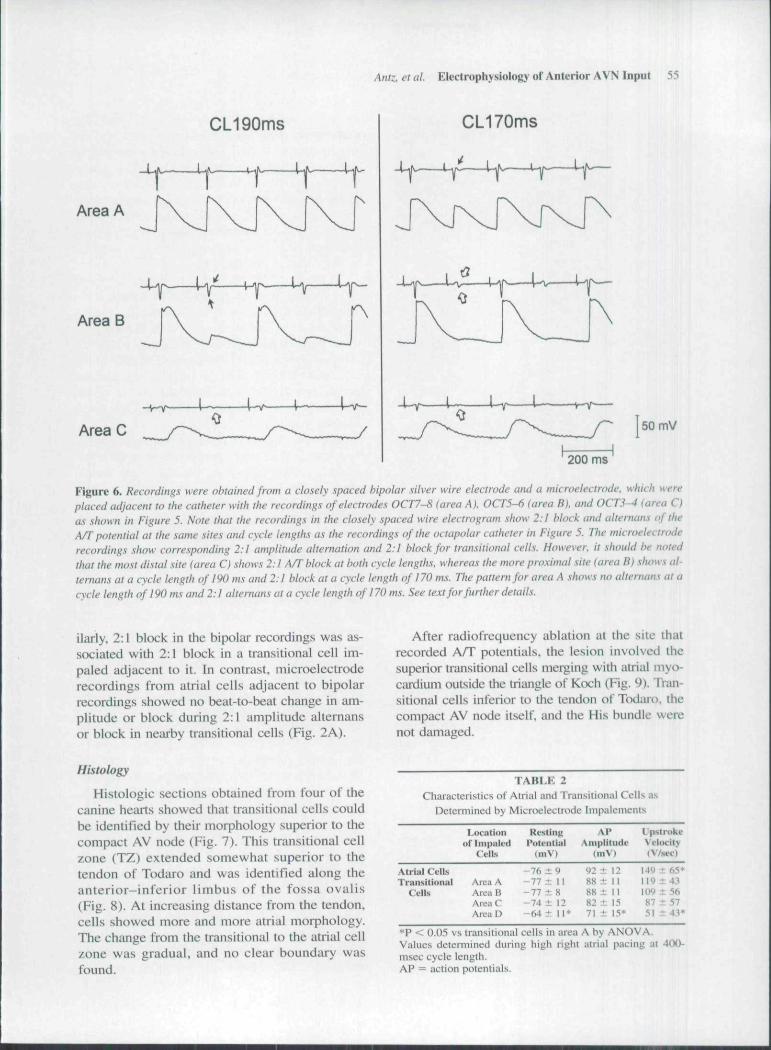
During decremental HRA pacing. 2:1 ampli-tude altemims eventually occuiTed in all biptilai clec-trogratns: however, it occurred at longer CLs atthe distal electrodes kx:ated near the tendon of To-dan> thiin in tbe most proximal electrtxie pair in themid-atrial region of tbe anterior limbus (185 ± 25vs 171 ± 2 0 msec; P < 0.05). Two to one blockalso occunied at a longer CL in tlie electrognun clos-est to the tendon of Todaro and later in tbe moreproximal pairs (161 ± 18 vs 150 ± 18 msec; P <0.05). It should be noted that 2:1 block was ob-served in tbe most proximal clectiogiiim (electrtxles7-8) in only I of 6 dogs. Recordings obtained fmmthe closely spaced bipohir wire electrode showed2:1 amplitude altemans and 2:1 block at the siune.sites and CLs at wbich it was recorded from the ad-jacent catheter electrodes (compare Figs. 5 and 6).
Distribution of Action Potentials Along the AnteriorLimbus of the Fossa Ovalis
Microelectrode recordings were obtained in theanterior interatrial septum adjacent to tbe electrodesof the octapolar catbeter that was placed along theanterior l imbus of the fossa ovalis , i.e.. allrecordings were obtained from cells located su-perior and just inferior to the tendon of Todaro(Fig. IB). In this region, intracellular potentialswere consistent with electrophysiologic criteria re-ported for transit ional ce l l s ' " ' " and atrialcells."^''' Atrial cells were triangular in shape, bada resting potential of -76 ± 9 mV. an action po-tential amplitude of 92 ± 12 mV. and an upstaikevelocity of 149 ± 65 VAsec measured at an atrialpacing CL of 400 msec (Table 2). During decre-mental pacing, atrial cells showed a slight but pi\>gressive reduction of tbeir action potential ampli-tude without change in the level of tbe restingpotential (Fig. 2A). These ceils did not exhibit 2:1amplitude altemans or 2:1 block at CLs > 100msec. In contrast, transitional cell action potentialsexhibited electrophysiologic characteristics that

54 Journal of Cardiovascular Electrophysiology Vol. S. No. /, January 1997
A) CL190ms
B) CL170ms
OCT
^ —
-f-v-- ~f~ .p __
100 ms '
Figure 5. Recordings obtained from an octapolar catheter placed along the anterior limbus of the fossa ovatis with its tipelectrode located within the apex of the triangle of Koch {.see Fig. IB) demonstrate different responses to the same stimulationinten'als along the bipotes of the octapotar catheter at a cycte tength {CD of 190 and 170 ms. (A) At a CL of 190 ms. a 2:1block of the A/lpotentiat was observed in the bipolar recordings from the two distal electrode pairs (electrodes 1-2 and3-4). A 2:1 atnplitude attemans was observed in the bipolar recording from electrodes 5-6. Init there was no change in themost proximal bipolar electrogram from electrodes 7-8. (B) At a 20-ms shorter cycle tengtti. the bipolar A/T etectrogramrecorded in etectrodes 5-(5 now shows clear 2:1 block. Electrograms from electrodes 7-8 now display a 2:1 A/T amplitude al-temans. Observe that the distal bipolar electrode pair displays a large ventricular and His-bundte potentiat and onty a verysmall A/T electrogram. Note that the spread of atrial activation occurs from the proximal to distal bipoles during HRA pac-ing, indicating that the distal electrodes are closer to the A V node.
were more node-like, with reduced upstroke ve-locities and decremental response to atrial pacing(Figs. 2B and 6).
Furthermore, tbere was a gradation of re-sponse of transitional cells depending on their dis-tance from the compact AV node: when the tni-eroelectrtxle impalements within the anterior lim-bus of tbe fossa ovalis approached the AV nodeduring an atrial pacing CL of 4(X) msec, the rest-ing membrane potential became less negative (-77± 11 mV in area A vs -64 ± 11 mV in area D; P< 0.05), the action potential amplitude decreased(88 ± 11 mV in area A vs 71 ± 15 mV in area D;P < 0.05), and a reduction in action potential up-stroke velocity of transitional cells was observed(119 ± 43 V/sec in area A vs 51 ±43 V/sec inarea D; P < 0.05; compare Table 2 and Fig. 6).
Transitional cells consistently showed a specificpattem of response to atrial pacing. At short atrialpacing CLs, transitional cells exhibited a decrease
in amplitude in every other action potential, whilethe amplitude of the preceding beats slightly in-creased (2:1 antplitude altemans). as shown in Fig-ure 2B. When the CL was further reduced, everyother action potential was lost (2:1 action poten-tial block). The progression to complete 2:1 ac-tion potential bkx;k at very short adial pacing CLswas recorded in all transitional cells locatedclose to tbe tendon of Todiiro. but only occasion-ally at sites near tbe proximal poles of the <x;ta-polar catheter.
Correlation Between Microelectrode and BipolarRecordings
Whenever 2:1 amplitude altemans in the cath-eter or in the close bipolar wire electrode wasrecorded, microelectrode recordings from an ad-jacent transitional cell showed 2:1 altemans con-cordant with the bipolar recordings (Fig. 6). Sim-
CL190ms
,4/1?:. et al. Electrophysiology of Anterior AVN Input 55
CL170ms
Area A
Area B
Area C 50 mV
200 ms
Figure 6. Recordings were obtained from a closely spaced bipolar silver wire electrode and a microelectrode, which wereplaced adjacent to the catheter with the recordings of electrodes OCT7-S (area A). OCT5-6 (area B), and 0CT3-^ (area C)as shown in Figure 5. Note that the recordings in the closely spaced wire etectrogram show 2:1 block and alternans of theA/T potential at the same sites and cycle lengths as the recordings of the octapolar catheter in Figure 5. The microelectroderecordinf-s show corresponding 2:1 amplitude altemation and 2:1 block for tramitional cells. However, it should be notedthat the most distal site (area C) shows 2:1 A/T block at both cycle lengths, whereas the more proximal site (area B) shows al-ternans at a cycle length of 190 ms and 2:1 block at a cycle length of 170 ms. The pattern for area A shows tw alternans at acycle length of 190 ms and 2:1 alternans at a cycle length of 170 ms. See text for further details.
ilarly, 2:1 block in the bipolar recordings was as-sociated with 2:1 block in a transitional cell im-paled adjacent to it. In contrast, microelectroderecordings from atrial cells adjacent to bipolarrecordings showed no beat-to-beat change in am-plitude or block during 2:1 amplitude altemansor block in nearby transitional cells (Fig. 2A).
After radiofrequency ablation at the site thatrecorded ATT potentials, the lesion involved thesuperior transitional cells merging with atrial myo-caidium outside the triangle of Koch (Fig. 9). Tran-sitional cells inferior to the tendon of Todaro, thecompact AV node itself, and the His bundle werenot damaged.
Histology
Histologic sections obtained from four of thecanine hearts showed that transitional cells couldbe identified by their morphology superior to thecompact AV node (Fig. 7). This transitional cellzone (TZ) extended somewhat superior to thetendon of Todaro and was identified along theanterior-inferior limbus of the fossa ovalis(Fig. 8). At increasing distance from the tendon,cells showed more and more atrial morphology.The change from the transitional to the atrial cellzone was gradual, and no clear boundary wasfound.
TABLE 2Characteristics of Airial and Transiiional Cells as
Determined by Microelectrode Impalements
Atrial CellsTranNitiunal
Cells
l (K'ati»nof Impaled
Cells
Area AAreaBAreaCArea D
RestingPotential
(mV)
-76 ± 9-77 ± 11-77 ± 8-74 ± 12-64 ± 1 1"
Al>Amplitude
|m\')
92 ± 1288± II88+1182 ± !571 t 15*
UpslntkeVi-locity(V/stC)
149 i 65"119 + 43\m ± 5687 ±5751 ± 43*
*P < O.O.'i vs transitional cells in area A by ANOVA.Values determined during high right atrial pacing at 4(X)-msec cycle length.AP = action potentials.

56 Journal of Cardiovascular Electrophysiology Vol. 8, No. 1. Jamiury 1997
^4%!:iffc^:l
v - • , < • •
Figure 7. haiL\itioiiai cfil.\ {IZ.) Lippiouiliiiiii ilu-AV iiaih-
(AVN) from .superior at the level ofthe [wnetfatiii}' himdk'(PB). Note the wave-like .shape of the trausitioiial cellsclo.se to the AVN. CFB = central fibrous body: V = ventrk-ular mvocarditim.
Discussion
Previous reports have described characteris-tics of transitional cells and nodal cells focusingon the AV node itseli" or the posterior AV nodalinput.'"" In his meticulous microelectrode studyof AV nodal activation in the rabbit, Billette" alsodescribed transitional cells, located just superiorand inferior to the tendon of Todaro, with differ-ent properties. He suggested that different typesof transitional cells were responsible for inho-mogeneous activation, but did not establish a spe-cific activation pattern in the anterior atrial sep-tum. For the posterior approach to the AV node,McGuire et al.' showed that cells near the tricus-pid annulus have nodal-type action potentials,whereas cells fiirther away from the annulus ex-hibit action potentials that are more "atrial" intype. Extracellular recordings showed doublepotentials simiiar to those described by Haissa-guerre et al.- (high-frequency followed by low-
frequency detlections). However, in that study, theresponse to decreasing atriai pacing CLs was notsystematically studied, particularly for the variouscell types other than tbe node-like action poten-tials recorded close to the tricuspid annulus. Thelatter showed Wenckebach responses with de-creasing CLs, similar to AV nodal cells, but his-tologically these subendocardial layers "were notarranged in an interweaving fashion as is foundin the compact AV node."'
In the study by McGuire et al..' cell impale-ments just inferior to the tendon of Todaro at thelevel of the CS ostium showed atriai cells. The re-sults ot the present study show a sitnilar arrange-ment superior to the tendon of Todaro within theanterior limbus of the fossa ovalis. The cells in thisarea were identified histologically and electro-physiological ly as transitional cells. As we movedsuperiorly from the tendon, more atrial cells wereencountered.
The most important result of the present study,however, is that, from this area, bipolar cathetersrecord specific potentials that show rate-depen-dent 2:1 amplitude altemans in vivo and ex vivoas well as 2:1 A/T block in the isolated periusedheart. The progressive shift of 2:1 amplitude al-temans and 2:1 block toward longer CLs as theAV node is approached from the inleratrial sep-tum is probably a manifestation of a progressivedecrease in intercellular coupling and, thereby, re-duction of source cun^ent. The presented data sug-gest tbat the 2:1 block of the A/T potential asrecorded in the bipolar electrograms at shortpacing CLs reflects the 1:1 conduction pattem inatrial cells and the 2:1 blocking pattem in transi-tional cells. However, the precise site at whichthis block occurs was not determined, because in-tercellular resistance and the transfer of currentacross specialized connections between adjacentcells were not determined. Future studies involv-ing current injection techniques in one cell anddetermination of electrical responses in adjacentcells may provide additional information on cell-to-cell interactions.
It should be mentioned that the bipolar elec-trograms at the proximal and distal electrodes ofthe octapolai' catheter showed distinct vaiiations inamplitude during 2:1 A/T altemans and bUx'k. Inthe proximal region (mid-atrial septum). Ihcbipolar A/T potential was usually large and had asecond component with a smaller amplitude andhigher frequency, wbich delayed and showed 2:1altemans in response to decreasing CLs, leavingthe amplitude of the larger potential relatively

Ant:., etal. Electrophysiology of Anterior AVN Input 57
Figure 8. In the inferior margin ofthe anterior limbus of the fossa ovalis (FO). transitional cells (TZ) are identified hy theirseparation into small fascicles and their wave-like shape. Transitional cells can be clearly followed superior to the loweredge of the FO. In the superior part ofthe anterior limbus ofthe FO. cells become atrial in morphology. Note that separationof cells is also due to interstitial edema, caused by the Langendorff preparation.
unaffected (Fig. 6. area A). However, in the dis-tal region (close to the tendon of Todaro), the ma-jor component of the A/T potential showed 2:1amplitude altemation and block in response to de-creasing CLs. These differences may reflect therelative distribution of the amount of atrial andtransitional tissue comprising these areas. As wasshown from the histologic studies, the distal re-gions are composed of mainly transitional cellsand few atrial cells, whereas the reverse was foundin the proximal (more superior) portion of the fastpathway input zone.
Experimental Preparation
The observation of 2:1 amplitude altemans invivo suggests that the findings of the present studywere not an artifact of the ex vivo preparation. In-deed, we have shown that electrophysioiogic pa-rameters remain stable in these Langendoi-ft" prepa-rations over at least 3.5 hours and that atrial andventricular refractory periods were the same as invivo. Another important finding was that AV nodalconduction was reduced in the denervated isolatedperfused heart, but retumed to in vivo values af-ter epinephrine administration, which provided ev-idence that the AV node in the isolated and per-fused canine hearts was not damaged, but withoutadrenergic drive from sympathetic nerves as wellas circulating catecholajnines.
Perfusion of the isolated heart does not have thelimitation of superiused preparations in which onlythe superiicial cell layers survive, and there maybe possible electrotonic interaction between the su-perficial layers and deeper hypoxic cells.-*' Ac-cording to available published data,"'' the use ofDAM to dampen contraction to allow stable mi-croelectrode impalements does not affect actionpotential morphology and electrophysioiogie prop-erties.
Basic and Clinical Implications
The seminal microelectrode studies of Mendezand Moe-' demonstrated the tKXurrencc of reentryin and around the AV node. These and other stud-ies from Moe et al.-- established tbe concept of adual system consisting of fast and slow AV nocialpathways whose conduction properties could bedissociated sufficiently to facilitate echo beats andAVNRT. With the advent of clinical electrophysi-ology. the evidence for dual AV nodal patJiwayswas demonstrated in man-' as well as the key roleof the AV node in AVNRT--*
Early on. a polemic iirose as to whether AVNRT,with its dichotomized fast and slow conductionpathways, was confined within the AV node or ad-jacent atrial tissue composed an upper commonpathway to complete the circuit. Experimetital stud-ies in the dog heart in vivo and in the isolated su-

58 Journal of Cardiovascular Klectroph.vsiology Vot. 8. No. I, Januan,-1997
• r/
Fijjure 9. Histotogy obtained from a dog fottowing ra-diofrcijuency ablation at the site at which A/T potentialswere recorded, with induction of an S-H interval prolonga-tion with no change in intraatrial conduction time {S—A in-teiTat). The histologic section shows a tine of demarcationbetween damaged atrial cells (note contraction handsabove the arrows) and the transitional zone (TZ). The latteris composed of some larger atrial cells {center) graduattyfusing with smatter transirionat cells. As.sociated intercettu-tar materiat is seen in the tower right. Ttw lesion does notinvolve transitional celts ctose to the A V node (bottom).
perfused rabbit AV junctional prepiiration providedevidence in favor of an intra-AV nodal reentry cir-cuit.-**" ' Other reports using the i.solaled rabbit heartalso provided convincing evidence for the partic-ipation of tissues adjacent to the AV node in in-duced AVNRT.^'-''
Clinically, the major proponents of the conceptof intra-AV nodal reentry accumulated, in man, anumber of electrophysiologic demonstrations thatwere interpreted to exclude the participation ofpathways outside (he AV node in AVNRT^' How-ever, support for this view began to erode wbenseveral surgical teams were able to cure AVNRTby placing multiple lesions around the AV nodewithout inducing loss of AV conduction." " Wilhtbe introduction of radiofrequency catheter abla-
tion, discrete lesions placed in tbe posterior AVjunction (slow pathway) at a distance from thecompact AV ntxie pennanently prevented tbe oc-currence of AVNRT without altering AV nodalconduction.'-
More recent studies have suggested tbat transi-tional, rather than atrial. tissues arc the major con-stituents of the slow pathway and that the transi-tional zone cells share similar properties as thenodal cells of the compact AV node.' Specifically,both tnuisitional and ntxlal cells show Wenckebachblock in response to decreasing CL. In light of thesurgical and catbeter ablation studies, several in-vestigators proposed an anatomic expansion of theAV nixle. which included the transitional zone tis-sues, thus maintaining the intra-AV nodal con-cept of AVNRT-^ '• •'• •"
The present study confirms and extends thefindings of previous reports from our labora-tory-"" that transitional tissue in the anterior AVjunction constitutes another major input to theAV node. Moteover. these transitional cells donot demonstrate Wenckebach conduction blockcommon to transitional cells of the slow path-way'"' and those of the compact AV node .""The anterior ttansitional cells exhibit 2:1 block(in relation to atrial cells) at CLs distinctly shorterthan lhe average AV WCL (see Re.still.s). Tbepresent data further suggest ihat the transi-tional cells composing the anterior approach tothe AV node are anatomically, as well as elec-trophysiologically, distinct frotn cells compos-ing the compact AV node and electrophysio-logicatly distinct from the transitional cells ofthe slow pathway.'^
The present finding that ablation of these ante-rior transitional ti.ssues significanlly prolongs lheS-H interval, i.e., AV nodal conduction time with-out altering the CL at which AV nodal Wencke-bacb block occurs, provides strong presumptiveevidence thai the anterior AV nodal input consti-tutes all or a major part of the so-called fast AVntKlal input. These findings iire con.sonant with tberecent posiulation by Sung et al.'^ and the earlierdemonstration of specific anterior and posterioratrial exit sites in patients witb AVNRT during ven-tricular pacing. '*
Since AVNRT was not induced in our experi-ments, the participation of these tissues in AVNRTcannot be verified. The abolition of AVNRT in pa-tients by ablation in this region eliminating fastpathway conduction suggests that these same tis-sues participate. Similarly, elimination ofAVNRT by ablation of the tissues posterior and

Antz, et al. Electrophysiolog)' of Anterior AVN Input 59
inferior in the septum joining the slow pathwayindicates that those tissues also participate. Thelink between the two inputs remains to be identi-fied.
Study Limitations
Vagal stimulation was used in tbis study in vivofor better identification of atrial potentials atshort CLs. However, vagal stimulation not onlyproduces high-degree AV nodal block, but alsochanges tbe atrial action potential (acceleration ofrepolarization and frequently hyperpolarization'^).It is unlikely that the observed 2:1 amplitude al-ternations as recorded from the His-bundle cathe-ter in our study was produced by \agal stimula-tion, because this pattem was still obsei ved aftertermination of stimulation and was never recordedfrom tbe electrodes in the HRA or the crux ofthe bean. Moreover, the recording of 2:1 ampli-tude alternans in the atrial potential was charac-teristic for the anterior approach to the AV nodeand was also observed ex vivo in the absence ofvagal stimuli.
Microelectrode recordings were obtained fromcells in the anterior interatrial septum. These cellswere arbitrarily divided into four different areasaccording to the position of the octapolar catheter.Although great care was taken to place the cathe-ter in identical anatomic positions in all experi-ments, some variation between experiments can-not be excluded.
Tissue swelling in tbe Langendorff prepara-tion made the histologic evaluation of myocar-dial architecture more difficult than in unma-nipulated hearts. There was an increase of in-terstitial space and a tendency of fascicles to besomewhat wavy, even in areas appearing remotefrom the identifiable transitional zone. The co-agulation of cells in tbe fresh ablation lesion wasless prominent and recognizable by virtue of con-traction bands lacking the hemorrbagic marginsseen in the in vivo prtwiuced lesions. Althoughwe could determine that the transitional cell zoneextended somewhat superior to the tendon of To-daro and tbat the radiofrequency lesion involvedsuperior transitional eells merging with atrialmyocardium without damaging structures infe-rior to the tendon of Todaro. the histology fromthe isolated and perfused canine hearts did notallow precise measurements of the lesion cir-cumference or of the superior extent of the tran-sitional cell zone within the anterior limbus ofthe fossa ovalis.
A definite correlation between the blocking pat-tern of transitional cells and fast pathway con-duction could not be established directly for thefollowing two reasons: (1) It was not possible todetermine the refractory periods of tbe fast path-way in the catecholamine-depleted isolated andperfused canine heart, because, ex vivo, none ofthe dogs showed dual AV nodal physiology (asdefined by AV nodal echo beats or abrupt changesin the A-H interval during atrial stimulationwith extrastimuli or decreasing CL) or VA con-duction: and (2) AV Wenckebach occurred atlonger CLs than the 2:1 amplitude alternans or2:1 blocking pattems. As a result, it was only pos-sible to coiTelate the bipolar potentials recordedin the anterior interatrial septum to the fast path-way in an indirect way (by fast pathway ablationwith consequent disappearance of these potentials,induction of S—H interval prolongation, and nochange in AV Wenckebach). A limitation is thatonly the S-H interval (instead of the A-H inter-val) could be determined in most dogs, becausethe onset of the atrial ptitential could not be clearlyidentified in the His-bundle recording obtainedfrom a catheter below the tricuspid valve. How-ever, in those dogs in which a clear atrial pt^en-tial was consistently recorded, the S-A intervalafter ablation was unchanged compared lo tbe pre-ablation value. As a result, S-H interval prolon-gation due to an intraatrial conduction defect couldbe excluded in these dogs, indicating successfulfast pathway ablation and subsequent prolonga-tion of AV nodal conduction time. i.e.. A-H in-terval. These findings were al.so confimied histo-logically since the compact AV node was not dam-aged by the radiofrequency current.
Conclusion
The transitional cell zone in the anterior inter-atrial septum exhibits specific rate-dependent elec-trophysiologic changes in bipolar electrograms andmicroelectrode recordings. Electrophysiologicchanges induced by radiofrequency ahlation ofthese potentials provide strong presumptive evi-dence that this area constitutes all or a major partof the fast AV nodal pathway.
Acknowledf-ment: The authors ihank Jonathan Bussey for his tech-nical assistance.
References
1. Jackman WM. Beckman KJ. McClelland JH, et al: Treat-ment of supraventricular tachycardia due Io atrioventric-

60 Journal of Cardiovascular Electrophysiology Vol. 8, No. 1, January 1997
ular nodal reentry by radiofrequency ablation of slow-pathway conduction. N EngI J Med 1992:327: 313-318.
2. Haissaguerre M. Gaita F. Fischer B. et al: Elimina-tion of atrioventricular nodal reentrant tachycardiausing discrete slow potentials to guide application ofradiofrequency energy. Circulation 1992:85:2162-2175.
3. McGuire MA, de Bakker JMT, Vermeulen JT, et al:Origin and significance of double potentials near theatrioventricular node. Correlation of extracellular po-tentials, intracellular potentials, and histology. Circula-tion 1994:89:2351-2360.
4. Scherlag BJ, Patterson E, Nakagawa H, et al: Changingconcepts of A-V nodal conduction: Basic and clinicalcorrelates. Primary Cardiol 1995:21:13-24.
5. Scherlag BJ. Abelleira JL. Samet P: Electrode catheterrecordings from the His bundle and left bundle in theintact dog. In Kao FF, Koizumi K, Vassalle M, eds:Re.search in Phy.siotogy. Aulo Gaggi, Bologna, 1971,pp. 223-238.
6. Kottkamp H, Hindricks G, Wiilems S, et al: Ananatomically and electrogram-guided stepwise ap-proach for effective and safe catheter ablation of thefast pathway for elimination of atrioventricular nodereentrant tachycardia. J Am Coll Cardiol 1995;25:974-981.
7. Kates RE, Yee YG. Hill I: Effect of albumin on theelectrophysiological stability of isolated perfusedrabbit hearts. J Cardiovasc Pharmacol 1989:13:168-172.
8. Li T. Sperelakis N. Teneick RE. et al: Effect.s of di-acetyl monoxime on cardiac excitation-contractioncoupling. J Pharmacol Exp Ther 1985:232:688-695.
9. Mazgalev T. Cbeng YN. Efimov 1. et al: Effects of 2,3-butanedione monoxime on the AV conduction in theisolated heart. (Abstract) Circulation l995:92(SupplI):I-301.
10. Rubio R, Ceballos G. Suarez J: Coronary flow stimu-lates auric ular-ventricular transmission in the isolatedperfused guinea pig heart. Am J Physiol 1995;269(Heart Circ Physiol 38):HI 177-Hl 185.
11. Billelte J: Atrioventricular nodal activation during peri-odic premature stimulation of the atrium. Am J Physiol1987:252(1 Pt 2):H163-H177.
12. Paes de Carvalho A. de Almeida DF: Spread of activitythrough the atrioventricular node. Circ Res I96O;8:801-809.
13. Anderson RH, Janse MJ, van Capelle FJ, et al: A com-bined morphological and electrophysiological study ofthe atrioventricular node of the rabbit heart. Circ Res1974;35:9()9-922.
14. Anderson RH. Becker AE. Brechenmacher C, et al:The human atrioventricular junctional area. A morpho-logical study of the A-V node and bundle. Eur J Car-diol 1975:3:11-25.
15. Meijier FL. Jan.se MJ: Morphology and electrophysiol-ogy of the mammalian atrioventricular node. PhysiolRev 1988:68:608-647.
16. Scherlag BJ, Patterson E. Lazzara R: Comparative
electrophysiology of anterior and posterior transitionalcells adjacent to the compact atrioventricular node.(Abstract). PACE 1995:18:879.
17. Hoffman BF. Cranefield PF: Electrophysioloj^y of theHeart. McGraw-Hill Book Company, Inc., New York,1960, pp. 42-74.
18. Wagner ML, Lazzara R. Weiss RM. et al: Specializedconducting fibers in the interatrial band. Circ Res1966:18:502-518.
19. Woods WT, Sherf L, James TN: Structure and functionof specific regions of the canine atrioventricular nude.Am J Physiol I982:243:H41-H5O.
20. Tranum-Jensen J, Janse MJ: Fine structural identifica-tion of individual cells subjected to microelectroderecording in pertused cardiac preparations. J Mol CellCardiol 1982:14:233-247.
21. Mendez C, Moe GK: Demonstration of a dual AV-noda! conduction system in the isolated rabbit heart.Circ Res 1955:19:.378-393.
22. Moe GK. Preston JB, Burlington H: Physiologic evi-dence for a dual AV transmission .system. Circ Res1956;4:357-375.
23. Rosen KM. Mehta A, Miller RA: Demonstration ofdual atrioventricular nodal pathways in man. Am JCardiol 1974:33:291-294.
24. Goldreyer BN. Bigger JT Jr: Site of reentry in paroxys-mal supraventricular tachycardia in man. Circulation1971:43:15-26.
25. Wallace AG. Mignone RJ: Physiologic evidence con-cerning the reentry hypothesis for ectopic beats. AmHeart J 1966:72:60-70.
26. Janse MJ, van Capelle FJ, Freud GE, et al: Circusmovement within the AV node as a basis for supraven-tricular tachycardia as shown by multiple microelec-trode recording in the isolated rabbit heart. Circ Resi97l;28:403-4l4.
27. Wit AL, Goldreyer BN, Damato AN: An in vitromodel of paroxysmal supraventricular tachycardia. Cir-culation 1971:43:862-875.
28. Mazgalev T. Dreifus LS, Bianchi J, et al: The mecha-nism of AV junctional reentry: role of the atrionodaljunction. Anai Rec 198 l;201:179-188.
29. linuma H. Dreifus LS, Mazgalev T, et al: Role of theperinodal region in atrioventricular nodal reentry: evi-dence in an isolated rabbit heati preparation. J Am CollCardiol 1983:2:465-473.
30. Josephson ME. Miller JM: Atrioventricular nodal reen-try: Evidence supporting an intranodal location. PACEl993:l6(Ptll):599-6l4.
31. Ross DL, Johnson DC, Denniss AR. et al: Curativesurgery for atrioventricular junction ("AV nodal") reen-trant tachycardia. J Am Coll Carxiiol 1983;6:1383-1392.
32. Guiraudon GM, Klein GJ, Sharma AD, et ai: Skele-tonization of the atrioventricular node for AV nodereentrant tachycardia: Experience with 32 patients.Ann Thorac Surg 1990:49:565-572.
33. Cox JL, Holman WL. Cain ME: Cryosurgical treat-ment of atrioventricular node reentrant tachycardia.Circulation I987;76:I329-I336.

Atitz. et al. Electrophysiology of Anterior AVN Input 61
34. Josephson ME. Miller JM: Atrioventricular node reen-try tachycardias, is the atrium a necessary link? InTouboul P. Waldo AL. eds: Atria! Arrhythmia: Cur-rent Concepts and Management. Mosby Inc., St. Louis,MO. ! 990, pp. 331-3.^9.
35. Spach MS. Josephson ME: Initiating reentry: The roleof nonuniform anisotropy in small circuits. J Cardio-vasc Electrophysiol 1994;5:182-209.
36. Hirao K. Scherlag BJ, Poty H, et al: Teehnique for se-lective ablatinn of the atrial extensions of the fast andslow atrio-ventricular nodal pathways using an epicar-dial approach. (Abstraet) Circulation l993;88:I-492.
37. Paes de Carvalho A: Cellular electrophysiology of theairial specialized tissues. In Paes de Carvalho A, deMellii WC. Hoffman BE. eds: The Specialized Tissuesof the Heart. Elsevier. Amsterdam. 1961. pp. I \f)-133.
38. Sung RJ. Lauer MR. Chun H: Alrioventricuiar nodereentry: Cunenl concepts and new perceptives. PACE1994;I7:14I3-I43O.
39. Sung RJ. Waxman HL, Saksena S. et al: Sequence ofretrograde atrial activation in patients with dual alrio-ventricular nodal pathways. Circulation 1981:64:1059-1067,





















