
Effects of ultraviolet radiation on periphyton in an alpine lake
6
Limnol. Oceanogr., 41(5), 1996, 1035-1040 0 1996, by the American Society of Limnology and Oceanography, Inc. Effects of ultraviolet radiation on periphyton in an alpine lake Rolf D. Vinebrooke and Peter R. Leavitt Department of Biology, University of Regina, Regina, Saskatchewan S4S OA2 Abstract Many alpine lakes have low concentrations of ultraviolet (UV) radiation-absorbing dissolved organic matter, yet receive higher UV radiation flux than low-elevation lakes. We tested whether ambient UV radiation affected periphyton development in a small alpine lake in Banff National Park, Canada. After 30 d, total periphytic biomass and chlorophyll accrual on artificial substrates were enhanced - 100% (t-test, P < 0.05) by removal of UV radiation (~400 nm). The inhibitory effect of UV radiation was species-specific, significantly suppressing (Bonferroni-adjusted t-test, P < 0.05) Achnanthes minutissima Kiitzing but not other colonists. Although taxa apparently differed in their sensitivity to UV radiation, periphyton commu- nities remained dominated by early successional taxa, especially A. minutissima (75% of total biovolume). In contrast, natural epilithic communities were dominated by cyanobacteria (Anabaena subcylindrica Borge, Calothrix sp.). These findings suggest that ambient UV radiation at alpine sites can suppress periphyton development by inhibiting littoral diatom production during the short ice-free season (July-September). Future climatic change may affect the ultraviolet (UV) radiation environment in aquatic ecosystems. For ex- ample, a warmer and drier climate in central North Amer- ica has reduced inputs and increased in-lake removal of dissolved organic matter (DOM), the primary attenuator of UV radiation in lakes (Schindler et al. 1990, 1992; Jones 1992). Low concentrations of DOM allow UV ra- diation to penetrate to greater depths (Scully and Lean 1994; Kirk 1994) and may combine with absolute in- creasesin UV-B (280-320 nm) radiation flux (Blumthaler and Ambach 1990) to adversely affect aquatic organisms (e.g. Williamson et al. 1994). Alpine lakes may offer insights into the long-term ef- fects of elevated UV radiation on aquatic organisms. UV-B and total irradiance increase by -20% and - 10% per 1,000 m of elevation (Caldwell et al. 1980; Blumthaler et al. 1992). In addition, absolute UV radiation flux in alpine regions has increased significantly over the last 10 yr (Blumthaler and Ambach 1990). Furthermore, many alpine lakes contain < 2 mg DOM liter - l and allow UV-B radiation to penetrate to a depth of several meters (Scully and Lean 1994). Transparent water, combined with high ambient UV irradiance, may maximize UV radiation dose to alpine organisms (e.g. Vincent et al. 1984). Shallow-water benthic communities in alpine lakes may be particularly sensitive to UV radiation. Periphyton on rocks (epilithon) includes immobile species that cannot seek low UV refuges, unlike sediment-dwelling periph- yton (Happey-Wood 1988; Vincent et al. 1993) or alpine phytoflagellates (Rodhe et al. 1966; Rott 1988), both of Acknowledgments This work was supported by a Natural Sciences and Engi- neering Research Council of Canada postgraduate scholarship to R.D.V. and a research grant to P.R.L. We thank David Schindler for the opportunity to conduct this research at his semipermanent camp in Banff National Park, Brian Parker for limnological data analysis, and David Lean for UV irradiance and attenuation data. Patti Sim and Craig Wilson provided valuable field assistance. which undergo vertical migration. As well, extensive scouring by ice (2 m thick), cold water (< 12OC), and short ice-free seasons (-90 d at 5O”N) limit the rate of periph- yton development, leading to prolonged succession and dominance by diatoms (Niederhauser and Schanz 1993; but seeStanford and Prescott 1988). Certain diatoms may be especially susceptible to UV radiation because of their small size (Garcia-Pichel 1994) and low concentrations of UV-absorbing pigments (Karentz et al. 199 1 b). Inhibition of algal photosynthesis by UV radiation has been extensively documented in laboratory studies (see Hader 1993; Vincent and Roy 1993). Potentially, UV radiation can reduce phytoplankton production by direct inhibition of photosystems 1 and 2 (Jokiel and York 1984; Neale et al. 1993), formation of nucleic acid lesions (Ka- rentz et al. 199 1 a), or production of lipid-oxidizing com- pounds such as peroxides and free oxygen radicals (Coo- per et al. 1989). However, it is unclear whether short- term physiological effects of UV radiation result in sig- nificant depression of in situ production, due to the pos- sibility of compensatory processes (Neale et al. 1993). For example, Smith et al. (1992) showed that elevated UV radiation associated with Antarctic ozone depletion reduced primary production by only 12%. Experiments that have demonstrated in situ suppression of algal growth by UV radiation have either used artificially enhanced UV irradiance (Worrest et al. 1978) or shallow systems (< 1 cm) that lack significant natural attenuation of UV radiation (Bothwell et al. 1993, 1994). We report on the influence of natural UV radiation on periphyton colonization in an alpine lake in Banff Na- tional Park, Canada (5 1”36’N, l15°50’W). Snowflake Lake is a remote ultraoligotrophic headwater lake situated 200 m above treeline at an elevation of 2,320 m as1(Table 1). The lake contains little terrestrial DOM, and the 1% attenuation depth for UV-B radiation is 2 m (Scully and Lean 1994). Pelagic food-web composition has been de- scribed by Paul and Schindler (1994). Gammarus Zacus- tris dominates the littoral zoobenthos, with densities of 1 O-50 individuals m-2 (Anderson and Donald 1978; 1035
Transcript of Effects of ultraviolet radiation on periphyton in an alpine lake

Limnol. Oceanogr., 41(5), 1996, 1035-1040 0 1996, by the American Society of Limnology and Oceanography, Inc.
Effects of ultraviolet radiation on periphyton in an alpine lake
Rolf D. Vinebrooke and Peter R. Leavitt Department of Biology, University of Regina, Regina, Saskatchewan S4S OA2
Abstract Many alpine lakes have low concentrations of ultraviolet (UV) radiation-absorbing dissolved organic
matter, yet receive higher UV radiation flux than low-elevation lakes. We tested whether ambient UV radiation affected periphyton development in a small alpine lake in Banff National Park, Canada. After 30 d, total periphytic biomass and chlorophyll accrual on artificial substrates were enhanced - 100% (t-test, P < 0.05) by removal of UV radiation (~400 nm). The inhibitory effect of UV radiation was species-specific, significantly suppressing (Bonferroni-adjusted t-test, P < 0.05) Achnanthes minutissima Kiitzing but not other colonists. Although taxa apparently differed in their sensitivity to UV radiation, periphyton commu- nities remained dominated by early successional taxa, especially A. minutissima (75% of total biovolume). In contrast, natural epilithic communities were dominated by cyanobacteria (Anabaena subcylindrica Borge, Calothrix sp.). These findings suggest that ambient UV radiation at alpine sites can suppress periphyton development by inhibiting littoral diatom production during the short ice-free season (July-September).
Future climatic change may affect the ultraviolet (UV) radiation environment in aquatic ecosystems. For ex- ample, a warmer and drier climate in central North Amer- ica has reduced inputs and increased in-lake removal of dissolved organic matter (DOM), the primary attenuator of UV radiation in lakes (Schindler et al. 1990, 1992; Jones 1992). Low concentrations of DOM allow UV ra- diation to penetrate to greater depths (Scully and Lean 1994; Kirk 1994) and may combine with absolute in- creases in UV-B (280-320 nm) radiation flux (Blumthaler and Ambach 1990) to adversely affect aquatic organisms (e.g. Williamson et al. 1994).
Alpine lakes may offer insights into the long-term ef- fects of elevated UV radiation on aquatic organisms. UV-B and total irradiance increase by -20% and - 10% per 1,000 m of elevation (Caldwell et al. 1980; Blumthaler et al. 1992). In addition, absolute UV radiation flux in alpine regions has increased significantly over the last 10 yr (Blumthaler and Ambach 1990). Furthermore, many alpine lakes contain < 2 mg DOM liter - l and allow UV-B radiation to penetrate to a depth of several meters (Scully and Lean 1994). Transparent water, combined with high ambient UV irradiance, may maximize UV radiation dose to alpine organisms (e.g. Vincent et al. 1984).
Shallow-water benthic communities in alpine lakes may be particularly sensitive to UV radiation. Periphyton on rocks (epilithon) includes immobile species that cannot seek low UV refuges, unlike sediment-dwelling periph- yton (Happey-Wood 1988; Vincent et al. 1993) or alpine phytoflagellates (Rodhe et al. 1966; Rott 1988), both of
Acknowledgments This work was supported by a Natural Sciences and Engi-
neering Research Council of Canada postgraduate scholarship to R.D.V. and a research grant to P.R.L.
We thank David Schindler for the opportunity to conduct this research at his semipermanent camp in Banff National Park, Brian Parker for limnological data analysis, and David Lean for UV irradiance and attenuation data. Patti Sim and Craig Wilson provided valuable field assistance.
which undergo vertical migration. As well, extensive scouring by ice (2 m thick), cold water (< 12OC), and short ice-free seasons (-90 d at 5O”N) limit the rate of periph- yton development, leading to prolonged succession and dominance by diatoms (Niederhauser and Schanz 1993; but see Stanford and Prescott 1988). Certain diatoms may be especially susceptible to UV radiation because of their small size (Garcia-Pichel 1994) and low concentrations of UV-absorbing pigments (Karentz et al. 199 1 b).
Inhibition of algal photosynthesis by UV radiation has been extensively documented in laboratory studies (see Hader 1993; Vincent and Roy 1993). Potentially, UV radiation can reduce phytoplankton production by direct inhibition of photosystems 1 and 2 (Jokiel and York 1984; Neale et al. 1993), formation of nucleic acid lesions (Ka- rentz et al. 199 1 a), or production of lipid-oxidizing com- pounds such as peroxides and free oxygen radicals (Coo- per et al. 1989). However, it is unclear whether short- term physiological effects of UV radiation result in sig- nificant depression of in situ production, due to the pos- sibility of compensatory processes (Neale et al. 1993). For example, Smith et al. (1992) showed that elevated UV radiation associated with Antarctic ozone depletion reduced primary production by only 12%. Experiments that have demonstrated in situ suppression of algal growth by UV radiation have either used artificially enhanced UV irradiance (Worrest et al. 1978) or shallow systems (< 1 cm) that lack significant natural attenuation of UV radiation (Bothwell et al. 1993, 1994).
We report on the influence of natural UV radiation on periphyton colonization in an alpine lake in Banff Na- tional Park, Canada (5 1”36’N, l15°50’W). Snowflake Lake is a remote ultraoligotrophic headwater lake situated 200 m above treeline at an elevation of 2,320 m as1 (Table 1). The lake contains little terrestrial DOM, and the 1% attenuation depth for UV-B radiation is 2 m (Scully and Lean 1994). Pelagic food-web composition has been de- scribed by Paul and Schindler (1994). Gammarus Zacus- tris dominates the littoral zoobenthos, with densities of 1 O-50 individuals m-2 (Anderson and Donald 1978;
1035
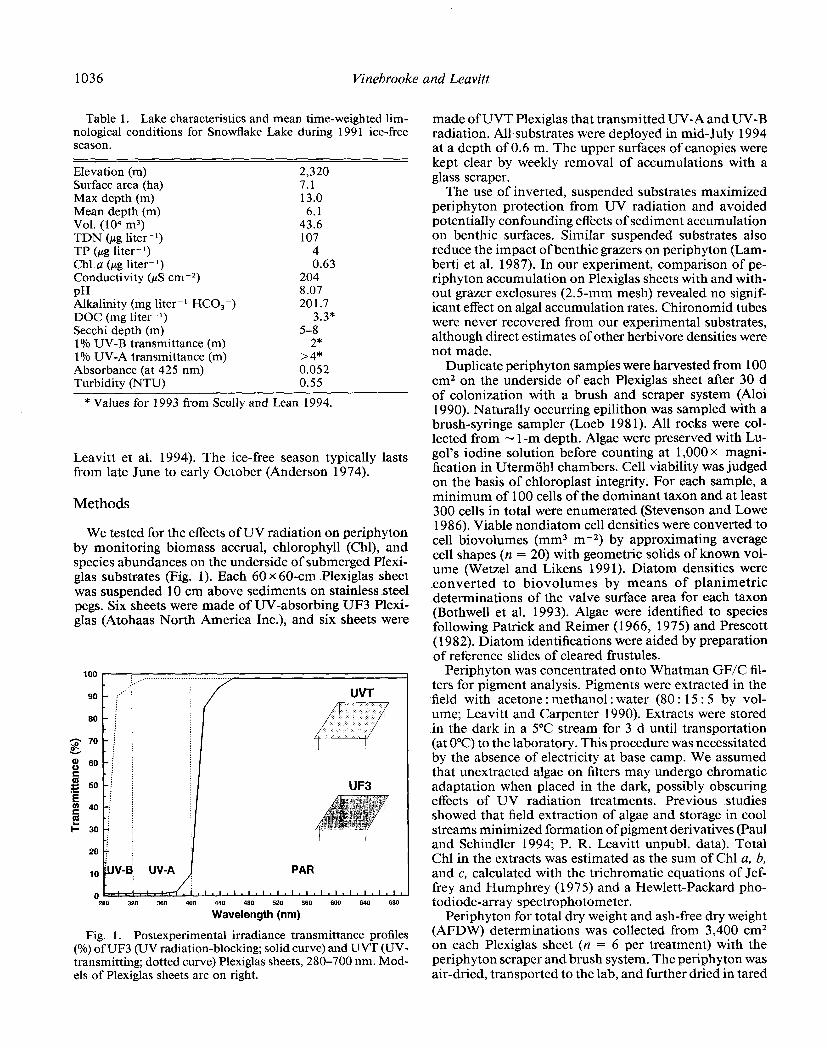
1036 Vinebrooke and Leavitt
Table 1. Lake characteristics and mean time-weighted lim- nological conditions for Snowflake Lake during 199 1 ice-free season.
Elevation (m) 2,320 Surface area (ha) 7.1 Max depth (m) 13.0 Mean depth (m) 6.1 Vol. (lo4 m3) 43.6 TDN (pg liter-‘) 107 TP (pg liter-‘) 4 Chl a (pg liter-‘) 0.63 Conductivity ($S cm-2) 204 PH 8.07 Alkalinity (mg liter-l HCO,-) 201.7 DOC (mg liter-‘) 3.3* Secchi depth (m) 5-8 1% UV-B transmittance (m) 2” 1% UV-A transmittance (m) >4* Absorbance (at 425 nm) 0.052 Turbidity (NTU) 0.55
* Values for 1993 from Scully and Lean 1994.
Leavitt et al. 1994). The ice-free season typically lasts from late June to early October (Anderson 1974).
Methods
We tested for the effects of UV radiation on periphyton by monitoring biomass accrual, chlorophyll (Chl), and species abundances on the underside of submerged Plexi- glas substrates (Fig. 1). Each 60 x60-cm Plexiglas sheet was suspended 10 cm above sediments on stainless steel pegs. Six sheets were made of UV-absorbing UF3 Plexi- glas (Atohaas North America Inc.), and six sheets were
80
‘;i 70 3 8 60
s .E 5o E 2 40
!! I- 30
. . . . . . . . . . . . . . . .
UF3
Fig. 1. Postexperimental irradiance transmittance profiles (%) of UF3 (UV radiation-blocking; solid curve) and UVT (UV- transmitting; dotted curve) Plexiglas sheets, 280-700 nm. Mod- els of Plexiglas sheets are on right.
made of UVT Plexiglas that transmitted UV-A and UV-B radiation. All substrates were deployed in mid-July 1994 at a depth of 0.6 m. The upper surfaces of canopies were kept clear by weekly removal of accumulations with a glass scraper.
The use of inverted, suspended substrates maximized periphyton protection from UV radiation and avoided potentially confounding effects of sediment accumulation on benthic surfaces. Similar suspended substrates also reduce the impact of benthic grazers on periphyton (Lam- berti et al. 1987). In our experiment, comparison of pe- riphyton accumulation on Plexiglas sheets with and with- out grazer exclosures (2.5-mm mesh) revealed no signif- icant effect on algal accumulation rates. Chironomid tubes were never recovered from our experimental substrates, although direct estimates of other herbivore densities were not made.
Duplicate periphyton samples were harvested from 100 cm2 on the underside of each Plexiglas sheet after 30 d of colonization with a brush and scraper system (Aloi 1990). Naturally occurring epilithon was sampled with a brush-syringe sampler (Loeb 198 1). All rocks were col- lected from - 1 -m depth. Algae were preserved with Lu- gol’s iodine solution before counting at 1,000 X magni- fication in Utermohl chambers. Cell viability was judged on the basis of chloroplast integrity. For each sample, a minimum of 100 cells of the dominant taxon and at least 300 cells in total were enumerated (Stevenson and Lowe 1986). Viable nondiatom cell densities were converted to cell biovolumes (mm3 m-2) by approximating average cell shapes (n = 20) with geometric solids of known vol- ume (Wetzel and Likens 1991). Diatom densities were converted to biovolumes by means of planimetric determinations of the valve surface area for each taxon (Bothwell et al. 1993). Algae were identified to species following Patrick and Reimer (1966, 1975) and Prescott ( 1982). Diatom identifications were aided by preparation of reference slides of cleared frustules.
Periphyton was concentrated onto Whatman GF/C fil- ters for pigment analysis. Pigments were extracted in the field with acetone : methanol : water (80 : 15 : 5 by vol- ume; Leavitt and Carpenter 1990). Extracts were stored in the dark in a 5°C stream for 3 d until transportation (at OOC) to the laboratory. This procedure was necessitated by the absence of electricity at base camp. We assumed that unextracted algae on filters may undergo chromatic adaptation when placed in the dark, possibly obscuring effects of UV radiation treatments. Previous studies showed that field extraction of algae and storage in cool streams minimized formation of pigment derivatives (Paul and Schindler 1994; P. R. Leavitt unpubl. data). Total Chl in the extracts was estimated as the sum of Chl a, b, and c, calculated with the trichromatic equations of Jef- frey and Humphrey (1975) and a Hewlett-Packard pho- todiode-array spectrophotometer.
Periphyton for total dry weight and ash-free dry weight (AFDW) determinations was collected from 3,400 cm2 on each Plexiglas sheet (n = 6 per treatment) with the periphyton scraper and brush system. The periphyton was air-dried, transported to the lab, and further dried in tared

UV radiation and periphyton 1037
crucibles (lOO”C, 24 h) to estimate dry tion. Samples were combusted (5 5O”C, AFDW.
Transmittance characteristics of UF3
mass accumula- 1 h) to estimate
and UVT Plexi- glas were determined at the end of the experiment. Small (2.5 x 0.9 cm) pieces of Plexiglas were cut from the sheets, mounted in quartz cuvettes in water, and irradiance trans- mission (280-700 nm) measured in a Hewlett-Packard photodiode-array spectrophotometer. Regular, direct es- timation of UV radiation dose was not possible at these remote sites. Consequently, we used previously published UV irradiance and attenuation data for Snowflake Lake in 1993 (Scully and Lean 1994) and our estimates of UV attenuation by Plexiglas to approximate the UV dose re- ceived by the periphyton. This estimate represents the maximum possible UV flux under cloudless conditions at solar noon in mid-July.
One-way ANOVA was used to test for the effects of UV radiation on accrual of total periphytic biomass, Chl, and cell biovolume. Species-specific responses to UV ra- diation were also quantified for the five dominant taxa by means of Bonferroni-adjusted t-tests (Sokal and Rohlf 1995). These species accounted for >95% of total cell biovolume in all treatments. Species biovolumes were log,,-transformed before analysis. Least-significant-dif- ference (LSD) multiple comparisons were used to deter- mine which taxa were significantly affected by UV radi- ation (Sokal and Rohlf 1995). The Shannon-Weiner index (H’) was used as a measure of the taxonomic diversity of periphyton communities.
Results and discussion
Control Plexiglas (UVT) transmitted near-ambient levels of UV radiation in comparison with UV-absorbing (UF3) sheets (Fig. 1). According to Scully and Lean (1994), Snowflake Lake received -60.0 W m-2 of integrated UV-A (320-400 nm) and - 3.0 W m-2 of integrated UV-B radiation at the lake surface under cloudless conditions in mid-July 1993. These integrated fluxes are reduced to a maximum of -20.0 W m-2 of UV-A and -0.6 of UV-B radiation at 50-cm depth. Allowing for differences in UV attenuation by Plexiglas, we estimate that periphyton un- der UV-transmitting Plexiglas was exposed to a midday maximum of - 19.6 W m-2 of UV-A and -0.6 of UV-B, and communities beneath UV-absorbing UF3 Plexiglas were exposed to -0.6 W m-2 of UV-A and -0.01 of UV-B radiation.
Periphyton accrual was reduced significantly by near- ambient levels of UV radiation (Fig. 2). Both dry weight (not shown) and AFDW were reduced - 50% (t-test; P < 0.02; 5 df) when exposed to radiation ~400 nm. Total Chl concentrations were significantly reduced (47%; t-test; P -c 0.001; 5 dl-) b y ambient levels of UV radiation, although ratios among the various chlorophylls remained constant. Total algal biovolume accrual was suppressed 66% by ambient UV radiation (t-test; P < 0.01; 5 df). Similar magnitudes of Chl, biomass, and cell biovolume responses and constant Chl ratios suggest that UV radi- ation did not cause extensive pigment bleaching. Instead,
UV Treatment
Fig. 2. Periphyton response to ambient (+UV) and reduced (-UV) UV radiation (~400 nm) during 1 month. A. Ash-free dry weight. B. Chlorophyll a + b + c. C. Total cell biovolume. Vertical bars show SE of the mean (n = 6).
these results indicate that ambient UV radiation directly inhibited periphyton development.
Suppression of periphyton accrual by UV radiation was due primarily to inhibition of diatom colonization and growth, particularly by Achnanthes minutissima Kiitzing, an immobile stalked species (Table 2, Fig. 3). Because abundances of most taxa declined with exposure to am- bient UV radiation (Fig. 3), there was little change in relative community composition. However, both Bon- ferroni-adjusted t-tests and LSD multiple comparisons showed that only changes in A. minutissima abundance were statistically significant (P < 0.05), suggesting that taxa differed in their sensitivity to UV radiation. Similarly species richness, but not Shannon-Weiner diversity (H’), was lower on UV-transmitting Plexiglas than on UV- absorbing substrates (Table 2).
High variability in periphyton response to UV radia- tion may result from species-specific sensitivity to UV exposure (Jokiel and York 1984). A. minutissima is an early colonist of substrates and is one of the few species that is consistently suppressed by ambient UV radiation at low elevations (Bothwell et al. 1993). High sensitivity

1038 Vinebrooke and Leavitt
Table 2. Mean and standard errors of species diversity, richness, and biovolume (mm3 m-2) for taxa whose abundance is > 1% of total biovolume. N = 6 for periphyton exposed to ambient (+UV) or reduced (-UV) UV radiation; N = 3 for natural rock substrates.
Treatment +uv -UV Natural
Diversity (H’) Species richness Diatoms
Achnanthes minutissima Cymbella microcephala Cymbella minuta Diatoma w&are Gomphonema parvulum Synedra amphicephala Synedra minuscula
Cyanobacteria Anabaena subcylindrica Calothrix sp. Gloeocapsa sp.
0.48 kO.09 12.5kO.7
318+ 109 2*2
18f17 0
31+29 5+4 3+2
14f12 0
19+10
0.47kO.09 15.5zk0.8
976f27 1025 23211
0 70*19
9+3 21+4
50+49 0
120+42
0.89kO.10 24k1.2
43+36 86+27
5f4 97f36
3+3 If0 6+5
242+63 786+121
50+32
ofA. minutissima to UV radiation may be a consequence of its immobility, small width (3 pm), and large surface area : volume ratio (1.7 1; Bothwell et al. 1993). Karentz et al. (199 1 b) reported that small diatoms exhibited great- er mortality and DNA damage than larger cells. Although some diatoms produce UV-absorbing mycosporinelike amino acids (Karentz et al. 199 1 b), Garcia-Pichel (1994) showed that such pigments require substantial metabolic investments and offer only limited benefits to oval cells < 10 pm wide. Instead, A. minutissima normally domi- nates diatom communities in colored subarctic (Moore 1979) and boreal lakes (Stockner and Armstrong 197 l), both habitats where UV radiation should be low (Scully and Lean 1994).
1,000
O +uv -UV
Fig. 3. Response of the five most abundant periphyton spe- cies to ambient (+UV) and reduced UV radiation (-UV) during 1 month. Means and error bars as in Fig. 2.
Slow rates of periphytic growth in cold, ultraoligotro- phic alpine waters may have contributed to the weak effect of UV radiation on periphyton community composition. Low species richness and diversity and high abundance of small adnate diatoms (Table 3) indicated that the ex- perimental periphyton was in the early stages of succes- sion (Hoagland et al. 1982; Niederhauser and Schanz 1993). Although eight of 10 dominant taxa were common to natural and experimental substrates (Table 2), the epi- lithon at -2-m depth was dominated by cyanobacteria (Calothrix sp., Anabaena subcylindrica Borge, Gloeocapsa sp.). Species composition during colonization is often more variable than in mature periphyton because of minor dif- ferences in initial conditions. Such high variability may be prolonged in alpine sites where succession can take years to complete (e.g. Stanford and Prescott 1988) and may obscure subtle effects of UV radiation on periphyton composition.
The magnitude of periphyton response to UV radiation in alpine lakes (Figs. 2, 3) was similar to that recorded in other in situ experiments despite differences in experi- mental protocols. Worrest et al. (1978) reported that di- atom growth was suppressed 20% in 4 weeks in experi- ments with artificial UV-B levels (1.4 W m-2) higher than those estimated for Snowflake Lake (0.6 W m-2). Both- well et al. (1993) also reported that short-term exposure ( < 14 d) to ambient UV radiation inhibited diatom growth by 30-40%. Although Bothwell et al. (1994) showed that long-term exposure (~28 d) can stimulate algal produc- tion by selectively eliminating benthic herbivores (chi- ronomids), their experiments were conducted in shallow troughs (1 cm deep). Such shallow water attenuates little UV radiation and may overestimate the impact of UV radiation on benthic invertebrates, particularly in com- parison to deeper waters. Additionally, their experiments excluded benthivorous fish, important predators on chi- ronomids in streams (e.g. Power 1990). Elimination of

UV radiation and periphyton 1039
herbivores by fish may re-establish inhibitory effects of UV radiation on periphyton accrual.
Although Snowflake Lake is naturally fishless, our ex- periments probably underestimated the importance of herbivores. Inverted, suspended substrates were not col- onized by chironomids and may be difficult to access for negatively buoyant herbivores such as G. Zacustris (Lam- berti et al. 1987). Consequently, we can say little about the relative importance of herbivory and UV radiation in regulating periphyton growth. Rather, our study pro- vides the first evidence that natural levels of UV radiation can significantly suppress in situ periphyton accumulation in lakes.
Two lines of evidence suggest that short-term experi- ments provide relevant information on the role of UV radiation in structuring periphyton in alpine lakes. First, colonization is quantitatively important in alpine lakes where winter ice and benthic scouring can remove pe- riphyton up to 2 m deep. In Snowflake Lake, 29% of the lake bottom lies within the 2-m isopleth-also the max- imum depth of UV-B penetration (Table 1). Within this shallow zone, mature periphyton is irregularly distrib- uted, and bare rock and mud predominate. Dense benthic mats are common only in central, deep waters of alpine lakes (Stanford and Prescott 1988; D. W. Schindler and R. Vinebrooke unpubl. data). As well, the short ice-free season (-90 d) and low water temperature (< 12°C) can significantly limit the rates of periphyton development, prolong colonization sequences throughout summer, and maximize the exposure of colonizing periphyton to UV radiation.
Paleoecological studies also support the relevance of in situ UV experiments to alpine lakes. Analyses of fossil communities from alpine and subalpine lakes demon- strate that diatoms are less abundant during historical periods when mountain lakes are above treeline than when treeline exceeds lake elevation (Hickman and Reasoner 1994; Leavitt et al. unpubl. data). In particular, A. mi- nutissima is common only when lakes are below treeline and terrestrial DOM inputs are high. This pattern is con- sistent with experiments that demonstrate the high sen- sitivity of A. minutissima to UV radiation (Bothwell et al. 1993; this study) and with the importance of this spe- cies in humic lakes of low UV penetration (Stockner and Armstrong 197 1; Moore 1979). Interestingly, these ob- servations suggest that A. minutissima may be a sensitive indicator of elevated UV radiation in western montane lakes. We are testing this hypothesis by quantifying changes in periphyton community composition along a 50-lake gradient of elevation, DOM content, and transparency to UV radiation. Preliminary survey data show that alpine lake sediments contain high concentrations of pigments similar to those produced by benthic cyanobacteria upon exposure to UV radiation (Leavitt et al. unpubl. data).
Conclusions
Our findings suggest that ambient UV radiation sup- presses periphyton development in alpine lakes by inhib-
iting diatom production. In lakes where the growing sea- son is short and waters are transparent, UV radiation can reduce primary production in the littoral zone and may offset predicted increases in production that arise from climatic warming and longer ice-free seasons. Although DOM in alpine lakes can exceed 2 mg liter - l, it is usually autochthonous in origin (Baron et al. 199 1) and less ef- fective than allochthonous DOM at absorbing UV radi- ation (McKnight et al. 1994). Consequently, we anticipate that UV radiation may remain an important control of periphytic development until suitable sources of terres- trial DOM develop.
References
ALOI, J. 1990. A critical review of recent freshwater periphyton field methods. Can. J. Fish. Aquat. Sci. 47: 656-670.
ANDERSON, R. S. 1974. Diurnal primary production patterns in seven lakes and ponds in Alberta (Canada). Oecologia 14: 1-17.
-, AND D. B. DONALD. 1978. Limnological survey of some lakes in the vicinity of the Cascade Trail, Banff Na- tional Park. Can. Wildl. Serv. Ms. Rep.
BARON, J., D. MCKNIGHT, AND A.S. DENNING. 1991. Sources of dissolved and particulate organic material in Loch Vale watershed, Rocky Mountain National Park, Colorado, USA. Biogeochemistry 15: 89-l 10.
BLUMTHALER, M., AND W. AMBACH. 1990. Indication ofin- creasing solar ultraviolet-B radiation flux in alpine regions. Science 248: 206-208.
- - AND W. REHWALD. 1992. Solar UV-A and UV-B rad;ation fluxes at two alpine stations at different altitudes. Theor. Appl. Climatol. 46: 39-44.
BOTHWELL, M. L., D. SHERBOT, AND C. M. POLLOCK. 1994. Ecosystem response to solar ultraviolet-B radiation: Influ- ence of trophic-level interactions. Science 265: 97-l 00.
- -, A. ROBERGE,AND R.J. DALEY. 1993. Influence of’natural ultraviolet radiation on lotic periphytic diatom community growth, biomass accrual, and species compo- sition: Short-term versus long-term effects. J. Phycol. 29: 24-35.
CALDWELL, M.M.,R. ROBBERECHT,AND W.D. BILLINGS. 1980. A steep latitudinal gradient of solar ultraviolet-B radiation in the arctic-alpine lift zone. Ecology 61: 600-6 11.
COOPER, W. J., D. R. S. LEAN, AND J. H. CARY. 1989. Spatial and temporal patterns of hydrogen peroxide in lake waters. Can. J. Fish. Aquat. Sci. 46: 1227-123 1.
GARCIA-PICHEL, F. 1994. A model for internal self-shading in planktonic organisms and its implications for the usefulness of ultraviolet sunscreens. Limnol. Oceanogr. 39: 1704-l 7 17.
H;~DER, D.-P. 1993. Risks of enhanced solar ultraviolet ra- diation for aquatic ecosystems. Prog. Phycol. Res. 9: l-45.
HAPPEY-WOOD, C. 1988. Vertical-migration patterns of fla- gellates in a community of freshwater benthic algae. Hy- drobiologia 161: 99-l 23.
HICKMAN, M., AND M. A. REASONER. 1994. Diatom responses to late Quaternary vegetation and climate change, and to deposition of two tephras in an alpine and a sub-alpine lake in Yoho National Park, British Columbia. J. Paleolimnol. 11: 173-188.
HOAGLAND, K. D., S. C. ROEMER, AND J. R. ROSOWSKI. 1982. Colonization and community structure of two periphyton assemblages, with emphasis on the diatoms (Bacillariophy- ceae). Am. J. Bot. 69: 188-213.

1040 Vinebrooke and Leavitt
JEFFREY, S. W., AND G. F. HUMPHREY. 1975. New spectro- scopic equations for determining chlorophylls a, b, c, and c2 in higher plants, algae and natural phytoplankton. Bio- them. Physiol. Pflanz. 167: 19 1-194.
JOKIEL, P. L., AND R. H. YORK. 1984. Importance of ultraviolet radiation in photoinhibition of microalgal growth. Limnol. Oceanogr. 29: 192-l 99.
JONES, R. I. 1992. The influence of humic substances on la- custrine planktonic food chains. Hydrobiologia 229: 73-9 1.
KARENTZ, K.,J.E. CLEAVER,AND D.L. MITCHELL. 1991 a. Cell survival characteristics and molecular responses of antarc- tic phytoplankton to ultraviolet-B radiation. J. Phycol. 27: 326-341.
-, F.S. MCEUEN, M.C. LAND,AND W.C. DUNLAP. 1991 b. Survey of mycosporine-like amino acids in antarctic marine organisms: Potential protection from ultraviolet exposure. Mar. Biol. 108: 157-166.
KIRK, J. T. 0. 1994. Optics of UV-B radiation in natural waters. Arch. Hydrobiol. 43: 1-15.
LAMBERTI, G. A., L. R. ASHKENAS, S. V. GREGORY, AND A. D. STEINMAN. 1987. Effects of three herbivores on periphy- ton communities in laboratory streams. J. N. Am. Benthol. Sot. 6: 92-l 04.
LEAVITT, P. R., AND S. R. CARPENTER. 1990. Aphotic pigment degradation in the hypolimnion: Implications for sedimen- tation studies and paleolimnology. Limnol. Oceanogr. 35: 520-535.
-, D. E. SCHINDLER, A. J. PAUL, A. K. HARDIE, AND D. W. SCHINDLER. 1994. Fossil pigment records of phyto- plankton in trout-stocked alpine lakes. Can. J. Fish. Aquat. Sci. 51: 241 l-2423.
LOEB, S. L. 198 1. An in situ method for measuring the primary productivity and standing crop of the epilithic periphyton community in lentic systems. Limnol. Oceanogr. 26: 394- 399.
M&NIGHT, D. M., E. D. ANDREWS, S. A. SPAULDING, AND G. R. AIKEN. 1994. Aquatic fulvic acids in algal-rich ant- arctic ponds. Limnol. Oceanogr. 39: 1972-1979.
MOORE, J. W. 1979. Distribution and abundance of attached littoral algae in 2 1 lakes and streams in the Northwest Ter- ritories. Can. J. Bot. 57: 568-577.
NEALE, P. J., J. J. CULLEN, M. P. LESSER, AND A. MELIS. 1993. Physiological bases for detecting and predicting photo- inhibition of aquatic photosynthesis by PAR and UV ra- diation, p. 61-77. In H. Y. Yamamoto and C. M. Smith [eds.], Photosynthetic responses to the environment. Am. Sot. Plant. Physiol.
NIEDERHAUSER, P., AND F. SCHANZ. 1993. Effects ofnutrient (N, P, C) enrichment upon the littoral diatom community of an oligotrophic high-mountain lake. Hydrobiologia 269/ 270: 453-462.
PATRICK, R., AND C. W. REIMER. 1966. The diatoms of the United States. V. 1. Acad. Nat. Sci. Phil. Monogr. 13.
-,AND- 1975. The diatoms of the United States. V. 2. Acad. Nat. Sci. Phil. Monogr. 13.
PAUL, A. J., AND D. W. SCHINDLER. 1994. Regulation of ro- tifers by predatory Calanoid copepods (subgenus Hespero- diaptomus) in lakes of the Canadian Rocky Mountains. Can. J. Fish. Aquat. Sci. 51: 2520-2528.
POWER, M. E. 1990. Effects of fish in river food webs. Science 250: 811-814.
PRESCOTT, G. W. 1982. Algae of the western Great Lakes area. Koeltz.
RODHE, W., J. E. HOBBIE, AND R. T. WRIGHT. 1966. Photot- rophy and heterotrophy in high mountain lakes. 2. Pro- duction. Int. Ver. Theor. Angew. Limnol. Verh. 16: 302- 313.
ROTT, E. 1988. Some aspects of the seasonal distribution of flagellates in mountain lakes. Hydrobiologia 161: 159-l 70.
SCHINDLER, D. W.,S. E. BAYLEY, ANDOTHERS. 1992. Natural and man-caused factors affecting the abundance and cycling ofdissolved organic substances in Precambrian shield lakes. Hydrobiologia 229: l-2 1.
-, K. G. BEATY, AND OTHERS. 1990. Effects of climatic warming on lakes of the central boreal forest. Science 250: 44-47.
SCULLY, N. M., AND D. R. S. LEAN. 1994. The attenuation of ultraviolet radiation in temperate lakes. Arch. Hydrobiol. 43: 135-144.
SMITH, R. C., AND OTHERS. 1992. Ozone depletion: Ultraviolet radiation and phytoplankton biology in antarctic waters. Science 255: 952-959.
SOKAL, R. R., AND F. J. ROHLF. 1995. Biometry, 3rd ed. Free- man.
STANFORD, J. A., AND G. W. PRESCOTT. 1988. Limnological features of a remote alpine lake in Montana, including a new species of Cladophora (Chlorophyta). J. N. Am. Ben- thol. Sot. 7: 140-15 1.
STEVENSON, R. J., AND R. L. LOWE. 1986. Sampling and in- terpretation of algal patterns for water quality assessments, p. 18-149. In B. G. Isom [ed.], Rationale for sampling and interpretation of ecological data in the assessment of fresh- water ecosystems. Am. Sot. Test. Mater. STP 894.
STOCKNER, J. G., AND F. A.J. ARMSTRONG. 1971. Periphyton of the Experimental Lakes Area, northwestern Ontario. J. Fish. Res. Bd. Can. 28: 2 15-229.
VINCENT, W. F., R. W. CASTENHOLZ, M. T. DOWNES, AND C. HOWARD-WILLIAMS. 1993. Antarctic cyanobacteria: Light, nutrients, and photosynthesis in the microbial mat envi- ronment. J. Phycol. 29: 745-755.
-, P. J. NEALE, AND P. J. RICHERSON. 1984. Photoinhi- bition: Algal responses to bright light during diel stratifi- cation and mixing in a tropical alpine lake. J. Phycol. 20: 201-211.
-, AND S. ROY. 1993. Solar ultraviolet-B radiation and aquatic primary production: Damage, protection, and re- covery. Environ. Rev. 1: 1-12.
WETZEL, R. G., AND G. E. LIKENS. 1991. Limnological anal- yses, 2nd ed. Springer.
WILLIAMSON, C. E.,H.E. ZAGARESE, P.C. SCHULZE, B. R. HAR- GREAVES, AND J. SEVA. 1994. The impact of short-term exposure to UV-B radiation on zooplankton communities in north temperate lakes. J. Plankton Res. 16: 205-218.
WORREST, R. C., H. VAN DYKE, AND B. E. THOMSON. 1978. Impact of enhanced simulated solar UV radiation upon a marine community. Photochem. Photobiol. 27: 47 l-478.




















