
Effects of three years of soil warming and shading on the rate of soil respiration: substrate...
10
Effects of three years of soil warming and shading on the rate of soil respiration: substrate availability and not thermal acclimation mediates observed response IAIN P. HARTLEY 1 , ANDREAS HEINEMEYER and PHIL INESON Biology Department, Stockholm Environment Institute (SEI-York centre), University of York, York YO10 5YW, UK Abstract In a number of recent field studies, the positive response of soil respiration to warming has been shown to decline over time. The two main differing hypotheses proposed to explain these results are: (1) soil microbial respiration acclimates to the increased temperature, and (2) substrate availability within the soil decreases with warming so reducing the rate of soil respiration. To investigate the relative merits of these two hypotheses, soil samples (both intact cores and sieved samples) from a 3-year grassland soil-warming and shading experiment were incubated for 4 weeks at three different temperatures under constant laboratory conditions. We tested the hypothesis that sieving the soils would reduce differences in substrate availability between warmed and control plot samples and would therefore result in similar respiration rates if microbial activity had not acclimated to soil warming. In addition, to further test the effect of substrate availability, we compared the respiration rates of soils taken from shaded and unshaded plots. Both soil warming and shading significantly reduced respiration rates in the intact cores, especially under higher incubation temperatures. However, sieving the soil greatly reduced these differences suggesting that substrate availability, and not microbial acclimation to the higher temperatures, played the dominant role in determining the response of heterotrophic soil respiration to warming. The effect of shading appeared to be mediated by reduced plant productivity affecting substrate availability within the soil and hence microbial activity. Given the lack of evidence for thermal acclimation of microbial respiration, there remains the potential for prolonged carbon losses from soils in response to warming. Keywords: acclimation, CO 2 , heterotrophic respiration, incubation, microbial community, positive feedback, root biomass, soil warming, substrate availability, temperature Received 8 November 2006; revised version received 8 February 2007 and accepted 5 February 2007 Introduction Only approximately 45% of the CO 2 released by anthro- pogenic activities has ended up in the atmosphere and it is believed that increased photosynthesis in terrestrial ecosystems has absorbed a large proportion of the remaining emissions (Houghton et al., 1998; Intergo- vernmental Panel on Climate Change, 2001). However, Cox et al. (2000) suggested that the positive effect of increasing global temperatures on soil respiration may result in terrestrial ecosystem losing soil carbon (C) and so enhance the rate of climate change by 35%. If these results are correct, then C-cycle feedbacks have the potential to undermine international attempts to curb global CO 2 emissions (Bellamy et al., 2005). Therefore, it is necessary to undertake a critical evaluation of the assumptions underlying this modelling study, the key one being that soil organic matter (SOM) decomposition will respond to long-term warming with a Q 10 of 2 (i.e. heterotrophic soil respiration will increase exponen- tially, with its rate doubling for every 10 1C rise in temperature). A number of recent studies have chal- lenged this assumption. Long-term monitoring in the field has identified temperature as a key determinant of soil respiration 1 Present address: School of Biological and Environmental Sciences, University of Stirling, Stirling FK9 4LA, UK. Correspondence: Iain P. Hartley, fax 1 44 1786 467843, e-mail: [email protected] Global Change Biology (2007) 13, 1761–1770, doi: 10.1111/j.1365-2486.2007.01373.x r 2007 The Authors Journal compilation r 2007 Blackwell Publishing Ltd 1761
Transcript of Effects of three years of soil warming and shading on the rate of soil respiration: substrate...

Effects of three years of soil warming and shading on therate of soil respiration: substrate availability and notthermal acclimation mediates observed response
I A I N P. H A R T L E Y 1 , A N D R E A S H E I N E M E Y E R and P H I L I N E S O N
Biology Department, Stockholm Environment Institute (SEI-York centre), University of York, York YO10 5YW, UK
Abstract
In a number of recent field studies, the positive response of soil respiration to warming
has been shown to decline over time. The two main differing hypotheses proposed to
explain these results are: (1) soil microbial respiration acclimates to the increased
temperature, and (2) substrate availability within the soil decreases with warming so
reducing the rate of soil respiration. To investigate the relative merits of these two
hypotheses, soil samples (both intact cores and sieved samples) from a 3-year grassland
soil-warming and shading experiment were incubated for 4 weeks at three different
temperatures under constant laboratory conditions. We tested the hypothesis that sieving
the soils would reduce differences in substrate availability between warmed and control
plot samples and would therefore result in similar respiration rates if microbial activity
had not acclimated to soil warming. In addition, to further test the effect of substrate
availability, we compared the respiration rates of soils taken from shaded and unshaded
plots. Both soil warming and shading significantly reduced respiration rates in the intact
cores, especially under higher incubation temperatures. However, sieving the soil greatly
reduced these differences suggesting that substrate availability, and not microbial
acclimation to the higher temperatures, played the dominant role in determining the
response of heterotrophic soil respiration to warming. The effect of shading appeared to
be mediated by reduced plant productivity affecting substrate availability within the soil
and hence microbial activity. Given the lack of evidence for thermal acclimation of
microbial respiration, there remains the potential for prolonged carbon losses from soils
in response to warming.
Keywords: acclimation, CO2, heterotrophic respiration, incubation, microbial community, positive
feedback, root biomass, soil warming, substrate availability, temperature
Received 8 November 2006; revised version received 8 February 2007 and accepted 5 February 2007
Introduction
Only approximately 45% of the CO2 released by anthro-
pogenic activities has ended up in the atmosphere and
it is believed that increased photosynthesis in terrestrial
ecosystems has absorbed a large proportion of the
remaining emissions (Houghton et al., 1998; Intergo-
vernmental Panel on Climate Change, 2001). However,
Cox et al. (2000) suggested that the positive effect of
increasing global temperatures on soil respiration may
result in terrestrial ecosystem losing soil carbon (C) and
so enhance the rate of climate change by �35%. If these
results are correct, then C-cycle feedbacks have the
potential to undermine international attempts to curb
global CO2 emissions (Bellamy et al., 2005). Therefore,
it is necessary to undertake a critical evaluation of the
assumptions underlying this modelling study, the key
one being that soil organic matter (SOM) decomposition
will respond to long-term warming with a Q10 of 2 (i.e.
heterotrophic soil respiration will increase exponen-
tially, with its rate doubling for every 10 1C rise in
temperature). A number of recent studies have chal-
lenged this assumption.
Long-term monitoring in the field has identified
temperature as a key determinant of soil respiration
1Present address: School of Biological and Environmental Sciences,
University of Stirling, Stirling FK9 4LA, UK.
Correspondence: Iain P. Hartley, fax 144 1786 467843,
e-mail: [email protected]
Global Change Biology (2007) 13, 1761–1770, doi: 10.1111/j.1365-2486.2007.01373.x
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd 1761

rates (e.g. Raich & Schlesinger, 1992; Lloyd & Taylor,
1994; Davidson et al., 1998; Rodeghiero & Cescatti,
2005), and laboratory incubations of both intact cores
(e.g. Fang & Moncrieff, 2001; Reichstein et al., 2005) and
sieved root-free samples (e.g. Bowden et al., 1998; Fang
et al., 2005) have also demonstrated a strong tempera-
ture dependence. Further, a meta-analysis of the results
from 17 different long-term field-warming experiments
identified a statistically significant increase in the rate of
soil respiration following warming (Rustad et al., 2001),
yet the magnitude of this increase was not as large as
predicted from short-term experiments. In addition, a
number of studies have observed that the initial in-
crease in the rate of soil respiration declines over time
(Rustad et al., 2001; Eliasson et al., 2005). These observa-
tions have led authors to conclude that soil respiration
may ‘acclimate’ to warming and so dampen any
potential positive feedback to global climate change
(Luo et al., 2001).
Two main explanations for soil respiration ‘acclima-
tion’ have been proposed. Firstly, soil warming may
induce a change in the composition of the microbial
community within soil (Zogg et al., 1997; Andrews et al.,
2000; Zhang et al., 2005) with the decomposition activ-
ities associated with the new community having a
different temperature dependence (Luo et al., 2001).
Alternatively, modelling studies have suggested that
soil respiration responses to increased temperature are
likely to be limited by substrate availability, with labile
C pools becoming depleted by increased activity in
response to warming, and so leading to a subsequent
reduction in the rate of soil respiration (Kirschbaum,
2004; Eliasson et al., 2005; Knorr et al., 2005). These two
conflicting hypotheses may have very different conse-
quences. If acclimation/adaptation of the microbial
community is central to the reduction in the rate
of respiration in warmed plots, then soil C stocks may
be preserved and the potential for C sequestration
in terrestrial ecosystems maintained. However, if sub-
strate limitation is the key driver then there remains
the potential for reduced sequestration of recently
plant-derived C and, ultimately, C losses from soils
(Eliasson et al., 2005).
A further complication exists when investigating the
responses of soil microbes to soil warming due to the
potentially large contribution (�50%) of root respiration
to belowground respiration (Hanson et al., 2000). Root
respiration and SOM decomposition may respond dif-
ferently to long-term changes in temperature (Fitter
et al., 1998; Huang et al., 2005) and difficulties associated
with separating these two terms (Hanson et al., 2000)
mean that direct measurements of the response of
heterotrophic respiration to increases in soil tempera-
ture are often difficult in situ. However, removing soil
samples from the field severs all connections to assim-
ilate supply and results in a rapid decline in the rate of
root respiration (Langley et al., 2005). Therefore, the
incubation of soils taken from soil warming experi-
ments can allow the effects on heterotrophic respiration
to be measured directly.
In addition, by breaking up aggregates and making
previously physically protected SOM available, sieving
can reduce differences in substrate availability between
soil samples and generally causes an increase in the rate
of respiration (Franzluebbers, 1999). In terms of redu-
cing substrate availability differences between samples,
sieving has an advantage over single substrate addi-
tions which, although providing an estimate of micro-
bial biomass, tend to promote the proliferation of
microbes especially adapted to the particular substrate
added (Pennanen et al., 2004). In addition the very rapid
pulse of respiration associated with substrate-induced
respiration was not suitable for the type of investigation
being carried out here.
The main aim of our study was to determine whether
microbial community acclimation/adaptation or sub-
strate depletion is the key driver of the reduction in the
positive warming response of soil respiration observed
in many long-term warming experiments (Rustad et al.,
2001). Intact cores and sieved samples taken from a full
factorial, 3-year grassland soil-warming and shading
experiment were incubated at three different tempera-
tures. The shading treatment altered plant growth rates
(Edwards et al., 2004), and hence, the amount of new C
entering the soil. It, therefore, provided a further op-
portunity to investigate the importance of substrate
availability in determining the respiration rates in the
intact cores vs. sieved samples. We tested the following
two main hypotheses: (1) previous experience of 3 years
of soil warming would reduce the respiration rates of
intact cores when incubated at a common temperature,
and (2) this response would be mediated through
changes in substrate availability within the soil and so
differences between warmed and ambient plot samples
would be lost when sieving made additional substrates
available (alternatively, if differences were maintained,
this would be consistent with the response being
mediated by thermal acclimation of the microbial com-
munity).
Materials and methods
Field experimental design
The soil incubated in this experiment was a sandy loam
(pH 6.5) which had been taken from the soil-warming
and shading experiment established in the experimen-
tal garden at the University of York. This experiment
1762 I . P. H A R T L E Y et al.
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770

consisted of twelve main 2 m2 plots which were seeded
with a mixture of northern grassland species in July
1998 (see Edwards et al., 2004), with Holcus lanatus L.
later becoming the dominant species. Each main plot
contained two subplots, one ambient temperature
subplot and a subplot in which the soil was warmed
to 3 1C above ambient (Fig. 1) using low-voltage heating
cables connected to a 1 m� 0.5 m steel grid with a
25mm� 25 mm mesh size (Ineson et al., 1998). The steel
grid resulted in the heat being spread evenly through-
out the plot and so minimized lateral thermal gradients.
Controllers, connected to arrays of thermistor probes
placed 2 cm below the heating meshes, maintained a
3 1C difference between warmed and ambient subplots.
Warming was switched on continuously from mid
January 2000 until the soils were sampled on 30th
January 2003.
Of the 12 pairs of plots, four were left unshaded and
the other eight plots were exposed to two levels of
shading (‘half shade’ and ‘full shade’; Fig. 1). The half
shade treatment blocked 70% of ambient photosynthetic
photon density and the full shade treatment blocked out
86%. These shading values were measured using PAR
sensors placed 75 cm above the ground, but shading
at leaf level may have been lower (Heinemeyer et al.,
2004).
Soil sampling
On 30th January 2003 the heating cables were removed
and four soil cores (68 mm in diameter and 100 mm in
depth) were taken from each plot. On the following day,
to avoid between-plot differences in soil moisture con-
tent, the cores were adjusted to water holding capacity
by adding deionized water. Three of the cores were left
intact and placed into 2 L jars and then incubated at
three different temperatures.
Two days after the soils were adjusted to water
holding capacity, the final core was dry-sieved first
through a 2 mm mesh and then through a 500 mm mesh
with roots being removed at each step. Approximately
10 g of sieved soil was added to each of three 125 cm3
bottles and these were then incubated at the three
different temperatures.
Soil incubation
The cores and sieved samples were incubated at 4, 8
and 12 1C for 4 weeks. The 4 1C treatment reflected the
mean soil temperature that the control plots had ex-
perienced over the preharvest month while 12 1C was
the maximum soil temperature recorded in the warmed
subplots over the preharvest month. During incubation,
the intact cores and sieved samples were maintained at
water holding capacity by weighing the bottles and jars
every 2 days and then adding the correct amount of
deionized water. Deionized water reservoirs were
maintained in each incubator to allow the water addi-
tions to be made at the correct temperature.
Respiration measurement
After 3 days of incubation at each temperature, the
respiration rates of the intact cores were measured.
The jars containing the intact cores were closed and
over-pressured with 50 cm3 of CO2-free air. The head-
space was initially sampled (t0) and again after 3 h of
incubation by removing 10 cm3 of headspace gas into
stoppered syringes. The jars were then opened until the
next set of measurements.
Two days after sieving, measurements of the respira-
tion rates in the sieved samples were made. As for the
measurements made on the intact cores, the serum
bottles were sealed and then over-pressured, but this
time using 25 cm3 of CO2-free air. Again, the headspace
was sampled at t0 and then after 3 h of incubation. The
serum bottles were then reopened until the next set of
measurements. Soil respiration rates were measured on
a further three occasions with a sampling interval of 1
week for both the intact cores and the sieved samples.
To measure the change in the headspace CO2 con-
centration over the period of incubation, gas samples
were injected into a gas sampling valve (Rheodyne Inc.,
Bensheim, Germany) connected to a 1 cm3 gas sampling
loop. The gas was then injected into a CO2-free air
Unshaded
UnshadedUnshaded
Unshaded
Full shade Full shade
Full shade
Full shade
Half shade
Half shade
Half shade
Half shade
Warmed
Ambient
1 : 1 1 : 2 1 : 3 1 : 4 1 : 5 1 : 6 1 : 7 1 : 8
2 : 1 2 : 2 2 : 3 2 : 4 2 : 5 2 : 6 2 : 7 2 : 8
3 : 1 3 : 2 3 : 3 3 : 4 3 : 5 3 : 6 3 : 7 3 : 8
1 m
Fig. 1 Diagram of the experimental set-up of the soil-warming
meshes and shading treatments in the experimental garden at
the University of York.
WA R M I N G E F F E C T S O N S O I L R E S P I R AT I O N 1763
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770

stream flowing through an infra-red gas analyser (model:
ADC-225 MK3, ADC Bioscientific Ltd, Herts, UK) and
peak heights were recorded on a chart recorder (model:
SE 120 BBC Goerz Metrawatt, Austria). Standards of 377
and 512 ppm were run between every set of 10 samples.
Root biomass and soil dry-weight analysis
At the end of the incubation, the intact cores were
broken up and sieved. Stones, fine and coarse roots
were removed and weighed, with fine roots being
defined as roots with a diameter o1 mm. The roots
and stones were oven-dried at 80 1C and weighed to
produce a measurement of root and stone content for
each of the incubated cores. Subsamples of soil from
each intact core and sieved sample were removed and
oven-dried at 105 1C to obtain a measure of soil moist-
ure content which was in turn used to calculate the dry
weight of soil in each sample.
C content and d13C ratio analysis
Root-free soil samples and H. lanatus leaves removed
from each plot were oven-dried at 60 and 105 1C
respectively, and then ground in a custom-built end-
over-end shaker, before being weighed into 6 mm� 4 mm
tin cups (Elemental Microanalysis Ltd, Okehampton,
UK). These samples were then run, with sugar stan-
dards, through a elemental analyser combustion unit
(Flash EA1112, ThermoFinnigan, CA, USA) coupled to
a isotope ratio mass spectrometer (Provac Services,
Cheshire, UK) to allow their d13C ratios and C contents
to be measured.
Data analysis
Root biomasses were expressed in terms of milligram
dry weight of root per gram dry weight of soil
(mg g dw�1). Headspace volumes, in the jars and Whea-
ton bottles, were measured allowing CO2 fluxes to be
expressed in microgram of C per gram dry weight of
soil per hour (mg C g dw�1 h�1). The sieved samples
taken from the first unshaded plot (subplots 1 : 1 and
1 : 2) respired at a rate well over twice as high as the
samples taken from all other plots. This was linked to
an ant colony which had developed in this specific plot,
and therefore, these samples were considered as out-
liers and were not included in the final analysis.
Statistical analyses were carried out using SPSS (SPSS
Science, Birmingham, UK). Repeated-measures ANOVAs,
using warmed and ambient subplots as the within-
subject variables and shading as the between-subject
factor, were used to investigate whether there were
significant effects of the warming and shading treat-
ments on (1) the respiration rates of the intact cores
and restructured samples, (2) root biomass, and (3) the
d13C ratio and C content of the H. lanatus leaves and
soil samples taken from the plots. Repeated-measures
ANOVAs were also used to investigate whether the
differences between warmed and ambient subplot soils
changed through time and, when significant differences
were observed, paired t-tests were used to investigate
which weeks differed significantly. Linear regressions
were used to investigate the relationship between root
biomass and the respiration rates of the intact cores.
Results
Intact cores
Respiration rate. Overall, significant differences were
observed between the respiration rates of the intact
cores taken from the warmed and ambient subplots,
and also between intact cores taken from the three
different shading treatments (Fig. 2). At 4 1C, signi-
ficant differences were only observed between
shading treatments, with samples taken from the full
shade treatment respiring at a rate significantly lower
than both unshaded (P 5 0.020) and half shade
(P 5 0.019) samples. For the samples incubated at 8 1C,
there was also no significant warming effect despite
cores taken from ambient plots respiring at on average
a 29% higher rate. However, the respiration rate of
samples taken from the unshaded plots was
significantly higher than that of samples taken from
the half shade (P 5 0.003) and full shade treatments
(P 5 0.001). However, at 12 1C a significant difference
Soi
l res
pira
tion
rate
(µg
C g
dw
−1 h
−1)
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
8°C12°C
Half shade Full shadeUnshadedAmbient Warmed Ambient Warmed Ambient Warmed
4°C
Fig. 2 The respiration rate measured in the intact cores taken
from the different warming and shading treatments and incu-
bated at the three different temperatures. The rates represent
the mean of the four measurement weeks. Error bars represent
11 SE (n 5 4).
1764 I . P. H A R T L E Y et al.
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770
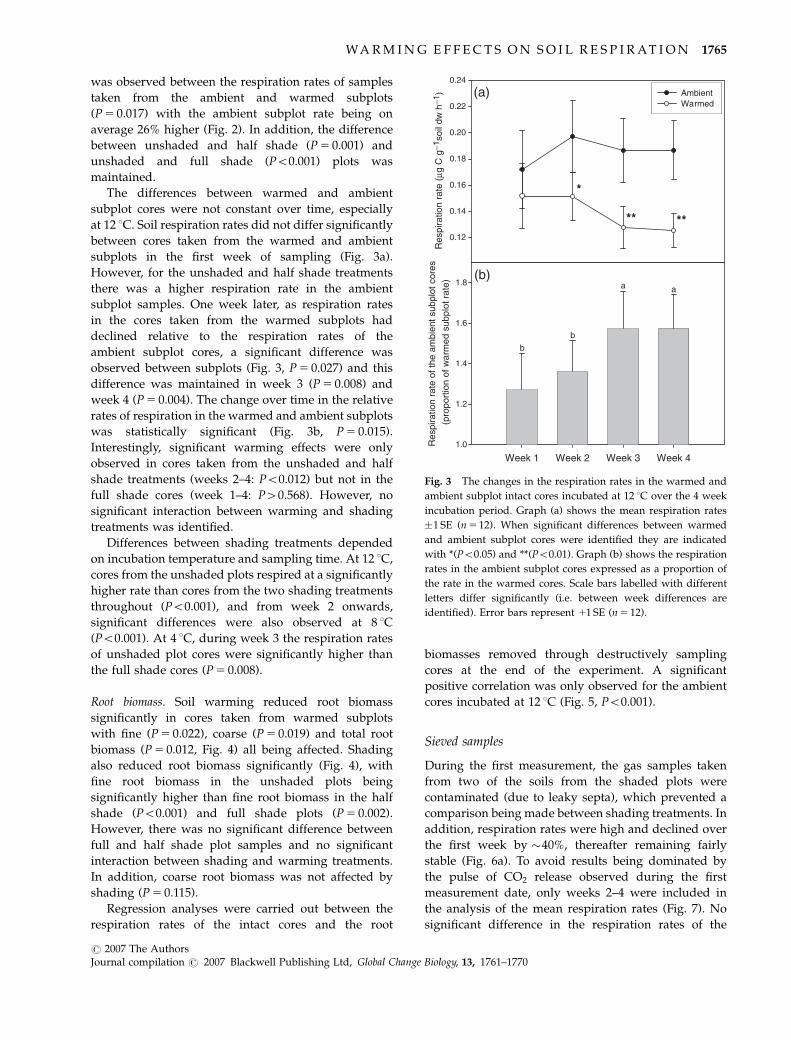
was observed between the respiration rates of samples
taken from the ambient and warmed subplots
(P 5 0.017) with the ambient subplot rate being on
average 26% higher (Fig. 2). In addition, the difference
between unshaded and half shade (P 5 0.001) and
unshaded and full shade (Po0.001) plots was
maintained.
The differences between warmed and ambient
subplot cores were not constant over time, especially
at 12 1C. Soil respiration rates did not differ significantly
between cores taken from the warmed and ambient
subplots in the first week of sampling (Fig. 3a).
However, for the unshaded and half shade treatments
there was a higher respiration rate in the ambient
subplot samples. One week later, as respiration rates
in the cores taken from the warmed subplots had
declined relative to the respiration rates of the
ambient subplot cores, a significant difference was
observed between subplots (Fig. 3, P 5 0.027) and this
difference was maintained in week 3 (P 5 0.008) and
week 4 (P 5 0.004). The change over time in the relative
rates of respiration in the warmed and ambient subplots
was statistically significant (Fig. 3b, P 5 0.015).
Interestingly, significant warming effects were only
observed in cores taken from the unshaded and half
shade treatments (weeks 2–4: Po0.012) but not in the
full shade cores (week 1–4: P40.568). However, no
significant interaction between warming and shading
treatments was identified.
Differences between shading treatments depended
on incubation temperature and sampling time. At 12 1C,
cores from the unshaded plots respired at a significantly
higher rate than cores from the two shading treatments
throughout (Po0.001), and from week 2 onwards,
significant differences were also observed at 8 1C
(Po0.001). At 4 1C, during week 3 the respiration rates
of unshaded plot cores were significantly higher than
the full shade cores (P 5 0.008).
Root biomass. Soil warming reduced root biomass
significantly in cores taken from warmed subplots
with fine (P 5 0.022), coarse (P 5 0.019) and total root
biomass (P 5 0.012, Fig. 4) all being affected. Shading
also reduced root biomass significantly (Fig. 4), with
fine root biomass in the unshaded plots being
significantly higher than fine root biomass in the half
shade (Po0.001) and full shade plots (P 5 0.002).
However, there was no significant difference between
full and half shade plot samples and no significant
interaction between shading and warming treatments.
In addition, coarse root biomass was not affected by
shading (P 5 0.115).
Regression analyses were carried out between the
respiration rates of the intact cores and the root
biomasses removed through destructively sampling
cores at the end of the experiment. A significant
positive correlation was only observed for the ambient
cores incubated at 12 1C (Fig. 5, Po0.001).
Sieved samples
During the first measurement, the gas samples taken
from two of the soils from the shaded plots were
contaminated (due to leaky septa), which prevented a
comparison being made between shading treatments. In
addition, respiration rates were high and declined over
the first week by �40%, thereafter remaining fairly
stable (Fig. 6a). To avoid results being dominated by
the pulse of CO2 release observed during the first
measurement date, only weeks 2–4 were included in
the analysis of the mean respiration rates (Fig. 7). No
significant difference in the respiration rates of the
Res
pira
tion
rate
(µg
C g
−1so
il dw
h−1
)
0.12
0.14
0.16
0.18
0.20
0.22
0.24
Ambient Warmed
*
** **
(a)
Week 1 Week 2 Week 3 Week 4
Res
pira
tion
rate
of t
he a
mbi
ent s
ubpl
ot c
ores
(pro
port
ion
of w
arm
ed s
ubpl
ot r
ate)
1.0
1.2
1.4
1.6
1.8(b)
b
b
a a
Fig. 3 The changes in the respiration rates in the warmed and
ambient subplot intact cores incubated at 12 1C over the 4 week
incubation period. Graph (a) shows the mean respiration rates
�1 SE (n 5 12). When significant differences between warmed
and ambient subplot cores were identified they are indicated
with *(Po0.05) and **(Po0.01). Graph (b) shows the respiration
rates in the ambient subplot cores expressed as a proportion of
the rate in the warmed cores. Scale bars labelled with different
letters differ significantly (i.e. between week differences are
identified). Error bars represent 11 SE (n 5 12).
WA R M I N G E F F E C T S O N S O I L R E S P I R AT I O N 1765
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770

samples taken from the warmed and ambient subplots
was observed at any of the three incubation tempera-
tures, even when excluding the full shade plots. At
12 1C the respiration measured in samples taken from
unshaded plots was significantly higher than in the half
shade (P 5 0.022) and full shade samples (P 5 0.009).
However, as with the intact cores, between weeks the
respiration rates of the warmed subplot samples de-
clined as a proportion of the respiration rates in the
ambient subplot samples but only when incubated at
12 1C (Fig. 6b, P 5 0.008). By week 3, there was a
significant difference between the rate of respiration in
the warmed and ambient subplot samples (Fig. 6a,
P 5 0.045) and this difference was maintained in week
4 (P 5 0.048).
Throughout weeks 2–4, the samples taken from the
unshaded plots respired at a significantly higher rate
than soil from the half and full shade plots but only
when incubated at 12 1C (Fig. 7).
C content and d13C ratios
The H. lanatus leaf samples taken from the warmed
subplots had a significantly more negative d13C ratio
than the samples from the ambient subplots (Table 1)
but no further significant warming or shading effects on
the C content or d13C ratio of the leaves or soil were
observed (Table 1).
Discussion
The effect of soil warming
Respiration rates. We hypothesized that prior experience
of three years of soil warming would reduce the
respiration rate of intact cores when incubated at a
common temperature. This hypothesis was supported
by the fact that warmed subplot cores respired at a
significantly lower rate than ambient subplot cores
when incubated at 12 1C (Fig. 2). The reduction in the
respiration rate of the warmed subplots samples was
on average 26%. This result is in broad agreement with
the findings of other long-term field experiments in
which soil warming has only resulted in a transient
stimulation of soil respiration (Luo et al., 2001; Rustad
et al., 2001; Eliasson et al., 2005). However, it should be
mentioned that during the incubation of the intact
cores, root death probably occurred which could have
affected substrate availability and microbial activity.
Therefore, the rates observed in the intact cores may
not have been completely indicative of the rates of
respiration occurring under undisturbed field conditions.
We hypothesized that, by breaking up soil
aggregates and making previously protected SOM
available, sieving would reduce the differences in
respiration rates between warmed and ambient
subplot soils if substrate availability rather than
thermal acclimation was responsible for the observed
reduction in the respiration rates of the warmed subplot
intact cores. In support of this hypothesis, overall the
respiration rates in the sieved samples taken from
the warmed subplots were only 6% lower than those
in ambient subplot samples when incubated at 12 1C
(Fig. 7), and significant differences were not observed
until after 3 weeks of incubation (Fig. 6a). Further, the
near identical rates of respiration early in the incubation
(Fig. 6b) suggest that microbial respiration had not
acclimated to the soil warming treatment. Over time,
the decline in the rate of respiration in both the intact
Unshaded Half shade Full shade
Tot
al r
oot b
iom
ass
(mg
g dw
−1)
0.0
0.2
0.4
0.6
0.8
1.6
1.4
1.2
1.0
AmbientWarmed
Fig. 4 The mean total root biomass extracted from the intact
cores taken from the different warming and shading treatments.
Root biomasses are expressed as mg dry weight of roots per
gram dry weight of soil (mg g dw�1). Error bars represent
11 SE (n 5 4).
Total root biomass (mg g dw−1)
0.0 0.5 1.0 1.5 2.0 2.5
Res
pira
tion
rate
(µg
C g
dw
−1 h
−1)
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
y = 0.099x + 0.129
R2 = 0.83
Fig. 5 The relationship between the total dry-weight of roots
taken from the ambient cores, incubated at 12 1C, and the mean
respiration rate of each core over the 4 week period. A linear
regression is plotted.
1766 I . P. H A R T L E Y et al.
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770
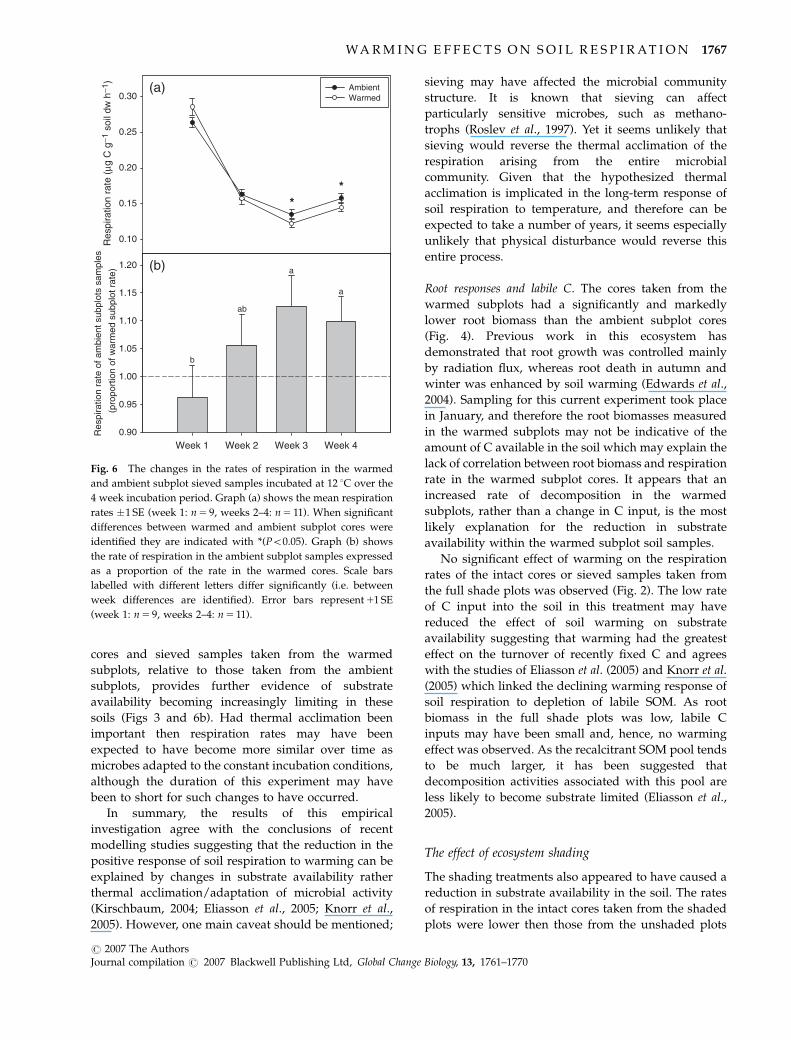
cores and sieved samples taken from the warmed
subplots, relative to those taken from the ambient
subplots, provides further evidence of substrate
availability becoming increasingly limiting in these
soils (Figs 3 and 6b). Had thermal acclimation been
important then respiration rates may have been
expected to have become more similar over time as
microbes adapted to the constant incubation conditions,
although the duration of this experiment may have
been to short for such changes to have occurred.
In summary, the results of this empirical
investigation agree with the conclusions of recent
modelling studies suggesting that the reduction in the
positive response of soil respiration to warming can be
explained by changes in substrate availability rather
thermal acclimation/adaptation of microbial activity
(Kirschbaum, 2004; Eliasson et al., 2005; Knorr et al.,
2005). However, one main caveat should be mentioned;
sieving may have affected the microbial community
structure. It is known that sieving can affect
particularly sensitive microbes, such as methano-
trophs (Roslev et al., 1997). Yet it seems unlikely that
sieving would reverse the thermal acclimation of the
respiration arising from the entire microbial
community. Given that the hypothesized thermal
acclimation is implicated in the long-term response of
soil respiration to temperature, and therefore can be
expected to take a number of years, it seems especially
unlikely that physical disturbance would reverse this
entire process.
Root responses and labile C. The cores taken from the
warmed subplots had a significantly and markedly
lower root biomass than the ambient subplot cores
(Fig. 4). Previous work in this ecosystem has
demonstrated that root growth was controlled mainly
by radiation flux, whereas root death in autumn and
winter was enhanced by soil warming (Edwards et al.,
2004). Sampling for this current experiment took place
in January, and therefore the root biomasses measured
in the warmed subplots may not be indicative of the
amount of C available in the soil which may explain the
lack of correlation between root biomass and respiration
rate in the warmed subplot cores. It appears that an
increased rate of decomposition in the warmed
subplots, rather than a change in C input, is the most
likely explanation for the reduction in substrate
availability within the warmed subplot soil samples.
No significant effect of warming on the respiration
rates of the intact cores or sieved samples taken from
the full shade plots was observed (Fig. 2). The low rate
of C input into the soil in this treatment may have
reduced the effect of soil warming on substrate
availability suggesting that warming had the greatest
effect on the turnover of recently fixed C and agrees
with the studies of Eliasson et al. (2005) and Knorr et al.
(2005) which linked the declining warming response of
soil respiration to depletion of labile SOM. As root
biomass in the full shade plots was low, labile C
inputs may have been small and, hence, no warming
effect was observed. As the recalcitrant SOM pool tends
to be much larger, it has been suggested that
decomposition activities associated with this pool are
less likely to become substrate limited (Eliasson et al.,
2005).
The effect of ecosystem shading
The shading treatments also appeared to have caused a
reduction in substrate availability in the soil. The rates
of respiration in the intact cores taken from the shaded
plots were lower then those from the unshaded plots
Res
pira
tion
rate
(µg
C g
−1 s
oil d
w h
−1)
0.10
0.15
0.20
0.25
0.30AmbientWarmed
**
Week 1 Week 2 Week 3 Week 4
Res
pira
tion
rate
of a
mbi
ent s
ubpl
ots
sam
ples
(pro
port
ion
of w
arm
ed s
ubpl
ot r
ate)
0.90
0.95
1.00
1.05
1.10
1.15
1.20 (b)
(a)
b
ab
a
a
Fig. 6 The changes in the rates of respiration in the warmed
and ambient subplot sieved samples incubated at 12 1C over the
4 week incubation period. Graph (a) shows the mean respiration
rates �1 SE (week 1: n 5 9, weeks 2–4: n 5 11). When significant
differences between warmed and ambient subplot cores were
identified they are indicated with *(Po0.05). Graph (b) shows
the rate of respiration in the ambient subplot samples expressed
as a proportion of the rate in the warmed cores. Scale bars
labelled with different letters differ significantly (i.e. between
week differences are identified). Error bars represent 11 SE
(week 1: n 5 9, weeks 2–4: n 5 11).
WA R M I N G E F F E C T S O N S O I L R E S P I R AT I O N 1767
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770

when incubated both at 8 and 12 1C (Fig. 2). It appears
that shading, through effects on plant productivity and
root biomass (Edwards et al., 2004), reduced substrate
availability in the soil, consequently reducing hetero-
trophic soil respiration especially at higher incubation
temperatures. This result is in agreement with the study
of Bond-Lamberty et al. (2004) that identified a strong
link between autotrophic and heterotrophic soil respira-
tion across forest sites and linked this to variations in
primary productivity. In addition, in situ, the impor-
tance of photosynthetic substrate availability in regulat-
ing belowground respiration is being increasingly
recognized (Craine et al., 1999; Hogberg et al., 2001;
Janssens et al., 2001; Hartley et al., 2006).
In the sieved samples incubated at 12 1C, respiration
rates in the shaded plot samples were significantly
lower than those in the unshaded plot samples
(Fig. 7). However, at 12 1C the magnitude of the differ-
ence between shaded and unshaded intact cores was
�45% and sieving reduced this to �15%. Again, it
appears that sieving reduced differences in substrate
availability between treatments.
C content and d13C ratios
The d13C ratio of H. lanatus leaves taken from the
warmed subplots was significantly more negative than
the d13C ratio of leaves taken from the ambient subplots
(Table 1). If soil-warming had resulted in an increase in
water-stress in the warmed subplot plants, then it
would have been expected that stomatal apertures
would have been decreased causing a reduction in
C-isotope discrimination and a consequent increase in
the d13C ratio (Smedley et al., 1991). Thus, no evidence
for soil warming resulting in water-stressed plants was
produced. Therefore, in agreement with Edwards et al.
(2004), there was little isotopic evidence of soil warming
affecting plant growth in this system.
Despite respiration measurements identifying a re-
duction in substrate availability in the soils taken from
the shaded plots and warmed subplots, no effect was
observed on the bulk C content of the soils (Table 1). This
result demonstrates how difficult it is to measure a
‘small change in a big number’ such as a treatment
effect on the amount of C stored in soils, as such
changes are likely to be within the error term of the
measurement techniques used (Valentini et al., 2000).
Further work is required that investigates the effect of
soil warming on the different SOM pools. Again, in
support of previous modelling exercises (Eliasson et al.,
2005; Knorr et al., 2005), a possible explanation for the
patterns observed in the study presented here, is that
changes in the size of the relatively small labile C pool
are responsible for the changes in respiration rates
and that these changes cannot be detected in the
bulk C content of the soil. Isotopic (Heath et al., 2005;
Trueman & Gonzalez-Meler, 2005; Trumbore, 2006)
and fractionation approaches (Cardon et al., 2001; Chris-
tensen, 2001) may be necessary to identify direct effects
of treatments on SOM turnover and storage.
Conclusions
Intact soil cores taken from plots exposed to three-years
of warming and shading, respired at lower rates than
cores taken from ambient subplots and unshaded plots.
Table 1 The effect of shading and warming on the d13C ratio and carbon (C) content (%C) of the soil and Holcus lanatus leaves
Unshaded Half shade Full shade Ambient Warmed
Leaf d13C �33.5 � 0.1 �33.8 � 0.2 �33.7 � 0.2 �33.4 � 0.1a �33.9 � 0.2b
Leaf %C 41.0 � 0.6 41.9 � 0.2 41.6 � 0.4 41.4 � 0.4 41.5 � 0.3
Soil d13C �25.5 � 0.1 �25.6 � 0.1 �25.5 � 0.1 �25.6 � 0.1 �25.5 � 0.1
Soil %C 4.6 � 0.1 4.4 � 0.1 4.5 � 0.1 4.4 � 0.0 4.5 � 0.1
The mean C content and d13C ratios are shown as well as �1 SE (n 5 4). Within a d13C ratio or C content measurement, warming
treatments labelled with different letters differ significantly.
Soi
l res
pira
tion
rate
(µg
C g
dw
−1 h
−1)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.204°C 8°C 12°C
Half shade Full shadeUnshadedAmbient Warmed Ambient Warmed Ambient Warmed
Fig. 7 The respiration rate measured in the sieved samples
from the different warming and shading treatments incubated
at the three different temperatures. The rates represent the mean
of the latter three measurement weeks, i.e. rates after the initial
pulse of respiration had declined. Error bars represent 11 SE
(n 5 3 unshaded, n 5 4 half and full shade).
1768 I . P. H A R T L E Y et al.
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770

However, sieving substantially reduced the differences
between warming and shading treatments. These
results suggest that different substrate availabilities
caused the differences observed in the intact core
respiration rates. Therefore, this response to warming,
mediated through changes in substrate availability and
not through active adaptation of the microbial commu-
nity, should not be referred to as ‘acclimation’. The
results presented here were also consistent with re-
sponses being largely due to changes in the substrate
availability in the recently fixed, labile SOM pool and
not due to changes in CO2 production associated with
the decomposition of the recalcitrant soil C pool. While
studies have demonstrated that microbial community
structure may be altered by soil warming, no study as
yet appears to have formally demonstrated thermal
acclimation of heterotrophic respiration. It must, there-
fore, be assumed that temperature does indeed increase
heterotrophic activity and that the main limiting factor
for a sustained response is the amount of substrate
available for decomposition. The magnitude of C losses
induced by warming a particular soil may be depen-
dent on the organic content and the chemical and
physical properties of the soil which regulate substrate
availability. Understanding how different soil types will
respond to warming remains of key importance in the
modelling of global C-cycle feedbacks.
Acknowledgements
We are extremely grateful for the opportunity to sample the soilsfrom the long-term experiment which was set up and run by E. J.Edwards and A. H. Fitter. During the main experimental periodthe plots were maintained by C. P. Lancaster and P. Scott. Thetemperature control system was designed and built by D. Ben-ham. The members of the Ineson, Fitter, Atkin and Hodgelaboratory groups helped during harvesting of the plots. I. P.H.’s PhD Studentship was funded by NERC and Forest Researchthrough the UK Centre for Terrestrial Carbon Dynamics (CTCD).
References
Andrews JA, Matamala R, Westover KM, Schlesinger WH (2000)
Temperature effects on the diversity of soil heterotrophs and
the d13C of soil-respired CO2. Soil Biology and Biochemistry,
32, 699–706.
Bellamy PH, Loveland PJ, Bradley RI, Lark RM, Kirk GJD (2005)
Carbon losses from soils across England and Wales 1978–2003.
Nature, 437, 245–248.
Bond-Lamberty B, Wang C, Gower ST (2004) A global relation-
ship between the heterotrophic and autotrophic components
of soil respiration? Global Change Biology, 10, 1756–1766.
Bowden RD, Newkirk KM, Rullo GM (1998) Carbon dioxide and
methane fluxes by a forest soil under laboratory-controlled
moisture and temperature conditions. Soil Biology and Biochem-
istry, 30, 1591–1597.
Cardon ZG, Hungate BA, Cambardella CA, Chapin III FS, Field
CB, Holland EA, Mooney HA (2001) Contrasting effects of
elevated CO2 on old and new soil carbon pools. Soil Biology and
Biochemistry, 33, 365–373.
Christensen BT (2001) Physical fractionation of soil and structural
and functional complexity in organic matter turnover. Eur-
opean Journal of Soil Science, 52, 345–353.
Cox PM, Betts RA, Jones CD, Spall SA, Totterdell IJ (2000)
Acceleration of global warming due to carbon-cycle feedbacks
in a coupled climate model. Nature, 408, 184–187.
Craine JM, Wedin DA, Chapin FS (1999) Predominance of
ecophysiological controls on soil CO2 flux in a Minnesota
grassland. Plant and Soil, 207, 77–86.
Davidson EA, Belk E, Boone RD (1998) Soil water content and
temperature as independent and confounded factors control-
ling soil respiration in a temperate mixed hardwood forest.
Global Change Biology, 4, 217–227.
Edwards EJ, Benham DG, Marland LA, Fitter AH (2004)
Root production is determined by radiation flux in a
temperate grassland community. Global Change Biology, 10,
209–227.
Eliasson PE, McMurtrie RE, Pepper DA, Stromgren M, Linder S,
Agren GI (2005) The response of heterotrophic CO2 flux to soil
warming. Global Change Biology, 11, 167–181.
Fang C, Moncrieff JB (2001) The dependence of soil CO2 efflux
on temperature. Soil Biology and Biochemistry, 33, 155–165.
Fang C, Smith P, Moncrieff JB, Smith JU (2005) Similar response
of labile and resistant soil organic matter pools to changes in
temperature. Nature, 433, 57–59.
Fitter AH, Graves JD, Self GK, Brown TK, Bogie DS, Taylor K
(1998) Root production, turnover and respiration under two
grassland types along an altitudinal gradient: influence of
temperature and soil radiation. Oecologia, 114, 20–30.
Franzluebbers AJ (1999) Potential C and N mineralization and
microbial biomass from increasingly disturbed soils of various
textures. Soil Biology and Biochemistry, 31, 1083–1090.
Hanson PJ, Edwards NT, Garten CT, Andrews JA (2000) Separ-
ating root and soil microbial contributions to soil respiration:
a review of methods and observations. Biogeochemistry, 48,
115–146.
Hartley IP, Armstrong AF, Murthy R, Barron-Gafford G, Ineson
P, Atkin OK (2006) The dependence of respiration on photo-
synthetic substrate supply and temperature: integrating leaf,
soil and ecosystem measurements. Global Change Biology, 12,
1954–1968.
Heath J, Ayres E, Possel M et al. (2005) Rising atmospheric CO2
reduces sequestration of root-derived soil carbon. Science,
309, 1711–1713.
Heinemeyer A, Ridgway KP, Edwards EJ, Benham DG, Young
JPW, Fitter AH (2004) Impact of soil warming and shading on
colonization and community structure of arbuscular mycor-
rhizal fungi in roots of a native grassland community. Global
Change Biology, 10, 52–64.
Hogberg P, Nordgren A, Buchmann N et al. (2001) Large-scale
forest girdling shows that current photosynthesis drives soil
respiration. Nature, 411, 789–792.
WA R M I N G E F F E C T S O N S O I L R E S P I R AT I O N 1769
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770

Houghton RA, Davidson EA, Woodwell GM (1998) Missing
sinks, feedbacks, and understanding the role of terrestrial
ecosystems in the global carbon balance. Global Biogeochemical
Cycles, 12, 25–34.
Huang X, Lakso AN, Eissenstat DM (2005) Interactive effects of
soil temperature and moisture on Concord grape root respira-
tion. Journal of Experimental Botany, 56, 2651–2660.
Ineson P, Benham DG, Poskitt J, Harrison AF, Taylor K, Woods C
(1998) Effects of climate change on nitrogen dynamics in
upland soils. 2. A soil warming study. Global Change Biology,
4, 153–161.
Intergovernmental Panel on Climate Change (2001) Climate
Change 2001: The Scientific Basis. Cambridge University Press,
Cambridge, UK.
Janssens IA, Lankreijer H, Matteucci G et al. (2001) Productivity
overshadows temperature in determining soil and ecosystem
respiration across European forests. Global Change Biology, 7,
269–278.
Kirschbaum MUF (2004) Soil respiration under prolonged soil
warming: are rate reductions caused by acclimation or sub-
strate loss? Global Change Biology, 10, 1870–1877.
Knorr W, Prentice IC, House JI, Holland EA (2005) Long-term
sensitivity of soil carbon turnover to warming. Nature, 433,
298–301.
Langley JA, Johnson NC, Koch GW (2005) Mycorrhizal status
influences the rate but not the temperature sensitivity of soil
respiration. Plant and Soil, 277, 335–344.
Lloyd J, Taylor JA (1994) On the temperature dependence of soil
respiration. Functional Ecology, 8, 315–323.
Luo Y, Wan S, Hui D, Wallace LL (2001) Acclimatization of soil
respiration to warming in a tall grass prairie. Nature, 413,
622–625.
Pennanen T, Caul S, Daniell TJ, Griffiths BS, Ritz K,
Wheatley RE (2004) Community-level responses of metaboli-
cally active soil microorganisms to the quantity and quality
of substrate inputs. Soil Biology and Biochemistry, 36,
841–848.
Raich JW, Schlesinger WH (1992) The global carbon dioxide flux
in soil respiration and its relationship to vegetation and
climate. Tellus B, 44, 81–99.
Reichstein M, Subke J-A, Angeli AC, Tenhunen JD (2005) Does
the temperature sensitivity of decomposition of soil organic
matter depend upon water content, soil horizon, or incubation
time? Global Change Biology, 11, 1754–1767.
Rodeghiero M, Cescatti A (2005) Main determinants of forest soil
respiration along an elevation/temperature gradient in the
Italian Alps. Global Change Biology, 11, 1024–1041.
Roslev P, Iversen N, Henriksen K (1997) Oxidation and assimila-
tion of atmospheric methane by soil methane oxidisers.
Applied and Environmental Microbiology, 63, 874–880.
Rustad LE, Campbell JL, Marion GM et al. (2001) A meta-analysis
of the response of soil respiration, net mineralization, and
aboveground plant growth to experimental ecosystem warm-
ing. Oecologia, 126, 543–562.
Smedley MP, Dawson TE, Comstock JP, Donovan LA, Sherrill DE,
Cook CS, Ehleringer JR (1991) Seasonal carbon isotope discri-
mination in a grassland community. Oecologia, 85, 314–320.
Trueman RJ, Gonzalez-Meler MA (2005) Accelerated below-
ground C cycling in a managed agriforest ecosystem exposed
to elevated carbon dioxide concentrations. Global Change
Biology, 11, 1258–1271.
Trumbore S (2006) Carbon respired by terrestrial ecosystems –
recent progress and challenges. Global Change Biology, 12, 141–
153.
Valentini R, Matteucci G, Dolman AJ et al. (2000) Respiration as
the main determinant of carbon balance in European forests.
Nature, 404, 861–864.
Zhang W, Parker KM, Luo Y, Wan S, Wallace LL, Hu S (2005) Soil
microbial responses to experimental warming and clipping in
a tallgrass prairie. Global Change Biology, 11, 266–277.
Zogg GP, Zak DR, Ringelberg DB, MacDonald NW, Pregitzer KS,
White DC (1997) Compositional and functional shifts in micro-
bial communities due to soil warming. Soil Science Society of
America Journal, 61, 475–481.
1770 I . P. H A R T L E Y et al.
r 2007 The AuthorsJournal compilation r 2007 Blackwell Publishing Ltd, Global Change Biology, 13, 1761–1770




















