
Effects of sex steroids on cell growth and C-myc oncogene expression in LN-CaP and DU145 prostatic...
14
International Urology and Nephrology 27 (1). pp. 67-80 (1995) Effects of Sex Steroids on Cell Growth and C-myc Oncogene Expression in LN-CaP and DU-145 Prostatic Carcinoma Cell Lines* F. K. ASADI, R. SHARIFI Department of Urology, University of Illinois Medical Center at Chicago, Chicago, Illinois (Accepted September 6, 1994) The prostate gland is dependent on androgens for the maintenance of its nor- mal growth and functional integrity. Initially, growth of the majority of prostate tu- mours can be manipulated by endocrine therapy. In the present study, we evaluated the effects of sex steroids on the cell growth and expression of the C-myc oncogene in two human prostatic adenocarcinoma cell lines. We found that dihydrotestos- terone increases the proliferation rate of prostatic cells, and amplification of C-myc oncogene is hormone-dependent. We also demonstrated a positive correlation be- tween the number of cells positive for C-myc oncogene and oncoprotein in hor- mone-treated prostate cell lines. Introduction Carcinoma of the prostate is the most common non-skin cancer in men, and the second leading cause of male cancer death in the United States [1]. The prostate gland is dependent on androgens for the maintenance of its normal growth and functional integrity. In addition, prostatic tumours are initially an- drogen responsive, and decrease in size in response to androgen ablation or anti-androgen therapy. The extent and duration of the response are variable, and ultimately, prostate cancer reverts to an androgen-independent state. The molecular mechanism of this loss of growth control by androgens is poorly un- derstood [2]. Oncogenic products have been demonstrated to have sequence homologies similar to proteins that are important mediators of hormone action on target cells. It is therefore likely that growth factors within the prostate may be related to oncogene products. Both human and animal systems have suggest- ed a role for oncogenes in prostatic carcinogenesis [3-12]. Elevated expression of the C-myc oncogene is often associated with cell proliferation, and the p62 C-myc oncogene product has been correlated closely with tumour grade [13]. Expression of the C-myc oncogene generally appears higher in prostatic cancer tissue than in benign hyperplasia [14]. While Rijnders and colleagues [11] dem- onstrated that the expression of C-myc oncogenes was independent of hor- * Supported by Internal Grant from Section of Urology at West Side Veterans Administration Hospital. VSP. Utrecht Akademiai Kiad6. Budapest
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Effects of sex steroids on cell growth and C-myc oncogene expression in LN-CaP and DU145 prostatic...

International Urology and Nephrology 27 (1). pp. 67-80 (1995)
Effects of Sex Steroids on Cell Growth and C-myc Oncogene Expression in LN-CaP and DU-145 Prostatic
Carcinoma Cell Lines*
F. K. ASADI, R. SHARIFI
Department of Urology, University of Illinois Medical Center at Chicago, Chicago, Illinois
(Accepted September 6, 1994)
The prostate gland is dependent on androgens for the maintenance of its normal growth and functional integrity. Initially, growth of the majority of prostate tumours can be manipulated by endocrine therapy. In the present study, we evaluated the effects of sex steroids on the cell growth and expression of the C-myc oncogene in two human prostatic adenocarcinoma cell lines. We found that dihydrotestosterone increases the proliferation rate of prostatic cells, and amplification of C-myc oncogene is hormone-dependent. We also demonstrated a positive correlation between the number of cells positive for C-myc oncogene and oncoprotein in hormone-treated prostate cell lines.
Introduction
Carcinoma of the prostate is the most common non-skin cancer in men, and the second leading cause of male cancer death in the United States [1]. The prostate gland is dependent on androgens for the maintenance of its normal growth and functional integrity. In addition, prostatic tumours are initially androgen responsive, and decrease in size in response to androgen ablation or anti-androgen therapy. The extent and duration of the response are variable, and ultimately, prostate cancer reverts to an androgen-independent state. The molecular mechanism of this loss of growth control by androgens is poorly understood [2]. Oncogenic products have been demonstrated to have sequence homologies similar to proteins that are important mediators of hormone action on target cells. It is therefore likely that growth factors within the prostate may be related to oncogene products. Both human and animal systems have suggested a role for oncogenes in prostatic carcinogenesis [3-12]. Elevated expression of the C-myc oncogene is often associated with cell proliferation, and the p62 C-myc oncogene product has been correlated closely with tumour grade [13]. Expression of the C-myc oncogene generally appears higher in prostatic cancer tissue than in benign hyperplasia [14]. While Rijnders and colleagues [11] demonstrated that the expression of C-myc oncogenes was independent of hor-
* Supported by Internal Grant from Section of Urology at West Side Veterans Administration Hospital.
VSP. Utrecht Akademiai Kiad6. Budapest

68 Asadi, Shariji: Sex-steroid modulation
monal treatment, a recent paper indicates that oestradiol produces an increase' in the level ofC-myc mRNA [15]. Griffiths and associates [16] have found transcripts of C-myc and p53 in benign adenoma and/or carcinoma tissue. By use of a recombinant retroviral vector, the C-myc oncogene is recently introduced singly or in combination with the ras oncogene into the foetal urogenital sinus, resulting in distinct phenotypes of prostatic pathology [17]. These observations prompted us to explore whether expression of C-myc oncogene was under hormonal control. A combination of immunohistochemistry, in situ hybridization, and Southern hybridization has been used to study the expression of the C-myc oncogene within two human prostatic cell lines. We found that C-myc oncogene and oncoprotein are present in LN-CaP and DU-145 prostatic carcinoma cell lines and in both prostatic carcinomatous tissues. With regard to hormonal effects, we showed that dihydrotestosterone (DHT) and estrogen increase expression of C-myc oncogene and oncoprotein; however, DHT appears to be more effective in eliciting C-myc expression.
Materials and methods
Cell culture
Two human adenocarcinoma cell lines, DU-145 and LN-CaP were obtained from the American Type Culture Collection (Rockville, MD). Both cell lines were cultured at 37°C in Roswell Park Memorial Institute (RPMI)-1640 medium, with added glutamine, supplemented with 10% (V N) dextran-coated charcoal stripped foetal bovine serum (FBS). The control cells received no hormones, but were maintained in RPMI-1640 supplemented with 10% stripped serum. The method used for the preparation of stripped serum was a modified procedure originally described by Drovin [18]. The treated cells in a period of one week received either 10-3 5a.-dihydrotestosterone or 200 pg/ml 1713-estradiol purchased from Sigma Chemical Co. (St. Louis, MO, 63178). The control cells for hormone experiments received also 0.2% solution of hormone solvent mixture (ethanol: propylene glycol 9: 1).
In vitro proliferation assay
A suspension of 1 x 103 cells/ml was seeded in tissue culture 6-well plates and incubated overnight in a CO2 incubator. The following day, 100 III of 5-bromo-2' -deoxyuridine (Amersharn Corp., Arlington Heights, IL) was added to each well. After one-hour incubation, cells were gently washed with phosphate buffer saline (PBS), fixed in 5% acetic acid in ethanol, washed in 0.1% Tween 20 in PBS, and incubated with 50 III of anti-5-bromo-2' -deoxyuridine monoclonal antibody for 30 minutes. This was followed by incubating with the secondary peroxidase rabbit anti-mouse antibody for 25 minutes at room temperature. Slides were then treated with the peroxidase substrate solution (O.l% DAB in Tris-HCl, pH 7.2, containing 0.005% hydrogen peroxide) for
International Urology and Nephrology 27, 1995

Asadi, Sharifi: Sex-steroid modulation 69
5 minutes. Five slides of each specimen were selected at random and counted. The percentage of positive cells was determined by counting a total of 250 and positive cells tallied.
In situ hybridization
Slides of cell cultures were immersed in 3% peroxide for five minutes, followed by rinsing in deionized distilled water. Slides were treated with 1 mM lavamisole (Sigma Chemical Co., St. Louis, MO) and rehydrated by sequential double washes in 95%, 80%, 45%, and 0% ethanol in water. To each slide 100 III of hybridization mixture was added. Hybridization mixtures were made from 50 III deionized formamide (Fisher Scientific, Fair Lawn, NJ), 10 III of 50% dextran sulfate (Sigma Chemical Co., St. Louis, MO), 10 11120 X SSPE (3.6 M sodium chloride, 0.2 M sodium phosphate, pH 7.4, 0.02 M EDTA, pH 7.4), 20 III of 5 pg/1l1 stock DNA probe (ENZO Biochem, New York, NY), and 10 III deionized water. For denaturation, slides were placed in a 14 cm glass Petri dish, and incubated in 0.2 N HCl solution at room temperature for 20 minutes. After denaturation, slides were washed in 0.05% Triton X-I00 and immersed in 50% formamide in 0.1 X SSC (1 M NaCl, 1 mM Na3 citrate - H20, pH 7.0) at 37°C for 10 minutes. After a formaldehyde bath, slides were washed in 0.05% Triton X-lOO, then 200 III of streptavidin and biotinylated horseradish peroxidase detection complex (ENZO Biochem, New York, NY) were added and incubation continued at room temperature for 20 minutes. Finally, slides were rinsed in wash buffer (1 X PBX, 5 mM EDTA), 400 III of substrate mixture (25 III of 1% H20 2, 20 III of aminoethylcarbazole, 1 1110.5 M sodium acetate, pH 4.9) was added to the specimen and incubated at room temperature for 10-15 minutes. The slides were then rinsed with wash buffer and cover slips were mounted on the slide and viewed by light microscopy at x400 magnification.
Southern experiment
Total DNA was purified and analyzed by the Southern procedure [19]. Digestion of DNA by restriction endonuclease was accomplished as recommended by the supplier (Boehringer Mannheim, Indianapolis, IN). DNA fragments were fractionated on 0.8% agarose gels, transferred to nylon membrane, hybridized to the nick-translated C-myc probe, and visualized by autoradiography. The blots were hybridized and washed at 42°C in 0.3 M NaCl.
Statistical analysis
Analysis of variance employing the BMDP Statistical package was used to analyze the cell growth data (available on the main frames of UIC computer). The program BMDP8V was applied to differentiate between fixed and random factors.
International Urology and Nephrology 27, 1995
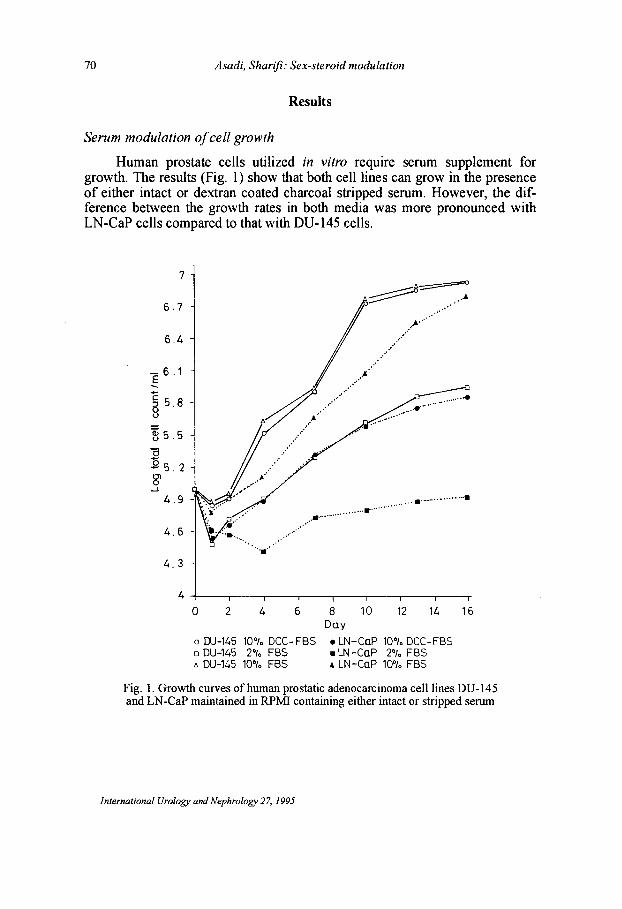
70 Asadi, Sharifi: Sex-steroid modulation
Results
Serum modulation of cell growth
Human prostate cells utilized in vitro require serum supplement for growth. The results (Fig. 1) show that both cell lines can grow in the presence of either intact or dextran coated charcoal stripped serum. However, the difference between the growth rates in both media was more pronounced with LN-CaP cells compared to that with DU-145 cells.
7
6.7
6.4
c 5 5.8 u
~ 5.5 -
a :§5.2-8'
...J
4.9
~ .....•.....•.. /./ •... t,··~
.' .. '
..... ......
.... ~ .
.' .'
.. ' .... '
",., .
.......•...••...........• .. ' ...............................
4.6 o ...•..
4.3 -
o 2 4 6
o DU-145 10"10 DCC- FBS o DU-145 2% F8S /l DU-145 10"10 FBS
8 10 12 14 Day
• LN-CaP 10°/. DCC-FBS • LN-CaP 2% F8S .4 LN-CaP 10% FBS
16
Fig. 1. Growth curves of human prostatic adenocarcinoma cell lines DU-145 and LN-CaP maintained in RPM! containing either intact or stripped serum
International Urology and Nephrology 27, 1995
C ::J 8
~
Asadi. Sharifi: Sex-steroid modulation 71
5.96 -
5.88
5.80
5.72
564 -
5.56 -
5.48 -
5.40
o DU-145 Control • DU-145 10-3M DHT t:. r:x.H45 10-4M DHT
Day
iii DU-145 10-5M DHT e DU-145 10-6M DHT 8l DU-145 10 - 7M DHT
Fig. 2
575
6-
5.9
58 -
5 7 -
5.6 -
5.5
54
5.3
5 2-
Day
o LN - Co P Control + LN-CoP 10-3MDHT x LN - CaP 10-4M DHT
",+
.. LN-CaP 1O-5M DHT <I) LN-CaP 10-6M DHT I3J LN-CaP 10-7M DHT
Fig. 3
5·50
545
5.40
.f:.
§ 550-8
5.35 -
530
~ 525-c 12 .§'525-
5.20 -
5 -r-I ---'--'--' -----r, 3 5 7
Day o DU-145 Control .. DU-145 100 pg Iml EST + DU-145 40 pg/ml EST • DU-145 150 pg/ml EST x DU-145 70 pg/ml EST ~ DU-145 200 pg/ml EST
Fig. 4
5·15 -
5. 1 0 ._ •. ::..:<;~F·:·· cr."
5.05 f'f
5rl----~I----~I----~, 1 3 5 7
Day o LN-CaP Control s LN -CaP 100 pg/ml EST o LN-CoP 40 pg/ml EST eLN-CaP 250 pg/ml EST f:. LN-CaP 70 pg/ml EST 8 LN-CoP 200 pg/ml EST
Fig. 5
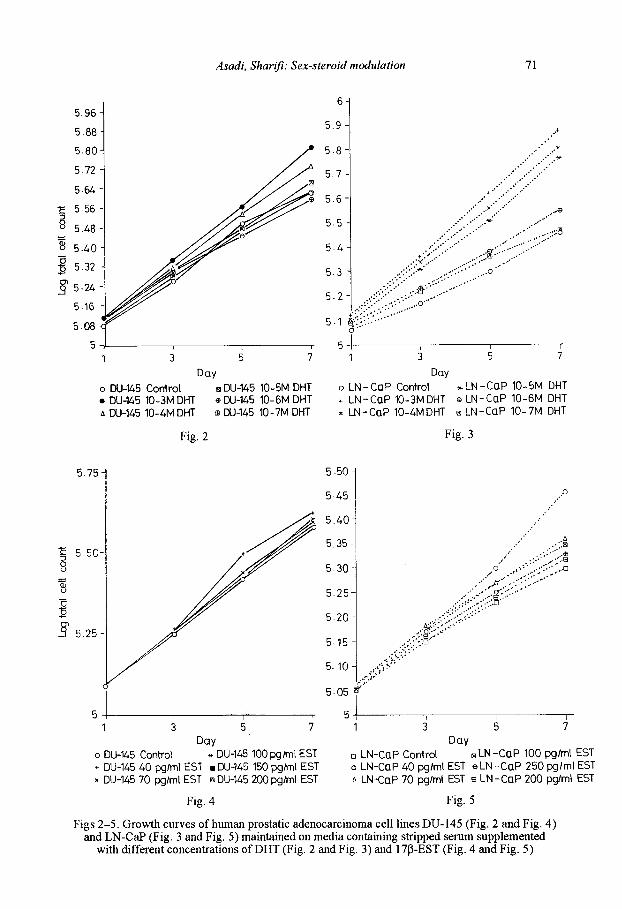
Figs 2-5. Growth curves of human prostatic adenocarcinoma cell lines DU-145 (Fig. 2 and Fig. 4) and LN-CaP (Fig. 3 and Fig. 5) maintained on media containing stripped serum supplemented
with different concentrations ofDHT (Fig. 2 and Fig. 3) and l7P-EST (Fig. 4 and Fig. 5)

72 Asadi, Sharifi: Sex-steroid modulation
Steroid modulation of cell growth
The growth rate ofOU-145 and LN-CaP cells treated with different concentrations ofDHT (10-3 M -10-7 M) and 17 M-EST (40 pg/ml to 200 pg/ml) are shown in Figs 2-5. Concentrations of 10-3 M and 10-4 M of DHT had the most stimulatory effect on the proliferation of DU-145 cells. LN-CaP cells showed the highest growth rate when treated with the highest concentration of DHT (10-3 M). On the other hand, there was no evident effect of EST concentrations on the growth ofOU-145 and LN-CaP cells and the log total cell counts were very close to that of the control cells.
Proliforation assay
Proliferation assay showed that the percentages of proliferating cells on day 7 were comparable between the two cell lines when untreated (18-20%) or increased by treatment with OHT (26-27%). The increase in per cent proliferation was statistically significant with DHT treatment for both cell lines (Table 1). Estrogen treatment, however, differentially decreased the percentage of proliferating cells (Table 1), as well as the total number of cells (data are not shown). Both the decrease from the control values as well as the difference between cell lines were statistically significant (P<O.OOOI).
Table 1 Percentage of cells positive for proliferations
DU-145
LN-CaP
Untreated controls
20.5± 1.4
l7.7±1.2
Number of proliferating cells (±SEM) (N=3)
Testosterone treated
26.3±1.3
26.6±1.3
Estradiol treated
13.7±0.4
6.9±O.9
Two-way analysis of variance establishes that there are significant differences between cell lines, among treatments, as well as in intergroup effect. A Duncan multiple range test shows significant (95% confidence) differences between all groups except for the two groups within each set of continuously underlined groups.
Pattern of oncogene and oncoprotein staining
In situ DNA-DNA hybridization revealed that both cell lines showed amplified C-myc oncogene (Fig. 6). Double immuno-enzyme staining of prostate cell lines for either oncoprotein and proliferation or oncoprotein and oncogene suggests that C-myc expression is related in some way to the cellular prolifera-
International Urology and Nephrology 27, 1995
Asadi. Sharifi: Sex-steroid modulation
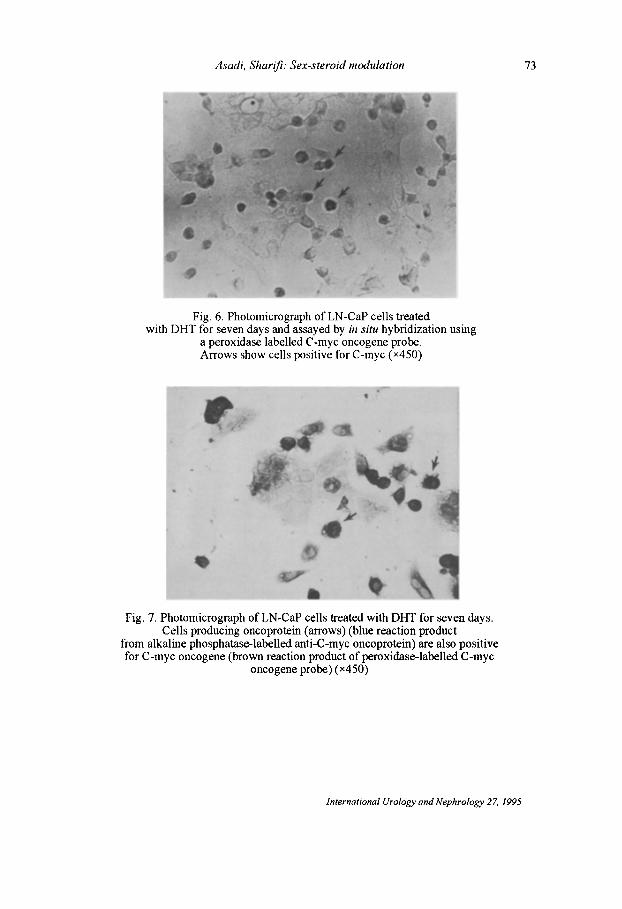
Fig. 6. Photomicrograph ofLN-CaP cells treated with DHT for seven days and assayed by in situ hybridization using
a peroxidase labelled C -myc oncogene probe. Arrows show cells positive for C-myc (x450)
1 • Fig. 7. Photomicrograph ofLN-CaP cells treated with DHT for seven days.
Cells producing oncoprotein (arrows) (blue reaction product from alkaline phosphatase-labelled anti-C-myc oncoprotein) are also positive for C-myc oncogene (brown reaction product of peroxidase-labelled C-myc
oncogene probe) (x450)
International Urology and Nephrology 27. 1995
73
74 Asadi, Sharifi: Sex-steroid modulation
tion of prostate cells (Fig. 7). We showed that DHT and EST increased the percentage of positive LN-CaP cells for C-myc oncogene from 19.3±O.2 'to 39.2±1.0 and to 26.02±2.0, respectively, and for DU-145 cells from 15.6±1.7 to 29.O±O.7 and to 21.5±1.9, respectively (Table 2). On the other hand, DHT increased the percentage of cells expressing C-myc oncoprotein from 6.2±O.9 to 14.6±O.7 for DU-145 and from 9.6±10 to 12.4±1.11 for LN-CaP (Table 3). Estradiol decreases the percentage of cells expressing C-myc oncoprotein from 9.6±1.1 to 6.1±1.1 for LN-CaP cells (Table 3). The results of the regression
Table 2 Percentage of cells positive for C-myc oncogene.
N=3 for control groups, N=6 for testosterone (DHT) and estrogen (EST) treatment groups
Control DHT EST
LN-CaP 19.3±O.2 30.2±l.O 26.2±2.0
DU-145 15.6±1.7 29.0±O.7 2 l.5± 1.9
DHT and EST cause an increase in C-myc oncogene expression in both cell types. The only difference between the two cell types is that in the DU-145 cells EST was less effective in increasing C-myc oncogene expression.
Table 3 Percentage of cells positive for C-myc protein.
N=3 for control groups, N=6 for testosterone (DHT) and estrogen (EST) treatment groups
Control DHT EST
LN-CaP 9.6±1.0 12.4 ±l.l 6.1±1.l
DU-145 6.2±O.9 14.65±O.7 6.8±0.4
ANOV A shows a significant hormone treatment effect with a significant interaction with cell type. A subsequent Duncan multiple range test shows that DHT causes an increase in DU-145 cells and EST causes a decrease in LN-CaP cells.
analysis done on the oncogene/oncoprotein data showed a negative correlation between cells positive for oncogene and oncoprotein in DU-145 and LN-CaP control cells. However, DHT and EST treated DU-145 cells and DHT treated LN -CaP cells showed a positive linear correlation between presence of oncogene and oncoprotein. Estradiol treated LN-CaP cells did not show any such correlation (Fig. 8a, b).
International Urology and Nephrology 27, 1995

c ~ 0 .... 0. 0 u c
0
c
~ a. 0 u c: 0
Asadi. Sharifi: Sex-steroid modulation
150
138 -
126 -
114 -
102 -
90 -
78 -
66
54 • .......... -...... .. --42 ....... >41-j,-" 30 • I
75 95 115 135 155 175 195
140
127
114 -
Oncogene • DU-145 Control • DU-145 DHT & DU-145 EST
(a)
I /'
• .r / /
•
215 235 255 275
• 101 /" 88 -
75 -
62 -
49 '-36 -
• & 23- ./ /.
• & 10 I
60 80 100 120 140
•
I
160 180 200 220 2L.O 260 Oncogene
• LN - Co P Control • LN-CoP DHT & LN -Co'p EST
(b)
Fig. 8. Relationship between cells positive for the presence of C-myc oncogene and oncoprotein in DU-145 (a) and LN-CaP (b) cells treated with either 10-3 M DHT
or 200 pg/ml EST for seven days
75
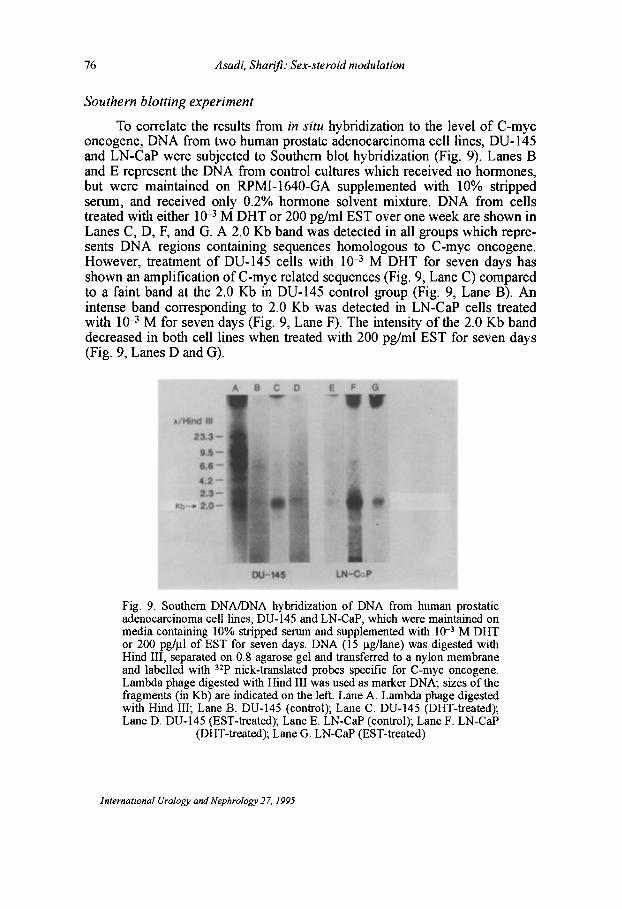
76 Asadi. Sharifi: Sex-steroid modulation
Southern blotting experiment
To correlate the results from in situ hybridization to the level of C-myc oncogene, DNA from two human prostate adenocarcinoma cell lines, DU-145 and LN-CaP were subjected to Southern blot hybridization (Fig. 9). Lanes B and E represent the DNA from control cultures which received no hormones, but were maintained on RPMI-1640-GA supplemented with 10% stripped serum, and received only 0.2% hormone solvent mixture. DNA from cells treated with either 10--3 M DHT or 200 pg/ml EST over one week are shown in Lanes C, D, F, and G. A 2.0 Kb band was detected in all groups which represents DNA regions containing sequences homologous to C-myc oncogene. However, treatment of DU-145 cells with 10-3 M DHT for seven days has shown an amplification ofC-myc related sequences (Fig. 9, Lane C) compared to a faint band at the 2.0 Kb in DU-145 control group (Fig. 9, Lane B). An intense band corresponding to 2.0 Kb was detected in LN-CaP cells treated with 10--3 M for seven days (Fig. 9, Lane F). The intensity of the 2.0 Kb band decreased in both cell lines when treated with 200 pg/ml EST for seven days (Fig. 9, Lanes D and G).
Fig. 9. Southern DNAIDNA hybridization of DNA from human prostatic adenocarcinoma cell lines, DU-145 and LN-CaP, which were maintained on media containing 10% stripped serum and supplemented with 10-3 M DHT or 200 pg/l.L1 of EST for seven days. DNA (15 Ilg/lane) was digested with Hind III, separated on 0.8 agarose gel and transferred to a nylon membrane and labelled with 32p nick-translated probes specific for C-myc oncogene. Lambda phage digested with Hind III was used as marker DNA; sizes of the fragments (in Kb) are indicated on the left. Lane A. Lambda phage digested with Hind III; Lane B. DU-145 (control); Lane C. DU-145 (DHT-treated); Lane D. DU-145 (EST-treated); Lane E. LN-CaP (control); Lane F. LN-CaP
(DHT-treated); Lane G. LN-CaP (EST-treated)
international Urology and Nephrology 27. 1995

Asadi, Sharifi: Sex-steroid modulation 77
Discussion
The prostate gland requires a permanent androgen stimulus for its differentiation and function. Prostatic tumour cells, however, can overcome androgen dependence and escape hormone ablation therapy. The C-myc oncogene is a highly conserved DNA sequence and is a cellular homologue of C-myc viral oncogenes. It encodes a nuclear protein that has been linked with cell proliferation and DNA synthesis. Expression of C-myc has been reported in prostatic carcinoma cell lines such as PC3, PC133, and PC 135 [11]. They have been reported in hyperplastic and carcinomatous prostatic tissue as well [14]. Hierowski et al. [20] have reported the potential dependence of human prostatic growth factor on stimulating thymidine incorporation into DNA. They have shown that stimulation was significantly enhanced by DHT and EST in the androgen sensitive LN-CaP cell lines, which express high level of androgen receptor [21]. We have used a fast growing colony from this LN-CaP cell line, designated LN-CaP-FGC, and have observed that growth of these cells is also stimulated by androgen (DHT), whereas estradiol was suppressive. It appears that the DU-145 cell line is apparently not hormone sensitive since it grows equally well in either 10% intact or stripped serum. The cell line DU-I45 does not express androgen receptor, although the androgen receptor gene is present [22]. The presence of a wide range of concentrations of DHT and estradiol was found to be without significant effect on the growth ofDU-I45 cells. However, these hormones show a significant effect on the growth ofLN-CaP cells. The stimulatory effect of DHT or inhibitory effect of 17P-EST on the mitotic index (PDT) of the LN -CaP cell line might indicate its degree of androgen sensitivity. We obtained similar results as those reported by Horoszewicz et al. [23], indicating that the LN-CaP cell line is responsive to hormonal treatment. We were also able to confirm the results reported by Tilley et al. [22] indicating that the DU-I45 cell line is not dependent on hormones for its survival.
Recent studies indicate that the C-myc gene is expressed throughout the cell cycle. Our data demonstrate the expression of the C-myc oncogene in both human prostatic adenocarcinoma cell lines as well as benign and malignant prostatic tumours. However, the level of expression of the C-myc oncogene is significantly higher in adenocarcinoma of the prostate than in benign prostatic hyperplasia (Figs 10 and II). Although p62 oncoprotein expression correlated with histologic differentiation and thus indirectly with survival, the prognostic value of p62 quantitation in prostate cancer will have to be determined in future studies. Elevated expression of the C-myc oncogene is often associated with cell proliferation. In our study, the expression of C-myc oncoprotein in proliferating cells confirms the relationship between cell proliferation and C-myc expression. Whether oncogene activation occurs at a late stage in prostate cancer or whether it occurs only as an apparently random, infrequent event, awaits the results of further studies.
To assess co-expression of C-myc oncogene and oncoprotein we studied prostate cell lines by in situ hybridization using a peroxidase-labelled C-myc
International Urology and Nephrology 27, 1995
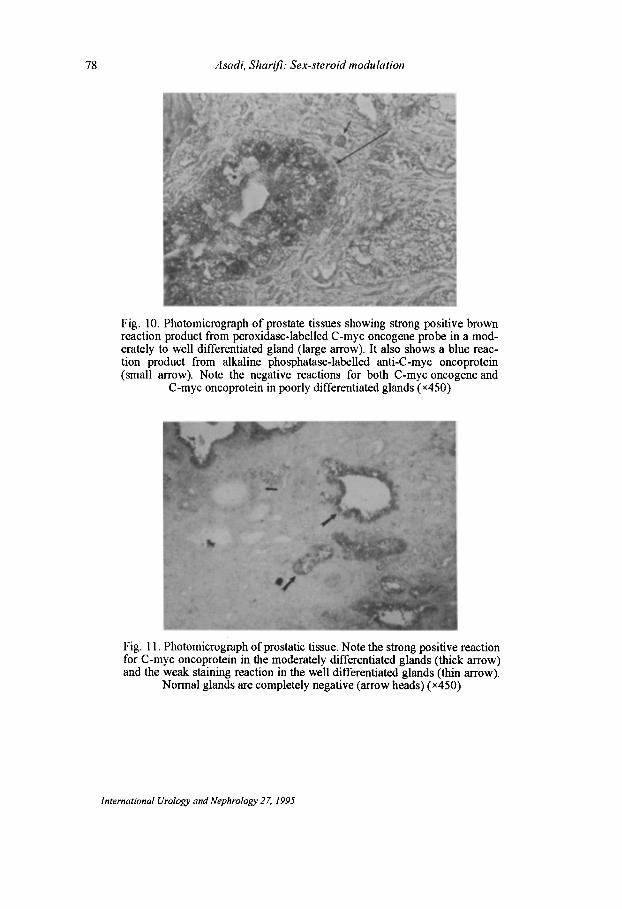
78 Asadi, Sharifi: Sex-steroid modulation
Fig. 10. Photomicrograph of prostate tissues showing strong positive brown reaction product from peroxidase-labelled C-myc oncogene probe in a moderately to well differentiated gland (large arrow). It also shows a blue reaction product from alkaline phosphatase-labelled anti-C-myc oncoprotein (small arrow). Note the negative reactions for both C-myc oncogene and
C-myc oncoprotein in poorly differentiated glands (x4S0)
Fig. 11. Photomicrograph of prostatic tissue. Note the strong positive reaction for C-myc oncoprotein in the moderately differentiated glands (thick arrow) and the weak staining reaction in the well differentiated glands (thin arrow).
Normal glands are completely negative (arrow heads) (x4S0)
International Urology and Nephrology 27, 1995

Asadi, Sharifi: Sex-steroid modulation 79
probe and by immunocytochemical staining with an alkaline phosphatasclabelled, mouse anti-p62 monoclonal antibody, simultaneously. Dihydrotestosterone and estradiol increased the percentage of cells positive for C-myc oncogene in both cell lines. On the other hand, the percentage of cells positive for oncoprotein increased with DHT treatment in DU-145 cells and decreased with EST treatment in LN-CaP cells. Regression analysis showed a negative correlation between the number of cells positive for oncogene and for oncoprotein in DU-145 and LN-CaP control cells. DHT- and EST-treated DU-145 cells and DHT -treated LN-CaP cells, however, showed a positive association between the presence of oncogene and oncoprotein. EST -treated LN -CaP cells did not show any such association. This suggests that translation may be linked to other intracellular controlling factors.
Southern blotting using a 32P-C-myc probe also showed a more intense band of 2.0 Kb corresponding to C-myc sequences in cells treated with steroids when compared with untreated control cells. The intensity of the DNA band from DHT -treated cells, however, was more prominent than the band from EST-treated cells. The difference between the in situ hybridization experiments and the Southern blotting experiments can be reconciled on the basis that in situ hybridization only reflected individual cell differences in a semiquantitative manner.
The data discussed in these studies strongly implicate a link between hormonal treatment and C-myc expression in human prostatic adenocarcinomas. The prognostic significance of elevated levels of C-myc expression in hormone-treated human prostatic adenocarcinoma warrants further investigation. In view of the claims that over-expression of C-myc and other oncogenes has prognostic significance, it is of great interest to identify the type of tumours that express these genes.
Acknowledgements
We are grateful to Drs. Joe Monaligod, Herbert Yamashiroya, and Paul Weiz-Carington for advice and discussion during the course of this work. We also thank B. Branham for help in preparing the manuscript.
References
1. Boring, C. C., Squires, T. S., Tong, T.: Cancer statistics. CA Cancer J. Clin., 42, 19 (1992).
2. Davies, P., Eaton, C. L.: Regulation of prostate growth. J. Endocrinol., 131, 5 (1991). 3. Peehl, D. M., Wehner, N., Stamey, T. A.: Activated Ki-ras oncogene in human prostatic
adenocarcinoma. Prostate, 10, 281 (1987). 4. Konishi, N., Enomoto, T., Buzard, G., Ohshima, M., Ward, J. M., Rice, J. M.: Ki-ras
activation and ras 21 expression in latent prostatic carcinomas in Japanese men. Cancer, 69,2293 (1992).
5. Carter, B. S., Epstein, J. E., Isaacs, W. B.: Ras gene mutations in prostate cancer. Cancer Res., 50, 6830 (1990).
6. Thompson, T. C., Southgate, J., Kitchener, G., Land, H.: Multistage carcinogenesis induced by ras and myc oncogenes in a reconstituted organ. Cell, 56, 917 (1989).
International Urology and Nephrology 27, 1995

80 Asadi. Sharifi: Sex-steroid modulation
7. Sukumar, S., Annstrong, B., Bruyntjes, J. P., Leave, I., Bosland, M. C.: Frequent activation of the Ki-ras oncogene at codon 12 in N-methyl-N-nitrosourea-induced rat·prostate adenocarcinomas and neurogenic sarcomas. Mol. Carcinogen. 4. 362 (1991).
8. Treiger, B., Isaacs, J.: Expression of a transfected v-H-ras oncogene in a Dunning rat prostate adenocarcinoma and the development of high metastatic ability. J. Urol .. 140. 1580 (1988).
9. Voeller, H. J., Wilding, G., Gelman, E. P.: v-H-ras expression confers honnone-independent in vitro growth to LN-CaP prostate carcinoma cells. Mol. Endocrinol .• 5. 209 (1991).
10. Viola, M. V., Fromowitz, F., Oraver, S., Deb, S., Finkel, G., Lundy, J., Hand, P., Thor, A., Schlom, J.: Expression of ras oncogene P21 in prostate cancer. N. Engl. J. Med .• 314. 133 (1986).
11. Rijnders, A., Van der Corput, J., Van Steenbrugge, G., Romign, J., Trapman, J.: Expression of cellular oncogenes in human prostatic carcinoma cell lines. Biochem. Biophys. Res. Comm. (13L) 2, 548 (1985).
12. Gumerlock, P. H., Poonamallee, U. R., Meyers, F. J., White, R. W. V.: Activated ras alleles in human carcinoma of the prostate are rare. Cancer Res .. 51, 1632 (1991).
13. Ellis, D., Leffers, S., Davies, J., Ng, A.: Multiple immunoperoxidase markers in benign hyperplasia and adenocarcinoma of the prostate. Am. J. Clin. Pathol.. 81. 279 (1984).
14. Fleming, W., Hamel, A., MacDonald, R.: Expression of the C-myc proto-oncogene in human prostatic carcinoma and benign prostatic hyperplasia. Cancer Res .• 46. 1535 (1986).
15. Kirland, J. L., Murthy, L., Stancel, G. M.: Progesterone inhibits the estrogen-induced expression of C-fos mRNA in the uterus. Endocrinol. 130. 3223 (1992).
16. Griffiths, J., Davies, P., Harper, M., Peeling, W., Pierrepoint, C.: The etiology and endocrinology of prostatic cancer. In: Rose, D. (ed.): Endocrinology of Cancer. CRC Press, 1979, p. 1.
17. Thompson, T. C., Tadmon, D., Timme, T. L., Merz, V. W., Egawa, S., Krebs, T., Scardino, P. T., Parr, S. H.: Experimental oncogene induced prostate cancer. Cancer Surv .. 11. 55 (1991).
18. Drovin, J.: Estradiol induced increase of the LH responsiveness to LH releasing hormone (LHRH) in rat anterior pituitary cells in culture. Endocrinol.. 99. 1477 (1983).
19. Southern, E.: Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Bioi., 98, 503 (1975).
20. Hierowski, M., McDonald, M., Dunn, L., Sullivan, J.: The partial dependency of human prostatic growth factors on steroid honnones in stimulating thymidine incorporation into DNA. J. Urol.. 138. 909 (1987).
21. Veldscholte, J., Voorhorst-ogink, M. M., Bolt-de Vries, J., Van Rooji, H. C. J., Trapman, J., Mulder, E.: Unusual specificity of the androgen receptor in the human prostate tumor cell line LN-Cap. Biochem. Biophys. Acta. 1052. 187 (1990).
22. Tilley, W. D., Wilson, C. M., Marcelli, M., Mcphaul, M. J.: Androgen receptor gene expression in human prostate carcinoma cell lines. Cancer Res .• 50. 5382 (1990).
23. Horoszewicz, J., Leong, S., Kawinski et aI.: LN-Cap model of human prostatic carcinoma. Cancer Res .. 43. 1809 (1983).
International Urology and Nephrology 27, 1995




















