
Effects of selenium on chromium (VI)-induced hepatotoxicity in adult rats
8
Experimental and Toxicologic Pathology 63 (2011) 541–548 Contents lists available at ScienceDirect Experimental and Toxicologic Pathology jo u rn al h omepage: www.elsevier.de/etp Effects of selenium on chromium (VI)-induced hepatotoxicity in adult rats Nejla Soudani a , Ibtissem Ben Amara a,1 , Mediha Sefi a,1 , Tahia Boudawara b , Najiba Zeghal a,∗ a Animal Physiology Laboratory, UR/08-73 Sfax Faculty of Sciences, BP1171, 3000 Sfax, Tunisia b Anatomopathology Laboratory, Habib Bourguiba Hospital, Sfax, Tunisia a r t i c l e i n f o Article history: Received 28 October 2009 Accepted 14 April 2010 Keywords: Selenium Potassium dichromate Rat Hepatotoxicity Histopathological studies a b s t r a c t Chromium, a major environmental pollutant, is known for its wide toxic manifestations. The present experiment pertains to the protective role of selenium (Se) against K 2 Cr 2 O 7 -induced hepatotoxicity. Female Wistar rats were divided into four groups of six each: group I served as controls which received standard diet; group II received in drinking water K 2 Cr 2 O 7 alone (700 ppm); group III received both K 2 Cr 2 O 7 and Se (0.5 Na 2 SeO 3 mg/kg of diet); group IV received Se (0.5 mg/kg of diet) for 3 weeks. Exposure of rats to chromium promoted oxidative stress with an increase in malondialdehyde (MDA) and a decrease in glutathione (GSH) levels. A decrease in glutathione peroxidase (GPx) and an increase in superoxide dismutase (SOD) and catalase (CAT) activities were observed. Se supplementation to the diet of group III improved all the parameters cited above. Yet, plasma transaminases (AST and ALT), lactate dehydrogenase (LDH) activities, cholesterol, triglycerides (TG) and low density lipoprotein-cholesterol (LDL-C) levels increased, while high density lipoprotein-cholesterol (HDL-C) decreased. Co-administration of Se to the diet of group III restored hepatic markers to near-normal values. The biochemical results confirmed the histopathological findings. Therefore, our investigation revealed that Se was effective in preventing K 2 Cr 2 O 7 -induced hepatotoxicity. © 2010 Elsevier GmbH. All rights reserved. 1. Introduction Chromium (Cr), a naturally occurring heavy metal, exists in many states, but the trivalent [Cr(III)] and hexavalent [Cr(VI)] forms are the most common in the environment (O’Brien et al., 2003). Yet, Cr(VI), a pollutant compound, is frequently found in brownfields, as well as in surface and subterranean waters throughout the industrialized world (Langard, 1990). The wide dis- tribution of this contaminant in the environment is the result of its extensive exploitation to produce stainless steel, wood treat- ment products, tanning of leather or pigments (Barceloux, 1999; Xu et al., 2004). Moreover, occupational exposure to K 2 Cr 2 O 7 , a hexavalent form of Cr(VI), is found among approximately several million industrial workers worldwide (Zhitkovich, 2002; OSHA, 2006). In fact, human beings exposed to K 2 Cr 2 O 7 under environ- mental or occupational settings show increased risk of asthma, nasal septum and skin ulcerations, allergic and contact dermati- tis, as well as respiratory cancers (Salnikow and Zhitkovich, 2008). Indeed, experimental animals exposed to K 2 Cr 2 O 7 exhibit acute tissue damage, including testicular lesions, kidney tubular necro- ∗ Corresponding author at: Animal Physiology Laboratory, Life Sciences Depart- ment, Sfax Faculty of Sciences, BP1171, 3000 Sfax, Tunisia. Tel.: +216 74 274 600; fax: +216 74 274 437. E-mail addresses: [email protected], naj [email protected] (N. Zeghal). 1 These authors contributed equally to this work. sis, and liver toxicity (Bagchi et al., 2002; Pedraza-Chaverri et al., 2005). Due to its role in the transformation of environmen- tal xenobiotics, the liver is at great risk of injury and induced hepatotoxicity as demonstrated by Bagchi et al. (1995a,b) and Bos ¸ gelmez et al. (2008) after the exposure of adult rats to hexava- lent chromium. According to O’Brien et al. (2003), Cr(VI) readily enters cells through the non-specific phosphate/sulfate ion trans- porters. In fact, K 2 Cr 2 O 7 undergoes an intracellular reduction, resulting in the generation of toxic Cr intermediates, which are considered as strong oxidants. ROS generated in excess by these reactions can cause injury to cellular proteins, lipids and DNA, leading to oxidative stress (Nordberg and Arner, 2001; Ueno et al., 2001). Since free radical generation is implicated in the pathogenesis of chromium toxicity, supplementation of antioxidants can be consid- ered as the alternative method for chelation therapy. Selenium (Se) is a rational candidate element for the prevention of liver injury. In fact, Diplock et al. (1986), Xiao et al. (2002) and Ognjanovic et al. (2008) have found that Se has detoxification effects on various heavy metals. Selenium, considered as an essential micronutri- ent for animals and humans (Klasing, 1998; Eisler, 2000), plays an important role in antioxidant defense systems. It is an active site of glutathione peroxidase (GPx). This selenoenzyme protects cells against damages induced by free radicals and also permits the regeneration of a membrane lipid molecule through reacyla- tion (McPherson, 1994). Selenium deficiency is usually associated with increased lipid peroxidation, which alters the integrity of cell 0940-2993/$ – see front matter © 2010 Elsevier GmbH. All rights reserved. doi:10.1016/j.etp.2010.04.005
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Effects of selenium on chromium (VI)-induced hepatotoxicity in adult rats

E
Na
b
a
ARA
KSPRHH
1
mf2ittimXhm2mntIt
mf
0d
Experimental and Toxicologic Pathology 63 (2011) 541– 548
Contents lists available at ScienceDirect
Experimental and Toxicologic Pathology
jo u rn al h omepage: www.elsev ier .de /e tp
ffects of selenium on chromium (VI)-induced hepatotoxicity in adult rats
ejla Soudania, Ibtissem Ben Amaraa,1, Mediha Sefia,1, Tahia Boudawarab, Najiba Zeghala,∗
Animal Physiology Laboratory, UR/08-73 Sfax Faculty of Sciences, BP1171, 3000 Sfax, TunisiaAnatomopathology Laboratory, Habib Bourguiba Hospital, Sfax, Tunisia
r t i c l e i n f o
rticle history:eceived 28 October 2009ccepted 14 April 2010
eywords:eleniumotassium dichromateatepatotoxicity
a b s t r a c t
Chromium, a major environmental pollutant, is known for its wide toxic manifestations. The presentexperiment pertains to the protective role of selenium (Se) against K2Cr2O7-induced hepatotoxicity.Female Wistar rats were divided into four groups of six each: group I served as controls which receivedstandard diet; group II received in drinking water K2Cr2O7 alone (700 ppm); group III received bothK2Cr2O7 and Se (0.5 Na2SeO3 mg/kg of diet); group IV received Se (0.5 mg/kg of diet) for 3 weeks. Exposureof rats to chromium promoted oxidative stress with an increase in malondialdehyde (MDA) and a decreasein glutathione (GSH) levels. A decrease in glutathione peroxidase (GPx) and an increase in superoxidedismutase (SOD) and catalase (CAT) activities were observed. Se supplementation to the diet of group III
istopathological studies improved all the parameters cited above. Yet, plasma transaminases (AST and ALT), lactate dehydrogenase(LDH) activities, cholesterol, triglycerides (TG) and low density lipoprotein-cholesterol (LDL-C) levelsincreased, while high density lipoprotein-cholesterol (HDL-C) decreased. Co-administration of Se to thediet of group III restored hepatic markers to near-normal values. The biochemical results confirmedthe histopathological findings. Therefore, our investigation revealed that Se was effective in preventing
oxici
K2Cr2O7-induced hepatot. Introduction
Chromium (Cr), a naturally occurring heavy metal, exists inany states, but the trivalent [Cr(III)] and hexavalent [Cr(VI)]
orms are the most common in the environment (O’Brien et al.,003). Yet, Cr(VI), a pollutant compound, is frequently found
n brownfields, as well as in surface and subterranean watershroughout the industrialized world (Langard, 1990). The wide dis-ribution of this contaminant in the environment is the result ofts extensive exploitation to produce stainless steel, wood treat-
ent products, tanning of leather or pigments (Barceloux, 1999;u et al., 2004). Moreover, occupational exposure to K2Cr2O7, aexavalent form of Cr(VI), is found among approximately severalillion industrial workers worldwide (Zhitkovich, 2002; OSHA,
006). In fact, human beings exposed to K2Cr2O7 under environ-ental or occupational settings show increased risk of asthma,
asal septum and skin ulcerations, allergic and contact dermati-
is, as well as respiratory cancers (Salnikow and Zhitkovich, 2008).ndeed, experimental animals exposed to K2Cr2O7 exhibit acuteissue damage, including testicular lesions, kidney tubular necro-∗ Corresponding author at: Animal Physiology Laboratory, Life Sciences Depart-ent, Sfax Faculty of Sciences, BP1171, 3000 Sfax, Tunisia. Tel.: +216 74 274 600;
ax: +216 74 274 437.E-mail addresses: [email protected], naj [email protected] (N. Zeghal).
1 These authors contributed equally to this work.
940-2993/$ – see front matter © 2010 Elsevier GmbH. All rights reserved.oi:10.1016/j.etp.2010.04.005
ty.© 2010 Elsevier GmbH. All rights reserved.
sis, and liver toxicity (Bagchi et al., 2002; Pedraza-Chaverri etal., 2005). Due to its role in the transformation of environmen-tal xenobiotics, the liver is at great risk of injury and inducedhepatotoxicity as demonstrated by Bagchi et al. (1995a,b) andBos gelmez et al. (2008) after the exposure of adult rats to hexava-lent chromium. According to O’Brien et al. (2003), Cr(VI) readilyenters cells through the non-specific phosphate/sulfate ion trans-porters. In fact, K2Cr2O7 undergoes an intracellular reduction,resulting in the generation of toxic Cr intermediates, which areconsidered as strong oxidants. ROS generated in excess by thesereactions can cause injury to cellular proteins, lipids and DNA,leading to oxidative stress (Nordberg and Arner, 2001; Ueno et al.,2001).
Since free radical generation is implicated in the pathogenesis ofchromium toxicity, supplementation of antioxidants can be consid-ered as the alternative method for chelation therapy. Selenium (Se)is a rational candidate element for the prevention of liver injury.In fact, Diplock et al. (1986), Xiao et al. (2002) and Ognjanovic etal. (2008) have found that Se has detoxification effects on variousheavy metals. Selenium, considered as an essential micronutri-ent for animals and humans (Klasing, 1998; Eisler, 2000), playsan important role in antioxidant defense systems. It is an activesite of glutathione peroxidase (GPx). This selenoenzyme protects
cells against damages induced by free radicals and also permitsthe regeneration of a membrane lipid molecule through reacyla-tion (McPherson, 1994). Selenium deficiency is usually associatedwith increased lipid peroxidation, which alters the integrity of cell
5 Toxico
m2
pcht
2
2
(fg
2
oucm(NtMA
ta
iwKoi(itdwJdi(ehpio1a
A
l
gBfasa
42 N. Soudani et al. / Experimental and
embranes and, consequently, affects cell functions (Valko et al.,004).
To our knowledge, there are no investigations carried out on therotective effect of selenium against K2Cr2O7-induced hepatotoxi-ity in rats. The present investigation evaluates the antioxidant andepatoprotective potential of Se against K2Cr2O7-induced oxida-ive damage, especially in the liver.
. Materials and methods
.1. Chemicals
Potassium dichromate (K2Cr2O7), was obtained from MerckDarmstadt, Germany). Sodium selenite (Na2SeO3) was purchasedrom Sigma (St. Louis, MO, USA). All other chemicals of analyticalrade were purchased from standard commercial suppliers.
.2. Animals and treatment
Female Wistar rats with initial mean body weight of 130 ± 10 g,btained from the Central Pharmacy (SIPHAT, Tunis, Tunisia), weresed in this study. They were maintained under standard laboratoryonditions (temperature 22 ± 2 ◦C; 12 h light–dark cycle). The ani-als had free access to water and commercial standard pellet diet
SICO, Sfax, Tunisia). The content of Se in standard diet (0.17 mga2SeO3/kg) was determined, after mineralization, by the Elec-
rothermic Atomic Absorption Spectrometry technique (ETAAS).easurements were performed on a PerkinElmer 5100/Zeeman
tomic Absorption Spectrometer with a 196-nm wavelength.The experimental procedures were carried out according to
he National Institute of Health Guidelines for Animal Care andpproved by the Ethical Committee of the Sfax Faculty of Sciences.
One week after acclimatization, rats were randomly dividednto four groups of six animals each: group I served as controls
hich received standard diet; group II received via drinking water2Cr2O7 alone (700 ppm, orally); group III received both 700 ppmf K2Cr2O7 and Se (0.5 mg/kg of diet) and group IV served as a pos-tive control and received Se (0.5 mg/kg of diet) as sodium seleniteNa2SeO3). The present study was designed to investigate the tox-city of K2Cr2O7 given to rats via drinking water. Se was added inhe diet (0.5 mg/kg of diet). K2Cr2O7 and Se were administrateduring 21 days. The dose of K2Cr2O7 and the period of treatmentere selected on the basis of previous studies (Trivedi et al., 1989;
unaid et al., 1995, 1996a,b; Kanojia et al., 1996, 1998). The seleniumose (0.5 mg/kg of diet) used in our experiment and in other find-
ngs gave high protection against stress conditions in several tissuesOgnjanovic et al., 2008; Ben Amara et al., 2009). According to Hotzt al. (1997), lower doses of selenium gave less protection whileigher doses were not much effective. During the experimentaleriod, food and water intake were monitored daily. The amount of
ngested diet was calculated as the difference between the weightf feed that remained in the food bin (Da) and the amount placed
day before (Db). These data were then used to calculate the dailyverage feed intake, according to the formula:
verage feed intake = (Db − Da)
Quantities of K2Cr2O7 and Se ingested by each rat were calcu-ated from water and diet intake, respectively (Table 2).
At the end of the experimental period, animals of differentroups were killed by cervical decapitation to avoid animal stress.lood was collected into EDTA tubes and centrifuged at 2200 × g
or 10 min. Plasma samples were drawn and stored at −20 ◦C untilnalysis. Livers were dissected out, cleaned and weighed. Someamples were rinsed and homogenized (10%, w/v) in an appropri-te buffer (pH = 7.4) and centrifuged. The resulting supernatants
logic Pathology 63 (2011) 541– 548
were used for biochemical assays. Other samples were immediatelyremoved, cleaned and fixed in 10% formalin solution and embeddedin paraffin for histological studies.
2.3. Biochemical assays
2.3.1. Lipid peroxidationThe concentration of malondialdehyde (MDA) in tissues, an
index of lipid peroxidation, was determined spectrophotometri-cally according to Draper and Hadley (1990). An aliquot of 0.5 mlof liver extract supernatant was mixed with 1 ml of trichloroaceticacid solution and centrifuged at 2500 × g for 10 min. One millil-itre of a solution containing 0.67% thiobarbituric acid (TBA) and0.5 ml of supernatant were incubated for 15 min at 90 ◦C and cooled.Absorbance of TBA–MDA complex was determined at 532 nm usinga spectrophotometer. Lipid peroxidation was expressed as nmoleof thiobarbituric acid reactive substances (TBARS), using 1,1,3,3-tetra-ethoxypropane as standard.
2.3.2. Protein quantificationProtein content in the liver was determined according to Lowry
et al. (1951) using Bovine serum albumin as a standard.
2.3.3. Determination of antioxidant enzyme activitiesCatalase (CAT) activity was assayed by the method of Aebi
(1984). Enzymatic reaction was initiated by adding an aliquot of20 �l of the homogenized tissue and the substrate (H2O2) to aconcentration of 0.5 M in a medium containing 100 mM phos-phate buffer (pH 7.4). Changes in absorbance were recorded at240 nm. CAT activity was calculated in terms of �mole H2O2 con-sumed/min/mg of protein.
Superoxide dismutase (SOD) activity was estimated accordingto Beauchamp and Fridovich (1971). The reaction mixture con-tained 50 mM of tissue homogenates in potassium phosphate buffer(pH 7.8), 0.1 mM EDTA, 13 mM l-methionine, 2 �M riboflavin and75 mM nitro blue tetrazolium (NBT). The developed blue colour inthe reaction was measured at 560 nm. Units of SOD activity wereexpressed, as the amount of enzyme required to inhibit the reduc-tion of NBT by 50% and the activity was expressed as units/mg ofprotein.
Glutathione peroxidase (GPx) activity was measured accordingto Flohe and Gunzler (1984). The enzyme activity was expressed asnmole of GSH oxidized/min/mg protein.
2.3.4. Liver glutathione (GSH) levelsGSH in the liver was determined by the method of Ellman
(1959) modified by Jollow et al. (1974). The method is based onthe development of a yellow colour when DTNB (5,5-dithiobtis-2nitro benzoic acid) was added to compounds containing sulfhydrylgroups. Five hundred microlitres of tissue homogenate in phos-phate buffer were added to 3 ml of 4% sulfosalicylic acid. Themixture was centrifuged at 1600 × g for 15 min. Five hundred millil-itres of supernatants were taken and added to Ellman’s reagent.The absorbance was measured at 412 nm after 10 min. Total GSHcontent was expressed as mg/mg of protein in the liver.
2.3.5. Biochemical markers in plasma and liverThe activities of plasma lactate dehydrogenase (LDH), aspar-
tate aminotransferase (AST), alanine aminotransferase (ALT), andtotal bilirubin (TB) were assayed spectrophotometrically accord-ing to the standard procedures using commercially availablediagnostic kits (Biomaghreb, Ariana, Tunisia, Ref 20012; 20043;
20047; 20102), respectively. Plasma lipid parameters such as totalcholesterol (TC), triacylglycerol (TG) and high density lipoprotein-cholesterol (HDL-C) levels were determined by enzymatic methodsusing commercial kits from Biomaghreb (Ariana Tunis, Tunisia, Ref
N. Soudani et al. / Experimental and Toxicologic Pathology 63 (2011) 541– 548 543
Table 1Body and absolute and relative liver weights of controls (CT) and rats treated with K2Cr2O7, selenium (Se) or their combination (K2Cr2O7 + Se).
Parameters and treatments Control K2Cr2O7 K2Cr2O7 + Se Se
Initial body weight (g) 135.11 ± 1.45 136.22 ± 1.20 137.11 ± 0.90 134.45 ± 0.40Final body weight (g) 161.2 ± 3.74 162.44 ± 3.67 164.61 ± 3.34 174.67 ± 3.38*
Absolute liver weight (g) 5.43 ± 0.47 5.66 ± 0.44 5.69 ± 0.62 6.18 ± 0.14**
Relative liver weight (g/100 g b.w.) 3.37 ± 0.26 3.48 ± 0.22 3.71 ± 0.62 3.54 ± 0.03*
Values are means ± SE for seven rats in each group.* p < 0.05, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.
** p < 0.01, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.
Table 2Daily food and water consumption of controls (CT) and rats treated with K2Cr2O7, selenium (Se) or their combination (K2Cr2O7 + Se).
Parameters and treatments Food consumption (g/day/rat) Water consumption(ml/day/rat)
Ingested K2Cr2O7
quantities (mg/day/rat)Ingested Se quantities(�g/day/rat)
Control 13.14 ± 2.36 20.16 ± 1.44 – 2.14 ± 0.41K2Cr2O7 12.27 ± 1.87 12.94 ± 1.78*** 9.71 ± 1.33 2.09 ± 0.37K2Cr2O7 ± Se 16.05 ± 3.52** 12.53 ± 2.24*** ,++ 9.40 ± 1.74 8.02 ± 1.76***
Se 15.64 ± 3.20 19.37 ± 3.20 – 7.82 ± 1.61***
Values are means ± SE for seven rats in each group.** p < 0.01, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.
***
2cd1
on −
2
l(tseEus
2
gCoib(ep
3
3
pas
p < 0.001, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.++ p < 0.01, K2Cr2O7 + Se group vs. K2Cr2O7 group.
0111, 20131, 20113), respectively. The low density lipoprotein-holesterol (LDL-C) fraction and atherogenic index (AI) wereetermined according to the Friedewald equation (Friedewald,972):
LDL-C = Total cholesterol concentration − Triglyceride concentrati
AI = TC − HDL-CHDL-C
.4. Histopathological studies
Some liver samples, intended for histological examination byight microscopy, were immediately fixed in formalin solution10%) and processed in a series of graded ethanol solutions. Thenhey were embedded in paraffin, serially sectioned at 5 �m andtained with hematoxylin–eosin. Six slides were prepared fromach liver. All sections were evaluated for the degree of liver injury.ach liver slide was examined and assigned for severity of changessing scores on a scale of none (−), mild (+), moderate (++) andevere (+++) damage.
.5. Statistical analysis
The data were analysed using the statistical package pro-ram Stat view 5 Soft Ware for Windows (SAS Institute, Berkley,A). Statistical analysis was performed using one-way analysisf variance (ANOVA) followed by Fisher’s protected least signif-cant difference (PLSD) test as a post hoc test for comparisonetween groups [treated groups (K2Cr2O7, K2Cr2O7 + Se, Se) vs.controls)] and [K2Cr2O7 + Se] vs. [(K2Cr2O7)]. All values werexpressed as mean ± S.E. Differences were considered significant if
< 0.05.
. Results
.1. Evaluation of body, absolute and relative organ weights
Death was not observed during the experimental period. Resultsresented in Table 1 indicate that the body and absolute and rel-tive liver weights of treated groups (K2Cr2O7, K2Cr2O7 + Se) wereimilar to those of negative controls, whereas, the body and liver
HDL-C concentration
weights of rats, which have received 0.5 selenium mg/kg of diet,significantly increased when compared to negative controls.
3.2. Food and water consumption
Food and water consumption by adult rats in control and testedgroups is recorded in Table 2. In K2Cr2O7 and (K2Cr2O7 + Se)-treatedgroups, water consumption significantly decreased (−36 and −38%)compared to that of negative controls. In animals fed with Se, therewas a significant increase by 21% in food intake.
3.3. Estimation of lipid peroxidation
Our results revealed, in K2Cr2O7-treated rats, a significantincrease in lipid peroxidation level in the liver, as evidenced by theenhanced MDA levels. In fact, in the liver homogenate of K2Cr2O7-treated rats, MDA levels significantly increased 2.5-fold comparedto those of control group. Supplementation of Se to the diet of(K2Cr2O7 + Se) group decreased MDA contents in the liver whencompared to K2Cr2O7 group. In positive controls (Se group), liverMDA contents were not significantly changed, when compared tonegative controls.
3.4. Liver glutathione (GSH) levels
A significant decrease of glutathione (GSH) levels in liver (−18%)was evident in rats exposed to K2Cr2O7 (Table 3). Supplementa-tion of Se ameliorated GSH levels in (K2Cr2O7 + Se) group whencompared to K2Cr2O7 group.
3.5. Enzymatic antioxidant status in liver
Antioxidant enzyme activities of CAT, SOD and GPx of controland treated groups are represented in Table 3. K2Cr2O7 treatmentled to a significant increase in CAT, SOD by 63 and 24% in K2Cr2O7group, respectively, compared to those of control group, while GPx
activity was significantly lower (−41%) in K2Cr2O7 group than thatof control. Treatment with Se restored the levels of enzymaticantioxidants to near-normal values when compared with K2Cr2O7-treated rats.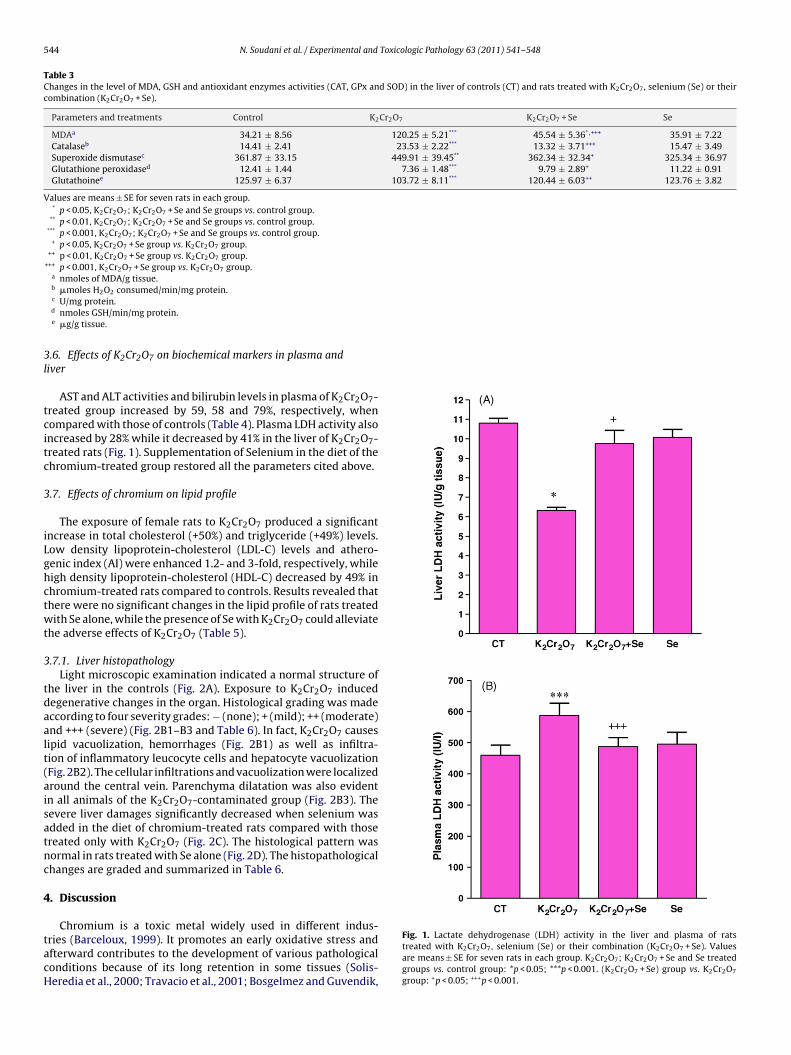
544 N. Soudani et al. / Experimental and Toxicologic Pathology 63 (2011) 541– 548
Table 3Changes in the level of MDA, GSH and antioxidant enzymes activities (CAT, GPx and SOD) in the liver of controls (CT) and rats treated with K2Cr2O7, selenium (Se) or theircombination (K2Cr2O7 + Se).
Parameters and treatments Control K2Cr2O7 K2Cr2O7 + Se Se
MDAa 34.21 ± 8.56 120.25 ± 5.21*** 45.54 ± 5.36*,+++ 35.91 ± 7.22Catalaseb 14.41 ± 2.41 23.53 ± 2.22*** 13.32 ± 3.71+++ 15.47 ± 3.49Superoxide dismutasec 361.87 ± 33.15 449.91 ± 39.45** 362.34 ± 32.34+ 325.34 ± 36.97Glutathione peroxidased 12.41 ± 1.44 7.36 ± 1.48*** 9.79 ± 2.89+ 11.22 ± 0.91Glutathoinee 125.97 ± 6.37 103.72 ± 8.11*** 120.44 ± 6.03++ 123.76 ± 3.82
Values are means ± SE for seven rats in each group.* p < 0.05, K2Cr2O7; K2Cr2O7 + Se and Se groups vs. control group.
** p < 0.01, K2Cr2O7; K2Cr2O7 + Se and Se groups vs. control group.*** p < 0.001, K2Cr2O7; K2Cr2O7 + Se and Se groups vs. control group.
+ p < 0.05, K2Cr2O7 + Se group vs. K2Cr2O7 group.++ p < 0.01, K2Cr2O7 + Se group vs. K2Cr2O7 group.
+++ p < 0.001, K2Cr2O7 + Se group vs. K2Cr2O7 group.a nmoles of MDA/g tissue.b �moles H2O2 consumed/min/mg protein.
3l
tcitc
3
iLghctwt
3
tdaalt(aisatnc
4
tacH
c U/mg protein.d nmoles GSH/min/mg protein.e �g/g tissue.
.6. Effects of K2Cr2O7 on biochemical markers in plasma andiver
AST and ALT activities and bilirubin levels in plasma of K2Cr2O7-reated group increased by 59, 58 and 79%, respectively, whenompared with those of controls (Table 4). Plasma LDH activity alsoncreased by 28% while it decreased by 41% in the liver of K2Cr2O7-reated rats (Fig. 1). Supplementation of Selenium in the diet of thehromium-treated group restored all the parameters cited above.
.7. Effects of chromium on lipid profile
The exposure of female rats to K2Cr2O7 produced a significantncrease in total cholesterol (+50%) and triglyceride (+49%) levels.ow density lipoprotein-cholesterol (LDL-C) levels and athero-enic index (AI) were enhanced 1.2- and 3-fold, respectively, whileigh density lipoprotein-cholesterol (HDL-C) decreased by 49% inhromium-treated rats compared to controls. Results revealed thathere were no significant changes in the lipid profile of rats treatedith Se alone, while the presence of Se with K2Cr2O7 could alleviate
he adverse effects of K2Cr2O7 (Table 5).
.7.1. Liver histopathologyLight microscopic examination indicated a normal structure of
he liver in the controls (Fig. 2A). Exposure to K2Cr2O7 inducedegenerative changes in the organ. Histological grading was madeccording to four severity grades: − (none); + (mild); ++ (moderate)nd +++ (severe) (Fig. 2B1–B3 and Table 6). In fact, K2Cr2O7 causesipid vacuolization, hemorrhages (Fig. 2B1) as well as infiltra-ion of inflammatory leucocyte cells and hepatocyte vacuolizationFig. 2B2). The cellular infiltrations and vacuolization were localizedround the central vein. Parenchyma dilatation was also evidentn all animals of the K2Cr2O7-contaminated group (Fig. 2B3). Theevere liver damages significantly decreased when selenium wasdded in the diet of chromium-treated rats compared with thosereated only with K2Cr2O7 (Fig. 2C). The histological pattern wasormal in rats treated with Se alone (Fig. 2D). The histopathologicalhanges are graded and summarized in Table 6.
. Discussion
Chromium is a toxic metal widely used in different indus-
ries (Barceloux, 1999). It promotes an early oxidative stress andfterward contributes to the development of various pathologicalonditions because of its long retention in some tissues (Solis-eredia et al., 2000; Travacio et al., 2001; Bosgelmez and Guvendik,Fig. 1. Lactate dehydrogenase (LDH) activity in the liver and plasma of ratstreated with K2Cr2O7, selenium (Se) or their combination (K2Cr2O7 + Se). Valuesare means ± SE for seven rats in each group. K2Cr2O7; K2Cr2O7 + Se and Se treatedgroups vs. control group: *p < 0.05; ***p < 0.001. (K2Cr2O7 + Se) group vs. K2Cr2O7
group: +p < 0.05; +++p < 0.001.

N. Soudani et al. / Experimental and Toxicologic Pathology 63 (2011) 541– 548 545
Table 4Enzyme and bilirubin activities in the plasma of controls (CT) and rats treated with K2Cr2O7, selenium (Se) or their combination (K2Cr2O7 + Se).
Parameters and treatments Control K2Cr2O7 K2Cr2O7 + Se Se
ALT (IU/l) 30.33 ± 4.91 47.95 ± 4.73*** 34.65 ± 5.72++ 29.75 ± 4.29AST (IU/l) 72.04 ± 7.94 114.81 ± 11.65*** 84.71 ± 9.09*,+++ 73.51 ± 15.45Bilirubin (mg/l) 0.66 ± 0.05 1.196 ± 0.05*** 0.81 ± 0.09*,+++ 0.62 ± 0.06
AST: aspartate transaminase, ALT: alanine transaminase, LDH: lactate dehydrogenase. Values are means ± SE for seven rats in each group.* p < 0.05, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.
*** p < 0.001, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.++ p < 0.01, K2Cr2O7 + Se group vs. K2Cr2O7 group.
+++ p < 0.001, K2Cr2O7 + Se group vs. K2Cr2O7 group.
Table 5Lipoprotein profiles of rats treated with K2Cr2O7, selenium (Se) or their combination (K2Cr2O7 + Se).
Parameters and treatments Control K2Cr2O7 K2Cr2O7 + Se Se
Total cholesterol (mg/dl) 93.71 ± 7.51 137.46 ± 14.22*** 89.95 ± 8.03+++ 96.72 ± 6.84Triglycerides (mg/dl) 85.99 ± 2.21 124.74 ± 5.41*** 86.46 ± 11.71*,+++ 86.17 ± 12.25LDL-cholesterol (mg/dl) 40.85 ± 9.33 94.61 ± 13.72*** 30.17 ± 7.76+++ 38.99 ± 8.51HDL-cholesterol (mg/dl) 35.73 ± 2.16 18.26 ± 2.79*** 42.98 ± 4.65**,+++ 40.49 ± 4.69Atherogenic index (AI) 1.64 ± 0.33 6.63 ± 1.11*** 1.105 ± 0.21*,+++ 1.42 ± 0.38
TG: triglycerides; HDL-C: high density lipoprotein-cholesterol; LDL-C: low density lipoprotein-cholesterol; AI: atherogenic index. Values are means ± SE for six rats in eachgroup.
* p < 0.05, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.
2Rcece
eTwtalivcomKie
cift(iwtpt
TG
S
** p < 0.01, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.*** p < 0.001, K2Cr2O7; K2Cr2O7 + Se and Se treated groups vs. control group.
+++ p < 0.001, K2Cr2O7 + Se group vs. K2Cr2O7 group.
004; Fatima et al., 2005; Acharya et al., 2006; Banu et al., 2008).ecent studies have been reported on the actions of agents againsthromium-induced hepatotoxicity (Anand, 2005). To our knowl-dge, our paper constitutes the first report on the exploration ofhromium hepatotoxicity in rats and the relative cytoprotectivefficiency of selenium against this metal.
In our experimental study results show no significant differ-nces in body and liver weights between K2Cr2O7 and control rats.his metal did not probably affect the appetite of rats, since thereere no differences between quantities of diet ingested by con-
rol and chromium-treated rats. While, exposure rats to seleniumlone (0.5 mg/kg of diet) during 21 days increased their body andiver weights. Thus could be explained by the increase of daily foodntake (+19%; see Table 2). Our results are in agreement with pre-ious findings of Ben Amara et al. (2009) who have found similarhanges in body weight of female rats exposed to the same dosef selenium. These authors explained the body weight gain of ani-als by an increase of their appetite. Other findings of Hawkes and
eim (2003) suggested that high doses of selenium supplementedn the diet of healthy volunteers human beings decreased energyxpenditure and consequently caused the weight gain.
Different mechanisms have been postulated to explainhromium-induced liver injury, such as lipid peroxidation andnteraction with membrane components (Bagchi et al., 1997). Inact, lipid peroxidation represents one of the most frequent reac-ions resulting from free radicals’ attack on biological structuresStohs and Bagchi, 1995). In our experiment, the MDA level signif-cantly increased in the liver of chromium-treated rats. Treatment
ith Se could prevent chromium-induced alteration probably byoxin elimination from the body or encapsulating Cr(VI), thusreventing the toxin from further oxidative injury. Likewise, Glu-athione (GSH), a crucial component of the antioxidant defense
able 6rading of the histopathological changes in the liver sections.
Groups Hemorrhage Infiltration of mononuclear cells L
Controls − − −K2Cr2O7 +++ ++ +
coring was done as follows: none (−), mild (+), moderate (++) and severe (+++).
mechanism, functions as a direct reactive free-radical scavenger(Romão et al., 2006). The decreased liver GSH level in K2Cr2O7-treated rats may be due to its consumption in the scavenging freeradicals by K2Cr2O7 (Bagchi et al., 1995a; Gunaratnam and Grant,2008). Our results are in agreement with previous studies of Anand(2005) who demonstrated that the exposure of rats to K2Cr2O7(100 mg/kg, per os) caused a significant decrease in liver GSH level.Our study showed that nutritional antioxidant Se improved GSHconcentration in the liver.
Selenium may reduce the effects of K2Cr2O7-metabolites bymeans of their interactive relations with intermediary metabolites.The most important antioxidant aspect of selenium is its function inthe active site of selenoenzyme glutathione peroxidase (Stadtman,1990). According to McPherson (1994), the last antioxidant enzymeand others such as superoxide dismutase (SOD) and catalase (CAT)constitute mutually a supportive team of defense against reactiveoxygen species. SOD is a metalloprotein which catalyses the dis-mutation of superoxide radicals (Fridovich, 1972; McCord et al.,1976). CAT is a hemoprotein which catalyses the reduction of H2O2to water and oxygen and thus protects the cell from oxidative dam-age by H2O2 and OH (Chance et al., 1952). GPx is a selenoenzymewhich plays a major role in the reduction of H2O2 and hydroper-oxide to non-toxic products (Bruce et al., 1982). Our experimentreveals an increase in SOD and CAT activities and a decrease in GPxactivity, indicating that H2O2 may be detoxified mainly by CAT inthe liver. H2O2 formed by SOD may be too high for detoxifyingby GPx. CAT acts as an antioxidant enzyme at high concentra-tions of H2O2, while GPx acts at lower H2O2 concentrations than
those of CAT (Jones et al., 1981), CAT may work more than GPx inthis tissue. The activities of these enzymes improved as a result ofSe-co-administration, suggesting the hepatoprotective role of thisoligoelement against heavy metal-induced oxidative damages asipid vacuolization Parenchyma dilatation Cell vacuolization
− −++ +++ ++
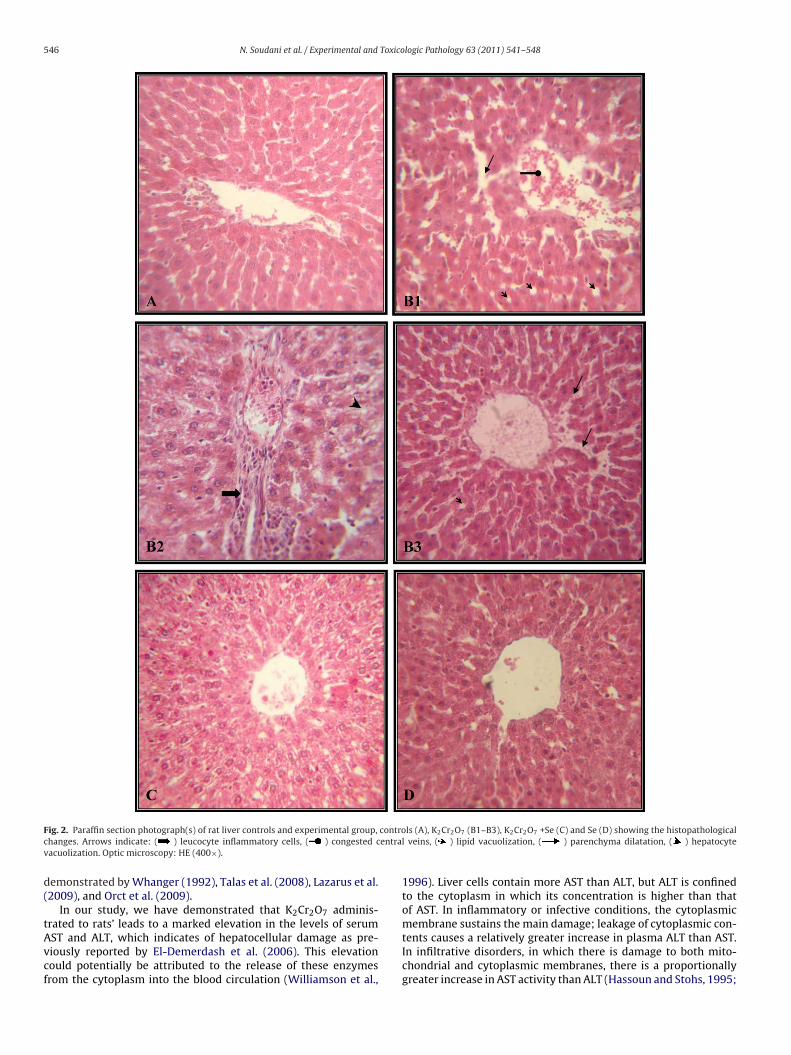
546 N. Soudani et al. / Experimental and Toxicologic Pathology 63 (2011) 541– 548
F controc entrav
d(
tAvcf
ig. 2. Paraffin section photograph(s) of rat liver controls and experimental group,
hanges. Arrows indicate: ( ) leucocyte inflammatory cells, ( ) congested cacuolization. Optic microscopy: HE (400×).
emonstrated by Whanger (1992), Talas et al. (2008), Lazarus et al.2009), and Orct et al. (2009).
In our study, we have demonstrated that K2Cr2O7 adminis-rated to rats’ leads to a marked elevation in the levels of serum
ST and ALT, which indicates of hepatocellular damage as pre-iously reported by El-Demerdash et al. (2006). This elevationould potentially be attributed to the release of these enzymesrom the cytoplasm into the blood circulation (Williamson et al.,ls (A), K2Cr2O7 (B1–B3), K2Cr2O7 +Se (C) and Se (D) showing the histopathologicall veins, ( ) lipid vacuolization, ( ) parenchyma dilatation, ( ) hepatocyte
1996). Liver cells contain more AST than ALT, but ALT is confinedto the cytoplasm in which its concentration is higher than thatof AST. In inflammatory or infective conditions, the cytoplasmicmembrane sustains the main damage; leakage of cytoplasmic con-
tents causes a relatively greater increase in plasma ALT than AST.In infiltrative disorders, in which there is damage to both mito-chondrial and cytoplasmic membranes, there is a proportionallygreater increase in AST activity than ALT (Hassoun and Stohs, 1995;
Toxico
AprDbwibtii(ipprst(oec
bsfillaftldasiliettclpvsKmpdidNogSpr1
cd(lta
N. Soudani et al. / Experimental and
kila et al., 1998). Also K2Cr2O7 hepatotoxicity is evidenced in theresent study by an increase of bilirubin activity in the plasma ofats. Our results are in agreement with previous findings of El-emerdash et al. (2006) who suggested that the increase in serumilirubin is a clear marker of hepatic dysfunction. Co-treatmentith selenium is effective in the prevention of oxidative damages
nduced by K2Cr2O7, objectified by lower liver ALT, AST and biliru-in concentrations than those of chromium-treated rats, indicatinghe important role of Se in protecting the integrity and function-ng of tissues and cells. Another biochemical marker employedn this study to evaluate liver function is lactate dehydrogenaseLDH) activity. LDH levels in the K2Cr2O7-treated group increasedn plasma and decreased in liver tissue. The elevated LDH levels inlasma may be attributed to a generalized increase in membraneermeability (Kaczor et al., 2005). On the other hand, it has beeneported that the decreased levels of LDH in the liver result fromuperoxide anions and hydroxyl radicals in the presence of transi-ion metal ions which cause oxidative damage to the cell membraneYadav et al., 1997). The co-administration of Se through the dietf K2Cr2O7-treated rats restored the levels of LDH; this could bexplained according to Ognjanovic et al. (2008) by the antiradi-al/antioxidant and metal-chelating efficacy of this trace element.
The information available about the effects of K2Cr2O7 oniochemical parameters, particularly on lipid metabolism iscanty. Indeed, K2Cr2O7 treatment causes changes in lipid pro-le; indeed, cholesterol, triglycerides (TG) and low density
ipoprotein-cholesterol (LDL-C) increased, while high densityipoprotein-cholesterol (HDL-C) decreased, suggesting that thebnormal activities of lipase enzymes seem to be one of the chiefactors responsible for the rise in serum triglycerides and choles-erol. It appears that enzymes inhibited by K2Cr2O7, are triglycerideipase, unspecific esterase and pyrophosphates. Also, hyperlipi-emia may be attributed to an increase in the synthesis of fattycids in the liver due probably to the incidence of liver cholesta-is (Owings and Georgeson, 2000). The observed abnormalitiesn lipoprotein profile may be due to an over-production of veryow density lipoprotein (VLDL) by the liver or to the decreasen the removal of VLDL and LDL from the circulation (Tsutsumit al., 1995). The liver is known as the site of cholesterol andriglycerides synthesis. The presence of Se with Cr(VI) restoreshe changes on lipidic profile to normal values. Furthermore, bio-hemical observations confirm histopathological studies on theiver. In fact, K2Cr2O7 causes hemorrhages, lipid vacuolization,arenchyma dilatation, infiltration of inflammatory leucocytes andacuolization hepatocyte cells. Similar changes in the hepatic tis-ue of Wistar rats have been reported by previous findings ofalayarasan et al. (2008). These changes could be the results ofembrane distribution induced by chromium. In fact, this metal
romotes an early oxidative stress. Afterwards it contributes to theevelopment of various pathological aspects in soft tissues includ-
ng liver. Moreover cytotoxic mediators in liver involving cells’amage as demonstrated by some researches (Fatima et al., 2005;ordberg and Arner, 2001; Ueno et al., 2001). Co-administrationf Se increased the regeneration of liver structure. Our results sug-ested that Se could reduce liver injury induced by K2Cr2O7. Indeed,e acted as an antioxidant functions and inhibited the oxidativerocesses of lipids and lipoproteins in cell membranes as it has beeneported by previous findings (McPherson, 1994; Murray et al.,988; Combs and Combs, 1984).
In conclusion, the present data records that K2Cr2O7 intoxi-ation enhances lipid peroxidation in rats liver, associated withecreased levels of GSH and altered antioxidant enzymes activities
GPx, SOD and CAT) and provokes impairments of liver histo-ogical aspects. Accordingly, care must be taken into accounto avoid mammalian and human exposure to K2Cr2O7. The co-dministration of Se with K2Cr2O7 attenuates the observed harmfullogic Pathology 63 (2011) 541– 548 547
effects on all the parameters cited above. On the basis of this study,it should be taken into consideration that the supplementation of anatural antioxidant such as selenium may act as a protective agentagainst the toxicity of K2Cr2O7.
Acknowledgments
The authors are indebted to Miss Dalenda Kchaou and Mr ChedliBouzid for their assistance in histological techniques. We also wishto extend our thanks to Mr Bejaoui Hafedh, teacher of English atSfax Faculty of Sciences, who has helped proofread and edit thispaper. This work was supported by the DGRST grants (Appui à laRecherche Universitaire de Base ARUB 99/UR/08-73), Tunisia.
References
Acharya UR, Mishra M, Tripathy RR, Mishra I. Testicular dysfunction and antioxida-tive defense system of Swiss mice after chromic acid exposure. Reprod Toxicol2006;22:87–91.
Aebi H. Catalase in vitro. Methods Enzymol 1984;105:121–6.Akila G, Rajakrishnan V, Viswanathan P, Rajashekaran KN, Menon VP. Effects of cur-
cumin on lipid profile and lipid peroxidation status in experimental hepaticfibrosis. Hepatol Res 1998;11:147–57.
Anand SS. Protective effect of vitamin B6 in chromium-induced oxidative stress inliver. J Appl Toxicol 2005;25:440–3.
Bagchi D, Balmoori J, Bagchi M, Ye X, Williams CB, Stohs SJ. Comparative effects ofTCDD, endrin, naphthalene and chromium (VI) on oxidative stress and tissuedamage in the liver and brain tissues of mice. Toxicology 2002;175:73–82.
Bagchi D, Hassoun EA, Bagchi M, Muldoon DF, Stohs SJ. Oxidative stress induced bychronic administration of sodium dichromate [Cr(VI)] to rats. Comp BiochemPhysiol C Pharmacol Toxicol Endocrinol 1995a;110:281–7.
Bagchi D, Hassoun EA, Bagchi M, Stohs SJ. Chromium induced excretion of urinarylipid metabolites, DNA damage, nitric oxide production, and generation of reac-tive oxygen species in Sprague–Dawley rats. Comp Biochem Physiol C Pharmacol1995b;110:177–87.
Bagchi D, Vuchetich PJ, Bagchi M, Hassoun EA, Tran MX, Tang L, et al. Induction ofoxidative stress by chronic administration of sodium dichromate [chromium VI]and cadmium chloride [cadmium II] to rats. Free Radic Biol Med 1997;22:471–8.
Banu SK, Samuel JB, Arosh JA, Burghardt RC, Aruldhas MM. Lactational exposureto hexavalent chromium delays puberty by impairing ovarian development,steroidogenesis and pituitary hormone synthesis in developing Wistar rats. Tox-icol Appl Pharmacol 2008;232:180–9.
Barceloux DG. Chromium. J Toxicol Clin Toxicol 1999;37:173–94.Beauchamp C, Fridovich I. Superoxide dismutase: improved assays and an assay
applicable to acrylamide gel. Anal Biochem 1971;44:276–87.Ben Amara I, Fetoui H, Guermazi F, Zeghal N. Dietary selenium addition improves
cerebrum and cerebellum impairments induced by methimazole in sucklingrats. Int J Dev Neurosci 2009;27:719–26.
Bosgelmez II, Guvendik G. Effects of taurine on oxidative stress parameters andchromium levels altered by acute hexavalent chromium exposure in mice kid-ney tissue. Biol Trace Elem Res 2004;102:209–25.
Bos gelmez II, Söylemezoglu T, Güvendik G. The protective and antidotal effects oftaurine on hexavalent chromium-induced oxidative stress in mice liver tissue.Biol Trace Elem Res 2008;125:46–58.
Bruce A, Freeman D, James C. Biology of disease-free radicals and tissue injury. LabInvest 1982;47:412–26.
Chance B, Greenstein DS, Roughton RJW. The mechanism of catalase action 1-steadystate analysis. Arch Biochem Biophys 1952;37:301–39.
Combs GF, Combs SB. The nutritional biochemistry of selenium. Ann Rev Nutr1984;4:257–80.
Diplock AT, Watkins WJ, Heurson M. Selenium and heavy metals. Ann Clin Res1986;18:55–60.
Draper HH, Hadley M. Malondialdehyde determination as index of lipid peroxida-tion. Method Enzymol 1990;86:421–31.
Eisler R. Selenium handbook of chemical risk assessment: health hazards to humans,plants, and animals, vol. 3. Boca Raton, FL: CRC Press; 2000. p. 1649–705.
El-Demerdash FM, Yousef MI, Elaswad FA. Biochemical study on the protectiverole of folic acid in rabbits treated with chromium (VI). J Environ Sci HealthB 2006;41:731–46.
Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys 1959;82:70–7.Fatima S, Arivarasu NA, Banday AA, Yusufi AN, Mahmood R. Effect of potassium
dichromate on renal brush border membrane enzymes and phosphate transportin rats. Hum Exp Toxicol 2005;24:631–8.
Flohe L, Gunzler WA. Assays of glutathione peroxidase. Method Enzymol1984;105:114–21.
Fridovich I. Superoxide radical and superoxide dismutase. Acc Chem Res1972;5:321–3.
Friedewald WT. Estimation of concentration of low-density lipoprotein choles-terol in plasma without use of the preparative ultracentrifuge. Clin Chem1972;18:499–502.

5 Toxico
G
H
H
H
J
J
J
J
J
K
K
K
K
KL
L
L
M
M
N
M
O
O
Yadav P, Sarkar S, Rhatnagar D. Action of Capparis deciduas against alloxan-induced
48 N. Soudani et al. / Experimental and
unaratnam M, Grant MH. Cr (VI) inhibits DNA, RNA and protein syntheses inhepatocytes: involvement of glutathione reductase, reduced glutathione andDT-diaphorase. Toxicol In Vitro 2008;22:879–86.
assoun EA, Stohs SJ. Comparative studies on oxidative stress as a mechanism for thefetotoxic of TCDD, endrin and lindane in C57BL/6J and DBA/2J mice. Teratology1995;51:186–92.
awkes WC, Keim NL. Dietary Selenium Intake Modulates Thyroid Hormone andEnergy Metabolism in Men. J Nutr 2003;133:3443–8.
otz CS, Fitzpatrick DW, Trick KD, L’Abbé MR. Dietary iodine and selenium interactto affect thyroid hormone metabolism of rats. J Nutr 1997;127:1214–8.
ollow DJ, Mitchell JR, Zampaglione N, Gillete JR. Bromobenzene induced liver necro-sis: protective role of glutathione and evidence for 3,4-bromobenzeneoxide asthe hepatotoxic intermediate. Pharmacology 1974;11:151–69.
ones DP, Eklöw L, Thor H, Orrenius S. Metabolism of hydrogen peroxide in iso-lated hepatocytes: relative contributions of catalase and glutathione peroxidasein decomposition of endogenously generated H2O2. Arch Biochem Biophys1981;210:505–16.
unaid M, Murthy RC, Saxena DK. Chromium fetotoxicity in mice during late preg-nancy. Vet Hum Toxicol 1995;37:320–3.
unaid M, Murthy RC, Saxena DK. Embryo-toxicity of orally administeredchromium in mice: exposure during the period of organogenesis. Toxicol Lett1996a;84:143–8.
unaid M, Murthy RC, Saxena DK. Embryo and fetotoxicity of chromium in preges-tationally exposed mice. Bull Environ Contam Toxicol 1996b;57:327–34.
aczor JJ, Ziolkowski W, Popinigis J, Tarnopolsky M. Anaerobic and aerobic enzymeactivities in human skeletal muscle from children and adults. Pediatr Res2005;57:331–5.
alayarasan S, Sriram N, Sureshkumar A, Sudhandiran G. Chromium (VI)-inducedoxidative stress and apoptosis is reduced by garlic and its derivative S-allylcysteine through the activation of Nrf2 in the hepatocytes of Wistar rats.J Appl Toxicol 2008;28:908–19.
anojia RK, Junaid M, Murthy RC. Chromium induced teratogenicity in female rat.Toxicol Lett 1996;89:207–14.
anojia RK, Junaid M, Murthy RC. Embryo and fetotoxicity of hexavalent chromium:a long-term study. Toxicol Lett 1998;95:165–72.
lasing KC. Comparative avian nutrition. New York: Oxford University Press; 1998.angard S. One hundred years of chromium and cancer: a review of epidemiological
evidence and selected case reports. Am J Ind Med 1990;17:189–215.azarus M, Orct T, Jurasoviæ J, Blanusa M. The effect of dietary selenium supple-
mentation on cadmium absorption and retention in suckling rats. Biometals2009;22:973–83.
owry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folinphenol reagent. J Biol Chem 1951;193:265–75.
cCord JM, Keele BB, Fridovich I. An enzyme based theory of obligate anaerobiosis.The physiological functions of superoxide dismutase. Proc Natl Acad Sci USA1976;68:1024–7.
cPherson A. Selenium vitamin E and biological oxidation. In: Cole DJ, Garnswor-thy PJ, editors. Recent advances in animal nutrition. Oxford: Butterworth andHeinemann’s; 1994. p. 3–30.
ordberg J, Arner ES. Reactive oxygen species, antioxidants, and the mammalianthioredoxin system. Free Radic Biol Med 2001;31:1287–312.
urray RK, Granner DK, Mayes PA, Rodwell VW. Harper’s biochemistry (a Langemedical book). international ed. Appleton and Lang Lange Medical Publication;1988. p. 574–76.
’Brien TJ, Ceryak S, Patierno SR. Complexities of chromium carcinogenesis: role
of cellular response, repair and recovery mechanisms. Mutat Res 2003;533:3–36.ccupational Safety and Health Administration (OSHA), Department of Labor.Occupational exposure to hexavalent chromium. Final rule Fed Regist2006;71:10099–385.
logic Pathology 63 (2011) 541– 548
Ognjanovic BI, Markovic SD, Pavlovic SZ, Zikic RV, Stajn AS, Saicic ZS. Effect of chroniccadmium exposure on antioxidant defense system in some tissues of rats: pro-tective effect of selenium. Physiol Res 2008;57:403–11.
Orct T, Lazarus M, Jurasovic J, Blanusa M, Piasek M, Kostial K. Influence of sele-nium dose on mercury distribution and retention in suckling rats. J Appl Toxicol2009;29:585–9.
Owings E, Georgeson K. Management of cholestasis in infants with very low birthweight. Sem Pediatr Surg 2000;9:96–102.
Pedraza-Chaverri JD, Barrera ON, Medina-Campos RC, Carvajal R, Hernandez-Pando NA, Macias-Ruvalcaba PD, et al. Time course study of oxidative andnitrosative stress and antioxidant enzymes in K2Cr2O7-induced nephrotoxicity.BMC Nephrol 2005;6:4.
Romão PRT, Tovar J, Fonseca SG, Moraes RH, Cruz AK, Hothersall JS, et al. Glu-tathione and the redox control system trypanothione/trypanothione reductaseare involved in the protection of Leishmania spp. against nitrosothiol-inducedcytotoxicity. Braz J Med Biol Res 2006;39:355–63.
Salnikow K, Zhitkovich A. Genetic and epigenetic mechanisms in metal carcino-genesis and cocarcinogenesis: nickel, arsenic, and chromium. Chem Res Toxicol2008;21:28–44.
Solis-Heredia MJ, Quintanilla-Vega B, Sierra-Santoyo A, Hernandez JM, Brambila E,Cebrian ME, et al. Chromium increases pancreatic metallothionein in the rat.Toxicology 2000;142:111–7.
Stadtman TC. Selenium biochemistry. Annu Rev Biochem 1990;59:111–27.Stohs SJ, Bagchi D. Oxidative mechanisms in the toxicity of metal ions. Free Radic
Biol Med 1995;18:321–36.Talas ZS, Orun I, Ozdemir I, Erdogan K, Alkan A, Yilmaz I. Antioxidative role of sele-
nium against the toxic effect of heavy metals (Cd+2, Cr+3) on liver of rainbowtrout (Oncorhynchus mykiss Walbaum 1792). Fish Physiol Biochem 2008;34:217–22.
Travacio M, Polo JM, Llesuy S. Chromium (VI) induces oxidative stress in the mousebrain. Toxicology 2001;162:139–48.
Trivedi B, Saxena DK, Murthy RC, Chandra SV. Embryotoxicity and fetotoxic-ity of orally administered hexavalent chromium in mice. Reprod Toxicol1989;3:275–8.
Tsutsumi K, Inoue Y, Shime A, Murase T. Correction of hypertriglyceridemia withlow and high-density lipoprotein cholesterol by the novel compound No 1886,a lipoprotein lipase promoting agent in STZ induced diabetic rats. Diabetes1995;44:414–7.
Ueno S, Kashimoto T, Susa N, Furukawa Y, Ishii M, Yokoi K, et al. Detection ofdichromate (VI)-induced DNA strand breaks and formation of paramagneticchromium in multiple mouse organs. Toxicol Appl Pharmacol 2001;170:56–62.
Valko M, Izakovic M, Mazur M, Rhodes CJ, Telser J. Role of oxygen radicals in DNAdamage and cancer incidence. Mol Cell Biochem 2004;266:37–56.
Whanger PD. Selenium in the treatment of heavy metals poisoning and chemicalcarcinogenesis. J Trace Elem Elect 1992;6:209–21.
Williamson LH, Andrews FM, Maykuth PL, White SL, Green EM. Biochemical changesin three-day-event horses at the beginning, middle and end of Phase C and afterPhase D. Equine Vet J Suppl 1996;22:92–8.
Xiao P, Jia XD, Zhong WJ, Jin XP, Nordberg G. Restorative effects of zinc and sele-nium on cadmium-induced kidney oxidative damage in rats. Biomed EnvironSci 2002;15:67–74.
Xu XR, Li HB, Li XY, Gu JD. Reduction of hexavalent chromium by ascorbic acid inaqueous solutions. Chemosphere 2004;57:609–13.
oxidative stress and diabetes in rat tissues. Pharm Res 1997;36:221–8.Zhitkovich A. Chromium: exposure, toxicity and biomonitoring approaches. In:
Biomarkers of environmentally associated disease: technologies, concepts, andperspectives. CRC Press; 2002. p. 269–87.




















