
Effects of ketamine on prefrontal and striatal regions in an overt verbal fluency task: a functional...
11
Psychopharmacology (2005) 183: 92–102 DOI 10.1007/s00213-005-0154-9 ORIGINAL INVESTIGATION Cynthia H. Y. Fu . Kathryn M. Abel . Matthew P. G. Allin . David Gasston . Sergi G. Costafreda . John Suckling . Steve C. R. Williams . Philip K. McGuire Effects of ketamine on prefrontal and striatal regions in an overt verbal fluency task: a functional magnetic resonance imaging study Received: 23 June 2005 / Accepted: 23 June 2005 / Published online: 15 October 2005 # Springer-Verlag 2005 Abstract Rationale: Glutamatergic dysfunction at N- methyl-D-aspartate (NMDA) receptors has been proposed as a neurochemical model for schizophrenia. A key fea- ture of this disorder is impairments in cognitive function. Objective: The present study sought to investigate the ef- fects of ketamine, an NMDA antagonist, on the perfor- mance and neural correlates of verbal fluency, a task that engages executive function. Methods: Ten healthy dextral male volunteers received intravenous placebo normal saline or ketamine (bolus of 0.23 mg/kg and infusion of 0.65 mg/ kg), administered in a double-blind, randomized order, during two functional magnetic resonance imaging ses- sions. During scanning, subjects performed a verbal fluency task. Two levels of cognitive load were examined in the task, and overt responses were acquired in order to measure subject performance on-line. Results: Ketamine induced symptoms in the healthy individuals comparable to an acute psychotic state. Although ketamine did not signif- icantly impair task performance relative to placebo, an in- teraction of task demand with ketamine was observed in the anterior cingulate, prefrontal, and striatal regions. Conclusions: The behavioural and functional effects of ketamine during verbal fluency in healthy individuals were comparable to those evident in patients with schizophrenia. The findings support a role for glutamatergic dysfunction in the pathophysiology of schizophrenia. Keywords Ketamine . N-methyl-D-aspartate . NMDA . Schizophrenia . Verbal fluency . Executive function . Functional magnetic resonance imaging . fMRI Introduction Psychotomimetic drugs that are noncompetitive antago- nists at the N-methyl-D-aspartate (NMDA) site, such as phenylcyclidine and ketamine, illustrate a neurochemical model for the syndrome of schizophrenia (Luby et al. 1959; Krystal et al. 1994; Adler et al. 1999). The NMDA receptor complex (Vincent et al. 1979; Zukin and Zukin 1979) is a class of receptors for the excitatory amino acid glutamate, the major excitatory neurotransmitter in the brain (Coyle et al. 2001). Phencyclidine (PCP) and its derivatives, cyclo- hexamine and ketamine, were introduced as general an- aesthetics in the 1950s (Collins et al. 1960; Corssen and Domino 1966). These ‘dissociative anaesthetics’ induce a behavioural dissociated state that resembles catatonia, as patients develop waxy flexibility and a flat expression with fixed, sightless staring. Moreover, a significant proportion of patients also developed psychotic symptoms of para- noia, hallucinations and thought disorder, as well as bizarre behaviour and agitation (Meyer et al. 1959; reviewed in Javitt and Zukin 1991). These observations led to proposals for NMDA antagonists to be a pharmacological model for schizophrenia (Luby et al. 1959; Ban et al. 1961). However, PCP produces neurotoxic effects in the rat brain (Olney et al. 1989) and is associated with prolonged psychosis (Luby et al. 1959; Meyer et al. 1959; Rainey and C. H. Y. Fu (*) . M. P. G. Allin . D. Gasston . S. G. Costafreda . S. C. R. Williams . P. K. McGuire Institute of Psychiatry, King’ s College London, De Crespigny Park, London, SE5 8AF, UK e-mail: [email protected] Tel.: +44-207-8485350 Fax: +44-207-8485229 K. M. Abel Department of Psychiatry and Behavioural Sciences, Manchester University, Williamson Building, Manchester, M13 9PL, UK J. Suckling Brain Mapping Unit, Department of Psychiatry, Addenbrooke’ s Hospital, University of Cambridge, Cambridge, CB2 2QQ, UK
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effects of ketamine on prefrontal and striatal regions in an overt verbal fluency task: a functional...

Psychopharmacology (2005) 183: 92–102DOI 10.1007/s00213-005-0154-9
ORIGINAL INVESTIGATION
Cynthia H. Y. Fu . Kathryn M. Abel .Matthew P. G. Allin . David Gasston .Sergi G. Costafreda . John Suckling .Steve C. R. Williams . Philip K. McGuire
Effects of ketamine on prefrontal and striatal regions in an overtverbal fluency task: a functional magnetic resonanceimaging studyReceived: 23 June 2005 / Accepted: 23 June 2005 / Published online: 15 October 2005# Springer-Verlag 2005
Abstract Rationale: Glutamatergic dysfunction at N-methyl-D-aspartate (NMDA) receptors has been proposedas a neurochemical model for schizophrenia. A key fea-ture of this disorder is impairments in cognitive function.Objective: The present study sought to investigate the ef-fects of ketamine, an NMDA antagonist, on the perfor-mance and neural correlates of verbal fluency, a task thatengages executive function. Methods: Ten healthy dextralmale volunteers received intravenous placebo normal salineor ketamine (bolus of 0.23 mg/kg and infusion of 0.65 mg/kg), administered in a double-blind, randomized order,during two functional magnetic resonance imaging ses-sions. During scanning, subjects performed a verbal fluencytask. Two levels of cognitive load were examined in thetask, and overt responses were acquired in order to measuresubject performance on-line. Results: Ketamine inducedsymptoms in the healthy individuals comparable to anacute psychotic state. Although ketamine did not signif-
icantly impair task performance relative to placebo, an in-teraction of task demand with ketamine was observedin the anterior cingulate, prefrontal, and striatal regions.Conclusions: The behavioural and functional effects ofketamine during verbal fluency in healthy individuals werecomparable to those evident in patients with schizophrenia.The findings support a role for glutamatergic dysfunctionin the pathophysiology of schizophrenia.
Keywords Ketamine . N-methyl-D-aspartate . NMDA .Schizophrenia . Verbal fluency . Executive function .Functional magnetic resonance imaging . fMRI
Introduction
Psychotomimetic drugs that are noncompetitive antago-nists at the N-methyl-D-aspartate (NMDA) site, such asphenylcyclidine and ketamine, illustrate a neurochemicalmodel for the syndrome of schizophrenia (Luby et al. 1959;Krystal et al. 1994; Adler et al. 1999). The NMDA receptorcomplex (Vincent et al. 1979; Zukin and Zukin 1979) is aclass of receptors for the excitatory amino acid glutamate,the major excitatory neurotransmitter in the brain (Coyleet al. 2001). Phencyclidine (PCP) and its derivatives, cyclo-hexamine and ketamine, were introduced as general an-aesthetics in the 1950s (Collins et al. 1960; Corssen andDomino 1966). These ‘dissociative anaesthetics’ induce abehavioural dissociated state that resembles catatonia, aspatients develop waxy flexibility and a flat expression withfixed, sightless staring. Moreover, a significant proportionof patients also developed psychotic symptoms of para-noia, hallucinations and thought disorder, as well as bizarrebehaviour and agitation (Meyer et al. 1959; reviewed inJavitt and Zukin 1991). These observations led to proposalsfor NMDA antagonists to be a pharmacological modelfor schizophrenia (Luby et al. 1959; Ban et al. 1961).However, PCP produces neurotoxic effects in the rat brain(Olney et al. 1989) and is associated with prolongedpsychosis (Luby et al. 1959; Meyer et al. 1959; Rainey and
C. H. Y. Fu (*) . M. P. G. Allin . D. Gasston .S. G. Costafreda . S. C. R. Williams .P. K. McGuireInstitute of Psychiatry,King’s College London,De Crespigny Park,London, SE5 8AF, UKe-mail: [email protected].: +44-207-8485350Fax: +44-207-8485229
K. M. AbelDepartment of Psychiatry and Behavioural Sciences,Manchester University,Williamson Building,Manchester, M13 9PL, UK
J. SucklingBrain Mapping Unit,Department of Psychiatry,Addenbrooke’s Hospital,University of Cambridge,Cambridge, CB2 2QQ, UK

Crowder 1975). In contrast, ketamine has about one-tenththe relative potency of PCP (Javitt and Zukin 1991) anddoes not have similar long-term consequences (Rainey andCrowder 1975; Schorn and Whitwan 1980).
Schizophrenia is characterized by positive psychoticsymptoms, negative withdrawal features, and cognitiveimpairments (Liddle 1987). In healthy individuals, keta-mine induces symptoms comparable to the positive symp-toms of schizophrenia, such as hallucinations, delusions,and thought disorder, in addition to the negative symptomsof apathy, affective blunting, and social withdrawal (Lubyet al. 1959; Breier et al. 1997; Vollenweider et al. 1997).Ketamine also produces impairments in cognitive function,namely in selective attention, symbolic thinking, episodicmemory, and verbal fluency (Ghoneim et al. 1985; Krystalet al. 1994; Malhotra et al. 1996; Newcomer et al. 1999;Hetem et al. 2000). In patients with schizophrenia, keta-mine effectively “rekindles” their core psychotic symptomsand induces cognitive impairments (Luby et al. 1959; Banet al. 1961; Lahti et al. 1995; Malhotra et al. 1997). Theeffects of NMDA antagonists are thus in contrast to dopa-minergic agonists which primarily elicit positive symptoms(Angrist et al. 1982; Carter et al. 1997) and implicate ahypoglutamatergic state or dysregulation of NMDA recep-tor function as a neurochemical aetiology of schizophrenia(Javitt and Zukin 1991; Jentsch and Roth 1999; Tamminga1999; Tsai and Coyle 2002).
Most studies in schizophrenia are confounded by thepresence of antipsychotic medication, and animal studiesare not able to reproduce the complexity and individual per-spective of psychotic symptoms (Fu and McGuire 1999).Investigation of the effects of ketamine in healthy individ-uals provides a psychopharmacological paradigm that elic-its symptoms comparable to those of schizophrenia andallows personal reports of psychotomimetic experiences.Functional neuroimaging studies of a ketamine-inducedpsychotic state in healthy volunteers have observed in-creased resting state activity in the anterior cingulate andprefrontal cortices (Breier et al. 1997; Vollenweider et al.1997; Holcomb et al. 2001), with the severity of psychoticsymptoms showing a correlation with activity in these re-gions (Breier et al. 1997; Vollenweider et al. 1997; Holcombet al. 2001). In patients with schizophrenia, ketamine simi-larly elicited increased resting state activity in the anteriorcingulate cortex (Lahti et al. 1995).
As cognitive impairments are a key feature of schizo-phrenia, we sought to study the neural effects of ketaminein a task which engages executive function. A classicalmeasure of executive function is the verbal fluency task(Benton and Hamsher 1994; Lezak 1995), and patientswith schizophrenia show impaired performance in verbalfluency (Allen et al. 1993), particularly during an acutepsychotic episode (Rice et al. 2002; Tso et al. 2002;Kravariti et al. 2005). In healthy individuals, ketamine alsoshows a specific impairment in verbal fluency (Krystalet al. 1994, 1998; Adler et al. 1998). Neuroimaging studieshave generally observed reduced activation in the anteriorcingulate and prefrontal cortices with verbal fluency tasks
in schizophrenia (Lewis et al. 1992; Yurgelun-Todd et al.1996; Curtis et al. 1998; Artiges et al. 2000). However,there is additional modulation of prefrontal activity by taskdemand and the severity of psychotic symptoms (Spenceet al. 1998; Crespo-Facorro et al. 1999; Callicott et al. 2000;Perlstein et al. 2001; Sommer et al. 2001; Fu et al. 2005).
In the present study, we used functional magnetic reso-nance imaging (fMRI) to examine the neural correlates ofa ketamine-induced psychotic state in healthy individualswhile they performed a verbal fluency task. We expectedthat a ketamine-induced psychotic state in healthy indi-viduals would be associated with similar behavioural andfunctional imaging abnormalities as evident in acutelypsychotic patients with schizophrenia. We hypothesizedthat a ketamine-induced psychosis would be associatedwith impaired performance on verbal fluency with alteredactivation in the anterior cingulate and prefrontal cortices.
Methods
Subjects
Ten healthy, dextral (Annett 1970), native English-speak-ing male volunteers [mean (SD), age 26.9 years (5.3); meanIQ 108.0 (6.7)] (Ammons and Ammons 1962), who werenaive to ketamine (and other compounds that bind toNMDA receptors, e.g.,. PCP) were recruited. All subjectswere screened with a semi-structured clinical interview bya psychiatrist (KA or CF). Exclusion criteria were a historyof psychiatric disorders including substance abuse, medicalillness, head injury, or a family history of psychosis. Fivesubjects were smokers, and none had smoked for at least1 h prior to the scan. All subjects provided written informedconsent, and ethical approval was provided by the Instituteof Psychiatry and South London and Maudsley NHS Trust.
Ketamine and placebo infusions
Subjects received either intravenous placebo (normal salinesolution) bolus over 30 s followed by a saline infusion oran intravenous ketamine bolus of 0.23 mg/kg over 30 sfollowed by an infusion of 0.65 mg/kg (Malhotra et al.1996) administered in a double-blind manner, during twoseparate scan sessions separated by a minimum of 1 day.The duration of the infusion was approximately 1 h assubjects participated in two tasks during the scan, presentedin a randomized order. Only the verbal fluency task isdiscussed in the present paper. The order of infusions wasrandomized across patients. Heart rate and blood pressurewere measured prior to and immediately following thebolus and at 10-min intervals throughout the scan. TheBrief Psychiatric Rating Scale (BPRS) (Overall and Gorham1962) and the observer version of the Clinician Adminis-tered Dissociative States Scale (CADSS) (Bremner et al.1991) were administered (KA or MA) at the beginning andend of each scan session.
93

Verbal fluency task
Subjects were required to overtly articulate a word be-ginning with a visually presented letter. Cognitive load wasmodulated by two levels of task difficulty: ‘easy’ and‘hard’ conditions which involved letters that differed withrespect to the ease with which volunteers can usuallygenerate words beginning with them. The easy conditionwas one of two sets of letter stimuli: T, L, B, R, S or T, C, B,P, S; and hard condition either, O, A, N, E, G or I, F, N, E, G(Fu et al. 2002). Incorrect responses were defined as wordsthat were proper names, repetitions or grammatical vari-ations of the previous word, and ‘pass’ responses (Bentonand Hamsher 1994; Lezak 1995). Each subject was pre-sented with a different set of easy and hard letter cues ineach of the two scan sessions. The order of presentationwas randomized between and within subjects. For theexperimental condition, seven presentations of each letterwere displayed at 4-s intervals in block, lasting a total of28 s. The experimental block alternated with the controlcondition of word repetition, consisting of a 28-s blockof seven presentations of the word ‘rest’ displayed at 4-sintervals, which subjects were required to read aloud. Thefive blocks of each control (A) and experimental (B) con-dition were presented in an alternating ABAB order.
fMRI image acquisition
T2*-weighted gradient echo single-shot echo-planar im-ages were acquired on a 1.5-T, neuro-optimized, IGE LXSystem (General Electric, Milwaukee, WI, USA) at theMaudsley Hospital, South London NHS Trust, UK. Twelvenon-contiguous axial planes (7 mm thickness, slice skip1 mm) parallel to the anterior commissure–posterior com-missure (AC–PC) line were collected over 1.1 s (TE 40 ms,70° flip angle) (Fu et al. 2002) (Fig. 1). The letter stimuluswas presented immediately after each acquisition, remain-ing visible for 750 ms (height 7 cm and subtending a 0.4°field of view). The time to repetition (TR) was 4.0 s, whichallowed subjects to make a single overt verbal response inrelative silence, a duration of 2.9 s. Head movement wasminimized by a forehead strap. Seventy-four volumes wereacquired for each condition. The term ‘clustered’ or ‘com-pressed’ has been applied to this form of image acquisition,which has an advantage of providing a period of relativesilence when the scanner is not acquiring images, thusallowing subjects to speak aloud and hear themselvesclearly in the absence of scanner noise (Amaro et al. 2002).Additionally, the TR was determined such that any con-comitant head movement would be resolved prior to thenext image acquisition (Amaro et al. 2002; Fu et al. 2002).
Verbal responses were recorded via an MRI-compatiblemicrophone on Cool Edit 2000 (Syntrillium SoftwareCorporation). In order to ensure that subjects heard theirresponses clearly, their voice was transmitted by an MRI-compatible microphone fixed at 5 cm anterior to the sub-ject’s mouth, amplified by a computer sound card, andrelayed back through an acoustic MRI sound system (Ward
Ray, Hampton Court, UK) and noise-insulated, stereoheadphones at a volume of 91±2 dB.
fMRI data analysis
The initial volume images of each time series, acquiredwhile the MR signal was reaching steady state (four images)and responsible for triggering the software program (fifthimage), were discarded. Subject movement was correctedby rigid-body realignment of the volumes to the seriesmean and residual magnetic history effects estimated by anautoregressive model followed by spatial smoothing with aGaussian kernel (standard deviation 3 mm). The amplitudeof response to each contrast of the design was estimated byleast-squares regression of a linear model: a convolution ofthe experimental design with two Poisson kernels (λ=2and 4 s). A map of responses derived under the null hy-pothesis of no experimental effect was generated by re-gression of the model after repeated permutation of thedata following orthogonal transformation into the waveletdomain (Bullmore et al. 2001).
Median observed and permuted responses, F*, werecalculated after affine mapping of individual maps intostandard stereotactic space (Talairach and Tournoux 1988).Group brain activation maps (GBAMs) were created byinference against the permutation distribution. Probabilisticthresholds were set such that the estimated number of type Ierror voxels across the entire volume was 10, equivalent toP<0.0001 (Brammer et al. 1997; Bullmore et al. 1999).
Image acquisition
Stimulus presentationand subject response
Scanner noise (1.1s) Silent portion (2.9s)
One block (7 trials; 28s)
BOLD responsefor each block
“Clustered” imageacquisitions
time
Fig. 1 Experimental design for verbal fluency task. Brain imageswere acquired in a blocked fMRI design, in a ‘clustered’ acquisitionover 1.1 s, with 2.9 s of relative silence during which the letterstimulus was presented (for 0.75 s) and subjects made their overtverbal response. The duration for each trial was 4 s, and each blockconsisted of seven trials of the same stimulus (total time of 28 sper block). A schematic of the blood oxygenation level-dependent(BOLD) response as it is sampled by the clustered acquisition se-quence is shown in the bottom figure. For each condition, experi-mental blocks of word generation alternated with control blocks ofword repetition
94

Differences between task or group were inferred by re-gression of the general linear model: F=α0+α1H+α2X+e,where F are the responses at a particular standard spacevoxel location, H are dummy codes for the condition orgroup, X is an optional covariate, and e is the residual error.Maps of the standardized coefficient of task difficulty,which is a measure of task demand, α1*, were tested forsignificance against a two-tailed distribution, generated byrepeated randomization of H, sampling the null hypothesisof no difference between groups. To improve sensitivity,spatial information was introduced by thresholding themaps of α1* such that only voxels exceeding the criticalvalue, CV0.05, corresponding to a voxelwise test at P<0.05were retained and contiguous supra-threshold voxels ag-gregated into three-dimensional clusters. The sum of (α1*−CV0.05) for each cluster, the spatial extent statistic, wasthen tested for significance against the identically derivedrandomization distribution at a probabilistic threshold suchthat the estimated number of false-positive clusters acrossthe entire volume was <1 (Bullmore et al. 2001).
GBAMS of the overall response (verbal fluency exper-imental condition vs word repetition control condition,irrespective of difficulty level) were generated for eachgroup. As the data were acquired in a blocked design foreach condition, the easy and hard conditions were con-trasted within each group using a categorical comparisonand coding the covariate X as the number of erroneousresponses produced.
In order to test our hypotheses, the analysis consisted ofa 2×2×2 factorial design with the three factors being task(verbal fluency, word repetition), drug (ketamine, placebo),and task demand (easy, hard). Group maps of the taskresponse (verbal fluency vs repetition, irrespective of taskdemand) were generated for each drug (ketamine andplacebo). The main effects of each factor and their inter-action were examined, covarying for the number of erro-neous responses.
Results
Behavioural effects of ketamine
Mean total BPRS and CADSS scores increased with ke-tamine: BPRS with placebo 0.9 (SD 0.7) and ketamine 9.6(SD 4.7) (F=26.9, df=1,13, P<0.001), and CADSS withplacebo 0.1 (SD 0.4) and ketamine 22.4 (SD 17.8) (F=12.7,df=1,13, P<0.003). These differences reflected increases indepersonalization, suspiciousness, thought disorder, elation,and psychomotor excitation with ketamine. The symptomswere typical of those observed with similar dosages of ke-tamine (Abel et al. 2003b). All subjects recovered fullywithin 1 h of completing the scan session.
Task performance
During the placebo infusion, subjects generated 32.5 (SD1.6) correct responses during the easy condition and 29.5
(SD 3.5) during the hard condition. With ketamine, therewere 33.4 (SD 1.2) words produced during the easycondition and 28.7 (SD 3.4) during the hard trials. A maineffect of task demand was observed with more correctresponses produced during the easy than the hard condition(repeated-measures ANOVA, F=28.7, df=1, 9, P<0.001).However, there was no main effect of drug (F=0.004, df=1,9, P=0.95) nor an interaction of task demand with drug(F=1.0, df=1, 9, P=0.34).
fMRI data
Activation during verbal fluency (main effect of task)
During both the placebo and ketamine conditions, verbalfluency (relative to word repetition) engaged a distributedset of regions, which included the anterior cingulate gyri,middle and inferior prefrontal gyri, insula, parietal andlateral temporal cortices, and cerebellum. The pattern ofactivation closely resembled that observed with the sametask in a different group of healthy individuals (Fu et al.2002) and more generally in other studies (Indefrey andLevelt 2000). A full list of coordinates is available uponrequest.
Main effect of task demand
No regions were significantly more activated during thehard than easy condition, with covariance for performance.
Main effect of ketamine
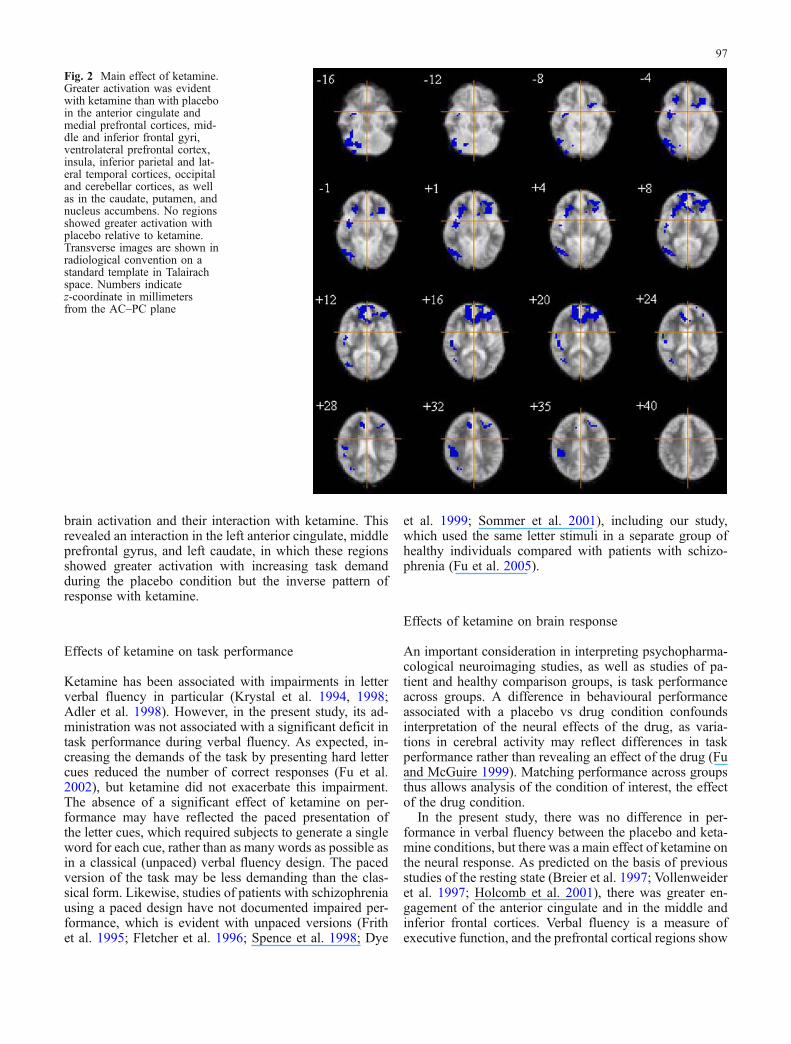
A main effect of drug (ketamine vs placebo, irrespective oftask difficulty) was evident in the anterior cingulate andmedial prefrontal cortices; middle and inferior frontal gyri,ventrolateral prefrontal cortex; insula; inferior parietal andlateral temporal cortices; occipital and cerebellar cortices;as well as in the caudate, putamen, and nucleus accumbens(P<0.03, estimated number of false-positive clusters <1)(Table 1, Fig. 2). All regions showed greater activationwith ketamine than with placebo. No regions showedgreater activation with placebo relative to ketamine.
Interaction effect of ketamine and task demand
A ketamine by task demand interaction was evident in twoclusters (P<0.02, estimated number of false-positive clusters<1) (Fig. 3). One large cluster (404 voxels) encompassedthe head of the left caudate (extending from {x, y, z}={−16,20, 8} to {−16, 20, 16) cluster size 36 voxels), left anteriorcingulate gyrus [(BA 32) {−27, 34, 20} to {−14, 25, 28)168 voxels], and left middle frontal gyrus [(BA 9) {−28,36, 24} to {−28, 36, 35}]. Post hoc analysis indicated thatthere was increased activation with increasing task demandduring the placebo infusion (F=6.6, df=1,9, P<0.03), but
95

the inverse relationship during the ketamine infusion withgreater activation during the easy than the hard condition(F=5.3, df=1,9, P<0.05). Moreover, there was a significanteffect of ketamine during the easy condition, with greateractivation elicited during the ketamine infusion relativeto placebo (F=14.7, df=1,9, P<0.004), but no differenceduring the hard condition (F=1.4, df=1,9, P=0.27) A sec-ond cluster (200 voxels) extended from the mid-cingulategyrus to the left precentral gyrus (centroid {−23, −11, 34}).This region showed no difference in activation with taskdemand during the placebo infusion, but greater activationwith increasing task demand in the presence of ketamine.
Discussion
To investigate the glutamatergic dysfunction model ofschizophrenia, the present study examined the neurocog-nitive effects of a ketamine-induced psychotic state inhealthy individuals. Previous studies have observed thatthe administration of ketamine is associated with impair-ments in cognitive function (Ghoneim et al. 1985; Krystalet al. 1994; Malhotra et al. 1996; Adler et al. 1999) and anincrease in resting state activity in anterior cingulate andprefrontal cortical regions (Vollenweider et al. 1997; Holcombet al. 2001). However, its effects on executive function andregional brain activation have largely been studied sepa-rately. In the present study, by combining fMRI with on-line measurement of behavioural responses, we were ableto examine the effect of ketamine on cognitive performanceand brain activation concurrently using a paradigm knownto be associated with both behavioural and functional neuralabnormalities in schizophrenia. As expected, when healthyvolunteers received ketamine, they rapidly exhibited per-ceptual abnormalities and reality distortions consistent withthe symptomatic effects described in previous studies withketamine and comparable to the symptoms of schizophre-nia (Luby et al. 1959; Krystal et al. 1994; Malhotra et al.1996; Adler et al. 1999). The design of the present studyalso allowed examination of the effects of cognitive load on
Table 1 Main effect of ketamine
Anatomical region (Brodmann’s area) Talairachcoordinates
Clustersize
x y z
Anterior cingulate gyrus(BA 32) L −5 25 24 4
L −9 40 16 420L −18 40 12 24R 10 25 35 132
11 35 24(BA 24/32) R 6 44 12 456
12 37 8Medial prefrontal cortex (BA 9) L −20 55 24 424
−9 40 20Middle prefrontal gyrus(BA 9/46) L −25 32 35 104
−35 28 24(BA 10) L −38 40 12 56Inferior frontal gyrus (BA 45) L −31 36 8 144
L −31 31 1 136R 28 22 1 100
Ventrolateral prefrontal cortex (BA 47) L −27 20 −4 104R 26 27 −4 56
Insula R 46 −12 4 20836 −7 −8
Precentral gyrus(BA 4) R 55 −8 24 24
55 −8 12(BA 6) R 48 −13 12 20Postcentral gyrus (BA 2) R 50 −21 28 48
50 −22 24Inferior parietal gyrus (BA 40) R 45 −32 35 204
50 −37 28Superior temporal gyrus (BA 42/22) R 48 −29 20 124
48 −13 8Middle temporal gyrus (BA 39) R 37 −66 28 52
42 −63 16Inferior temporal gyrus (BA 37) R 48 −66 −8 44Occipital gyrus (BA 18/19) R 29 −68 32 8
29 −68 28R 41 −74 12 504
44 −80 −8R 33 −91 −8 20
Lingual gyrus (BA 18/19) R 18 −76 −4 818 −76 −8
Caudate nucleus L −12 18 4 28−11 15 −1
Putamen L −25 15 −8 40Putamen/globus pallidus L −30 34 4 172
−32 23 −1R 28 21 4 140
26 27 −1Nucleus accumbens R 35 −7 −12 32
27 −5 −20
Anatomical region (Brodmann’s area) Talairachcoordinates
Clustersize
x y z
Cerebellum R 34 −67 −16 33632 −67 −20
R 19 −74 −12 20R 28 −90 −20 24
All regions showed greater activation with ketamine relative toplacebo. No regions showed greater activation with placebo relativeto ketamine. Cluster size indicates the number of voxels in eachcluster. The Talairach and Tournoux coordinates for the centroid ofeach cluster are presented. If a cluster volume extended over two ormore slices, the most dorsal and ventral coordinates are presentedBA Brodmann’s area, R right hemisphere, L left hemisphere
Table 1 (continued)
96
brain activation and their interaction with ketamine. Thisrevealed an interaction in the left anterior cingulate, middleprefrontal gyrus, and left caudate, in which these regionsshowed greater activation with increasing task demandduring the placebo condition but the inverse pattern ofresponse with ketamine.
Effects of ketamine on task performance
Ketamine has been associated with impairments in letterverbal fluency in particular (Krystal et al. 1994, 1998;Adler et al. 1998). However, in the present study, its ad-ministration was not associated with a significant deficit intask performance during verbal fluency. As expected, in-creasing the demands of the task by presenting hard lettercues reduced the number of correct responses (Fu et al.2002), but ketamine did not exacerbate this impairment.The absence of a significant effect of ketamine on per-formance may have reflected the paced presentation ofthe letter cues, which required subjects to generate a singleword for each cue, rather than as many words as possible asin a classical (unpaced) verbal fluency design. The pacedversion of the task may be less demanding than the clas-sical form. Likewise, studies of patients with schizophreniausing a paced design have not documented impaired per-formance, which is evident with unpaced versions (Frithet al. 1995; Fletcher et al. 1996; Spence et al. 1998; Dye
et al. 1999; Sommer et al. 2001), including our study,which used the same letter stimuli in a separate group ofhealthy individuals compared with patients with schizo-phrenia (Fu et al. 2005).
Effects of ketamine on brain response
An important consideration in interpreting psychopharma-cological neuroimaging studies, as well as studies of pa-tient and healthy comparison groups, is task performanceacross groups. A difference in behavioural performanceassociated with a placebo vs drug condition confoundsinterpretation of the neural effects of the drug, as varia-tions in cerebral activity may reflect differences in taskperformance rather than revealing an effect of the drug (Fuand McGuire 1999). Matching performance across groupsthus allows analysis of the condition of interest, the effectof the drug condition.
In the present study, there was no difference in per-formance in verbal fluency between the placebo and keta-mine conditions, but there was a main effect of ketamine onthe neural response. As predicted on the basis of previousstudies of the resting state (Breier et al. 1997; Vollenweideret al. 1997; Holcomb et al. 2001), there was greater en-gagement of the anterior cingulate and in the middle andinferior frontal cortices. Verbal fluency is a measure ofexecutive function, and the prefrontal cortical regions show
Fig. 2 Main effect of ketamine.Greater activation was evidentwith ketamine than with placeboin the anterior cingulate andmedial prefrontal cortices, mid-dle and inferior frontal gyri,ventrolateral prefrontal cortex,insula, inferior parietal and lat-eral temporal cortices, occipitaland cerebellar cortices, as wellas in the caudate, putamen, andnucleus accumbens. No regionsshowed greater activation withplacebo relative to ketamine.Transverse images are shown inradiological convention on astandard template in Talairachspace. Numbers indicatez-coordinate in millimetersfrom the AC–PC plane
97

the greatest distinction between healthy and schizophrenicindividuals (Fu and McGuire 1999). In addition, the degreeof medial prefrontal activity has been correlated withseverity of ‘ego identity’ impairments, such as a loss of‘ego boundaries’ or sense of self, in healthy individualsreceiving ketamine (Vollenweider et al. 1997). Severalrecent studies have examined the neural effects of ketaminewith activation paradigms. Cognitive tasks, involving verbalworking memory and episodic memory, have found a maineffect of ketamine relative to placebo in the regions en-compassed by the tasks, including the anterior cingulateand prefrontal cortical areas (Honey et al. 2004, 2005;Northoff et al. 2005), whereas an emotional processing taskrevealed an effect of ketamine primarily in the visual cor-tices, a region involved in affective facial processing (Abelet al. 2003a). In each of these paradigms, ketamine mod-ulated cerebral activity that was specific to the task utilized.
With our verbal fluency task, ketamine also augmentedsubcortical activations in the caudate and putamen as wellas in the cerebellum. Increased resting activity in the pu-tamen following the administration of ketamine has beencorrelated with a ‘schizophrenic syndrome’ that includes
measures of apathy, thought disorder, and hallucinations(Vollenweider et al. 1997). Although ketamine was alsoassociated with increased resting cerebellar activity in thesame study (Vollenweider et al. 1997), it has more fre-quently been linked to a decrease in cerebellar activity(Lahti et al. 1995; Breier et al. 1997; Holcomb et al. 2001).In the present study, subjects were engaged in a cognitivetask that engages the cerebellum (Petersen et al. 1988; Fu etal. 2002), whereas in previous studies they were scanned atrest (Lahti et al. 1995; Holcomb et al. 2001) or whileperforming a relatively undemanding continuous perfor-mance task (Breier et al. 1997). NMDA receptors have awide distribution throughout the cerebral cortex; however,ketamine binds to a site within the channel of the receptorcomplex which must first be depolarized (‘activated’) priorto binding (Cotman and Monaghan 1987). The cerebellumhas a role not only in motor control, but also in languageand memory processes (Desmond and Fiez 1998), and theright cerebellar hemisphere, as in the present study, hasbeen particularly involved in studies of verbal fluency(Petersen et al. 1988; Desmond and Fiez 1998; Fu et al.2002). The current finding implicates a specific augmenta-
Fig. 3 Interaction of ketamineby task demand. One cluster(coloured red) encompassed thehead of the left caudate, leftanterior cingulate (BA 32), andleft middle frontal (BA 9) gyri.This region showed increasedactivation with increasing taskdemand (from the ‘easy’ to‘hard’ conditions) during theplacebo infusion, but the inverserelationship during the ketamineinfusion (as shown in graph). Asecond cluster extended fromthe mid-cingulate gyrus to theleft precentral gyrus (colouredpurple). This cluster showed nodifference in activation with taskdemand during the placeboinfusion, but had greateractivation with increasing taskdemand in the presence ofketamine. Transverse images areshown in radiological conven-tion on a standard template inTalairach space. Numbers indi-cate z-coordinate in millimetersfrom the AC–PC plane
98

tion by ketamine of right cerebellar activation during itsrecruitment in the verbal fluency task.
Increasing the demands of verbal fluency (by presentinghard rather than easy letter cues) is normally associatedwith greater engagement of the left anterior cingulate gyrus(Fu et al. 2002). Consistent with this observation, duringthe placebo condition, increasing task demand was asso-ciated with greater activation in a cluster that included theleft anterior cingulate gyrus and adjacent portions of theleft middle frontal gyrus and caudate. Generating wordsfrom hard letters places more demands on executive func-tion and error monitoring that implicate these regions(Smith and Jonides 1999; Duncan and Owen 2000). How-ever, in the presence of ketamine, increasing the task de-mand had the opposite effect on activation in this cluster,namely, a reduction in activation. There was an inversepattern of response with ketamine in which prefrontal andstriatal regions showed reduced activation with increasingtask demand.
Several circuits connect the prefrontal cortex to the basalganglia in reciprocal and parallel pathways. Of particularrelevance to the present study are the projections from theanterior cingulate and middle frontal cortices to the caudate(Alexander et al. 1990). Glutamate is the major excitatoryneurotransmitter of cortical–subcortical projections as wellas cortico-cortical interconnections, and the prefrontal cor-tex and basal ganglia have a high density of NMDA re-ceptors (Monaghan and Cotman 1985). Glutamate as theexcitatory neurotransmitter is balanced by gamma aminobutyric acid (GABA) which is the main inhibitory neuro-transmitter (Cotman and Monaghan 1987). Two conse-quences of the blocking of NMDA receptors by ketamineare a marked release of glutamate and a reduction in localfeedback inhibition (Newcomer and Krystal 2001). Withinthe striatum, this would disrupt the inhibitory cortico–striato–thalamic feedback loops resulting in activation ofthe prefrontal cortex (Vollenweider et al. 1997). It is sug-gested that glutamate increases neuronal firing rates butdecreases its variability, which produces an impaired signal-to-noise ratio (Svensson et al. 1995). The glutamate anddopamine neurotransmitter systems also have reciprocalinterrelationships (Carlsson and Carlsson 1990). Ketamineincreases dopamine release in the striatum and prefrontalcortex, which is associated with impaired cognitive perfor-mance (Verma and Moghaddam 1996) and the develop-ment of psychotic symptoms (Breier et al. 1997; Smith etal. 1998; Vollenweider et al. 2000), and the extent ofdopamine release is comparable to the excessive responseobserved in patients with schizophrenia (Kegeles et al.2000). Enhanced prefrontal cortical activation by ketaminemay thus be a result of direct NMDA receptor antagonismor an indirect effect of other monoaminergic neurotrans-mitter activity or a combination of these effects within thecontext of the cognitive task that subjects were performing.
However, caudate and prefrontal cortical activation de-creased with increasing task demand in the presence ofketamine. Dopamine agonists also increase prefrontal neu-ronal firing and are believed to improve their signal-to-
noise ratio (Seamans et al. 1998). In an fMRI study ofverbal working memory, Mattay et al. (2000) found that theprefrontal cortical response to the dopamine agonist dex-troamphetamine was determined by its ‘baseline’ activity.Subjects who had high initial prefrontal activation showedthe smallest increase, whereas those with a lower initialactivation had the greatest subsequent increase in activa-tion (Mattay et al. 2000). It is proposed that dopamineproduces its optimal response in prefrontal function andactivity within a narrow range, and at insufficient or ex-cessive amounts, prefrontal function and activity is reduced(Williams and Goldman-Rakic 1995; Murphy et al. 1996;Arnsten 1998). As the hard trials normally recruit greateranterior cingulate activity than the easy trials (Fu et al.2002), perhaps the additional effect of ketamine exceededthe optimal range of function, and thus further activationwas not observed. This interaction with cognitive load isconsistent with the effect of ketamine observed in a work-ing memory paradigm, in which ketamine was associatedwith increased prefrontal activation when task demandswere low, but had little effect on activation when demandswere high (Honey et al. 2004). These results were in-terpreted in terms of ketamine shifting an inverted U-shaped curve denoting the relationship between activityand task demand (Honey et al. 2004). A similar mechanismcould have accounted for the effects of ketamine in thepresent study.
Relationship to brain response during verbal fluencyin schizophrenia
Overall, ketamine was associated with increased activationin a distributed set of cortical and subcortical regions im-plicated in verbal fluency. As a neurochemical model forschizophrenia, it may seem surprising that ketamine isassociated with increased cerebral activation, as ‘hypo-frontality’ has been the more traditional finding (Fu andMcGuire 1999). However, several neuroimaging studies inschizophrenia have reported increased prefrontal activa-tion, particularly in patients with positive symptoms (Volkowet al. 1986; Cleghorn et al. 1992; Ebmeier et al. 1993;McGuire et al. 1993; Shergill et al. 2000). As well, somestudies of activation during cognitive tasks have foundno difference in prefrontal activity between patients withschizophrenia and healthy controls (Frith et al. 1995;Fletcher et al. 1996; Curtis et al. 1999; Dye et al. 1999;Spence et al. 2000; Sommer et al. 2001; Fu et al. 2005) andeven hyperfrontality in patients (Stevens et al. 1998;Manoach et al. 1999). Moreover, Lahti et al. (1995) observedincreased prefrontal activity in a group of schizophrenicpatients in remission who were re-experiencing their usualpsychotic symptoms while receiving ketamine. The neuraleffects of ketamine in healthy individuals, however, maynot necessarily be fully comparable to the abnormalities oflongstanding disrupted brain circuit as in schizophrenia.Furthermore, prolonged exposure to PCP is more fre-quently associated with the severe, persistent psychotic
99

symptoms that are typical of schizophrenia, and Jentschand Roth (1999) argue that repeated administration ofNMDA receptor antagonists provides a more valid modelof schizophrenia than acute administration of a singlechallenge.
Limitations
The present study was a preliminary investigation of thebehavioural and neural effects of ketamine in healthy in-dividuals while they performed the executive function taskof verbal fluency. The sample size was small (ten subjects),and all the subjects were male, which has a risk of a type IIerror and limits the generalizability of the findings. Aswell, there were only two levels of task difficulty, whichlimits interpretation of the effects of cognitive load on ce-rebral responses. With a greater range in task demand andsubject performance, more precise delineation of the ce-rebral load response effects would be possible. Moreover,the task itself is composed of several neurocognitive pro-cesses, including response selection, error monitoring, andthe manipulation of generated items (Indefrey and Levelt2000), which is reflected in the distribution of its functionalneuroanatomy. The number of the processes inherent in thepresent task limits the specificity of the neurocognitiveanalysis. All subjects received the same dosage of keta-mine that is estimated to produce plasma ketamine levelsof 0.1–0.25 mg/l (Krystal et al. 1994), whereas a range inketamine levels may reveal dose-dependent modulation ofthe neural system involved in the cognitive task (Honeyet al. 2004).
Summary
Much evidence has supported dysfunction in prefrontalcortico-striatal circuits in schizophrenia (Goldman-Rakicand Selemon 1997). These regions have been linked to thepositive symptoms and cognitive impairments observedin schizophrenia (Goldman-Rakic and Selemon 1997;Halligan and David 2001). In healthy individuals, verbalfluency is associated with increased activation in these re-gions, with a further increase in anterior cingulate acti-vation with increasing task demand (Fu et al. 2002). Inpatients with schizophrenia, verbal fluency has revealedimpaired activation in the prefrontal cortex (Lewis et al.1992; Yurgelun-Todd et al. 1996; Curtis et al. 1998; Artigeset al. 2000), with suggestions of additional dysfunction inacutely psychotic patients (Artiges et al. 2000; Fu et al.2005). In the present study, ketamine induced a temporarypsychotic state in healthy volunteers, which was associatedwith increased activation in cortical and subcortical re-gions associated with verbal fluency. Furthermore, an in-teraction with task demand was revealed in the anteriorcingulate, middle prefrontal cortex, and caudate, which areparticularly implicated in the pathophysiology of schizo-phrenia. The specificity of the interaction offers further
support for a role for glutamatergic dysfunction in thesymptoms of schizophrenia and concomitant cognitiveimpairments.
Acknowledgements This work was supported by a WellcomeTrust Fellowship to CF and from the Mason Medical Foundation toKMA. We would also like to thank the radiographers at the MRICenter, Maudsley and South London NHS Trust, for their expertassistance. The experiments in the present study comply with thecurrent laws of the UK, in which they were performed.
References
Abel KM, Allin MPG, Kucharska-Pietura K et al (2003) Ketaminealters neural processing of facial emotion recognition in healthymen: an fMRI study. Neuroreport 14:387–391
Abel KM, Allin MPG, Hemsley DR, Geyer MA (2003) Low doseketamine induces prepulse inhibition in healthy men. Neuro-pharmacology 44:729–737
Adler CM, Goldberg TE, Malhotra AK et al (1998) Effects ofketamine on thought disorder, working memory, and semanticmemory in healthy volunteers. Biol Psychiatry 43:811–816
Adler CM, Malhotra AK, Elman I et al (1999) Comparison ofketamine-induced thought disorder in healthy volunteers andthought disorder in schizophrenia. Am J Psychiatry 156:1646–1649
Alexander GE, Crutcher MD, DeLong MR (1990) Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculo-motor, “prefrontal” and “limbic” functions. Prog Brain Res85:119–146
Allen HA, Liddle PF, Frith CD (1993) Negative features, retrievalprocesses and verbal fluency in schizophrenia. Br J Psychiatry163:769–775
Amaro E Jr, Williams SCR, Shergill SS et al (2002) Acoustic noiseand functional magnetic resonance imaging: current strategiesand future prospects. J Magn Reson Imaging 16:497–510
Ammons R, Ammons C (1962) Quick Test Missoula, MT: psy-chological test specialists
Angrist BM, Peselow E, Rubenstein M et al (1982) Partialimprovement in negative schizophrenic symptoms after am-phetamine psychosis: preliminary observations. Biol Psychiatry2:95–107
Annett MA (1970) A classification of hand preference by associ-ation analysis. Br J Psychol 61:303–321
Arnsten AF (1998) The biology of being frazzled. Science 280:1711–1712
Artiges E, Martinot JL, Verdys M et al (2000) Altered hemisphericfunctional dominance during word generation in negativeschizophrenia. Schizophr Bull 26:709–721
Ban TA, Lohrenz JJ, Lehmann HE (1961) Observations on theaction of Sernyl—a new psychotropic drug. Can PsychiatrAssoc J 6:150–156
Benton AL, Hamsher KD (1994) Multilingual aphasia examination.Oxford University Press, New York
Brammer MJ, Bullmore ET, Simmons A et al (1997) Generic brainactivation mapping in functional magnetic resonance imaging:a nonparametric approach. Magn Reson Imaging 15:763–770
Breier A, Malhotra AK, Pinals DA et al (1997) Association ofketamine-induced psychosis with focal activation of the pre-frontal cortex in healthy volunteers. Am J Psychiatry 154:805–811
Bremner JD, Krystal JH, Putnam FW et al (1991) Measurement ofdissociative states with the clinician administered dissociativestates scale. J Trauma Stress 11:25–136
Bullmore ET, Suckling J, Overmeyer S et al (1999) Global, voxel,and cluster tests, by theory and permutation, for a differencebetween two groups of structural MR images of the brain. IEEETrans Med Imag 18:32–42
100

Bullmore ET, Long C, Suckling J et al (2001) Colored noise andcomputational inference in neurophysiological (fMRI) timeseries analysis: resampling methods in time and wavelet do-mains. Hum Brain Mapp 12:61–78
Callicott JH, Bertolino A, Mattay VS et al (2000) Physiologicaldysfunction of the dorsolateral prefrontal cortex in schizophre-nia revisited. Cereb Cortex 10:1078–1092
Carlsson M, Carlsson A (1990) Interactions between glutamatergicand monoaminergic systems within the basal ganglia–implica-tions for schizophrenia and Parkinson’s disease. Trends Neu-rosci 13:272–276
Carter CS, Barch D, Cohen JD et al (1997) CNS catecholamines andcognitive dysfunction in schizophrenia. Schizophr Res 24:211
Cleghorn JM, Franco S, Szechtman B et al (1992) Towards a brainmap of auditory hallucinations. Am J Psychiatry 149:1062–1069
Collins VJ, Gorospe CA, Rovenstine EA (1960) Intravenousnonbarbiturate, nonnarcotic analgesics: preliminary studies. I.cyclohexylamines. Anesth Analg 39:303–306
Corssen G, Domino EF (1966) Dissociative anesthesia: furtherpharmacologic studies and first clinical experience with thephencyclidine derivative CI-581. Anesth Analg 45:29–40
Cotman CW, Monaghan DT (1987) Anatomical organization ofexcitatory amino acid receptors and their properties. Adv ExpMed Biol 203:237–252
Coyle JT, Leski M, Morrison J (2001) Diverse role of L-glutamateacid in brain signal transduction. In: Neuropsychopharmacol-ogy: fifth generation of progress. Lippincott, New York
Crespo-Facorro B, Paradiso S, Andreasen NC et al (1999) Recallingword lists reveals “cognitive dysmetria” in schizophrenia: apositron emission tomography study. Am J Psychiatry 156:386–392
Curtis VA, Bullmore ET, Brammer MJ et al (1998) Attenuatedfrontal activation during a verbal fluency task in patients withschizophrenia. Am J Psychiatry 155:1056–1063
Curtis VA, Bullmore ET, Morris RG et al (1999) Attenuated frontalactivation in schizophrenia may be task dependent. SchizophrRes 37:35–44
Desmond JE, Fiez JA (1998) Neuroimaging studies of the cere-bellum: language, learning and memory. Trends Cogn Neurosci2:355–362
Duncan J, Owen AM (2000) Common regions of the human frontallobe recruited by diverse cognitive demands. Trends Neurosci23:475–483
Dye SM, Spence SA, Bench CJ et al (1999) No evidence for leftsuperior temporal dysfunction in asymptomatic schizophreniaand bipolar disorder. PET study of verbal fluency. Br J Psy-chiatry 175:367–374
Ebmeier KP, Blackwood DH, Murray C et al (1993) Single-photonemission computed tomography with 99mTc-exametazime inunmedicated schizophrenic patients. Biol Psychiatry 33:487–495
Fletcher PC, Frith CD, Grasby PM et al (1996) Local and distributedeffects of apomorphine on fronto-temporal function in acuteunmedicated schizophrenia. J Neurosci 16:7055–7062
Frith CD, Friston KJ, Herold S et al (1995) Regional brain activityin chronic schizophrenic patients during the performance of averbal fluency task. Br J Psychiatry 167:343–349
Fu CHY, McGuire PK (1999) Functional neuroimaging in psychi-atry. Philos Trans R Soc Lond B 354:1359–1370
Fu CHY, Morgan K, Suckling J et al (2002) An fMRI study of overtletter verbal fluency using a clustered acquisition sequence:greater anterior cingulate activation with increased task de-mand. Neuroimage 17:871–879
Fu CHY, Suckling J, Williams SCR et al (2005) Effects of psychoticstate and task demand on prefrontal function in schizophrenia:an fMRI study of overt verbal fluency. Am J Psychiatry 162:485–494
Ghoneim MM, Hinrichs JV, Mewaldt SP et al (1985) Ketamine:behavioral effects of subanesthetic doses. J Clin Psychophar-macol 5:70–77
Goldman-Rakic PS, Selemon LD (1997) Function and anatomicalaspects of prefrontal pathology in schizophrenia. SchizophrBull 23:437–458
Halligan PW, David AS (2001) Cognitive neuropsychiatry: towardsa scientific psychopathology. Nat Neurosci Rev 2:209–215
Hetem LA, Danion JM, Diemunsch P et al (2000) Effect of a sub-anesthetic dose of ketamine on memory and conscious aware-ness in healthy volunteers. Psychopharmacology (Berl) 152:283–288
Holcomb HH, Lahti AC, Medoff DR et al (2001) Sequentialregional cerebral blood flow brain scans using PETwith H2(15)O demonstrate ketamine actions in CNS dynamically. Neuro-psychopharmacology 25:165–172
Honey GD, Honey RAE, O’Loughlin C et al (2005) Ketaminedisrupts frontal and hippocampal contribution to encoding andretrieval of episodic memory: an fMRI study. Cereb Cortex15:749–759
Honey RAE, Honey GD, O’Loughlin C et al (2004) Acute ketamineadministration alters the brain responses to executive demandsin a verbal working memory task: an fMRI study. Neuropsy-chopharmacology 29:1203–1214
Indefrey P, Levelt WJM (2000) The neural correlates of languageproduction. In: Gazzaniga MS (ed) The new cognitive neuro-sciences, 2nd edn. MIT Press, Cambridge, MA, pp 845–865
Javitt DC, Zukin SR (1991) Recent advances in the phencyclidinemodel of schizophrenia. Am J Psychiatry 148:1301–1308
Jentsch JD, Roth RH (1999) The neuropsychopharmacology ofphencyclidine: from NMDA receptor hypofunction to the do-pamine hypothesis of schizophrenia. Neuropsychopharmacolo-gy 20:201–225
Kegeles LS, Abi-Dargham A, Zea-Ponce Y et al (2000) Modulationof amphetamine-induced striatal dopamine release by ketaminein humans: implications for schizophrenia. Biol Psychiatry48:627–640
Kravariti E, Dixon T, Frith C et al (2005) Association of symptomsand executive function in schizophrenia and bipolar disorder.Schizophr Res 74:221–231
Krystal JK, Karper LP, Seibyl JP et al (1994) Subanesthetic effectsof the noncompetitive NMDA receptor antagonist, ketamine, inhumans: psychotomimetic, perceptual, cognitive, and neuroen-docrine responses. Arch Gen Psychiatry 51:199–214
Krystal JH, Karper LP, Bennett A et al (1998) Interactive effects ofsubanesthetic ketamine and subhypnotic lorazepam in humans.Psychopharmacology 135:213–229
Lahti AC, Holcomb HH, Medoff DR et al (1995) Ketamine activatespsychosis and alters limbic blood flow in schizophrenia. Neu-roreport 6:869–872
Lewis SW, Ford RA, Syed GM et al (1992) A controlled study of99mTc-HMPAO single-photon emission imaging in chronicschizophrenia. Psychol Med 22:27–35
Lezak MD (1995) Neuropsychological assessment, 3rd edn. OxfordUniversity Press, New York, pp 544–546
Liddle PF (1987) The symptoms of chronic schizophrenia. A re-examination of the positive–negative dichotomy. Br J Psychi-atry 151:145–151
Luby ED, Cohen BD, Rosenbaum F et al (1959) Study of a newschizophrenomimetic drug, Sernyl. Arch Neurol Psychiatry 81:363–369
Malhotra AK, Pinals DA, Weingartner H et al (1996) NMDAreceptor function and human cognition: the effect of ketaminein healthy volunteers. Neuropsychopharmacology 14:301–307
Malhotra AK, Pinals DA, Adler CM et al (1997) Ketamine-inducedexacerbation of psychotic symptoms and cognitive impairmentin neuroleptic-free schizophrenics. Neuropsychopharmacolgy17:141–150
Manoach DS, Press DZ, Thangaraj V et al (1999) Schizophrenicsubjects activate dorsolateral prefrontal cortex during a workingmemory task, as measured by fMRI. Biol Psychiatry 45:1128–1137
101

Mattay VS, Callicott JH, Bertolino A et al (2000) Efects of dex-troamphetamine on cognitive performance and cortical activa-tion. Neuroimage 12:268–275
McGuire PK, Shah GMS, Murray RM (1993) Increased blood flowin Broca’s area during auditory hallucinations in schizophrenia.Lancet 342:703–706
Meyer JS, Greifenstein F, DeVault M (1959) A new drug causingsymptoms of sensory deprivation. J Nerv Ment Dis 129:54–61
Monaghan DT, Cotman CW (1985) Distribution of N-methyl-D-aspartate-sensitive L-[3H]glutamate-binding sites in rat brain. JNeurosci 5:2909–2919
Murphy BL, Arnsten AF, Goldman-Rakic PS et al (1996) Increaseddopamine turnover in the prefrontal cortex impairs spatialworking memory performance in rats and monkeys. Proc NatlAcad Sci U S A 93:1325–1329
Newcomer JW, Farber NB, Jevtovic-Todorovic V et al (1999)Ketamine-induced NMDA receptor hypofunction as a model ofmemory impairment and psychosis. Neuropsychopharmacolo-gy 20:106–118
Newcomer JW, Krystal JH (2001) NMDA receptor regulation ofmemory and behavior in humans. Hippocampus 11:529–542
Northoff G, Richter A, Bermpohl F et al (2005) NMDA hypofunc-tion in the posterior cingulate as a model for schizophrenia: anexploratory ketamine administration study in fMRI. SchizophrRes 72:235–248
Olney JW, Labruyere J, Price MT (1989) Pathological changesinduced in cerebrocortical neurons by phenylcyclidine and re-lated drugs. Science 244:1360–1362
Overall JE, Gorham DR (1962) The brief psychiatric rating scale.Psychol Rep 10:799–812
Perlstein WM, Carter CS, Noll DC et al (2001) Relation ofprefrontal cortex dysfunction to working memory and symp-toms in schizophrenia. Am J Psychiatry 158:1105–1113
Petersen SE, Fox PT, Posner MI et al (1988) Positron emissiontomographic studies of the cortical anatomy of single-wordprocessing. Nature 331:585–589
Rainey JM Jr, Crowder MK (1975) Prolonged psychosis attributedto phencyclidine: report of three cases. Am J Psychiatry 132:1076–1078
Rice CD, Done DJ, Manly T et al (2002) Schizophrenic patientswith symptoms show more impairment than those withoutsymptoms on an ecologically valid test of executive function.Schizophr Res 53:133
Schorn TO, Witwam JG (1980) Are there long-term effects ofketamine on the central nervous system? Br J Anaesth 52:967–968
Seamans JK, Floresco SB, Phillips AG (1998) D1 receptormodulation of hippocampal–prefrontal cortical circuits inte-grating spatial memory with executive functions in the rat. JNeurosci 18:1613–1621
Shergill SS, Brammer MJ, Williams SC et al (2000) Mappingauditory hallucinations in schizophrenia using functional mag-netic resonance imaging. Arch Gen Psychiatry 57:1033–1088
Smith EE, Jonides J (1999) Storage and executive processes in thefrontal lobes. Science 283:1657–1661
Smith GS, Schloesser R, Brodie JD et al (1998) Glutamatemodulation of dopamine measured in vivo with positron emis-sion tomography (PET) and 11C-raclopride in normal humansubjects. Neuropsychopharmacology 18:18–25
Sommer IEC, Ramsey NF, Kahn RS (2001) Language lateralisationin schizophrenia, an fMRI study. Schizophr Res 52:57–67
Spence SA, Hirsch SR, Brooks DJ et al (1998) Prefrontal cortexactivity in people with schizophrenia and control subjects:evidence from positron emission tomography for remission of“hypofrontality” with recovery from acute schizophrenia. Br JPsychiatry 172:316–323
Spence SA, Liddle PF, Stefan MD et al (2000) Functional anatomyof verbal fluency in people with schizophrenia and those atgenetic risk. Br J Psychiatry 176:52–60
Stevens AA, Goldman-Rakic PS, Gore JC et al (1998) Corticaldysfunction in schizophrenia during auditory word and toneworking memory demonstrated by functional magnetic reso-nance imaging. Arch Gen Psychiatry 55:1097–1103
Svensson TH, Mathe JM, Andersson JL et al (1995) Mode of actionof atypical neuroleptics in relation to the phencyclidine modelof schizophrenia: role of 5-HT2 receptor and alpha 1-adreno-ceptor antagonism. J Clin Psychopharmacol 11–18S
Talairach J, Tournoux P (1988) Co-planar stereotaxic atlas of thehuman brain. Thieme, New York
Tamminga C (1999) Glutamatergic aspects of schizophrenia. Br JPsychiatry Suppl 37:12–15
Tsai G, Coyle JT (2002) Glutamatergic mechanisms in schizophre-nia. Annu Rev Pharmacol Toxicol 42:165–179
Tso IF, Chan RCK, Chen EYH et al (2002) Longitudinal profiles ofneurocognitive function in first-episode psychosis. SchizophrRes 53:121
Verma A, Moghaddam B (1996) NMDA receptor antagonists im-pair prefrontal cortex function as assessed via spatial delayedalternation performance in rats: modulation by dopamine. JNeurosci 16:373–379
Vincent JP, Kartalovski B, Geneste P et al (1979) Interaction ofphencyclidine (“angel dust”) with a specific receptor in ratbrain membranes. Proc Natl Acad Sci U S A 76:4678–4682
Volkow ND, Brodie JD, Wolf AP et al (1986) Brain metabolism inpatients with schizophrenia before and after acute neurolepticsadministration. J Neurol Neurosurg Psychiatry 49:1199–1202
Vollenweider FX, Leenders KL, Scharfetter C et al (1997) Metabolichyperfrontality and psychopathology in the ketamine model ofpsychosis using positron emission tomography (PET) and [18F]fluorodeoxyglucose (FDG). Eur Neuropsychopharmacol 7:9–24
Vollenweider FX, Vontobel P, Oye I et al (2000) Effects of (S)-ketamine on striatal dopamine: a [11C]raclopride PET study ofa model psychosis in humans. J Psychiatr Res 34:35–43
Williams GV, Goldman-Rakic PS (1995) Modulation of memoryfields by dopamine D1 receptors in prefrontal cortex. Nature6:572–575
Yurgelun-Todd D, Waternaux C, Cohen B et al (1996) Functionalmagnetic resonance imaging of schizophrenic patients duringword production. Am J Psychiatry 153:200–205
Zukin SR, Zukin RS (1979) Specific [3H]phencyclidine binding inrat central nervous system. Proc Natl Acad Sci U S A 76:5372–5376
102
















![Overt [-R] subjects in infinitival complements from Spanish and Italian as bound variables](https://static.fdokumen.com/doc/165x107/6321cf90807dc363600a25c9/overt-r-subjects-in-infinitival-complements-from-spanish-and-italian-as-bound.jpg)



