
Effects of Freezing, Thawing, and Storing Human Liver Microsomes on Cytochrome P450 Activity
25
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 331, No. 2, July 15, pp. 145–169, 1996 Article No. 0294 Effects of Freezing, Thawing, and Storing Human Liver Microsomes on Cytochrome P450 Activity Robin E. Pearce,* Christin J. McIntyre,* Ajay Madan,* Uma Sanzgiri,* Alison J. Draper,* Peter L. Bullock,* ,1 Dorn C. Cook,² L. Alayne Burton,‡ Jason Latham,‡ Cheryl Nevins,‡ and Andrew Parkinson* ,2 *Department of Pharmacology, Toxicology, and Therapeutics, Center for Environmental and Occupational Health, University of Kansas Medical Center, Kansas City, Kansas 66160-7417; ²Human Biologics Incorporated, P.O. Box 50862, Phoenix, Arizona 85076; and ‡XenoTech, L.L.C. 3800 Cambridge, Kansas City, Kansas 66103 Received February 7, 1996; and in revised form April 22, 1996 P450 enzymes. In study B, the concentration of cyto- chromes P450 and b 5 , the activity of NADPH–cyto- The stability of cytochrome P450 enzymes, cyto- chrome c reductase, and the activity of individual cyto- chrome b 5 , and NADPH–cytochrome c reductase was chrome P450 enzymes were determined in 10 samples examined in (A) human liver samples frozen in liquid of human liver microsomes stored at 0807C for approx- nitrogen and stored at 0807C, (B) human liver micro- imately 0, 1, or 2 years. The sample-to-sample variation somes suspended in 250 mM sucrose and stored at in the concentration and activity of cytochrome P450, 0807C, and (C) human liver microsomes subjected to cytochrome b 5 , and NADPH – cytochrome c reductase as many as 10 cycles of thawing and freezing. In study was nominally affected by long-term storage of human A, microsomes from five human livers were prepared liver microsomes at 0807C, indicating there was no dif- from fresh (unfrozen) tissue and from tissue that was ferential loss of cytochrome P450 activity, cytochrome stored frozen at 0807C for 1, 2, 4, or 6 months. The b 5 concentration, or NADPH – cytochrome c reductase apparent concentration of cytochromes P450 and b 5 activity. In study C, microsomes from a pool of human and the activity of NADPH – cytochrome c reductase livers were subjected to 1, 2, 3, 5, 7, or 10 cycles of decreased 20–40% as a result of freezing the liver, re- freezing at 0807C followed by thawing at room temper- gardless of whether the liver was stored for 1 or 6 ature. Freezing/thawing liver microsomes for up to 10 months. Similar decreases were observed in the activi- cycles did not convert cytochrome P450 to P420, nor ties of cytochrome P450 enzymes belonging to several did it cause significant loss of CYP1A2, CYP2A6, gene families, namely CYP1A2 (7-ethoxyresorufin O- CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4/5, or dealkylation and caffeine N3-demethylation), CYP2A6 CYP4A9/11 activity. Overall, these results suggest that (coumarin 7-hydroxylation), CYP2C9 (tolbutamide our current methods for storing and processing hu- methylhydroxylation), CYP2C19 (S-mephenytoin 4*- man liver are well suited to preserving microsomal hydroxylation), CYP2D6 (dextromethorphan O-de- P450 enzyme activity. q 1996 Academic Press, Inc. methylation), CYP2E1 (chlorzoxazone 6-hydroxyla- Key Words: cytochrome P450; human liver micro- tion), CYP3A4/5 (testosterone 6b-hydroxylation), and somes; stability. CYP4A9/11 (lauric acid 12-hydroxylation). Freezing human liver did not convert cytochrome P450 to its inactive form, cytochrome P420, but it increased the contamination of liver microsomes with hemoglobin Liver microsomes contain numerous P450 en- or other heme-containing proteins, which resulted in zymes that play an important role in the biotransfor- a uniform decrease in the specific activity of cyto- mation of xenobiotics, such as drugs and environ- chromes P450 and b 5 and in the specific activity of all mental chemicals. 3 Marked species differences in the function and regulation of P450 enzymes underscore 1 Present address: Phoenix International, Life Sciences Incorpo- rated, St. Laurent (Montreal), Quebec, H4R 2N6 Canada. 2 To whom correspondence should be addressed at Department of 3 Experimental Procedures are presented as a Miniprint Supple- Pharmacology, Toxicology, and Therapeutics, University of Kansas Medical Center, Kansas City, KS 66160-7417. Fax: (913) 588-7330. ment at the end of this paper. 145 0003-9861/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved. AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Effects of Freezing, Thawing, and Storing Human Liver Microsomes on Cytochrome P450 Activity

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 331, No. 2, July 15, pp. 145–169, 1996Article No. 0294
Effects of Freezing, Thawing, and Storing Human LiverMicrosomes on Cytochrome P450 Activity
Robin E. Pearce,* Christin J. McIntyre,* Ajay Madan,* Uma Sanzgiri,* Alison J. Draper,*Peter L. Bullock,*,1 Dorn C. Cook,† L. Alayne Burton,‡ Jason Latham,‡Cheryl Nevins,‡ and Andrew Parkinson*,2
*Department of Pharmacology, Toxicology, and Therapeutics, Center for Environmental and Occupational Health,University of Kansas Medical Center, Kansas City, Kansas 66160-7417; †Human Biologics Incorporated,P.O. Box 50862, Phoenix, Arizona 85076; and ‡XenoTech, L.L.C. 3800 Cambridge, Kansas City, Kansas 66103
Received February 7, 1996; and in revised form April 22, 1996
P450 enzymes. In study B, the concentration of cyto-chromes P450 and b5, the activity of NADPH–cyto-The stability of cytochrome P450 enzymes, cyto-chrome c reductase, and the activity of individual cyto-chrome b5, and NADPH–cytochrome c reductase waschrome P450 enzymes were determined in 10 samplesexamined in (A) human liver samples frozen in liquidof human liver microsomes stored at0807C for approx-nitrogen and stored at 0807C, (B) human liver micro-imately 0, 1, or 2 years. The sample-to-sample variationsomes suspended in 250 mM sucrose and stored atin the concentration and activity of cytochrome P450,0807C, and (C) human liver microsomes subjected tocytochrome b5, and NADPH–cytochrome c reductaseas many as 10 cycles of thawing and freezing. In studywas nominally affected by long-term storage of humanA, microsomes from five human livers were preparedliver microsomes at0807C, indicating there was no dif-from fresh (unfrozen) tissue and from tissue that wasferential loss of cytochrome P450 activity, cytochromestored frozen at 0807C for 1, 2, 4, or 6 months. Theb5 concentration, or NADPH–cytochrome c reductaseapparent concentration of cytochromes P450 and b5
activity. In study C, microsomes from a pool of humanand the activity of NADPH–cytochrome c reductaselivers were subjected to 1, 2, 3, 5, 7, or 10 cycles ofdecreased 20–40% as a result of freezing the liver, re-freezing at0807C followed by thawing at room temper-gardless of whether the liver was stored for 1 or 6ature. Freezing/thawing liver microsomes for up to 10months. Similar decreases were observed in the activi-cycles did not convert cytochrome P450 to P420, norties of cytochrome P450 enzymes belonging to severaldid it cause significant loss of CYP1A2, CYP2A6,gene families, namely CYP1A2 (7-ethoxyresorufin O-CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4/5, ordealkylation and caffeine N3-demethylation), CYP2A6CYP4A9/11 activity. Overall, these results suggest that(coumarin 7-hydroxylation), CYP2C9 (tolbutamideour current methods for storing and processing hu-methylhydroxylation), CYP2C19 (S-mephenytoin 4*-man liver are well suited to preserving microsomalhydroxylation), CYP2D6 (dextromethorphan O-de-P450 enzyme activity. q 1996 Academic Press, Inc.methylation), CYP2E1 (chlorzoxazone 6-hydroxyla-
Key Words: cytochrome P450; human liver micro-tion), CYP3A4/5 (testosterone 6b-hydroxylation), andsomes; stability.CYP4A9/11 (lauric acid 12-hydroxylation). Freezing
human liver did not convert cytochrome P450 to itsinactive form, cytochrome P420, but it increased thecontamination of liver microsomes with hemoglobin
Liver microsomes contain numerous P450 en-or other heme-containing proteins, which resulted inzymes that play an important role in the biotransfor-a uniform decrease in the specific activity of cyto-mation of xenobiotics, such as drugs and environ-chromes P450 and b5 and in the specific activity of allmental chemicals.3 Marked species differences in thefunction and regulation of P450 enzymes underscore1 Present address: Phoenix International, Life Sciences Incorpo-
rated, St. Laurent (Montreal), Quebec, H4R 2N6 Canada.2 To whom correspondence should be addressed at Department of
3 Experimental Procedures are presented as a Miniprint Supple-Pharmacology, Toxicology, and Therapeutics, University of KansasMedical Center, Kansas City, KS 66160-7417. Fax: (913) 588-7330. ment at the end of this paper.
1450003-9861/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

146 PEARCE ET AL.
TA
BL
EI
Info
rmat
ion
onH
um
anL
iver
Don
ors
and
Sam
ples Dat
ean
dti
me
of
Dru
g,al
coh
ol,
and
Liv
erL
iver
Fre
shor
mg
mic
roso
mal
Don
orS
exA
geR
ace
Cau
seof
deat
hto
bacc
ou
seC
ross
-cla
mp
deli
very
proc
essi
ng
froz
enli
ver
prot
ein
/gli
ver
1F
57C
auca
sian
Intr
acer
ebra
lbl
eed
No
alco
hol
use
,n
onsm
oker
5/17
/91
5/17
/91
5/17
/91
Fre
sh6.
605
:00
18:0
018
:00
2F
53C
auca
sian
Intr
acer
ebra
lbl
eed
Pen
toba
rbit
al8
hpr
ior
to6/
26/9
16/
27/9
18/
22/9
1F
roze
n20
.7li
ver
rem
oval
,n
oal
coh
ol15
:47
14:4
08:
30u
se,
smok
ed1
pack
/day
for
un
know
nn
um
ber
ofye
ars
3M
33C
auca
sian
Gu
nsh
otw
oun
dto
Hea
vyal
coh
olu
se,
7/03
/91
7/05
/91
8/21
/91
Fro
zen
10.3
the
hea
dn
onsm
oker
,co
cain
ean
d10
:00
17:0
08:
30m
arij
uan
au
se4
M41
Cau
casi
anG
un
shot
wou
nd
toN
oal
coh
olu
se,
non
smok
er8/
16/9
18/
17/9
18/
17/9
1F
resh
9.3
the
hea
d15
:25
11:0
011
:00
5M
42C
auca
sian
Su
bdu
ral
No
alco
hol
use
,n
onsm
oker
9/07
/91
9/08
/91
9/26
/91
Fro
zen
7.1
hem
atom
a22
:50
17:0
008
:00
6F
51C
auca
sian
Myo
card
ial
Occ
asio
nal
alco
hol
use
,9/
24/9
19/
25/9
19/
25/9
1F
resh
10.5
infa
rcti
on;
non
smok
er12
:30
10:0
010
:00
intr
acer
ebra
lbl
eed
7M
45C
auca
sian
Su
bdu
ral
blee
dH
eavy
alco
hol
use
,10
/05/
9110
/06/
9110
/22/
91F
resh
11.0
un
know
nsm
okin
g00
:50
09:0
08:
308
F50
Cau
casi
anIn
trac
ereb
ral
blee
dN
oal
coh
olu
se,
smok
ed2
10/2
0/91
10/2
1/91
10/2
1/91
Fre
sh9.
5pa
cks/
day
forú
10ye
ars
12:5
008
:00
08:0
09
F11
mon
ths
Cau
casi
anC
ereb
ral
edem
aN
oal
coh
olu
se,
non
smok
er10
/25/
9110
/26/
911/
08/9
2F
roze
n12
.920
:46
09:3
00:
800
10F
54H
ispa
nic
Su
bara
chn
oid
Mod
erat
eal
coh
olu
se(3
12/0
2/91
12/0
2/91
1/09
/92
Fro
zen
6.7
hem
mor
hag
edr
inks
/day
),n
onsm
oker
09:4
309
:43
0:80
011
M54
Afr
ican
-B
rain
inu
ury
–D
aily
alco
hol
inta
ke,
12/1
1/91
12/1
2/91
2/27
/92
Fro
zen
9.1
Am
eric
anm
ult
iple
trau
ma
non
smok
er15
:17
09:4
508
:00
12F
36C
auca
sian
Gu
nsh
otw
oun
dto
Mod
erat
eal
coh
ol,
1/01
/92
1/02
/92
2/26
/92
Fro
zen
8.3
the
hea
du
nkn
own
smok
ing
18:0
013
:30
08:0
013
F52
Cau
casi
anIn
trac
ereb
ral
blee
dO
ccas
ion
alal
coh
ol,
2/08
/92
2/09
/92
2/28
/92
Fro
zen
6.1
un
know
nsm
okin
g08
:56
08:0
008
:00
AID ARCH 9497 / 6b1b$$9497 06-21-96 08:09:43 arcas AP: Archives

147STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 1. Stability of human liver and human liver microsomes: Cytochrome b5. For each enzyme analyzed (Figs. 1–13), the graph in theupper left corner shows the effects of freezing and storing human liver at 0807C for up to 6 months (study A). Due to large sample-to-sample differences in P450 activities, the data (mean of five samples) are expressed as a percentage of the values determined with microsomesprepared from fresh human liver. The graph in the lower left corner shows the effects of storing 10 samples of human liver microsomes at0807C for 1 or 2 years on P450 activities (study B). The graph in the lower right corner depicts the correlation between P450 activitiesdetermined in 1992 and those determined 1 (1993) or 2 (1994) years later (study B). The graph in the upper right corner shows the effectsof freezing and thawing pooled human liver microsomes (liver sample Nos. 2, 3, 5, 6, and 11) for up to 10 cycles (study C). (In all graphs,bars without error bars depict the mean of duplicate determinations; bars with error bars depict the mean of three determinations { SD).Cytochrome b5 concentrations were determined in human liver microsomes prepared and stored as described under Experimental Procedures.
the importance of studying biotransformation reac- chrome P450 and other enzymes in the in vitro bio-transformation of xenobiotics (1–3). These studiestions in the primary species of interest, which for
most drugs and environmental chemicals is the hu- are generally carried out with microsomes preparedfrom the livers of brain-dead organ donors, whichman. Clinical (i.e., in vivo) studies of xenobiotic bio-
transformation in humans are subject to several con- cannot be transplanted because a recipient cannotbe identified or because the donor liver is deemedstraints (ethics, cost, time, etc.), for which reason
considerable emphasis has been placed on the devel- unsuitable (generally because the liver is fatty orcontains small blood clots that might increase theopment of in vitro test systems. Human liver micro-
somes are now one of the most widely used in vitro risk of thromboembolism or because the donor isolder than 65 years). These livers are neverthelesstest systems for characterizing the role of cyto-
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives
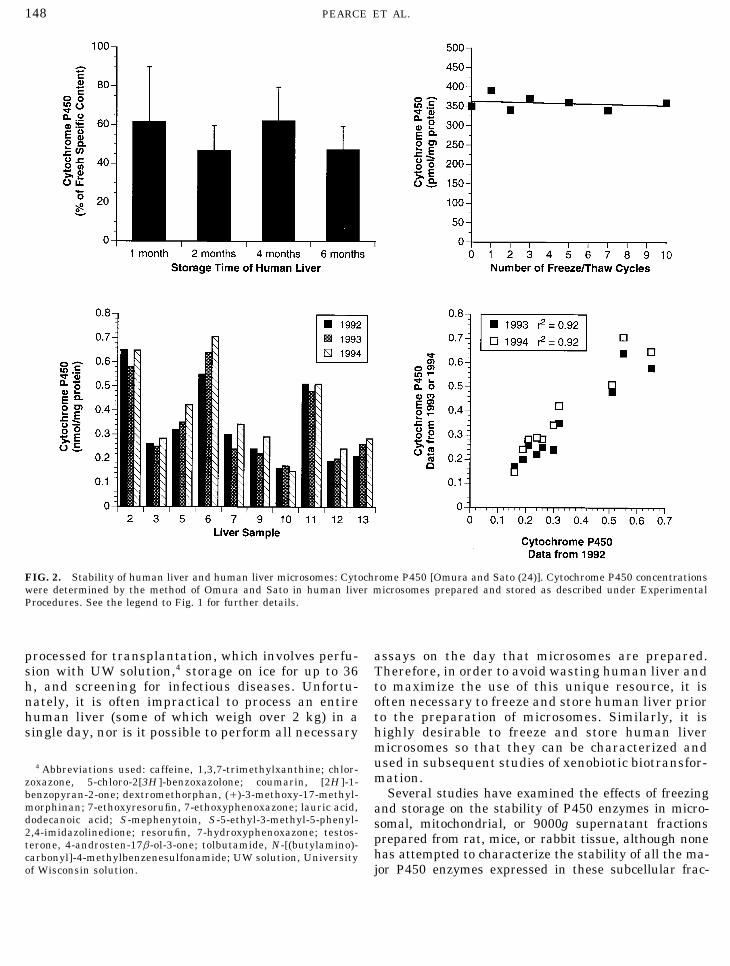
148 PEARCE ET AL.
FIG. 2. Stability of human liver and human liver microsomes: Cytochrome P450 [Omura and Sato (24)]. Cytochrome P450 concentrationswere determined by the method of Omura and Sato in human liver microsomes prepared and stored as described under ExperimentalProcedures. See the legend to Fig. 1 for further details.
processed for transplantation, which involves perfu- assays on the day that microsomes are prepared.Therefore, in order to avoid wasting human liver andsion with UW solution,4 storage on ice for up to 36
h, and screening for infectious diseases. Unfortu- to maximize the use of this unique resource, it isoften necessary to freeze and store human liver priornately, it is often impractical to process an entire
human liver (some of which weigh over 2 kg) in a to the preparation of microsomes. Similarly, it ishighly desirable to freeze and store human liversingle day, nor is it possible to perform all necessarymicrosomes so that they can be characterized andused in subsequent studies of xenobiotic biotransfor-4 Abbreviations used: caffeine, 1,3,7-trimethylxanthine; chlor-mation.zoxazone, 5-chloro-2[3H ]-benzoxazolone; coumarin, [2H ]-1-
Several studies have examined the effects of freezingbenzopyran-2-one; dextromethorphan, (/)-3-methoxy-17-methyl-morphinan; 7-ethoxyresorufin, 7-ethoxyphenoxazone; lauric acid, and storage on the stability of P450 enzymes in micro-dodecanoic acid; S-mephenytoin, S-5-ethyl-3-methyl-5-phenyl- somal, mitochondrial, or 9000g supernatant fractions2,4-imidazolinedione; resorufin, 7-hydroxyphenoxazone; testos- prepared from rat, mice, or rabbit tissue, although noneterone, 4-androsten-17b-ol-3-one; tolbutamide, N-[(butylamino)-
has attempted to characterize the stability of all the ma-carbonyl]-4-methylbenzenesulfonamide; UW solution, Universityof Wisconsin solution. jor P450 enzymes expressed in these subcellular frac-
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

149STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 3. Stability of human liver and human liver microsomes: Cytochrome P450 [Matsubara et al. (25)]. Cytochrome P450 concentrationswere determined by the method of Matsubara et al. in human liver microsomes prepared and stored as described under ExperimentalProcedures. See the legend to Fig. 1 for further details.
tions. About 30–70% of P450 activity is apparently lost 20), whereas others have reported no differential loss ofP450 activity (5, 13, 16, 19).when subcellular fractions are prepared from frozen tis-
sue compared with those prepared from fresh (unfrozen) The effects of freezing and long-term storage of humanliver on the activity of microsomal enzymes have receivedtissue (4–6). Cytochrome P450 activity is lost more rap-
idly from liver microsomes stored at or above freezing surprisingly little attention. Powis et al. (21) reportedthat P450 activities in human liver microsomes are differ-than from those stored at lower temperatures (6–8), a
phenomenon partially attributed to the conversion of cy- entially affected by cold storage, suggesting that someP450 enzymes are less stable than others to freezing andtochrome P450 into cytochrome P420 (9). Similarly, cyto-
chrome P450 activity is lost more rapidly from liver mi- storage. Although undesirable, differential loss of P450activity would not preclude the use of frozen human livercrosomes stored at0207C (6, 10–12) compared with those
stored at temperatures below 0707C (6, 13–20). Several microsomes for in vitro studies of xenobiotic biotransfor-mation provided the loss was gradual, limited in extent,investigators have reported that individual P450 en-
zymes lose their activity at different rates when liver and relatively constant from one sample to the next. Un-der these conditions the sample-to-sample variation inmicrosomes are frozen and stored (6, 10–12, 15, 17, 18,
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

150 PEARCE ET AL.
TABLE II
Regression Coefficients (r2) for the Sample-to-Sample Variation in the Levels of Cytochrome P450, Cytochrome b5, andEnzyme Activities in Human Liver Microsomes
7-EthoxyresorulinCytochrome P450 Cytochrome P450 NADPH–cytochrome O-dealkylation(Omura and Sato) (Matsubara et al.) Cytochrome b5 c reductase CYP1A2
Cytochrome P450 (Omura and Sato) 0.948 0.400 0.089 0.160Cytochrome P450 (Matsurbara et al.) 0.948 0.398 0.106 0.098Cytochrome b5 0.400 0.398 0.267 0.273NADPH–cytochrome c reductase 0.089 0.106 0.267 0.0017-Ethoxyresorufin O-dealkylation
CYP1A2 0.160 0.098 0.273 0.001Caffeine N3-demethylation CYP1A2 0.231 0.162 0.245 0.001 0.951Coumarin 7-hydroxylation CYP2A6 0.141 0.069 0.053 0.016 0.100Tolbutamide methylhydroxylation
CYP2C9 0.503 0.502 0.189 0.294 0.013S-Mephenytoin 4*-hydroxylation
CYP2C19 0.001 0.003 0.151 0.078 0.009Dextromethorphan O-demethylation
CYP2D6 0.008 0.010 0.171 0.402 0.084Chlorzoaxazone 6-hydroxylation
CYP2E1 0.375 0.369 0.071 0.101 0.002Testosterone 6b-hydroxylation
CYP3A4/5 0.808 0.866 0.301 0.266 0.016Lauric acid 12-hydroxylation
CYP4A9/11 0.288 0.203 0.107 0.209 0.009
Note. Values in bold are statistically significant at the 1% level of significance.
P450 enzyme activity would not vary appreciably over per-gram-of-liver basis varied approximately three-fold and ranged from 6.1 mg/g (sample 13) to 20.7time which would enable a bank of human liver micro-
somes to be used for reaction phenotyping by correlation mg/g (sample 2), as shown in Table I. The results ofthe initial measurements of cytochrome P450 andanalysis (1–3).
As there appears to be limited information on the sta- b5 concentration, NADPH–cytochrome c reductaseactivity, and P450 activities (discussed below) arebility of enzymes in general, and human P450 enzymes
in particular, we have undertaken studies to examine the represented by the solid black bars (data for 1992)in the lower left panels of Figs. 1–13.stability of cytochrome P450 enzymes in various gene
(sub)families, cytochrome b5, and NADPH–cytochrome cCytochrome P450, Cytochrome b5, and NADPH–reductase in (A) human liver samples frozen in liquid
Cytochrome c Reductasenitrogen and stored at0807C and (B) human liver micro-somes suspended in 250 mM sucrose and stored at0807C. The concentration of cytochrome b5 in the 10 samplesIn addition, we have examined whether P450 enzyme was determined by the method of Omura and Sato (24),activities are differentially lost when human liver micro- and the results are shown in Fig. 1. The samples con-somes are subjected to as many as 10 cycles of thawing tained between 0.24 and 0.43 nmol cytochrome b5/mgand freezing. protein, which is normal for human liver microsomes
(1, 45–47). The concentration of cytochrome P450 wasRESULTS determined by the method of Omura and Sato (24) and
by the method of Matsubara et al. (25). The resultsHuman Liver Microsomesare summarized in Figs. 2 and 3. The concentration of
The microsomes used in this study were prepared cytochrome P450 ranged from approximately 0.16 tofrom 13 human liver samples obtained from various 0.65 nmol/mg protein, which is normal for human liverorgan banks in 1991–1992.5 The available informa- microsomes (1, 45–47). There was excellent agreementtion on each of the donors is summarized in Table I. (r2Å 0.948) between the cytochrome P450 values deter-The amount of microsomal protein recovered on a mined by the method of Omura and Sato (24) and those
determined by the method of Matsubara et al. (25), as5 UNOS identification numbers are available upon request. shown in Table II. In contrast, there was no striking
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

151STABILITY OF HUMAN LIVER CYTOCHROME P450
TABLE II—Continued
Caffeine Coumarin Tobutamide S-Mephenytoin Dextromethorphan Chlorzoxazone Testosterone Lauric acidN3-demethylation 7-hydroxylation methylhydroxylation 4*-hydroxylation O-demethylation 6-hydroxylation 6b-hydroxylation 12-hydroxylation
CYP1A2 CYP2A6 CYP2C9 CYP2C19 CYP2D6 CYP2E1 CYP3A4/5 CYP4A9/11
0.231 0.141 0.503 0.001 0.008 0.375 0.808 0.2880.162 0.069 0.502 0.003 0.010 0.369 0.866 0.2030.245 0.053 0.189 0.151 0.171 0.071 0.301 0.1070.001 0.016 0.294 0.078 0.402 0.101 0.266 0.209
0.951 0.100 0.013 0.009 0.084 0.002 0.016 0.0090.147 0.003 0.006 0.128 0.001 0.047 0.007
0.147 0.001 0.317 0.001 0.132 0.082 0.146
0.003 0.001 0.014 0.160 0.521 0.602 0.396
0.006 0.317 0.014 0.167 0.101 0.002 0.202
0.128 0.001 0.160 0.167 0.011 0.095 0.127
0.001 0.132 0.521 0.101 0.011 0.286 0.194
0.047 0.082 0.602 0.002 0.095 0.286 0.371
0.007 0.146 0.396 0.202 0.127 0.194 0.371
correlation between the concentration of cytochrome are catalyzed by the same enzyme, namely CYP1A2. Theclose association between these two activities suggestsP450 and the concentration of cytochrome b5 (Table II).
The rate of cytochrome c reduction by the 10 samples that CYP1A1 contributes negligibly to the O-dealkylationof 7-ethoxyresorufin. This interpretation is consistentof human liver microsomes is shown in Fig. 4. NADPH–
cytochrome c reductase activity ranged from 111 to with the observation that human liver microsomes in-variably contain undetectable levels of CYP1A1 (1–3).232 nmol/mg protein/min, which can be considered a
normal range (1, 45–47). There were no striking corre- The sample-to-sample variation in 7-ethoxyresorufin O-dealkylase and caffeine N3-demethylase activity was notlations between NADPH–cytochrome c reductase ac-
tivity and the concentration of either cytochrome b5 or highly correlated with any other P450 enzyme activities(Table II).cytochrome P450 (Table II).
CYP2A6 ActivityCYP1A1/2 Activity Coumarin 7-hydroxylase activity has previously been
shown to be a marker for CYP2A6 activity in humansCYP1A2 catalyzes both the O-dealkylation of 7-ethox-(1–3, 30). The rate of coumarin 7-hydroxylation catalyzedyresorufin and the N3-demethylation of caffeine in hu-by the 10 samples of human liver microsomes varied 9-man liver microsomes (1–3). CYP1A1, which does notfold (120 to 1081 pmol/mg protein/min), as shown in Fig.appear to be expressed in human liver microsomes (1–3),7. This range of activity is comparable to the 17-fold sam-is as active as CYP1A2 at catalyzing the O-dealkylation ofple-to-sample variation in coumarin 7-hydroxylase activ-7-ethoxyresorufin, but is considerably less active (õ15%)ity and immunoreactive CYP2A6 levels previously re-at catalyzing the N3-demethylation of caffeine (51). Theported (30). The sample-to-sample variation was notrates of 7-ethoxyresorufin O-dealkylation and caffeinehighly correlated with any other P450 enzyme activitiesN3-demethylation catalyzed by the 10 samples of human(Table II).liver microsomes are shown in Figs. 5 and 6, respectively.
The rate of 7-ethoxyresorufin O-dealkylation varied 9- CYP2C9 Activityfold from sample-to-sample (10 to 95 pmol/mg protein/ Tolbutamide methylhydroxylation was measured tomin). The rate of caffeine N3-demethylation varied 27- determine the catalytic activity of CYP2C9 (49–51).6fold (7 to 189 pmol/mg protein/min). As shown in TableII, these two activities were highly correlated with one 6 Several allelic variants of CYP2C9 have been described, which
are generated by one or more amino acid substitutions at positionsanother (r2 Å 0.951), which suggests that both reactions
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

152 PEARCE ET AL.
FIG. 4. Stability of human liver and human liver microsomes: NADPH–cytochrome c reductase. NADPH–cytochrome c reductase activitieswere determined in human liver microsomes prepared and stored as described under Experimental Procedures. See the legend to Fig. 1for further details.
As shown in Fig. 8, the rate of conversion of tolbuta- samples of human liver microsomes (53, 54). The rateof S-mephenytoin 4*-hydroxylation varied 16-foldmide to hydroxymethyltolbutamide by the 10 samples
of human liver microsomes varied threefold from sam- from sample to sample (7 to 113 pmol/mg protein/min), as shown in Fig. 9. The sample-to-sample varia-ple to sample (103 to 275 pmol/mg protein/min) and
was highly correlated with testosterone 6b-hydroxy- tion in S-mephenytoin 4*-hydroxylation was nothighly correlated with any other P450 enzyme activi-lase (r2 Å 0.602) activity, but not with any other P450
enzyme activities (Table II). ties (Table II).
CYP2C19 Activity CYP2D6 ActivityS-Mephenytoin 4*-hydroxylase activity was mea- The activity of CYP2D6 present in the 10 samples of
sured to determine the activity of CYP2C19 in the 10 human liver microsomes was determined by the rateof dextromethorphan O-demethylation (55). The rateof dextromethorphan O-demethylation catalyzed by the144, 358, 359, and 417 (3). One of the allelic variants of CYP2C9
(Tyr358 and Gly417) is known as CYP2C10 (Cys358 and Asp417) (52). 10 samples of human liver microsomes varied 56-fold
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

153STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 5. Stability of human liver and human liver microsomes: 7-Ethoxyresorufin O-dealkylation (CYP1A2). 7-Ethoxyresorufin (10 mM)was incubated with human liver microsomes (0.1 mg/ml) for 2 min, as described under Experimental Procedures. See the legend to Fig. 1for further details.
(5 to 281 pmol/mg protein/min), as shown in Fig. 10. CYP3A4/5 ActivityThe sample-to-sample variation in dextromethorphan Testosterone 6b-hydroxylation was measured to de-O-demethylation was not highly correlated with any termine the catalytic activity of the CYP3A enzymesother P450 enzyme activities (Table II). in human liver microsomes (56).7 The rates of testoster-
one 6b-hydroxylation by the 10 samples of human liverCYP2E1 Activity microsomes are shown in Fig. 12. The rate of testoster-
one 6b-hydroxylation varied 15-fold from sample toThe rate of conversion of chlorzoxazone to 6-hydroxy-sample (861 to 12,500 pmol/mg protein/min). The sam-5-chloro-2[3H ]-benzoxazolone by the 10 samples of hu-ple-to-sample variation in testosterone 6b-hydroxyla-man liver microsomes was measured to determine thetion was highly correlated with total P450 concentra-catalytic activity of CYP2E1 (36). The rate of chlorzoxa-
zone 6-hydroxylation varied fivefold from sample tosample (1450 to 6860 pmol/mg protein/min), as shown 7 All human livers appear to express CYP3A4 (and/or its allelicin Fig. 11. The sample-to-sample variation in chlorzox- variant CYP3A3), whereas only 10–30% express CYP3A5 (57, 58).azone 6-hydroxylation was not highly correlated with CYP3A7 is considered a fetal enzyme, although it is expressed in
some adult livers (59).any other P450 enzyme activities (Table II).
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

154 PEARCE ET AL.
FIG. 6. Stability of human liver and human liver microsomes: Caffeine N3-dealkylation (CYP1A2). Caffeine (1 mM) was incubated withhuman liver microsomes (2.0 mg/ml) for 60 min, as described under Experimental Procedures. See the legend to Fig. 1 for further details.
tion (r2 Å 0.808, Omura and Sato, 1964; r2 Å 0.866, Stability of P450 Enzymes in Human Liver SamplesStored at 0807CMatsubara et al., 1976) and with tolbutamide methyl-
hydroxylase (r2 Å 0.602), but not with any other P450 Microsomes from five human livers were preparedenzyme activities (Table II). from fresh (unfrozen) tissue and from tissue that was
stored frozen at 0807C for 1, 2, 4, or 6 months. As aCYP4A9/11 Activity result of freezing the livers, the concentration of spec-
trally apparent cytochrome P450 decreased 40 to 60%The 12-hydroxylation of lauric acid was measuredto determine the catalytic activity of CYP4A9/11 in when measured by the method of Omura and Sato (24)
(Fig. 2, upper left panel), but decreased only 20 to 40%human liver microsomes (60, 61). The rate of lauricacid 12-hydroxylation varied fourfold in the 10 sam- when corrected for hemoglobin contamination by the
method of Matsubara et al. (25) (Fig. 3, upper leftples of human liver microsomes (310 to 1100 pmol/mg protein/min), as shown in Fig. 13. The sample-to- panel), regardless of whether the liver was stored for
1 or 6. A 20–35% decrease in the concentration of cyto-sample variation in lauric acid 12-hydroxylation wasnot highly correlated with any other P450 enzyme chrome b5 (Fig. 1, upper left panel) and a 30–35% de-
crease in the activity of NADPH–cytochrome c reduc-activities (Table II).
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

155STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 7. Stability of human liver and human liver microsomes: Coumarin 7-hydroxylation (CYP2A6). Coumarin (50 mM) was incubatedwith human liver microsomes (0.01 to 0.1 mg/ml) for 10 min, as described under Experimental Procedures. See the legend to Fig. 1 forfurther details.
tase (Fig. 4, upper left panel) were observed in micro- amined over a 2-year period. Although individual samplesexhibited slight variations between values determined insomes prepared from frozen liver compared with1992, 1993, and 1994, no significant changes in the concen-microsomes prepared from fresh liver. Decreases of thetration of spectrally apparent cytochrome P450 or cyto-same magnitude were observed in the activity of allchrome b5, the activity of NADPH–cytochrome c reduc-P450 enzymes measured, namely CYP1A2 (20–34%,tase, or the activity of P450 enzymes belonging to variousFigs. 5 and 6), CYP2A6 (25–36%, Fig. 7), CYP2C9 (26–gene (sub)families were observed as a result of storing38%, Fig. 8), CYP2C19 (25–44%, Fig. 9), CYP2D6 (26–human liver microsomes at0807C for up to 2 years (Figs.38%, Fig. 10), CYP2E1 (22–33%, Fig. 11), CYP3A4/51–13, lower left panels). There was excellent agreement(31–39%, Fig. 12), and CYP4A9/11 (27–36%, Fig. 13).between the cytochrome P450 concentrations determinedin 1992 and those determined in 1993 and 1994 by theStability of P450 Enzymes in Human Livermethod of Omura and Sato (24) (r2 Å 0.92 for both) andMicrosomes Stored up to 2 Years at 0807Cby the method of Matsubara et al. (25) (r2 Å 0.88 and
The stability of P450 enzymes present in microsomes 0.90, respectively), as shown in Figs. 2 and 3 (lower rightpanels), respectively. Similarly, the activities of CYP1A2,prepared from 10 different human liver samples was ex-
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

156 PEARCE ET AL.
FIG. 8. Stability of human liver and human liver microsomes: Tolbutamide methylhydroxylation (CYP2C9). Tolbutamide (1 mM) wasincubated with human liver microsomes (1.0 mg/ml) for 30 min, as described under Experimental Procedures. See the legend to Fig. 1 forfurther details.
2A6, 2C9, 2C19, 2D6, 2E1, and 3A4/5 determined in 1992 late significantly with the activity of CYP4A9/11 deter-mined in 1992 (r2Å 0.63), as shown in Fig. 13 (lower rightwere highly correlated (r2 Å 0.82 to 0.99) with the activi-
ties of these same enzymes determined in 1993 and 1994, panel), and was highly correlated (r2 Å 0.82) with theactivity of CYP4A9/11 determined in 1993. In some cases,as shown in Figs. 5–12 (lower right panels). The concen-
tration of cytochrome b5 and the activity of NADPH–cyto- the values determined in one year were systematicallyhigher or lower than those determined in another year,chrome c reductase determined in 1993 and 1994 were
significantly correlated with their respective values deter- the reason for which is discussed later.mined in 1992 (r2 Å 0.65 to 0.74) (Figs. 1 and 4, lowerright panels), but produced a high correlation between the Stability of P450 Enzymes in Human Livervalues obtained in 1993 and 1994 (r2 Å 0.88 and 0.97, Microsomes Subjected to Freeze/Thawing Cyclesrespectively). The sample-to-sample variation in the activ-ity of CYP4A9/11 determined in 1993 was not significantly The upper right panels of Figs. 1–13 show the effects
of subjecting microsomes pooled from five human sam-correlated with the activity of CYP4A9/11 determined in1992 (r2Å 0.53). However, the sample-to-sample variation ples to 1, 2, 3, 5, 7, or 10 cycles of freezing at 0807C
followed by thawing at room temperature on the con-in the activity of CYP4A9/11 determined in 1994 did corre-
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

157STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 9. Stability of human liver and human liver microsomes: S-Mephenytoin 4*-hydroxylation (CYP2C19). S-Mephenytoin (400 mM) wasincubated with human liver microsomes (2.0 mg/ml) for 30 min, as described under Experimental Procedures. See the legend to Fig. 1 forfurther details.
centration and activity of cytochrome P450, the concen- NADPH–cytochrome c reductase at three levels: (A) inhuman liver samples frozen in liquid nitrogen andtration of cytochrome b5, and the activity of NADPH–
cytochrome c reductase. Freezing and thawing human stored at 0807C, (B) in human liver microsomes sus-pended in 250 mM sucrose and stored at 0807C, andliver microsomes for up to 10 cycles caused no detect-
able loss of spectrally apparent cytochrome P450, nor (C) in human liver microsomes subjected to as manyas 10 cycles of thawing and freezing.did it convert cytochrome P450 to P420, the catalyti-
cally inactive form of cytochrome P450. Freezing and When microsomes were prepared from frozen liver,the concentration and activity of all P450 enzymes uni-thawing human liver microsomes for up to 10 cycles
caused no detectable loss of cytochrome b5 or NADPH– formly decreased by Ç30% compared with the concen-tration and activity of these enzymes in microsomescytochrome c reductase activity, nor did it cause sig-
nificant loss of activity of CYP1A1, 2A6, 2C9, 2C19, prepared from fresh (unfrozen) tissue, regardless ofhow long the frozen liver was stored at 0807C (1 to 62D6, 2E1, 3A4/5, or 4A9/11.months). These results could be interpreted as evidence
DISCUSSION that freezing human liver causes a partial inactivationof all microsomal P450 enzymes, or they could be inter-This study was undertaken to determine the stability
of cytochrome P450 enzymes, cytochrome b5, and preted as evidence that microsomes prepared from fro-
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

158 PEARCE ET AL.
FIG. 10. Stability of human liver and human liver microsomes: Dextromethorphan O-demethylation (CYP2D6). Dextromethorphan (80mM) was incubated with human liver microsomes (1.0 mg/ml) for 30 min, as described under Experimental Procedures. See the legend toFig. 1 for further details.
zen liver are contaminated with additional protein, When the concentration of cytochrome P450 was de-termined by the method of Omura and Sato (24), allwhich decreases the specific activity of all microsomal
enzymes. Several results support the latter interpreta- microsomal samples contained a 420-nm absorbingchromophore, regardless of whether the microsomestion. First, freezing human liver prior to isolating mi-
crosomes caused a 20–40% decrease in the concentra- were prepared from frozen or fresh liver. The concen-tration of this 420-nm absorbing chromophore variedtion of cytochrome b5 (Fig. 1) and the activity of
NADPH–cytochrome c reductase (Fig. 4) and carboxyl- from sample to sample and was highly correlated (r2 Å0.99) with the degree to which the microsomal samplesesterase (results not shown). It seems unlikely that
these diverse microsomal enzymes would all be inacti- were contaminated with hemoglobin (or other ferrousheme-containing proteins), as shown in Fig. 14. Fur-vated to the same extent by freezing human liver. Sec-
ond, freezing human liver did not increase the concen- thermore, the concentration of the 420-nm absorbingchromophore was low or undetectable when the concen-tration of cytochrome P420, the inactive form of cyto-
chrome P450, although this finding requires further tration of cytochrome P450 was determined by themethod of Matsubara et al. (25), which compensatesexplanation.
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

159STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 11. Stability of human liver and human liver microsomes: Chlorzoxazone 6-hydroxylation (CYP2E1). Chlorzoxazone (500 mM) wasincubated with human liver microsomes (0.4 mg/ml) for 30 min, as described under Experimental Procedures. See the legend to Fig. 1 forfurther details.
for the presence of hemoglobin. These results suggest tinguish cytochrome P420 from methemoglobin, whichis likely to be present in microsomes contaminated withthat the liver microsomes used in this study contained
little or no cytochrome P420, although they were con- hemoglobin. These results suggest that little or no deg-radation of cytochrome P450 occurred by freezing andtaminated to varying degrees with hemoglobin or some
other 420-nm absorbing chromophore. storing human liver at 0807C, but caused hemoglobinand perhaps other additional proteins (disrupted fromFigure 15 shows hemoglobin and cytochrome P450
spectra determined by the method of Matsubara et al. subcellular organelles and blood) to be recovered in themicrosomal fraction. As shown in Table III, 30–40%(25) with microsomes prepared from fresh and frozen
liver. Liver microsomes from fresh liver contained low more protein was recovered per gram of human liverin microsomes prepared from frozen tissue, regardlesslevels of hemoglobin and little or no cytochrome P420
(Fig. 15, spectrum A). Freezing human liver increased of the length of storage at0807C, compared with micro-somes prepared from fresh tissue. This additional pro-the concentration of hemoglobin in microsomes (Fig.
15, spectrum C). It also appeared to increase the con- tein can account for the uniform decrease in the specificcontent of cytochromes P450 and b5 and in the specificcentration of cytochrome P420, although it must be
recalled that Matsubara et al.’s (25) method cannot dis- activity of all P450 enzymes and NADPH–cytochrome
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

160 PEARCE ET AL.
FIG. 12. Stability of human liver and human liver microsomes: Testosterone 6b-hydroxylation (CYP3A4/5). Testosterone (250 mM) wasincubated with human liver microsomes (0.2 mg/ml) for 8 min, as described under Experimental Procedures. See the legend to Fig. 1 forfurther details.
c reductase in liver microsomes prepared from frozen bin contamination by incubating liver microsomes withalbumin and dithioerythreitol, but without success (re-human liver when compared with liver microsomes
prepared from fresh liver. sults not shown). We have recently determined thatthis same phenomenon occurs when rat liver is frozenThe additional protein associated with microsomes
prepared from frozen liver does not detract from the prior to the preparation of microsomes (results notshown).usefulness of these liver microsomes. These additional
proteins do not appear to alter the stability of P450 The observation that liver microsomes prepared fromfrozen liver have uniformly lower P450 enzyme activi-enzymes during long-term storage of liver microsomes.
Because it is often impractical to process an entire hu- ties than those prepared from fresh liver has practicalimplications. If several batches of microsomes are toman liver (which may weigh over 2 kg) in a single day,
freezing human liver prior to the preparation of micro- be prepared from a single liver, batch-to-batch varia-tion can be minimized by freezing the entire liver, sosomes is virtually unavoidable. Contamination of mi-
crosomes with hemoglobin and other proteins may also that all batches of microsomes undergo a dilution ofP450 enzymes by hemoglobin and other proteins thatbe unavoidable. We have attempted to reduce hemoglo-
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

161STABILITY OF HUMAN LIVER CYTOCHROME P450
FIG. 13. Stability of human liver and human liver microsomes: Lauric acid 12-hydroxylation (CYP4A9/11). [14C]Lauric acid (100 mM, 50nCi/ml) was incubated with human liver microsomes (0.4 mg/ml) for 10 min, as described under Experimental Procedures. See the legendto Fig. 1 for further details.
associate with the microsomal membranes after human 2C8, 2C19, 2E1, or CYP3A4/5 (30 and unpublished re-sults). In some cases, the values determined in one yearliver is frozen.
Storage of liver microsomes at 0807C for up to 2 were consistently higher or lower than those deter-mined in another year. In most cases, these differencesyears had little or no effect on the concentration of
cytochrome b5, the concentration of cytochrome P450, could be attributed to year-by-year fluctuations in stan-dard values, which artificially introduce systematic in-or the activity of NADPH–cytochrome c reductase and
P450 enzymes belonging to various gene (sub)families creases or decreases in enzymatic rates. For example,the rates of 7-ethoxyresorufin O-dealkylation deter-(Figs. 1–13, lower left panel). The sample-to-sample
variation in P450 enzyme activity was nominally af- mined in 1993 were consistently higher than those de-termined in 1992 or 1994, which were comparable tofected by long-term storage of human liver microsomes,
indicating that there was no differential loss of P450 each other. We ascribe this variation in CYP1A2 activ-ity to experimental error because there was no corre-enzyme activity. In addition, proteolytic fragments of
P450 enzymes were not detected on Western immu- sponding change in caffeine N3-demethylation, whichis also catalyzed by CYP1A2. Furthermore, it seemsnoblots of human liver microsomes probed with various
polyclonal antibodies that recognize CYP2A6, 1A2, unlikely that P450 enzyme activity will increase when
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

162 PEARCE ET AL.
FIG. 14. Apparent concentration of a 420-nm absorbing chromo-phore and hemoglobin in human liver microsomes. The concentrationof a 420-nm absorbing chromophore in human liver microsomes wasdetermined from the difference spectrum of cytochrome P450 ac-cording to the method of Omura and Sato (24). The concentration ofhemoglobin was determined as described under Experimental Proce-dures.
human liver microsomes are stored for 1 year at0807C.FIG. 15. Cytochrome P450 and hemoglobin difference spectra ob-
NADPH–cytochrome c reductase activity determined tained with human liver microsomes. Cytochrome P450 and hemo-in 1993 was also consistently higher than that deter- globin difference spectra of microsomes prepared from fresh (B and
A, respectively) or frozen human liver stored at 0807C for 1 monthmined in 1992 or 1994. In this case, the temperature(sample 6) (D and C, respectively) determined according to theof the reaction may be the cause of a systematic errormethod of Matsubara et al. (25), as described under Experimentalbecause the reactions were carried out at room temper-Procedures. Similar cytochrome P450 and hemoglobin difference
ature, which was not recorded and may have varied spectra were also obtained with microsomes prepared from fresh orconsiderably between the annual measurements of frozen liver samples Nos. 1, 4, 7, and 8.NADPH–cytochrome c reductase activity. Inasmuch asthese annual variations were systematic in nature, thesample-to-sample variation in enzyme activity was well differential loss of P450 enzyme activity from micro-preserved over the 2-year period. somes stored in the EDTA-free buffer. These results
Several laboratories have reported differential loss suggests that EDTA improves the stability of P450 en-of P450 enzyme activity from microsomes stored at0807C (6, 10–12, 15, 17), which contrasts with the re-sults of this study. Our results are supported by two
TABLE IIIstudies that demonstrated uniform stability of rat orAmount of Protein Recovered in the Microsomal Fractionrabbit cytochrome P450 activity in microsomes pre-
from Fresh (Unfrozen) Human Liver and from Humanpared from both hepatic and extrahepatic tissues andLiver Stored Frozen at 0807C for 1, 2, 4, or 6 Monthsstored at 080 or 01967C for up to 2.5 months (5, 13).
In both of these studies, as in our study, microsomesRecovery (mg microsomal protein per gram of liver)were prepared or suspended in a buffer containing
EDTA, which does not appear to have been used in the Sample Fresh 1 Month 2 Months 4 Months 6 Monthsstudies reporting differential loss of microsomal P450
1 6.6 12.5 11.2 11.0 12.6activity. Tredger and Chhabra compared several P450-4 9.3 11.9 11.9 14.0 13.2catalyzed reactions in rat liver microsomes that were6 10.5 16.7 16.7 16.7 16.0prepared with and without EDTA in the microsomal7 11.0 12.7 12.3 12.8 12.3
suspension buffer and subsequently stored in liquid 8 9.5 14.2 14.1 11.6 13.3nitrogen (01967C) for 28 days (5). They reported uni-
Mean 9.4 13.6 13.2 13.2 13.5form stability of P450 enzymes in microsomes sus-SD {1.7 {1.9 {2.2 {2.3 {1.5
pended in the EDTA-containing buffer, but observed
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

163STABILITY OF HUMAN LIVER CYTOCHROME P450
zymes in liver microsomes, perhaps by chelating cal- such as cytochrome b5, CYP2C9, CYP2E1, CYP4A9/11,and carboxylesterase, show limited sample-to-samplecium and thereby inhibiting calcium-dependent prote-
ases and phospholipases. variation, even though the time between procuring andprocessing human liver varies considerably from oneThe stability of P450 enzymes during repeated freez-
ing and thawing of human liver microsomes was an sample to the next. If P450 enzyme activities do declineafter the livers are surgically removed, it appears thatunexpected observation. It was anticipated that sub-
jecting liver microsomes to a large number of freeze/ this process is completely halted by freezing the liveror by isolating microsomes.thaw cycles (up to 10) would provide some insight into
the relative instability of the major P450 enzymes inhuman liver microsomes. The fact that none of the ACKNOWLEDGMENTSP450 enzymes examined lost activity through 10 cycles
This work was supported by Grants ES03765 and GM37044, andof freezing and thawing should extend the usefulness by Training Grant ES07079, from the National Institutes of Health.of human liver microsomes for in vitro studies of xeno-biotic biotransformation. However, it is important to REFERENCESrecognize that our result do not preclude the possibility
1. Wrighton, S. A., Vandenbranden, M., Stevens, J. C., Shipley,that other enzymes, including P450 enzymes not mea-L. A., Ring, B. J., Rettie, A. E., and Cashman, J. R. (1993) Drugsured in this study, might be inactivated by repeated Metab. Rev. 25, 453–484.
freezing and thawing of human liver microsomes. 2. Guengerich, F. P. (1989) Annu. Rev. Pharmacol. Toxicol. 29,The observation that human liver microsomes can 241–264.
be stored at 0807C for 2 years with little or no loss of 3. Parkinson, A. (1995) in Casarett and Doull’s Toxicology. TheP450 enzyme activity needs to be extrapolated with Basic Science of Poisons (Klaassen, C. D., Ed.), 5th ed., pp. 113–
186, McGraw-Hill, New York.caution. First, our results should not be interpreted as4. Reinke, L. A., and Stohs, S. J. (1976) Res. Commun. Chem. Pa-evidence that all microsomal samples and all micro-
thol. Pharmacol. 14, 295–303.somal enzymes are stable when stored at 0807C for5. Tredger, J. M., and Chhabra, R. S. (1976) Drug Metab. Dispos.this length of time. It is possible that the method of
4, 451–459.preparing human liver microsomes influences P4506. Bartosek, I., Dolfini, E., Ghersa, P., Guaitani, A., Villa, P., andstability, as the study by Tredger and Chhabra sug- Villa, S. (1980) J. Pharmacol. Methods 3, 191–200.
gests (5). Therefore, P450 enzymes may not be uni- 7. Leadbeater, L., and Davies, D. R. (1964) Biochem. Pharmacol.formly stable in human liver microsomes prepared by 13, 1607–1617.another procedure, nor can it be assumed that the pro- 8. Wade, A. E., Wu, B., and Greene, F. E. (1972) Toxicol. Appl.cedures described in this study will uniformly preserve Pharmacol. 22, 503–512.P450 enzyme activities in liver microsomes from other 9. Hewick, D. S., and Fouts, J. R. (1970) Biochem. Pharmacol. 19,
457–472.species, although limited studies with rats, mice, dogs,10. Levin, W., Alvares, A., Jacobson, M., and Kuntzman, R. (1969)and cynomolgus monkeys suggest this will be the case
Biochem. Pharmacol. 18, 883–889.(unpublished results). It is also possible that our proce-11. Crankshaw, D. L., Erikson, R. R., and Holtzman, J. L. (1990)dures for preparing and storing human liver micro-
Biochem. Pharmacol. 39, 393–396.somes fail to preserve the activity of some of the P45012. Litterst, C. L., Mimnaugh, E. G., Regan, R. L., and Gram, T. E.enzymes not measured in this study (such as CYP2B6,
(1974) Biochem. Pharmacol. 23, 2391–2394.CYP2C8, and CYP2C18), as well as other xenobiotic-
13. Danner-Rabovsky, J., and Groseclose, R. D. (1982) J. Toxicol.biotransforming enzymes (such as the notoriously la- Environ. Health 10, 601–611.bile flavin-containing monooxygenases). The results of 14. Bode, C., and Meinel, A. (1982) J. Clin. Chem. Clin. Biochem.this study raise the possibility that intact human liver 20, 9–13.and isolated microsomes may be stable indefinitely 15. Kim, S. Y., and Park, K. H. (1988) Korean Biochem. J. 21, 505–
511.when stored at 0807C, although long-term studies willbe required to test this possibility. Finally, this study 16. Ohmiya, Y., and Mehendale, H. M. (1984) Jpn. J. Pharmacol.
36, 256–257.focuses on the stability of microsomal P450 enzymes17. Bulger, W. H., and Stohs, S. J. (1975) Res. Commun. Chem.in human liver only from the time the liver was re-
Pathol. Pharmacol. 11, 467–474.ceived and processed and does not address whether18. Tauc, M., Hermann, M., Dansette, P. M., and Vandecasteele,significant changes occurred between the time the liver
J. P. (1984) Mut. Res. 125, 123–133.was procured and processed. However, it is noteworthy19. Hubbard, S. A., and Davis, P. J. B. (1990) Mutagenesis 5, 7–8.that, in general, the P450 enzymes in human liver mi-20. Bauer, C., Corsi, C., and Paolini, M. (1994) Teratogen. Carcino-crosomes vary independently from one another, which gen. Mutagen. 14, 13–22.
argues against a uniform loss of P450 enzyme activity 21. Powis, G., Jardine, I., Van Dyke, R., Weinshilboum, R., Moore,during the time the livers are in transit from surgery to D., Wilke, T., Rhodes, W., Nelson, R., Benson, L., and Szumlan-
ski, C. (1988) Drug Metab. Dispos. 16, 582–589.the laboratory. Furthermore, some enzyme activities,
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

164 PEARCE ET AL.
22. Lu, A. Y. H., and Levin, W. (1972) Biochem. Biophys. Res. Com- 43. Romano, M. C., Straub, K. M., Yodis, L. A. P., Eckardt, R. D.,and Newton, J. F. (1988) Anal. Biochem. 170, 83–93.mun. 46, 1334–1339.
44. Bruning, J. L., and Kintz, B. L. (1987) in Computational Hand-23. BCA Protein Assay Reagent (1991) in Technical Bulletin 23225Xbook of Statistics, 3rd ed., pp. 176–180, Harper Collins, IL.from Pierce Chemical Co., pp. 1–16.
45. Shimada, T., Yamazaki, H., Mimura, M., Inui, Y., and Guenger-24. Omura, T., and Sato, R. (1964) J. Biol. Chem. 239, 2379–2385.ich, F. P. (1994) J. Pharmacol. Exp. Ther. 270, 414–423.
25. Matsubara, T., Koike, M., Touchi, A., Tochino, Y., and Sugeno,46. Guengerich, F. P., and Turvy, C. G. (1991) J. Pharmacol. Exp.K. (1976) Anal. Biochem. 75, 596–603.
Ther. 256, 1189–1194.26. Guengerich, F. P. (1989) in Principles and Methods of Toxicology, 47. Forrester, L. M., Henderson, C. J., Glancey, M. J., Back, D. J.,
2nd ed., pp. 777–814, Raven Press, New York. Park, K. B., Ball, S. E., Kitteringham, N. R., McLaren, A. W.,27. Phillips, A. H., and Langdon, R. G. (1962) J. Biol. Chem. 237, Miles, J. S., Skett, P., and Wolf, C. R. (1992) Biochem. J. 281,
2652–2660. 359–368.28. Burke, M. D., and Mayer, R. T. (1974) Drug Metab. Dispos. 2, 48. Gu, L., Gonzalez, F. J., Kalow, W., and Tang, B. K. (1992) Phar-
583–588. macogenetics 2, 73–77.49. Brian, W. R., Srivastava, P. K., Umbenhauer, D. R., and Lloyd,29. Dutton, D. R., and Parkinson, A. (1989) Arch. Biochem. Biophys.
R. (1989) Biochemistry 28, 4993–4999.268, 617–629.50. Srivastava, P. K., Yun, C-H., Beaune, P. H., Ged, C., and Gueng-30. Pearce, R., Greenway, D., and Parkinson, A. (1992) Arch. Bio-
erich, F. P. (1991) Mol. Pharmacol. 40, 69–79.chem. Biophys. 298, 211–225.51. Relling, M. V., Aoyama, T., Gonzalez, F. J., and Meyer, U. A.31. Greenlee, W. F., and Poland, A. (1978) J. Pharmacol. Exp. Ther.
(1990) J. Pharmacol. Exp. Ther. 252, 442–447.205, 596–605.52. Sandhu, P., Baba, T., and Guengerich, F. P. (1993) Arch. Bio-32. Naline, E., Flouvat, B., Advenier, C., and Pays, M. (1987) J.
chem. Biophys. 306, 443–450.Chromatogr. 419, 177–189.53. Wrighton, S. A., Stevens, J. C., Becker, G. W., and VandenBran-
33. Meier, U. T., Dayer, P., Male, P. J., Kronbach, T., and Meyer, den, M. (1993) Arch. Biochem. Biophys. 306, 240–245.U. R. (1985) Clin. Pharmacol. Ther. 38, 488–494.
54. Goldstein, J. A., Faletto, M. B., Romkes-Sparks, M., Sullivan,34. Knodell, R. G., Hall, S. D., Wilkinson, G. R., and Guengerich, T., Kitareewan, S., Raucy, J. L., Lasker, J. M., and Ghanayem,
F. P. (1987) J. Pharmacol. Exp. Ther. 241, 1112–1119. B. I. (1994) Biochemistry 33, 1743–1752.35. Kronbach, T. (1991) Methods Enzymol. 206, 509–517. 55. Schmid, B., Bircher, J., Preisig, R., and Kupfer, A. (1985) Clin.
Pharmacol. Ther. 38, 618–624.36. Peter, R., Bocker, R., Beaune, P. H., Iwasaki, M., Guengerich,F. P., and Yang, C. S. (1990) Chem. Res. Toxicol. 3, 566–573. 56. Waxman, D. J., Attisano, C., Guengerich, F. P., and Lapenson,
D. P. (1988) Arch. Biochem. Biophys. 263, 424–436.37. Wood, A. W., Ryan, D. E., Thomas, P. E., and Levin, W. (1983)57. Wrighton, S. A., Brian, W. R., Sari, M-A., Iwasaki, M., Guenger-J. Biol. Chem. 258, 8839–8847.
ich, F. P., Raucy, J. L., Molowa, D. T., and VandenBranden, M.38. Sonderfan, A. J., Arlotto, M. P., Dutton, D. R., McMillen, S. K.,(1990) Mol. Pharmacol. 38, 207–213.and Parkinson, A. (1987) Arch. Biochem. Biophys. 255, 27–41.
58. Wrighton, S. A., Ring, B. J., Watkins, P. B., and VandenBranden,39. Sonderfan, A. J., and Parkinson, A. (1988) Arch. Biochem. Bio- M. (1989). Mol. Pharmacol. 36, 97–105.
phys. 265, 208–218.59. Wrighton, S. A., and VandenBranden, M. (1989) Arch. Biochem.
40. Arlotto, M. P., Greenway, D. J., and Parkinson, A. (1989) Arch. Biophys. 268, 144–151.Biochem. Biophys. 270, 441–457. 60. Dirven, H. A. M., Peters, J. G. P., Gibson, G. G., Peters,
41. Smith, S., Korzekwa, K. R., Aoyama, T., Gonzalez, F. J., Dar- W. H. M., and Jongeneelen, F. J. (1991) Biochem. Pharmacol.byshire, J. F., Sugiyama, K., and Gillette, J. R. (1992) Drug 42, 1841–1844.Metab. Dispos. 20, 566–571. 61. Kawashima, H., Kusunose, E., Kikuta, Y., Kinoshita, H., Ta-
naka, S., Yamamoto, S., Kishimoto, T., and Kusunose, M. (1994)42. Giera, D. D., and van Lier, R. B. (1991) Fundam. Appl. Toxicol.16, 348–355. J. Biochem. (Tokyo) 116, 74–80.
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

165STABILITY OF HUMAN LIVER CYTOCHROME P450
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

166 PEARCE ET AL.
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

167STABILITY OF HUMAN LIVER CYTOCHROME P450
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives

168 PEARCE ET AL.

169STABILITY OF HUMAN LIVER CYTOCHROME P450
AID ARCH 9497 / 6b1b$$$261 06-21-96 08:09:43 arcas AP: Archives




















