
Collagen synthesis in an established liver connective tissue cell line (GRX) during induction of the...
11
Int. J. Exp. Path. (I992) 73, 125-I35 Effects of retinol on proliferation, cell adherence and extracellular matrix synthesis in a liver myofibroblast or lipocyte cell line (GRX) Rogerio Margis*, Maircia Pinheiro-Margis, Luiz-Claiudio F. da Silva and Radovan Borojevic Departamento de Bioquimica, Instituto de Quimica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil Received for publication 9 April I99I Accepted for publication 20 September I99I Summary. We have studied the effect of retinol on an established murine cell line (GRX), representative of liver connective tissue cells. This cell line has myofibroblast characteristics; under retinol treatment it is induced into the lipocyte (Ito-cell) phenotype. Retinol decreased the proliferation rate in the entire cell population. It increased cell adherence to the substrate, which was correlated with the increased secretion of fibronectin. Collagen secretion was specifically decreased, whilst the total protein secretion remained stable. Heparan sulphate was decreased in the pericellular compartment, but other glycosaminoglycans were not affected by retinol treatment. Modulations of pericellular components induced by retinol may alter the relations among liver mesenchymal cells, and may be related to vitamin-A-induced modifications of the homoeostasis of hepatic connective tissue and hepatic fibrosis. Keywords: retinol, lipocyte, Ito-cell, myofibroblast, collagen, glycosaminoglycan, fibronectin, hepatic fibrosis In normal liver, the ratio between connective tissue cells and hepatocytes is low. However, many chronic injuries of hepatic tissue lead to the formation of extensive fibrosis, with large increases in both connective tissue cell populations and extracellular matrix. Acti- vation of liver connective tissue cells (LCTC) is thus associated, and probably causally related, with the development of hepatic fibrosis. The two major connective tissue cell popu- lations of hepatic lobules are described as peri-sinusoidal cells with myofibroblastic characteristics and lipocytes (fat-storing cells, Ito-cells). They are frequently consi- dered to be two phenotypes of a single hepatic connective tissue cell lineage (Biou- lac-Sage & Balabaud I985). Hepatic lipo- cytes were observed to be associated with collagen fibres in normal and fibrotic hepatic parenchyma (Kent et al. 1 9 76). In an in-vivo study of lipocytes, Clement et al. (I 984) have demonstrated synthesis of extracellular matrix molecules, involving fibronectin and collagen types I, III and IV. In-vitro studies have also indicated that lipocytes are one of * Present address: Escola Tcnica Federal de Quimica, Rio de Janeiro, Brazil. Correspondence: Professor Radovan Borojevic, Caixa Postal 68021, 2I944 Rio de Janeiro, Brazil. 125
Transcript of Collagen synthesis in an established liver connective tissue cell line (GRX) during induction of the...

Int. J. Exp. Path. (I992) 73, 125-I35
Effects of retinol on proliferation, cell adherence andextracellular matrix synthesis in a liver myofibroblast or
lipocyte cell line (GRX)
Rogerio Margis*, Maircia Pinheiro-Margis, Luiz-Claiudio F. da Silva andRadovan Borojevic
Departamento de Bioquimica, Instituto de Quimica, Universidade Federal do Rio de Janeiro,Rio de Janeiro, Brazil
Received for publication 9 April I99IAccepted for publication 20 September I99I
Summary. We have studied the effect of retinol on an established murine cell line (GRX),representative of liver connective tissue cells. This cell line has myofibroblast characteristics;under retinol treatment it is induced into the lipocyte (Ito-cell) phenotype. Retinol decreasedthe proliferation rate in the entire cell population. It increased cell adherence to the substrate,which was correlated with the increased secretion of fibronectin. Collagen secretion wasspecifically decreased, whilst the total protein secretion remained stable. Heparan sulphatewas decreased in the pericellular compartment, but other glycosaminoglycans were notaffected by retinol treatment. Modulations of pericellular components induced by retinol mayalter the relations among liver mesenchymal cells, and may be related to vitamin-A-inducedmodifications of the homoeostasis of hepatic connective tissue and hepatic fibrosis.
Keywords: retinol, lipocyte, Ito-cell, myofibroblast, collagen, glycosaminoglycan, fibronectin,hepatic fibrosis
In normal liver, the ratio between connectivetissue cells and hepatocytes is low. However,many chronic injuries of hepatic tissue leadto the formation of extensive fibrosis, withlarge increases in both connective tissue cellpopulations and extracellular matrix. Acti-vation of liver connective tissue cells (LCTC)is thus associated, and probably causallyrelated, with the development of hepaticfibrosis.The two major connective tissue cell popu-
lations of hepatic lobules are described asperi-sinusoidal cells with myofibroblastic
characteristics and lipocytes (fat-storingcells, Ito-cells). They are frequently consi-dered to be two phenotypes of a singlehepatic connective tissue cell lineage (Biou-lac-Sage & Balabaud I985). Hepatic lipo-cytes were observed to be associated withcollagen fibres in normal and fibrotic hepaticparenchyma (Kent et al. 19 76). In an in-vivostudy of lipocytes, Clement et al. (I 984) havedemonstrated synthesis of extracellularmatrix molecules, involving fibronectin andcollagen types I, III and IV. In-vitro studieshave also indicated that lipocytes are one of
* Present address: Escola Tcnica Federal de Quimica, Rio de Janeiro, Brazil.Correspondence: Professor Radovan Borojevic, Caixa Postal 68021, 2I944 Rio de Janeiro, Brazil.
125

the major potential sources of collagen andassociated extracellular matrix molecules inliver tissue (Senoo et a]. I 984; Friedman et al.I985; Gressner & Haarman I988; Gressneretal. I989).An increase in number of lipocytes is
known to be associated with hepatic cir-rhosis due to hypervitaminosis A (Jacques etal. 19 79). Conversely, an increase in numberof liver myofibroblasts, associated with aproportional decrease in number of lipocytes,was observed in chronic alcohol intoxicationin both humans and experimental models(Minato et al. I983; Mak et al. I984). Thetransition of lipocytes into activated colla-gen-secreting myofibroblasts has thereforebeen proposed as one mechanism leading toincreased fibrogenesis in alcohol-induced cir-rhosis and fibrosis (Mak et al. I984). Theexpression of lipocyte versus myofibroblastphenotype therefore may be one of theprincipal control points for the homoeostasisof normal and pathological secretion ofextracellular matrix in liver.We have established a continuous cell
line, termed GRX, from fibro-granulomatouslesions induced in mouse liver by schistoso-mal infection (Borojevic et al. I 985a). It hasbeen maintained since then by serial pass-ages and has been recloned several times: ithas now undergone more than 500 popula-tion doublings. Nevertheless, GRX cell line isdependent upon a high concentration ofserum (maximal growth at 30% foetalbovine serum), it does not grow in soft agar,and is not tumorigenic in syngeneic mice,indicating that this is not a fully transformedcell line. It has myofibroblastic character-istics, secretes a complex extracellularmatrix, and displays in vitro biological andbiochemical characteristics of primary celllines isolated from human or mouse fibroticlivers (Borojevic et al. I985b; Monteiro &Borojevic I987). Like the in-vivo inductionof lipocyte hyperplasia by hypervitaminosisA, retinol treatment of GRX cells inducestheir conversion into the lipocyte phenotype(Margis & Borojevic I989a; Borojevic et al.I990). Differential study of lipid accumu-
lation and analysis of induction of enzymesinvolved in lipid synthesis have shown thatthe conversion of GRX cells displaying themyofibroblast phenotype into lipocytesrepresents induction of the full expression ofa different phenotype, and not simply ametabolic response ofthe cell line to a specificstimulus (Margis & Borojevic I989a; Boroje-vic et al. I990). Since this conversion hasbeen related to the control of liver fibrosis, wehave studied the in-vitro modification ofproliferation, cell adherence and secretion ofthe three major components of extracellularmatrix (collagen, glycosaminoglycans andfibronectin) in GRX cells, together with theirretinol-induced conversion from myofibro-blasts into lipocytes.
Materials and methods
Cell cultures
GRX-I 23 cells were obtained from the Rio deJaneiro Cell Bank (BIO-RIO, Rio de Janeiro,Brazil). They were routinely cultivated inDulbecco's medium (low glucose, SigmaChemical Co., St Louis, MO) supplementedwith Io% foetal bovine serum (Cultilab,Campinas, SP, Brazil) and 3 g/l of HEPES(Sigma), at 3 70C, under normal air atmos-phere.
Induction of the fat-storing phenotype
Freshly trypsinized cells were plated (5 x IO'cells/2 5 cm2 flask) and left to adhere for 2 h.They were washed with balanced salt solu-tion (BSS) and incubated in the mediumdescribed above, supplemented with IO 'PMall-trans retinol (Sigma), dissolved in abso-lute ethanol. Control cells were incubatedin the standard medium containing o.i%ethanol.
Cellular growth and proliferation rate
An aliquot of 3 x Io5 cells were plated per 25cm2 tissue culture flasks. They were left toadhere for 2 h and subsequently incubated in
I26 R. Margis et al.

Effect of retinol on liver myofibroblaststhe control or the retinol-containing media.Every 2 days, three flasks of control andretinol-treated cells were trypsinized andcells were counted.The proliferation rate of individual clones
was estimated by plating cells at cloningdensity (250 cells per 25 cm2 flask). Cellswere left to adhere overnight and subse-quently, the number of cells in 150 differentclones was monitored daily, in control andretinol-treated cultures, under a Leitzinverted microscope equipped with phasecontrast.
Estimation of cell adherence
The adherence of cells to the plastic substratewas estimated by their resistance to trypsini-zation. Io5 cells were plated per well, in 24-well plates (Nunclon). They were left toadhere for 2 h, and incubated for 36 h incontrol or retinol-containing media. Cellswere then washed twice with calcium andmagnesium-free BSS (CMF-BSS), and incu-bated with i ml/well of trypsin solution(0.I25 mg/ml trypsin and 0.025 mg/mlEDTA in CMF-BSS), with constant agitationon a gyratory shaker at 6o r.p.m. Trypsiniza-tion was interrupted after every 2 min intriplicate wells, by fixation with i ml ofsaturated aqueous solution of picric acid,containing 4% formalin. When all wells weretrypsinized and fixed, the fixative withdetached cells was discarded and the cellsthat remained adherent were quantified by amodified Coomassie Brilliant Blue R-250method (CBBR-250) (Margis & Borojevici989b).
Collagen quantificationCells were plated as described for assessingcell growth and proliferation, and left toattain early confluence (approximately 106cells/flask). Cell cultures were washed andincubated with 4 ml of the standard orretinol-containing medium, supplementedwith i pCi "4C-proline (273 mCi/mM specificactivity, New England Nuclear, Boston, Ma),
I00 pg/ml ascorbic acid and 50 pg/ml beta-amino-propionitrile (Sigma).
Procollagen was precipitated from cellculture supernatants as described by Ram-shaw et al. (I984). Briefly, 3 ml of super-natant was neutralized with NaOH supple-mented with 50 ,ug of carrier collagen type I.Collagen was subsequently precipitated byincubating with io% polyethylene glycolPEG-400 (Sigma), for 24 h at 40C. Theprecipitate was solubilized with 0.2 ml gla-cial acetic acid. Radioactivity was quantifiedby liquid scintillation counting in a BeckmanLS I803 counter, using the automaticquenching compensation program withinternal gamma-emitting radiation source,to express the results in d.p.m.
Total protein was precipitated from i ml ofsupernatant with trichloroacetic acid (TCA,12.5% w/v, final concentration) for 24 h at4°C. The precipitate was centrifuged (I 5 000g, I0 min), washed in io% TCA and centri-fuged again. The pellet was solubilized informic acid and radioactivity quantified asabove.
Fibronectin analysisCells were plated as described for collagensynthesis quantification. They were pre-treated for 24 h with retinol or with standardmedium and incubated for 4 h with 50 pCi of35S-methionine (generously supplied by Pro-fessor M. A. Rabello, Universidade Federal doRio de Janeiro) in i ml of standard medium.Cells were washed with BSS and disrupted byaddition of 200 p1 of standard urea lysisbuffer. Samples were centrifuged and super-natants stored at- 20°C for later analysis.
Fibronectin present in the cellular layerwas analysed by: (a) standard 5% polyacryl-amide gel electrophoresis (PAGE-SDS) in adiscontinuous gel buffer (Laemmli 1970),and (b) two dimensional isoelectric focusingfollowed by PAGE-SDS, described by O'Farrel(I974), using ampholines in the pH 5-7 and3.5-Io ranges (4:I).
Fibronectin was differentiated from otherproteins by immunoblotting (Symington
I27
R. Margis et al.
I984), using goat polyclonal anti-fibronec-tin and peroxidase-coupled anti-goat-IgGantibodies (both from Sigma). Radiolabelledfibronectin was quantified after exposure ofdried gels to X-Omat radiography film(Kodak, Sao Paulo, Brazil) and by densito-metric analysis of autoradiograms.
Glycosaminoglycans analysis
Cell cultures in early confluence were incu-bated in fresh Eagle's medium without sul-phate (Eurobio, Paris, France) containingio% foetal bovine serum, with or without io/IM all-trans retinol. Each 25 cm2 cultureflask was supplemented with 40 ,uCi H23"SO4(Instituto de Pesquisas Energeticas e Nuc-leares, Sao Paulo, Brazil), for 48 h at 3 70C ina humidified atmosphere containing 5%CO2. At the completion of this labellingperiod cells attained full confluence. Mediawere decanted and cells were washed twicewith phosphate-buffered saline (PBS, pH7.4). Sulphated glycosaminoglycans wereisolated from the cell culture as previouslydescribed (Silva et al. I990). Briefly, cellswere detached by mild trypsinization with 2ml CMF-BSS containing 0.05% trypsin and0.05 EDTA, for I0 min at room temperature.Cells were separated from the supernatant bycentrifugation: this supernatant containedpericellular glycosaminoglycans. The pelletwas subsequently digested with 3 ml of thesame trypsin solution, for 24 h at 3 7TC,releasing the intracellular glycosaminogly-cans. Both fractions were centrifuged andthe glycosaminoglycans in the supernatantwere precipitated with 95% ethanol andmaintained at - io'C for 24 h. The precipi-tates were collected by centrifugation,washed with 8o% ethanol, vacuum driedand dissolved in 0.2 ml distilled water.
35S-sulphated glycosaminoglycans wereidentified by their electrophoretic mobilitiesin agarose gels, in parallel with knownstandards, and by the pattern of their enzy-matic degradation with chondroitinases ACand ABC (Silva et al. I989).
Three independent experiments were done
in quadruplicate and results were analysedby Student's t-test: P-values equal or inferiorto O.OI were considered significant.
Results
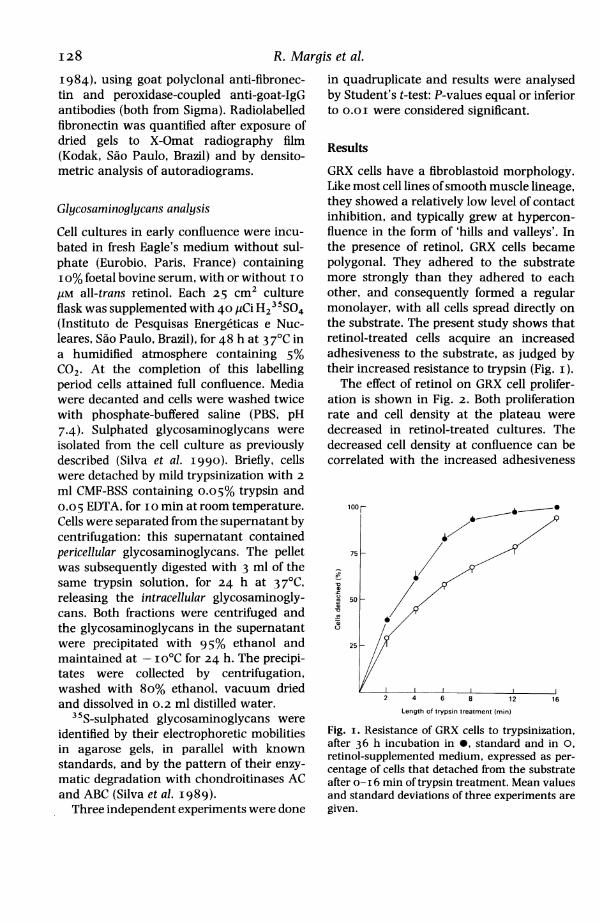
GRX cells have a fibroblastoid morphology.Like most cell lines ofsmooth muscle lineage,they showed a relatively low level of contactinhibition, and typically grew at hypercon-fluence in the form of 'hills and valleys'. Inthe presence of retinol, GRX cells becamepolygonal. They adhered to the substratemore strongly than they adhered to eachother, and consequently formed a regularmonolayer, with all cells spread directly onthe substrate. The present study shows thatretinol-treated cells acquire an increasedadhesiveness to the substrate, as judged bytheir increased resistance to trypsin (Fig. i).The effect of retinol on GRX cell prolifer-
ation is shown in Fig. 2. Both proliferationrate and cell density at the plateau weredecreased in retinol-treated cultures. Thedecreased cell density at confluence can becorrelated with the increased adhesiveness
75 - /
50
25
25-~~~~~~~~~
2 4 6 8 12 16
Length of trypsin treatment (min)
Fig. i. Resistance of GRX cells to trypsinization,after 36 h incubation in 0, standard and in 0,retinol-supplemented medium, expressed as per-centage of cells that detached from the substrateafter o- I 6 min of trypsin treatment. Mean valuesand standard deviations of three experiments aregiven.
I28

Effect of retinol on liver myofibroblasts
10
(O 0
2 4 6
Time in culture (days)
Fig. 2. Proliferation of GRX cells in 4and in 0, retinol-supplemented mediio days of culture. Results are exprecells x io0, and represent mean valuedard deviations of four experiments.
and consequent spreading on theThe proliferation rate was reduce(start of the culture period, in bot]high density cultures. This cot
either a decreased proliferation r
whole cell population, or the indunon-proliferative subpopulationnally differentiated cells. This que
addressed in the study of prolificloning density. This showed thation decrease to affect the entirelation, since no differentiationsubpopulations was detected (Fig4 days of culture, a net decreadivision was observed in retinol-treThis inhibition of cell proliferationdependent and increased steadily20 gM retinol (results not showconcentrations of retinol were ncGRX cell as determined by the trdye exclusion test.
Collagen secretion by GRX celwas influenced by the cell densretinol treatment modified cell prand consequently cell density icultures, we had to establish wi
observed differences in collagen secretionwere due directly to the presence of retinol orto the decrease of cell density. This was done
_91_~~ by monitoring the relationship between celldensity and the collagen secreted into thesupernatant by GRX cells under standardconditions, in the presence of beta-amino-propionitrile (Fig. 4). The relative inhibitionof collagen synthesis in the presence ofretinol is shown in Fig. 5. This inhibition wasspecifically of collagen synthesis since totalprotein synthesis did not change throughoutthe 8 days of culture. The ratio betweencollagen and total protein decreased from
8 10 50% at 2 days to less than 20% after I 2 daysof retinol treatment.
*, standard Modifications of protein synthesis inducedrnm, during in GRX cells by retinol were studied by both2ssed as log one and two-dimensional SDS-PAGE electro-Ss and stan- phoresis of proteins from cells and their
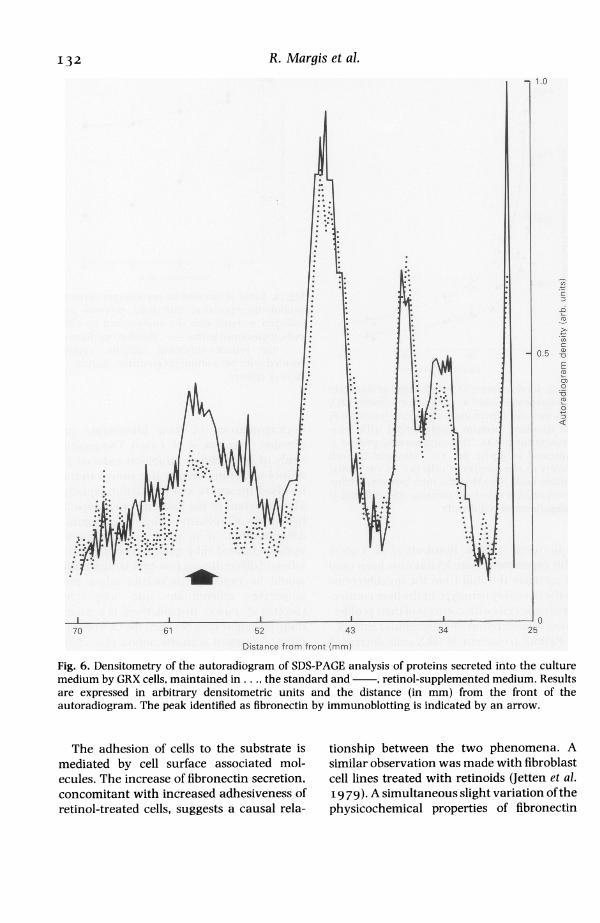
supernatants, after incubation with 35S-methionine. The synthesis of several un-identified proteins was modified. Fibronectin
substrate. was identified by immunoblotting. Densito-d from the metric analysis of autoradiograms of electro-h low and phoretic separations of synthesized proteins,ild reflect and the subsequent integration of surfacesate of the corresponding to the fibronectin band, hasiction of a shown that the incorporation of methionineof termi- into fibronectin was increased in retinol-zstion was treated cells (Fig. 6). Under standard celleration at culture conditions, the fibronectin bandve prolifer- appeared as a symmetrical peak, while incell popu- retinol-treated ones, a subfraction of fibro-into two nectin with a slightly higher molecular mass3). After could be identified (Fig. 6). Similarly, in two-
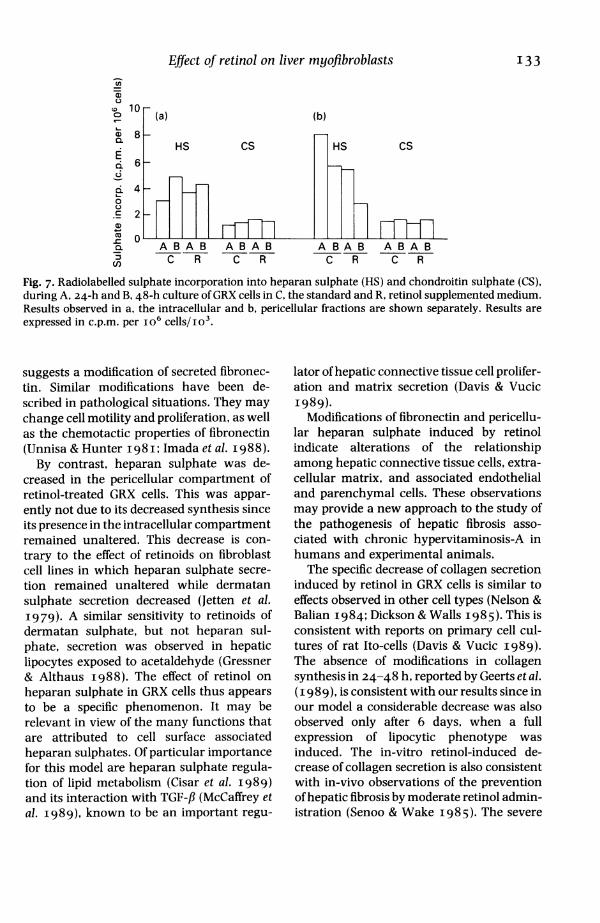
ise of one dimensional electrophoresis, the first isoelec-mated cells. tric-focusing run detected a slight shift of theiwas dose fibronectin spot to the basic pH (not shown).from i to Total glycosaminoglycan synthesis, asrn). These estimated by 35S incorporation, was signifi-t toxic to cantly decreased by retinol treatment. Theypan-blue decrease (P<O.OI) involved essentially the
pericellular fraction of glycosaminoglycans[Is in vitro since incorporation into the intracellular;ity. Since fraction remained approximately constantoliferation throughout 48 h. GRX 123 cells in effectn parallel synthesized heparan sulphate and chondroi-iether the tin 4-sulphate. While the 35S incorporation
I29

R. Margis et al.
60
40
20
'I)en
0
0
6z
60
40
20
(b)
0
0~~~
0 1 2 4 8 16 32
No. of cells
Fig. 3. Proliferation ofGRX cells at cloning density, in 0, standard and 0, retinol-supplemented mediumduring a, I; b, 2; c, 3; d, 4 days of culture. The number of clones containing from I to 128 cells is shownfor each day.
into chondroitin remained constantthroughout the experiment, its incorpora-tion into heparan was decreased (Fig. 7).
Discussion
Liver is a major site for uptake, storage andmobilization of retinol. Although hepato-cytes have been shown to incorporate andmetabolize retinol from chylomicrons, more
than 90% of stored retinoids in the liver are
found in lipocytes (Knook et al. I989).Studies of primary in-vitro cultures of rat
lipocytes have shown that retinol modifies
their proliferation rate and their metabolism(Davis et al. I987; Davis & Vucic I988).However, in primary cultures of hepaticmyofibroblasts, only a minor effect of retinolwas observed and a full expression of thelipocyte phenotype was not described (Geertset al. I 989). The cloned murine cell line GRX,used in the present study, was establishedfrom fibrotic lesions in liver: it expressesmorphological and biochemical featureswhich are characteristic of hepatic myofibro-blasts (Borojevic et al. i98 5a). These cells aresensitive to retinol, which induces a fullexpression ofthe lipocyte phenotype, and hasa pleotropic effect on cell metabolism (Margis
I30

Effect of retinol on liver myofibroblasts
3 6Time in culture (days)
-.A.
0 50 100 150
Cells per 25 cm2 x 105
Fig. 4. Incorporation of radiolabelled proline intoA, total proteins and ,, collagen secreted by GRX
cells into the supernatant during 24-h culture inthe standard medium supplemented with beta-aminopropionitrile. The incorporated proline isexpressed as d.p.m. per IOS cells, and the celldensity as the number of cells per 25 cm2 tissueculture flask. Note that the ratio between prolineincorporation into total proteins and collagen isindependent of cell density.
& Borojevic I989a; Borojevic et a). I990).This experimental model has now been usedto correlate the shift from the myofibroblastto the lipocyte phenotype in the liver connec-tive tissue cells with controls of their prolifer-ation and secretion of extracellular matrix.
Retinol treatment of GRX cells decreasedthe cell density at confluence and theiroverall proliferation rate. The former effectmay be related directly to increased adhe-siveness: the increased spreading of cells onthe substrate induces a more efficient contactinhibition, lowering the final cell density ofthe culture. The same phenomena cannot beinvolved in proliferation decrease, since thiswas observed also in low density cultures.We have shown that the conversion of
hepatic myofibroblasts into lipocytes is a full
Fig. 5. Ratio (expressed as percentage) betweenproline incorporation into total proteins andcollagen secreted into the supernatant by GRXcells, maintained in the , standard medium or
... the retinol-containing medium, supple-mented with beta-aminopropionitrile, during I2days of culture.
re-programming of their phenotypic ex-
pression (Borojevic et al. I990). The presentstudy of proliferation inhibition induced byretinol at cloning density has shown that itinvolved the entire cell population equally.The induction of the lipocytic phenotype inhepatic myofibroblasts is thus not a terminaldifferentiation; if it were, a proliferativeundifferentiated subpopulation plus a popu-lation of differentiated non-proliferating cellswould be expected, as occurs when pre-adipocytes differentiate into adipocytes(Scott et al. I982). Instead, there is a poten-tially reversible shift between the two pheno-types, consistent with the notion of a singlepopulation of connective tissue cells in hepa-tic lobules. The relative proportion of the twophenotypes is probably coordinated by exter-nal conditions, such as vitamin-A status andthe presence of potentially fibrogenic mol-ecules in hepatic tissue. The vitamin-A-induced 'hyperplasia' of fat-storing cells inliver is consequently due not to theirincreased proliferation but to the increasedconversion of liver connective tissue cellsinto the lipocyte phenotype.
C-
-6
a)
'a)
m
60-
o 40
o20
c
2a-
I
.. 1 -
;.I..
9 12
I31
R. Margis et al.
; . ; e. | . i .;, r2 OF,: -"I ' .'' , :1'
f~~~~~~~~~~~~~~~~~~~~0.'.i.r ;
T:l
j~~~~~~~~~i* -r. . - !R, ;, r, , .......... - E ... ;'m't *\rl. WM.@*;. ;.rFig. 6. Densitometry of the autoradiogram of SDS-PAGE analysis of proteins secreted into the culturemedium by GRX cells, maintained in . . ., the standard and , retinol-supplemented medium. Resultsare expressed in arbitrary densitometric units and the distance (in mm) from the front of theautoradiogram. The peak identified as fibronectin by immunoblotting is indicated by an arrow.
The adhesion of cells to the substrate ismediated by cell surface associated mol-ecules. The increase of fibronectin secretion,concomitant with increased adhesiveness ofretinol-treated cells, suggests a causal rela-
tionship between the two phenomena. Asimilar observation was made with fibroblastcell lines treated with retinoids (Jetten et al.19 79). A simultaneous slight variation of thephysicochemical properties of fibronectin
132
Effect of retinol on liver myofibroblasts
Ca)10O 10 (a) (b)
DLa> C R CHS Cs HS Csc6
0
.22
_L -A BAB A BA B A BA B A BA B:3 C R C R CR C R
Fig. 7. Radiolabelled sulphate incorporation into heparan sulphate (HS) and chondroitin sulphate (CS),during A, 24-h and B, 48-h culture of GRX cells in C, the standard and R, retinol supplemented medium.Results observed in a, the intracellular and b, pericellular fractions are shown separately. Results areexpressed in c.p.m. per io6 cells/io'.
suggests a modification of secreted fibronec-tin. Similar modifications have been de-scribed in pathological situations. They maychange cell motility and proliferation, as wellas the chemotactic properties of fibronectin(Unnisa & Hunter I98I; Imada et al. I988).By contrast, heparan sulphate was de-
creased in the pericellular compartment ofretinol-treated GRX cells. This was appar-ently not due to its decreased synthesis sinceits presence in the intracellular compartmentremained unaltered. This decrease is con-trary to the effect of retinoids on fibroblastcell lines in which heparan sulphate secre-tion remained unaltered while dermatansulphate secretion decreased (Jetten et al.I979). A similar sensitivity to retinoids ofdermatan sulphate, but not heparan sul-phate, secretion was observed in hepaticlipocytes exposed to acetaldehyde (Gressner& Althaus I988). The effect of retinol onheparan sulphate in GRX cells thus appearsto be a specific phenomenon. It may berelevant in view of the many functions thatare attributed to cell surface associatedheparan sulphates. Of particular importancefor this model are heparan sulphate regula-tion of lipid metabolism (Cisar et al. I989)and its interaction with TGF-fl (McCaffrey etal. I989), known to be an important regu-
lator of hepatic connective tissue cell prolifer-ation and matrix secretion (Davis & VucicI989).
Modifications of fibronectin and pericellu-lar heparan sulphate induced by retinolindicate alterations of the relationshipamong hepatic connective tissue cells, extra-cellular matrix, and associated endothelialand parenchymal cells. These observationsmay provide a new approach to the study ofthe pathogenesis of hepatic fibrosis asso-ciated with chronic hypervitaminosis-A inhumans and experimental animals.The specific decrease of collagen secretion
induced by retinol in GRX cells is similar toeffects observed in other cell types (Nelson &Balian I 984; Dickson & Walls I98 5). This isconsistent with reports on primary cell cul-tures of rat Ito-cells (Davis & Vucic i989).The absence of modifications in collagensynthesis in 24-48 h, reported by Geerts et al.(i 989), is consistent with our results since inour model a considerable decrease was alsoobserved only after 6 days, when a fullexpression of lipocytic phenotype wasinduced. The in-vitro retinol-induced de-crease of collagen secretion is also consistentwith in-vivo observations of the preventionof hepatic fibrosis by moderate retinol admin-istration (Senoo & Wake I985). The severe
133

I34 R. Margis et al.
liver fibrosis and cirrhosis observed in ele-vated chronic hypervitaminosis-A remain tobe explained. They are apparently not di-rectly related to retinol modulation of colla-gen synthesis in liver connective tissue cellsbut should involve more complex cell andtissue controls of extracellular matrix de-posits.
Acknowledgements
This investigation has received financialsupport from CNPq, FINEP and FAPERJ,Brazil and from the Commission of theEuropean Communities (CEE STD221 7.M.(B)).
ReferencesBIOIJLAC-SAGE P. & BALABAIJD C. (i 9 8 5) La cellule
perisinusoidale (ou cellule de Ito). Gastroenterol.Clin. Biol. 9, 312-322.
BOROJEVIc R., GIJARAGNA R.M., MARGIS R. & DIJTRAH.S. (90ggo) In-vitro induction of the fat-storingphenotype in a liver connective tissue cellline-GRX. In Vitro Cell. Develop. Biol. 2 5, 36 I-368.
BOROJEVIc R., MONTEIRO A.N.A., VINHAS S.A.,DOMONT G.B., MOURAO P.A.S., EMONARD H.,GRIMALDI JR. G. & GRIMAUDJ.A. (i985a) Estab-lishment of a continuous cell line from fibroticschistosomal granulomas in mice livers. InVitro Cell. Develop. Biol. 21, 3 82-390.
BOROJEVIc R., VINHAS S.A., MONTEIRO A.N.A.,DOMONT G.B., ZYNGIER F.R. & GRIMAUD J.A.(I985b) Liver connective tissue cells isolatedfrom human schistosomal fibrosis or alcoholiccirrhosis represent a modified phenotype ofsmooth muscle cells. Biol. Cell 5 3, 2 3 I1-2 3 8.
CISAR L.A., HOOGERWERF A.J., CuPP M., RAPPORTC.A. & BENSADOUN A. (I989) Secretion anddegradation of lipoprotein lipase in culturedadipocytes. J. Biol. Chem. 264, 1767-I774.
CLEMENT B., EMONARD H., RISSEL M., DRUGUET M.,GRIMAUD J.A., HERBAGE D., BOIJREL M. & GuIL-LOIJZO M. (j1984) Cellular origin of collagen andfibronectin in liver. Cell. Mol. Biol. 30, 489-496.
DAVIS B.H., PRATT B.M. & MADRI J.A. (I987)Retinol and extracellular collagen matricesmodulate hepatic Ito cell collagen phenotypeand cellular retinol binding protein levels. 1.Biol. Chemn. 262, 1O280-IO286.
DAVIS B.H. & VuJcic A. (1988) The effect of retinolon Ito cell proliferation in vitro. Hepatology 8,788-793.
DAVIS B.H. & VuJcic A. (1989) Transforminggrowth factor-beta modulates hepatic Ito cellproliferation, collagen synthesis and vitamin Ametabolism in vitro. In Cells of Hepatic Sinusoid.Volume 2. eds E. Wisse, D.L. Knook & K. Decker.Rijswijk, The Netherlands: Kupffer Cell Founda-tion, pp. 39-42.
DICKSON I. & WALLS J. (I 98 5) Vitamin A and boneformation. Effect of an excess of retinol on bonecollagen synthesis in vitro. Biochem. J. 226,789-795.
FRIEDMANN S.L., ROLL F.J., BOYLES J. & BISSEL M.(1985) Hepatic lipocytes: the principal colla-gen-producing cells of normal rat liver. Proc.NatI Acad. Sci. USA 82, 868I-8685.
GEERTS A., VRIJSEN R., SCHELLINCK P. & WISSE E.(I989) Retinol affects the phenotype and pro-tein synthesis of fat storing cell derived myo-fibroblasts in vitro. In Cells of Hepatic Sinusoid.Volume 2. Eds E. Wisse, D.L. Knook & K. Decker.Rijswijk, The Netherlands: Kupffer Cell Founda-tion, pp. 20-24.
GRESSNER A.M. & ALTHAIJS M. (I988) Effect ofethanol, acetaldehyde, and lactate on proteog-lycan synthesis and proliferation of cultured ratliver fat-storing cells. Gastroenterology 94, 79 7-807.
GRESSNER A.M. & HAARMANN R. (1988) Hyaluro-nic acid synthesis and secretion by rat liver fatstoring cells (perisinusoidal lipocytes) in cul-ture. Biochem. Biophys. Res. Commun. 15 I,222-229.
GRESSNER A.M., HAARMANN R., SCHAFER S. &ZERBE 0. (I989) The role of fat storing cells inmatrix proteoglycan and hyaluronate produc-tion in relation to Kupffer cells and hepatocytes.In Cells of Hepatic Sinusoid. Volume 2. Eds E.Wisse, D.L. Knook & K. Decker. Rijswijk, TheNetherlands: Kupffer Cell Foundation, pp. 64-68.
IMADA S., SIJGIYAMA Y. & IMADA M. (I988)Fibronectin phosphorylation by ecto-proteinkinase. Exp. Cell Res. 179, 554-564.
JACQIJES E.A., BIJSCHMANN R.J. & LAYDEN T.J.( r979) The histopathologic progression of vita-min A induced hepatic injury. Gastroenterology76, 599-602.
JETTEN A.M., JETTEN M.E.R., SHAPIRO S.S. & POONJ.P. (I979) Characterization of the action ofretinoids on mouse fibroblast cell lines. Exp. CellRes. 119, 289-299.
KENT G.. GAY S., INOUYE T., BAHU R., MINICK O.T. &POPPER H. (1976) Vitamin-A containing lipo-

EJfect of retinol on liver myofibroblasts I35cytes and formation of type III collagen in liverinjury. Proc. Nati Acad. Sci. USA 73, 3719-3722.
KNOOK D.L., BLANER W.S., BROUWER A. & HEN-DRIKS H.F.J. (I989) The role of non-parenchy-mal cells in liver retinoid metabolism. In Cells ofHepatic Sinusoid. Volume 2. Eds E. Wisse, D.L.Knook & K. Decker. Rijswijk, The Netherlands:Kupffer Cell Foundation, pp. i6-i 9.
LAEMMLI U.K. (I970) Cleavage of structural pro-teins during the assembly of the head ofbacteriophage T4. Nature 227, 680-685.
MAK K., LEO M.A. & LIEBER C.S. (I984) Alcoholicliver injury in baboons: transformation of lipo-cytes to transitional cells. Gastroenterology 87,I88-200.
MARGIS R. & BOROJEVIc R. (I989a) Retinoid-mediated induction ofthe fat-storing phenotypein a liver connective tissue cell line (GRX).Biochim. Biophys. Acta IoII, 1-5.
MARGIS R. & BOROJEVIc R. (I989b) Quantificationof attached cells in tissue culture plates and onmicrocarriers. Anal. Biochem. I 8 I, 209-21 1.
MCCAFFREY T.A., FALCONE D.J., BRAYTON C.F.,AGARWAL L.A., WELT F.G.P. & WEBSTER B.B.(I989) Transforming growth factor-# activityis potentiated by heparin via dissociation ofthe transforming growth factor B/A2 macro-globulin inactive complex. 1. Cell Biol. IO9,441-448.
MINATO Y., HASUMURA Y. & TAKEUCHI J. (I983)The role of fat-storing cells in Disse spacefibrogenesis in alcoholic liver disease. Hepa-tology 3, 559-566.
MONTEIRO A.N.A. & BOROJEVIc R. (I987) In vitroformation of fibrous septa by liver connectivetissue cells. In Vitro Cell. Develop. Biol. 23, 10-14.
NELSON D.L. & BALIAN G. (I984) The effect ofretinoic acid on collagen synthesis by human
dermal fibroblasts. Collagen Rel. Res. 4, II9-I28.
O'FARELL P.H. (I974) High resolution two-dimensional electrophoresis of proteins. 1. Biol.Chem. 250, 4007-402I.
RAMSHAW J.A.M., BATEMAN J.F. & COLE W.G.(I984) Precipitation of collagen by polyethyl-eneglycols. Anal. Biochem. 141, 361-365.
ScoTT R.E., HOERL B.J., WILLE JR. J.J., FLORINE D.L.,KRAWIsz B.R. & YIJN K. (I982) Coupling ofpreadipocyte growth arrest and differentiation.A cell cycle model for the physiological controlof cell proliferation. 1. Cell Biol. 94, 400-405.
SENOO H., HATA R., NAGAI Y. & WAKE K. (I984)Stellate cells (Vitamin A-storing cells) are theprimary site of collagen synthesis in non-parenchymal cells in the liver. Biomed. Res. 5,451-458.
SENOO H. & WAKE K. (I985) Suppression ofexperimental hepatic fibrosis by administrationof vitamin A. Lab. Invest. 52, I82-194.
SILVA L.C.F., MOURAO P.A.S. & BOROJEVIc R.(1989) Patterns of sulphated glycosaminogly-cans synthesis and accumulation in hepaticgranulomas induced by schistosomal infection.Exp. Mol. Pathol. 50, 411-420.
SILVA L.C.F., BOROJEVIc R. & MOURAO P.A.S.(I990) Identification of cells responsible forsynthesis of sulphated glycosaminoglycans inschistosome-induced hepatic granulomas. Int.J. Exp. Pathol. 71, 845-856.
SYMINGTON J. (I984) Electrophoretic transfer ofproteins from two-dimensional gels to sheetsand their detection. In Two-Dimensional GelElectrophoresis of Proteins. Eds J.E.C. Celis &R. Bravo. Academic Press, New York. pp. I 2 7-i68.
UNNISA I. & HUNTER T. (I 98 I) Structural compari-son of fibronectins from normal and trans-formed cells. J. Biol. Chem. 256, 7671-7677.




















