
THE EFFECT OF MARINATION AND COOKING ON THE MECHANICAL PROPERTIES OF INTRAMUSCULAR CONNECTIVE TISSUE
19
THE EFFECT OF MAFUNATION AND COOKING ON THE MECHANICAL PROPERTIES OF INTRAMUSCULAR CONNECTIVE TISSUE GRAHAM J. LEWIS and PETER P. PURSLOWI Muscle Biology Department, AFRC Insitute of Food Research, Lungford, Bristol BS18 7DY, Great Britain Received for Publication on July 30, 1990 Accepted for Publication November 26, 1990 ABSTRACT Effects of acid marination and cooking on the mechanical properties of in- tramuscular connective tissue and whole muscle were investigated. R e transverse strength of cooked muscle was decreased by marinating and at all cooking temperatures between 40°C and 90°C. During marination the samples increased in cross-sectional area perpendicular to the direction of testing, and this increase was largely retained during cooking. A decrease in transverse strength brought about by marination could only be partly explained on the basis of this swelling thus diluting out the load-bearing connective tissue structures. Marinating also reduced the strength of isolated perimysia and produced a reversible increase in its initial stifiess, apparently due to straightening out of collagen fibers during swelling. Real weakening of the load bearing components occurred only when the samples were subsequently cooked to above 60°C. Apparent weakening could be attributed to a dilution of the components due to swelling. INTRODUCTION Acidic marination, i.e., the soaking of meat or fish in an acidic solution, is a commonly used culinary technique which aims to improve tenderness. Connec- tive tissue-rich muscles are often tough and are therefore the cheaper parts of a carcass, and the tenderizing effect of marinating could be a commercially impor- tant means of upgrading especially these tough muscles. To exploit this phenomena most efficiently, a more detailed understanding of the mechanism of marinating is required, especially in relation to the collagenous contribution to meat toughness. 'Current address is Muscle and Collagen Research Group, Dept. of Veterinary Medicine, University of Bristol Veterinary School, Langford, Bristol BS18 7DY, Great Britain Journal of Muscle Foods 2 (1991) 177-195. A11 Rights Reserved. @Copyright 1991 by Food & Nutrition Press, Inc., Trumbull, Connecticut. 177
Transcript of THE EFFECT OF MARINATION AND COOKING ON THE MECHANICAL PROPERTIES OF INTRAMUSCULAR CONNECTIVE TISSUE

THE EFFECT OF MAFUNATION AND COOKING ON THE MECHANICAL PROPERTIES OF INTRAMUSCULAR CONNECTIVE TISSUE
GRAHAM J. LEWIS and PETER P. PURSLOWI
Muscle Biology Department, AFRC Insitute of Food Research, Lungford, Bristol BS18 7DY, Great Britain
Received for Publication on July 30, 1990 Accepted for Publication November 26, 1990
ABSTRACT
Effects of acid marination and cooking on the mechanical properties of in- tramuscular connective tissue and whole muscle were investigated. R e transverse strength of cooked muscle was decreased by marinating and at all cooking temperatures between 40°C and 90°C. During marination the samples increased in cross-sectional area perpendicular to the direction of testing, and this increase was largely retained during cooking. A decrease in transverse strength brought about by marination could only be partly explained on the basis of this swelling thus diluting out the load-bearing connective tissue structures. Marinating also reduced the strength of isolated perimysia and produced a reversible increase in its initial stifiess, apparently due to straightening out of collagen fibers during swelling.
Real weakening of the load bearing components occurred only when the samples were subsequently cooked to above 60°C. Apparent weakening could be attributed to a dilution of the components due to swelling.
INTRODUCTION
Acidic marination, i.e., the soaking of meat or fish in an acidic solution, is a commonly used culinary technique which aims to improve tenderness. Connec- tive tissue-rich muscles are often tough and are therefore the cheaper parts of a carcass, and the tenderizing effect of marinating could be a commercially impor- tant means of upgrading especially these tough muscles. To exploit this phenomena most efficiently, a more detailed understanding of the mechanism of marinating is required, especially in relation to the collagenous contribution to meat toughness. 'Current address is Muscle and Collagen Research Group, Dept. of Veterinary Medicine, University of Bristol Veterinary School, Langford, Bristol BS18 7DY, Great Britain
Journal of Muscle Foods 2 (1991) 177-195. A11 Rights Reserved. @Copyright 1991 by Food & Nutrition Press, Inc., Trumbull, Connecticut. 177

178 G.J. LEWIS AND P.P. PURSLOW
It has been proposed (Offer and Knight 1988) that three potential mechanisms of tenderization due to marination are
(1) pH-induced swelling dilutes the amount of load-resisting material, (2) accelerated or additional proteolytic weakening of (a) muscle fibers,
and/or (b) connective tissue; and (3) increased rates of collagen solubilization upon cooking.
The evidence for each of these possibilities is detailed by Offer and Knight (1988).
In order to examine the contibution from the first mechanism it is necessary to relate changes in the strength of the meat to the amount of swelling induced, and specifically to the swelling in the plane perpendicular to the direction of applied stress. Several previous studies have addressed these aspects. Wenham and Locker (1976), measured dimensional changes on marination and related them to measured toughness. However, due to the large sample size, sufficient penetration by the marinade was not achieved and the meat pieces in their study increased in volume by only 15% and in weight by only 19% after 43h in 1.5% acetic acid. Nevertheless, a three fold increase in tenderness was found. Gault (1984) reported weight gains of about 100% on marination, but measured only total weight gain and not dimensional changes. Again, substantial reductions in the shear strength due to marinating were observed.
The results of Wenham and Locker (1976) and of Gault (1984) clearly imply that meat swelling resulted in some dilution of the load-bearing components, but in addition there may have been direct effects of the marinating on the intrinsic strength of these components. Because of the difficulty in defining the exact pat- tern of applied stress in shear press tests, it is difficult by use of these tests to separate and quantify the precise contribution of dilution effects as opposed to weakening of myofibrillar and collagenous components to overall toughness.
Previous work also indicates that weakening of the intramuscular connective tissue during marination may also occur, although the evidence is not clear, Wenham and Locker (1974) inferred that marinating had a greater effect on the tenderness of muscles with a high collagen content. However, this conclusion was reached on the basis of comparing different muscles used in two different experiments. The meat pieces in the two experiments differed in size as well as in the method of cooking and so were not strictly comparable. Gault (1985) felt that the relationship between meat tenderness and the degree of pH-induced swelling was not unduly affected by the connective tissue content of the muscle, although he reported that the form of the relationship between toughness and degree of swelling did differ between muscles of different collagen content.
As well as its direct contribution to meat toughness, intramuscular connective tissue also influences meat texture indirectly by its shrinkage on cooking; this squeezes out fluid released from the heated myofibrils, so affecting their proper- ties and hence contribution to texture. Changes in the thermally-induced

MARINATION EFFECTS ON CONNECTIVE TISSUE 179
shrinkage of collagen at low pH may affect the toughness of marinated meat and also after cooking yields by this means. Privalov (1982) noted that collagen in solution suffers a 6°C depression in its denaturation temperature at pH 2.2 com- pared to pH 7.0. Any decrease in thermal denaturation temperatures may result in the mechanical changes brought on in meat by cooking at lower temperatures in marinated than in unmarinated meat.
Perimysial connective tissue has previously been shown to have an important role in meat toughness (Purslow 1985). Tensile tests on meat pulled across the muscle fiber direction measure the lateral strength of the perimysial network without a contribution from muscle fibers (Bouton and Harris 1972), and so can be used to judge the effects of any treatment on the connective tissue component of meat toughness. Use of tensile testing allows the strength values to be cor- rected for changes in cross-sectional area during marinating and cooking and so make it possible to distinguish between changes in the strength resulting from dilution of load-resisting components and those resulting from direct effects on the components.
The intent of this study was to clarify whether the increased tenderness of meat caused by marinating prior to cooking was entirely due to the dilution of load- bearing components or due in part to direct effects on the muscle fiber and con- nective tissue components. In order to determine this, we have measured the ten- sile strength of normal and marinated meat both parallel and perpendicular to the muscle fiber direction, after cooking to a range of temperatures. Perpendicular strength measuring essentially the strength of the collagenous perimysial net- work, and parallel strength, determined by both myofibrillar and collagenous components, have been corrected for changes in dimensions on marination in order to quantify the dilution effect. The tensile strength and stiffness of isolated perimysial strips have also been measured at high and low pH to show directly the effect of pH on the intrinsic properties of this component.
MATERIALS AND METHODS
Transverse Breaking Strength of Meat
Both sernitendinosus muscles of an 18 month-old Fresian-cross heifer were ex- cised 24 h postmortem. The muscles were cleaned of external fat, vacuum- packaged and stored at -20°C. When frozen, each muscle was cut on a band saw, first in half across the fiber direction then into 7mm thick longitudinal slices. The slices were individually vacuum-packaged and again stored at -20 "C. There were enough slices to provide one from each muscle for each of 14 treatments. A template and a sharp knife were used to cut 2 blocks measuring 5Omm by 5Omm from each of the frozen slices. The blocks were allowed to thaw at room temperature.

180 G.J. LEWIS AND P.P. PURSLOW
For each temperature treatment, two blocks (one originating from each mus- cle) were each placed into 500 mL of 1 % acetic acid that had been pre-chilled to 4°C. The blocks were supported in the solution on a wire mesh platform. This enabled the marinade to be gently circulated by a magnetic stirrer underneath the platform without damaging the meat pieces. The control blocks for each treat- ment were simply kept in sealed plastic bags at the same temperature. After 24 h, the blocks were removed, individually vacuum packed and cooked in a preheated water bath at 40 O , 50 O , 60 O , 70 O, 80 " or 90 "C for 1 h, or in the case of the uncooked treatment, used immediately. Cooking was arrested by placing the packed slices in running cold water for 10 min. After thawing, after marina- tion and after cooking, the blocks were surfacedried then weighed and their length, width and thickness recorded. Dumbelled tensile test pieces were cut from the raw or cooked blocks using a 5mm-wide template and a sharp scalpel. The test pieces were measured in tension on an Instron 6022 Testing Machine with an extension rate of 50mm/min and a gauge length of 30 mm between the pneumatic grips. Tensile strengths were taken as peak load before fracture divid- ed by the cross-sectional area of the specimens.
Longitudinal Breaking Strength of Meat
Another pair of semitendinosus muscles was prepared in the same way as detailed above. In this case, one block measuring 50 mm wide by 100 mm long was cut from each of the frozen slices. The blocks were then marinated as before. For each temperature treatment, one block originating from each muscle was placed into 500 mL of 1 % acetic acid prechilled to 4 "C. The blocks were then cooked at the same range of temperatures as above for 1 h, or in the case of the uncooked treatment, used immediately. Dumbelled test specimens were cut from the blocks, orientated along the fiber direction, using a 5 mm-wide template and a sharp scalpel.
In the transverse strength experiment, detailed above, problems were en- countered in determining the strength of the marinated samples as their slimy and gel-like texture made gripping the ends for testing difficult, especially after cooking to higher temperatures. As the strength of meat parallel to the muscle fiber direction is much higher, this was even more of a problem in this experi- ment. Consequently, liquid C0,cooled 'cryo-grips' were employed in place of the pneumatic grips. As such grips are an innovation in mechanical tests on meat, brief details of their construction and performance are given in the appen- dix. Tensile tests were performed as for the transverse tests above, except that the gauge length of the meat blocks was twice that used previously, i.e., 60 mm instead of 30 mm.

MARINATION EFFECTS ON CONNECTIVE TISSUE 181
Stiffness of Isolated Perimysium
Another semitendinosus muscle was prepared as above except that the frozen muscle was cut on a band saw into 7 mm thick transverse slices. A ribbon of perimysial connective tissue was carefully dissected from around the primary bundles of one of the slices. Ribbons with an initial length of 2.9 mm and a width of 2 mm, were tested in the microscope-mounted tensile testing machine describ- ed by Lewis and Purslow (1989). The long axis of the specimens (direction of stretching) corresponded to the direction transverse to the muscle fibers in the meat. The specimen was immersed in 0.9% NaCl before and during testing, and cyclically loaded a number of times at a rate of 6.5 mmlmin, ensuring that the upper limit of extension did not cause any irreversible damage to the material. The load-extension curves were automatically recorded. The bathing medium was then quickly exchanged for 1% acid in 0.9% NaCl. The load extension behavior was again determined at intervals up to 30 min to determine any time- dependent changes in the properties of the tissue due to low pH. The bathing medium was then replaced with 0.9% NaCl and the process repeated to see if the changes in mechanical properties were reversible.
Breaking Strength of Isolated Perimysia
An additional sernirendinosus muscle was prepared as stated above. Ribbons of perimysial connective tissue were dissected out of the slices as before and then dumbelled to leave a central parallel-sided part 5 mm long by 2 mm wide. These were soaked at room temperature in either 0.9% NaCl or 1 % acetic acid in 0.9% NaCl for 1 h. No consistent dimensional changes were seen in the test pieces during these treatments; however, there may be a significant margin of error in such observations due to the high compliance of the material. Specimens were then subjected to tensile test in the microscope-mounted tensile testing machine. The bathing media used during the test was the same as that used previously to soak the specimen. These samples were extended at a rate of 6.5 mm/min to breaking point and the peak breaking loads noted. After the test, the central parallel-sided part of each test piece was cut away and retained. These were then freeze dried and a value of breaking strength determined in terms of breaking load per dry weight of the standard length of specimen.
RESULTS AND DISCUSSION
Changes in the Meat Blocks During Marination
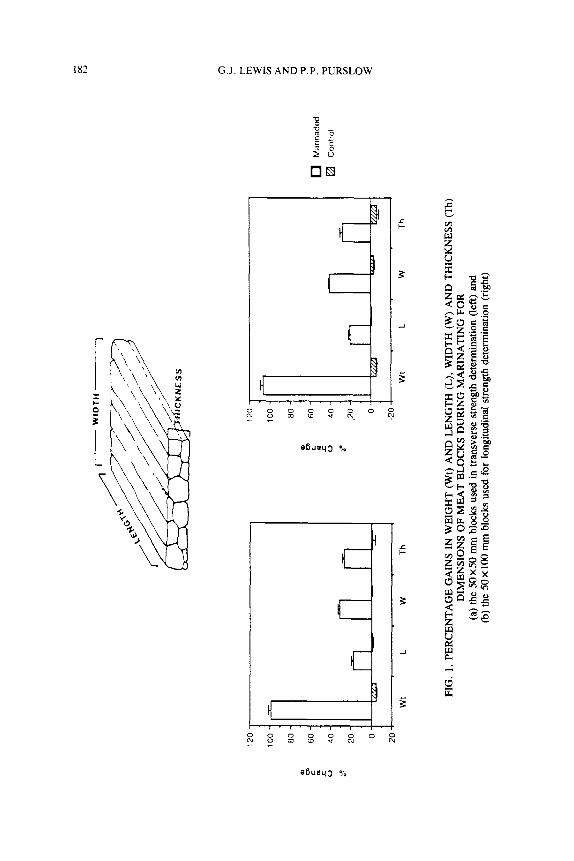
Figure 1 summarizes the dimensional and weight changes induced by marina- tion. The samples increased in weight by approximately 100% during the 24 h.
120 '
0
C
m S cn
0 s
20 !
wt
L W
Th
120
,
u C
W
S
0
cn
a"
-" .
Wt
L W
Th
FIG
. 1. P
ER
CE
NT
AG
E G
AIN
S IN
WEI
GH
T (W
t) A
ND
LEN
GTH
(L
), W
IDTH
(W
) AN
D T
HIC
KN
ESS (Th)
DIM
EN
SIO
NS
OF
ME
AT
BLO
CK
S D
UR
lNG
MA
RIN
AT
ING
FO
R
(a) t
he 5
0 x50
mm
blo
cks u
sed
in tr
ansv
erse
stre
ngth
det
erm
inat
ion
(left)
and
(b
) the
50
x 10
0 m
m b
lock
s use
d fo
r lo
ngitu
dina
l str
engt
h de
term
inat
ion
(rig
ht)
Mar
inad
ed
P
Con
trol
2

183 MARINATION EFFECTS ON CONNECTIVE TISSUE
In terms of dimensional changes, there appeared to be a difference between the degree of swelling in the two directions lateral to the muscle fiber direction, the swelling in the width dimension being the larger in both experiments. Although the blocks did not remain regular after thawing and this provided a source of er- ror in the measuring of width, thickness and length, the measured volume change shows reasonable agreement with the overall weight change. The anisotropic swelling transverse to the muscle fiber axis may be due to the fiber bundles being wider in one direction than the other before swelling. Locker and Daines (1976) reported that, under alkali conditions, several muscles exhibit anisotropic swelling perpendicular to the muscle fiber direction. Histological evidence for a preferred direction in the perimysial network architecture was on- ly found for one of muscles (M.psom). However, shear tests carried out in two orthogonal directions parallel to the fibers did suggest a difference in the strength of the connective tissue component in these directions, which might be due to an anisotropy in the perimysial architecture. Rowe (1977) showed that fiber bundles in beef M. semitendinosus are initially elongated in one dimension of their cross-section and this corresponds with the observed anisotropy of swell- ing reported here.
It is interesting to note that a length increase of about 20% occurred on marinating. The turgid appearance of the meat suggests that this increase in length could occur due to the swelling in the lateral directions being restricted by the perimysial sheaths, so forcing the swelling to be accommodated by length changes. Gault (1985) infers that the resistance to swelling provided by in- tramuscular connective tissue is an important factor in defining the amount of fluid uptake, from the observation that the degree of swelling appeared to reflect the differences in connective tissue content of the 3 muscles he studied.
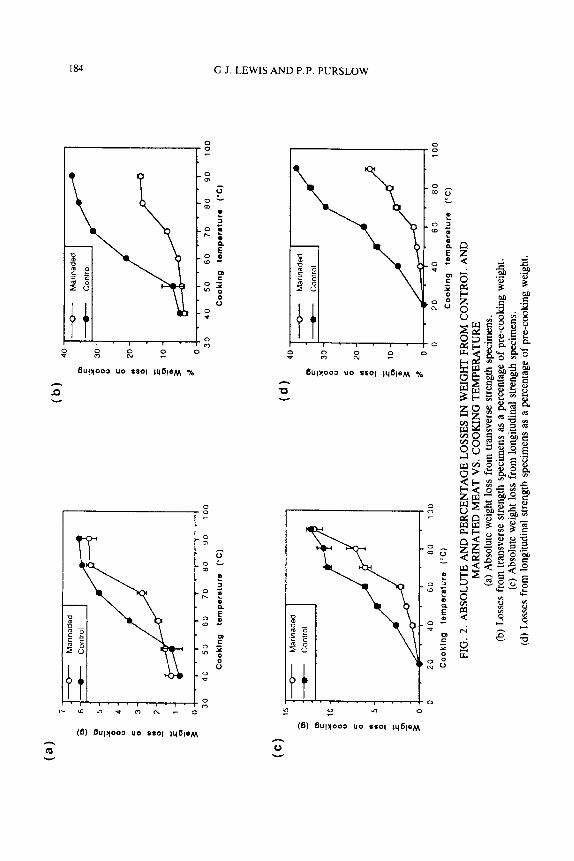
Cooking Losses from Control and Marinated Meat
Figure 2 reflects the weight loss at various cooking temperatures, both in ab- solute terms and as a percentage of the weight of specimens before cooking (but after marination for the non-control samples), for the meat blocks used in the determination of transverse strength (Fig. 2a and 2b) and longitudinal strength (Fig. 2c and 2d). The results for both transverse and longitudinal specimens are shown because these have different shapes, and cooking losses may differ because of this condition. The absolute weight loss on cooking was much the same for the marinated and untreated blocks (Fig. 2a and 2c) so that the weight gained during marination was largely retained during cooking. Roughly similar absolute weight losses between control and marinated (i.e., swollen) meat at each temperature of cooking results in the percentage weight loss from the marinated meat being less at all cooking temperatures. The fluid content of marinated meat is thus higher than that of control samples at all cooking temperatures.
o-
l.
,.
,
. ,
. ,
. ,
. ~
r
30
40
5
0
60
7
0
80
90
Co
ok
lng
te
mp
era
ture
("
C)
:; 10 - 01
m - I 0 u
--O
-
Mar
inad
ed
----t
Con
trol
5
. ,
. ,
. ,
. ,
. ,
. ,
.
.* M
arin
aded
*-
Con
lrol
-0-
Mar
inad
ed
(''
15
(dl
40
-
Con
trol
E!
m.
m
-
o 30-
- -
x
10 -
0.
u
.
C.
0
u C
O 20-
- x I
80
v
.1
.1
.1
.1
2 5
-
... -
.
-.
L m
10
- -
.c m.
-
0
0
20
40
60
80
1
00
2
0
40
60
80
10
0
Co
ok
lng
le
rnp
era
lure
("
C)
Co
ok
lng
te
mp
ers
ture
("
C)
FIG
. 2. A
BSO
LUTE
AN
D P
ERC
ENTA
GE
LOSS
ES I
N W
EIG
HT
FRO
M C
ON
TRO
L A
ND
M
AR
INA
TED
ME
AT
VS.
CO
OK
ING
TEM
PER
ATU
RE
(a) A
bsol
ute
wei
ght l
oss
from
tran
sver
se st
reng
th s
peci
men
s. (b
) Los
ses
from
tran
sver
se st
reng
th sp
ecim
ens a
s a
perc
enta
ge o
f pre
-coo
king
wei
ght.
(c) A
bsol
ute
wei
ght l
oss
from
long
itudi
nal s
tren
gth
spec
imen
s. (d
) Los
ses
from
long
itudi
nal
stren
gth
spec
imen
s as
a pe
rcen
tage
of
pre-
cook
ing
wei
ght.
... $2 0 i- Q E
?
m
P
ycr

m n 3
0.10
- m a 5 rn I 0.
05
c
0 - t
0.00
5
m
t C m
+ Ma
rinad
ed
--t
Con
trol
;:*
I.
,.
,.
m
m t m - X
U
Mar
inad
ed
0001
. ,
. ,
. ,
. ,
. 0
20
40
6
0
80
1
00
C
oo
kln
g
tem
pe
ratu
re
('C)
U
Mar
inad
ed
06
2 . ---C
C
onuo
l I f;
04
m
a Y t m
00 0
20
40
60
8
0
1
Co
oki
ng
te
mp
era
ture
("
C)
I
(dl
0.
61
]--t - Co
ntro
l M
arin
aded
I P
I
5 I-----
/I
0 2
0
40
60
80
10
0
Co
okl
ng
te
mp
era
ture
("
C)
FIG
. 3. B
REA
KIN
G S
TREN
GTH
OF
CO
NTR
OL
AN
D M
AR
INA
TED
MEA
T V
S. COOKING T
EMPE
RA
TUR
E M
easu
red
stre
ngth
s (a
) per
pend
icul
ar a
nd (
c) pa
ralle
l to
the
mus
cle
fiber
axi
s. B
reak
ing
stre
ngth
cor
rect
ed fo
r cr
oss-
sect
iona
l are
a cha
nge d
ue to
mar
inat
ing
are
show
n in
(b) f
or th
e pe
rpen
dicu
lar
and
(d) t
he p
aral
lel d
irec
tions
.

186 G.J. LEWIS AND P.P. PURSLOW
Transverse Breaking Strength of Meat
The form of the relationship between breaking strength and cooking temperature in the untreated meat is shown in Fig. 3a and is similar to that reported previously (Bouton and Harris 1972). There is an increase in strength with cooking to 50 "C followed by a decrease with cooking to 60 "C. A further increase in strength occurs with cooking to 70°C and then a fall in strength above 80 "C.
The strength of the test specimens from the marinated meat was substantially less than that of nonmarinated controls at all cooking temperatures. This cor- responds to some extent to the decreased shear force values observed by Gault (1985) and by Wenham and Locker (1976) on marinated meat after cooking. However, these authors could infer only indirectly how much of the tenderizing effect was due to changes in the myofibrillar component of toughness as opposed to the connective tissue component because their shear force measurements of toughness were determined by both components. In our transverse strength tests, the influence of acetic acid on the perimysial connective tissue component is unequivocally demonstrated.
The decreased strength of the perimysial collagen network indicated by Fig. 3 could be due to any of the three mechanisms outlined in the introduction. The results of Gault (1985) suggest that the tenderizing mechanism is one of pH- induced swelling diluting out the amount of load-bearing material. Focusing on the perimysium, this corresponds to a smaller number of perimysial strands be- ing present per unit cross-sectional area in the marinated meat because the mus- cle fiber bundles have swollen. Assuming that perimiysial connective tissue is stronger than the swollen myofibrillar material, fewer perimysial strands per unit area would result in a lower breaking load per unit area (i.e., strength), even if the nature of the perimysial strands themselves had not been altered at low pH. In this case, the decrease in transverse strength between control and marinated meat at each temperature should be eliminated if the strength values are cor- rected for the swelling in the plane perpendicular to the testing direction that has taken place on marinating. Strength values were corrected for swelling to test this hypothesis. Because the swelling of the meat blocks was anisometric, it should be most accurate to correct for swelling changes in the plane that defines the cross-section of the transverse strength of the test pieces, i.e., by correcting for dimensional changes in thickness x length in the original (cooked blocks), rather than correcting for increased weight. The result of this correction pro- cedure is shown in Fig. 3b. Compensation for dimensional changes on marina- tion did not completely account for the differences between treated and untreated strength. Although the errors inevitable in measuring an easily deformable meat block could be partly responsible for this, it appears that there is a real weaken- ing of the perimysial connective tissue due to marinating. The tensile testing of isolated perimysium was performed to check this possibility, as discussed below.

MARINATION EFFECTS ON CONNECTIVE TISSUE 187
There is a slight suggestion, especially from Fig. 3a, that as well as diluting out the loadbearing perimysial collagen network, marination also tends to shift the pattern of changes in strength with cooking temperature downwards by ap- proximately 5 "C. This may be seen as a possible shift to the left of the peaks and troughs in the strength vs. temperature curve of marinated meat compared to control material in Fig. 3a.
Longitudinal Breaking Strength of Meat
Figure 3c indicates that the breaking strength of test specimens from the marinated blocks is less than that of those from the control blocks at all cooking temperatures. However, the strengths only begin to diverge markedly above 60 "C, the control specimens becoming stronger and the marinated specimens weaker. At 90 "C the marinated blocks were noticeably fragile in the direction perpendicular to the muscle fiber direction. Longitudinal gaps appeared whenever the blocks were handled, suggesting a severe degradation of the perimysium or of the endomysial-perimysial junction. In the transverse strength experiment, it proved impractical using conventional pneumatic grips to grip the 90 "C specimens due to this fragility.
To determine to what extent these differences are due to dimensional changes in the cooked blocks and to what extent due to alterations in the strength of the load-bearing components, it is again necessary to adjust the results for dimen- sional changes. The data were therefore corrected for dimensional changes in the cross-sectional area perpendicular to the direction of strain (width X thickness) in the cooked control and marinated blocks (Fig. 3d) to give a corrected breaking strength. The strengths at a cooking temperature of 60 "C or less were now seen to be virtually the same in the control and marinated specimens. Above 60 "C, the curves are still seen to diverge. The increase in this corrected strength of the unmarinated meat above 60 "C with increasing cooking temperature may be in- terpreted on the basis that the rate of accelerated conditioning has been found to increase to a maximum at 60°C but is virtually absent above this temperature (Davey and Gilbert 1976).
The corrected strength of the marinated meat is seen in Fig. 3d to decrease more or less steadily with increasing temperature up to 90°C. The corrected strength of the unmarinated meat is more or less constant with increasing cook- ing temperature to 60 "C above which is increased. The result of this is that only above 60 "C does the corrected strength of the marinated meat fall below that of the unmarinated samples. The reduction in the strength of meat due to marinating, when it is cooked to 60°C or less, is due entirely to the swelling ef- fect of marinating which is retained during cooking.

188 G.J. LEWIS AND P.P. PURSLOW
Stiffness of Isolated Perimysium
Figure 4 relates the load-extention behavior of an isolated perimysial specimen. The curve marked 0 was obtained with the specimen in 0.9% NaCl solution only. Curves marked 7 , 10 and 30 illustrate the load-extension behavior at 7, 10 and 30 min after the introduction of the 1% acetic acid in 0.9% NaCl bathing medium. In NaCl alone, the load-extension curve (curve 0) shows an in- itial compliance or ‘toe’ region, becoming stiffer at higher loads. This is thought to result for the straightening of crimps and reorientation of the crossed lattice of collagen fibers in the material (Lewis and Purslow 1989). As the time course of marination proceeded, the length of this toe region diminished, approaching a minimum at about 50 min. On removing the acetic acid, the toe region was re- established, approaching it original state after about 100 min.
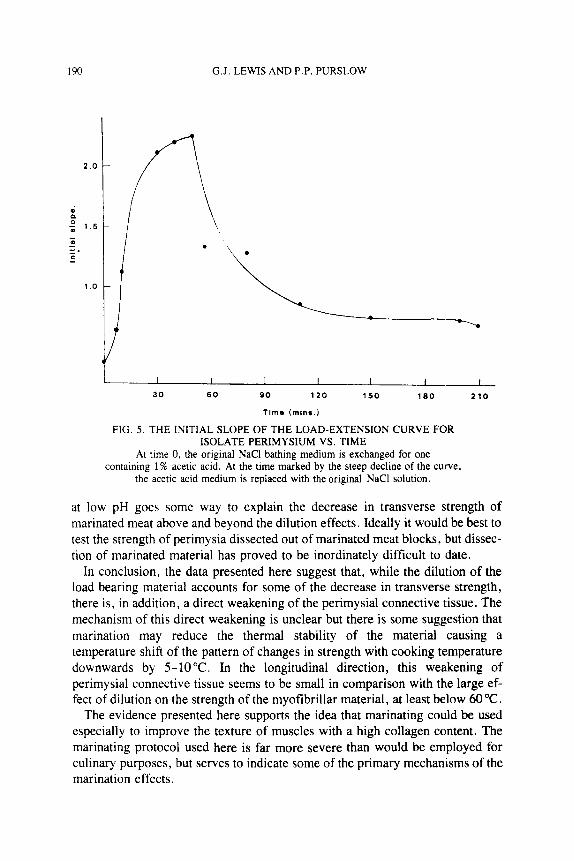
The process can also be illustrated by plotting the initial slope of the toe region with time (Fig. 5 ) . From this figure, it can be seen that the stiffening process began slowly, reaching a maximum rate between 7 and 10 min after introduction of the acetic acid, and then began to approach an equilibrium stiffness at some time after 50 min. On removing the acetic acid, the stiffness fell rapidly at first and then slowly approached an equilibrium value. This final equilibrium value is slightly above the initial toe region stiffness at the beginning of the experiment. Figure 5 shows that neither the stiffening process on adding the acetic acid or the reverse on its removal went to completion in the time duration of this experi- ment. The reversible loss of the toe region on lowering the pH, without a con- comitant change in the stiffness of the load-extention curve at loads above 5g (Fig. 4), strongly suggests that the changes in initial stiffness are due to the straightening out of the crimp in the collagen fibers, possibly due to their swell- ing laterally in the acetic acid. This will be investigated at a future stage by polarized light microscopy observations on the degree of crimping seen in perimysial collagen fibers at various pH values. The results in Fig. 4 and 5 show that perimysial connective tissue, by becoming stiffer at low extensions, actually increases its resistance to lateral swelling of the myofibrils at low pH. This may be responsible for some of the swelling capacity being expressed in longitudinal swelling of the muscle fibers, although this can only be a matter of conjecture at this stage.
Breaking Strength of Isolated Perimysia
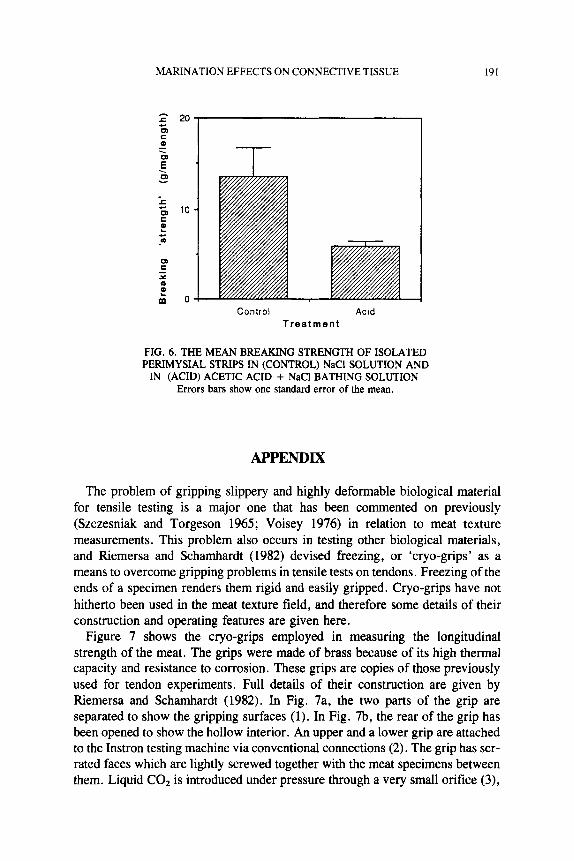
Figure 6 shows the mean breaking strengths of isolated strips of perimysium in NaCl only or in acetic acid + NaC1. The mean breaking strength of the NaCl on- ly (control) test specimens was significantly greater (P C 0.05) than those that had been marinated in acetic acid. None of the specimens in acetic acid had strengths above 8 .3 g/(mg/2.% length) while 5 out of 9 of the control specimens were stronger than this and, in one instance, as high as 32

MARINATION EFFECTS ON CONNECTIVE TISSUE 189
30 10 7 0
1 2
Extension (rnrn)
FIG. 4. LOAD-EXTENSION CURVES FOR ISOLATED PERIMYSIUM. Curve marked 0; specimen in NaCl solution only. Curves marked 7, 10,30 show the load-extension behaviour of the same specimen 7. 10 and 30 min after the introduction of an acetic acid + NaCl medium.
g/(mg/2.9mm length). The variation in the control specimen strengths was noticeably higher than in the marinated specimens. Acetic acid seemed to reduce the strength of all specimens to a minimum value. The conclusion must be that the marinating with 1 % acetic acid in 0.9% NaCl reduces the breaking strength of isolated perimysia. This gives some support to the inference from Fig. 3a that the dilution effect of swelling on the transverse strength of meat does not entirely explain the magnitude of the decrease in strength seen. Although the precise composition of marinating solutions used in the experiments on whole meat and isolated perimysium were different, the observation that perimysium is weaker
190 G.J. LEWIS AND P.P. PURSLOW
30 6 0 9 0 1 2 0 150 180 2 1 0
Time (rnlna.)
FIG. 5 . THE INITIAL SLOPE OF THE LOAD-EXTENSION CURVE FOR ISOLATE PERIMYSIUM VS. TIME
At time 0, the original NaCl bathing medium is exchanged for one containing 1 SJO acetic acid. At the time marked by the steep decline of the curve,
the acetic acid medium is replaced with the original NaCl solution.
at low pH goes some way to explain the decrease in transverse strength of marinated meat above and beyond the dilution effects. Ideally it would be best to test the strength of perimysia dissected out of marinated meat blocks, but dissec- tion of marinated material has proved to be inordinately difficult to date.
In conclusion, the data presented here suggest that, while the dilution of the load bearing material accounts for some of the decrease in transverse strength, there is, in addition, a direct weakening of the perimysial connective tissue. The mechanism of this direct weakening is unclear but there is some suggestion that marination may reduce the thermal stability of the material causing a temperature shift of the pattern of changes in strength with cooking temperature downwards by 5-10°C. In the longitudinal direction, this weakening of perimysial connective tissue seems to be small in comparison with the large ef- fect of dilution on the strength of the myofibrillar material, at least below 60 "C.
The evidence presented here supports the idea that marinating could be used especially to improve the texture of muscles with a high collagen content. The marinating protocol used here is far more severe than would be employed for culinary purposes, but serves to indicate some of the primary mechanisms of the marination effects.
MARINATION EFFECTS ON CONNECTIVE TISSUE
r ^ m e ul C Q) - . . r ul - 5 m ’0 2 e 00
m
Y a - e m O
Control Acid Treatment
191
FIG. 6 . THE MEAN BREAKING STRENGTH OF ISOLATED PERIMYSIAL STRIPS IN (CONTROL) NaCl SOLUTION AND
Errors bars show one standard error of the mean. IN (ACID) ACETIC ACID + NaCl BATHING SOLUTION
APPENDIX
The problem of gripping slippery and highly deformable biological material for tensile testing is a major one that has been commented on previously (Szczesniak and Torgeson 1965; Voisey 1976) in relation to meat texture measurements. This problem also occurs in testing other biological materials, and Riemersa and Schamhardt (1982) devised freezing, or ‘cryo-grips’ as a means to overcome gripping problems in tensile tests on tendons. Freezing of the ends of a specimen renders them rigid and easily gripped. Cryo-grips have not hitherto been used in the meat texture field, and therefore some details of their construction and operating features are given here.
Figure 7 shows the cryo-grips employed in measuring the longitudinal strength of the meat. The grips were made of brass because of its high thermal capacity and resistance to corrosion. These grips are copies of those previously used for tendon experiments. Full details of their construction are given by Riemersa and Schamhardt (1982). In Fig. 7a, the two parts of the grip are separated to show the gripping surfaces (1). In Fig. 7b, the rear of the grip has been opened to show the hollow interior. An upper and a lower grip are attached to the Instron testing machine via conventional connections (2). The grip has ser- rated faces which are lightly screwed together with the meat specimens between them. Liquid C 0 2 is introduced under pressure through a very small orifice (3),

I92 G.J. LEWIS AND P.P. PURSLOW
FIG. 7. CRYO-GRIPS EMPLOYED IN THE MEASUREMENT OF LONGITUDINAL MEAT STRENGTH (a) The serrated plates between which the meat is held. (b) Rear view of grip showing hollow chamber in which COl snow is formed. Numbers refer to features mentioned in the text.
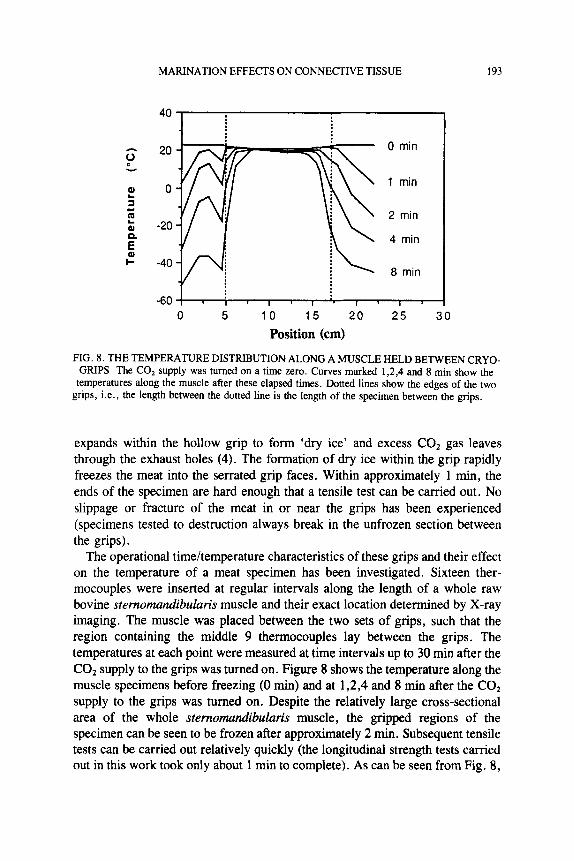
MARINATION EFFECTS ON CONNECTIVE TISSUE 193
40
20
0
-20
-40
-60 0 5 1 0 1 5 20 25 30
Position (cm) FIG. 8. THE TEMPERATURE DISTRIBUTION.ALONG A MUSCLE HELD BETWEEN CRYO-
GRIPS The CO, supply was turned on a time zero. Curves marked 1,2,4 and 8 min show the temperatures along the muscle after these elapsed times. Dotted lines show the edges of the two
grips, i.e., the length between the dotted line is the length of the specimen between the grips.
expands within the hollow grip to form ‘dry ice’ and excess C02 gas leaves through the exhaust holes (4). The formation of dry ice within the grip rapidly freezes the meat into the serrated grip faces. Within approximately 1 min, the ends of the specimen are hard enough that a tensile test can be carried out. No slippage or fracture of the meat in or near the grips has been experienced (specimens tested to destruction always break in the unfrozen section between the grips).
The operational timehemperature characteristics of these grips and their effect on the temperature of a meat specimen has been investigated. Sixteen ther- mocouples were inserted at regular intervals along the length of a whole raw bovine stemomundibuluris muscle and their exact location determined by X-ray imaging. The muscle was placed between the two sets of grips, such that the region containing the middle 9 thermocouples lay between the grips. The temperatures at each point were measured at time intervals up to 30 min after the COz supply to the grips was turned on. Figure 8 shows the temperature along the muscle specimens before freezing (0 min) and at 1,2,4 and 8 min after the COz supply to the grips was turned on. Despite the relatively large cross-sectional area of the whole stemomundibuluris muscle, the gripped regions of the specimen can be seen to be frozen after approximately 2 min. Subsequent tensile tests can be carried out relatively quickly (the longitudinal strength tests carried out in this work took only about 1 min to complete). As can be seen from Fig. 8,

194 G.J. LEWIS AND P.P. PURSLOW
there was very little cooling of the central portion of the meat specimen at times up to 8 min, i.e., long afier the time in which a test could be completed. The breaking strength of the unfrozen mid-region therefore gives a fair measurement of the strength of the material at normal room temperature.
REFERENCES
BOUTON. P.E. and HARRIS, P.V. 1972. The effects of cooking temperature and time on some mechanical properties of meat. J. Food Sci. 37. 140-144.
DAVEY, C.L. and GILBERT, K.V. 1976. The temperature coefficient of beef ageing. J. Sci. Fd. Agric. 27, 244-250.
GAULT, N.F.S. 1984. The influence of acetic acid concentration on the effi- ciency of marinading as a process for tenderizing beef. Proc. 30th Meeting of Europeari Meat Research Workers, pp. 184-185.
GAULT, N.F.S. 1985. The relationship between water-holding capacity and cooked meat tenderness in some beef muscles as influenced by acidic condi- tions below the ultimate pH. Meat Sci. 15, 15-30.
LEWIS, G.J. and PURSLOW, P.P. 1989. The strength and stiffness of perimysial connective tissue isolated from cooked beef muscle. Meat Sci. 26,
OFFER. G. and KNIGHT, P. 1988. The structural basis of water-holding in meat. Part 1 : General principles and water uptake in meat processing. In Development in Meat Science - 4 (R. Lawrie, ed.) pp. 63-171, Elsevier, Lon- don.
LOCKER, R.H. and DAINES, G.J. 1976. Transverse anisotropy in beef mus- cle. J . Sci. Fd. Agric. 27, 186-192.
PRIVALOV, P.L. 1982. Stability of proteins. Proteins which do not present a single cooperative system. Adv. Pror. Chem. 35, 1-104.
PURSLOW, P.P. 1985. The physical basis of meat texture: observations on the fracture behaviour of cooked bovine M. semitendinosus. Meat Sci. 12,39-60.
ROWE. R.W. 1977. The influence of the collagen fiber network of muscle on the compliance and tensile strength of muscle samples subjected to loads at right angles to the muscle fiber axis. Meat Sci. 1, 135-148.
RIEMERSA, D.J. and SCHAMHARDT, H.C. 1982. Technical note: The cryo- jaw, a clamp designed for in vitro rheology studies of horse digital flexor ten- dons. J . Biomechanics 15, 619-620.
SZCZESNIAK, A.S. and TORGESON, K.W. 1965. Methods of meat texture measurement viewed from the background of factors affecting tenderness. Ad\,. Food Res. 14, 33-165.
255-269.

MARINATION EFFECTS ON CONNECTIVE TISSUE I95
WENHAM, L.M. and LOCKER, R.H. 1976. The effect of marinading on beef.
VOISEY, P.W. 1976. Engineering assessment and critique of instruments used J . Sci. Fd. Agri. 27, 1079-1084.
for meat tenderness evaluation. J. Texture Studies 7, 11-48.




















