
Construct Validation of Experimental Manipulations in Social ...
Upload
independentCategory
view
1download
0

rience a nonsteady supply ofanother limiting resource, even in"steady-state" experiments. Changes in light availability, lightquality, and light-dark cycles favor certain species and altercommunity structure (Wall and Briand 1979; Chisholm 1981;Glover et al. 1986; Lewis et al. 1986). However, interactionsbetween light availability and nutrient limitation have rarelybeen tested by direct experimental manipulation (Ward andWetzel 1980).
The dominance of small phytoplankton species in certainlakes is thought to be the outcome of competition for limitingnutrients. Correlative and experimental data indicate that P lim-itation (e.g., high N:P ratios) favors smaller cells in freshwa-ters, particularly species of the cyanobacterium Sy nechoc occus(Suttle et al. 1987; Stockner and Shortreed 1988; Watson andMcCauley 1988;Wehr 1989). At P levels >0.2 pM, the sizespectrum usually shifts toward larger cells (Stockner andShortreed 1988; Wehr 1989). This balance among cell sizesmay also be affected by light, as well as nutrients. In order toresolve whether resource competition is important in any com-munity, one must determine both the effect of resource limi-tation and the intensity of resource depletion (Goldberg 1990).Among phytoplankton species, the mechanism behind differentpatterns in size structure is apparently a greater rate of inorganicPOI uptake in smaller cells during P-limited conditions (Leanand White 1983; Suttle and Harrison 1988).
In this paper, I test whether two key resources, P supply andlight availability, interactively limit the growth of phytoplank-ton and affect community structure in an oligotrophic lake.Experiments are run in situ with whole phytoplankton com-munities; treatments are single pulses of inorganic P and irra-diance shifts, using incubations at two depths. Treatments are
applied in a combined, or factorial, manner in order to assess
the separate and interactive effects of two important resourcegradients on the phytoplankton community. I evaluate effectsof competition in terms of changes in (l) size structure,(2) species composition, and (3) resource depletion.
Materials and Methods
Study Site
Experiments were carried out in Rye Lake, a moderately deeplake located in southeastern New York State (41'04'N,13"44'W). The lake basin was connected to Kensico Lake inthe early 1900's with the formation of Kensico Reservoir(890 ha, Z^u* - 50 m). The surface area of Rye Lake itself is-260 ha and maximum depth is -40 m. The system lies withina crystalline, gneiss-schist bedrock and levels of solublereactive P (SRP) and soluble total P are consistently low (<0.05and 0.25 pM, respectively). The basin stratifies in spring, andautumn turnover usually occurs in late October or November.Other physical and chemical features of the Kensico systemwere described previously (Wehr l99l). Samples wereprocessed at the laboratories of the Louis Calder Center,Fordham University (Armonk, NY), located -8 km from thestudy site.
Field Procedures and Experimental Design
Experiments were run on two 4-d periods, 23-27 September(during stratification) and2-6 November (after turnover) 1990.Subsequent physical and chemical analysis revealed that on theNovember dates, temperatures were isothermal (eliminatingtemperature effects in experimental treatments), but chemical
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
conditions were not uniform with depth. For the purposes ofthis paper, the term "after turnover" will be retained. On eachstarting date, a triplicate set of water chemistry and phytoplank-ton samples were collected from I and 15 m, and temperatureand light profiles were measured. A set of phytoplankton sam-ples for size-fractionated biomass estimates was also collectedand measured. The methods used were the same as outlinedbelow for experiment harvest periods. Light was measuredusing a Li-Cor (LI-185A) PAR meter fitted with a submersiblecosine probe, and temperature was recorded using a YSI tem-perature/O, meter.
A three-way factorial design was used in which communitydepth (l or 15 m), incubation depth (1 or 15 m), and a singleP pulse (0 or * 2 pM) were factor treatments (a total of eighttreatments x three replicates). The intent of this design wasto examine the separate and interacting effects of depth, light,and nutrients on the size structure and competition among phy-toplankton in a clearwater, oligotrophic system. The lake is Plimited, with relatively high levels of inorganic N (-30 pMNO3, 0.5 pM NH"; see also Wehr l99l). Effects of thesemanipulations on nutrient (POo, total P, NO3, NH) depletionwere also measured.
Water was collected with a peristaltic pump from the 1- and15-m depths (-90 and 17o surtace irradiance; labeled 1o) intotriplicate 4-L containers, which served as experimental micro-cosms. Prior to each experiment, larger zooplankton wereremoved using 202-pm Nitex mesh (to minimize container-exaggerated herbivory effects). Microcosms were attached toan anchored float near the deepest part of Rye Lake. Half themicrocosms were fertilized with a single pulse of KH,PO4(stock[P] : 4 mM) using syringes fitted with0.2-pmpore sizesterile filter units (Gelman acrodisc) to a final concentration of2 pM P. They were then fully mixed and attached to the floats.Bags were positioned at either 0.5 or 14 m, so that light pen-etration in the bags approximated levels measured in situ at the1- and 15-m depths, respectively.
After 4 d of incubation, bags were removed and phytoplank-ton were collected with a pump fitted with an in-line filter holdercontaining 20-pm Nitex mesh. Filters and the remaining filtratewere kept cold (4'C) and dark until returned to the laboratory.Water collected for nutrient analysis and phytoplankton iden-tification was pumped without prefiltration. Nutrient sampleswere immediately postfiltered (0.45-p,m pore size), stored coldon the boat, and frozen within 3 h of collection.
Laboratory Procedures
Size categories of phytoplankton follow earlier definitions(Sieburth et al. 1978; Wehr 1989). Briefly, microplankton arecells or colonies 20-200 pm, nanoplankton are 2-20 pm, andpicoplankton are 0.2-2 pm. The filtering methods (includingcare to prevent disruption of more fragile organisms) have beendescribed previously (Wehr 1989, 1990). Chlorophyll a (Chl a)was extracted by placing filters in vials containing 3-4 mL ofneutral 9OVo acetone (with MgCO.), grinding with a metal spat-ula, and extracting cold (<4"C) in the dark for 18-24 h.Extracts were made up to 5 mL and centrifuged; Chl o was thenmeasured spectrophotometrically (Shimadzu model UV-160)after correction for pheophytin a (Lorenzen 196'l; APHA 1985).
Whole phytoplankton samples were split in half; one aliquotwas preserved with Lugol's iodine and the other was preservedwith glutaraldehyde to final concentration of ZVo and filteredonto 25-mm black-stained Poretics filters (0.2-pm pore size).
931
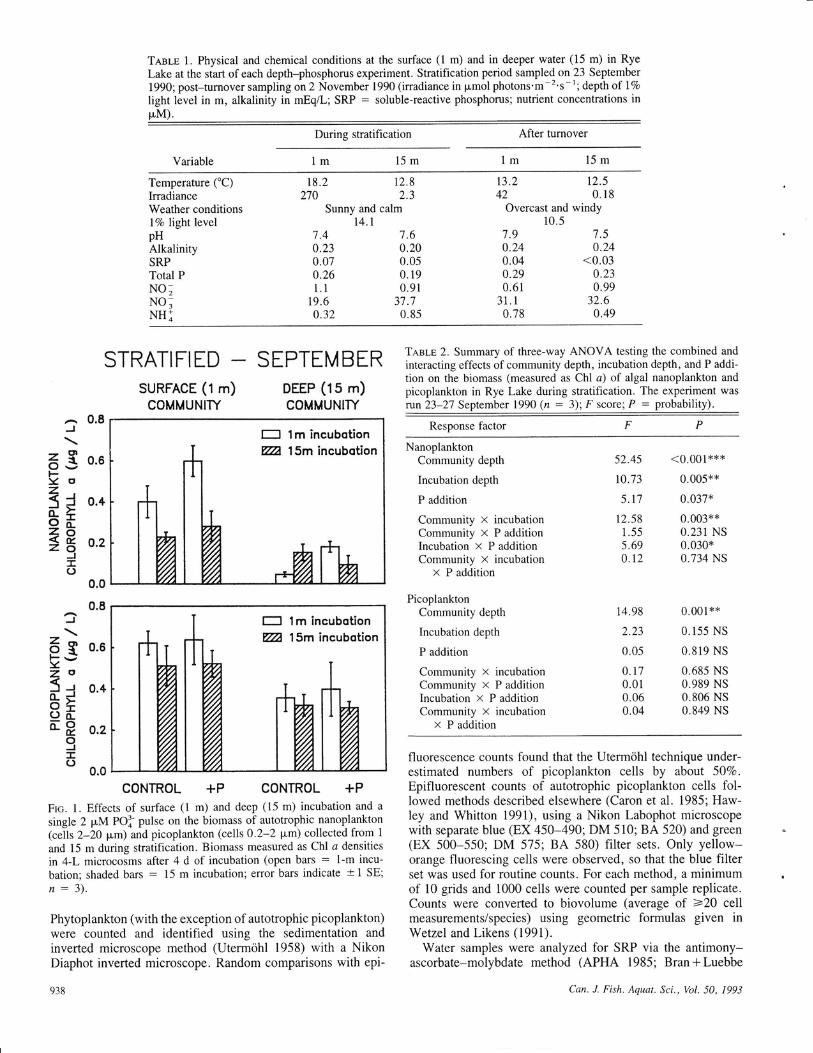
Tnsle 1. Physical and chemical conditions at the surface (l m) and in deeper water (15 m) in RyeLake at the start of each depth-phosphorus experiment. Stratification period sampled on 23 September
1990; post-turnover sampling on 2 November 1990 (irradiance in pmol photons'm-2's-1; depth of 17o
light level in m, alkalinity in mEq/L; SRP : soluble-reactive phosphorus; nutrient concentrations inpM).
During stratification After turnover
Variable lm 15m 1m 15m
Temperature ('C)IrradianceWeather conditionsl7o light levelpHAlkalinitySRPTotal PNO;NO,NHI
Sunny and calmt4.t
Overcast and windy10.5
t8.22't0
t2.82.3
7.60.200.050. 19
0.913"t.'l0.85
t3.242
7.90.240.040.290.61
31.10.78
t2.50.18
't.5
0.24<0.03
0.230.99
32.60.49
7.40.230.070.261.1
t9.60.32
0.8J
- ctrz<()*>FYOz{itL4otrz6<oa2.6
J-o
0.6
0.4
0.2
0.0
3p_t:<201 __,dJo+()dEP
oJIo
STRATI FI IDSURFACE (1 m)
COMMUNITY
SEPTEM BERDEEP (1s m)COMMUNITY
0.6
0.4
o.2
CONTROL +P CONTROL +PFrc. l Effects of surface (l m) and deep (15 m) incubation and a
single 2 pM PO]- pulse on the biomass of autotrophic nanoplankton
lceils 2-20 pm) and picoplankton (cells 0.2-2 pm) collected from 1
and 15 m during stratification. Biomass measured as Chl a densities
in 4-L microcosms after 4 d of incubation (open bars : l-m incu-
bation; shaded bars : 15 m incubation; error bars indicate -r 1 SE;
n=3).
Phytoplankton (with the exception of autotrophic picoplankton)were counted and identified using the sedimentation and
inverted microscope method (Utermohl 1958) with a NikonDiaphot inverted microscope. Random comparisons with epi-
938
Tnsle 2. Summary of three-way ANOVA testing the combined andinteracting effects of community depth, incubation depth, and P addi-tion on the biomass (measured as Chl a) of algal nanoplankton and
picoplankton in Rye Lake during stratification. The experiment was
^" 2:-ZZ S.pt.-U"t
Response factor
NanoplanktonCommunity depth
Incubation depth
P addition
Community x incubationCommunity x P additionIncubation x P additionCommunity x incubation
x P addition
PicoplanktonCommunity depth
Incubation depth
P addition
Community x incubationCommunity x P additionIncubation x P additionCommunity x incubation
x P addition
fluorescence counts found that the Utermohl technique under-estimated numbers of picoplankton cells by about 507o.Epifluorescent counts of autotrophic picoplankton cells fol-lowed methods described elsewhere (Caron et al. 1985; Haw-ley and Whitton 1991), using a Nikon Labophot microscopewith separate blue (EX 450-490; DM 510;BA 520) and green(EX 500-550; DM 575; BA 580) filter sets. Only yellow-orange fluorescing cells were observed, so that the blue filterset was used for routine counts. For each method, a minimumof 10 grids and 1000 cells were counted per sample replicate.Counts were converted to biovolume (average of >20 cellmeasurementsispecies) using geometric formulas given inWetzel and Likens (1991).
Water samples were analyzed for SRP via the antimony-ascorbate-molybdate method (APHA 1985; Bran*Luebbe
52.45
t0.73
5.1'7
12.581.555.690.12
14.98
2.23
0.05
0.170.010.060.04
<0.001*'r{'i
0.005**
0.037*
0.003**0.231 NS0.030*0.734 NS
0.001**
0.155 NS
0.819 NS
0.685 NS0.989 NS0.806 NS0.849 NS
l-] 1m incubotion@ 15m incubotion
El I m incubotionw 15m incubotion
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
+ 2 pM P1m COMMUNITY l5m COMMUNITY
TOT_P
}TO P ADDED1m COMMUNITY l5m COMMUNIfi
FJ
b0{{
tu
II
\6hob-t
.t
0.,I
0
,-1
bo-t
z
80
70
60
50
40
30
20
l00
TIME (DAYS)
1m COMMUNITY
TIME (DAYS)
15m COMMUNITY
TIME (DAYS)
1m COMMUNITY
TIME (DAYS)
15m COMMUNITY600
500
400
200
,J
hnrt
z
600
500
400
300
200
100
0
I"----:ANO3-N
NO3-N
NH4-N 1 NH4-N100
o r,rE (DAYS) 4
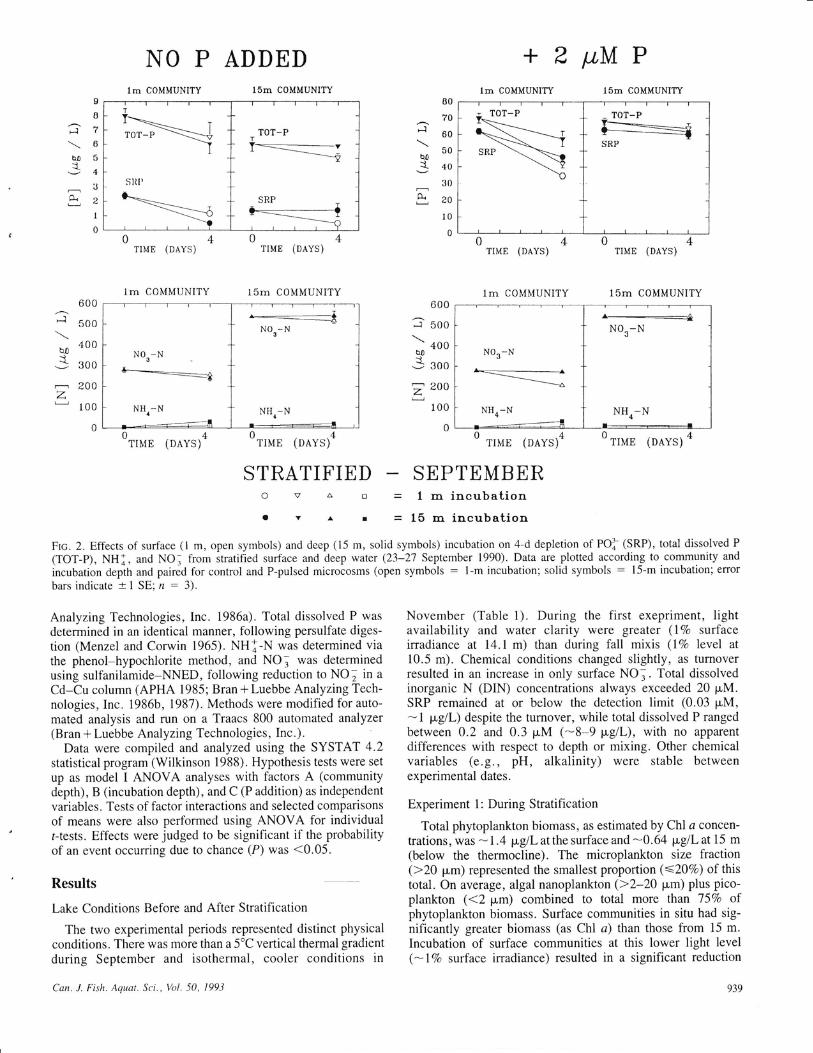
Frc. 2. Effects of surface (l m, open symbols) and deep (15 m, solid symbols) incubation on 4-d depletion of POI'(SRP), total dissolved P
(TOT-P), NHl, and NO. from itratified surface and deep water (23-27 September 1990). Data are plotted according to community and
incubation depth and paired for control and P-pulsed microcosms (open symbols : l-m incubation; solid symbols : l5-m incubation; error
bars indicate +1 SE; n : 3).
or,tE 1ra,vs;a
STRATIFIEDOvao
aval
Analyzing Technologies, Inc. 1986a). Total dissolved P wasdetermined in an identical manner, following persulfate diges-tion (Menzel and Corwin 1965). NHI-N was determined viathe phenol-hypochlorite method, and NOi was determinedusing sulfanilamide-NNED, following reduction to NOJ in a
Cd-Cu column (APHA 1985; Bran * Luebbe Analyzing Tech-nologies, Inc. 1986b, 1987). Methods were modified for auto-mated analysis and run on a Traacs 800 automated analyzet(Bran * Luebbe Analyzing Technologies, Inc.).
Data were compiled and analyzed using the SYSTAT 4.2statistical program (Wilkinson 1988). Hypothesis tests were set
up as model I ANOVA analyses with factors A (communitydepth), B (incubation depth), and C (P addition) as independentvariables. Tests offactor interactions and selected comparisonsof means were also performed using ANOVA for individualt-tests. Effects were judged to be significant if the probabilityof an event occurring due to chance (P) was <0.05.
Results
Lake Conditions Before and After Stratification
The two experimental periods represented distinct physicalconditions. There was more than a 5"C vertical thermal gradient
during September and isothermal, cooler conditions in
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
0 .4TIME (DAYS)
SEPTEMBER= 1 m incubation
= 15 m. incubation
November (Table l). During the first exepriment, lightavailability and water clarity were greater (|Vo surfaceirradiance at 14.1 m) than during fall mixis (l7o level at10.5 m). Chemical conditions changed slightly, as tumoverresulted in an increase in only surface NO, . Total dissolvedinorganic N (DIN) concentrations always exceeded 20 pM.SRP remained at or below the detection limit (0.03 pM,- I p"glL) despite the turnover, while total dissolved P rangedbetween 0.2 and 0.3 p.M (-8*9 pg/L), with no apparentdifferences with respect to depth or mixing. Other chemicalvariables (e.g., pH, alkalinity) were stable betweenexperimental dates.
Experiment 1: During Stratification
Total phytoplankton biomass, as estimated by Chl 4 concen-trations, was - L4 p,glLatthesurface and -0.64 pg/L at 15 m(below the thermocline). The microplankton size fraction(>20 pm) represented the smallest proportion (<207o) of thistotal. On average, algal nanoplankton ()2-20 pm) plus pico-plankton (<2 pm) combined to total more than 75Vo ofphytoplankton biomass. Surface communities in situ had sig-nificantly greater biomass (as Chl a) than those from 15 m.Incubation of surface communities at this lower light level(-l7o surtace irradiance) resulted in a significant reduction
NO _N3-r-i
NH -N1
s39
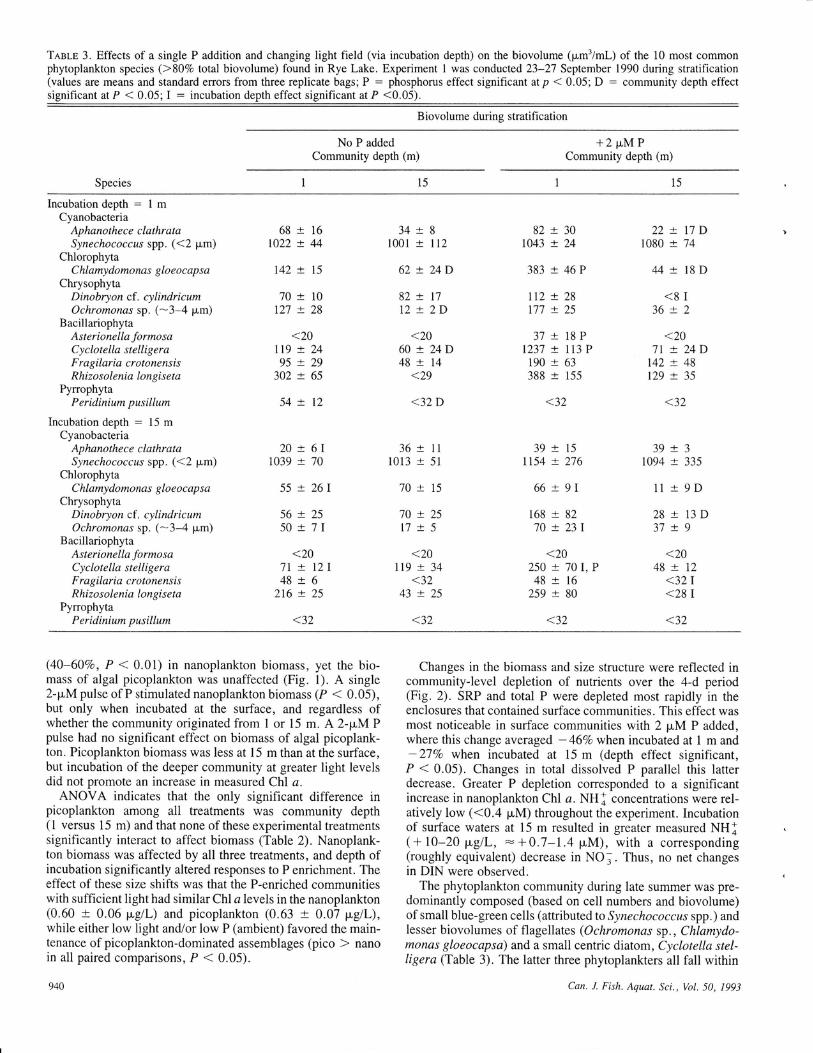
Tnsr-B 3. Effects of a single P addition and changing light field (via incubation depth) on the biovolume (pm'/ml-) of the 10 most commonphytoplankton species (>80Vo total biovolume) found in Rye Lake. Experiment I was conducted 23-27 September 1990 during stratification(values are means and standard errors from three replicate bags; P = phosphorus effect significant atp < 0.05; D : community depth effectsignificant at P < 0.05; I = incubation depth effect significant at P <0.05).
Biovolume during stratifi cation
No P addedCommunity depth (m)
+2pMPCommunity depth (m)
Species 15t5
lncubationdepth: I mCyanobacteria
Aphanothece clathrataSynechococcu,s spp. (<2 pm)
ChlorophytaChlamydomonas gloe ocap sa
ChrysophytaDinobryon cf . cylindricumOchromonas sp. (-3*4 pm)
BacillariophytaAsterionella formosaCyclotella stelligeraF ragilaria crotone nsisRhizosolenia Iongise ta
PyrrophytaPeridinium pusillum
Incubation depth : 15 mCyanobacteria
Aphanothece clathrataSynechococcus spp. (<2 pm)
ChlorophytaChlamydomonas gloeocap sa
ChrysophytaDinobryon cf . cylindricumOchromonas sp. (-3-4 pm)
BacillariophytaAsterionella formosaCyclotella stelligeraF ra gilaria c ro t o ne nsisRhizosolenia longis eta
PyrrophytaPeridinium pusillum
68 -f 16LQ22 -r 44
142 -r 15
70 -f l0127 +- 28
<20119 -r 2495 -+ 29
302 + 65
20 -ts
1039 -f
55 -f
6I70
26t
<2071 4'l2I48'r 6
216 -r 25
<32
36 -rt0t3 -'-
<a 7)
39+15ll54 -r 276
66 -r 9l
<32
22 'r 1'l D1080 t 74
44+l8D
<207t -r 24D
142 + 48129 'r 35
<32
39-r31094 + 335
11 +9D
<2048 -r 12
<321<28 r
<32
34-f81001 + 112
82t043
383
l2t77
371237
190388
2825
18Pl13 P63r55
+30-r 24
-r 46P
++
++++
28 + 13D3'7 -r 9
168 -l_ 8270 -r 23 I
<20250 -r 70 r, P48+16
259 -f 80
54 'r 12
-r 24D
+17-r 2D
<2060 -r 24D48 -r 14
<29
<32D
62
82t2
<8I36'r 2
70 'f
70 -f1"7 'r
56 -r 2550 -F 7I
1l51
15
255
<20119 -r 34
<3243 -r 25
1-) 1
(40-60Vo, P < 0.01) in nanoplankton biomass, yet the bio-mass of algal picoplankton was unaffected (Fig. l). A single2-pM pulse of P stimulated nanoplankton biomass (P < 0.05),but only when incubated at the surface, and regardless ofwhether the community originated from I or 15 m. A 2-pM Ppulse had no significant effect on biomass of algal picoplank-ton. Picoplankton biomass was less at l5 m than at the surface,but incubation of the deeper community at greater light levelsdid not promote an increase in measured Chl a.
ANOVA indicates that the only significant difference inpicoplankton among all treatments was community depth( I versus 15 m) and that none of these experimental treatmentssignificantly interact to affect biomass (Table 2). Nanoplank-ton biomass was affected by all three treatments, and depth ofincubation significantly altered responses to P enrichment. Theeffect of these size shifts was that the P-enriched communitieswith sufficient light had similar Chl a levels in the nanoplankton(0.60 -r 0.06 pgll-) and picoplankton (0.63 + 0.07 pClL),while either low light and/or low P (ambient) favored the main-tenanca of picoplankton-dominated assemblages (pico ) nanoin all paired comparisons, P < 0.05).
940
Changes in the biomass and size structure were reflected incommunity-level depletion of nutrients over the 4-d period(Fig. 2). SRP and total P were depleted most rapidly in theenclosures that contained surface communities. This effect wasmost noticeable in surface communities with 2 pM P added,where this change averaged -46Vo when incubated at I m and*2'7Vo when incubated at 15 m (depth effect significant,P < 0.05). Changes in total dissolved P parallel this latterdecrease. Greater P depletion corresponded to a significantincrease in nanoplankton Chl a. NH] concentrations were rel-atively low (<0.4 pM) throughout the experiment. Incubationof surface waters at 15 m resulted in greater measured NH j(+ 10-20 p"glL, - +0.7-1.4 pM), with a corresponding(roughly equivalent) decrease in NO,. Thus, no net changesin DIN were observed.
The phytoplankton community during late summer was pre-dominantly composed (based on cell numbers and biovolume)ofsmall blue-green cells (attributedto Synechococcus spp.) andlesser biovolumes of flagellates (Ochromonas sp., Chlamydo-monas gloeocapsa) and a small centric diatom, Cyclotella stel-ligera (Table 3). The latter three phytoplankters all fall within
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
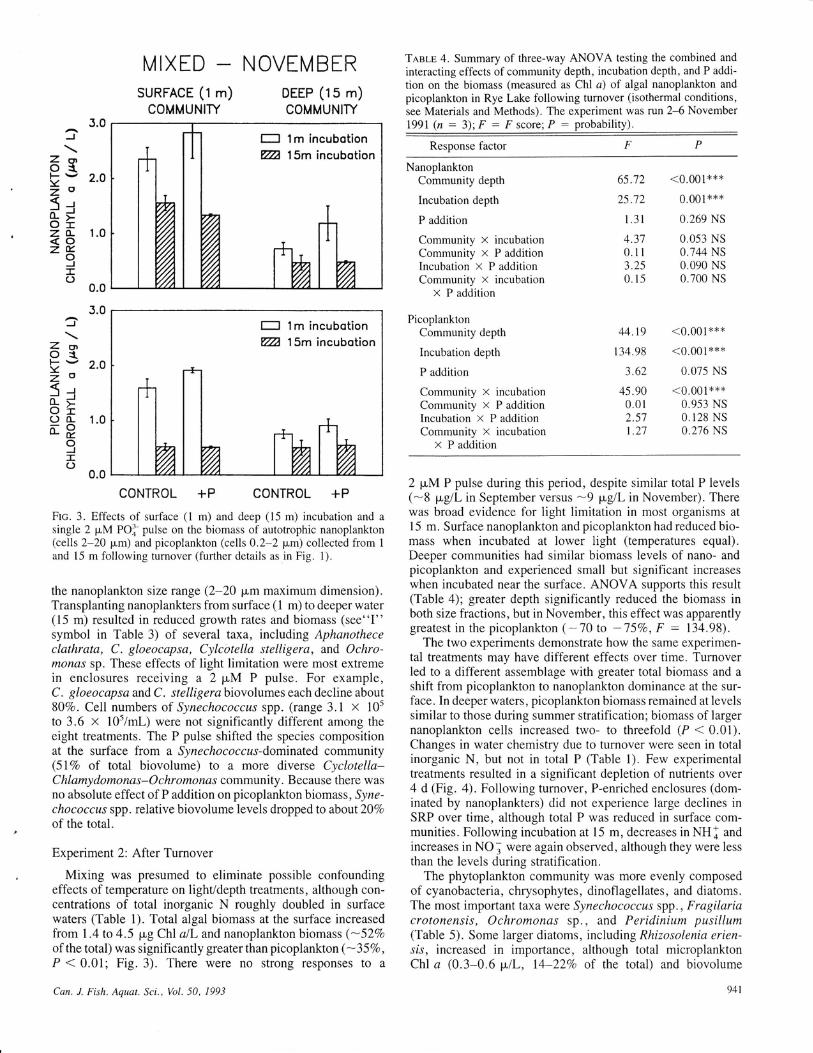
l-] 1m incubotionvl 15m incubotion
CONTROL +P CONTROL +P
Ftc. 3. Effects of surface (l m) and deep (15 m) incubation and a
single 2 pM POI pulse on the biomass of autotrophic nanoplankton(cells 2-20 pm) and picoplankton (cells 0.2-2 pm) collected from 1
and 15 m following turnover (further details as in Fig. 1).
the nanoplankton size runge (2-20 y,m maximum dimension).Transplanting nanoplankters from surface (l m) to deeper water(15 m) resulted in reduced growth rates and biomass (see"I"symbol in Table 3) of several taxa, including Aphanothececlathrata, C. gloeocapsa, Cylcotella stelligera, and Ochro-monas sp. These effects of light limitation were most extremein enclosures receiving a 2 p.M P pulse. For example,C. gloeocapsa and C. stelligera biovolumes each decline aboul80Vo. Cell numbers of Synechococcus spp. (range 3.1 x lOsto 3.6 x 10s/mL; were not significantly different among theeight treatments. The P pulse shifted the species compositionat the surface from a Synechococcus-dominated community(5lVo of total biovolume) to a more diverse CyclotelLa-Chlamydomonas-O chromonas community. Because there wasno absolute effect of P addition on picoplankton biomass, Syne-chococcus spp. relative biovolume levels droppedto about2}Voof the total.
Experiment 2: After Turnover
Mixing was presumed to eliminate possible confoundingeffects of temperature on light/depth treatments, although con-centrations of total inorganic N roughly doubled in surfacewaters (Table 1). Total algal biomass at the surface increasedfrom 1.4to4.5 pg ChlalL andnanoplanktonbiomass (-52Voof the total) was significantly greater than picoplankton(-35V0,P < 0.01; Fig.3). There were no strong responses to a
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
TnsLn 4. Summary of three-way ANOVA testing the combined and
interacting effects of community depth, incubation depth, and P addi-tion on the biomass (measured as Chl c) of algal nanoplankton and
picoplankton in Rye Lake following turnover (isothermal conditions,see Materials and Methods). The experiment was run 2-6 Novemberl99l (n : 3); F : F score; P : probability).
Response factor
NanoplanktonCommunity depth
Incubation depth
P addition
Community x incubationCommunity x P additionIncubation x P additionCommunity x incubation
x P addition
PicoplanktonCommunity depth
Incubation depth
P addition
Community x incubationCommunity x P additionIncubation x P additionCommunity x incubation
x P addition
2 pM P pulse during this period, despite similar total P levels(-8 pgll- in September versus -9 pg/L in November). Therewas broad evidence for light limitation in most organisms atI 5 m. Surface nanoplankton and picoplankton had reduced bio-mass when incubated at lower light (temperatures equal).Deeper communities had similar biomass levels of nano- andpicoplankton and experienced small but significant increaseswhen incubated near the surface. ANOVA supports this result(Table 4); greater depth significantly reduced the biomass inboth size fractions, but in November, this effect was apparentlygreatest in the picoplankton ( -70 to -757o, F : 134.98).
The two experiments demonstrate how the same experimen-tal treatments may have different effects over time. Turnoverled to a different assemblage with greater total biomass and a
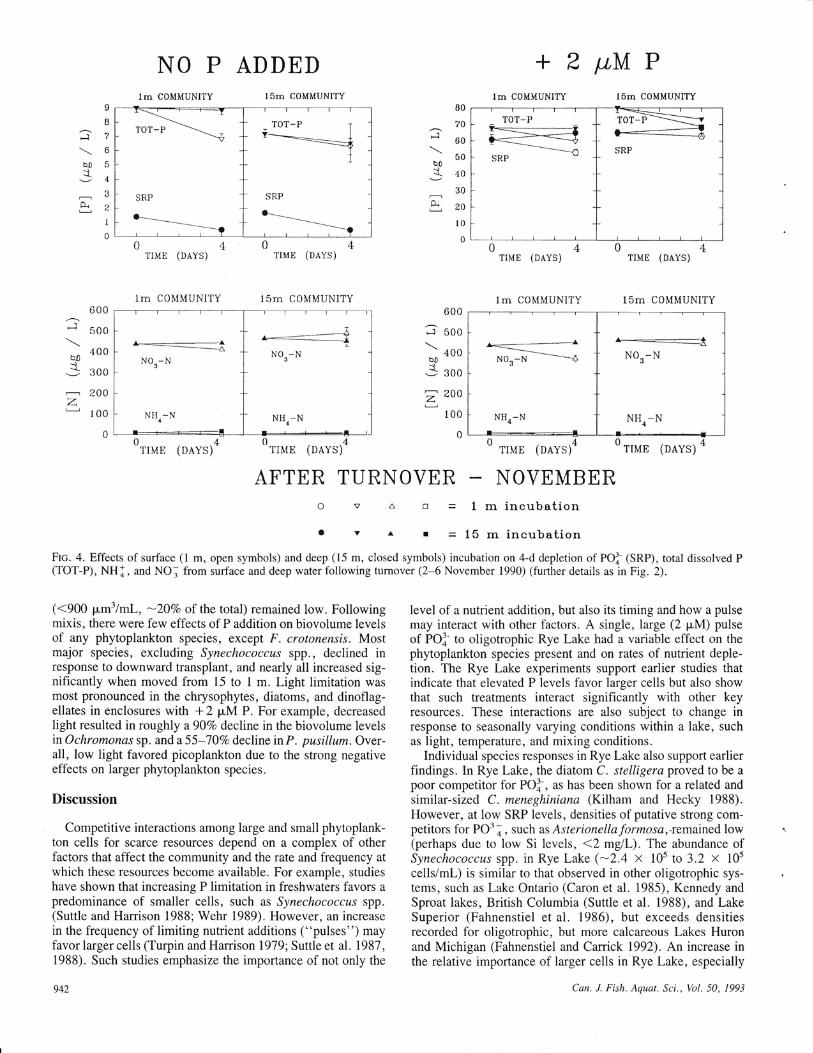
shift from picoplankton to nanoplankton dominance at the sur-face. In deeper waters, picoplankton biomass remained at levelssimilar to those dunng summer stratification; biomass of largernanoplankton cells increased two- to threefold (P < 0.01).Changes in water chemistry due to turnover were seen in totalinorganic N, but not in total P (Table 1). Few experimentaltreatments resulted in a significant depletion of nutrients over4 d (Fig. 4). Following turnover, P-enriched enclosures (dom-inated by nanoplankters) did not experience large declines inSRP over time, although total P was reduced in surface com-munities. Following incubation at l5 m, decreases in NH ] andincreases in NO. were again observed, although they were lessthan the levels during stratification.
The phytoplankton community was more evenly composedof cyanobacteria, chrysophytes, dinoflagellates, and diatoms.The most important taxa were Synechococcus spp. , Fragilariacrotonensis, Ochromonas sp., and Peridinium pusillum(Table 5). Some larger diatoms, including Rhizosolenia erien-sls, increased in importance, although total microplanktonChl a (0.3-0.6 p"lL, l4-22%o of the total) and biovolume
MIXED NOVEMBERSURFACE (1 m)
COMMUNITYDEEP (15 m)COMMUNITY
2.0
1.0
J\
Zaoi_FVv' 70{ --r.L<of. zo-- <ozEo
JIC)
3Zaoi.v- 2.07o{ --ro-ioa()L 1.0-osM
oJx()
0.0
3.0
65.12 <0.001r<,r.{.
25.12 0.001'rr<*
1.31 0.269 NS
4.37 0.053 NS0.11 0.744 NS3.25 0.090 NS0.15 0.700 NS
44.19 <0.001'6**
1 34,98 <0.001+**
3.62 0.075 NS
45.90 <0.001***0.01 0.953 NS2.5'7 0. r28 NS1.21 0.276 NS
l-] 1m incubotion71 15m incubotion
941
TOT-P
SRP 1' SRP
15m COMMUNITY
TIME (DAYS)
15rn COMMUNITY
or,ru lnnvs;a
AFTER TURNOVER
+ 2 pM Plm COMMUNITY 15m COMMUNITY
TOT-P TOT_P
SRP
NO P ADDED1m COMMUNITY
Fl
b0I
A
II
\6bo {>t
-*43
F.
I0
3 soo\
+oobo
-] eoo
z 200
100
F-l
h0e
z
80
70
60
50
40
30
20
10
0
rrME (DAYS)
1m COMMUNITY
o4TIME (DAYS)
TIME (DAYS)
1m COMMUNITY
rrMn (DAYS)
15m COMMUNITY600
500
400
300
200
100
0
No3-N -- 4 T No3-N
NH4-N T NH4-N
o v ^ r =lbrnincubationFIc. 4. Effects of surface (l m, open symbols) and deep (15 m, closed symbols) incubation on 4-d depletion of PO]- (SRP), total dissolved P(TOT-P), NHl, and NOI from surface and deep water following turnover (2-6 November 1990) (further details as in Fig. 2).
(<900 pm3/ml, -2OVo of the total) remained low. Followingmixis, there were few effects of P addition on biovolume levelsof any phytoplankton species, except F. crotonensis. Mostmajor species, excluding Synechococca.r spp., declined inresponse to downward transplant, and nearly all increased sig-nificantly when moved from 15 to I m. Light limitation wasmost pronounced in the chrysophytes, diatoms, and dinoflag-ellates in enclosures with + 2 pM P. For example, decreasedlight resulted in roughly a90Va decline in the biovolume levelsinOchromonas sp. and a55-10Vo decline inP. pusillum. Over-all, low light favored picoplankton due to the strong negativeeffects on larger phytoplankton species.
Discussion
Competitive interactions among large and small phytoplank-ton cells for scarce resources depend on a complex of otherfactors that affect the community and the rate and frequency atwhich these resources become available. For example, studieshave shown that increasing P limitation in freshwaters favors apredominance of smaller cells, such as Synechococca.r spp.(Suttle and Harrison 1988; Wehr 1989). However, an increasein the frequency of limiting nutrient additions ("pulses") mayfavor larger cells (Turpin and Harrison 1979; Suttle et al. 1987,1988). Such studies emphasize the importance of not only the
942
o ,,r, lonvs;4 o rlun (DAYS) 4
NOVEMBER1 m incubation
level of a nutrient addition, but also its timing and how a pulsemay interact with other factors. A single, large (2 pM) pulseof PO] to oligotrophic Rye Lake had a variable effect on thephytoplankton species present and on rates of nutrient deple-tion. The Rye Lake experiments support earlier studies thatindicate that elevated P levels favor larger cells but also showthat such treatments interact significantly with other keyresources. These interactions are also subject to change inresponse to seasonally varying conditions within a lake, suchas light, temperature, and mixing conditions.
Individual species responses in Rye Lake also support earlierfindings. In Rye Lake, the diatom C. stelligera proved to be apoor competitor for PO] , as has been shown for a related andsimilar-sized C. meneghiniana (Kllham and Hecky 1988).However, at low SRP levels, densities of putative strong com-petitors for PO3;, such as Asterionellaformosa,-remained low(perhaps due to low Si levels, (2 mg/L). The abundance ofSynechococcus spp. in Rye Lake (-2.4 x 105 to 3.2 x lOs
cells/ml) is similar to that observed in other oligotrophic sys-tems, such as Lake Ontario (Caron et al. 1985), Kennedy andSproat lakes, British Columbia (Suttle et al. 1988), and LakeSuperior (Fahnenstiel et al. 1986), but exceeds densitiesrecorded for oligotrophic, but more calcareous Lakes Huronand Michigan (Fahnenstiel and Canick 1992). An increase inthe relative importance of larger cells in Rye Lake, especially
_aNO _N
NO-N I 3"tNH1-N + NH.-N
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
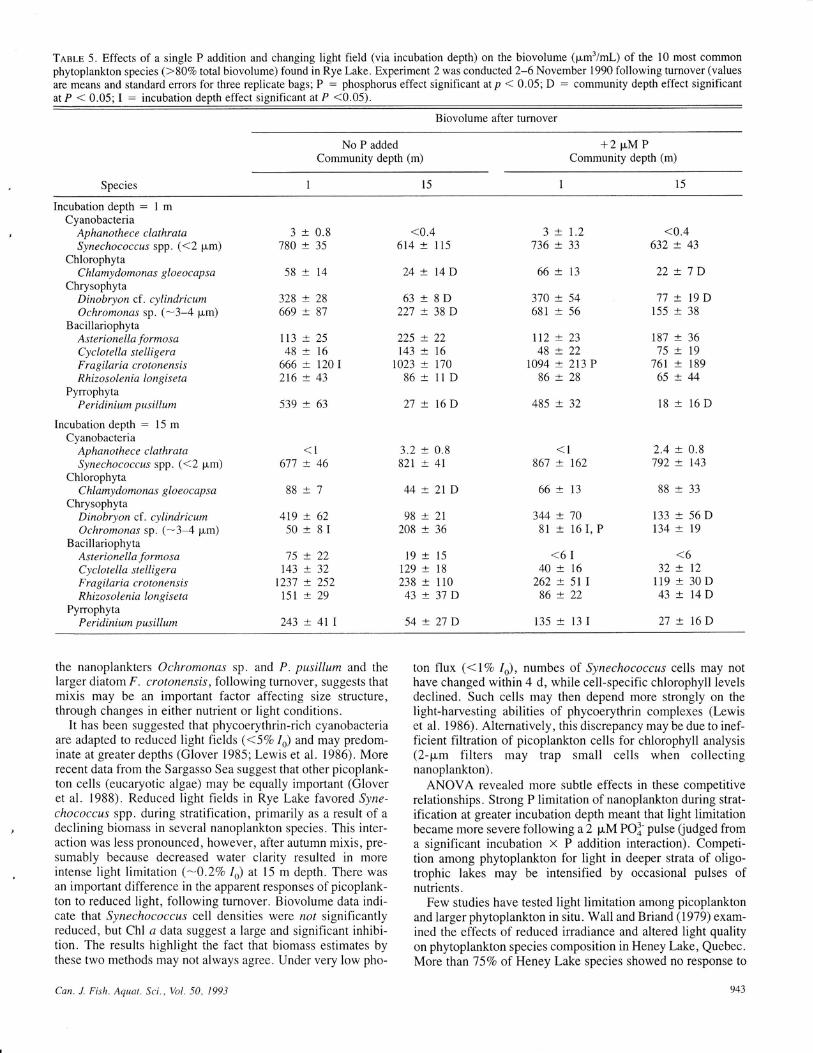
TaaI-e 5. Effects of a single P addition and changing light field (via incubation depth) on the biovolume (pm3/ml-) of the 10 most commonphytoplankton species (>807o total biovolume) found in Rye Lake. Experiment 2 was conducted 2-6 November 1990 following turnover (values
aremeansandstandarderrorsforthreereplicatebags; P: phosphoruseffectsignificantatp<0.05; D: communitydeptheffectsignificantutP < 0.05, I : in.ubutiot
Biovolume after tumover
No P addedCommunity depth (m)
+2pMPCommunity depth (m)
Species l5l5
Incubationdepth: lmCyanobacteria
Aphanothece clathrataSynechococcus spp. (<2 pm)
ChlorophytaC hlamydomonas gloeocap sa
ChIysophytaDinobryon cl. cylindricumOchromonas sp. (-3-4 pm)
BacillariophytaAsterionella formosaCycLotella steLligeraF ra gilaria crotone nsi s
Rhizos ole nia longis e taPynophyta
Peridinium pusiLlum
Incubation depth : 15 mCyanobacteria
Aphanothece cLathrataSynechococcus spp. (<2 pm)
ChlorophytaChlamydomonas gloeocap sa
ChrysophytaDinobryon cf . cylindricumOchromonas sp. (-3-4 pn)
BacillariophytaAsterionella formosaCyclotella stelligeraF ra gila r ia c rotone nsisRhizos ole nia Iongis e ta
PyrrophytaPeridinium pusiLlum
J
780
58
328669
11348
666216
3
736
66
+ 0.8+35
+14
+ 19D-r 38
+- 54+56
-r 23+22-+ 213 P-+ 28
63 -F 8D227 -r 38 D
++
++++
<0.4614 + 115
24 -r 14n
19 -f 15
129 + 18
238 -r 11043i3'lD
54+27D
-r 1.2-+ 33
-+ 13
<0.4632 + 43
22+'lD
539 :t
288',1
25t6120 I43
63
3-70
681
11248
t09486
4d) 'ts 32
7'7
155
18775
76165
18
225 + 22143 -r 16
1023 + 17086 -r 11D
2'l -r t6 D
3.2 + 0.8821 1- 41
44 -t zlD
r- 36-r 19-f 189+44
-f 16D
<l867 -ts 162
<1677 + 46
2.4 + 0.8792 1- 143
88 -f 3388-r7 66+ 13
344 -r 7081 + 16I,
<6 I40 -r 16
262 -+ 5l I86!22
135 + 13I 2'7 -r 16 D
4t950
75143
t237151
133 + 56D134 + 19
<632 4- l2
119 -r 30 D43 4- 74D
98!21208 + 36
-r 62+ 8I
+- 22-r 32-r 252-+ 2g
243 1- 4l I
the nanoplankters Ochromonas sp. and P. pusillum and thelarger diatom F. crotonensis, following turnover, suggests thatmixis may be an important factor affecting size structure,through changes in either nutrient or light conditions.
It has been suggested that phycoerythrin-rich cyanobacteriaare adapted to reduced light fields (<57o I) and may predom-inate at greater depths (Glover 1985; Lewis et al. 1986). Morerecent data from the Sargasso Sea suggest that other picoplank-ton cells (eucaryotic algae) may be equally important (Gloveret al. 1988). Reduced light fields in Rye Lake favored Syne-chococcus spp. during stratification, primarily as a result of a
declining biomass in several nanoplankton species. This inter-action was less pronounced, however, after autumn mixis, pre-sumably because decreased water clarity resulted in moreintense light limitation (-0.2% 1o) at 15 m depth. There wasan important difference in the apparent responses ofpicoplank-ton to reduced light, following turnover. Biovolume data indi-cate that Synechococcus cell densities were not significantlyreduced, but Chl c data suggest a large and significant inhibi-tion. The results highlight the fact that biomass estimates bythese two methods may not always agree. Under very low pho-
Can. J. Fish. Aquat. Sci., Vol. 50, 1993
ton flux (<lVo Io), numbes of Synechococcrrr cells may nothave changed within 4 d, while cell-specific chlorophyll levelsdeclined. Such cells may then depend more strongly on thelighrharvesting abilities of phycoerythrin complexes (Lewiset al. 1986). Alternatively, this discrepancy may be due to inef-ficient filtration of picoplankton cells for chlorophyll analysis(2-pm filters may trap small cells when collectingnanoplankton).
ANOVA revealed more subtle effects in these competitiverelationships. Strong P limitation of nanoplankton during strat-ification at greater incubation depth meant that light limitationbecame more severe following a 2 pM PO] pulse (udged froma significant incubation x P addition interaction). Competi-tion among phytoplankton for light in deeper strata of oligo-trophic lakes may be intensified by occasional pulses ofnutrients.
Few studies have tested light limitation among picoplanktonand larger phytoplankton in situ. Wall and Briand (1979) exam-ined the effects of reduced irradiance and altered light qualityon phytoplankton species composition in Heney Lake, Quebec.More than l5Va of Heney Lake species showed no response to

experimental manipulations , but certain species of green algae,colonial cyanobacteria, and diatoms were favored under "high-light" conditions (total irradiance >5Vo IJ.Two taxa (Peri-dinium sp., Gomphosphaeria lacustris) were competitivelyfavored at reduced light (<57o 1). It was not clear how nutrientlevels may have affected the results at Heney Lake or whethersmaller coccoid cyanobacteria were important. The most fre-quently occurring species in Heney were large, colonial, het-erocystous cyanobacteria; systems typified by these organismstend to have low densities of picoplankton (Watson andMcCauley 1988; Wehr 1991). In Rye Lake, light limitation wasat times a function of nutrient availability. Competition for lightmay only proceed in oligotrophic lakes following pulses of lim-iting nutrients or following holomixis. Given the demonstratedimportance of light limitation to many phytoplankters and theinteractive effects that can result following nutrient pulses, itshould be recognized in future studies that the alleviation ofone limiting resource (e.g., PO] ) may induce competition fora secondary resource. Autotrophic picoplankton, and in partic-ular PE-rich Synechococcr.r spp., may outcompete larger cellsat greater depths in clear, oligotrophic lakes because these cellsare less affected by P limitation than other species (e.g., Suttleand Harrison 1988; Wehr 1989) but also because of greatercompetitive abilities under low light levels. This may be par-ticularly important during brief periods of nutrient pulses whenan increase in the numbers of larger phytoplankton cells reduceswater clarity. SynechococcruJ spp. biovolume and total pico-plankton biomass (as Chl a) were unaffected by cross-trans-plants between the 1- and 15-m depths during late summer,suggesting that irradiance levels approaching l%o Iomay be suf-ficient to support autotrophic picoplankton in oligotrophiclakes.
Interacting effects of light and P limitation can also haveindirect effects on nutrient cycling and turnover. In Rye Lake,reduced irradiance (incubation at 15 m) gives Synechococcus acompetitive advantage. This resulted in increased rates of Pdepletion and apparent regeneration of NH I . Following a2 p"MPO] pulse, nanoplankter species predominate, and P depletionrates decline. Size structure of a phytoplankton community canhave dramatic effects on nutrient depletion and cycling (Leanand White 1983). In oligotrophic Flathead Lake (Montana),experimental data suggest that pelagic microorganisms domi-nate uptake and regeneration of NH| and PO]- during strati-fication (Dodds et al. 1991). Recognition of the factors thatregulate the competitive interactions among microbial speciestherefore is important for understanding nutrient dynamics inlakes. In Rye Lake, there is evidence for a kind of microbialcascade in which competition for light and/or P initiates a shifttowards smaller cells. This in tum alters the rates of cyclingand turnover of key nutrients within the water column.
Acknowledgements
This study was funded in part by the NSF grant DIR-9002145 andthe Louis Calder Center, Fordham University. I gratefully acknowl-edge Ms. Carol Stepczuk at the Kensico Laboratory of the NYCDepartment of Environmental Protection for logistic support andapproval to work at Rye Lake. I would also like to thank Ms. JeanetteSamaritan and Mr. Bryan Milstead for their helpful assistance in thefield.
References
AvsntcnN PusI-rc HEer-rH AssocrerroN (APHA). 1985. Standard methods forthe analysis of water and wastewater. 16th ed. APHA, Washington, DC.
944
BRAN+LUEBBE ANnlyzrNc TecuNoLocrEs, INc. 1986a. Ortho phosphate inwater and seawater. Industrial Method No- 812-86T.
BRAN+LUEBBE ANALYZTNG TEcHNoLocrEs, Inc. 1986b. Ammonia in waterand seawater. Industrial Method No. 804-86T.
BRAN + LUEBBE ANALYZTNG TEcHNoLocrEs, INc. 1987. Nitrate/nitrite in waterand seawater. Industrial Method No. 818-87T.
CARoN, D.A., F.R. PrcK, nNo D.R.S. LEAN. 1985. Chroococcoid cyanobac-teria in Lake Ontario: vertical and seasonal distribution during 1982. J.Phycol.2l: l'71-l'75.
Cutssolv, S.W. 1981. Temporalpatternsof celldivisioninunicellularalgae.In T. Platt [ed.] Physiological bases ofphytoplankton ecology. Can. Bull.Fish. Aquat. Sci.2l0: 150-181.
DoDDs, W. K., J.C. Pnrscu, nno B. K. Er-r-rs. I 99 l Seasonal uptake and regen-eration of inorganic nitrogen and phosphorus in a large oligotrophic lake:size-fractionation and antibiotic treatment. J. Plankton Res. l3: 1339-I 358.
FTHNET'rsrrEL, G.L., rNn H.J. Crnnrcr. 1992. Phototrophic picoplankton inLakes Huron and Michigan: abundance, distribution, composition, andcontribution to biomass and production. Can. J. Fish. Aquat. Sci. 43:379-388.
F,qsNrNsrrpr-, G.L., L. Srcro-Go,qo, D. ScAVrA, AND E.F. Sroenrilsn. 1986.Importance of picoplankton in Lake Superior. Can. J. Fish. Aquat. Sci.43:235-240.
Gr-ovEn, H. E. I 985. The physiology and ecology of the marine cyanobacterialgenus Synechococcus. Adv. Aquat. Microbiol. 3: 49-107 .
Glovrn, H.E., M.D. KlllEn, eNo R.L. Gu[leno. 1986. Light quality andoceanic ultraphytoplankters. Nature (Lond.) 319: 142-143.
Glovsn, H.E., B.B. PR6ZELrN, L. Cevpnell, AND M. WyMAN. 1988. Pico-and ultraplankton Sargasso Sea communities: variability and comparativedistributions of Synechococcus spp. and algae. Mar. Ecol. Prog. Ser. 49:121 -139.
Golonenc, D.E. 1990. Components of resource competition in plant com-munities, p.27-49.1z J.B. Grace and D. Tilman [ed.] Perspective onplant competition. Academic Press, Inc., San Diego, CA.
HAwLEy, G.R., eNo B.A. WsnroN. 1991. Surveyof algal picoplankton fromlakes in five continents. Verh. Int. Ver. Limnol. 24:1220-1222.
Krlnlu, P., AND R.E. HEcKy. 1988. Comparative ecology of marine and fresh-water phytoplankton. Limnol. Oceanogr. 33:'7'7 6-795.
LEAN, D.R.S., rNo E. WHnr. 1983. Chemical and radiotracer measurementsofphosphorus uptake by lake plankton. Can. J. Fish. Aquat. Sci. 40: 147-155.
LEwrs, M.R., R.E. WARNoCK, AND T. PLATT. 1986. Photosynthetic responseof marine picoplankton at low photon flux. 1n T. Platt and W.K.W. Lifed.] Photosynthetic picoplankton. Can. Bull. Fish. Aquat. Sci. 214: 235-250.
LonENzrn, C.l. 1961. Determination ofchlorophyll and pheo-pigments: spec-trophotometric equations. Limnol. Oceano gr. 12: 343*346...
MeNzEI-, D.W., aNo N. CoRwrN. 1965. The measurement of total phosphorusin seawater based upon the liberation of organically bound fractions bypersulfate oxidation. Limnol. Oceano gr. l0: 280-282.
Rtcrleps, R.E. 1990. Ecology. 3rd ed. W.H. Freeman & Co., New York,NY.
Slssunrg, J. McN., V. SruErrcm, eNo J. LrNz. 1978. Pelagic ecosystemstructure: heterotrophic compartments of the plankton and their relation-ship to plankton size fractions. Limnol. Oceanogr.23: 1256-1263.
Sot'lurn, U. 1985. Comparisons between steady state and non-steady statecompetition: experiments with natural phytoplankton. Limnol. Oceanogr.30: 335-346.
Soltltrn, U. [ED.] 1989. Phytoplankton ecology: succession in plankton com-munities. Springer-Verlag, New York, NY.
SrocrNEn, J.G. 1988. Phototrophic picoplankton: an overview from marineand freshwater ecosystems. Limnol. Oceano gr. 33 7 65-77 5.
SrocrNrn, J.G., eNo K.S. SHonrnmD. 1988. Response of Anabaena andSynechococcus to manipulation of nitrogen:phosphorus ratios in a lakefertilization experiment. Limnol. Oceanogr. 33: I 348- 1 361.
Surrlr, C.A., eNo P.J. HARRrsoN. 1988. Ammonium and phosphate uptakerates, N:P supply ratios, and evidence for N and P limitation in someoligotrophic lakes. Limnol. Oceanogr. 33: 186-202.
SurrLE, C.A., J.G. SrocxNtn, eNo P.J. HARRrsoN. 1987. Effects of nurrientpulses on community structure and cell size of a freshwater phytoplanktonassemblages in culture. Can. J. Fish. Aquat. Sci. 44 1768-l'174.
Surrle, C.A., J.G. SrocxNEn, K.S. Suonrnlpo, nNo P.J. HrnnrsoN. 1988.Time courses of size-fractionated phosphate uptake: are larger cells bettercompetitors for pulses of phosphate than smaller cells? Oecologia 74: 57 1-516.
TunprN, D.H., eNo P.J. HARRrsoN. 1979. Limiting nutrient patchiness and itsrole in phytoplankton ecology. J. Exp. Mar. Biol. Ecol. 39: l5l-166.
Can. J. Fish. Aquat. Sci., Vol. 50, 1993

UTERMOHL, H. I 958. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Int. Ver. Limnol. Mitt. 9: l-38.
Wnll, D. , AND F. BRIAND. 1979. Response of lake phytoplankton communitiesto in situ manipulations of light intensity and colour. J. Plankton Res. l:103*l 12.
W,cno, A. K., AND R. G. WETZEL. I 980. Interactions of light and nitrogen sourceamong planktonic bluegreen algae. Arch. Hydrobiol. 90: l-25.
Wnrsox, S.,,qNo E. McCeulev. 1988. Contrasting patterns of net- and nano-plankton production and biomass among lakes. Can. J. Fish. Aquat. Sci.45:915-920.
Wesn, J.D. 1989. Experimental tests of nutrient limitation in freshwaterpico-plankton. Appl. Environ. Microbiol. 55: 1605-l6l l.
Weun, J.D. 1990. Predominance of picoplankton and nanoplankton ineutrophicCalder Lake. Hydrobiologia 203: 35-44.
WEHR, J.D. 1991. Nutrient- and grazer-mediated effects on picoplankton andsize structure in phytoplankton communitres. Int. Rev. Gesamten Hydro-biol.76: 643-656.
Wntzel, R.G.,,cNo G.E. LrxrNs. 1991. Limnological analyses.2nd ed.Springer-Verlag, New York, NY.
Wtt-xtNsoN, L. 1988. SYSTAT: the system for statistics. Systat, Inc., Evans-ton, IL.
Can. J. Fish. Aquat. Sci., Vol. 50, 1993 945
Copyright © 2022 FDOKUMEN




















