Influence of Top-Down and Bottom-Up Manipulations on the R-BT065 Subcluster of -Proteobacteria, an...
10
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 2005, p. 2381–2390 Vol. 71, No. 5 0099-2240/05/$08.000 doi:10.1128/AEM.71.5.2381–2390.2005 Copyright © 2005, American Society for Microbiology. All Rights Reserved. Influence of Top-Down and Bottom-Up Manipulations on the R-BT065 Subcluster of -Proteobacteria, an Abundant Group in Bacterioplankton of a Freshwater Reservoir Karel S ˇ imek, 1,2 * Karel Horn ˇa ´k, 1,2 Jan Jezbera, 1,2 Michal Mas ˇı ´n, 1 Jir ˇı ´ Nedoma, 1 Josep M. Gasol, 3 and Michael Schauer 4 Hydrobiological Institute of the Academy of Sciences of the Czech Republic, Na sa ´dka ´ch 7, CZ-37005 C ˇ eske ´ Bude ˇjovice, Czech Republic 1 ; Faculty of Biological Sciences, University of South Bohemia, CZ-37005 C ˇ eske ´ Bude ˇjovice, Czech Republic 2 ; Institut de Cie `ncies del Mar-CMIMA, CSIC, Pg. Marı ´tim de la Barceloneta, 37-49, E-08003 Barcelona, Spain 3 ; and Institute for Limnology of the Austrian Academy of Sciences, A-5310 Mondsee, Austria 4 Received 23 June 2004/Accepted 24 November 2004 We studied the effects of nutrient availability and protistan grazing on bacterial dynamics and community composition (BCC) in different parts of the canyon-shaped R ˇ ı ´mov reservoir (Czech Republic). The effects of protistan grazing on BCC were examined using a size fractionation approach. Water from the dam area with only bacteria (<0.8 m), bacteria and heterotrophic nanoflagellates (<5 m), or whole water were incubated in situ inside dialysis bags. Top-down or predator manipulations (size fractionation) were also combined with bottom-up or resource manipulations, i.e., transplantation of samples to the middle and upper inflow parts of the reservoir with increased phosphorus availability. Significant genotypic shifts in BCC occurred with transplantation as indicated by denaturing gradient gel electrophoresis. Using different probes for fluorescence in situ hybridization, we found that 10 to 50% of total bacteria were members of the phylogenetically small cluster of -proteobacteria (targeted with the probe R-BT065). These rod-shaped cells of very uniform size were vulnerable to predation but very fast growing and responded markedly to the different experimental manipulations. In all the grazer-free treatments, the members of the R-BT065 cluster showed the highest net growth rates of all studied bacterial groups. Moreover, their relative abundance was highly correlated with bacterial bulk parameters and proportions of bacteria with high nucleic acid (HNA) content. In contrast, increasing protistan bacterivory yielded lower proportions of R-BT065-positive and HNA bacteria substituted by increasing proportions of the class Actinobacteria, which profited from the enhanced protistan bacterivory. Our knowledge of bacterioplankton community composition (BCC) in a broad variety of aquatic systems has significantly advanced with the use of various molecular techniques (see, e.g., references 14, 15, 16 and 51). However, much less is known about which factors regulate BCC in general and of individual bacterial groups in particular (4, 35, 37). Further- more, the factors regulating BCC in highly dynamic aquatic systems or those with a large gradient of environmental con- ditions, such as river estuaries (3, 5, 7) or typical canyon- shaped reservoirs (11, 19, 26, 43), have received even less attention. Canyon-shaped reservoirs are highly heterogeneous ecosys- tems. Often they have relatively short water retention times and display strong downstream longitudinal gradients in the factors potentially controlling bacterial dynamics and BCC. Thus, very distinct environmental conditions can be found in the same water body and at the same time. This heterogeneity has been explored to study how BCC is affected by various resource supply rates or grazing pressure encountered along such gradients (11, 43). Top-down manipulations have been seen to alter the balance between bacterial production and mortality in previous studies conducted in the R ˇ ı ´mov reservoir (44, 46), generating shifts in BCC (45). Distinct top-down manipulations generated different BCC (46), illustrating the specific role of both protistan selective grazing and various bacterial growth strategies (also cf. references 17, 21, and 31) in shaping overall BCC. One general pattern that emerges from this previous work is that the impact of protistan grazing on BCC tends to be sig- nificant only under nutrient-limited conditions. Under such conditions, grazing pressure targeted to certain bacterial groups exceeds their growth rates, and changes in BCC result (43). In contrast, under resource-rich conditions (i.e., at the reservoir inflow), physiological shift-ups and accelerated bac- terial growth are observed, and the changes induced by in- creased resource supply overwhelm losses due to grazing (43). However, some groups of bacteria have the potential for in- creases in abundance in spite of the strong grazing pressure, in particular those strains having a “fast growth and division strat- egy” (cf. reference 31). The members of the R-BT065 cluster (a small group of the Rhodoferax sp. BAL47 cluster [51]; for the phylogenetic affil- iation, see reference 46) can be an example of such bacterial * Corresponding author. Mailing address: Hydrobiological Institute AS CR, Na sa ´dka ´ch 7, CZ-37005 C ˇ eske ´ Bude ˇjovice, Czech Republic. Phone: 420 38 7775873. Fax: 420 38 5310248. E-mail: ksimek@hbu .cas.cz. 2381
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Influence of Top-Down and Bottom-Up Manipulations on the R-BT065 Subcluster of -Proteobacteria, an...

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 2005, p. 2381–2390 Vol. 71, No. 50099-2240/05/$08.00�0 doi:10.1128/AEM.71.5.2381–2390.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Influence of Top-Down and Bottom-Up Manipulations on theR-BT065 Subcluster of �-Proteobacteria, an AbundantGroup in Bacterioplankton of a Freshwater ReservoirKarel Simek,1,2* Karel Hornak,1,2 Jan Jezbera,1,2 Michal Masın,1 Jirı Nedoma,1
Josep M. Gasol,3 and Michael Schauer4
Hydrobiological Institute of the Academy of Sciences of the Czech Republic, Na sadkach 7, CZ-37005Ceske Budejovice, Czech Republic1; Faculty of Biological Sciences, University of South Bohemia,
CZ-37005 Ceske Budejovice, Czech Republic2; Institut de Ciencies delMar-CMIMA, CSIC, Pg. Marıtim de la Barceloneta, 37-49, E-08003
Barcelona, Spain3; and Institute for Limnology of theAustrian Academy of Sciences, A-5310
Mondsee, Austria4
Received 23 June 2004/Accepted 24 November 2004
We studied the effects of nutrient availability and protistan grazing on bacterial dynamics and communitycomposition (BCC) in different parts of the canyon-shaped Rımov reservoir (Czech Republic). The effects ofprotistan grazing on BCC were examined using a size fractionation approach. Water from the dam area withonly bacteria (<0.8 �m), bacteria and heterotrophic nanoflagellates (<5 �m), or whole water were incubatedin situ inside dialysis bags. Top-down or predator manipulations (size fractionation) were also combined withbottom-up or resource manipulations, i.e., transplantation of samples to the middle and upper inflow parts ofthe reservoir with increased phosphorus availability. Significant genotypic shifts in BCC occurred withtransplantation as indicated by denaturing gradient gel electrophoresis. Using different probes for fluorescencein situ hybridization, we found that 10 to 50% of total bacteria were members of the phylogenetically smallcluster of �-proteobacteria (targeted with the probe R-BT065). These rod-shaped cells of very uniform sizewere vulnerable to predation but very fast growing and responded markedly to the different experimentalmanipulations. In all the grazer-free treatments, the members of the R-BT065 cluster showed the highest netgrowth rates of all studied bacterial groups. Moreover, their relative abundance was highly correlated withbacterial bulk parameters and proportions of bacteria with high nucleic acid (HNA) content. In contrast,increasing protistan bacterivory yielded lower proportions of R-BT065-positive and HNA bacteria substitutedby increasing proportions of the class Actinobacteria, which profited from the enhanced protistan bacterivory.
Our knowledge of bacterioplankton community composition(BCC) in a broad variety of aquatic systems has significantlyadvanced with the use of various molecular techniques (see,e.g., references 14, 15, 16 and 51). However, much less isknown about which factors regulate BCC in general and ofindividual bacterial groups in particular (4, 35, 37). Further-more, the factors regulating BCC in highly dynamic aquaticsystems or those with a large gradient of environmental con-ditions, such as river estuaries (3, 5, 7) or typical canyon-shaped reservoirs (11, 19, 26, 43), have received even lessattention.
Canyon-shaped reservoirs are highly heterogeneous ecosys-tems. Often they have relatively short water retention timesand display strong downstream longitudinal gradients in thefactors potentially controlling bacterial dynamics and BCC.Thus, very distinct environmental conditions can be found inthe same water body and at the same time. This heterogeneityhas been explored to study how BCC is affected by variousresource supply rates or grazing pressure encountered along
such gradients (11, 43). Top-down manipulations have beenseen to alter the balance between bacterial production andmortality in previous studies conducted in the Rımov reservoir(44, 46), generating shifts in BCC (45). Distinct top-downmanipulations generated different BCC (46), illustrating thespecific role of both protistan selective grazing and variousbacterial growth strategies (also cf. references 17, 21, and 31)in shaping overall BCC.
One general pattern that emerges from this previous work isthat the impact of protistan grazing on BCC tends to be sig-nificant only under nutrient-limited conditions. Under suchconditions, grazing pressure targeted to certain bacterialgroups exceeds their growth rates, and changes in BCC result(43). In contrast, under resource-rich conditions (i.e., at thereservoir inflow), physiological shift-ups and accelerated bac-terial growth are observed, and the changes induced by in-creased resource supply overwhelm losses due to grazing (43).However, some groups of bacteria have the potential for in-creases in abundance in spite of the strong grazing pressure, inparticular those strains having a “fast growth and division strat-egy” (cf. reference 31).
The members of the R-BT065 cluster (a small group of theRhodoferax sp. BAL47 cluster [51]; for the phylogenetic affil-iation, see reference 46) can be an example of such bacterial
* Corresponding author. Mailing address: Hydrobiological InstituteAS CR, Na sadkach 7, CZ-37005 Ceske Budejovice, Czech Republic.Phone: 420 38 7775873. Fax: 420 38 5310248. E-mail: [email protected].
2381

opportunists. They can be abundant (up to 50% of total bac-teria) and are found in many European and North Americanlakes (51, 52). The R-BT065 cluster contains sequences re-trieved through cloning from lakes Grosse Fuchskuhle andGossenkollesee (15), the Columbia River (6), and the RımovReservoir, with a minimum similarity within the cluster of�96% (46). Little is known about the ecology and the trophicrole of the members of this cluster in any environment. Ourpreliminary data indicate that they are very fast-growing rod-shaped cells of uniform size which are vulnerable to predation.While proportions of this group are stable or slightly increasewith enhanced protistan bacterivory, they grow particularlywell in bacterivore-free treatments (43, 46). The R-BT065 clus-ter must involve one or more bacterial phylotypes with a “fastgrowth and division strategy,” thus representing true opportu-nistic strategists among the reservoir bacterioplankton.
In this study, we examined in detail the dynamics and growthcapabilities of the members of the R-BT065 cluster as well asthe overall shifts in BCC and activity structure under severaldifferent scenarios. Bacterioplankton were subjected to shiftsin both bottom-up and top-down control in the canyon-shapedRımov reservoir. Shifts in BCC were monitored by using flu-orescence in situ hybridization (FISH) with oligonucleotideprobes and by denaturing gradient gel electrophoresis(DGGE). We estimated the proportions of bacteria with highnucleic acid (HNA) and low nucleic acid content cytometri-cally and rates of grazing-induced bacterial mortality. Ourgoals were (i) to assess short-term variability in BCC as afunction of resource supply and predation rates, (ii) to deter-mine the role of the R-BT065 cluster in changes in BCC andoverall bacterioplankton dynamics; and (iii) to examine howthe dynamics of the members of this cluster were related tochanges in the proportions of HNA bacteria in the assemblage.
MATERIALS AND METHODS
Study site and experimental design. The experiment was conducted in thecanyon-shaped, mesoeutrophic Rımov Reservoir (South Bohemia [see reference46 for details]). On 20 May 2002, 40-liter samples from the dam area (chlorophylla concentration, 4.5 �g liter�1) were collected from a depth of 0.5 m and mixedin a 50-liter plastic container. The experiment was conducted from 20 to 24 Mayat three experimental sites along the longitudinal profile of the reservoir (as-signed as dam, middle, and river) during the clear-water phase of the lake. Watertemperatures were 18 to 20°C at the dam and middle parts and 13 to 16°C at theriver inflow. The experimental design combined top-down and bottom-up ma-nipulations with plankton samples (see reference 43 for details). Briefly, dupli-cated, unfiltered �5-�m and �0.8-�m treatments were incubated in the dam sitein dialysis bags with the latter two size fractions also incubated in parallel inbottles (see below). Also, �5-�m and �0.8-�m treatments originating from thedam site were transplanted into middle and river sites and incubated in dialysisbags. With this protocol, we ensured varied grazing pressures on the bacteria aswell as nutrient supply rates.
We used a size fractionation approach to alter levels of bacterivory in naturalpopulations (for details, see references 43 and 46) so that bacterioplankton wassubjected to either negligible (�0.8 �m) or high (�5 �m) levels of protistanbacterivory. To assess the influence of resource availability, aliquots of size-fractionated plankton samples (see below) collected at the dam area were incu-bated in both dialysis bags (relatively free exchange of low-molecular-weightcompounds) and bottles (no nutrient penetration). This was done at the DAMsite characterized by moderate primary production (PP) of 16 �g of C liter�1 h�1
and a rather low concentration of dissolved reactive phosphorus (DRP) of 2 �gliter�1. A parallel set of samples of the �0.8-�m and �5-�m fractions wastransplanted and incubated in dialysis bags in the middle part of the reservoir(highest PP, 31 �g of C liter�1 h�1; enhanced DRP concentration, 23 �g liter�1),and finally, a third set of dialysis bags was incubated in the river inflow of the
reservoir (moderate PP of 14 �g of C liter�1 h�1 but the highest concentrationof DRP at 59 �g liter�1).
The size fractions represented (i) “bacterivore-free” treatment via filtrationthrough 0.8-�m-pore-size filters (47-mm diameter; OSMONIC Inc., Livermore,Calif.) which were assumed to remove all bacterivorous protists, (ii) “increasedbacterivory” treatment via filtration through 5.0-�m-pore-size filters which re-moved the predators of heterotrophic nanoflagellates (HNF), and, in the DAMsite also, (iii) control, untreated sample. For more details concerning filtrationprocedures, see the work of Simek et al. (44, 46). The fractions incubated indialysis bags are designated throughout the text as �5-�m, �0.8-�m, and “un-filtered” (UNF) treatments, respectively, and were completed with an identifi-cation of the site of the incubation, i.e., “dam,” “middle,” or “river.” These waterfractions were placed in �2.5-liter pretreated (deionized water rinsed andboiled) dialysis tubes (diameter, 75 mm; molecular weight cutoff, 12,000 to16,000 Da; Poly Labo, Switzerland). The samples incubated at the dam site in2-liter glass bottles are also named with an identification of the type of theincubation, i.e., “bottle.” All dialysis bag and bottle treatments were incubated ata depth of 0.5 m, oriented horizontally in open Plexiglas holders. Samples (�300to 450 ml) were taken from each dialysis bag and bottle at 0, 24, 48, 72, and 96 h.
Bacterial abundance and biomass. Samples were fixed with formaldehyde (2%[vol/vol] final concentration), stained with DAPI (4�,6�-diamidino-2-phenylin-dole) (final concentration, 0.1 �g ml�1, wt/vol), and enumerated by epifluores-cence microscopy (Olympus BX 60). Cell volumes were measured using theLUCIA D (Lucia 3.52 [resolution, 750 by 520 pixels, 256 grey levels]; LaboratoryImaging, Prague, Czech Republic) semiautomatic image analysis system as pre-viously described in detail by Posch et al. (32). Bacterial carbon biomass wascalculated according to the method of Norland (29).
Bacterial production. Bacterial production was measured via thymidine incor-poration modified from the method described by Riemann and Søndergaard (34)and described in detail by Simek et al. (46). Site-specific empirical conversionfactors between the thymidine incorporation rate and bacterial cell productionrate were determined using data from the duplicate �0.8-�m treatments. Thecell production rate was calculated from the slope of the increase of ln bacterialabundance over time (0, 24, and 48 h). Our determined ECFs, 0.84 1018, 1.58 1018, 1.36 1018, and 1.16 1018 cells mol�1 of thymidine incorporated forbottle incubations and dam, middle, and river dialysis bag incubations, respec-tively, were used for calculations.
PCR and DGGE analysis. The bacterial 16S rRNA genes in the samples wereamplified by filter PCR according to the protocol of Kirchman et al. (22). Samplefixation and filtration were performed as for preparing filters for FISH onpolycarbonate filters. As templates, small pieces (�5 mm2) of the 0.2-�m poly-carbonate filters were used. The reactions (50-�l volume) contained 400 �Meach deoxynucleoside triphosphate, 0.3 �M each primer, 3 mM MgCl2, 1 PCRbuffer, 1.25 units of Taq DNA Polymerase (QIAGEN), and 1.5 �l of bovineserum albumin (stock solution, 10 mg ml�1; SERVA). We used the bacterialspecific primer 358f (5�-CCT ACG GGA GGC AGC AG-3�), with a 40-bp GCclamp, and the universal primer 907r (5�-CCG TCA ATT CCT TTR AGTTT-3�), which amplifies a 550-bp DNA fragment of bacterial 16S rRNA (27). ThePCR was performed with a Primus 96plus thermal cycler (MWG Biotech) usingthe following program: initial denaturation step at 94°C for 5 min, 30 cycles ofdenaturation (at 94°C for 1 min), annealing (at 55°C for 1 min), and extension (at72°C for 2.5 min), and a final extension step at 72°C for 5 min. PCR productswere verified and quantified by agarose gel electrophoresis with a standard in thegel (Low DNA Mass ladder; GIBCO BRL). The products of three PCRs werepooled for each sample.
DGGE was carried out with a DCode Universal Mutation Detection system(Bio-Rad) as described previously by Muyzer et al. (27). We used 6% polyacryl-amide gels with a gradient of DNA-denaturing agent from 20% to 80% (100%denaturing agent is 7 M urea and 40% deionized formamide). Comparableamounts of PCR product were loaded for each sample, and the gel was run at 100V for 16 h at 60°C in 1 TAE buffer (40 mM Tris [pH 7.4], 20 mM sodiumacetate, 1 mM EDTA). The gel was stained with the nucleic acid stain SybrGold(Molecular Probes) for 45 min, rinsed with 1 TAE buffer, removed from theglass plate to a UV transparent gel scoop, and visualized in a UV transillumi-nator (LTF-Labortechnik, Wasserburg, Germany) with BioCapt software (LFT-Labortechnik). High-resolution images (768 by 568 pixels, 8-bit dynamic range)were saved as computer files (see reference 38 for details).
FISH with rRNA-targeted oligonucleotide probes. Analysis of planktonic BCCwas carried out by in situ hybridization with group-specific Cy3-labeled oligonu-cleotide probes on membrane filters (1). To assess proportions of the Actinobac-teria group (HGC69a probe), a modified catalyzed reporter deposition-FISHprotocol was applied (see references 30 and 39 for details). Six different group-specific oligonucleotide probes (ThermoHybaid, Germany) were targeted to the
2382 SIMEK ET AL. APPL. ENVIRON. MICROBIOL.

domain Bacteria (EUB338), to � and subclasses of the class Proteobacteria (theBET42a and GAM42a probes, respectively), to a narrower subcluster of the�-proteobacteria (R-BT065), to the Cytophaga/Flavobacterium group (CF319a),and to the Actinobacteria group (HGC69a). After the whole procedure, the filtersections were stained with DAPI and the proportions of hybridized bacterial cellswere enumerated using an epifluorescent microscope (Olympus AX70 Provis).
Flow cytometric sample analysis. Subsamples were preserved with 1% para-formaldehyde plus 0.05% glutaraldehyde (final concentration), frozen in liquidnitrogen, and stored at �80°C. The samples were later thawed, stained for a fewminutes with Syto13 (Molecular Probes) at 2.5 �M, and run in a desktop FAC-SCalibur flow cytometer (Becton Dickinson). We used MilliQ water as sheathfluid, samples were run at low speed (approximately 18 �l min�1), yellow-green0.92-�m Polysciences latex beads were used as internal standards, and the fixedsamples were diluted two to four times with MilliQ water to keep the rate ofparticle passage below 500 particles per second to avoid coincidence (12). Bac-teria were detected by their signature in a plot of side scatter versus greenfluorescence, and we used the percentage of HNA (%HNA) bacteria as a simpleway of describing the activity structure of the bacterial community (12, 13).
Protozoan grazing and enumeration. Protozoan grazing on bacteria was esti-mated using fluorescently labeled bacterioplankton (FLB) (42) concentratedfrom the reservoir water (see reference 46 for details). HNF and ciliate (onlyunfiltered treatment) FLB uptake rates were determined in short-term FLBdirect-uptake experiments with FLBs equal to 10 to 15% of natural bacterialconcentration. Subsamples for protozoan enumeration and tracer ingestion de-terminations (see reference 46 for details) were fixed with Lugol’s formaldehydedecolorization technique (42). Five-milliliter (for flagellates) or 20-ml (for cili-ates) subsamples were stained with DAPI, filtered through 1-�m black Poreticsfilters, and inspected via epifluorescence microscopy. To estimate total proto-zoan grazing, we multiplied the average uptake rates of ciliates and flagellates bytheir in situ abundances as previously described (46).
Chemical parameters. Dissolved organic carbon was analyzed on a ShimadzuTOC-5000A analyzer, and total and dissolved reactive phosphorus were deter-mined spectrophotometrically according to a method described previously byKopaeek and Hejzlar (23).
Statistical analysis. After arc-sine data transformation, two-way analysis ofvariance (ANOVA) was used to compare changes in the proportions of the
respective genotypic groups of bacteria (as a percentage of total DAPI-stainedbacteria) at 24, 48, 72, and 96 h induced by the top-down manipulation (i.e., sizefractionation) and by the different bottom-up supply rates in the dam, middle,and river sites. Using duplicate data on cell number increase in �0.8-�m treat-ments, net doubling time and growth rate of different bacterial subgroups werecalculated for intervals with exponential growth using nonlinear regression. Wethen tested (F test) for significant differences in growth rates of the bacteriatargeted by the R-BT065 probe compared to those of total bacterioplankton andother phylogenetic groups of bacteria. All statistics analyses were performedusing GraphPad Prism (GraphPad Software, San Diego, Calif.).
RESULTS
Bacterial and flagellate community dynamics and activity.In all treatments, bacterial numbers and biomass covariedclosely but with remarkably lower values found in the bottleincubation (Fig. 1). In the grazer-free (�0.8-�m) dam, middle,and river dialysis bag treatments, bacterial numbers reachedsimilar maxima and doubling times of 16 to 20 h (Table 1).However, the same treatment, incubated at the dam site in anonpermeable bottle, showed much lower maximal abundanceand longer bacterial doubling times of �42 h.
Different trends were evident in the �5-�m treatments (Fig.1). The increase in bacterial abundance observed during thefirst 48 to 72 h stopped, and then, with HNF exponentialgrowth during the second half of the experiment (Fig. 2),bacterial abundances decreased markedly at 96 h. The onlyexception was the river �5-�m treatment, where bacterialnumbers grew throughout the whole period, probably due tothe inefficient grazing control by slow-growing HNF (cf. Fig. 1and 2). The unfiltered treatment (dam UNF) showed fairly
FIG. 1. Time course changes in bacterial abundance and biomass in different size fractionation treatments of �0.8 �m, �5 �m, and UNFreservoir water collected and exposed in dialysis bags at the dam area of the reservoir compared to the changes in the same �0.8-�m and �5-�mtreatments but incubated in bottles or transplanted upstream to the middle and river inflow of the reservoir and incubated there in dialysis bags.Values are means for two replicate treatments. Error values are not shown to allow for a better visualization of the changes.
VOL. 71, 2005 HIGHLY ACTIVE SMALL SUBCLUSTER OF �-PROTEOBACTERIA 2383

similar trends in all microbial parameters as observed in thecorresponding dam �5-�m treatment (Fig. 1 and 2), except forslightly lower values of bacterial biomass and production thatmight be related to the presence of metazooplankton grazing(20 cladocerans liter�1 [data not shown]).
Bacterial production (BP) in the grazer-free (�0.8-�m)treatments (Fig. 2) roughly reflected nutrient availability in theincubation site or treatment. Thus, in the bottle, BP was lowestand showed a decreasing trend. In contrast, much higher val-ues and overall increasing trends were found in all dialysis bagincubations, with maxima of BP gradually increasing from themore P-limited dam site, through the middle site, and to theP-rich river site.
In all grazer-enhanced (�5-�m) and dam UNF treatments,changes in BP were strongly affected by the grazer population
dynamics and bacterivory (Fig. 2). Thus, in most cases, BPincreased only during the first 48 to 72 h and then markedlydropped due to grazing by the fast-growing HNF populations,peaking at 96 h (�10 103 to 59 103 individuals ml�1). Theonly exception was the river �5-�m treatment, where HNFreached only �4 103 individuals ml�1 and grazed �10% ofBP at 96 h. This fact, together with the nutrient-rich conditionsin the river site, yielded a continuous increase of BP through-out the study period. In contrast, in all other grazer-presenttreatments, total bacterivory approached (dam �5 �m), orexceeded, the level of BP at 96 h, sometimes severalfold (Fig.2); see, e.g., these parameters in the bottle �5-�m treatment.
Changes in BCC. During the course of the study, the ma-nipulations resulted in treatment- and site-specific changes inBCC as shown by both the fingerprinting (DGGE) and rRNA-
FIG. 2. Time course changes in bacterial production (Bact. prod.) and total protistan bacterivory in different size fractionation treatments ofreservoir water collected at the dam area and incubated in bottles or in dialysis bags at the dam, middle, and river parts of the reservoir (for moredetails, see the Fig. 1 legend). Values are means for two replicate treatments. Note that “total protistan grazing” refers only to HNF bacterivoryin all but the unfiltered treatment incubated at the dam site (DAM UNF), where �20% of total grazing was caused by the ciliates present in thesample (data not shown).
TABLE 1. Net doubling times of ungrazed bacterioplankton and different bacterial subgroupsa
Incubationsite
Doubling time (h)Interval (h)
Bacterioplankton R-BT065 BET42a GAM42a CF319a HGC69a
Dam 20** 8 13** 11 23* 23* 0–48Bottle 42 19 29 60 32 35 0–48Middle 20** 9 13 8 12* 18 0–48River 16 9 11 10 14 12 24–72
a Shown are net doubling times of ungrazed bacterioplankton and different bacterial subgroups in the predator-free treatments (�8 �m) exposed at the dam areain dialysis bags (dam) and bottles compared to those transplanted and incubated in dialysis bags in the middle and river inflow parts of the reservoir. The values aremeans for two duplicate treatments. Rates were calculated for intervals with exponential growth using nonlinear fit of the duplicated data followed by testing significantdifferences in doubling times of the bacteria targeted by the R-BT065 probe compared to other bacterial groups. *, P � 0.05; **, P � 0.01.
2384 SIMEK ET AL. APPL. ENVIRON. MICROBIOL.

targeted oligonucleotide probes (Fig. 3 and 4). The initialDGGE banding pattern indicated that size fractionation itselfhad a negligible effect on BCC (Fig. 3). However, the incuba-tion of the different size fractions and their transplantationupstream into areas differing in primary production and Pavailability induced marked shifts in BCC.
In the control samples at time zero, 19 DGGE bands weredetected. At the end of the experiment, we found between 8and 20 DGGE bands in each sample. The smallest band num-ber was commonly found in both �0.8-�m and �5-�m riverincubated treatments, suggesting a resource supply effect ondiversity. Another general pattern was that the bacterivore-free treatments differed from the corresponding bacterivore-enhanced treatments, with the most marked differences de-tected within the bottle and middle treatments, where HNFexerted the strongest grazing pressure in the �5-�m treat-ments.
For the six different rRNA-targeted oligonucleotide probeswe used in this study, cells detected by CF319a and HGC69ashowed no clear trend in any treatment or sample group exceptfor increases in proportions of HGC69a-positive cells in thebottle �5-�m treatment (Fig. 4). Commonly, cells detectedwith the general probe EUB338 and group-specific probesBET42a, R-BT065, and GAM42a showed initial increasesreaching maximal proportions at 48 or 72 h, and then, theyvariably decreased towards the end. However, the proportionof cells detected with probe GAM42a also showed differenttrends: in the bottle �0.8-�m treatment, they slowly de-creased, while in the river �0.8-�m treatment, they markedlyincreased during the second half of incubation.
A few additional common trends were also observed in allincubation sites. For instance, the proportion of HGC69a-detected cells (Fig. 4) was generally much higher in grazer-enhanced (�5 �m, 16% � 5% [mean � standard deviation])than in the grazer-free (�0.8 �m, 8% � 3%) treatments. In thelatter treatments, cells detected with the specific R-BT065
probe for a small subcluster of �-proteobacteria mostly over-grew other �-proteobacteria (BET42a probe) within the first48 h (e.g., middle and river in Fig. 4). Between 48 and 96 h ofthe experiment, we observed marked but not significantlyhigher proportions of the R-BT065 subcluster in the absence ofgrazing (34% � 12%) compared to that in its presence (20% �9%). The fast growth of the R-BT065-positive cells also yieldedthe shortest net doubling time of all studied groups (Table 1)that was significantly (P � 0.05; F test) faster than that of othergroups of bacteria at dam and middle sites, while these differ-ences were statistically insignificant in river and bottle incuba-tions. The R-BT065 subcluster displayed large growth poten-tial and responded highly significantly (two-way ANOVA) toboth types of manipulation, while cells detected with theprobes CF319a and GAM42a responded either to transplan-tation or to different grazing levels, respectively (for details,see Table 2). The members of the class Actinobacteria(HGC96a probe) were significantly affected by the manipula-tion of the grazing pressure (Table 2, cf. Fig. 4).
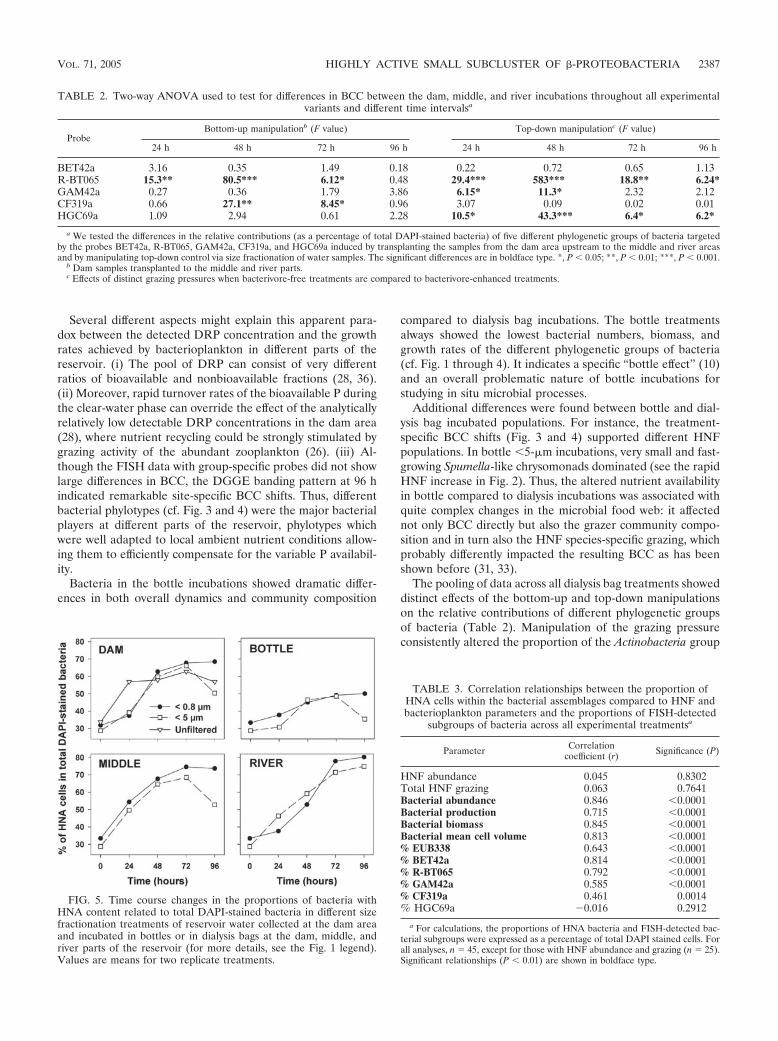
Bacterial cytometric fingerprints. In general, the %HNAincreased during the experiments, largely coinciding with in-creases in bacterial abundance and production (cf. Fig. 1, 2,and 5). In the grazer-free treatments, HNA cells were initially�30% of total DAPI-stained bacteria (Fig. 5) and reachedproportions of 50, 68, 75, and 80% in bottle, dam, middle andriver, respectively, at 96 h. Interestingly, all grazer-presenttreatments showed initially similar trends as did bacterivore-free ones, but between 72 and 96 h, along with markedlyincreasing protistan bacterivory, HNA bacteria diminished inall but the river �5-�m treatment (Fig. 5), the one with rela-tively low HNF bacterivory (Fig. 2).
Statistical relationships. When data were pooled across alltreatments, no significant relationship was found between theproportions of HNA bacteria and HNF abundance or bacte-rivory rates (Table 3). However, the %HNA bacteria was sig-nificantly and positively related to bacterial abundance, pro-duction, biomass, and mean cell volume (MCV). Thisindicated that the HNA bacteria are relatively large and likelythe most active ones, shaping the overall dynamics of thebacterial assemblage. More specifically, we also detected thatamong the phylogenetic bacterial groups studied, changes inthe proportions of cells targeted by BET42a and R-BT065probes explained the largest part of the variability in %HNAbacteria (66 and 63%, respectively) (Fig. 6B and C). Moreover,when only bacterivore-free treatments were considered, �70%of the variability in %HNA bacteria was explained by changesin the proportions of the narrow R-BT065 cluster (Fig. 6C).Overall, all but one probe-defined bacterial group (Actinobac-teria) significantly correlated with %HNA bacteria, and theimportance of the relationship (as the r value in Table 3)roughly corresponded to the net growth rate achieved in thebacterivore free-treatments shown in Table 1.
The clear importance of �-proteobacteria, particularly of theR-BT065 subcluster, as one of the forces driving the dynamicsof the reservoir bacterioplankton was also evidenced by thehighly significant correlations of these probe-defined bacterialsubpopulations and the basic bacterioplankton parametersshown in detail in Table 4.
FIG. 3. Negative image of the bacterial 16S rRNA gene fragmentsseparated by DGGE from the different size fractionation treatments of�0.8 �m and �5 �m at time zero compared to changes in bandingpatterns of samples incubated in bottles and in dialysis bags at the dam,middle, and river parts of the reservoir at the end of the incubation.
VOL. 71, 2005 HIGHLY ACTIVE SMALL SUBCLUSTER OF �-PROTEOBACTERIA 2385

DISCUSSION
Grazing has an effect on bacterial community composition. Ourresults show this through changes in the banding pattern in theDGGE (Fig. 3) and, as reported in previous studies, changes inthe relative importance of defined groups as determined usingFISH (cf. references 21, 44, and 46). However, by altering re-source through transplantation, our data also demonstrate theeffects of bottom-up factors in structuring BCC (Fig. 3).
We anticipated that the markedly different DRP concentra-tions at the three incubation sites would have a large influenceon various bacterial parameters. By comparing the two mostdistinct sites in terms of DRP concentrations, i.e., dam andriver, this assumption was only partially confirmed. For in-
stance, bulk bacterioplankton production and growth rate andphylotype-specific growth rates of bacteria were highest in theriver site, with no significant differences in growth rates com-pared to those of the fastest-growing cells, those detected withthe R-BT065 probe (Table 1). In contrast, R-BT065-positivecells grew in dam �0.8-�m treatment significantly faster thanother lineages, but the bulk bacterioplankton production andgrowth rate (Fig. 2 and Table 1) were only slightly lower thanthose of the river �0.8-�m treatment. Overall, these smalldifferences did not fully reflect the large site-specific differ-ences in DRP availability that were suggested in a previousexperiment to have marked effects on the dynamics of thereservoir bacterioplankton and virioplankton (43, 49).
FIG. 4. Time course changes in the phylogenetic composition of the bacterioplankton community in different size fractionation treatments ofreservoir water collected at the dam area and incubated in bottles or in dialysis bags at the dam, middle, and river parts of the reservoir (for moredetails, see the Fig. 1 legend). Shown are proportions of total FISH detection rates by the general bacterial probe EUB338, �-proteobacteria(BET42a), and the cells detected by the probe R-BT065, -proteobacteria (GAM42a), the Cytophaga/Flavobacterium group (CF319a), and theActinobacteria group (HGC69a). Values are means of two replicate treatments.
2386 SIMEK ET AL. APPL. ENVIRON. MICROBIOL.
Several different aspects might explain this apparent para-dox between the detected DRP concentration and the growthrates achieved by bacterioplankton in different parts of thereservoir. (i) The pool of DRP can consist of very differentratios of bioavailable and nonbioavailable fractions (28, 36).(ii) Moreover, rapid turnover rates of the bioavailable P duringthe clear-water phase can override the effect of the analyticallyrelatively low detectable DRP concentrations in the dam area(28), where nutrient recycling could be strongly stimulated bygrazing activity of the abundant zooplankton (26). (iii) Al-though the FISH data with group-specific probes did not showlarge differences in BCC, the DGGE banding pattern at 96 hindicated remarkable site-specific BCC shifts. Thus, differentbacterial phylotypes (cf. Fig. 3 and 4) were the major bacterialplayers at different parts of the reservoir, phylotypes whichwere well adapted to local ambient nutrient conditions allow-ing them to efficiently compensate for the variable P availabil-ity.
Bacteria in the bottle incubations showed dramatic differ-ences in both overall dynamics and community composition
compared to dialysis bag incubations. The bottle treatmentsalways showed the lowest bacterial numbers, biomass, andgrowth rates of the different phylogenetic groups of bacteria(cf. Fig. 1 through 4). It indicates a specific “bottle effect” (10)and an overall problematic nature of bottle incubations forstudying in situ microbial processes.
Additional differences were found between bottle and dial-ysis bag incubated populations. For instance, the treatment-specific BCC shifts (Fig. 3 and 4) supported different HNFpopulations. In bottle �5-�m incubations, very small and fast-growing Spumella-like chrysomonads dominated (see the rapidHNF increase in Fig. 2). Thus, the altered nutrient availabilityin bottle compared to dialysis incubations was associated withquite complex changes in the microbial food web: it affectednot only BCC directly but also the grazer community compo-sition and in turn also the HNF species-specific grazing, whichprobably differently impacted the resulting BCC as has beenshown before (31, 33).
The pooling of data across all dialysis bag treatments showeddistinct effects of the bottom-up and top-down manipulationson the relative contributions of different phylogenetic groupsof bacteria (Table 2). Manipulation of the grazing pressureconsistently altered the proportion of the Actinobacteria group
FIG. 5. Time course changes in the proportions of bacteria withHNA content related to total DAPI-stained bacteria in different sizefractionation treatments of reservoir water collected at the dam areaand incubated in bottles or in dialysis bags at the dam, middle, andriver parts of the reservoir (for more details, see the Fig. 1 legend).Values are means for two replicate treatments.
TABLE 2. Two-way ANOVA used to test for differences in BCC between the dam, middle, and river incubations throughout all experimentalvariants and different time intervalsa
ProbeBottom-up manipulationb (F value) Top-down manipulationc (F value)
24 h 48 h 72 h 96 h 24 h 48 h 72 h 96 h
BET42a 3.16 0.35 1.49 0.18 0.22 0.72 0.65 1.13R-BT065 15.3** 80.5*** 6.12* 0.48 29.4*** 583*** 18.8** 6.24*GAM42a 0.27 0.36 1.79 3.86 6.15* 11.3* 2.32 2.12CF319a 0.66 27.1** 8.45* 0.96 3.07 0.09 0.02 0.01HGC69a 1.09 2.94 0.61 2.28 10.5* 43.3*** 6.4* 6.2*
a We tested the differences in the relative contributions (as a percentage of total DAPI-stained bacteria) of five different phylogenetic groups of bacteria targetedby the probes BET42a, R-BT065, GAM42a, CF319a, and HGC69a induced by transplanting the samples from the dam area upstream to the middle and river areasand by manipulating top-down control via size fractionation of water samples. The significant differences are in boldface type. *, P � 0.05; **, P � 0.01; ***, P � 0.001.
b Dam samples transplanted to the middle and river parts.c Effects of distinct grazing pressures when bacterivore-free treatments are compared to bacterivore-enhanced treatments.
TABLE 3. Correlation relationships between the proportion ofHNA cells within the bacterial assemblages compared to HNF andbacterioplankton parameters and the proportions of FISH-detected
subgroups of bacteria across all experimental treatmentsa
Parameter Correlationcoefficient (r) Significance (P)
HNF abundance 0.045 0.8302Total HNF grazing 0.063 0.7641Bacterial abundance 0.846 �0.0001Bacterial production 0.715 �0.0001Bacterial biomass 0.845 �0.0001Bacterial mean cell volume 0.813 �0.0001% EUB338 0.643 �0.0001% BET42a 0.814 �0.0001% R-BT065 0.792 �0.0001% GAM42a 0.585 �0.0001% CF319a 0.461 0.0014% HGC69a �0.016 0.2912
a For calculations, the proportions of HNA bacteria and FISH-detected bac-terial subgroups were expressed as a percentage of total DAPI stained cells. Forall analyses, n � 45, except for those with HNF abundance and grazing (n � 25).Significant relationships (P � 0.01) are shown in boldface type.
VOL. 71, 2005 HIGHLY ACTIVE SMALL SUBCLUSTER OF �-PROTEOBACTERIA 2387

targeted by the HGC69a probe (Fig. 4). The cells from thislineage were generally very small (0.054 to 0.085 �m3), andmoreover, in all treatments, they seem to be discriminatedagainst by HNF. The latter finding is consistent with several
reports (18, 31) documenting grazing resistance of some acti-nobacterial phylotypes. This phenomenon could be related totheir small cell size but also partially to some size-independenttraits, as has been shown for the MWH-Mo1 strain describedpreviously by Hahn et al. (18). The actinobacterial phylotypesin the reservoir were small cells, and their relative abundancewas not correlated with %HNA bacteria; they showed growthrates comparable to the average values of the whole bacterio-plankton. Thus, while apparently avoided by HNF, the rela-tively short doubling time (12 to 23 h [Table 1]) gave them acompetitive advantage so that they were overrepresented inthe presence of grazing. For example, in the heavily grazedbottle �5-�m treatment, they reached as much as 30% of totalbacteria (Fig. 4). However, the proportions of Actinobacteriashowed enhanced variability when �0.8- and �5-�m treat-ments are compared at time zero (Fig. 4), which could bepartially related to the initial sample manipulation via sizefractionation.
This actinobacterial group is considered to be ubiquitous inlakes of various trophic state, size, and geographical location(15, 18, 48, 51). The known or cultivated phylotypes affiliatedto the freshwater Actinobacteria group are small- to medium-size cells (see, e.g., reference 18), and they seem to be anautochthonous and phylogenetically highly diverse componentof freshwater microbial assemblages (50). However, the role ofthe phylotypes in their habitats remains largely unknown, andeven identical actinobacterial sequence types collected fromvery different locations displayed different growth characteris-tics when growing in the same medium (18).
To our knowledge, there is no similar study dealing specif-ically with the ecological role of the lineage detected by theR-BT065 probe (a subcluster of the cosmopolitan clusterRhodoferax sp. BAL47) in a single environment. This groupplayed a main role in the reservoir bacterioplankton (Fig. 4)since its contribution to total bacteria was tightly correlated tomost bacterioplankton bulk parameters and responded veryclearly to both top-down and bottom-up manipulations (cf.Tables 1 through 4). Apparently, this lineage involves one ormore species representing the true example of an opportunisticbacterial strategy in the system. Similar ecological capabilitieshave been shown for a freshwater lineage beta II in the lakeGrosse Fukchskuhle (4) and several groups of marine -Pro-teobacteria like Alteromonas, Pseudoalteromonas, and Vibrio,which consistently react to enrichment and grazing in labora-tory experiments (2, 9).
In this study, as well as in a previous one (46), the increasesin �-proteobacteria (BET42a probe) in the grazer-free treat-ments were almost completely attributable to fast-growing cellsdetected by the R-BT065 probe, displaying a conspicuouslyshort doubling time of only 8 to 9 h (Table 1) in the dialysis bagtreatments. The cells of this lineage were generally larger (0.09to 0.22 �m3) rod-shaped cells of rather uniform morphology,certainly vulnerable to predation, without any obvious trait ofmorphological grazing resistance behavior (see, e.g., reference17 for a review). Thus, we suggest that the large growth po-tential of the members of this lineage is the key factor allowingthem to efficiently compensate for grazing losses in the grazer-enhanced treatments and to rapidly overgrow other lineages ingrazer-free treatments (Fig. 4).
A growing amount of evidence suggests that the HNA bac-
FIG. 6. Relationships between the proportions of bacteria withHNA content related to total DAPI-stained bacteria and proportionsof probe-defined bacterial subpopulations with data pooled from alltreatments (n � 45). (A) EUB338-detectable cells; (B) BET42a-de-tectable cells; (C) R-BT065-detectable cells. Full lines depict linearregressions for all data, and the r2 value shows the coefficient ofdetermination of the regressions. In C, the open symbols are thosefrom bacterivore-free treatments only (n � 20) fitted with the dashedregression line; closed symbols are for treatments with the presence ofgrazers. All regressions are significant (P � 0.0001).
TABLE 4. Correlation relationships between the proportions of�-Proteobacteria (the BET42a probe-detected cells) and of thesubcluster of �-Proteobacteria (R-BT065 probe-detected cells)
as components of the bacterial community versus basicbacterioplankton parameters throughout all treatmentsa
Parameter
% BET42a % R-BT065
Correlationcoefficient
(r)
Significance(P)
Correlationcoefficient
(r)
Significance(P)
Bacterial abundance 0.615 �0.0001 0.810 �0.0001Bacterial biomass 0.587 �0.0001 0.753 �0.0001Bacterial mean cell
volume0.661 �0.0001 0.575 �0.0001
Bacterial production 0.529 0.0004 0.546 0.0002
a For calculations, the proportions of FISH-detected bacterial subgroups wereexpressed as a percentage of total DAPI-stained bacteria. For all analyses, n �45.
2388 SIMEK ET AL. APPL. ENVIRON. MICROBIOL.

teria are the most active members of the bacterial community,responsible for the major part of bacterial production (13, 24,40). However, in natural systems, bacterial production can alsobe affected by the selective removal of possibly larger dividingand metabolically more active cells (41). Thus, growth of graz-ers can be initially stimulated by enhanced proportions oflarger HNA bacteria (47), but generally, in longer incubations,it yields lower proportions of metabolically active or HNAbacteria (see reference 8 and also cf. Fig. 5). Correspondingly,in our study, the proportions of R-BT065-positive cells tightlycorrelated with bacterial production, biomass, and %HNAbacteria, with the latter correlation even higher when com-puted for bacterivore-free treatments only (Fig. 6). However,our data did not show any significant correlation between HNFparameters and %HNA bacteria, while apparently, the inten-sive HNF grazing markedly decreased the %HNA bacteria bythe end of incubation (Fig. 5). A longer experiment wouldprobably have reflected this possibility.
Interestingly, at the beginning of the experiment, when HNFwere efficiently top-down controlled during the clear-waterphase, proportions of HNA bacteria were very low (�30% oftotal bacteria) in all treatments. This might suggest that clado-ceran filtration (which is responsible for the clear-water phase)removed primarily larger bacteria (20), presumably the pool ofHNA bacteria, as HNA bacteria are the largest component ofthe bacterial community (reference 25 and also see Table 3 inthis study). After removing zooplankton from our size-frac-tionated samples, the initially very small bacterial MCV (�0.08�m3) grew markedly together with increasing proportions ofHNA and R-BT065-positive bacteria in all treatments. Theonly exception was bottle �0.8-�m treatment, where MCVincreased only slightly while the proportions of R-BT065-pos-itive cells stayed largely invariable (Fig. 4).
More evidence of the high growth and metabolic rates, aswell as of ecological flexibility, of R-BT065-positive cells wasfound in a similar manipulation experiment conducted in thereservoir in May 2003. FISH combined with microautora-diography showed that the cells detected with the R-BT065probe remained a very important group in the assemblage,and they were the only phylogenetic group that showed amarked increase in the percentage of metabolically activecells ([3H]leucine uptake from �60 to 95%), even underheavy grazing pressure in �5-�m treatments (K. Hornak,unpublished data).
Overall, when data for all our treatments are pooled,changes in numbers of cells detected with the R-BT065 probeexplained �89% (r2 � 0.891, P � 0.0001) of the variability inthe numbers of HNA bacteria. This finding suggests an impor-tant role of this subgroup in the reservoir bacterioplanktondynamics and underlines the need to further identify the par-ticular members of this small but important lineage whosemembers were found in various freshwater environments (15,51). Thus, though we have strong evidence concerning the roleof this prominent lineage in overall bacterioplankton dynamics(cf. Fig. 4 and Tables 2 to 4), the relationship with HNA cellsis based only on the correlation approach. As yet, there havenot been any attempts to identify which groups or species aremajor component in the fraction of the HNA in freshwaters, incontrast to marine studies (see, e.g., references 25 and 50).
Interestingly, an enrichment experiment with North Sea bac-
terioplankton (9) and dilution experiments with water from theEnglish Channel (10) clearly documented that manipulatingnutrient supply yielded a dramatic community shift with HNAbacteria dominated mainly by opportunistic groups of -pro-teobacteria that usually account for only a minor part of theoriginal nonmanipulated marine samples. In contrast, our typ-ical freshwater group of opportunistic phylotypes belonging tothe R-BT065 cluster has always been a very abundant group inthe nonmanipulated reservoir bacterioplankton (references 26,43, and 46 and also see Fig. 4 in this paper).
In this study, we detected a small subcluster of �-proteobac-teria of importance in bacterioplankton dynamics in the reser-voir. For example, they appear to be responsible for carryingout most of the bacterial production processes. The membersof this lineage are extremely fast-growing cells, vulnerable toprotist predation without detectable MCV changes in the pres-ence of predators but having the ability to sensitively respondto variable experimental manipulations. Interestingly, whilethe typical marine opportunistic strategists found among-proteobacteria (9) are proportionally less important in nat-ural assemblages, the target for selective HNF predation (2),the members of the R-BT065 cluster, constituted both season-ally as well as spatially a remarkable part of the reservoirbacterioplankton assemblages, accounting for 50 to 100% ofthe total �-proteobacteria (26, 43, 46). The isolation of theparticular members of the R-BT065 cluster and characteriza-tion of its phylogenetic diversity are beyond the scope of thisstudy.
ACKNOWLEDGMENTS
This study was largely supported by the Grant Agency of the CzechRepublic under research grants 206/05/0007 and 206/02/0003 awardedto K.S. and partly by the project MSM 123100004 (structure andfunctioning of the system water-soil-plant), by projects DGICYTREN2001-2120/MAR and EVK3-CT-2002-00078 awarded to J.M.G.,and by a cooperation project between ASCR and CSIC. M.S. wassupported by the Austrian Science Foundation (project no. P15655).
We thank J. Dolan for valuable comments and English correction, J.Vrba for assistance with experimental sampling in the reservoir, J. He-jzlar for chemical analysis, and S. Smrckova and R. Mala for excellenttechnical assistance.
REFERENCES
1. Alfreider, A., J. Pernthaler, R. Amann, B. Sattler, F.-O. Glockner, A. Ville,and R. Psenner. 1996. Community analysis of the bacterial assemblages inthe winter cover and pelagic layers of a high mountain lake using in situhybridization. Appl. Environ. Microbiol. 62:2138–2144.
2. Beardsley, C., J. Pernthaler, W. Wosniok, and R. Amann. 2003. Are readilyculturable bacteria in coastal North Sea waters suppressed by selective graz-ing mortality? Appl. Environ. Microbiol. 69:2624–2630.
3. Bouvier, T. C., and P. A. del Giorgio. 2002. Compositional changes in free-living bacterial communities along a salinity gradient in two temperate es-tuaries. Limnol. Oceanogr. 47:453–470.
4. Burkert, U., F. Warnecke, D. Babenzien, E. Zwirnmann, and J. Pernthaler.2003. Members of a readily enriched �-proteobacterial clade are common insurface waters of a humic lake. Appl. Environ. Microbiol. 69:6550–6559.
5. Cottrell, M. T., and D. L. Kirchman. 2004. Single-cell analysis of bacterialgrowth, cell size, and community structure in the Delaware estuary. Aquat.Microb. Ecol. 34:39–149.
6. Crump, B. C., E. V. Armbrust, and J. A. Baross. 1999. Phylogenetic analysisof particle-attached and free-living bacterial communities in the ColumbiaRiver, its estuary, and the adjacent coastal ocean. Appl. Environ. Microbiol.65:3192–3204.
7. del Giorgio, P. A., and T. C. Bouvier. 2002. Linking the physiologic andphylogenetic successions in free-living bacterial communities along an estu-arine salinity gradient. Limnol. Oceanogr. 47:471–486.
8. del Giorgio, P. A., J. M. Gasol, D. Vaque, P. Mura, S. Agostı, and C. M.Duarte. 1996. Bacterioplankton community structure: protists control net
VOL. 71, 2005 HIGHLY ACTIVE SMALL SUBCLUSTER OF �-PROTEOBACTERIA 2389

production and the proportion of active bacteria in a coastal marine com-munity. Limnol. Oceanogr. 41:1169–1179.
9. Eilers, H., J. Pernthaler, and R. Amann. 2000. Succession of pelagic marinebacteria during enrichment: a close look at cultivation-induced shifts. Appl.Environ. Microbiol. 66:4634–4640.
10. Fuchs, B. M., M. V. Zubkov, K. Sahm, P. H. Burkill, and R. Amann. 2000.Changes in community composition during dilution cultures of marine bac-terioplankton as assessed by flow cytometric and molecular biological tech-niques. Environ. Microbiol. 2:191–201.
11. Gasol, J. M., M. Comerma, J.-C. Garcia, J. Armengol, E. O. Casamayor, P.Kojecka, and K. Simek. 2002. A transplant experiment to identify the factorscontrolling bacterial abundance, activity, production and community com-position in a eutrophic canyon-shaped reservoir. Limnol. Oceanogr. 47:62–77.
12. Gasol, J. M., and P. A. del Giorgio. 2000. Using flow cytometry for countingnatural plaktonic bacteria and understanding the structure of planktonicbacterial communities. Sci. Mar. 64:197–224.
13. Gasol, J. M., U. L. Zweifel, F. Peters, J. A. Fuhrman, and A. Hagstrom. 1999.Significance of size and nucleic acid content heterogeneity as measured byflow cytometry in natural planktonic bacteria. Appl. Environ. Microbiol.65:475–4483.
14. Glockner, F. O., B. M. Fuchs, and R. Amann. 1999. Bacterioplankton com-positions of lakes and oceans: a first comparison based on fluorescence insitu hybridization Appl. Environ. Microbiol. 65:3721–3726.
15. Glockner, F. O., E. Zaichikov, N. Belkova, L. Denissova, J. Pernthaler, A.Pernthaler, and R. Amann. 2000. Comparative 16S rRNA analysis of lakebacterioplankton reveals globally distributed phylogenetic clusters includingan abundant group of Actinobacteria. Appl. Environ. Microbiol. 66:5053–5065.
16. Hahn, M. W. 2003. Isolation of strains belonging to the cosmopolitanPolynucleobacter necessarius cluster from freshwater habitats located in threeclimatic zones. Appl. Environ. Microbiol. 69:5248–5254.
17. Hahn, M. W., and M. G. Hofle. 2001. Grazing of protozoa and its effect onpopulations of aquatic bacteria. FEMS Microbiol. Ecol. 35:113–121.
18. Hahn, M. W., H. Lunsdorf, Q. Wu, M. Schauer, M. G. Hofle, J. Boenigk, andP. Stadler. 2003. Isolation of novel ultramicrobacteria classified as Acti-nobacteria from five freshwater habitats in Europe and Asia. Appl. Environ.Microbiol. 69:1442–1451.
19. Jezbera, J., J. Nedoma, and K. Simek. 2003. Longitudinal changes in pro-tistan bacterivory and bacterial production in two differently loaded canyon-shaped reservoirs. Hydrobiologia 504:115–130.
20. Jurgens, K. 1994. Impact of Daphnia on planktonic microbial food webs—areview. Mar. Microb. Food Webs 8:295–324.
21. Jurgens, K., J. Pernthaler, S. Schalla, and R. Amann. 1999. Morphologicaland compositional changes in a planktonic bacterial community in responseto enhanced protozoan grazing. Appl. Environ. Microbiol. 65:1241–1250.
22. Kirchman, D. L., L. Yu, B. M. Fuchs, and R. Amann. 2001. Structure ofbacterial communities in aquatic systems as revealed by filter PCR. Aquat.Microb. Ecol. 26:13–22.
23. Kopacek, J., and J. Hejzlar. 1993. Semi-micro determination of total phos-phorus in fresh waters with perchloric acid digestion. Int. J. Environ. Anal.Chem. 53:173–183.
24. Lebaron, P., P. Servais, H. Agogue, C. Courties, and F. Joux. 2001. Does thehigh nucleic acid content of individual bacterial cells allow us to discriminatebetween active cells and inactive cells in aquatic systems? Appl. Environ.Microbiol. 67:1775–1782.
25. Lebaron, P., P. Servais, A.-C. Baudoux, M. Bourrain, C. Courties, and N.Parthuisot. 2002. Variations of bacterial-specific activity with cell size andnucleic acid content assessed by flow cytometry. Appl. Environ. Microbiol.68:131–140.
26. Masın, M., K. Simek, J. Jezbera, J. Nedoma, V. Straskrabova, and J. Hej-zlar. 2003. Changes in bacterial community composition and microbial ac-tivities along the longitudinal axis of two differently loaded canyon-shapedreservoirs. Hydrobiologia 504:99–113.
27. Muyzer, G., T. Brinkhoff, U. Nubel, C. Santegoeds, H. Schafer, and C.Wawer. 1997. Denaturing gradient gel electrophoresis (DGGE) in microbialecology, p. 1–27. In A. D. L. Akkermans, J. D. van Elsas, and F. J. de Bruijn(ed.), Molecular microbial ecology manual, 2nd ed., vol. 3. Kluwer AcademicPublishers, Dordrecht, The Netherlands.
28. Nedoma, J., P. Porcalova, J. Komarkova, and V. Vyhnalek. 1993. Phosphorusdeficiency diagnostics in the eutrophic Rımov reservoir. Water Sci. Technol.28:75–84.
29. Norland, S. 1993. The relationship between biomass and volume of bacteria,p. 303–308. In P. Kemp, B. F. Sherr, E. B. Sherr, and J. Cole (ed.), Handbookof methods in aquatic microbial ecology. Lewis, Boca Raton, Fla.
30. Pernthaler, A., J. Pernthaler, and R. Amann. 2002. Fluorescence in situhybridization and catalyzed reporter deposition for the identification ofmarine bacteria. Appl. Environ. Microbiol. 68:3094–3101.
31. Pernthaler, J., T. Posch, K. Simek, J. Vrba, U. Nubel, F.-O. Glockner, R.Psenner, and R. Amann. 2001. Predator-specific enrichment of actinobacte-
ria from a cosmopolitan freshwater clade in a mixed continuous culture.Appl. Environ. Microbiol. 67:2145–2155.
32. Posch, T., J. Pernthaler, A. Alfreider, and R. Psenner. 1997. Cell-specificrespiratory activity of aquatic bacteria studied with the tetrazolium reductionmethod, Cyto-Clear slides, and image analysis. Appl. Environ. Microbiol.63:867–873.
33. Posch, T., K. Simek, J. Vrba, S. Pernthaler, J. Nedoma, B. Sattler, B.Sonntag, and R. Psenner. 1999. Predator-induced changes of bacterial size-structure and productivity studied on an experimental microbial community.Aquat. Microb. Ecol. 18:235–246.
34. Riemann, B., and M. Søndergaard. 1986. Carbon dynamics in eutrophic,temperate lakes. Elsevier Science Publishers, Amsterdam, The Netherlands.
35. Riemann, L., G. F. Steward, and F. Azam. 2000. Dynamics of bacterialcommunity composition and activity during a mesocosm diatom bloom.Appl. Environ. Microbiol. 66:578–587.
36. Rigler, F. H. 1966. Radiobiological analysis of inorganic phosphorus in lakewater. Vehr. Int. Verein. Limnol. 16:465–470.
37. Schafer, H., L. Bernard, C. Courties, P. Lebaron, P. Servais, R. Pukall, E.Stackebrandt, M. Troussellier, T. Guindulain, J. Vives-Rego, and G.Muyzer. 2001. Microbial community dynamics in Mediterranean nutrient-enriched seawater mesocosms: changes in the genetic diversity of bacterialpopulations. FEMS Microbiol. Ecol. 34:243–253.
38. Schauer, M., V. Balague, C. Pedros-Alio, and R. Massana. 2003. Seasonalchanges in the taxonomic composition of bacterioplankton in a coastal oli-gotrophic system. Aquat. Microb. Ecol. 31:163–174.
39. Sekar, R., A. Pernthaler, J. Pernthaler, F. Warnecke, T. Posch, and R.Amann. 2003. An improved protocol for quantification of freshwater Acti-nobacteria by fluorescence in situ hybridization. Appl. Environ. Microbiol.69:2928–2935.
40. Servais, P., C. Courties, P. Lebaron, and M. Trousellier. 1999. Couplingbacterial activity measurements with cell sorting by flow cytometry. Microb.Ecol. 38:180–189.
41. Sherr, B. F., E. B. Sherr, and J. McDaniel. 1992. Effect of protistan grazingon the frequency of dividing cells in bacterioplankton assemblage. Appl.Environ. Microbiol. 58:2381–2385.
42. Sherr, E. B., and B. F. Sherr. 1993. Protistan grazing rates via uptake offluorescently labelled prey, p. 695–701. In P. Kemp, B. F. Sherr, E. B. Sherr,and J. Cole (ed.), Handbook of methods in aquatic microbial ecology. Lewis,Boca Raton, Fla.
43. Simek, K., K. Hornak, M. Masın, U. Christaki, J. Nedoma, M. G. Wein-bauer, and J. R. Dolan. 2003. Comparing the effects of resource enrichmentand grazing on a bacterioplankton community of a meso-eutrophic reservoir.Aquat. Microb. Ecol. 31:123–135.
44. Simek, K., P. Kojecka, J. Nedoma, P. Hartman, J. Vrba, and D. R. Dolan.1999. Shifts in bacterial community composition associated with differentmicrozooplankton size fractions in a eutrophic reservoir. Limnol. Oceanogr.44:1634–1644.
45. Simek, K., J. Nedoma, J. Pernthaler, T. Posch, and J. R. Dolan. 2002.Altering the balance between bacterial production and protistan bacterivorytriggers shifts in freshwater bacterial community composition. Antonie Leeu-wenhoek 81:453–463.
46. Simek, K., J. Pernthaler, M. G. Weinbauer, K. Hornak, J. R. Dolan, J.Nedoma, M. Masin, and R. Amann. 2001. Changes in bacterial communitycomposition, dynamics, and viral mortality rates associated with enhancedflagellate grazing in a mesoeutrophic reservoir. Appl. Environ. Microbiol.67:2723–2733.
47. Vaque, D., E. O. Casamayor, and J. M. Gasol. 2001. Dynamics of wholecommunity bacterial production and grazing losses in seawater incubationsas related to the changes in the proportions of bacteria with different DNAcontent. Aquat. Microb. Ecol. 25:163–177.
48. Warnecke, F., R. Amann, and J. Pernthaler. 2004. Actinobacterial 16S rRNAgenes from freshwater habitats cluster in four distinct lineages. Environ.Microbiol. 6:242–253.
49. Weinbauer, M. G., K. Simek, U. Christaki, and J. Nedoma. 2003. Comparingthe effects of resource enrichment and grazing on viral production in ameso-eutrophic reservoir. Aquat. Microb. Ecol. 31:137–144.
50. Zubkov, M. V., B. M. Fuchs, P. H. Burkill, and R. Amann. 2001. Comparisonof cellular and biomass specific activities of dominant bacterioplanktongroups in stratified waters of the Celtic Sea. Appl. Environ. Microbiol.67:5210–5218.
51. Zwart, G., B. C. Crump, M. P. Kamst-van Agterveld, F. Hagen, and S. K.Han. 2002. Typical freshwater bacteria: an analysis of available 16S rRNAgene sequences from plankton of lakes and rivers. Aquat. Microb. Ecol.28:141–155.
52. Zwart, G., E. J. van Hannen, M. P. Kamst-van Agterveld, K. Van der Gucht,E. S. Lindstrom, J. Van Wichelen, T. Lauridsen, B. C. Crump, S. K. Han,and S. Declerck. 2003. Rapid screening for freshwater bacterial groups byusing reverse line blot hybridization. Appl. Environ. Microbiol. 69:5875–5883.
2390 SIMEK ET AL. APPL. ENVIRON. MICROBIOL.