
The Impact of Clinical Pathway for Congestive Heart Failure ...

The American Journal of Pathology Vol 179 No 1 July 2011
Copyright copy 2011 American Society for Investigative Pathology
Published by Elsevier Inc All rights reserved
DOI 101016jajpath201103036
Stem Cells Tissue Engineering and Hematopoietic Elements
Effects of Age and Heart Failure on Human Cardiac
Stem Cell Function
Daniela Cesselli Antonio P BeltramiFederica DrsquoAurizio Patrizia MarconNatascha Bergamin Barbara ToffolettoMaura Pandolfi Elisa Puppato Laura Marinodagger
Sergio SignoreDagger Ugolino Livisect Roberto Verardopara
Silvano Piazzapara Luigi Marchionni
Claudia Fiorini Claudio Schneiderpara
Toru HosodaDagger Marcello RotaDagger Jan KajsturaDagger
Piero AnversaDagger Carlo A Beltrami andAnnarosa LeriDagger
From the Center for Regenerative Medicine and the Department
of Cardiac Surgerysect Udine Medical School Udine Italy the
Department of Experimental Medicinedagger Naples Second Medical
School Naples Italy the Departments of Anesthesia and
Medicine and Division of Cardiovascular MedicineDagger Brigham
and Womenrsquos Hospital Harvard Medical School Boston
Massachusetts the National Biotechnology Laboratory (LNCIB)para
Padriciano Trieste Italy the Sidney Kimmel Comprehensive
Cancer Center Johns Hopkins University School of Medicine
Baltimore Maryland and the Montefiore Medical Center
Albert Einstein College of Medicine New York New York
Currently it is unknown whether defects in stemcell growth and differentiation contribute to myo-cardial aging and chronic heart failure (CHF) andwhether a compartment of functional human car-diac stem cells (hCSCs) persists in the decompen-sated heart To determine whether aging and CHFare critical determinants of the loss in growth re-serve of the heart the properties of hCSCs wereevaluated in 18 control and 23 explanted heartsAge and CHF showed a progressive decrease infunctionally competent hCSCs Chronological agewas a major predictor of five biomarkers of hCSCsenescence telomeric shortening attenuated telome-rase activity telomere dysfunction-induced foci andp21Cip1 and p16INK4a expression CHF had similarconsequences for hCSCs suggesting that defects inthe balance between cardiomyocyte mass and thepool of nonsenescent hCSCs may condition the evo-lution of the decompensated myopathy A correlationwas found previously between telomere length in cir-
culating bone marrow cells and cardiovascular dis-
eases but that analysis was restricted to average telo-mere length in a cell population neglecting the factthat telomere attrition does not occur uniformly in allcells The present study provides the first demonstra-tion that dysfunctional telomeres in hCSCs are bio-markers of aging and heart failure The biomarkers ofcellular senescence identified here can be used todefine the birth date of hCSCs and to sort young cellswith potential therapeutic efficacy (Am J Pathol 2011179349ndash366 DOI 101016jajpath201103036)
The recognition that the human heart possesses a com-partment of c-Kit-positive cardiac stem cells (CSCs) thatcan regenerate myocytes and coronary vessels offers theunique opportunity to reconstitute the damaged myocar-dium restoring in part the physiological and anatomicalcharacteristics of the normal heart Human CSCs (hCSCs)can be isolated from small tissue samples and after theirexpansion in vitro can be delivered back to the samepatient in an attempt to repair the failing heart1ndash5 Thisdiscovery has laid the groundwork for the use of hCSCsin the treatment of the human disease Preclinical studieshave been completed and two phase I clinical trials arein progress [NCT00474461 (last accessed December 222010) and NCT00893360 (last accessed January 18
Supported by Regione Friuli Venezia Giulia (art 23 LR 262005 Progettodi ricerca SeNDndashSearch for New Drugs to CAB) MIUR (PRIN 2006 pr2006060854 to CAB) and by NIH grants P01-AG-023071 (PA) R01-AG-026107 (AL) R01-HL-65577 (AL) P01-HL-092868 (PA) and R01-AG-37490 (AL)
Accepted for publication March 3 2011
DC and APB contributed equally to the present work
Supplemental material for this article can be found at httpajpamjpatholorg or at doi 101016jajpath201103036
A guest editor acted as editor-in-chief for this manuscript No person atThomas Jefferson University or Albert Einstein College of Medicine wasinvolved in the peer review process or final disposition for this article
Address reprint requests to Daniela Cesselli MD PhD Center forRegenerative Medicine Udine Medical School Padiglione 13 PiazzaleS Maria della Misericordia 15 33100 Udine Italy or Annarosa LeriMD Departments of Anesthesia and Medicine and CardiovascularDivision Brigham and Womenrsquos Hospital Harvard Medical School 75Francis Street Boston MA 02115 E-mail danielacesselliuniudit or
aleripartnersorg
349
350 Cesselli et alAJP July 2011 Vol 179 No 1
2011) at httpwwwclinical trialsgov] Nonetheless ag-ing cardiac hypertrophy ischemic myocardial injuryand metabolic disorders together with genetic and envi-ronmental factors may dramatically affect the growth anddifferentiation behavior of resident hCSCs6ndash9 This possi-bility raises two critical questions First is heart failure astem cell disease characterized by severe depletion of thehCSC pool Second does a compartment of functionallycompetent hCSCs persist in the decompensated heart anddo these cells have potential therapeutic implications
As is the case for other stem cells10 the life cycle ofhCSCs is regulated by telomerase activity and telomerelength2 Telomerase is a reverse transcriptase that ex-tends the 3= chromosomal ends by using its own RNA asa template11 Telomerase activity delays but cannot pre-vent telomere erosion which is dictated by down-regu-lation of telomerase oxidative stress and loss of telom-ere-related proteins12 Shortening of telomeres beyond acritical length triggers cellular senescence which corre-sponds to irreversible growth arrest in the G1 phase withloss of specialized functions including stem cell prolifer-ation migration and differentiation Despite suggestiveevidence in humans and in genetically manipulatedmice13ndash15 that telomere shortening is a major variable ofcellular senescence and organ aging it remains to bedemonstrated whether hCSCs with critically shortened ordysfunctional telomeres undergo replicative senescenceand apoptosis and whether loss of telomere integrity isone of the variables involved in the decline of stem cellfunction in the failing human heart
These issues were addressed in the present study andthe consequences of aging and ventricular decompensa-tion on the clonogenicity multipotency and migratory prop-erties of hCSCs were characterized based on analysis ofnormal donor hearts and explanted hearts in end-stageheart failure Moreover the telomere-telomerase axis andthe presence of telomere dysfunction-induced foci (TIFs) incombination with markers of DNA damage response andreplicative senescence were determined to obtain a com-mon denominator for the processes that regulate the growthand death of hCSCs
Materials and Methods
Human Samples
Discarded atrial specimens weighing 3 to 6 g werecollected over a period of 5 years from donor hearts atthe time of transplantation and from explanted hearts ofpatients undergoing cardiac transplantation at the Car-diac Surgery Unit of the University Hospital of Udine inItaly The clinical data are presented in SupplementalTables S1 and S2 (available at httpajpamjpatholorg)Informed consent was obtained in accordance with theDeclaration of Helsinki and with approval by the Indepen-dent Ethics Committee of the University of Udine
Histological analysis of samples from donor hearts didnot show pathological changes Samples were used forthe isolation of c-Kit-positive hCSCs2ndash4 Fragments were
also fixed in formalin and embedded in paraffin2ndash4 for the
identification of stem cell niches and for the quantitativeanalysis of lineage-negative hCSCs81617 Lineage-neg-ative hCSCs were defined as cells positive for c-Kit andnegative for CD45 to exclude the bone marrow originand the mast cell phenotype and negative for GATA4and -sarcomeric actin (-SA) to exclude commitment tocardiac lineages The localization of OCT34 and NANOGwas determined in CD45-negative c-Kit-positive hCSCsMagnitude of sampling is presented in Supplemental Ta-ble S3 (available at httpajpamjpatholorg) antibodiesare listed in Table 1
Spectral Analysis
Spectral analysis was performed with a TCS-SP2 confo-cal microscope (Leica Wetzlar Germany) using thelambda acquisition mode myocardial sections were la-beled by c-Kit OCT34 and NANOG conjugated withAlexa Fluor 555 Cy5 and Alexa Fluor 555 respectivelyMyocytes were stained with -SA (Cy5) and nuclei withDAPI The emission signal for Alexa Fluor 555 was ex-cited at 543 nm with an argon laser and the fluores-cence intensity was recorded generating a lambdastack ranging from 558 to 798 nm at 49-nm intervalsThe emission signal for Cy5 was excited at 633 nm witha heliumneon laser and the fluorescence intensitywas recorded generating a lambda stack ranging from653 to 798 nm at 6-nm intervals Lens and correspond-ing numerical aperture were 63 and 14 respectivelySampling consisted of 30 c-Kit-positive OCT34-posi-tive and NANOG-positive cells Additionally 10 cellsnegative for these markers and present in the samesamples were used as control to discriminate back-ground autofluorescence from specific labeling Eachdetermination was restricted to the region of interestwithin each hCSC For each region of interest a graphplotting mean pixel intensity and the emission wave-length of the lambda stack was generated This proto-col has repeatedly been used in our laboratory17ndash20
Human Cardiac Stem Cells
In all cases atrial samples were used for the collectionof hCSCs this approach was followed to obtain a directcomparison between control and failing hearts Twoprotocols were used for the isolation of hCSCs enzy-matic dissociation of the samples with collagenase andprimary explant technique These protocols were de-veloped in the original documentation of hCSCs2 andwere retained here for continuity and comparison Aspreviously2 these two methodologies yielded compa-rable results up to 20 to 25 population doublings effi-ciency and viability of hCSCs were superimposableCollagenase treatment was not found to affect thesevariables The phenotype of the small cell populationwas defined by flow-cytometry and immunolabeling24
Single-cell suspensions were labeled with directly con-jugated antibodies (Table 1) specific for hematopoieticand endothelial cell (EC) lineages Conjugated isotypeantibodies were used as negative control To exclude
the presence of erythrocytes the samples of freshly
thiocyan
Human Cardiac Stem Cell Senescence 351AJP July 2011 Vol 179 No 1
isolated cells were exposed to BD fluorescence-acti-vated cell sorting (FACS) lysing solution the flow cy-tometric analysis was performed with a BD FACScansystem (BD Biosciences San Jose CA) or a CyAnsystem (Beckman Coulter Fullerton CA) For immuno-labeling hCSCs were fixed in 4 paraformaldehydefor 15 minutes at room temperature and the expressionof c-Kit OCT34 NANOG and SOX2 was evalu-ated2417 For clonal analysis a FACSAria cell sorter(BD-Biosciences) was used to seed single c-Kit-posi-tive hCSCs into 96-well Terasaki plates24
RT-PCR
Total RNA was extracted from nonconfluent cultures ofhCSCs at passage 3 using TRIzol reagent (InvitrogenCarlsbad CA) After treatment with DNase I (AmbionApplied Biosystems Austin TX) first-strand cDNAsynthesis was performed with 1 g total RNA usingrandom hexanucleotides and M-MLV reverse trans-
Table 1 Antibodies for FACS Immunolabeling and Western Blo
Antigen Antibody
FACSc-kit APC-conjugated mouse mAbc-kit PE-conjugated mouse mAbCD34 FITC-conjugated mouse mAbCD44 FITC-conjugated mouse mAbCD45 FITC-conjugated mouse mAbCD45 Pacific Blue-conjugated mouse mACD49a PE- conjugated mouse mAbCD73 PE-conjugated mouse mAbCD90 PE-conjugated mouse mAbCD133 PE-conjugated mouse mAbCD146 PE-conjugated mouse mAbCD105 PE-conjugated mouse mAbABCG2 PE-conjugated mouse mAbKDR PE-conjugated mouse mAb
Immunolabelingc-kit Goat pAbc-kit Rabbit pAbOct34 Rabbit pAbOct34 Goat pAbOct34 Mouse mAbNanog Mouse mAbNanog Rabbit pAbNkx25 Goat pAbGATA-6 Rabbit pAbEts1 Mouse mAbvWF Rabbit pAb-SMA Mouse mAb-CA Mouse mAb-SA Mouse mAbconnexin 43 Rabbit pAbp16INK4A Mouse mAbp21 Mouse mAbphospho-p53ser15 Mouse mAbH2AX Mouse mAb53BP1 Rabbit pAb
Western blottingphospho-p53ser15 Mouse mAbRan Mouse mAb
A488 and A555 Alexa Fluor 488 and 555 APC allophycocyanin Cypolyclonal antibody PE phycoerythrin TRITC tetramethylrhodamine isomeric actin -SMA -smooth muscle actin
criptase (Invitrogen) PCR amplification was performed
in a final volume of 50 L using 80 to 150 ng cDNA 10mmolL Tris-HCl pH 90 15 mmolL MgCl2 02 mmolLdNTPs 25 pmol of each primer and 2 U TaqI polymer-ase (Amersham GE Healthcare Little Chalfont UK)The PCR conditions were as follows 94degC for 2 min-utes 30 to 45 cycles at 94degC for 60 seconds 60degC for60 seconds and 72degC for 90 seconds The optimalconditions and the number of cycles were determinedto allow amplification of samples within the linearphase of the PCR The reaction products were ana-lyzed on 18 agarose gels19 Primers are listed inTable 2
Expression of c-Kit (KIT) was measured by quantitativereal-time PCR performed using a LightCycler 480 real-time PCR system (Roche Applied Sciences IndianapolisIN) Primers were designed from available human se-quences using primer analysis software Primer3 version040 (available at httpwwwroche-applied-sciencecomsisrtpcruplindexjspidUP030000) (Table 2) A dual-color assay with Hydrolysis Probe 2 and HPRT as internal
Source Fluorochrome Pseudocolor
BD APCBD PEBD FITCBD FITCBD FITCDako Pacific BlueBD PEBD PEBD PEMiltenyi Biotec PEBD PESerotec PEBD PESerotec PE
RampD A488 A555 Green redDako A488 A555 Green redAbcam A488 Yellow redSanta Cruz A555 Yellow redSanta Cruz Cy5 Yellow redAbcam A555 Cy5 Red whiteAbcam A488 MagentaSanta Cruz Cy5 WhiteSanta Cruz Cy5 YellowSanta Cruz Cy5 MagentaSigma-Aldrich A488 YellowDako A488 MagentaSigma TRITC RedSigma-Aldrich TRITC Cy5 Red whiteSanta Cruz A488 GreenNovocastra A488 GreenCalbiochem A555 YellowCell Signaling A555 RedUpstate A488 GreenCell Signaling A555 Red
Cell SignalingSigma-Aldrich
ne 5 FITC fluorescein isothiocyanate mAb monoclonal antibody pAbate vWf von Willebrand factor -CA -cardiac actinin -SA -sarco-
tting
b
5 cyani
control was used for normalization (Universal Probe Li-
CT34
352 Cesselli et alAJP July 2011 Vol 179 No 1
brary Roche) LightCycler 480 Basic software (Roche)used the second-derivative maximum method to identifythe crossing point (Cp) Glioma cell line A172 was usedas positive control
Immunofluorescence and Confocal Microscopy
The expression of pluripotency genes and cardiac cyto-plasmic proteins was determined by immunolabeling (Ta-ble 1) Image acquisition was performed with a LeicaTCS-SP2 confocal laser microscope using a 63 oilimmersion objective (numerical aperture 140) or a 40oil immersion objective (numerical aperture 125)
Migration Assay
Migration analysis was performed using a Cultrex 24-wellcell migration assay (Trevigen Gaithersburg MD) Ap-proximately 200000 cells were seeded per well Growthmedium containing 10 fetal bovine serum was used aschemoattractant
Tumorigenicity of hCSCs
The tumorigenic potential of hCSCs was established byin vitro and in vivo protocols In vitro the soft agar assaywas used 1 105 hCSCs were seeded in a 05 softagar solution placed on top of a 1 hard agar Thenumber of colonies grown within the semisolid mediumwas measured 28 days later For the in vivo analysis 1 106 hCSCs were injected subcutaneously in NODScid mice (HARLAN Italy SRL San Pietro al NatisoneItaly) SKOV-3 ovarian cancer cells were used as pos-itive control (Sigma-Aldrich St Louis MO) Animalswere kept in pathogen-free conditions and tumorgrowth was evaluated monthly for a period of 6 monthsor until SKOV-3 ovarian cancer cells generated a tumor1 cm in diameter
Measurements of Ca2 Oscillations in hCSCs
hCSCs were loaded with 10 molL Fluo-3 AM dye (In-vitrogen) and were placed on the stage of a two-photonmicroscope a BX51WI Olympus microscope (OlympusTokyo Japan) coupled with a Bio-Rad Radiance 2100MP
Table 2 PCR Primers and Product Length
Gene Forward primer
KIT 5=-CGTGGAAAAGAGAAAACAGTCA-3=POU5F1dagger 5=-GAGGATCACCCTGGGATATAC-3=NANOG 5=-AATACCTCAGCCTCCAGCAGATG-3=SOX2 5=-GTTACCTCTTCCTCCCACTCCA-3=KLF4 5=-GGGAGAAGACACTGCGTCA-3=GAPDH 5=-CGACCACTTTGTCAAGCTCA-3=HPRT 5=-CGTGATTAGTGATGATGAACCAG-3=
The protein product for the KIT gene is known also as c-kitdaggerThe protein product for POU5F1 is known also as OCT3 OCT4 or O
system (Bio-Rad Laboratories Hercules CA) Cells were
bathed with Tyrodersquos solution containing (in mmolL)NaCl 140 KCl 54 MgCl2 1 HEPES 5 glucose 55 andCaCl2 20 (pH 74 adjusted with NaOH) The Fluo-3 wasexcited at 900 to 960 nm with a Tsunami mode-lockedTisapphire femtosecond laser (Spectra-Physics New-port Corporation Irvine CA) and the emission signal wascollected at 535 nm Series of images were acquired at10-second intervals for a period of 33 minutes Changesof intracellular Ca2 in individual hCSCs were deter-mined by measuring the fluorescent signal of Fluo-3 Ineach cell the oscillations in fluorescence with time weregraphically visualized using ImageJ (NIH Bethesda MD)and Microsoft Office Excel 2003 software These traceswere used to assess the number amplitude and durationof Ca2 oscillations in hCSCs Fluo-3 signals were ex-pressed as normalized fluorescence (FF0) The durationof Ca2 oscillations was measured from the onset of therising phase to the return to baseline21 Caffeine (10mmolL Sigma-Aldrich) was used to assess ryanodinereceptor function2223
Telomere Repeat Amplification Protocol Assay
The detection of telomerase activity was performed usinga TRAPeze kit (ChemiconndashMillipore Temecula CA) forthe telomere repeat amplification protocol (TRAP) assaywith a radiolabeling detection method Samples werelabeled with 32P Radiographic films were digitized with afilm scanner (SnapScan 1236 Agfa-Gevaert MortselBelgium) and a G3 Macintosh computer (Apple Cuper-tino CA) Telomerase activity was measured on digitalimages by ImageJ software version 142q (NIH) as unitsof total product generated (TPG) calculated as TPG [(x x0)c 100][(r r0)cR] where x is the signal of theregion of the gel lane corresponding to the TRAP productladder bands from non-heat-treated samples x0 is thesignal of the region of the gel lane corresponding to theTRAP product ladder bands from heat-treated samples ris the signal of the region of the gel lane corresponding tothe TRAP product ladder bands from TSR8 quantitationcontrol r0 is the signal of the region of the gel lanecorresponding to the TRAP product ladder bands from1 CHAPS lysis buffer-only control c is the signal fromthe internal standard (S-IC) in non-heat-treated samplesand cR is the signal from the internal standard (S-IC) inTSR8 quantitation control Each unit of TPG corresponds
Reverse primerAmplicon size
(bp)
=-CACCGTGATGCCAGCTATTA-3= 70=-CGATACTGGTTCGCTTTCTCTTT-3= 295=-CTGCGTCACACCATTGCTATTCT-3= 149=-CACCCCTCCCATTTCCCTCGTT-3= 260=-GGAAGCACTGGGGGAAGT-3= 88=-AGGGGTCTACATGGCAACTG-3= 228=-CGAGCAAGACGTTCAGTCCT-3= 129
5555555
to the number of TS primers in a quantity of 1 103
Human Cardiac Stem Cell Senescence 353AJP July 2011 Vol 179 No 1
amol or 600 molecules extended with at least four telo-meric repeats by telomerase enzyme present in the ex-tract
Telomere Length in hCSCs
Telomere length was measured by flow FISH (a combi-nation of flow cytometry and fluorescence in situ hybrid-ization)24 Suspensions consisting of a 11 mixture ofhCSCs and control cells were hybridized in the presenceor absence of fluorescein-conjugated peptide nucleicacid (PNA) telomere probe (DakoCytomation GlostrupDenmark) The average telomeric fluorescence per ge-nome in the two cell classes was calculated as followsrelative telomere length RTL [(mean FL1 sample cellswith probe mean FL1 sample cells without probe) DNA index of control cells 100][(mean FL1 controlcells with probe mean FL1 control cells withoutprobe) DNA index of sample cells]
Telomere Dysfunction-Induced Foci
Telomere dysfunction-induced foci which were definedby the colocalization of 53BP1 with telomeres were an-alyzed using a Leica DMI 6000B microscope connectedto a Leica DFC350FX camera hCSCs were considered tobe TIF-positive when at least 50 of 53BP1 spots colo-calized with telomere hybridization signals The numberof TIFs per hCSC was also measured Sampling con-sisted of more than 100 hCSCs in each case (see Sup-plemental Table S3 at httpajpamjpatholorg)
Microarray Analysis
RNA Extraction cDNA Labeling and MicroarrayHybridization
hCSCs obtained from age- and sex-matched donorand explanted hearts (n 3 each) were used for theanalysis of the transcriptional profile Total RNA wasextracted with TRIzol reagent (Invitrogen) and sub-jected to DNase I treatment After RNA quality controlmicroarray cDNA targets were prepared according tothe two-step indirect fluorescent labeling procedure25
starting from 10 g total RNA First-strand cDNA syn-thesis reaction was performed with unmodified randomprimers in the presence of aminoallyl-dUTP The result-ing primary amino groups were then coupled to theN-hydroxysuccinimide ester of the fluorescent dye Foreach age- and sex- matched pair one sample waslabeled with Cy5 and the other with Cy3
LNCIB 28K cDNA microarray slides (supporting28000 cDNA clones hereafter called features) wereused in the large-scale transcript expression profilingstudies these were prepared by spotting onto AmershamType 7 Star reflective microarray slides the purified PCRproducts of 19741 LNCIB human cDNA clones26 and8160 cDNA clones selected from the human sequence-verified IMAGE clone collection (Research GeneticsInvitrogen)27 This clone set consisted of approximately
13620 unique human genes Purified cDNA inserts were
resuspended in 50 dimethyl sulfoxide (Sigma-Aldrich)and printed on the slides using a SDDC-2 ChipWriter Promicroarrayer (Virtek Vision International Waterloo ONCanada) GAPDH cDNA and -actin cDNA were used aspositive array controls and the corresponding purifiedPCR products were printed at different concentrations onthe slides Plant cDNAs and printing buffer 50 dimethylsulfoxide were used as negative controls
Competitive hybridizations were performed under cov-erslips at 63degC overnight in the presence of 3 standardsaline citrate 02 SDS 100 ngL human Cot-1 DNAand 330 ngL yeast tRNA using a semiautomated hy-bridization station (Advalytix ArrayBooster BeckmanCoulter Biomedical Munich Germany) After a washingslides were scanned at a resolution of 10 m per pixelusing an Axon GenePix 4000B microarray scanner (AxonInstrumentsndashMolecular Devices Sunnyvale CA) Imagesfor the Cy3 and Cy5 channel were acquired in a 16-bittagged-image file format (TIFF) and were analyzed usingGenePix 40 analysis software (Axon Instruments) Im-ages were manually flagged to identify artifacts and badspots
Low-level analysis of microarray data were performedin the R (version 28)Bioconductor (version 23) environ-ment using the limma microarray package (available athttpwwwR-projectorg last accessed January 2009)28
Expression data were loaded from the GenePix imageanalysis output files Features displaying in at least onechannel median fluorescence intensity lower than back-ground plus two standard deviations were flagged asbad spots Fluorescent intensity of the red and greenchannels was background-subtracted29 and the corre-sponding expression ratio (Cy5Cy3) was log-trans-formed (base 2) Within each array normalization wasaccomplished by the print-tip LOWESS method indepen-dently from each cDNA collection Normalized ratios fromreplicate expression measurements within the same ar-ray were averaged to produce a final expression matrix(FEM) to be used in the subsequent analyses The FEMwas composed of 21069 features and three age- andsex-matched paired samples
Unsupervised hierarchical cluster analysis (average-linkage clustering)30 was applied to the filtered expres-sion matrix Flagged features features not present in allsamples and features with fold changes equal to one inat least one sample were filtered out By this approach7868 features were identified For both genes and sam-ples uncorrelated distance was used as dissimilaritymeasure Cluster results were visualized using JavaTreeView31
Differential Gene Expression
The differential gene expression between hCSCs ob-tained from donor and explanted hearts was establishedby functions and methods implemented in the RBiocon-ductor environment A fixed-effects linear model was fit-ted for each individual feature to estimate expressiondifferences An empirical Bayes approach was applied tomoderate standard errors of M values32 For each ana-
lyzed feature moderated t-statistics log-odds ratios of
354 Cesselli et alAJP July 2011 Vol 179 No 1
differential expression (B-statistics) and raw and ad-justed P values (false discovery rate control by the Ben-jamini and Hochberg method33) were obtained
Functional Annotation Analysis
Differentially expressed gene lists were subjected tofunctional analysis using the DAVIDease Web-basedtool version 2008 (NIH-NIAID Bethesda MD) LNCIB28K unique associated genes were used as a back-ground list To highlight biological processes relevant tothe investigated sample groups an Ingenuity Pathwaysanalysis was performed using IPA software version 7(Ingenuity Systems Redwood City CA) For core analy-sis only genes with absolute expression value 17-foldchange (donor versus explanted heart-derived hCSCs)were considered The resulting annotations (networksbiofunctions canonical pathways) were ranked depend-ing on the number of significantly expressed genes andwere ordered by the score representing the probability offinding the observed molecules enriched in a given an-notation by chance score log(P value) The graphi-cal representation of the biofunctions was obtained byuse of the IPA Knowledge Base according to full-textarticles and information databases available online (httpwwwingenuitycomproductspathways_analysishtml lastaccessed June 2009) The spotted genes were deter-mined to be up-regulated or down-regulated from themicroarray low-level analysis
Statistical Analysis
Data are reported as means SD Differences weredetermined by two-tailed unpaired Studentrsquos t-test Linearregressions were calculated by the least squaresmethod Comparisons between slopes were made by theanalysis of covariance34 Microarray analysis was per-formed as described above
Results
Human Myocardium
Over a period of 5 years atrial tissue was obtained from63 donor hearts at the time of transplantation and from121 explanted failing hearts Atrial samples were col-lected in anatomical areas lacking apparent macro-scopic alterations By enzymatic digestion and primaryexplant technique24 a pool of small cardiac cells wasobtained in 166 of 184 samples tested which corre-sponds to a 90 rate of success Cell isolation was notfeasible in samples processed 48 hours after collectionIn successful isolations the cells were expanded andstored at passage 2 to passage 3 For the present studythe properties of hCSCs were analyzed in a subset of 41consecutive samples acquired from 18 donor and 23explanted hearts (see Supplemental Table S1 at httpajpamjpatholorg) These hearts were collected duringthe period from 2007 to 2009 at the University Hospital ofUdine In these cases cell isolation was performed within
48 hours Cell isolation and expansion was similar and
no difficulty was encountered at any stage of the cultureprotocol
Men were more numerous than women in the studysamples and ischemic cardiomyopathy predominatedThese variables may have introduced a partially biasedsampling however this reflects the sex differences andetiology of the disease in patients undergoing transplan-tation Donor subjects were younger than patients under-going transplantation who were diagnosed with ischemichypertrophic and idiopathic dilated cardiomyopathy or val-vular defects and acromegaly All patients undergoingtransplantation were class IV in the New York Heart Asso-ciation functional classification (NYHA IV) The duration ofthe underlying disease and the time from the onset ofheart failure to transplantation varied from 1 to 30 yearsand from 1 to 12 years respectively (see SupplementalTable S1 at httpajpamjpatholorg) With the exception ofone patient affected by hypertrophic cardiomyopathythe echocardiographic and hemodynamic parameterswere similar across subjects documenting severe ven-tricular dysfunction (see Supplemental Table S2 at httpajpamjpatholorg)
Human Cardiac Stem Cells
Structures with the characteristics of stem cell nicheswere identified in tissue sections of human atrial myocar-dium (Figure 1A) in which stem cells are preferentiallylocated16 Importantly c-Kit-positive hCSCs expressedOCT34 NANOG KLF4 and SOX2 which are implicatedin the self-renewing and pluripotent state of stem cells35
These properties were identified in c-Kit-positive hCSCsdistributed in atria from both donor and explanted hearts(Figures 1 BndashD and 2E) strengthening support for thenotion that hCSCs retain a highly immature embryonic-like cell phenotype
hCSCs were negative for CD45 as well as for thetranscription factor GATA4 and the myocyte cytoplasmicprotein -SA Quantitatively the number of c-Kit-positivelineage-negative hCSCs was 32-fold higher in atria ofdonor hearts than of explanted hearts (Figure 1E) Thisdifference was at variance with previous findings in theventricular myocardium of control and failing humanhearts in which cardiac decompensation was coupledwith an increase in hCSC number8 Thus regional differ-ences in hCSC number appear to be present in the failinghuman heart
In the atria of donor hearts there was 1 hCSC per 2000myocytes a frequency that is approximately fivefoldhigher than in the normal human left ventricle8 In severefailure the number of hCSCs in the atria was approxi-mately threefold lower than in the left ventricle8 The for-mer difference may reflect the site of storage of hCSCs inthe atria16 with the latter difference reflecting growthactivation of ventricular hCSCs in proximity of areas ofinjury Scarring and acute damage are distributed exclu-sively to the failing ventricle8
To validate the specificity of the recorded signalsspectral analysis was performed1720 The emissionspectra for c-Kit OCT34 NANOG and -SA were
clearly distinct from the emission spectra for tissue auto-
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

350 Cesselli et alAJP July 2011 Vol 179 No 1
2011) at httpwwwclinical trialsgov] Nonetheless ag-ing cardiac hypertrophy ischemic myocardial injuryand metabolic disorders together with genetic and envi-ronmental factors may dramatically affect the growth anddifferentiation behavior of resident hCSCs6ndash9 This possi-bility raises two critical questions First is heart failure astem cell disease characterized by severe depletion of thehCSC pool Second does a compartment of functionallycompetent hCSCs persist in the decompensated heart anddo these cells have potential therapeutic implications
As is the case for other stem cells10 the life cycle ofhCSCs is regulated by telomerase activity and telomerelength2 Telomerase is a reverse transcriptase that ex-tends the 3= chromosomal ends by using its own RNA asa template11 Telomerase activity delays but cannot pre-vent telomere erosion which is dictated by down-regu-lation of telomerase oxidative stress and loss of telom-ere-related proteins12 Shortening of telomeres beyond acritical length triggers cellular senescence which corre-sponds to irreversible growth arrest in the G1 phase withloss of specialized functions including stem cell prolifer-ation migration and differentiation Despite suggestiveevidence in humans and in genetically manipulatedmice13ndash15 that telomere shortening is a major variable ofcellular senescence and organ aging it remains to bedemonstrated whether hCSCs with critically shortened ordysfunctional telomeres undergo replicative senescenceand apoptosis and whether loss of telomere integrity isone of the variables involved in the decline of stem cellfunction in the failing human heart
These issues were addressed in the present study andthe consequences of aging and ventricular decompensa-tion on the clonogenicity multipotency and migratory prop-erties of hCSCs were characterized based on analysis ofnormal donor hearts and explanted hearts in end-stageheart failure Moreover the telomere-telomerase axis andthe presence of telomere dysfunction-induced foci (TIFs) incombination with markers of DNA damage response andreplicative senescence were determined to obtain a com-mon denominator for the processes that regulate the growthand death of hCSCs
Materials and Methods
Human Samples
Discarded atrial specimens weighing 3 to 6 g werecollected over a period of 5 years from donor hearts atthe time of transplantation and from explanted hearts ofpatients undergoing cardiac transplantation at the Car-diac Surgery Unit of the University Hospital of Udine inItaly The clinical data are presented in SupplementalTables S1 and S2 (available at httpajpamjpatholorg)Informed consent was obtained in accordance with theDeclaration of Helsinki and with approval by the Indepen-dent Ethics Committee of the University of Udine
Histological analysis of samples from donor hearts didnot show pathological changes Samples were used forthe isolation of c-Kit-positive hCSCs2ndash4 Fragments were
also fixed in formalin and embedded in paraffin2ndash4 for the
identification of stem cell niches and for the quantitativeanalysis of lineage-negative hCSCs81617 Lineage-neg-ative hCSCs were defined as cells positive for c-Kit andnegative for CD45 to exclude the bone marrow originand the mast cell phenotype and negative for GATA4and -sarcomeric actin (-SA) to exclude commitment tocardiac lineages The localization of OCT34 and NANOGwas determined in CD45-negative c-Kit-positive hCSCsMagnitude of sampling is presented in Supplemental Ta-ble S3 (available at httpajpamjpatholorg) antibodiesare listed in Table 1
Spectral Analysis
Spectral analysis was performed with a TCS-SP2 confo-cal microscope (Leica Wetzlar Germany) using thelambda acquisition mode myocardial sections were la-beled by c-Kit OCT34 and NANOG conjugated withAlexa Fluor 555 Cy5 and Alexa Fluor 555 respectivelyMyocytes were stained with -SA (Cy5) and nuclei withDAPI The emission signal for Alexa Fluor 555 was ex-cited at 543 nm with an argon laser and the fluores-cence intensity was recorded generating a lambdastack ranging from 558 to 798 nm at 49-nm intervalsThe emission signal for Cy5 was excited at 633 nm witha heliumneon laser and the fluorescence intensitywas recorded generating a lambda stack ranging from653 to 798 nm at 6-nm intervals Lens and correspond-ing numerical aperture were 63 and 14 respectivelySampling consisted of 30 c-Kit-positive OCT34-posi-tive and NANOG-positive cells Additionally 10 cellsnegative for these markers and present in the samesamples were used as control to discriminate back-ground autofluorescence from specific labeling Eachdetermination was restricted to the region of interestwithin each hCSC For each region of interest a graphplotting mean pixel intensity and the emission wave-length of the lambda stack was generated This proto-col has repeatedly been used in our laboratory17ndash20
Human Cardiac Stem Cells
In all cases atrial samples were used for the collectionof hCSCs this approach was followed to obtain a directcomparison between control and failing hearts Twoprotocols were used for the isolation of hCSCs enzy-matic dissociation of the samples with collagenase andprimary explant technique These protocols were de-veloped in the original documentation of hCSCs2 andwere retained here for continuity and comparison Aspreviously2 these two methodologies yielded compa-rable results up to 20 to 25 population doublings effi-ciency and viability of hCSCs were superimposableCollagenase treatment was not found to affect thesevariables The phenotype of the small cell populationwas defined by flow-cytometry and immunolabeling24
Single-cell suspensions were labeled with directly con-jugated antibodies (Table 1) specific for hematopoieticand endothelial cell (EC) lineages Conjugated isotypeantibodies were used as negative control To exclude
the presence of erythrocytes the samples of freshly
thiocyan
Human Cardiac Stem Cell Senescence 351AJP July 2011 Vol 179 No 1
isolated cells were exposed to BD fluorescence-acti-vated cell sorting (FACS) lysing solution the flow cy-tometric analysis was performed with a BD FACScansystem (BD Biosciences San Jose CA) or a CyAnsystem (Beckman Coulter Fullerton CA) For immuno-labeling hCSCs were fixed in 4 paraformaldehydefor 15 minutes at room temperature and the expressionof c-Kit OCT34 NANOG and SOX2 was evalu-ated2417 For clonal analysis a FACSAria cell sorter(BD-Biosciences) was used to seed single c-Kit-posi-tive hCSCs into 96-well Terasaki plates24
RT-PCR
Total RNA was extracted from nonconfluent cultures ofhCSCs at passage 3 using TRIzol reagent (InvitrogenCarlsbad CA) After treatment with DNase I (AmbionApplied Biosystems Austin TX) first-strand cDNAsynthesis was performed with 1 g total RNA usingrandom hexanucleotides and M-MLV reverse trans-
Table 1 Antibodies for FACS Immunolabeling and Western Blo
Antigen Antibody
FACSc-kit APC-conjugated mouse mAbc-kit PE-conjugated mouse mAbCD34 FITC-conjugated mouse mAbCD44 FITC-conjugated mouse mAbCD45 FITC-conjugated mouse mAbCD45 Pacific Blue-conjugated mouse mACD49a PE- conjugated mouse mAbCD73 PE-conjugated mouse mAbCD90 PE-conjugated mouse mAbCD133 PE-conjugated mouse mAbCD146 PE-conjugated mouse mAbCD105 PE-conjugated mouse mAbABCG2 PE-conjugated mouse mAbKDR PE-conjugated mouse mAb
Immunolabelingc-kit Goat pAbc-kit Rabbit pAbOct34 Rabbit pAbOct34 Goat pAbOct34 Mouse mAbNanog Mouse mAbNanog Rabbit pAbNkx25 Goat pAbGATA-6 Rabbit pAbEts1 Mouse mAbvWF Rabbit pAb-SMA Mouse mAb-CA Mouse mAb-SA Mouse mAbconnexin 43 Rabbit pAbp16INK4A Mouse mAbp21 Mouse mAbphospho-p53ser15 Mouse mAbH2AX Mouse mAb53BP1 Rabbit pAb
Western blottingphospho-p53ser15 Mouse mAbRan Mouse mAb
A488 and A555 Alexa Fluor 488 and 555 APC allophycocyanin Cypolyclonal antibody PE phycoerythrin TRITC tetramethylrhodamine isomeric actin -SMA -smooth muscle actin
criptase (Invitrogen) PCR amplification was performed
in a final volume of 50 L using 80 to 150 ng cDNA 10mmolL Tris-HCl pH 90 15 mmolL MgCl2 02 mmolLdNTPs 25 pmol of each primer and 2 U TaqI polymer-ase (Amersham GE Healthcare Little Chalfont UK)The PCR conditions were as follows 94degC for 2 min-utes 30 to 45 cycles at 94degC for 60 seconds 60degC for60 seconds and 72degC for 90 seconds The optimalconditions and the number of cycles were determinedto allow amplification of samples within the linearphase of the PCR The reaction products were ana-lyzed on 18 agarose gels19 Primers are listed inTable 2
Expression of c-Kit (KIT) was measured by quantitativereal-time PCR performed using a LightCycler 480 real-time PCR system (Roche Applied Sciences IndianapolisIN) Primers were designed from available human se-quences using primer analysis software Primer3 version040 (available at httpwwwroche-applied-sciencecomsisrtpcruplindexjspidUP030000) (Table 2) A dual-color assay with Hydrolysis Probe 2 and HPRT as internal
Source Fluorochrome Pseudocolor
BD APCBD PEBD FITCBD FITCBD FITCDako Pacific BlueBD PEBD PEBD PEMiltenyi Biotec PEBD PESerotec PEBD PESerotec PE
RampD A488 A555 Green redDako A488 A555 Green redAbcam A488 Yellow redSanta Cruz A555 Yellow redSanta Cruz Cy5 Yellow redAbcam A555 Cy5 Red whiteAbcam A488 MagentaSanta Cruz Cy5 WhiteSanta Cruz Cy5 YellowSanta Cruz Cy5 MagentaSigma-Aldrich A488 YellowDako A488 MagentaSigma TRITC RedSigma-Aldrich TRITC Cy5 Red whiteSanta Cruz A488 GreenNovocastra A488 GreenCalbiochem A555 YellowCell Signaling A555 RedUpstate A488 GreenCell Signaling A555 Red
Cell SignalingSigma-Aldrich
ne 5 FITC fluorescein isothiocyanate mAb monoclonal antibody pAbate vWf von Willebrand factor -CA -cardiac actinin -SA -sarco-
tting
b
5 cyani
control was used for normalization (Universal Probe Li-
CT34
352 Cesselli et alAJP July 2011 Vol 179 No 1
brary Roche) LightCycler 480 Basic software (Roche)used the second-derivative maximum method to identifythe crossing point (Cp) Glioma cell line A172 was usedas positive control
Immunofluorescence and Confocal Microscopy
The expression of pluripotency genes and cardiac cyto-plasmic proteins was determined by immunolabeling (Ta-ble 1) Image acquisition was performed with a LeicaTCS-SP2 confocal laser microscope using a 63 oilimmersion objective (numerical aperture 140) or a 40oil immersion objective (numerical aperture 125)
Migration Assay
Migration analysis was performed using a Cultrex 24-wellcell migration assay (Trevigen Gaithersburg MD) Ap-proximately 200000 cells were seeded per well Growthmedium containing 10 fetal bovine serum was used aschemoattractant
Tumorigenicity of hCSCs
The tumorigenic potential of hCSCs was established byin vitro and in vivo protocols In vitro the soft agar assaywas used 1 105 hCSCs were seeded in a 05 softagar solution placed on top of a 1 hard agar Thenumber of colonies grown within the semisolid mediumwas measured 28 days later For the in vivo analysis 1 106 hCSCs were injected subcutaneously in NODScid mice (HARLAN Italy SRL San Pietro al NatisoneItaly) SKOV-3 ovarian cancer cells were used as pos-itive control (Sigma-Aldrich St Louis MO) Animalswere kept in pathogen-free conditions and tumorgrowth was evaluated monthly for a period of 6 monthsor until SKOV-3 ovarian cancer cells generated a tumor1 cm in diameter
Measurements of Ca2 Oscillations in hCSCs
hCSCs were loaded with 10 molL Fluo-3 AM dye (In-vitrogen) and were placed on the stage of a two-photonmicroscope a BX51WI Olympus microscope (OlympusTokyo Japan) coupled with a Bio-Rad Radiance 2100MP
Table 2 PCR Primers and Product Length
Gene Forward primer
KIT 5=-CGTGGAAAAGAGAAAACAGTCA-3=POU5F1dagger 5=-GAGGATCACCCTGGGATATAC-3=NANOG 5=-AATACCTCAGCCTCCAGCAGATG-3=SOX2 5=-GTTACCTCTTCCTCCCACTCCA-3=KLF4 5=-GGGAGAAGACACTGCGTCA-3=GAPDH 5=-CGACCACTTTGTCAAGCTCA-3=HPRT 5=-CGTGATTAGTGATGATGAACCAG-3=
The protein product for the KIT gene is known also as c-kitdaggerThe protein product for POU5F1 is known also as OCT3 OCT4 or O
system (Bio-Rad Laboratories Hercules CA) Cells were
bathed with Tyrodersquos solution containing (in mmolL)NaCl 140 KCl 54 MgCl2 1 HEPES 5 glucose 55 andCaCl2 20 (pH 74 adjusted with NaOH) The Fluo-3 wasexcited at 900 to 960 nm with a Tsunami mode-lockedTisapphire femtosecond laser (Spectra-Physics New-port Corporation Irvine CA) and the emission signal wascollected at 535 nm Series of images were acquired at10-second intervals for a period of 33 minutes Changesof intracellular Ca2 in individual hCSCs were deter-mined by measuring the fluorescent signal of Fluo-3 Ineach cell the oscillations in fluorescence with time weregraphically visualized using ImageJ (NIH Bethesda MD)and Microsoft Office Excel 2003 software These traceswere used to assess the number amplitude and durationof Ca2 oscillations in hCSCs Fluo-3 signals were ex-pressed as normalized fluorescence (FF0) The durationof Ca2 oscillations was measured from the onset of therising phase to the return to baseline21 Caffeine (10mmolL Sigma-Aldrich) was used to assess ryanodinereceptor function2223
Telomere Repeat Amplification Protocol Assay
The detection of telomerase activity was performed usinga TRAPeze kit (ChemiconndashMillipore Temecula CA) forthe telomere repeat amplification protocol (TRAP) assaywith a radiolabeling detection method Samples werelabeled with 32P Radiographic films were digitized with afilm scanner (SnapScan 1236 Agfa-Gevaert MortselBelgium) and a G3 Macintosh computer (Apple Cuper-tino CA) Telomerase activity was measured on digitalimages by ImageJ software version 142q (NIH) as unitsof total product generated (TPG) calculated as TPG [(x x0)c 100][(r r0)cR] where x is the signal of theregion of the gel lane corresponding to the TRAP productladder bands from non-heat-treated samples x0 is thesignal of the region of the gel lane corresponding to theTRAP product ladder bands from heat-treated samples ris the signal of the region of the gel lane corresponding tothe TRAP product ladder bands from TSR8 quantitationcontrol r0 is the signal of the region of the gel lanecorresponding to the TRAP product ladder bands from1 CHAPS lysis buffer-only control c is the signal fromthe internal standard (S-IC) in non-heat-treated samplesand cR is the signal from the internal standard (S-IC) inTSR8 quantitation control Each unit of TPG corresponds
Reverse primerAmplicon size
(bp)
=-CACCGTGATGCCAGCTATTA-3= 70=-CGATACTGGTTCGCTTTCTCTTT-3= 295=-CTGCGTCACACCATTGCTATTCT-3= 149=-CACCCCTCCCATTTCCCTCGTT-3= 260=-GGAAGCACTGGGGGAAGT-3= 88=-AGGGGTCTACATGGCAACTG-3= 228=-CGAGCAAGACGTTCAGTCCT-3= 129
5555555
to the number of TS primers in a quantity of 1 103
Human Cardiac Stem Cell Senescence 353AJP July 2011 Vol 179 No 1
amol or 600 molecules extended with at least four telo-meric repeats by telomerase enzyme present in the ex-tract
Telomere Length in hCSCs
Telomere length was measured by flow FISH (a combi-nation of flow cytometry and fluorescence in situ hybrid-ization)24 Suspensions consisting of a 11 mixture ofhCSCs and control cells were hybridized in the presenceor absence of fluorescein-conjugated peptide nucleicacid (PNA) telomere probe (DakoCytomation GlostrupDenmark) The average telomeric fluorescence per ge-nome in the two cell classes was calculated as followsrelative telomere length RTL [(mean FL1 sample cellswith probe mean FL1 sample cells without probe) DNA index of control cells 100][(mean FL1 controlcells with probe mean FL1 control cells withoutprobe) DNA index of sample cells]
Telomere Dysfunction-Induced Foci
Telomere dysfunction-induced foci which were definedby the colocalization of 53BP1 with telomeres were an-alyzed using a Leica DMI 6000B microscope connectedto a Leica DFC350FX camera hCSCs were considered tobe TIF-positive when at least 50 of 53BP1 spots colo-calized with telomere hybridization signals The numberof TIFs per hCSC was also measured Sampling con-sisted of more than 100 hCSCs in each case (see Sup-plemental Table S3 at httpajpamjpatholorg)
Microarray Analysis
RNA Extraction cDNA Labeling and MicroarrayHybridization
hCSCs obtained from age- and sex-matched donorand explanted hearts (n 3 each) were used for theanalysis of the transcriptional profile Total RNA wasextracted with TRIzol reagent (Invitrogen) and sub-jected to DNase I treatment After RNA quality controlmicroarray cDNA targets were prepared according tothe two-step indirect fluorescent labeling procedure25
starting from 10 g total RNA First-strand cDNA syn-thesis reaction was performed with unmodified randomprimers in the presence of aminoallyl-dUTP The result-ing primary amino groups were then coupled to theN-hydroxysuccinimide ester of the fluorescent dye Foreach age- and sex- matched pair one sample waslabeled with Cy5 and the other with Cy3
LNCIB 28K cDNA microarray slides (supporting28000 cDNA clones hereafter called features) wereused in the large-scale transcript expression profilingstudies these were prepared by spotting onto AmershamType 7 Star reflective microarray slides the purified PCRproducts of 19741 LNCIB human cDNA clones26 and8160 cDNA clones selected from the human sequence-verified IMAGE clone collection (Research GeneticsInvitrogen)27 This clone set consisted of approximately
13620 unique human genes Purified cDNA inserts were
resuspended in 50 dimethyl sulfoxide (Sigma-Aldrich)and printed on the slides using a SDDC-2 ChipWriter Promicroarrayer (Virtek Vision International Waterloo ONCanada) GAPDH cDNA and -actin cDNA were used aspositive array controls and the corresponding purifiedPCR products were printed at different concentrations onthe slides Plant cDNAs and printing buffer 50 dimethylsulfoxide were used as negative controls
Competitive hybridizations were performed under cov-erslips at 63degC overnight in the presence of 3 standardsaline citrate 02 SDS 100 ngL human Cot-1 DNAand 330 ngL yeast tRNA using a semiautomated hy-bridization station (Advalytix ArrayBooster BeckmanCoulter Biomedical Munich Germany) After a washingslides were scanned at a resolution of 10 m per pixelusing an Axon GenePix 4000B microarray scanner (AxonInstrumentsndashMolecular Devices Sunnyvale CA) Imagesfor the Cy3 and Cy5 channel were acquired in a 16-bittagged-image file format (TIFF) and were analyzed usingGenePix 40 analysis software (Axon Instruments) Im-ages were manually flagged to identify artifacts and badspots
Low-level analysis of microarray data were performedin the R (version 28)Bioconductor (version 23) environ-ment using the limma microarray package (available athttpwwwR-projectorg last accessed January 2009)28
Expression data were loaded from the GenePix imageanalysis output files Features displaying in at least onechannel median fluorescence intensity lower than back-ground plus two standard deviations were flagged asbad spots Fluorescent intensity of the red and greenchannels was background-subtracted29 and the corre-sponding expression ratio (Cy5Cy3) was log-trans-formed (base 2) Within each array normalization wasaccomplished by the print-tip LOWESS method indepen-dently from each cDNA collection Normalized ratios fromreplicate expression measurements within the same ar-ray were averaged to produce a final expression matrix(FEM) to be used in the subsequent analyses The FEMwas composed of 21069 features and three age- andsex-matched paired samples
Unsupervised hierarchical cluster analysis (average-linkage clustering)30 was applied to the filtered expres-sion matrix Flagged features features not present in allsamples and features with fold changes equal to one inat least one sample were filtered out By this approach7868 features were identified For both genes and sam-ples uncorrelated distance was used as dissimilaritymeasure Cluster results were visualized using JavaTreeView31
Differential Gene Expression
The differential gene expression between hCSCs ob-tained from donor and explanted hearts was establishedby functions and methods implemented in the RBiocon-ductor environment A fixed-effects linear model was fit-ted for each individual feature to estimate expressiondifferences An empirical Bayes approach was applied tomoderate standard errors of M values32 For each ana-
lyzed feature moderated t-statistics log-odds ratios of
354 Cesselli et alAJP July 2011 Vol 179 No 1
differential expression (B-statistics) and raw and ad-justed P values (false discovery rate control by the Ben-jamini and Hochberg method33) were obtained
Functional Annotation Analysis
Differentially expressed gene lists were subjected tofunctional analysis using the DAVIDease Web-basedtool version 2008 (NIH-NIAID Bethesda MD) LNCIB28K unique associated genes were used as a back-ground list To highlight biological processes relevant tothe investigated sample groups an Ingenuity Pathwaysanalysis was performed using IPA software version 7(Ingenuity Systems Redwood City CA) For core analy-sis only genes with absolute expression value 17-foldchange (donor versus explanted heart-derived hCSCs)were considered The resulting annotations (networksbiofunctions canonical pathways) were ranked depend-ing on the number of significantly expressed genes andwere ordered by the score representing the probability offinding the observed molecules enriched in a given an-notation by chance score log(P value) The graphi-cal representation of the biofunctions was obtained byuse of the IPA Knowledge Base according to full-textarticles and information databases available online (httpwwwingenuitycomproductspathways_analysishtml lastaccessed June 2009) The spotted genes were deter-mined to be up-regulated or down-regulated from themicroarray low-level analysis
Statistical Analysis
Data are reported as means SD Differences weredetermined by two-tailed unpaired Studentrsquos t-test Linearregressions were calculated by the least squaresmethod Comparisons between slopes were made by theanalysis of covariance34 Microarray analysis was per-formed as described above
Results
Human Myocardium
Over a period of 5 years atrial tissue was obtained from63 donor hearts at the time of transplantation and from121 explanted failing hearts Atrial samples were col-lected in anatomical areas lacking apparent macro-scopic alterations By enzymatic digestion and primaryexplant technique24 a pool of small cardiac cells wasobtained in 166 of 184 samples tested which corre-sponds to a 90 rate of success Cell isolation was notfeasible in samples processed 48 hours after collectionIn successful isolations the cells were expanded andstored at passage 2 to passage 3 For the present studythe properties of hCSCs were analyzed in a subset of 41consecutive samples acquired from 18 donor and 23explanted hearts (see Supplemental Table S1 at httpajpamjpatholorg) These hearts were collected duringthe period from 2007 to 2009 at the University Hospital ofUdine In these cases cell isolation was performed within
48 hours Cell isolation and expansion was similar and
no difficulty was encountered at any stage of the cultureprotocol
Men were more numerous than women in the studysamples and ischemic cardiomyopathy predominatedThese variables may have introduced a partially biasedsampling however this reflects the sex differences andetiology of the disease in patients undergoing transplan-tation Donor subjects were younger than patients under-going transplantation who were diagnosed with ischemichypertrophic and idiopathic dilated cardiomyopathy or val-vular defects and acromegaly All patients undergoingtransplantation were class IV in the New York Heart Asso-ciation functional classification (NYHA IV) The duration ofthe underlying disease and the time from the onset ofheart failure to transplantation varied from 1 to 30 yearsand from 1 to 12 years respectively (see SupplementalTable S1 at httpajpamjpatholorg) With the exception ofone patient affected by hypertrophic cardiomyopathythe echocardiographic and hemodynamic parameterswere similar across subjects documenting severe ven-tricular dysfunction (see Supplemental Table S2 at httpajpamjpatholorg)
Human Cardiac Stem Cells
Structures with the characteristics of stem cell nicheswere identified in tissue sections of human atrial myocar-dium (Figure 1A) in which stem cells are preferentiallylocated16 Importantly c-Kit-positive hCSCs expressedOCT34 NANOG KLF4 and SOX2 which are implicatedin the self-renewing and pluripotent state of stem cells35
These properties were identified in c-Kit-positive hCSCsdistributed in atria from both donor and explanted hearts(Figures 1 BndashD and 2E) strengthening support for thenotion that hCSCs retain a highly immature embryonic-like cell phenotype
hCSCs were negative for CD45 as well as for thetranscription factor GATA4 and the myocyte cytoplasmicprotein -SA Quantitatively the number of c-Kit-positivelineage-negative hCSCs was 32-fold higher in atria ofdonor hearts than of explanted hearts (Figure 1E) Thisdifference was at variance with previous findings in theventricular myocardium of control and failing humanhearts in which cardiac decompensation was coupledwith an increase in hCSC number8 Thus regional differ-ences in hCSC number appear to be present in the failinghuman heart
In the atria of donor hearts there was 1 hCSC per 2000myocytes a frequency that is approximately fivefoldhigher than in the normal human left ventricle8 In severefailure the number of hCSCs in the atria was approxi-mately threefold lower than in the left ventricle8 The for-mer difference may reflect the site of storage of hCSCs inthe atria16 with the latter difference reflecting growthactivation of ventricular hCSCs in proximity of areas ofinjury Scarring and acute damage are distributed exclu-sively to the failing ventricle8
To validate the specificity of the recorded signalsspectral analysis was performed1720 The emissionspectra for c-Kit OCT34 NANOG and -SA were
clearly distinct from the emission spectra for tissue auto-
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

thiocyan
Human Cardiac Stem Cell Senescence 351AJP July 2011 Vol 179 No 1
isolated cells were exposed to BD fluorescence-acti-vated cell sorting (FACS) lysing solution the flow cy-tometric analysis was performed with a BD FACScansystem (BD Biosciences San Jose CA) or a CyAnsystem (Beckman Coulter Fullerton CA) For immuno-labeling hCSCs were fixed in 4 paraformaldehydefor 15 minutes at room temperature and the expressionof c-Kit OCT34 NANOG and SOX2 was evalu-ated2417 For clonal analysis a FACSAria cell sorter(BD-Biosciences) was used to seed single c-Kit-posi-tive hCSCs into 96-well Terasaki plates24
RT-PCR
Total RNA was extracted from nonconfluent cultures ofhCSCs at passage 3 using TRIzol reagent (InvitrogenCarlsbad CA) After treatment with DNase I (AmbionApplied Biosystems Austin TX) first-strand cDNAsynthesis was performed with 1 g total RNA usingrandom hexanucleotides and M-MLV reverse trans-
Table 1 Antibodies for FACS Immunolabeling and Western Blo
Antigen Antibody
FACSc-kit APC-conjugated mouse mAbc-kit PE-conjugated mouse mAbCD34 FITC-conjugated mouse mAbCD44 FITC-conjugated mouse mAbCD45 FITC-conjugated mouse mAbCD45 Pacific Blue-conjugated mouse mACD49a PE- conjugated mouse mAbCD73 PE-conjugated mouse mAbCD90 PE-conjugated mouse mAbCD133 PE-conjugated mouse mAbCD146 PE-conjugated mouse mAbCD105 PE-conjugated mouse mAbABCG2 PE-conjugated mouse mAbKDR PE-conjugated mouse mAb
Immunolabelingc-kit Goat pAbc-kit Rabbit pAbOct34 Rabbit pAbOct34 Goat pAbOct34 Mouse mAbNanog Mouse mAbNanog Rabbit pAbNkx25 Goat pAbGATA-6 Rabbit pAbEts1 Mouse mAbvWF Rabbit pAb-SMA Mouse mAb-CA Mouse mAb-SA Mouse mAbconnexin 43 Rabbit pAbp16INK4A Mouse mAbp21 Mouse mAbphospho-p53ser15 Mouse mAbH2AX Mouse mAb53BP1 Rabbit pAb
Western blottingphospho-p53ser15 Mouse mAbRan Mouse mAb
A488 and A555 Alexa Fluor 488 and 555 APC allophycocyanin Cypolyclonal antibody PE phycoerythrin TRITC tetramethylrhodamine isomeric actin -SMA -smooth muscle actin
criptase (Invitrogen) PCR amplification was performed
in a final volume of 50 L using 80 to 150 ng cDNA 10mmolL Tris-HCl pH 90 15 mmolL MgCl2 02 mmolLdNTPs 25 pmol of each primer and 2 U TaqI polymer-ase (Amersham GE Healthcare Little Chalfont UK)The PCR conditions were as follows 94degC for 2 min-utes 30 to 45 cycles at 94degC for 60 seconds 60degC for60 seconds and 72degC for 90 seconds The optimalconditions and the number of cycles were determinedto allow amplification of samples within the linearphase of the PCR The reaction products were ana-lyzed on 18 agarose gels19 Primers are listed inTable 2
Expression of c-Kit (KIT) was measured by quantitativereal-time PCR performed using a LightCycler 480 real-time PCR system (Roche Applied Sciences IndianapolisIN) Primers were designed from available human se-quences using primer analysis software Primer3 version040 (available at httpwwwroche-applied-sciencecomsisrtpcruplindexjspidUP030000) (Table 2) A dual-color assay with Hydrolysis Probe 2 and HPRT as internal
Source Fluorochrome Pseudocolor
BD APCBD PEBD FITCBD FITCBD FITCDako Pacific BlueBD PEBD PEBD PEMiltenyi Biotec PEBD PESerotec PEBD PESerotec PE
RampD A488 A555 Green redDako A488 A555 Green redAbcam A488 Yellow redSanta Cruz A555 Yellow redSanta Cruz Cy5 Yellow redAbcam A555 Cy5 Red whiteAbcam A488 MagentaSanta Cruz Cy5 WhiteSanta Cruz Cy5 YellowSanta Cruz Cy5 MagentaSigma-Aldrich A488 YellowDako A488 MagentaSigma TRITC RedSigma-Aldrich TRITC Cy5 Red whiteSanta Cruz A488 GreenNovocastra A488 GreenCalbiochem A555 YellowCell Signaling A555 RedUpstate A488 GreenCell Signaling A555 Red
Cell SignalingSigma-Aldrich
ne 5 FITC fluorescein isothiocyanate mAb monoclonal antibody pAbate vWf von Willebrand factor -CA -cardiac actinin -SA -sarco-
tting
b
5 cyani
control was used for normalization (Universal Probe Li-
CT34
352 Cesselli et alAJP July 2011 Vol 179 No 1
brary Roche) LightCycler 480 Basic software (Roche)used the second-derivative maximum method to identifythe crossing point (Cp) Glioma cell line A172 was usedas positive control
Immunofluorescence and Confocal Microscopy
The expression of pluripotency genes and cardiac cyto-plasmic proteins was determined by immunolabeling (Ta-ble 1) Image acquisition was performed with a LeicaTCS-SP2 confocal laser microscope using a 63 oilimmersion objective (numerical aperture 140) or a 40oil immersion objective (numerical aperture 125)
Migration Assay
Migration analysis was performed using a Cultrex 24-wellcell migration assay (Trevigen Gaithersburg MD) Ap-proximately 200000 cells were seeded per well Growthmedium containing 10 fetal bovine serum was used aschemoattractant
Tumorigenicity of hCSCs
The tumorigenic potential of hCSCs was established byin vitro and in vivo protocols In vitro the soft agar assaywas used 1 105 hCSCs were seeded in a 05 softagar solution placed on top of a 1 hard agar Thenumber of colonies grown within the semisolid mediumwas measured 28 days later For the in vivo analysis 1 106 hCSCs were injected subcutaneously in NODScid mice (HARLAN Italy SRL San Pietro al NatisoneItaly) SKOV-3 ovarian cancer cells were used as pos-itive control (Sigma-Aldrich St Louis MO) Animalswere kept in pathogen-free conditions and tumorgrowth was evaluated monthly for a period of 6 monthsor until SKOV-3 ovarian cancer cells generated a tumor1 cm in diameter
Measurements of Ca2 Oscillations in hCSCs
hCSCs were loaded with 10 molL Fluo-3 AM dye (In-vitrogen) and were placed on the stage of a two-photonmicroscope a BX51WI Olympus microscope (OlympusTokyo Japan) coupled with a Bio-Rad Radiance 2100MP
Table 2 PCR Primers and Product Length
Gene Forward primer
KIT 5=-CGTGGAAAAGAGAAAACAGTCA-3=POU5F1dagger 5=-GAGGATCACCCTGGGATATAC-3=NANOG 5=-AATACCTCAGCCTCCAGCAGATG-3=SOX2 5=-GTTACCTCTTCCTCCCACTCCA-3=KLF4 5=-GGGAGAAGACACTGCGTCA-3=GAPDH 5=-CGACCACTTTGTCAAGCTCA-3=HPRT 5=-CGTGATTAGTGATGATGAACCAG-3=
The protein product for the KIT gene is known also as c-kitdaggerThe protein product for POU5F1 is known also as OCT3 OCT4 or O
system (Bio-Rad Laboratories Hercules CA) Cells were
bathed with Tyrodersquos solution containing (in mmolL)NaCl 140 KCl 54 MgCl2 1 HEPES 5 glucose 55 andCaCl2 20 (pH 74 adjusted with NaOH) The Fluo-3 wasexcited at 900 to 960 nm with a Tsunami mode-lockedTisapphire femtosecond laser (Spectra-Physics New-port Corporation Irvine CA) and the emission signal wascollected at 535 nm Series of images were acquired at10-second intervals for a period of 33 minutes Changesof intracellular Ca2 in individual hCSCs were deter-mined by measuring the fluorescent signal of Fluo-3 Ineach cell the oscillations in fluorescence with time weregraphically visualized using ImageJ (NIH Bethesda MD)and Microsoft Office Excel 2003 software These traceswere used to assess the number amplitude and durationof Ca2 oscillations in hCSCs Fluo-3 signals were ex-pressed as normalized fluorescence (FF0) The durationof Ca2 oscillations was measured from the onset of therising phase to the return to baseline21 Caffeine (10mmolL Sigma-Aldrich) was used to assess ryanodinereceptor function2223
Telomere Repeat Amplification Protocol Assay
The detection of telomerase activity was performed usinga TRAPeze kit (ChemiconndashMillipore Temecula CA) forthe telomere repeat amplification protocol (TRAP) assaywith a radiolabeling detection method Samples werelabeled with 32P Radiographic films were digitized with afilm scanner (SnapScan 1236 Agfa-Gevaert MortselBelgium) and a G3 Macintosh computer (Apple Cuper-tino CA) Telomerase activity was measured on digitalimages by ImageJ software version 142q (NIH) as unitsof total product generated (TPG) calculated as TPG [(x x0)c 100][(r r0)cR] where x is the signal of theregion of the gel lane corresponding to the TRAP productladder bands from non-heat-treated samples x0 is thesignal of the region of the gel lane corresponding to theTRAP product ladder bands from heat-treated samples ris the signal of the region of the gel lane corresponding tothe TRAP product ladder bands from TSR8 quantitationcontrol r0 is the signal of the region of the gel lanecorresponding to the TRAP product ladder bands from1 CHAPS lysis buffer-only control c is the signal fromthe internal standard (S-IC) in non-heat-treated samplesand cR is the signal from the internal standard (S-IC) inTSR8 quantitation control Each unit of TPG corresponds
Reverse primerAmplicon size
(bp)
=-CACCGTGATGCCAGCTATTA-3= 70=-CGATACTGGTTCGCTTTCTCTTT-3= 295=-CTGCGTCACACCATTGCTATTCT-3= 149=-CACCCCTCCCATTTCCCTCGTT-3= 260=-GGAAGCACTGGGGGAAGT-3= 88=-AGGGGTCTACATGGCAACTG-3= 228=-CGAGCAAGACGTTCAGTCCT-3= 129
5555555
to the number of TS primers in a quantity of 1 103
Human Cardiac Stem Cell Senescence 353AJP July 2011 Vol 179 No 1
amol or 600 molecules extended with at least four telo-meric repeats by telomerase enzyme present in the ex-tract
Telomere Length in hCSCs
Telomere length was measured by flow FISH (a combi-nation of flow cytometry and fluorescence in situ hybrid-ization)24 Suspensions consisting of a 11 mixture ofhCSCs and control cells were hybridized in the presenceor absence of fluorescein-conjugated peptide nucleicacid (PNA) telomere probe (DakoCytomation GlostrupDenmark) The average telomeric fluorescence per ge-nome in the two cell classes was calculated as followsrelative telomere length RTL [(mean FL1 sample cellswith probe mean FL1 sample cells without probe) DNA index of control cells 100][(mean FL1 controlcells with probe mean FL1 control cells withoutprobe) DNA index of sample cells]
Telomere Dysfunction-Induced Foci
Telomere dysfunction-induced foci which were definedby the colocalization of 53BP1 with telomeres were an-alyzed using a Leica DMI 6000B microscope connectedto a Leica DFC350FX camera hCSCs were considered tobe TIF-positive when at least 50 of 53BP1 spots colo-calized with telomere hybridization signals The numberof TIFs per hCSC was also measured Sampling con-sisted of more than 100 hCSCs in each case (see Sup-plemental Table S3 at httpajpamjpatholorg)
Microarray Analysis
RNA Extraction cDNA Labeling and MicroarrayHybridization
hCSCs obtained from age- and sex-matched donorand explanted hearts (n 3 each) were used for theanalysis of the transcriptional profile Total RNA wasextracted with TRIzol reagent (Invitrogen) and sub-jected to DNase I treatment After RNA quality controlmicroarray cDNA targets were prepared according tothe two-step indirect fluorescent labeling procedure25
starting from 10 g total RNA First-strand cDNA syn-thesis reaction was performed with unmodified randomprimers in the presence of aminoallyl-dUTP The result-ing primary amino groups were then coupled to theN-hydroxysuccinimide ester of the fluorescent dye Foreach age- and sex- matched pair one sample waslabeled with Cy5 and the other with Cy3
LNCIB 28K cDNA microarray slides (supporting28000 cDNA clones hereafter called features) wereused in the large-scale transcript expression profilingstudies these were prepared by spotting onto AmershamType 7 Star reflective microarray slides the purified PCRproducts of 19741 LNCIB human cDNA clones26 and8160 cDNA clones selected from the human sequence-verified IMAGE clone collection (Research GeneticsInvitrogen)27 This clone set consisted of approximately
13620 unique human genes Purified cDNA inserts were
resuspended in 50 dimethyl sulfoxide (Sigma-Aldrich)and printed on the slides using a SDDC-2 ChipWriter Promicroarrayer (Virtek Vision International Waterloo ONCanada) GAPDH cDNA and -actin cDNA were used aspositive array controls and the corresponding purifiedPCR products were printed at different concentrations onthe slides Plant cDNAs and printing buffer 50 dimethylsulfoxide were used as negative controls
Competitive hybridizations were performed under cov-erslips at 63degC overnight in the presence of 3 standardsaline citrate 02 SDS 100 ngL human Cot-1 DNAand 330 ngL yeast tRNA using a semiautomated hy-bridization station (Advalytix ArrayBooster BeckmanCoulter Biomedical Munich Germany) After a washingslides were scanned at a resolution of 10 m per pixelusing an Axon GenePix 4000B microarray scanner (AxonInstrumentsndashMolecular Devices Sunnyvale CA) Imagesfor the Cy3 and Cy5 channel were acquired in a 16-bittagged-image file format (TIFF) and were analyzed usingGenePix 40 analysis software (Axon Instruments) Im-ages were manually flagged to identify artifacts and badspots
Low-level analysis of microarray data were performedin the R (version 28)Bioconductor (version 23) environ-ment using the limma microarray package (available athttpwwwR-projectorg last accessed January 2009)28
Expression data were loaded from the GenePix imageanalysis output files Features displaying in at least onechannel median fluorescence intensity lower than back-ground plus two standard deviations were flagged asbad spots Fluorescent intensity of the red and greenchannels was background-subtracted29 and the corre-sponding expression ratio (Cy5Cy3) was log-trans-formed (base 2) Within each array normalization wasaccomplished by the print-tip LOWESS method indepen-dently from each cDNA collection Normalized ratios fromreplicate expression measurements within the same ar-ray were averaged to produce a final expression matrix(FEM) to be used in the subsequent analyses The FEMwas composed of 21069 features and three age- andsex-matched paired samples
Unsupervised hierarchical cluster analysis (average-linkage clustering)30 was applied to the filtered expres-sion matrix Flagged features features not present in allsamples and features with fold changes equal to one inat least one sample were filtered out By this approach7868 features were identified For both genes and sam-ples uncorrelated distance was used as dissimilaritymeasure Cluster results were visualized using JavaTreeView31
Differential Gene Expression
The differential gene expression between hCSCs ob-tained from donor and explanted hearts was establishedby functions and methods implemented in the RBiocon-ductor environment A fixed-effects linear model was fit-ted for each individual feature to estimate expressiondifferences An empirical Bayes approach was applied tomoderate standard errors of M values32 For each ana-
lyzed feature moderated t-statistics log-odds ratios of
354 Cesselli et alAJP July 2011 Vol 179 No 1
differential expression (B-statistics) and raw and ad-justed P values (false discovery rate control by the Ben-jamini and Hochberg method33) were obtained
Functional Annotation Analysis
Differentially expressed gene lists were subjected tofunctional analysis using the DAVIDease Web-basedtool version 2008 (NIH-NIAID Bethesda MD) LNCIB28K unique associated genes were used as a back-ground list To highlight biological processes relevant tothe investigated sample groups an Ingenuity Pathwaysanalysis was performed using IPA software version 7(Ingenuity Systems Redwood City CA) For core analy-sis only genes with absolute expression value 17-foldchange (donor versus explanted heart-derived hCSCs)were considered The resulting annotations (networksbiofunctions canonical pathways) were ranked depend-ing on the number of significantly expressed genes andwere ordered by the score representing the probability offinding the observed molecules enriched in a given an-notation by chance score log(P value) The graphi-cal representation of the biofunctions was obtained byuse of the IPA Knowledge Base according to full-textarticles and information databases available online (httpwwwingenuitycomproductspathways_analysishtml lastaccessed June 2009) The spotted genes were deter-mined to be up-regulated or down-regulated from themicroarray low-level analysis
Statistical Analysis
Data are reported as means SD Differences weredetermined by two-tailed unpaired Studentrsquos t-test Linearregressions were calculated by the least squaresmethod Comparisons between slopes were made by theanalysis of covariance34 Microarray analysis was per-formed as described above
Results
Human Myocardium
Over a period of 5 years atrial tissue was obtained from63 donor hearts at the time of transplantation and from121 explanted failing hearts Atrial samples were col-lected in anatomical areas lacking apparent macro-scopic alterations By enzymatic digestion and primaryexplant technique24 a pool of small cardiac cells wasobtained in 166 of 184 samples tested which corre-sponds to a 90 rate of success Cell isolation was notfeasible in samples processed 48 hours after collectionIn successful isolations the cells were expanded andstored at passage 2 to passage 3 For the present studythe properties of hCSCs were analyzed in a subset of 41consecutive samples acquired from 18 donor and 23explanted hearts (see Supplemental Table S1 at httpajpamjpatholorg) These hearts were collected duringthe period from 2007 to 2009 at the University Hospital ofUdine In these cases cell isolation was performed within
48 hours Cell isolation and expansion was similar and
no difficulty was encountered at any stage of the cultureprotocol
Men were more numerous than women in the studysamples and ischemic cardiomyopathy predominatedThese variables may have introduced a partially biasedsampling however this reflects the sex differences andetiology of the disease in patients undergoing transplan-tation Donor subjects were younger than patients under-going transplantation who were diagnosed with ischemichypertrophic and idiopathic dilated cardiomyopathy or val-vular defects and acromegaly All patients undergoingtransplantation were class IV in the New York Heart Asso-ciation functional classification (NYHA IV) The duration ofthe underlying disease and the time from the onset ofheart failure to transplantation varied from 1 to 30 yearsand from 1 to 12 years respectively (see SupplementalTable S1 at httpajpamjpatholorg) With the exception ofone patient affected by hypertrophic cardiomyopathythe echocardiographic and hemodynamic parameterswere similar across subjects documenting severe ven-tricular dysfunction (see Supplemental Table S2 at httpajpamjpatholorg)
Human Cardiac Stem Cells
Structures with the characteristics of stem cell nicheswere identified in tissue sections of human atrial myocar-dium (Figure 1A) in which stem cells are preferentiallylocated16 Importantly c-Kit-positive hCSCs expressedOCT34 NANOG KLF4 and SOX2 which are implicatedin the self-renewing and pluripotent state of stem cells35
These properties were identified in c-Kit-positive hCSCsdistributed in atria from both donor and explanted hearts(Figures 1 BndashD and 2E) strengthening support for thenotion that hCSCs retain a highly immature embryonic-like cell phenotype
hCSCs were negative for CD45 as well as for thetranscription factor GATA4 and the myocyte cytoplasmicprotein -SA Quantitatively the number of c-Kit-positivelineage-negative hCSCs was 32-fold higher in atria ofdonor hearts than of explanted hearts (Figure 1E) Thisdifference was at variance with previous findings in theventricular myocardium of control and failing humanhearts in which cardiac decompensation was coupledwith an increase in hCSC number8 Thus regional differ-ences in hCSC number appear to be present in the failinghuman heart
In the atria of donor hearts there was 1 hCSC per 2000myocytes a frequency that is approximately fivefoldhigher than in the normal human left ventricle8 In severefailure the number of hCSCs in the atria was approxi-mately threefold lower than in the left ventricle8 The for-mer difference may reflect the site of storage of hCSCs inthe atria16 with the latter difference reflecting growthactivation of ventricular hCSCs in proximity of areas ofinjury Scarring and acute damage are distributed exclu-sively to the failing ventricle8
To validate the specificity of the recorded signalsspectral analysis was performed1720 The emissionspectra for c-Kit OCT34 NANOG and -SA were
clearly distinct from the emission spectra for tissue auto-
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

CT34
352 Cesselli et alAJP July 2011 Vol 179 No 1
brary Roche) LightCycler 480 Basic software (Roche)used the second-derivative maximum method to identifythe crossing point (Cp) Glioma cell line A172 was usedas positive control
Immunofluorescence and Confocal Microscopy
The expression of pluripotency genes and cardiac cyto-plasmic proteins was determined by immunolabeling (Ta-ble 1) Image acquisition was performed with a LeicaTCS-SP2 confocal laser microscope using a 63 oilimmersion objective (numerical aperture 140) or a 40oil immersion objective (numerical aperture 125)
Migration Assay
Migration analysis was performed using a Cultrex 24-wellcell migration assay (Trevigen Gaithersburg MD) Ap-proximately 200000 cells were seeded per well Growthmedium containing 10 fetal bovine serum was used aschemoattractant
Tumorigenicity of hCSCs
The tumorigenic potential of hCSCs was established byin vitro and in vivo protocols In vitro the soft agar assaywas used 1 105 hCSCs were seeded in a 05 softagar solution placed on top of a 1 hard agar Thenumber of colonies grown within the semisolid mediumwas measured 28 days later For the in vivo analysis 1 106 hCSCs were injected subcutaneously in NODScid mice (HARLAN Italy SRL San Pietro al NatisoneItaly) SKOV-3 ovarian cancer cells were used as pos-itive control (Sigma-Aldrich St Louis MO) Animalswere kept in pathogen-free conditions and tumorgrowth was evaluated monthly for a period of 6 monthsor until SKOV-3 ovarian cancer cells generated a tumor1 cm in diameter
Measurements of Ca2 Oscillations in hCSCs
hCSCs were loaded with 10 molL Fluo-3 AM dye (In-vitrogen) and were placed on the stage of a two-photonmicroscope a BX51WI Olympus microscope (OlympusTokyo Japan) coupled with a Bio-Rad Radiance 2100MP
Table 2 PCR Primers and Product Length
Gene Forward primer
KIT 5=-CGTGGAAAAGAGAAAACAGTCA-3=POU5F1dagger 5=-GAGGATCACCCTGGGATATAC-3=NANOG 5=-AATACCTCAGCCTCCAGCAGATG-3=SOX2 5=-GTTACCTCTTCCTCCCACTCCA-3=KLF4 5=-GGGAGAAGACACTGCGTCA-3=GAPDH 5=-CGACCACTTTGTCAAGCTCA-3=HPRT 5=-CGTGATTAGTGATGATGAACCAG-3=
The protein product for the KIT gene is known also as c-kitdaggerThe protein product for POU5F1 is known also as OCT3 OCT4 or O
system (Bio-Rad Laboratories Hercules CA) Cells were
bathed with Tyrodersquos solution containing (in mmolL)NaCl 140 KCl 54 MgCl2 1 HEPES 5 glucose 55 andCaCl2 20 (pH 74 adjusted with NaOH) The Fluo-3 wasexcited at 900 to 960 nm with a Tsunami mode-lockedTisapphire femtosecond laser (Spectra-Physics New-port Corporation Irvine CA) and the emission signal wascollected at 535 nm Series of images were acquired at10-second intervals for a period of 33 minutes Changesof intracellular Ca2 in individual hCSCs were deter-mined by measuring the fluorescent signal of Fluo-3 Ineach cell the oscillations in fluorescence with time weregraphically visualized using ImageJ (NIH Bethesda MD)and Microsoft Office Excel 2003 software These traceswere used to assess the number amplitude and durationof Ca2 oscillations in hCSCs Fluo-3 signals were ex-pressed as normalized fluorescence (FF0) The durationof Ca2 oscillations was measured from the onset of therising phase to the return to baseline21 Caffeine (10mmolL Sigma-Aldrich) was used to assess ryanodinereceptor function2223
Telomere Repeat Amplification Protocol Assay
The detection of telomerase activity was performed usinga TRAPeze kit (ChemiconndashMillipore Temecula CA) forthe telomere repeat amplification protocol (TRAP) assaywith a radiolabeling detection method Samples werelabeled with 32P Radiographic films were digitized with afilm scanner (SnapScan 1236 Agfa-Gevaert MortselBelgium) and a G3 Macintosh computer (Apple Cuper-tino CA) Telomerase activity was measured on digitalimages by ImageJ software version 142q (NIH) as unitsof total product generated (TPG) calculated as TPG [(x x0)c 100][(r r0)cR] where x is the signal of theregion of the gel lane corresponding to the TRAP productladder bands from non-heat-treated samples x0 is thesignal of the region of the gel lane corresponding to theTRAP product ladder bands from heat-treated samples ris the signal of the region of the gel lane corresponding tothe TRAP product ladder bands from TSR8 quantitationcontrol r0 is the signal of the region of the gel lanecorresponding to the TRAP product ladder bands from1 CHAPS lysis buffer-only control c is the signal fromthe internal standard (S-IC) in non-heat-treated samplesand cR is the signal from the internal standard (S-IC) inTSR8 quantitation control Each unit of TPG corresponds
Reverse primerAmplicon size
(bp)
=-CACCGTGATGCCAGCTATTA-3= 70=-CGATACTGGTTCGCTTTCTCTTT-3= 295=-CTGCGTCACACCATTGCTATTCT-3= 149=-CACCCCTCCCATTTCCCTCGTT-3= 260=-GGAAGCACTGGGGGAAGT-3= 88=-AGGGGTCTACATGGCAACTG-3= 228=-CGAGCAAGACGTTCAGTCCT-3= 129
5555555
to the number of TS primers in a quantity of 1 103
Human Cardiac Stem Cell Senescence 353AJP July 2011 Vol 179 No 1
amol or 600 molecules extended with at least four telo-meric repeats by telomerase enzyme present in the ex-tract
Telomere Length in hCSCs
Telomere length was measured by flow FISH (a combi-nation of flow cytometry and fluorescence in situ hybrid-ization)24 Suspensions consisting of a 11 mixture ofhCSCs and control cells were hybridized in the presenceor absence of fluorescein-conjugated peptide nucleicacid (PNA) telomere probe (DakoCytomation GlostrupDenmark) The average telomeric fluorescence per ge-nome in the two cell classes was calculated as followsrelative telomere length RTL [(mean FL1 sample cellswith probe mean FL1 sample cells without probe) DNA index of control cells 100][(mean FL1 controlcells with probe mean FL1 control cells withoutprobe) DNA index of sample cells]
Telomere Dysfunction-Induced Foci
Telomere dysfunction-induced foci which were definedby the colocalization of 53BP1 with telomeres were an-alyzed using a Leica DMI 6000B microscope connectedto a Leica DFC350FX camera hCSCs were considered tobe TIF-positive when at least 50 of 53BP1 spots colo-calized with telomere hybridization signals The numberof TIFs per hCSC was also measured Sampling con-sisted of more than 100 hCSCs in each case (see Sup-plemental Table S3 at httpajpamjpatholorg)
Microarray Analysis
RNA Extraction cDNA Labeling and MicroarrayHybridization
hCSCs obtained from age- and sex-matched donorand explanted hearts (n 3 each) were used for theanalysis of the transcriptional profile Total RNA wasextracted with TRIzol reagent (Invitrogen) and sub-jected to DNase I treatment After RNA quality controlmicroarray cDNA targets were prepared according tothe two-step indirect fluorescent labeling procedure25
starting from 10 g total RNA First-strand cDNA syn-thesis reaction was performed with unmodified randomprimers in the presence of aminoallyl-dUTP The result-ing primary amino groups were then coupled to theN-hydroxysuccinimide ester of the fluorescent dye Foreach age- and sex- matched pair one sample waslabeled with Cy5 and the other with Cy3
LNCIB 28K cDNA microarray slides (supporting28000 cDNA clones hereafter called features) wereused in the large-scale transcript expression profilingstudies these were prepared by spotting onto AmershamType 7 Star reflective microarray slides the purified PCRproducts of 19741 LNCIB human cDNA clones26 and8160 cDNA clones selected from the human sequence-verified IMAGE clone collection (Research GeneticsInvitrogen)27 This clone set consisted of approximately
13620 unique human genes Purified cDNA inserts were
resuspended in 50 dimethyl sulfoxide (Sigma-Aldrich)and printed on the slides using a SDDC-2 ChipWriter Promicroarrayer (Virtek Vision International Waterloo ONCanada) GAPDH cDNA and -actin cDNA were used aspositive array controls and the corresponding purifiedPCR products were printed at different concentrations onthe slides Plant cDNAs and printing buffer 50 dimethylsulfoxide were used as negative controls
Competitive hybridizations were performed under cov-erslips at 63degC overnight in the presence of 3 standardsaline citrate 02 SDS 100 ngL human Cot-1 DNAand 330 ngL yeast tRNA using a semiautomated hy-bridization station (Advalytix ArrayBooster BeckmanCoulter Biomedical Munich Germany) After a washingslides were scanned at a resolution of 10 m per pixelusing an Axon GenePix 4000B microarray scanner (AxonInstrumentsndashMolecular Devices Sunnyvale CA) Imagesfor the Cy3 and Cy5 channel were acquired in a 16-bittagged-image file format (TIFF) and were analyzed usingGenePix 40 analysis software (Axon Instruments) Im-ages were manually flagged to identify artifacts and badspots
Low-level analysis of microarray data were performedin the R (version 28)Bioconductor (version 23) environ-ment using the limma microarray package (available athttpwwwR-projectorg last accessed January 2009)28
Expression data were loaded from the GenePix imageanalysis output files Features displaying in at least onechannel median fluorescence intensity lower than back-ground plus two standard deviations were flagged asbad spots Fluorescent intensity of the red and greenchannels was background-subtracted29 and the corre-sponding expression ratio (Cy5Cy3) was log-trans-formed (base 2) Within each array normalization wasaccomplished by the print-tip LOWESS method indepen-dently from each cDNA collection Normalized ratios fromreplicate expression measurements within the same ar-ray were averaged to produce a final expression matrix(FEM) to be used in the subsequent analyses The FEMwas composed of 21069 features and three age- andsex-matched paired samples
Unsupervised hierarchical cluster analysis (average-linkage clustering)30 was applied to the filtered expres-sion matrix Flagged features features not present in allsamples and features with fold changes equal to one inat least one sample were filtered out By this approach7868 features were identified For both genes and sam-ples uncorrelated distance was used as dissimilaritymeasure Cluster results were visualized using JavaTreeView31
Differential Gene Expression
The differential gene expression between hCSCs ob-tained from donor and explanted hearts was establishedby functions and methods implemented in the RBiocon-ductor environment A fixed-effects linear model was fit-ted for each individual feature to estimate expressiondifferences An empirical Bayes approach was applied tomoderate standard errors of M values32 For each ana-
lyzed feature moderated t-statistics log-odds ratios of
354 Cesselli et alAJP July 2011 Vol 179 No 1
differential expression (B-statistics) and raw and ad-justed P values (false discovery rate control by the Ben-jamini and Hochberg method33) were obtained
Functional Annotation Analysis
Differentially expressed gene lists were subjected tofunctional analysis using the DAVIDease Web-basedtool version 2008 (NIH-NIAID Bethesda MD) LNCIB28K unique associated genes were used as a back-ground list To highlight biological processes relevant tothe investigated sample groups an Ingenuity Pathwaysanalysis was performed using IPA software version 7(Ingenuity Systems Redwood City CA) For core analy-sis only genes with absolute expression value 17-foldchange (donor versus explanted heart-derived hCSCs)were considered The resulting annotations (networksbiofunctions canonical pathways) were ranked depend-ing on the number of significantly expressed genes andwere ordered by the score representing the probability offinding the observed molecules enriched in a given an-notation by chance score log(P value) The graphi-cal representation of the biofunctions was obtained byuse of the IPA Knowledge Base according to full-textarticles and information databases available online (httpwwwingenuitycomproductspathways_analysishtml lastaccessed June 2009) The spotted genes were deter-mined to be up-regulated or down-regulated from themicroarray low-level analysis
Statistical Analysis
Data are reported as means SD Differences weredetermined by two-tailed unpaired Studentrsquos t-test Linearregressions were calculated by the least squaresmethod Comparisons between slopes were made by theanalysis of covariance34 Microarray analysis was per-formed as described above
Results
Human Myocardium
Over a period of 5 years atrial tissue was obtained from63 donor hearts at the time of transplantation and from121 explanted failing hearts Atrial samples were col-lected in anatomical areas lacking apparent macro-scopic alterations By enzymatic digestion and primaryexplant technique24 a pool of small cardiac cells wasobtained in 166 of 184 samples tested which corre-sponds to a 90 rate of success Cell isolation was notfeasible in samples processed 48 hours after collectionIn successful isolations the cells were expanded andstored at passage 2 to passage 3 For the present studythe properties of hCSCs were analyzed in a subset of 41consecutive samples acquired from 18 donor and 23explanted hearts (see Supplemental Table S1 at httpajpamjpatholorg) These hearts were collected duringthe period from 2007 to 2009 at the University Hospital ofUdine In these cases cell isolation was performed within
48 hours Cell isolation and expansion was similar and
no difficulty was encountered at any stage of the cultureprotocol
Men were more numerous than women in the studysamples and ischemic cardiomyopathy predominatedThese variables may have introduced a partially biasedsampling however this reflects the sex differences andetiology of the disease in patients undergoing transplan-tation Donor subjects were younger than patients under-going transplantation who were diagnosed with ischemichypertrophic and idiopathic dilated cardiomyopathy or val-vular defects and acromegaly All patients undergoingtransplantation were class IV in the New York Heart Asso-ciation functional classification (NYHA IV) The duration ofthe underlying disease and the time from the onset ofheart failure to transplantation varied from 1 to 30 yearsand from 1 to 12 years respectively (see SupplementalTable S1 at httpajpamjpatholorg) With the exception ofone patient affected by hypertrophic cardiomyopathythe echocardiographic and hemodynamic parameterswere similar across subjects documenting severe ven-tricular dysfunction (see Supplemental Table S2 at httpajpamjpatholorg)
Human Cardiac Stem Cells
Structures with the characteristics of stem cell nicheswere identified in tissue sections of human atrial myocar-dium (Figure 1A) in which stem cells are preferentiallylocated16 Importantly c-Kit-positive hCSCs expressedOCT34 NANOG KLF4 and SOX2 which are implicatedin the self-renewing and pluripotent state of stem cells35
These properties were identified in c-Kit-positive hCSCsdistributed in atria from both donor and explanted hearts(Figures 1 BndashD and 2E) strengthening support for thenotion that hCSCs retain a highly immature embryonic-like cell phenotype
hCSCs were negative for CD45 as well as for thetranscription factor GATA4 and the myocyte cytoplasmicprotein -SA Quantitatively the number of c-Kit-positivelineage-negative hCSCs was 32-fold higher in atria ofdonor hearts than of explanted hearts (Figure 1E) Thisdifference was at variance with previous findings in theventricular myocardium of control and failing humanhearts in which cardiac decompensation was coupledwith an increase in hCSC number8 Thus regional differ-ences in hCSC number appear to be present in the failinghuman heart
In the atria of donor hearts there was 1 hCSC per 2000myocytes a frequency that is approximately fivefoldhigher than in the normal human left ventricle8 In severefailure the number of hCSCs in the atria was approxi-mately threefold lower than in the left ventricle8 The for-mer difference may reflect the site of storage of hCSCs inthe atria16 with the latter difference reflecting growthactivation of ventricular hCSCs in proximity of areas ofinjury Scarring and acute damage are distributed exclu-sively to the failing ventricle8
To validate the specificity of the recorded signalsspectral analysis was performed1720 The emissionspectra for c-Kit OCT34 NANOG and -SA were
clearly distinct from the emission spectra for tissue auto-
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

Human Cardiac Stem Cell Senescence 353AJP July 2011 Vol 179 No 1
amol or 600 molecules extended with at least four telo-meric repeats by telomerase enzyme present in the ex-tract
Telomere Length in hCSCs
Telomere length was measured by flow FISH (a combi-nation of flow cytometry and fluorescence in situ hybrid-ization)24 Suspensions consisting of a 11 mixture ofhCSCs and control cells were hybridized in the presenceor absence of fluorescein-conjugated peptide nucleicacid (PNA) telomere probe (DakoCytomation GlostrupDenmark) The average telomeric fluorescence per ge-nome in the two cell classes was calculated as followsrelative telomere length RTL [(mean FL1 sample cellswith probe mean FL1 sample cells without probe) DNA index of control cells 100][(mean FL1 controlcells with probe mean FL1 control cells withoutprobe) DNA index of sample cells]
Telomere Dysfunction-Induced Foci
Telomere dysfunction-induced foci which were definedby the colocalization of 53BP1 with telomeres were an-alyzed using a Leica DMI 6000B microscope connectedto a Leica DFC350FX camera hCSCs were considered tobe TIF-positive when at least 50 of 53BP1 spots colo-calized with telomere hybridization signals The numberof TIFs per hCSC was also measured Sampling con-sisted of more than 100 hCSCs in each case (see Sup-plemental Table S3 at httpajpamjpatholorg)
Microarray Analysis
RNA Extraction cDNA Labeling and MicroarrayHybridization
hCSCs obtained from age- and sex-matched donorand explanted hearts (n 3 each) were used for theanalysis of the transcriptional profile Total RNA wasextracted with TRIzol reagent (Invitrogen) and sub-jected to DNase I treatment After RNA quality controlmicroarray cDNA targets were prepared according tothe two-step indirect fluorescent labeling procedure25
starting from 10 g total RNA First-strand cDNA syn-thesis reaction was performed with unmodified randomprimers in the presence of aminoallyl-dUTP The result-ing primary amino groups were then coupled to theN-hydroxysuccinimide ester of the fluorescent dye Foreach age- and sex- matched pair one sample waslabeled with Cy5 and the other with Cy3
LNCIB 28K cDNA microarray slides (supporting28000 cDNA clones hereafter called features) wereused in the large-scale transcript expression profilingstudies these were prepared by spotting onto AmershamType 7 Star reflective microarray slides the purified PCRproducts of 19741 LNCIB human cDNA clones26 and8160 cDNA clones selected from the human sequence-verified IMAGE clone collection (Research GeneticsInvitrogen)27 This clone set consisted of approximately
13620 unique human genes Purified cDNA inserts were
resuspended in 50 dimethyl sulfoxide (Sigma-Aldrich)and printed on the slides using a SDDC-2 ChipWriter Promicroarrayer (Virtek Vision International Waterloo ONCanada) GAPDH cDNA and -actin cDNA were used aspositive array controls and the corresponding purifiedPCR products were printed at different concentrations onthe slides Plant cDNAs and printing buffer 50 dimethylsulfoxide were used as negative controls
Competitive hybridizations were performed under cov-erslips at 63degC overnight in the presence of 3 standardsaline citrate 02 SDS 100 ngL human Cot-1 DNAand 330 ngL yeast tRNA using a semiautomated hy-bridization station (Advalytix ArrayBooster BeckmanCoulter Biomedical Munich Germany) After a washingslides were scanned at a resolution of 10 m per pixelusing an Axon GenePix 4000B microarray scanner (AxonInstrumentsndashMolecular Devices Sunnyvale CA) Imagesfor the Cy3 and Cy5 channel were acquired in a 16-bittagged-image file format (TIFF) and were analyzed usingGenePix 40 analysis software (Axon Instruments) Im-ages were manually flagged to identify artifacts and badspots
Low-level analysis of microarray data were performedin the R (version 28)Bioconductor (version 23) environ-ment using the limma microarray package (available athttpwwwR-projectorg last accessed January 2009)28
Expression data were loaded from the GenePix imageanalysis output files Features displaying in at least onechannel median fluorescence intensity lower than back-ground plus two standard deviations were flagged asbad spots Fluorescent intensity of the red and greenchannels was background-subtracted29 and the corre-sponding expression ratio (Cy5Cy3) was log-trans-formed (base 2) Within each array normalization wasaccomplished by the print-tip LOWESS method indepen-dently from each cDNA collection Normalized ratios fromreplicate expression measurements within the same ar-ray were averaged to produce a final expression matrix(FEM) to be used in the subsequent analyses The FEMwas composed of 21069 features and three age- andsex-matched paired samples
Unsupervised hierarchical cluster analysis (average-linkage clustering)30 was applied to the filtered expres-sion matrix Flagged features features not present in allsamples and features with fold changes equal to one inat least one sample were filtered out By this approach7868 features were identified For both genes and sam-ples uncorrelated distance was used as dissimilaritymeasure Cluster results were visualized using JavaTreeView31
Differential Gene Expression
The differential gene expression between hCSCs ob-tained from donor and explanted hearts was establishedby functions and methods implemented in the RBiocon-ductor environment A fixed-effects linear model was fit-ted for each individual feature to estimate expressiondifferences An empirical Bayes approach was applied tomoderate standard errors of M values32 For each ana-
lyzed feature moderated t-statistics log-odds ratios of
354 Cesselli et alAJP July 2011 Vol 179 No 1
differential expression (B-statistics) and raw and ad-justed P values (false discovery rate control by the Ben-jamini and Hochberg method33) were obtained
Functional Annotation Analysis
Differentially expressed gene lists were subjected tofunctional analysis using the DAVIDease Web-basedtool version 2008 (NIH-NIAID Bethesda MD) LNCIB28K unique associated genes were used as a back-ground list To highlight biological processes relevant tothe investigated sample groups an Ingenuity Pathwaysanalysis was performed using IPA software version 7(Ingenuity Systems Redwood City CA) For core analy-sis only genes with absolute expression value 17-foldchange (donor versus explanted heart-derived hCSCs)were considered The resulting annotations (networksbiofunctions canonical pathways) were ranked depend-ing on the number of significantly expressed genes andwere ordered by the score representing the probability offinding the observed molecules enriched in a given an-notation by chance score log(P value) The graphi-cal representation of the biofunctions was obtained byuse of the IPA Knowledge Base according to full-textarticles and information databases available online (httpwwwingenuitycomproductspathways_analysishtml lastaccessed June 2009) The spotted genes were deter-mined to be up-regulated or down-regulated from themicroarray low-level analysis
Statistical Analysis
Data are reported as means SD Differences weredetermined by two-tailed unpaired Studentrsquos t-test Linearregressions were calculated by the least squaresmethod Comparisons between slopes were made by theanalysis of covariance34 Microarray analysis was per-formed as described above
Results
Human Myocardium
Over a period of 5 years atrial tissue was obtained from63 donor hearts at the time of transplantation and from121 explanted failing hearts Atrial samples were col-lected in anatomical areas lacking apparent macro-scopic alterations By enzymatic digestion and primaryexplant technique24 a pool of small cardiac cells wasobtained in 166 of 184 samples tested which corre-sponds to a 90 rate of success Cell isolation was notfeasible in samples processed 48 hours after collectionIn successful isolations the cells were expanded andstored at passage 2 to passage 3 For the present studythe properties of hCSCs were analyzed in a subset of 41consecutive samples acquired from 18 donor and 23explanted hearts (see Supplemental Table S1 at httpajpamjpatholorg) These hearts were collected duringthe period from 2007 to 2009 at the University Hospital ofUdine In these cases cell isolation was performed within
48 hours Cell isolation and expansion was similar and
no difficulty was encountered at any stage of the cultureprotocol
Men were more numerous than women in the studysamples and ischemic cardiomyopathy predominatedThese variables may have introduced a partially biasedsampling however this reflects the sex differences andetiology of the disease in patients undergoing transplan-tation Donor subjects were younger than patients under-going transplantation who were diagnosed with ischemichypertrophic and idiopathic dilated cardiomyopathy or val-vular defects and acromegaly All patients undergoingtransplantation were class IV in the New York Heart Asso-ciation functional classification (NYHA IV) The duration ofthe underlying disease and the time from the onset ofheart failure to transplantation varied from 1 to 30 yearsand from 1 to 12 years respectively (see SupplementalTable S1 at httpajpamjpatholorg) With the exception ofone patient affected by hypertrophic cardiomyopathythe echocardiographic and hemodynamic parameterswere similar across subjects documenting severe ven-tricular dysfunction (see Supplemental Table S2 at httpajpamjpatholorg)
Human Cardiac Stem Cells
Structures with the characteristics of stem cell nicheswere identified in tissue sections of human atrial myocar-dium (Figure 1A) in which stem cells are preferentiallylocated16 Importantly c-Kit-positive hCSCs expressedOCT34 NANOG KLF4 and SOX2 which are implicatedin the self-renewing and pluripotent state of stem cells35
These properties were identified in c-Kit-positive hCSCsdistributed in atria from both donor and explanted hearts(Figures 1 BndashD and 2E) strengthening support for thenotion that hCSCs retain a highly immature embryonic-like cell phenotype
hCSCs were negative for CD45 as well as for thetranscription factor GATA4 and the myocyte cytoplasmicprotein -SA Quantitatively the number of c-Kit-positivelineage-negative hCSCs was 32-fold higher in atria ofdonor hearts than of explanted hearts (Figure 1E) Thisdifference was at variance with previous findings in theventricular myocardium of control and failing humanhearts in which cardiac decompensation was coupledwith an increase in hCSC number8 Thus regional differ-ences in hCSC number appear to be present in the failinghuman heart
In the atria of donor hearts there was 1 hCSC per 2000myocytes a frequency that is approximately fivefoldhigher than in the normal human left ventricle8 In severefailure the number of hCSCs in the atria was approxi-mately threefold lower than in the left ventricle8 The for-mer difference may reflect the site of storage of hCSCs inthe atria16 with the latter difference reflecting growthactivation of ventricular hCSCs in proximity of areas ofinjury Scarring and acute damage are distributed exclu-sively to the failing ventricle8
To validate the specificity of the recorded signalsspectral analysis was performed1720 The emissionspectra for c-Kit OCT34 NANOG and -SA were
clearly distinct from the emission spectra for tissue auto-
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

354 Cesselli et alAJP July 2011 Vol 179 No 1
differential expression (B-statistics) and raw and ad-justed P values (false discovery rate control by the Ben-jamini and Hochberg method33) were obtained
Functional Annotation Analysis
Differentially expressed gene lists were subjected tofunctional analysis using the DAVIDease Web-basedtool version 2008 (NIH-NIAID Bethesda MD) LNCIB28K unique associated genes were used as a back-ground list To highlight biological processes relevant tothe investigated sample groups an Ingenuity Pathwaysanalysis was performed using IPA software version 7(Ingenuity Systems Redwood City CA) For core analy-sis only genes with absolute expression value 17-foldchange (donor versus explanted heart-derived hCSCs)were considered The resulting annotations (networksbiofunctions canonical pathways) were ranked depend-ing on the number of significantly expressed genes andwere ordered by the score representing the probability offinding the observed molecules enriched in a given an-notation by chance score log(P value) The graphi-cal representation of the biofunctions was obtained byuse of the IPA Knowledge Base according to full-textarticles and information databases available online (httpwwwingenuitycomproductspathways_analysishtml lastaccessed June 2009) The spotted genes were deter-mined to be up-regulated or down-regulated from themicroarray low-level analysis
Statistical Analysis
Data are reported as means SD Differences weredetermined by two-tailed unpaired Studentrsquos t-test Linearregressions were calculated by the least squaresmethod Comparisons between slopes were made by theanalysis of covariance34 Microarray analysis was per-formed as described above
Results
Human Myocardium
Over a period of 5 years atrial tissue was obtained from63 donor hearts at the time of transplantation and from121 explanted failing hearts Atrial samples were col-lected in anatomical areas lacking apparent macro-scopic alterations By enzymatic digestion and primaryexplant technique24 a pool of small cardiac cells wasobtained in 166 of 184 samples tested which corre-sponds to a 90 rate of success Cell isolation was notfeasible in samples processed 48 hours after collectionIn successful isolations the cells were expanded andstored at passage 2 to passage 3 For the present studythe properties of hCSCs were analyzed in a subset of 41consecutive samples acquired from 18 donor and 23explanted hearts (see Supplemental Table S1 at httpajpamjpatholorg) These hearts were collected duringthe period from 2007 to 2009 at the University Hospital ofUdine In these cases cell isolation was performed within
48 hours Cell isolation and expansion was similar and
no difficulty was encountered at any stage of the cultureprotocol
Men were more numerous than women in the studysamples and ischemic cardiomyopathy predominatedThese variables may have introduced a partially biasedsampling however this reflects the sex differences andetiology of the disease in patients undergoing transplan-tation Donor subjects were younger than patients under-going transplantation who were diagnosed with ischemichypertrophic and idiopathic dilated cardiomyopathy or val-vular defects and acromegaly All patients undergoingtransplantation were class IV in the New York Heart Asso-ciation functional classification (NYHA IV) The duration ofthe underlying disease and the time from the onset ofheart failure to transplantation varied from 1 to 30 yearsand from 1 to 12 years respectively (see SupplementalTable S1 at httpajpamjpatholorg) With the exception ofone patient affected by hypertrophic cardiomyopathythe echocardiographic and hemodynamic parameterswere similar across subjects documenting severe ven-tricular dysfunction (see Supplemental Table S2 at httpajpamjpatholorg)
Human Cardiac Stem Cells
Structures with the characteristics of stem cell nicheswere identified in tissue sections of human atrial myocar-dium (Figure 1A) in which stem cells are preferentiallylocated16 Importantly c-Kit-positive hCSCs expressedOCT34 NANOG KLF4 and SOX2 which are implicatedin the self-renewing and pluripotent state of stem cells35
These properties were identified in c-Kit-positive hCSCsdistributed in atria from both donor and explanted hearts(Figures 1 BndashD and 2E) strengthening support for thenotion that hCSCs retain a highly immature embryonic-like cell phenotype
hCSCs were negative for CD45 as well as for thetranscription factor GATA4 and the myocyte cytoplasmicprotein -SA Quantitatively the number of c-Kit-positivelineage-negative hCSCs was 32-fold higher in atria ofdonor hearts than of explanted hearts (Figure 1E) Thisdifference was at variance with previous findings in theventricular myocardium of control and failing humanhearts in which cardiac decompensation was coupledwith an increase in hCSC number8 Thus regional differ-ences in hCSC number appear to be present in the failinghuman heart
In the atria of donor hearts there was 1 hCSC per 2000myocytes a frequency that is approximately fivefoldhigher than in the normal human left ventricle8 In severefailure the number of hCSCs in the atria was approxi-mately threefold lower than in the left ventricle8 The for-mer difference may reflect the site of storage of hCSCs inthe atria16 with the latter difference reflecting growthactivation of ventricular hCSCs in proximity of areas ofinjury Scarring and acute damage are distributed exclu-sively to the failing ventricle8
To validate the specificity of the recorded signalsspectral analysis was performed1720 The emissionspectra for c-Kit OCT34 NANOG and -SA were
clearly distinct from the emission spectra for tissue auto-
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
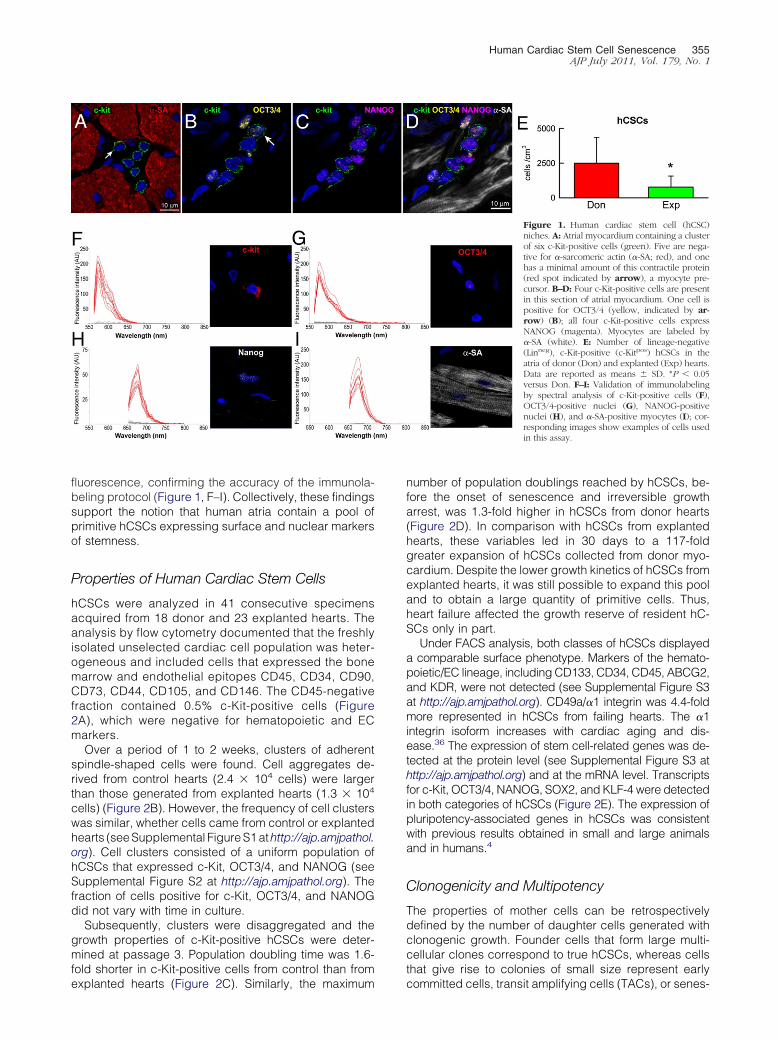
Human Cardiac Stem Cell Senescence 355AJP July 2011 Vol 179 No 1
fluorescence confirming the accuracy of the immunola-beling protocol (Figure 1 FndashI) Collectively these findingssupport the notion that human atria contain a pool ofprimitive hCSCs expressing surface and nuclear markersof stemness
Properties of Human Cardiac Stem Cells
hCSCs were analyzed in 41 consecutive specimensacquired from 18 donor and 23 explanted hearts Theanalysis by flow cytometry documented that the freshlyisolated unselected cardiac cell population was heter-ogeneous and included cells that expressed the bonemarrow and endothelial epitopes CD45 CD34 CD90CD73 CD44 CD105 and CD146 The CD45-negativefraction contained 05 c-Kit-positive cells (Figure2A) which were negative for hematopoietic and ECmarkers
Over a period of 1 to 2 weeks clusters of adherentspindle-shaped cells were found Cell aggregates de-rived from control hearts (24 104 cells) were largerthan those generated from explanted hearts (13 104
cells) (Figure 2B) However the frequency of cell clusterswas similar whether cells came from control or explantedhearts (see Supplemental Figure S1 at httpajpamjpatholorg) Cell clusters consisted of a uniform population ofhCSCs that expressed c-Kit OCT34 and NANOG (seeSupplemental Figure S2 at httpajpamjpatholorg) Thefraction of cells positive for c-Kit OCT34 and NANOGdid not vary with time in culture
Subsequently clusters were disaggregated and thegrowth properties of c-Kit-positive hCSCs were deter-mined at passage 3 Population doubling time was 16-fold shorter in c-Kit-positive cells from control than from
explanted hearts (Figure 2C) Similarly the maximum
number of population doublings reached by hCSCs be-fore the onset of senescence and irreversible growtharrest was 13-fold higher in hCSCs from donor hearts(Figure 2D) In comparison with hCSCs from explantedhearts these variables led in 30 days to a 117-foldgreater expansion of hCSCs collected from donor myo-cardium Despite the lower growth kinetics of hCSCs fromexplanted hearts it was still possible to expand this pooland to obtain a large quantity of primitive cells Thusheart failure affected the growth reserve of resident hC-SCs only in part
Under FACS analysis both classes of hCSCs displayeda comparable surface phenotype Markers of the hemato-poieticEC lineage including CD133 CD34 CD45 ABCG2and KDR were not detected (see Supplemental Figure S3at httpajpamjpatholorg) CD49a1 integrin was 44-foldmore represented in hCSCs from failing hearts The 1integrin isoform increases with cardiac aging and dis-ease36 The expression of stem cell-related genes was de-tected at the protein level (see Supplemental Figure S3 athttpajpamjpatholorg) and at the mRNA level Transcriptsfor c-Kit OCT34 NANOG SOX2 and KLF-4 were detectedin both categories of hCSCs (Figure 2E) The expression ofpluripotency-associated genes in hCSCs was consistentwith previous results obtained in small and large animalsand in humans4
Clonogenicity and Multipotency
The properties of mother cells can be retrospectivelydefined by the number of daughter cells generated withclonogenic growth Founder cells that form large multi-cellular clones correspond to true hCSCs whereas cellsthat give rise to colonies of small size represent early
Figure 1 Human cardiac stem cell (hCSC)niches A Atrial myocardium containing a clusterof six c-Kit-positive cells (green) Five are nega-tive for -sarcomeric actin (-SA red) and onehas a minimal amount of this contractile protein(red spot indicated by arrow) a myocyte pre-cursor BndashD Four c-Kit-positive cells are presentin this section of atrial myocardium One cell ispositive for OCT34 (yellow indicated by ar-row) (B) all four c-Kit-positive cells expressNANOG (magenta) Myocytes are labeled by-SA (white) E Number of lineage-negative(Linneg) c-Kit-positive (c-Kitpos) hCSCs in theatria of donor (Don) and explanted (Exp) heartsData are reported as means SD P 005versus Don FndashI Validation of immunolabelingby spectral analysis of c-Kit-positive cells (F)OCT34-positive nuclei (G) NANOG-positivenuclei (H) and -SA-positive myocytes (I) cor-responding images show examples of cells usedin this assay
committed cells transit amplifying cells (TACs) or senes-
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
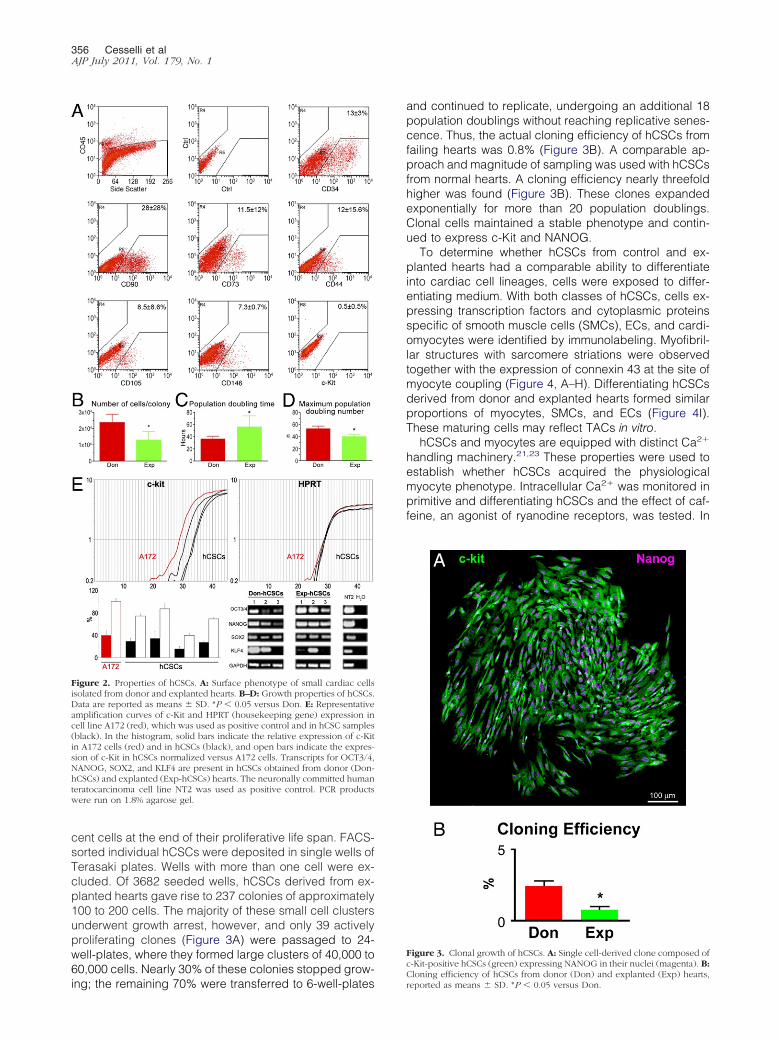
356 Cesselli et alAJP July 2011 Vol 179 No 1
cent cells at the end of their proliferative life span FACS-sorted individual hCSCs were deposited in single wells ofTerasaki plates Wells with more than one cell were ex-cluded Of 3682 seeded wells hCSCs derived from ex-planted hearts gave rise to 237 colonies of approximately100 to 200 cells The majority of these small cell clustersunderwent growth arrest however and only 39 activelyproliferating clones (Figure 3A) were passaged to 24-well-plates where they formed large clusters of 40000 to60000 cells Nearly 30 of these colonies stopped grow-
Figure 2 Properties of hCSCs A Surface phenotype of small cardiac cellsisolated from donor and explanted hearts BndashD Growth properties of hCSCsData are reported as means SD P 005 versus Don E Representativeamplification curves of c-Kit and HPRT (housekeeping gene) expression incell line A172 (red) which was used as positive control and in hCSC samples(black) In the histogram solid bars indicate the relative expression of c-Kitin A172 cells (red) and in hCSCs (black) and open bars indicate the expres-sion of c-Kit in hCSCs normalized versus A172 cells Transcripts for OCT34NANOG SOX2 and KLF4 are present in hCSCs obtained from donor (Don-hCSCs) and explanted (Exp-hCSCs) hearts The neuronally committed humanteratocarcinoma cell line NT2 was used as positive control PCR productswere run on 18 agarose gel
ing the remaining 70 were transferred to 6-well-plates
and continued to replicate undergoing an additional 18population doublings without reaching replicative senes-cence Thus the actual cloning efficiency of hCSCs fromfailing hearts was 08 (Figure 3B) A comparable ap-proach and magnitude of sampling was used with hCSCsfrom normal hearts A cloning efficiency nearly threefoldhigher was found (Figure 3B) These clones expandedexponentially for more than 20 population doublingsClonal cells maintained a stable phenotype and contin-ued to express c-Kit and NANOG
To determine whether hCSCs from control and ex-planted hearts had a comparable ability to differentiateinto cardiac cell lineages cells were exposed to differ-entiating medium With both classes of hCSCs cells ex-pressing transcription factors and cytoplasmic proteinsspecific of smooth muscle cells (SMCs) ECs and cardi-omyocytes were identified by immunolabeling Myofibril-lar structures with sarcomere striations were observedtogether with the expression of connexin 43 at the site ofmyocyte coupling (Figure 4 AndashH) Differentiating hCSCsderived from donor and explanted hearts formed similarproportions of myocytes SMCs and ECs (Figure 4I)These maturing cells may reflect TACs in vitro
hCSCs and myocytes are equipped with distinct Ca2
handling machinery2123 These properties were used toestablish whether hCSCs acquired the physiologicalmyocyte phenotype Intracellular Ca2 was monitored inprimitive and differentiating hCSCs and the effect of caf-feine an agonist of ryanodine receptors was tested In
Figure 3 Clonal growth of hCSCs A Single cell-derived clone composed ofc-Kit-positive hCSCs (green) expressing NANOG in their nuclei (magenta) B
Cloning efficiency of hCSCs from donor (Don) and explanted (Exp) heartsreported as means SD P 005 versus Don
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
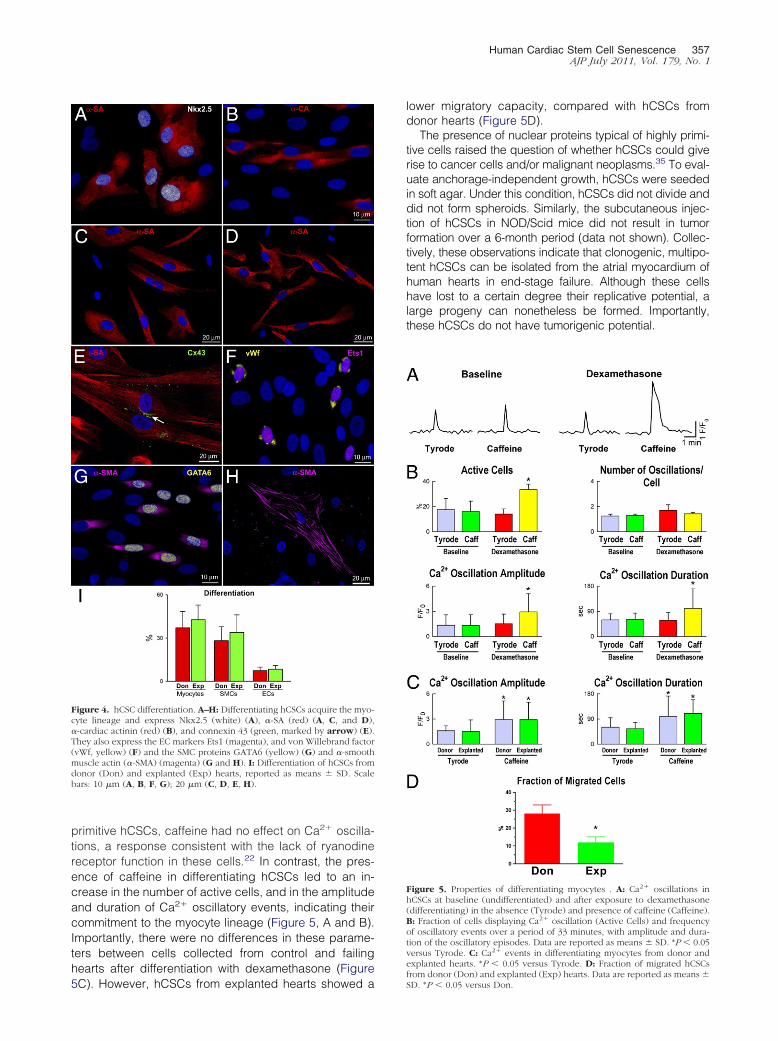
Human Cardiac Stem Cell Senescence 357AJP July 2011 Vol 179 No 1
primitive hCSCs caffeine had no effect on Ca2 oscilla-tions a response consistent with the lack of ryanodinereceptor function in these cells22 In contrast the pres-ence of caffeine in differentiating hCSCs led to an in-crease in the number of active cells and in the amplitudeand duration of Ca2 oscillatory events indicating theircommitment to the myocyte lineage (Figure 5 A and B)Importantly there were no differences in these parame-ters between cells collected from control and failinghearts after differentiation with dexamethasone (Figure
Figure 4 hCSC differentiation AndashH Differentiating hCSCs acquire the myo-cyte lineage and express Nkx25 (white) (A) -SA (red) (A C and D)-cardiac actinin (red) (B) and connexin 43 (green marked by arrow) (E)They also express the EC markers Ets1 (magenta) and von Willebrand factor(vWf yellow) (F) and the SMC proteins GATA6 (yellow) (G) and -smoothmuscle actin (-SMA) (magenta) (G and H) I Differentiation of hCSCs fromdonor (Don) and explanted (Exp) hearts reported as means SD Scalebars 10 m (A B F G) 20 m (C D E H)
5C) However hCSCs from explanted hearts showed a
lower migratory capacity compared with hCSCs fromdonor hearts (Figure 5D)
The presence of nuclear proteins typical of highly primi-tive cells raised the question of whether hCSCs could giverise to cancer cells andor malignant neoplasms35 To eval-uate anchorage-independent growth hCSCs were seededin soft agar Under this condition hCSCs did not divide anddid not form spheroids Similarly the subcutaneous injec-tion of hCSCs in NODScid mice did not result in tumorformation over a 6-month period (data not shown) Collec-tively these observations indicate that clonogenic multipo-tent hCSCs can be isolated from the atrial myocardium ofhuman hearts in end-stage failure Although these cellshave lost to a certain degree their replicative potential alarge progeny can nonetheless be formed Importantlythese hCSCs do not have tumorigenic potential
Figure 5 Properties of differentiating myocytes A Ca2 oscillations inhCSCs at baseline (undifferentiated) and after exposure to dexamethasone(differentiating) in the absence (Tyrode) and presence of caffeine (Caffeine)B Fraction of cells displaying Ca2 oscillation (Active Cells) and frequencyof oscillatory events over a period of 33 minutes with amplitude and dura-tion of the oscillatory episodes Data are reported as means SD P 005versus Tyrode C Ca2 events in differentiating myocytes from donor andexplanted hearts P 005 versus Tyrode D Fraction of migrated hCSCs
from donor (Don) and explanted (Exp) hearts Data are reported as means SD P 005 versus Don
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
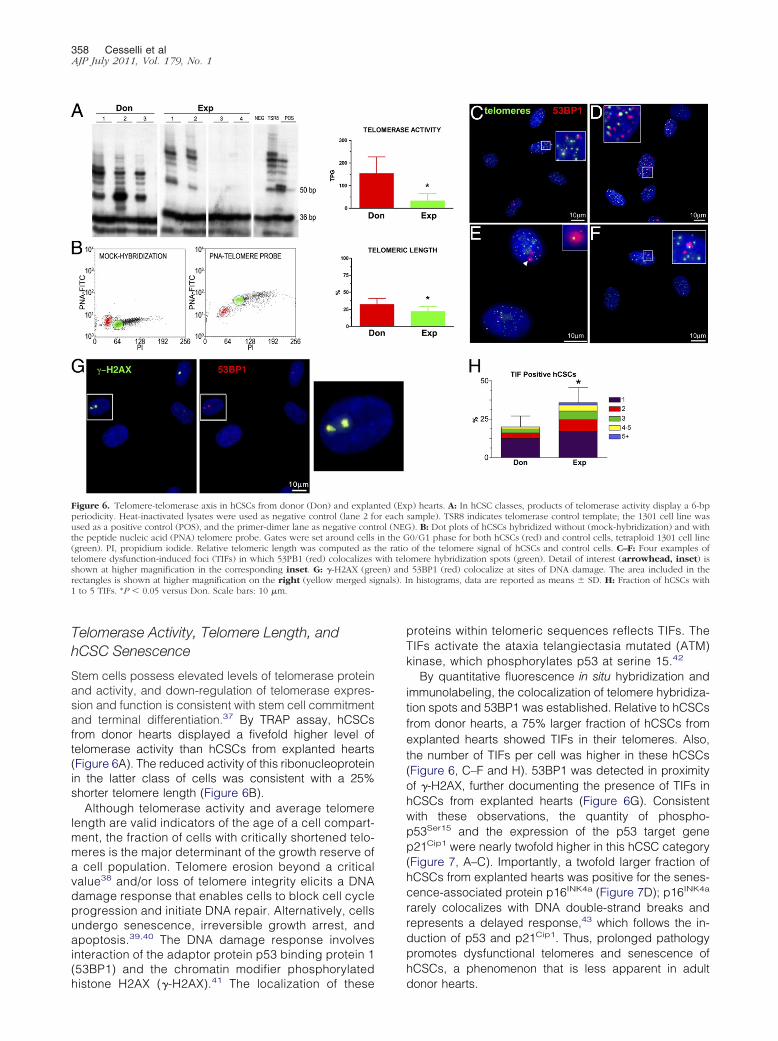
gnals) I
358 Cesselli et alAJP July 2011 Vol 179 No 1
Telomerase Activity Telomere Length andhCSC Senescence
Stem cells possess elevated levels of telomerase proteinand activity and down-regulation of telomerase expres-sion and function is consistent with stem cell commitmentand terminal differentiation37 By TRAP assay hCSCsfrom donor hearts displayed a fivefold higher level oftelomerase activity than hCSCs from explanted hearts(Figure 6A) The reduced activity of this ribonucleoproteinin the latter class of cells was consistent with a 25shorter telomere length (Figure 6B)
Although telomerase activity and average telomerelength are valid indicators of the age of a cell compart-ment the fraction of cells with critically shortened telo-meres is the major determinant of the growth reserve ofa cell population Telomere erosion beyond a criticalvalue38 andor loss of telomere integrity elicits a DNAdamage response that enables cells to block cell cycleprogression and initiate DNA repair Alternatively cellsundergo senescence irreversible growth arrest andapoptosis3940 The DNA damage response involvesinteraction of the adaptor protein p53 binding protein 1(53BP1) and the chromatin modifier phosphorylated
Figure 6 Telomere-telomerase axis in hCSCs from donor (Don) and explanperiodicity Heat-inactivated lysates were used as negative control (lane 2 foused as a positive control (POS) and the primer-dimer lane as negative contthe peptide nucleic acid (PNA) telomere probe Gates were set around cells(green) PI propidium iodide Relative telomeric length was computed astelomere dysfunction-induced foci (TIFs) in which 53PB1 (red) colocalizesshown at higher magnification in the corresponding inset G -H2AX (grerectangles is shown at higher magnification on the right (yellow merged si1 to 5 TIFs P 005 versus Don Scale bars 10 m
histone H2AX (-H2AX)41 The localization of these
proteins within telomeric sequences reflects TIFs TheTIFs activate the ataxia telangiectasia mutated (ATM)kinase which phosphorylates p53 at serine 1542
By quantitative fluorescence in situ hybridization andimmunolabeling the colocalization of telomere hybridiza-tion spots and 53BP1 was established Relative to hCSCsfrom donor hearts a 75 larger fraction of hCSCs fromexplanted hearts showed TIFs in their telomeres Alsothe number of TIFs per cell was higher in these hCSCs(Figure 6 CndashF and H) 53BP1 was detected in proximityof -H2AX further documenting the presence of TIFs inhCSCs from explanted hearts (Figure 6G) Consistentwith these observations the quantity of phospho-p53Ser15 and the expression of the p53 target genep21Cip1 were nearly twofold higher in this hCSC category(Figure 7 AndashC) Importantly a twofold larger fraction ofhCSCs from explanted hearts was positive for the senes-cence-associated protein p16INK4a (Figure 7D) p16INK4a
rarely colocalizes with DNA double-strand breaks andrepresents a delayed response43 which follows the in-duction of p53 and p21Cip1 Thus prolonged pathologypromotes dysfunctional telomeres and senescence ofhCSCs a phenomenon that is less apparent in adult
p) hearts A In hCSC classes products of telomerase activity display a 6-bpsample) TSR8 indicates telomerase control template the 1301 cell line was) B Dot plots of hCSCs hybridized without (mock-hybridization) and with0G1 phase for both hCSCs (red) and control cells tetraploid 1301 cell lineof the telomere signal of hCSCs and control cells CndashF Four examples ofmere hybridization spots (green) Detail of interest (arrowhead inset) is53BP1 (red) colocalize at sites of DNA damage The area included in the
n histograms data are reported as means SD H Fraction of hCSCs with
ted (Exr eachrol (NEGin the G
the ratiowith teloen) and
donor hearts
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
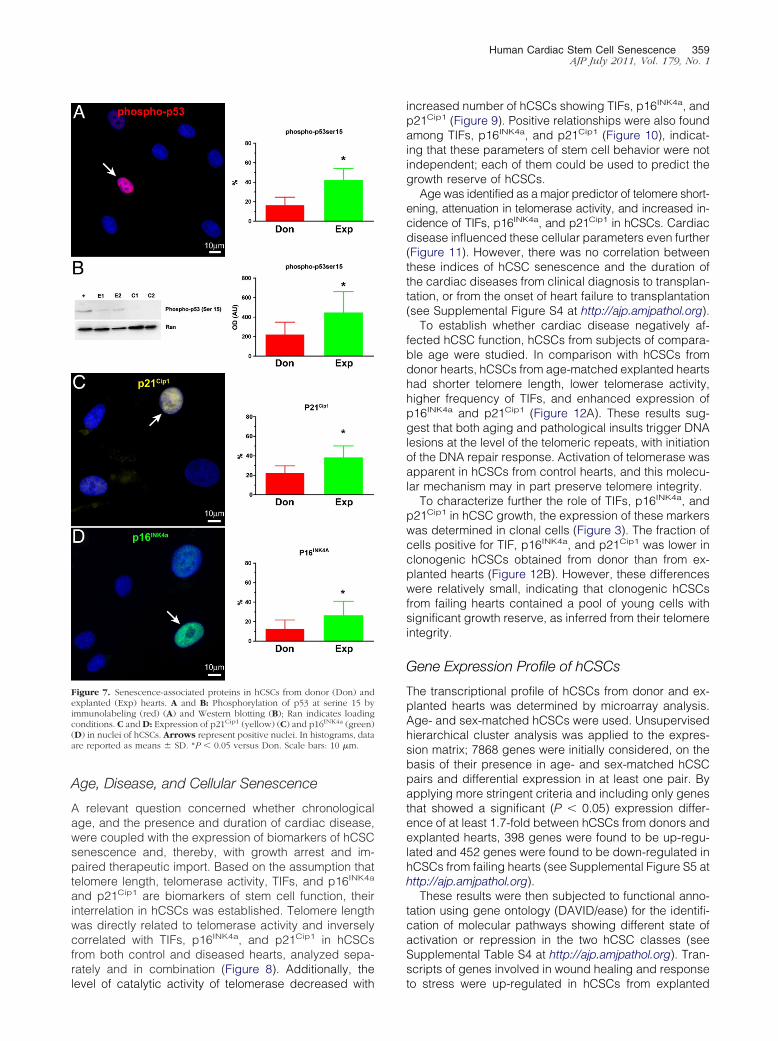
Human Cardiac Stem Cell Senescence 359AJP July 2011 Vol 179 No 1
Age Disease and Cellular Senescence
A relevant question concerned whether chronologicalage and the presence and duration of cardiac diseasewere coupled with the expression of biomarkers of hCSCsenescence and thereby with growth arrest and im-paired therapeutic import Based on the assumption thattelomere length telomerase activity TIFs and p16INK4a
and p21Cip1 are biomarkers of stem cell function theirinterrelation in hCSCs was established Telomere lengthwas directly related to telomerase activity and inverselycorrelated with TIFs p16INK4a and p21Cip1 in hCSCsfrom both control and diseased hearts analyzed sepa-rately and in combination (Figure 8) Additionally the
Figure 7 Senescence-associated proteins in hCSCs from donor (Don) andexplanted (Exp) hearts A and B Phosphorylation of p53 at serine 15 byimmunolabeling (red) (A) and Western blotting (B) Ran indicates loadingconditions C and D Expression of p21Cip1 (yellow) (C) and p16INK4a (green)(D) in nuclei of hCSCs Arrows represent positive nuclei In histograms dataare reported as means SD P 005 versus Don Scale bars 10 m
level of catalytic activity of telomerase decreased with
increased number of hCSCs showing TIFs p16INK4a andp21Cip1 (Figure 9) Positive relationships were also foundamong TIFs p16INK4a and p21Cip1 (Figure 10) indicat-ing that these parameters of stem cell behavior were notindependent each of them could be used to predict thegrowth reserve of hCSCs
Age was identified as a major predictor of telomere short-ening attenuation in telomerase activity and increased in-cidence of TIFs p16INK4a and p21Cip1 in hCSCs Cardiacdisease influenced these cellular parameters even further(Figure 11) However there was no correlation betweenthese indices of hCSC senescence and the duration ofthe cardiac diseases from clinical diagnosis to transplan-tation or from the onset of heart failure to transplantation(see Supplemental Figure S4 at httpajpamjpatholorg)
To establish whether cardiac disease negatively af-fected hCSC function hCSCs from subjects of compara-ble age were studied In comparison with hCSCs fromdonor hearts hCSCs from age-matched explanted heartshad shorter telomere length lower telomerase activityhigher frequency of TIFs and enhanced expression ofp16INK4a and p21Cip1 (Figure 12A) These results sug-gest that both aging and pathological insults trigger DNAlesions at the level of the telomeric repeats with initiationof the DNA repair response Activation of telomerase wasapparent in hCSCs from control hearts and this molecu-lar mechanism may in part preserve telomere integrity
To characterize further the role of TIFs p16INK4a andp21Cip1 in hCSC growth the expression of these markerswas determined in clonal cells (Figure 3) The fraction ofcells positive for TIF p16INK4a and p21Cip1 was lower inclonogenic hCSCs obtained from donor than from ex-planted hearts (Figure 12B) However these differenceswere relatively small indicating that clonogenic hCSCsfrom failing hearts contained a pool of young cells withsignificant growth reserve as inferred from their telomereintegrity
Gene Expression Profile of hCSCs
The transcriptional profile of hCSCs from donor and ex-planted hearts was determined by microarray analysisAge- and sex-matched hCSCs were used Unsupervisedhierarchical cluster analysis was applied to the expres-sion matrix 7868 genes were initially considered on thebasis of their presence in age- and sex-matched hCSCpairs and differential expression in at least one pair Byapplying more stringent criteria and including only genesthat showed a significant (P 005) expression differ-ence of at least 17-fold between hCSCs from donors andexplanted hearts 398 genes were found to be up-regu-lated and 452 genes were found to be down-regulated inhCSCs from failing hearts (see Supplemental Figure S5 athttpajpamjpatholorg)
These results were then subjected to functional anno-tation using gene ontology (DAVIDease) for the identifi-cation of molecular pathways showing different state ofactivation or repression in the two hCSC classes (seeSupplemental Table S4 at httpajpamjpatholorg) Tran-scripts of genes involved in wound healing and response
to stress were up-regulated in hCSCs from explanted
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
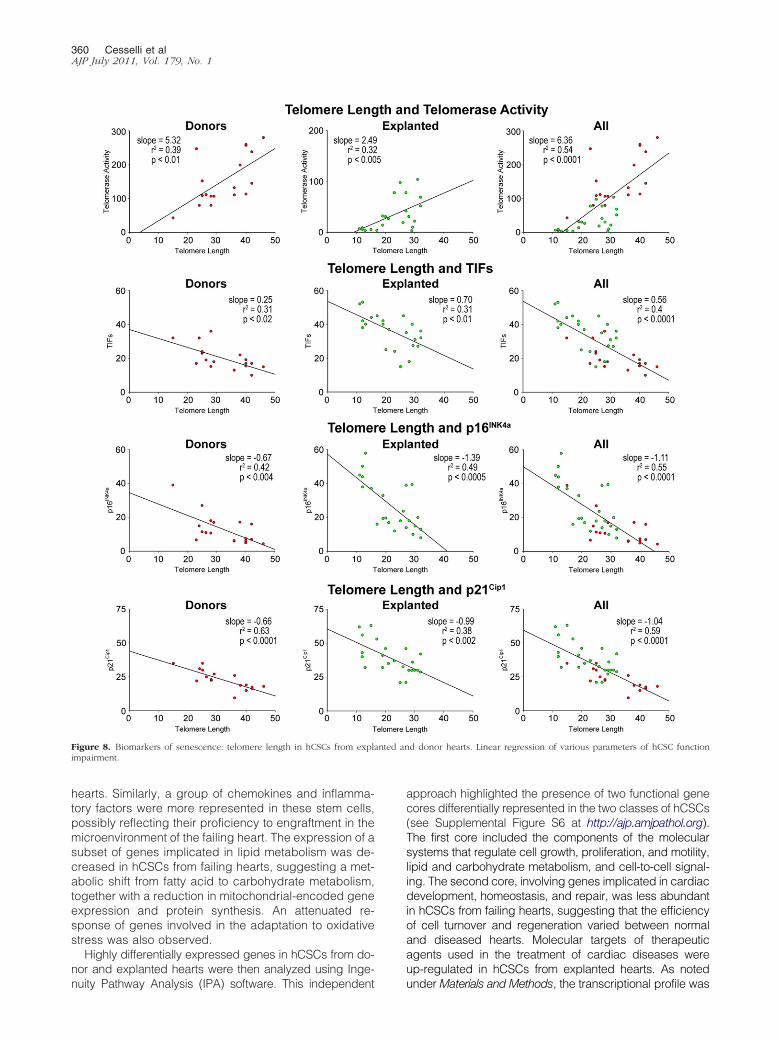
anted an
360 Cesselli et alAJP July 2011 Vol 179 No 1
hearts Similarly a group of chemokines and inflamma-tory factors were more represented in these stem cellspossibly reflecting their proficiency to engraftment in themicroenvironment of the failing heart The expression of asubset of genes implicated in lipid metabolism was de-creased in hCSCs from failing hearts suggesting a met-abolic shift from fatty acid to carbohydrate metabolismtogether with a reduction in mitochondrial-encoded geneexpression and protein synthesis An attenuated re-sponse of genes involved in the adaptation to oxidativestress was also observed
Highly differentially expressed genes in hCSCs from do-nor and explanted hearts were then analyzed using Inge-
Figure 8 Biomarkers of senescence telomere length in hCSCs from explimpairment
nuity Pathway Analysis (IPA) software This independent
approach highlighted the presence of two functional genecores differentially represented in the two classes of hCSCs(see Supplemental Figure S6 at httpajpamjpatholorg)The first core included the components of the molecularsystems that regulate cell growth proliferation and motilitylipid and carbohydrate metabolism and cell-to-cell signal-ing The second core involving genes implicated in cardiacdevelopment homeostasis and repair was less abundantin hCSCs from failing hearts suggesting that the efficiencyof cell turnover and regeneration varied between normaland diseased hearts Molecular targets of therapeuticagents used in the treatment of cardiac diseases wereup-regulated in hCSCs from explanted hearts As noted
d donor hearts Linear regression of various parameters of hCSC function
under Materials and Methods the transcriptional profile was
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
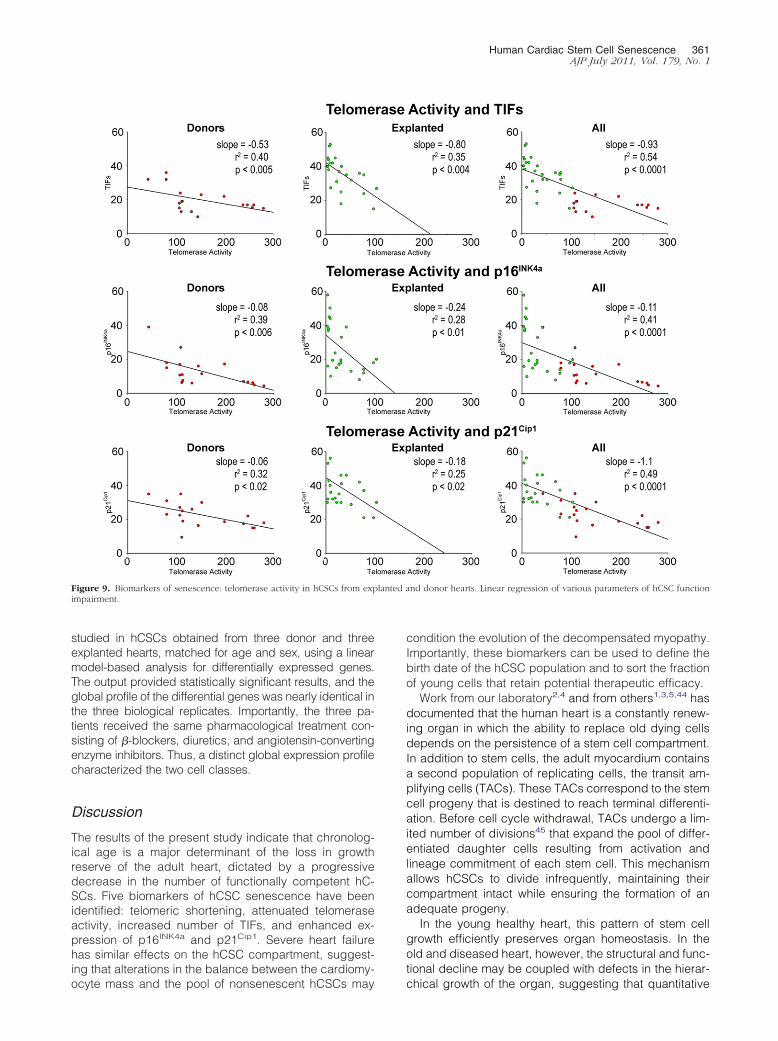
lanted a
Human Cardiac Stem Cell Senescence 361AJP July 2011 Vol 179 No 1
studied in hCSCs obtained from three donor and threeexplanted hearts matched for age and sex using a linearmodel-based analysis for differentially expressed genesThe output provided statistically significant results and theglobal profile of the differential genes was nearly identical inthe three biological replicates Importantly the three pa-tients received the same pharmacological treatment con-sisting of -blockers diuretics and angiotensin-convertingenzyme inhibitors Thus a distinct global expression profilecharacterized the two cell classes
Discussion
The results of the present study indicate that chronolog-ical age is a major determinant of the loss in growthreserve of the adult heart dictated by a progressivedecrease in the number of functionally competent hC-SCs Five biomarkers of hCSC senescence have beenidentified telomeric shortening attenuated telomeraseactivity increased number of TIFs and enhanced ex-pression of p16INK4a and p21Cip1 Severe heart failurehas similar effects on the hCSC compartment suggest-ing that alterations in the balance between the cardiomy-
Figure 9 Biomarkers of senescence telomerase activity in hCSCs from expimpairment
ocyte mass and the pool of nonsenescent hCSCs may
condition the evolution of the decompensated myopathyImportantly these biomarkers can be used to define thebirth date of the hCSC population and to sort the fractionof young cells that retain potential therapeutic efficacy
Work from our laboratory24 and from others13544 hasdocumented that the human heart is a constantly renew-ing organ in which the ability to replace old dying cellsdepends on the persistence of a stem cell compartmentIn addition to stem cells the adult myocardium containsa second population of replicating cells the transit am-plifying cells (TACs) These TACs correspond to the stemcell progeny that is destined to reach terminal differenti-ation Before cell cycle withdrawal TACs undergo a lim-ited number of divisions45 that expand the pool of differ-entiated daughter cells resulting from activation andlineage commitment of each stem cell This mechanismallows hCSCs to divide infrequently maintaining theircompartment intact while ensuring the formation of anadequate progeny
In the young healthy heart this pattern of stem cellgrowth efficiently preserves organ homeostasis In theold and diseased heart however the structural and func-tional decline may be coupled with defects in the hierar-
nd donor hearts Linear regression of various parameters of hCSC function
chical growth of the organ suggesting that quantitative
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
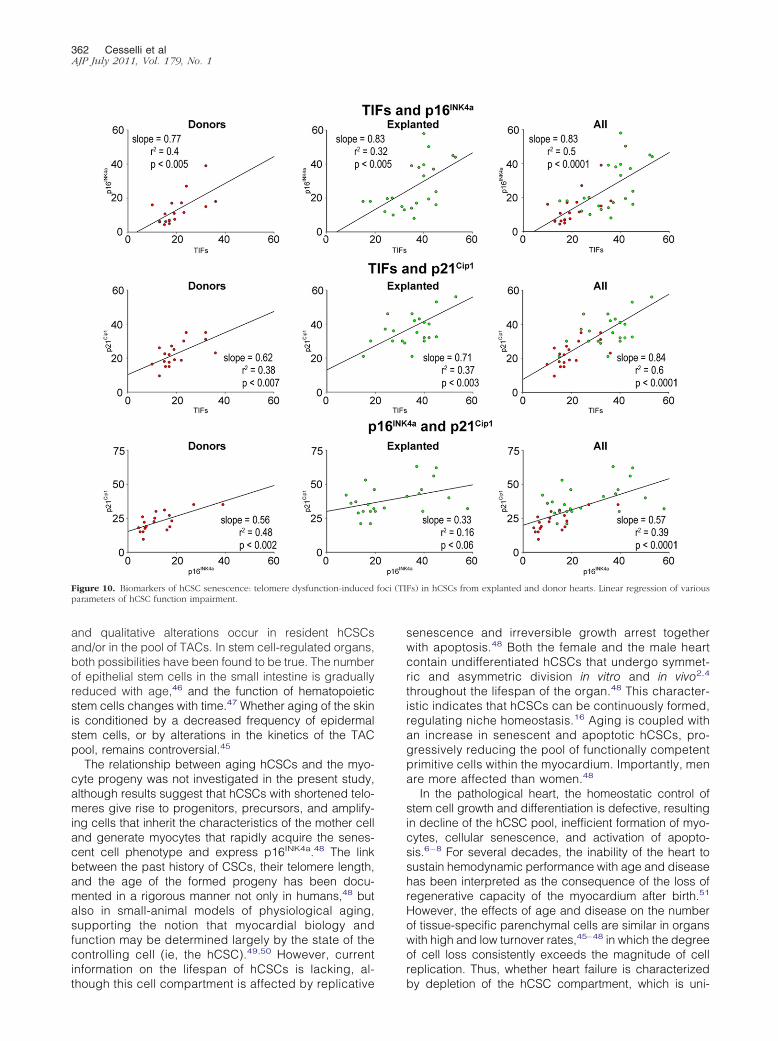
foci (TI
362 Cesselli et alAJP July 2011 Vol 179 No 1
and qualitative alterations occur in resident hCSCsandor in the pool of TACs In stem cell-regulated organsboth possibilities have been found to be true The numberof epithelial stem cells in the small intestine is graduallyreduced with age46 and the function of hematopoieticstem cells changes with time47 Whether aging of the skinis conditioned by a decreased frequency of epidermalstem cells or by alterations in the kinetics of the TACpool remains controversial45
The relationship between aging hCSCs and the myo-cyte progeny was not investigated in the present studyalthough results suggest that hCSCs with shortened telo-meres give rise to progenitors precursors and amplify-ing cells that inherit the characteristics of the mother celland generate myocytes that rapidly acquire the senes-cent cell phenotype and express p16INK4a48 The linkbetween the past history of CSCs their telomere lengthand the age of the formed progeny has been docu-mented in a rigorous manner not only in humans48 butalso in small-animal models of physiological agingsupporting the notion that myocardial biology andfunction may be determined largely by the state of thecontrolling cell (ie the hCSC)4950 However currentinformation on the lifespan of hCSCs is lacking al-
Figure 10 Biomarkers of hCSC senescence telomere dysfunction-inducedparameters of hCSC function impairment
though this cell compartment is affected by replicative
senescence and irreversible growth arrest togetherwith apoptosis48 Both the female and the male heartcontain undifferentiated hCSCs that undergo symmet-ric and asymmetric division in vitro and in vivo24
throughout the lifespan of the organ48 This character-istic indicates that hCSCs can be continuously formedregulating niche homeostasis16 Aging is coupled withan increase in senescent and apoptotic hCSCs pro-gressively reducing the pool of functionally competentprimitive cells within the myocardium Importantly menare more affected than women48
In the pathological heart the homeostatic control ofstem cell growth and differentiation is defective resultingin decline of the hCSC pool inefficient formation of myo-cytes cellular senescence and activation of apopto-sis6ndash8 For several decades the inability of the heart tosustain hemodynamic performance with age and diseasehas been interpreted as the consequence of the loss ofregenerative capacity of the myocardium after birth51
However the effects of age and disease on the numberof tissue-specific parenchymal cells are similar in organswith high and low turnover rates45ndash48 in which the degreeof cell loss consistently exceeds the magnitude of cellreplication Thus whether heart failure is characterized
Fs) in hCSCs from explanted and donor hearts Linear regression of various
by depletion of the hCSC compartment which is uni-
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
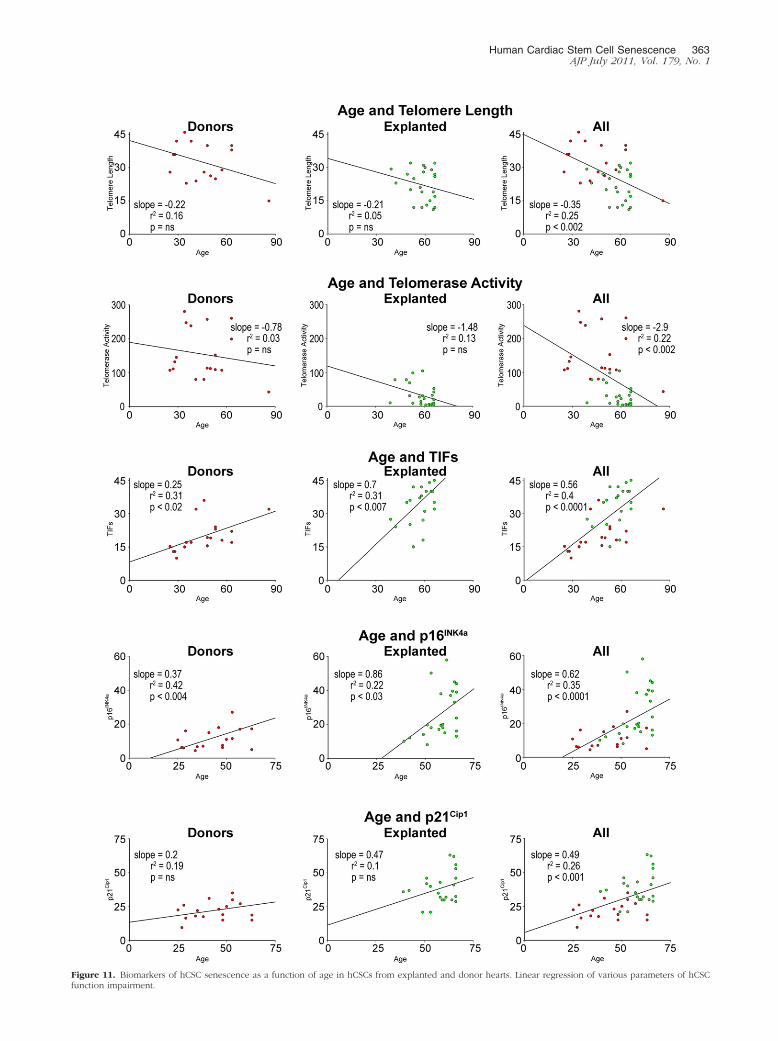
Human Cardiac Stem Cell Senescence 363AJP July 2011 Vol 179 No 1
Figure 11 Biomarkers of hCSC senescence as a function of age in hCSCs from explanted and donor hearts Linear regression of various parameters of hCSCfunction impairment
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
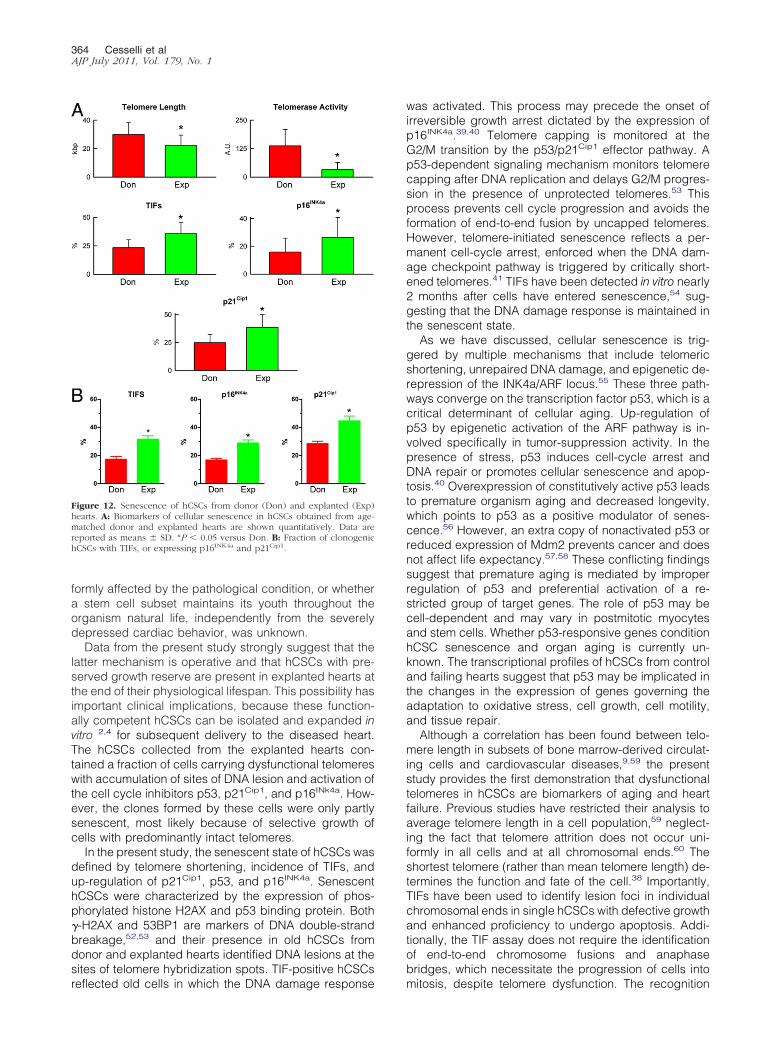
364 Cesselli et alAJP July 2011 Vol 179 No 1
formly affected by the pathological condition or whethera stem cell subset maintains its youth throughout theorganism natural life independently from the severelydepressed cardiac behavior was unknown
Data from the present study strongly suggest that thelatter mechanism is operative and that hCSCs with pre-served growth reserve are present in explanted hearts atthe end of their physiological lifespan This possibility hasimportant clinical implications because these function-ally competent hCSCs can be isolated and expanded invitro 24 for subsequent delivery to the diseased heartThe hCSCs collected from the explanted hearts con-tained a fraction of cells carrying dysfunctional telomereswith accumulation of sites of DNA lesion and activation ofthe cell cycle inhibitors p53 p21Cip1 and p16INk4a How-ever the clones formed by these cells were only partlysenescent most likely because of selective growth ofcells with predominantly intact telomeres
In the present study the senescent state of hCSCs wasdefined by telomere shortening incidence of TIFs andup-regulation of p21Cip1 p53 and p16INK4a SenescenthCSCs were characterized by the expression of phos-phorylated histone H2AX and p53 binding protein Both-H2AX and 53BP1 are markers of DNA double-strandbreakage5253 and their presence in old hCSCs fromdonor and explanted hearts identified DNA lesions at thesites of telomere hybridization spots TIF-positive hCSCs
Figure 12 Senescence of hCSCs from donor (Don) and explanted (Exp)hearts A Biomarkers of cellular senescence in hCSCs obtained from age-matched donor and explanted hearts are shown quantitatively Data arereported as means SD P 005 versus Don B Fraction of clonogenichCSCs with TIFs or expressing p16INK4a and p21Cip1
reflected old cells in which the DNA damage response
was activated This process may precede the onset ofirreversible growth arrest dictated by the expression ofp16INK4a3940 Telomere capping is monitored at theG2M transition by the p53p21Cip1 effector pathway Ap53-dependent signaling mechanism monitors telomerecapping after DNA replication and delays G2M progres-sion in the presence of unprotected telomeres53 Thisprocess prevents cell cycle progression and avoids theformation of end-to-end fusion by uncapped telomeresHowever telomere-initiated senescence reflects a per-manent cell-cycle arrest enforced when the DNA dam-age checkpoint pathway is triggered by critically short-ened telomeres41 TIFs have been detected in vitro nearly2 months after cells have entered senescence54 sug-gesting that the DNA damage response is maintained inthe senescent state
As we have discussed cellular senescence is trig-gered by multiple mechanisms that include telomericshortening unrepaired DNA damage and epigenetic de-repression of the INK4aARF locus55 These three path-ways converge on the transcription factor p53 which is acritical determinant of cellular aging Up-regulation ofp53 by epigenetic activation of the ARF pathway is in-volved specifically in tumor-suppression activity In thepresence of stress p53 induces cell-cycle arrest andDNA repair or promotes cellular senescence and apop-tosis40 Overexpression of constitutively active p53 leadsto premature organism aging and decreased longevitywhich points to p53 as a positive modulator of senes-cence56 However an extra copy of nonactivated p53 orreduced expression of Mdm2 prevents cancer and doesnot affect life expectancy5758 These conflicting findingssuggest that premature aging is mediated by improperregulation of p53 and preferential activation of a re-stricted group of target genes The role of p53 may becell-dependent and may vary in postmitotic myocytesand stem cells Whether p53-responsive genes conditionhCSC senescence and organ aging is currently un-known The transcriptional profiles of hCSCs from controland failing hearts suggest that p53 may be implicated inthe changes in the expression of genes governing theadaptation to oxidative stress cell growth cell motilityand tissue repair
Although a correlation has been found between telo-mere length in subsets of bone marrow-derived circulat-ing cells and cardiovascular diseases959 the presentstudy provides the first demonstration that dysfunctionaltelomeres in hCSCs are biomarkers of aging and heartfailure Previous studies have restricted their analysis toaverage telomere length in a cell population59 neglect-ing the fact that telomere attrition does not occur uni-formly in all cells and at all chromosomal ends60 Theshortest telomere (rather than mean telomere length) de-termines the function and fate of the cell38 ImportantlyTIFs have been used to identify lesion foci in individualchromosomal ends in single hCSCs with defective growthand enhanced proficiency to undergo apoptosis Addi-tionally the TIF assay does not require the identificationof end-to-end chromosome fusions and anaphasebridges which necessitate the progression of cells into
mitosis despite telomere dysfunction The recognition
Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

Human Cardiac Stem Cell Senescence 365AJP July 2011 Vol 179 No 1
that TIFs define a population of hCSCs that increases withchronological age and with the concomitant presence ofheart failure emphasizes the critical role that intrinsiccellular factors at the level of resident stem cells have inthe characterization of the human disease TheoreticallyTIF incidence may discriminate eugeric from pathogericprocesses (ie physiological aging as such versus phys-iological aging in the presence of cardiovascular dis-eases commonly found in the elderly population) How-ever the lack of overlap between the age of donor andexplanted hearts in the present study made this compar-ison problematic and actual distinction of physiologicaland pathological aging was not possible Donor heartsare generally obtained from young individuals and heartfailure affects mostly patients 65 years of age and older
References
1 Smith RR Barile L Cho HC Leppo MK Hare JM Messina E Giaco-mello A Abraham MR Marbaacuten E Regenerative potential of cardio-sphere-derived cells expanded from percutaneous endomyocardialbiopsy specimens Circulation 2007 115896ndash908
2 Bearzi C Rota M Hosoda T Tillmanns J Nascimbene A De AngelisA Yasuzawa-Amano S Trofimova I Siggins RW Lecapitaine NCascapera S Beltrami AP DrsquoAlessandro DA Zias E Quaini F Ur-banek K Michler RE Bolli R Kajstura J Leri A Anversa P Humancardiac stem cells Proc Natl Acad Sci USA 2007 10414068ndash14073
3 Beltrami AP Cesselli D Bergamin N Marcon P Rigo S Puppato EDrsquoAurizio F Verardo R Piazza S Pignatelli A Poz A Baccarani UDamiani D Fanin R Mariuzzi L Finato N Masolini P Burelli S BelluzziO Schneider C Beltrami CA Multipotent cells can be generated invitro from several adult human organs (heart liver and bone marrow)Blood 2007 1103438ndash3446
4 Bearzi C Leri A Lo Monaco F Rota M Gonzalez A Hosoda T PepeM Qanud K Ojaimi C Bardelli S DrsquoAmario D DrsquoAlessandro DAMichler RE Dimmeler S Zeiher AM Urbanek K Hintze TH KajsturaJ Anversa P Identification of a coronary vascular progenitor cell inthe human heart Proc Natl Acad Sci USA 2009 10615885ndash15890
5 Itzhaki-Alfia A Leor J Raanani E Sternik L Spiegelstein D Netser SHolbova R Pevsner-Fischer M Lavee J Barbash IM Patient charac-teristics and cell source determine the number of isolated humancardiac progenitor cells Circulation 2009 1202559ndash2566
6 Chimenti C Kajstura J Torella D Urbanek K Heleniak H Colussi CDi Meglio F Nadal-Ginard B Frustaci A Leri A Maseri A Anversa PSenescence and death of primitive cells and myocytes lead to pre-mature cardiac aging and heart failure Circ Res 2003 93604ndash613
7 Urbanek K Quaini F Tasca G Torella D Castaldo C Nadal-Ginard BLeri A Kajstura J Quaini E Anversa P Intense myocyte formationfrom cardiac stem cells in human cardiac hypertrophy Proc NatlAcad Sci USA 2003 10010440ndash10445
8 Urbanek K Torella D Sheikh F De Angelis A Nurzynska D SilvestriF Beltrami CA Bussani R Beltrami AP Quaini F Bolli R Leri AKajstura J Anversa P Myocardial regeneration by activation of mul-tipotent cardiac stem cells in ischemic heart failure Proc Natl AcadSci USA 2005 1028692ndash8697
9 Dimmeler S Leri A Aging and disease as modifiers of efficacy of celltherapy Circ Res 2008 1021319ndash1330
10 Sahin E Depinho RA Linking functional decline of telomeres mito-chondria and stem cells during ageing Nature 2010 464520ndash528
11 Blackburn EH The end of the (DNA) line Nat Struct Biol 20007847ndash850
12 de Lange T How telomeres solve the end-protection problem Sci-ence 2009 326948ndash952
13 Lansdorp PM Lessons from mice without telomerase J Cell Biol1997 139309ndash312
14 Xin ZT Beauchamp AD Calado RT Bradford JW Regal JA ShenoyA Liang Y Lansdorp PM Young NS Ly H Functional characteriza-
tion of natural telomerase mutations found in patients with hemato-logic disorders Blood 2007 109524ndash532
15 Alter BP Baerlocher GM Savage SA Chanock SJ Weksler BBWillner JP Peters JA Giri N Lansdorp PM Very short telomere lengthby flow fluorescence in situ hybridization identifies patients with dys-keratosis congenita Blood 2007 1101439ndash1447
16 Urbanek K Cesselli D Rota M Nascimbene A De Angelis A HosodaT Bearzi C Boni A Bolli R Kajstura J Anversa P Leri A Stem cellniches in the adult mouse heart Proc Natl Acad Sci USA 20061039226ndash9231
17 Hosoda T DrsquoAmario D Cabral-Da-Silva MC Zheng H Padin-IruegasME Ogorek B Ferreira-Martins J Yasuzawa-Amano S Amano KIde-Iwata N Cheng W Rota M Urbanek K Kajstura J Anversa P LeriA Clonality of mouse and human cardiomyogenesis in vivo Proc NatlAcad Sci USA 2009 10617169ndash17174
18 DrsquoAlessandro DA Kajstura J Hosoda T Gatti A Bello R Mosna FBardelli S Zheng H DrsquoAmario D Padin-Iruegas ME Carvalho ABRota M Zembala MO Stern D Rimoldi O Urbanek K Michler RE LeriA Anversa P Progenitor cells from the explanted heart generateimmunocompatible myocardium within the transplanted donor heartCirc Res 2009 1051128ndash1140
19 Boni A Urbanek K Nascimbene A Hosoda T Zheng H Delucchi FAmano K Gonzalez A Vitale S Ojaimi C Rizzi R Bolli R Yutzey KERota M Kajstura J Anversa P Leri A Notch1 regulates the fate ofcardiac progenitor cells Proc Natl Acad Sci USA 2008 10515529ndash15534
20 Kajstura J Urbanek K Perl S Hosoda T Zheng H Ogoacuterek BFerreira-Martins J Goichberg P Rondon-Clavo C Sanada FDrsquoAmario D Rota M Del Monte F Orlic D Tisdale J Leri AAnversa P Cardiomyogenesis in the adult human heart Circ Res2010 107305ndash315
21 Ferreira-Martins J Rondon-Clavo C Tugal D Korn JA Rizzi R Padin-Iruegas ME Ottolenghi S De Angelis A Urbanek K Ide-Iwata NDrsquoAmario D Hosoda T Leri A Kajstura J Anversa P Rota M Spon-taneous calcium oscillations regulate human cardiac progenitor cellgrowth Circ Res 2009 105764ndash774
22 Rota M Boni A Urbanek K Padin-Iruegas ME Kajstura TJ Fiore GKubo H Sonnenblick EH Musso E Houser SR Leri A Sussman MAAnversa P Nuclear targeting of Akt enhances ventricular function andmyocyte contractility Circ Res 2005 951332ndash1341
23 Rota M Hosoda T De Angelis A Arcarese ML Esposito G Rizzi RTillmanns J Tugal D Musso E Rimoldi O Bearzi C Urbanek KAnversa P Leri A Kajstura J The young mouse heart is composed ofmyocytes heterogeneous in age and function Circ Res 2007 101387ndash399
24 Baerlocher GM Vulto I de Jong G Lansdorp PM Flow cytometry andFISH to measure the average length of telomeres (flow FISH) NatProtoc 2006 12365ndash2376
25 Xiang CC Kozhich OA Chen M Inman JM Phan QN Chen YBrownstein MJ Amine-modified random primers to label probes forDNA microarrays Nat Biotechnol 2002 20738ndash742
26 Dalla E Mignone F Verardo R Marchionni L Marzinotto S LazarevicD Reid JF Marzio R Klaric E Licastro D Marcuzzi G Gambetta RPierotti MA Pesole G Schneider C Discovery of 342 putative newgenes from the analysis of 5=-end-sequenced full-length-enrichedcDNA human transcripts Genomics 2005 85739ndash751
27 Lennon G Auffray C Polymeropoulos M Soares MB The IMAGEConsortium an integrated molecular analysis of genomes and theirexpression Genomics 1996 33151ndash152
28 Smyth GK Limma linear models for microarray data Bioinformaticsand Computational Biology Solutions using R and Bioconductor Ed-ited by RC Gentleman VJ Carey W Huber RA Irizarry S Dudoit NewYork Springer 2005 pp 397ndash420
29 Edwards D Non-linear normalization and background correction inone-channel cDNA microarray studies Bioinformatics 2003 19825ndash 833
30 Eisen MB Spellman PT Brown PO Botstein D Cluster analysis anddisplay of genome-wide expression patterns Proc Natl Acad Sci USA1998 9514863ndash14868
31 Saldanha AJ Java Treeviewndashextensible visualization of microarraydata Bioinformatics 2004 203246ndash3248
32 Smyth GK Linear models and empirical Bayes methods for assess-ing differential expression in microarray experiments Stat Appl Genet
Mol Biol 2004 3Article3
366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579

366 Cesselli et alAJP July 2011 Vol 179 No 1
33 Benjamini Y Hochberg Y Controlling the false discovery rate apractical and powerful approach to multiple testing J R Stat SocSeries B Stat Methodol 1995 57289ndash300
34 Berenson ML Levine DM Rindskopf D Simple linear regression andcorrelation Applied Statistics Edited by ML Berenson DM Levine DRindskopf Englewood Cliffs Prentice Hall 1998 pp 362ndash418
35 Beltrami AP Cesselli D Beltrami CA Pluripotency rush Molecularcues for pluripotency genetic reprogramming of adult stem cellsand widely multipotent adult cells Pharmacol Ther 2009 12423ndash30
36 Burgess ML McCrea JC Hedrick HL Age-associated changes incardiac matrix and integrins Mech Ageing Dev 2001 1221739ndash1756
37 Hiyama E Hiyama K Telomere and telomerase in stem cells Br JCancer 2007 961020ndash1024
38 Hemann MT Strong MA Hao LY Greider CW The shortest telomerenot average telomere length is critical for cell viability and chromo-some stability Cell 2001 10767ndash77
39 Sharpless NE DePinho RA How stem cells age and why this makesus grow old Nat Rev Mol Cell Biol 2007 8703ndash713
40 Campisi J drsquoAdda di Fagagna F Cellular senescence when badthings happen to good cells Nat Rev Mol Cell Biol 2007 8729ndash740
41 drsquoAdda di Fagagna F Reaper PM Clay-Farrace L Fiegler H Carr PVon Zglinicki T Saretzki G Carter NP Jackson SP A DNA damagecheckpoint response in telomere-initiated senescence Nature 2003426194ndash198
42 Gire V Roux P Wynford-Thomas D Brondello JM Dulic V DNAdamage checkpoint kinase Chk2 triggers replicative senescenceEMBO J 2004 232554ndash2563
43 Jacobs JJ de Lange T p16INK4a as a second effector of the telo-mere damage pathway Cell Cycle 2005 41364ndash1368
44 Castaldo C Di Meglio F Nurzynska D Romano G Maiello C Ban-cone C Muumlller P Boumlhm M Cotrufo M Montagnani S CD117-positivecells in adult human heart are localized in the subepicardium andtheir activation is associated with laminin-1 and alpha6 integrin ex-pression Stem Cells 2008 261723ndash1731
45 Blanpain C Fuchs E Epidermal homeostasis a balancing act of stemcells in the skin Nat Rev Mol Cell Biol 2009 10207ndash217
46 Martin K Kirkwood TB Potten CS Age changes in stem cells ofmurine small intestinal crypts Exp Cell Res 1998 241316ndash323
47 Rossi DJ Bryder D Zahn JM Ahlenius H Sonu R Wagers AJWeissman IL Cell intrinsic alterations underlie hematopoietic stem
cell aging Proc Natl Acad Sci USA 2005 1029194ndash9199
48 Kajstura J Gurusamy N Ogoacuterek B Goichberg P Clavos-Rondon CHosoda T DrsquoAmario D Bardelli S Beltrami AP Cesselli D Bussani Rdel Monte F Quaini F Rota M Beltrami CA Buchholz B Leri AAnversa P Myocyte turnover in the aging human heart Circ Res2010 1071374ndash1386
49 Torella D Rota M Nurzynska D Musso E Monsen A Shiraishi I ZiasE Walsh K Rosenzweig A Sussman MA Urbanek K Nadal-GinardB Kajstura J Anversa P Leri A Cardiac stem cell and myocyteaging heart failure and insulin-like growth factor-1 overexpressionCirc Res 2004 94514ndash524
50 Gonzalez A Rota M Nurzynska D Misao Y Tillmanns J Ojaimi CPadin-Iruegas ME Muumlller P Esposito G Bearzi C Vitale S Dawn BSanganalmath SK Baker M Hintze TH Bolli R Urbanek K Hosoda TAnversa P Kajstura J Leri A Activation of cardiac progenitor cellsreverses the failing heart senescent phenotype and prolongs lifes-pan Circ Res 2008 102597ndash606
51 Murry CE Field LJ Menascheacute P Cell-based cardiac repair reflec-tions at the 10-year point Circulation 2005 1123174ndash3183
52 Herbig U Ferreira M Condel L Carey D Sedivy JM Cellular senes-cence in aging primates Science 2006 3111257
53 Thanasoula M Escandell JM Martinez P Badie S Muntildeoz P BlascoMA Tarsounas M p53 prevents entry into mitosis with uncappedtelomeres Curr Biol 2010 20521ndash526
54 Takai H Smogorzewska A de Lange T DNA damage foci at dys-functional telomeres Curr Biol 2003 131549ndash1556
55 Collado M Blasco MA Serrano M Cellular senescence in cancer andaging Cell 2007 130223ndash233
56 Maier B Gluba W Bernier B Turner T Mohammad K Guise TSutherland A Thorner M Scrable H Modulation of mammalian lifespan by the short isoform of p53 Genes Dev 2004 18306ndash319
57 Garciacutea-Cao I Garciacutea-Cao M Martiacuten-Caballero J Criado LM Klatt PFlores JM Weill JC Blasco MA Serrano M ldquoSuper p53rdquo mice exhibitenhanced DNA damage response are tumor resistant and age nor-mally EMBO J 2002 216225ndash6235
58 Mendrysa SM OrsquoLeary KA McElwee MK Michalowski J EisenmanRN Powell DA Perry ME Tumor suppression and normal aging inmice with constitutively high p53 activity Genes Dev 2006 2016ndash21
59 Brouilette SW Whittaker A Stevens SE van der Harst P Goodall AHSamani NJ Telomere length is shorter in healthy offspring of subjectswith coronary artery disease support for the telomere hypothesisHeart 2008 94422ndash425
60 Aubert G Lansdorp PM Telomeres and aging Physiol Rev 200888557ndash579
Copyright © 2022 FDOKUMEN













![[Are de novo acute heart failure and acutely worsened chronic heart failure two subgroups of the same syndrome?]](https://static.fdokumen.com/doc/165x107/63272f3a57d70b68cb099e92/are-de-novo-acute-heart-failure-and-acutely-worsened-chronic-heart-failure-two.jpg)






