
Fermentation processes: Historical development, components ...

Effect of moisture content in polyurethane foams as support
for solid-substrate fermentation of Lecanicillium lecanii
on the production profiles of chitinases
Marıa del Carmen Marin-Cervantes, Yoyi Matsumoto,Laura Ramırez-Coutino, Zaizy Rocha-Pino,
Gustavo Viniegra, Keiko Shirai *
Universidad Autonoma Metropolitana, Biotechnology Department, Laboratory of Biopolymers,
Avenue San Rafael Atlixco No. 186. Col. Vicentina, C.P. 09340, Mexico City, Mexico
Received 26 January 2007; received in revised form 31 August 2007; accepted 15 October 2007
www.elsevier.com/locate/procbio
Process Biochemistry 43 (2008) 24–32
Abstract
Finely minced (MPUF) and roughly cut (CPUF) polyurethane foam was used as inert support for the growth and chitinases production of
Lecanicillium lecanii by solid-substrate fermentation. L. lecanii growths on CPUF produced loose and disperse mycelia throughout the polymer
honeycomb, but MPUF resulted in dense aggregates at the ends of the polymer branches. Despite similar growth rates, m, and maximum biomass
concentration, Xmax, there were significant differences in the enzyme production. Highest enzyme titers (emax) without glucose supplementation
showed best results for>85% moisture content. emax, of exo-chitinase was 45-fold higher in CPUF than MPUF. Endochitinases, emax, were similar
for CPUF and MPUF. Catabolic repression of enzyme production depended on moisture level, being stronger for lower moisture contents for exo-
chitinases and milder or insignificant for endo-chitinases. Biomass yield coefficients of enzymes, Ye/x, were higher with MPUF than with CPUF for
endo-chitinases, but the reverse was found for exo-chitinases.
# 2007 Elsevier Ltd. All rights reserved.
Keywords: Lecanicillium lecanii; Polyurethane foam; Chitinases; Entomopathogenic fungi
1. Introduction
Verticillium are gathered mycoparasitic and entomopatho-
genic species that produce extracelullar enzymes, such as
chitinases. Several strains of Lecanicillium lecanii and
Verticillium fungicola were evaluated as chitinase producers,
and it is reported the activity to induce morphological changes
in phytopathogen fungi [1].
The strains of Verticillium fungicola USDA 4519 and those
of Lecanicillium lecanii USDA 974, USDA 2460, and ATCC
26854 showed the highest activities. Natural polymeric
substrates, such as chitin, have been used to support growth
of Lecanicillium lecanii as source of nutrients as well as inducer
[2]. Chitin is not degraded inside the cell due to its insolubility,
size, molecular complexity and heterogeneous composition but
* Corresponding author. Tel.: +52 5 5804 49 21; fax: +52 5 5804 47 12.
E-mail address: [email protected] (K. Shirai).
1359-5113/$ – see front matter # 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.procbio.2007.10.009
fungi secrete chitinases with different specificity, endochiti-
nases and exochitinases, which are able to transform or
hydrolyse chitin [3].
Verticillium lecanii (Lecanicillium lecanii) has been culti-
vated in solid substrate fermentation (SSF) using chitin as carbon
source. SSF produced a highly concentrated enzymatic extract
with very active chitinases [2]. The readily available sugar cane
bagasse was used as support for fungal growth in that study;
however, such SSF showed problems of substrate sterilization,
temperature and pH control, as well contamination of the system,
which was mainly detected at long process time. Besides, the
enzymatic extracts carried impureness from the organic support,
which involved a harder purification process.
The substrates in SSF, which are usually by products of agro-
industry, have some disadvantages, such as excessive thickness
of the substrate layer, low porosity, or inadequate internal
structures that disturbed the aeration, heat removal and
inefficient nutrient uptake. In most of these systems it is not
possible to separate residual solid substrate from biomass,
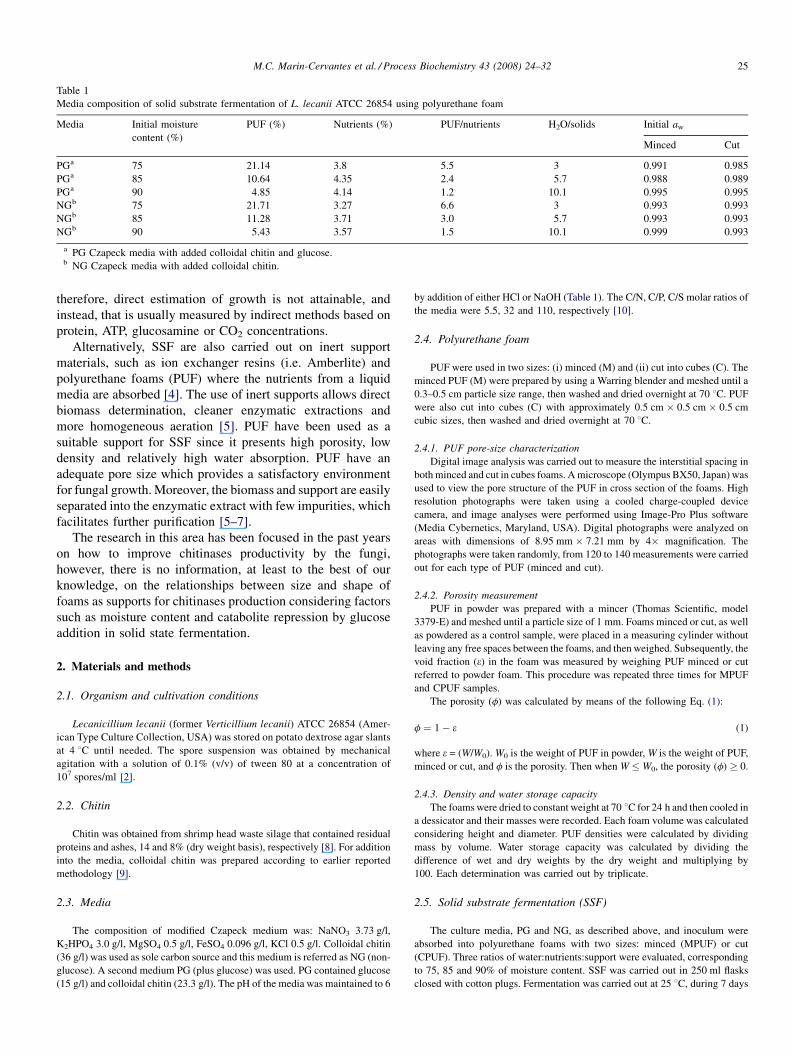
Table 1
Media composition of solid substrate fermentation of L. lecanii ATCC 26854 using polyurethane foam
Media Initial moisture
content (%)
PUF (%) Nutrients (%) PUF/nutrients H2O/solids Initial aw
Minced Cut
PGa 75 21.14 3.8 5.5 3 0.991 0.985
PGa 85 10.64 4.35 2.4 5.7 0.988 0.989
PGa 90 4.85 4.14 1.2 10.1 0.995 0.995
NGb 75 21.71 3.27 6.6 3 0.993 0.993
NGb 85 11.28 3.71 3.0 5.7 0.993 0.993
NGb 90 5.43 3.57 1.5 10.1 0.999 0.993
a PG Czapeck media with added colloidal chitin and glucose.b NG Czapeck media with added colloidal chitin.
M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–32 25
therefore, direct estimation of growth is not attainable, and
instead, that is usually measured by indirect methods based on
protein, ATP, glucosamine or CO2 concentrations.
Alternatively, SSF are also carried out on inert support
materials, such as ion exchanger resins (i.e. Amberlite) and
polyurethane foams (PUF) where the nutrients from a liquid
media are absorbed [4]. The use of inert supports allows direct
biomass determination, cleaner enzymatic extractions and
more homogeneous aeration [5]. PUF have been used as a
suitable support for SSF since it presents high porosity, low
density and relatively high water absorption. PUF have an
adequate pore size which provides a satisfactory environment
for fungal growth. Moreover, the biomass and support are easily
separated into the enzymatic extract with few impurities, which
facilitates further purification [5–7].
The research in this area has been focused in the past years
on how to improve chitinases productivity by the fungi,
however, there is no information, at least to the best of our
knowledge, on the relationships between size and shape of
foams as supports for chitinases production considering factors
such as moisture content and catabolite repression by glucose
addition in solid state fermentation.
2. Materials and methods
2.1. Organism and cultivation conditions
Lecanicillium lecanii (former Verticillium lecanii) ATCC 26854 (Amer-
ican Type Culture Collection, USA) was stored on potato dextrose agar slants
at 4 8C until needed. The spore suspension was obtained by mechanical
agitation with a solution of 0.1% (v/v) of tween 80 at a concentration of
107 spores/ml [2].
2.2. Chitin
Chitin was obtained from shrimp head waste silage that contained residual
proteins and ashes, 14 and 8% (dry weight basis), respectively [8]. For addition
into the media, colloidal chitin was prepared according to earlier reported
methodology [9].
2.3. Media
The composition of modified Czapeck medium was: NaNO3 3.73 g/l,
K2HPO4 3.0 g/l, MgSO4 0.5 g/l, FeSO4 0.096 g/l, KCl 0.5 g/l. Colloidal chitin
(36 g/l) was used as sole carbon source and this medium is referred as NG (non-
glucose). A second medium PG (plus glucose) was used. PG contained glucose
(15 g/l) and colloidal chitin (23.3 g/l). The pH of the media was maintained to 6
by addition of either HCl or NaOH (Table 1). The C/N, C/P, C/S molar ratios of
the media were 5.5, 32 and 110, respectively [10].
2.4. Polyurethane foam
PUF were used in two sizes: (i) minced (M) and (ii) cut into cubes (C). The
minced PUF (M) were prepared by using a Warring blender and meshed until a
0.3–0.5 cm particle size range, then washed and dried overnight at 70 8C. PUF
were also cut into cubes (C) with approximately 0.5 cm � 0.5 cm � 0.5 cm
cubic sizes, then washed and dried overnight at 70 8C.
2.4.1. PUF pore-size characterization
Digital image analysis was carried out to measure the interstitial spacing in
both minced and cut in cubes foams. A microscope (Olympus BX50, Japan) was
used to view the pore structure of the PUF in cross section of the foams. High
resolution photographs were taken using a cooled charge-coupled device
camera, and image analyses were performed using Image-Pro Plus software
(Media Cybernetics, Maryland, USA). Digital photographs were analyzed on
areas with dimensions of 8.95 mm � 7.21 mm by 4� magnification. The
photographs were taken randomly, from 120 to 140 measurements were carried
out for each type of PUF (minced and cut).
2.4.2. Porosity measurement
PUF in powder was prepared with a mincer (Thomas Scientific, model
3379-E) and meshed until a particle size of 1 mm. Foams minced or cut, as well
as powdered as a control sample, were placed in a measuring cylinder without
leaving any free spaces between the foams, and then weighed. Subsequently, the
void fraction (e) in the foam was measured by weighing PUF minced or cut
referred to powder foam. This procedure was repeated three times for MPUF
and CPUF samples.
The porosity (f) was calculated by means of the following Eq. (1):
f ¼ 1� e (1)
where e = (W/W0). W0 is the weight of PUF in powder, W is the weight of PUF,
minced or cut, and f is the porosity. Then when W �W0, the porosity (f) � 0.
2.4.3. Density and water storage capacity
The foams were dried to constant weight at 70 8C for 24 h and then cooled in
a dessicator and their masses were recorded. Each foam volume was calculated
considering height and diameter. PUF densities were calculated by dividing
mass by volume. Water storage capacity was calculated by dividing the
difference of wet and dry weights by the dry weight and multiplying by
100. Each determination was carried out by triplicate.
2.5. Solid substrate fermentation (SSF)
The culture media, PG and NG, as described above, and inoculum were
absorbed into polyurethane foams with two sizes: minced (MPUF) or cut
(CPUF). Three ratios of water:nutrients:support were evaluated, corresponding
to 75, 85 and 90% of moisture content. SSF was carried out in 250 ml flasks
closed with cotton plugs. Fermentation was carried out at 25 8C, during 7 days

M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–3226
sampling by duplicates for each condition. Solid substrate cultures were
collected from each flask, mixed with their same weight in water and pressur-
ized to 1000 psi. Cultures were centrifuged at 14,000 � g at 4 8C for 15 min.
Spores and mycelia were separated by filtration and used as crude enzyme for
assays.
2.5.1. Determination of fungal biomass
L. lecanii growth was measured by dry weight determinations. A statistical
programme STATISTICA 6.0 (Stat Soft, Inc., USA) was used to estimating the
specific growth rate by fitting the biomass data, X, to the logistic Eq. (2):
X ¼ Xmax
1þ ððXmax=X0Þ � 1Þe�mmaxt(2)
where X was the biomass determined by dry weight after time (t); X0 was the
initial biomass, Xmax was the maximal biomass (t!1), and mmax was the
specific growth rate. The X0, Xmax, and mmax values were estimated using a non-
linear estimation programme. Ye/x is the yield coefficient for enzyme on total
biomass produced given by the Eq. (3):
Ye=x ¼½emax�Xmax
(3)
where emax is the maximum enzyme activity that was determined from the
experimental data [7].
2.5.2. Estimation of glucose consumption with Gompertz model
The glucose concentration in SSF was measured by the reducing sugar
method [11]. The Gompertz model was applied to glucose concentration data
during time and the constants Smax, b and k in the culture media with added
glucose were estimated for enzymes production by using the non-lineal
estimation program. In this model, S is the shift of glucose concentration as
function of time t during SSF and it was considered as S = S0 � S(t), where S0 is
the initial amount of glucose added to the media and S(t) is the glucose
consumed at time (t). S is obtained according to the following Eq. (4):
S ¼ Smax expð�b expð�ktÞÞ (4)
where Smax is the maximum glucose consumed (t!1), b is a constant related
to the initial conditions (when t = 0, then S = Smaxexp(�b)) and k is the
consumption rate.
2.6. b-N-Acetyl hexosaminidase activity
The presence of b-N-acetyl hexosaminidase activity was determined in the
extracts from SSF using p-nitrophenyl-b-N-acetyl glucosamine (Sigma Che-
mical Co., St. Louis, MO) as substrate. 200 ml of crude enzyme was added to
200 ml of a citrate-phosphate buffer (0.2 M, pH 5.6). The solution was mixed
with 200 ml of p-nitrophenyl-b-N-acetyl glucosamine (1 mg/ml) and then
incubated with agitation (180 rpm) at 37 8C for 1 h. The reaction was quenched
by addition of 1 ml of NaOH 0.02 M. The released p-nitrophenol was evaluated
by a UV–vis spectrophotometry (Jenway 6305, Essex UK) at 400 nm. The
enzymatic unit was defined as the amount of enzyme that releases 1 mmol of p-
nitrophenol per minute [12].
2.7. Endochitinase activity
The endochitinase activity was measured by reduction of turbidity of
suspensions of 1% (w/v) of colloidal chitin in phosphate buffer (50 mM)
and NaN3 (0.01 M) at pH 6.7. 500 ml of crude enzyme was added to 500 ml
Table 2
Characteristics of polyurethane foams used in this study
Type of PUF Porosity (f) Void fraction (e)
Minced PUF 0.912 � 0.010 0.0875 � 0.0098
Cut PUF 0.955 � 0.05 0.0453 � 0.0051
of colloidal chitin. The mixture was incubated at 30 8C and stirred continuously
at 180 rpm for 24 h. Then it was diluted with 5 ml of water and the absorbance
was determined at 510 nm. The enzymatic unit was defined as the amount of
enzyme that decreases 5% the turbidity of a colloidal chitin suspension [13].
2.8. Statistical analysis
A randomized design was carried out in quadruplicate for experiments with
different moisture content levels, PUF sizes and glucose addition in SSF. The
programme SPSS version 8.0 (SPSS, Inc., USA 1997) computed the analysis of
variance with enzyme activity and biomass as response variables. The means
were compared with Tukey Kramer multiple means comparison test (P � 0.05).
2.9. Scanning electron microscopy
SSF samples were prepared by immersion in 5% (v/v) glutaraldehyde for
24 h at 4 8C and post-treated with OsO4 1% (w/v) for 2 h at 4 8C. Then they
were dehydrated in a graded alcohol series and covered with carbon and gold
prior to examination in the Scanning Electron Microscope (JEOL JSM-5900 LV,
Tokyo).
3. Results and discussion
Minced (MPUF) and cut PUF (CPUF), approximated
density of 0.018 g/cm3, were evaluated on their properties to
retain the culture media due to their capillarity and porosity.
The water storage capacity of MPUF and CPUF were
determined as 19.8 and 21.4 ml/g, respectively (Table 2).
CPUF presented honeycomb of polygonal cells of the
polymer while MPUF, which cells of the polymer were
destroyed during the mince process, was observed as irregular
forms. The averages of the interstitial areas were different for
cut and for minced PUF; the former was of 73.77 mm2 and the
latter 99.48 mm2, while the porosity of the cut one was 0.955
and for the minced 0.912. According to these data, MPUF
presented a decrease in the capillarity due to the decreased
porosity that had significant effect on the absorption of the
culture media compared with the CPUF (Table 2). The CPUF
was subjected to less mechanical force causing minor
destruction of pores than MPUF and therefore, it presents
higher porosity than the MPUF (Fig. 1). The effect of surface
tension (capillarity) is to draw liquid back into the borders. Due
to the existence of highly curve borders in CPUF, the capillarity
pressure was higher than in the MPUF thin films (Fig. 1).
3.1. Lecanicillium lecanii growth using PUF in SSF
The direct measurement of the biomass is one of the
advantages of using PUF in SSF with regard to other supports
[5], however the use of colloidal chitin in the medium hinders
this determination for its insolubility and retention in this type
of supports. The chitin fraction absorbed in the PUF was
Maximum water
absorption (ml/g PUF)
Interstitial spacing (mm2)
19.8 99.48 � 6.83
21.4 73.77 � 4.59

Fig. 1. Stereomicroscope micrographs of polyurethane foams (PUF) (4�): (a and b) cut PUF; (c and d) minced PUF.
Fig. 2. Biomass production of Lecanicillium lecanii ATCC26854 in SSF using
minced (M) and cut (C) PUF at various moisture content: (a) culture media
without addition of glucose: (*) M 75%, (&) C 75%, (^) M 85%, (~) C 85%,
(—) M 90%, (+) C 90%. (b) Culture media with addition of glucose: (*) M
75%, (&) C 75%, (^) M 85%, (~) C 85%, (—) M 90%, (+) C 90%. The
experimental data are mean of four determinations.
M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–32 27
measured and considered for the biomass calculation. The
curves of growth of Lecanicillium lecanii in MPUF and CPUF
with different moisture contents and glucose addition are shown
in Fig. 2.
As it was expected, the supplementation of glucose to the
media as well as the increase of moisture content favored the
production of biomass. The biomass in chitin media (NG) was
significantly lower in both types of PUF and at all moisture
content levels tested (Fig. 2).
The growth parameters were estimated on the type of PUF,
moisture content and chitin media only or with added glucose.
The estimated kinetic parameters showed that for all the
experiments the production of maximum biomasses concen-
trations (Xmax) increased with the addition of glucose, which
was the main source of variation rather than the type of support.
The highest Xmax were determined with 90% of moisture
content in PG medium, 0.193 and 0.15, for MPUF and CPUF,
respectively. However, in PG media there is no correlation
between the type of support and the production of biomass,
whereas, Xmax were higher in CPUF than in MPUF for all the
moisture contents in NG media. The maximum specific growth
rate (mmax) was the highest with 75% of moisture content in
CPUF (Table 3). Although the interstitial spacing in MPUF was
significantly higher than in CPUF (Table 2); the growth was not
significantly different in both PUFs (Fig. 2). This is in
agreement with the Van de Lagemaat and Pyle [7] report, where
it was suggested that size of PUF did not have a significant
effect on biomass production of Penicillium glabrum, the
growth was mainly limited by the substrate rather than PUF
pore size.

Table 3
Estimated growth kinetic parameters of L. lecanii ATCC26854 and enzyme production in SSF using PUF (minced or cut) at 75, 85 and 90% of moisture contents in
media with or without addition of glucose
Moisture content (%, w/w) Polyurethane foam size Culture media Xmax (g/g PUF) X0 (g/g PUF) mmax (h�1) R2
75 Ca NGc 0.012 0.00009 0.32 0.94
75 Mb NGc 0.011 0.00006 0.19 0.94
75 Ca PGd 0.023 0.00026 0.22 0.96
75 Mb PGd 0.050 0.00445 0.08 0.92
85 Ca NGc 0.032 0.00075 0.18 0.99
85 Mb NGc 0.013 0.00014 0.27 0.90
85 Ca PGd 0.068 0.00680 0.05 0.90
85 Mb PGd 0.047 0.00059 0.20 0.90
90 Ca NGc 0.059 0.00057 0.20 0.97
90 Mb NGc 0.044 0.00202 0.14 0.96
90 Ca PGd 0.150 0.01471 0.07 0.90
90 Mb PGd 0.193 0.00575 0.06 0.92
a C cut PUF.b M minced PUF.c NG Czapeck media with added colloidal chitin.d PG Czapeck media with added colloidal chitin and glucose.
M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–3228
The distribution of the fungal cells in MPUF with NG media
was observed as loose and disperse, whereas in CPUF the
growth was observed in loosely packed hyphae as shown in
Fig. 3a–c. The fungal growths in CPUF covered the surface by
formation of a net of intertwinement hyphae in PG media while
in MPUF L. lecanii grew forming aggregates in the ends of the
branches made from the sectioned cells of the PUF (Fig. 3 b–d).
The formation of filamentous mycelia in PG media might be
due to the media composition, which contained glucose that
favors this fungal morphology. However it is remarkable that
mycelial aggregation was observed in MPUF maybe due to
losses of water during SSF.
Fig. 3. Scanning electron micrographs of the Lecanicillium lecanii ATCC 26854 g
addition of glucose; (b) with addition of glucose; and minced PUF: (c) without ad
3.1.1. Glucose consumption during SSF
The glucose consumption through the time of cultivation in
the SSF was determined at several moisture contents and the
results are shown in Fig. 4. The glucose consumption in SSF
with CPUF was faster during the first 24 h compared to MPUF.
The glucose was depleted at 72 h with CPUF at any moisture
content, while the sugar consumptions with MPUF were
achieved at 96 h with 85 and 95% of moisture content. The
experimental evidences point out that the PUF size and
moisture contents were significant for the rate of glucose
consumption. The maximum glucose consumed (Smax) was
lower with CPUF than MPUF. The highest Smax was determined
rowths with colloidal chitin at 85% moisture content on cut PUF: (a) without
dition of glucose; (d) with addition of glucose.

Fig. 4. Glucose concentration during SSF of Lecanicillium lecanii ATCC26854
in minced (M) and cut (C) PUF at various moisture content: symbols (*) M
75%, (&) C 75%, (^) M 85%, (~) C 85%, (—) M 90%, (+) C 90%.
Fig. 5. pH determination of Lecanicillium lecanii ATCC 26854 in SSF using
minced (M) and cut (C) PUF in media with different moisture contents: (a) culture
media without addition of glucose: (*) M 75%, (&) C 75%, (^) M 85%, (~) C
85%, (—) M 90%, (+) C 90%. (b) Culture media with addition of glucose: (*) M
75%, (&) C 75%, (^) M 85%, (~) C 85%, (—) M 90%, (+) C 90%.
M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–32 29
at 90% moisture content with MPUF as support. Smax also
increased with the moisture content; however, the consumption
rates (k) were similar in all the experiments (Table 4).
3.1.2. Effect of the pH on fungal growth
The pH is an important environmental factor for fungal
growth and chitinases production and it changes due to the acid
or basic compounds produced by microbial use of nitrogen
sources and the modification of the solid substrate [14]. In SSF,
it is not possible to control the pH by addition of acids or bases
to the medium since homogeneous distribution cannot be
achieved when the media is already impregnated in the support.
Agitation is not recommendable at this stage due to
contamination risk and possible destruction of mycelia;
therefore, buffered substrates are usually added into the media
for that purpose.
In this study, the media were adjusted to an initial pH of 6, and
after fungal growth, the NG medium was slightly alkaline, ca. 7,
due to the assimilation of nitrogen source from chitin or salts
(Fig. 5a). In the PG medium, glucose was readily consumed and
the chitin assimilation followed, then the pH increased (Fig. 4,
Fig. 5b). For instance, the pH variation in the PG medium at 85%
of moisture content in CPUF was different from the earlier since
it presented a pH decrease to ca. 5 (Fig. 5b). However, after 72 h,
the pH increased to ca. 8 (Fig. 5b).
The maintenance of pH on near values of 6 during culture
had also a positive effect on the fungal growth in NG media,
since mmax were higher than those found in PG media (Table 3).
Table 4
Estimated parameters of glucose consumption during SSF of Lecanicillium lecani
Moisture content (%, w/w) Polyurethane foam size Smax
75 Ca 0.053
75 Mb 0.054
85 Ca 0.120
85 Mb 0.123
90 Ca 0.281
90 Mb 0.301
a C cut PUF.b M minced PUF.
Neither moisture content nor the PUF sizes were significant,
thus the factor that affected significantly the pH was the
addition of glucose.
3.1.3. Effect of moisture content on fungal growth
The moisture content can be defined as the water contained in
the medium able to dissolve nutrients, products, and moisturize
spores thus it promotes germination [14]. Fungi can grow in a
wide range of aw, 0.61–0.98, in SSF they grow well at aw of 0.96–
0.98 [15]. In this study, the foams presented 21.4 ml/g PUF and
19.8 ml/g of PUF of water absorption for cut and minced,
respectively (Table 2). This high water retention of PUF allowed
aw among 0.985–0.999, which are in the range for optimal fungal
growth. However it is important to consider that high retention of
water in the support can also favors the contamination by bacteria
or yeasts, as well as oxygen diffusion.
i using polyurethane foam as inert support
(g/g PUF) b (g/g PUF) k (h�1) R2
4.65 0.054 0.98825
3.02 0.055 0.98634
5.92 0.050 0.97790
2.51 0.045 0.95280
3.67 0.030 0.96352
2.76 0.052 0.97404
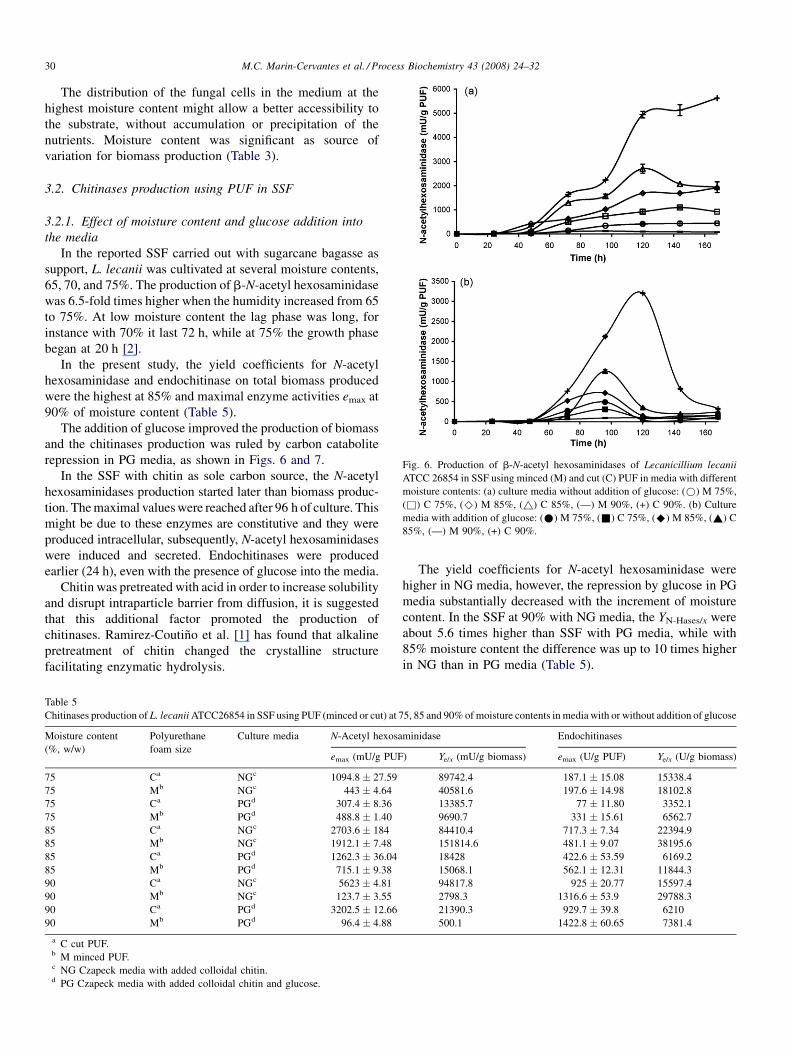
Fig. 6. Production of b-N-acetyl hexosaminidases of Lecanicillium lecanii
ATCC 26854 in SSF using minced (M) and cut (C) PUF in media with different
moisture contents: (a) culture media without addition of glucose: (*) M 75%,
(&) C 75%, (^) M 85%, (~) C 85%, (—) M 90%, (+) C 90%. (b) Culture
media with addition of glucose: (*) M 75%, (&) C 75%, (^) M 85%, (~) C
85%, (—) M 90%, (+) C 90%.
M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–3230
The distribution of the fungal cells in the medium at the
highest moisture content might allow a better accessibility to
the substrate, without accumulation or precipitation of the
nutrients. Moisture content was significant as source of
variation for biomass production (Table 3).
3.2. Chitinases production using PUF in SSF
3.2.1. Effect of moisture content and glucose addition into
the media
In the reported SSF carried out with sugarcane bagasse as
support, L. lecanii was cultivated at several moisture contents,
65, 70, and 75%. The production of b-N-acetyl hexosaminidase
was 6.5-fold times higher when the humidity increased from 65
to 75%. At low moisture content the lag phase was long, for
instance with 70% it last 72 h, while at 75% the growth phase
began at 20 h [2].
In the present study, the yield coefficients for N-acetyl
hexosaminidase and endochitinase on total biomass produced
were the highest at 85% and maximal enzyme activities emax at
90% of moisture content (Table 5).
The addition of glucose improved the production of biomass
and the chitinases production was ruled by carbon catabolite
repression in PG media, as shown in Figs. 6 and 7.
In the SSF with chitin as sole carbon source, the N-acetyl
hexosaminidases production started later than biomass produc-
tion. The maximal values were reached after 96 h of culture. This
might be due to these enzymes are constitutive and they were
produced intracellular, subsequently, N-acetyl hexosaminidases
were induced and secreted. Endochitinases were produced
earlier (24 h), even with the presence of glucose into the media.
Chitin was pretreated with acid in order to increase solubility
and disrupt intraparticle barrier from diffusion, it is suggested
that this additional factor promoted the production of
chitinases. Ramirez-Coutino et al. [1] has found that alkaline
pretreatment of chitin changed the crystalline structure
facilitating enzymatic hydrolysis.
Table 5
Chitinases production of L. lecanii ATCC26854 in SSF using PUF (minced or cut) at 7
Moisture content
(%, w/w)
Polyurethane
foam size
Culture media N-Acetyl hexosa
emax (mU/g PUF
75 Ca NGc 1094.8 � 27.59
75 Mb NGc 443 � 4.64
75 Ca PGd 307.4 � 8.36
75 Mb PGd 488.8 � 1.40
85 Ca NGc 2703.6 � 184
85 Mb NGc 1912.1 � 7.48
85 Ca PGd 1262.3 � 36.04
85 Mb PGd 715.1 � 9.38
90 Ca NGc 5623 � 4.81
90 Mb NGc 123.7 � 3.55
90 Ca PGd 3202.5 � 12.66
90 Mb PGd 96.4 � 4.88
a C cut PUF.b M minced PUF.c NG Czapeck media with added colloidal chitin.d PG Czapeck media with added colloidal chitin and glucose.
The yield coefficients for N-acetyl hexosaminidase were
higher in NG media, however, the repression by glucose in PG
media substantially decreased with the increment of moisture
content. In the SSF at 90% with NG media, the YN-Hases/x were
about 5.6 times higher than SSF with PG media, while with
85% moisture content the difference was up to 10 times higher
in NG than in PG media (Table 5).
5, 85 and 90% of moisture contents in media with or without addition of glucose
minidase Endochitinases
) Ye/x (mU/g biomass) emax (U/g PUF) Ye/x (U/g biomass)
89742.4 187.1 � 15.08 15338.4
40581.6 197.6 � 14.98 18102.8
13385.7 77 � 11.80 3352.1
9690.7 331 � 15.61 6562.7
84410.4 717.3 � 7.34 22394.9
151814.6 481.1 � 9.07 38195.6
18428 422.6 � 53.59 6169.2
15068.1 562.1 � 12.31 11844.3
94817.8 925 � 20.77 15597.4
2798.3 1316.6 � 53.9 29788.3
21390.3 929.7 � 39.8 6210
500.1 1422.8 � 60.65 7381.4
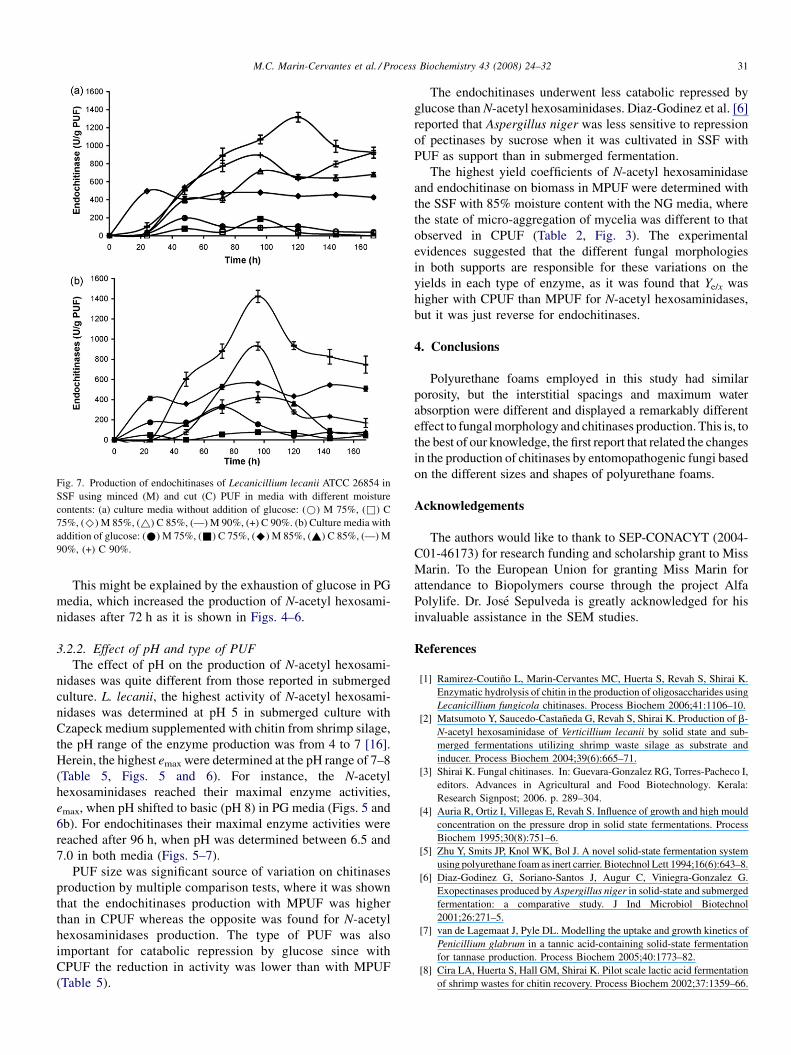
Fig. 7. Production of endochitinases of Lecanicillium lecanii ATCC 26854 in
SSF using minced (M) and cut (C) PUF in media with different moisture
contents: (a) culture media without addition of glucose: (*) M 75%, (&) C
75%, (^) M 85%, (~) C 85%, (—) M 90%, (+) C 90%. (b) Culture media with
addition of glucose: (*) M 75%, (&) C 75%, (^) M 85%, (~) C 85%, (—) M
90%, (+) C 90%.
M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–32 31
This might be explained by the exhaustion of glucose in PG
media, which increased the production of N-acetyl hexosami-
nidases after 72 h as it is shown in Figs. 4–6.
3.2.2. Effect of pH and type of PUF
The effect of pH on the production of N-acetyl hexosami-
nidases was quite different from those reported in submerged
culture. L. lecanii, the highest activity of N-acetyl hexosami-
nidases was determined at pH 5 in submerged culture with
Czapeck medium supplemented with chitin from shrimp silage,
the pH range of the enzyme production was from 4 to 7 [16].
Herein, the highest emax were determined at the pH range of 7–8
(Table 5, Figs. 5 and 6). For instance, the N-acetyl
hexosaminidases reached their maximal enzyme activities,
emax, when pH shifted to basic (pH 8) in PG media (Figs. 5 and
6b). For endochitinases their maximal enzyme activities were
reached after 96 h, when pH was determined between 6.5 and
7.0 in both media (Figs. 5–7).
PUF size was significant source of variation on chitinases
production by multiple comparison tests, where it was shown
that the endochitinases production with MPUF was higher
than in CPUF whereas the opposite was found for N-acetyl
hexosaminidases production. The type of PUF was also
important for catabolic repression by glucose since with
CPUF the reduction in activity was lower than with MPUF
(Table 5).
The endochitinases underwent less catabolic repressed by
glucose than N-acetyl hexosaminidases. Diaz-Godinez et al. [6]
reported that Aspergillus niger was less sensitive to repression
of pectinases by sucrose when it was cultivated in SSF with
PUF as support than in submerged fermentation.
The highest yield coefficients of N-acetyl hexosaminidase
and endochitinase on biomass in MPUF were determined with
the SSF with 85% moisture content with the NG media, where
the state of micro-aggregation of mycelia was different to that
observed in CPUF (Table 2, Fig. 3). The experimental
evidences suggested that the different fungal morphologies
in both supports are responsible for these variations on the
yields in each type of enzyme, as it was found that Ye/x was
higher with CPUF than MPUF for N-acetyl hexosaminidases,
but it was just reverse for endochitinases.
4. Conclusions
Polyurethane foams employed in this study had similar
porosity, but the interstitial spacings and maximum water
absorption were different and displayed a remarkably different
effect to fungal morphology and chitinases production. This is, to
the best of our knowledge, the first report that related the changes
in the production of chitinases by entomopathogenic fungi based
on the different sizes and shapes of polyurethane foams.
Acknowledgements
The authors would like to thank to SEP-CONACYT (2004-
C01-46173) for research funding and scholarship grant to Miss
Marin. To the European Union for granting Miss Marin for
attendance to Biopolymers course through the project Alfa
Polylife. Dr. Jose Sepulveda is greatly acknowledged for his
invaluable assistance in the SEM studies.
References
[1] Ramirez-Coutino L, Marin-Cervantes MC, Huerta S, Revah S, Shirai K.
Enzymatic hydrolysis of chitin in the production of oligosaccharides using
Lecanicillium fungicola chitinases. Process Biochem 2006;41:1106–10.
[2] Matsumoto Y, Saucedo-Castaneda G, Revah S, Shirai K. Production of b-
N-acetyl hexosaminidase of Verticillium lecanii by solid state and sub-
merged fermentations utilizing shrimp waste silage as substrate and
inducer. Process Biochem 2004;39(6):665–71.
[3] Shirai K. Fungal chitinases. In: Guevara-Gonzalez RG, Torres-Pacheco I,
editors. Advances in Agricultural and Food Biotechnology. Kerala:
Research Signpost; 2006. p. 289–304.
[4] Auria R, Ortiz I, Villegas E, Revah S. Influence of growth and high mould
concentration on the pressure drop in solid state fermentations. Process
Biochem 1995;30(8):751–6.
[5] Zhu Y, Smits JP, Knol WK, Bol J. A novel solid-state fermentation system
using polyurethane foam as inert carrier. Biotechnol Lett 1994;16(6):643–8.
[6] Diaz-Godinez G, Soriano-Santos J, Augur C, Viniegra-Gonzalez G.
Exopectinases produced by Aspergillus niger in solid-state and submerged
fermentation: a comparative study. J Ind Microbiol Biotechnol
2001;26:271–5.
[7] van de Lagemaat J, Pyle DL. Modelling the uptake and growth kinetics of
Penicillium glabrum in a tannic acid-containing solid-state fermentation
for tannase production. Process Biochem 2005;40:1773–82.
[8] Cira LA, Huerta S, Hall GM, Shirai K. Pilot scale lactic acid fermentation
of shrimp wastes for chitin recovery. Process Biochem 2002;37:1359–66.

M.C. Marin-Cervantes et al. / Process Biochemistry 43 (2008) 24–3232
[9] Pegg GF. Chitinase From Tomato ce:italicLycopersicon esculentum/ce:i-
talic Methods in Enzymol, vol. 161. New York: Academic Press; 1988. pp.
484–487.
[10] Matsumoto Y, Revah S, Saucedo-Castaneda G, Shirai K. Chitinases
production in solid state fermentation and submerged fermentation by
Verticillium lecanii with silage shrimp as substrate. In: Uragami T, Kurita
K, Fukamizo T, editors. Chitin and Chitosan in Life Sciences. Tokyo:
Kodansha Scientific Ltd.; 2001. p. 403–6.
[11] Miller GL. Use of dinitrosalicylic acid reagent for determination of
reducing sugars. Anal Chem 1959;31:426–8.
[12] Ohtakara A. Chitinase and b-N-acetyl hexosaminidase from Pycnoporus
cinnabarinus. Methods in Enzymol, vol. 161. New York: Academic Press;
1988. pp. 462–467.
[13] Tronsmo A, Harman GE. Detection and quantification of N-acetyl-b-D-
glucosaminidase, chitobiosidase, and endochitinase in solutions and on
gels. Anal Biochem 1993;208:74–9.
[14] Mitchell DA, Berovic M, Krieger N. Biochemical engineering aspects of
solid-state bioprocessing. In: Scheper Th., editor. Advances in Bio-
chemical Engineering Biotechnology. Berlin: Springer-Verlag; 2000. p.
65–138.
[15] Tengerdy RP. Solid substrate fermentation. Trends Biotechnol 1985;3
(4):96–9.
[16] Matsumoto Y. Production of fungal chitinases by solid-state fermentation.
Mexico: Universidad Autonoma Metropolitana; M. Ph. in Biotechnology
thesis; 2001, p. 115.
Copyright © 2022 FDOKUMEN




















