
Effect of mine tailing on the spatial variability of soil nematodes from lead pollution in La Union...
12
Effect of mine tailing on the spatial variability of soil nematodes from lead pollution in La Union (Spain) José Antonio Rodríguez Martín a, ⁎, Carmen Gutiérrez b , Miguel Escuer b ,M a Teresa García-González b , Raquel Campos-Herrera b , Nancy Águila b a Depto. Medio Ambiente, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (I.N.I.A), Carretera de A Coruña 7.5 km, 28040 Madrid, Spain b Instituto de Ciencias Agrarias, ICA-CSIC, Serrano, 115, 28006 Madrid, Spain HIGHLIGHTS • High lead contents from mine tailing diminish soil biodiversity. • Spatial relationship showing the soil Pb pollution impact of mines on soil health. • SOM and nutrients are the most influen- tial factors on soil nematodes in La Union. • Nematode populations have well adapted to high Pb after a long mining period. GRAPHICAL ABSTRACT abstract article info Article history: Received 23 July 2013 Received in revised form 22 November 2013 Accepted 16 December 2013 Available online xxxx Keywords: Mine tailing Multivariate geostatistics Nematode trophic groups Heavy metal pollution Metabolic footprints The Cartagena–La Union mining district, exploited since the end of the 3rd century BC, was one of the world's largest lead producers in the 19th century. Although activity ceased in 1991, today mining residues pose a huge pollution problem. This study characterises lead contents (total and DPTA) and other soil parameters (N, P, K, pH, SOM, CaCO 3 , granulometric fraction, etc.) using multivariate geostatistical methods in relation to nematode diversity. In this work, trophic groups and metabolic footprints of soil nematodes were measured using 193 samples from the mining, natural and agricultural areas in this district. We explored the relationship between soil health and nematode communities. High lead concentrations were quantified: mean 8500 mg kg -1 for total and 340 mg kg -1 for DPTA in this mining area. Although nematode diversity was broad (81 taxa), their di- versity, abundance and metabolic footprints significantly reduced in the mining area. Significant differences in the nematode community structure were observed, and the relative abundance of predators was sensitive to mine and agricultural activities, whilst omnivores reduced only in the agricultural area, and bacterial feeders exhibited a dif- ferential response to both anthropogenic disturbances. The total abundance of nematodes, trophic groups and c–p groups correlated negatively with soil Pb contents, and a positive relationship was found with SOM and N, P and K contents. © 2013 Elsevier B.V. All rights reserved. 1. Introduction For a considerable time, soil was considered a means with a practi- cally unlimited capacity to accumulate pollutants without immediately Science of the Total Environment 473–474 (2014) 518–529 ⁎ Corresponding author. Tel.: +34 91 3476795. E-mail address: [email protected] (J.A. Rodríguez Martín). 0048-9697/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.scitotenv.2013.12.075 Contents lists available at ScienceDirect Science of the Total Environment journal homepage: www.elsevier.com/locate/scitotenv
Transcript of Effect of mine tailing on the spatial variability of soil nematodes from lead pollution in La Union...

Science of the Total Environment 473–474 (2014) 518–529
Contents lists available at ScienceDirect
Science of the Total Environment
j ourna l homepage: www.e lsev ie r .com/ locate /sc i totenv
Effect ofmine tailing on the spatial variability of soil nematodes from leadpollution in La Union (Spain)
José Antonio Rodríguez Martín a,⁎, Carmen Gutiérrez b, Miguel Escuer b, Ma Teresa García-González b,Raquel Campos-Herrera b, Nancy Águila b
a Depto. Medio Ambiente, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (I.N.I.A), Carretera de A Coruña 7.5 km, 28040 Madrid, Spainb Instituto de Ciencias Agrarias, ICA-CSIC, Serrano, 115, 28006 Madrid, Spain
H I G H L I G H T S G R A P H I C A L A B S T R A C T
• High lead contents from mine tailingdiminish soil biodiversity.
• Spatial relationship showing the soil Pbpollution impact of mines on soil health.
• SOM and nutrients are the most influen-tial factors on soil nematodes in La Union.
• Nematodepopulationshavewell adaptedto high Pb after a long mining period.
⁎ Corresponding author. Tel.: +34 91 3476795.E-mail address: [email protected] (J.A. Rodríguez Martín
0048-9697/$ – see front matter © 2013 Elsevier B.V. All rihttp://dx.doi.org/10.1016/j.scitotenv.2013.12.075
a b s t r a c t
a r t i c l e i n f oArticle history:Received 23 July 2013Received in revised form 22 November 2013Accepted 16 December 2013Available online xxxx
Keywords:Mine tailingMultivariate geostatisticsNematode trophic groupsHeavy metal pollutionMetabolic footprints
The Cartagena–La Union mining district, exploited since the end of the 3rd century BC, was one of the world'slargest lead producers in the 19th century. Although activity ceased in 1991, today mining residues pose ahuge pollution problem. This study characterises lead contents (total and DPTA) and other soil parameters(N, P, K, pH, SOM, CaCO3, granulometric fraction, etc.) using multivariate geostatistical methods in relation tonematode diversity. In this work, trophic groups and metabolic footprints of soil nematodes were measuredusing 193 samples from the mining, natural and agricultural areas in this district. We explored the relationshipbetween soil health and nematode communities. High lead concentrations were quantified: mean 8500 mg kg−1
for total and 340 mg kg−1 for DPTA in thismining area. Although nematode diversitywas broad (81 taxa), their di-versity, abundance and metabolic footprints significantly reduced in the mining area. Significant differences in thenematode community structure were observed, and the relative abundance of predators was sensitive to mine andagricultural activities, whilst omnivores reduced only in the agricultural area, and bacterial feeders exhibited a dif-ferential response to both anthropogenic disturbances. The total abundance of nematodes, trophic groups and c–pgroups correlated negatively with soil Pb contents, and a positive relationship was found with SOM and N, P and Kcontents.
© 2013 Elsevier B.V. All rights reserved.
).
ghts reserved.
1. Introduction
For a considerable time, soil was considered a means with a practi-cally unlimited capacity to accumulate pollutants without immediately

519J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
posing harmful effects for the environment or human health. Howeverwe now realise that this is not true. The general public is aware of theharmful potential of some soil trace elements, commonly known asheavy metals. The natural concentration of heavy metals in soilsdepends primarily on geological parent material composition (Alloway,1995; García Navarro et al., 2011). The chemical composition of parentmaterial and weathering processes naturally condition the concentrationof different heavy metals in soils (Roca-Perez et al., 2010). Furthermore,variability of heavy metals in soil is also associated with variability ofthe physico-chemical properties of soil. pH, organicmatter, clayminerals,metal oxides, oxidation reduction reactions, ionic exchange processes, oradsorption, desorption and complexation phenomena are the mainedaphic characteristics that relate to metal retention in soil (Ramos-Miras et al., 2011; Roca-Perez et al., 2010). The natural concentration ofheavymetals in soil is generally very low and tends to remainwithin nar-row limits to ensure an optimum ecological equilibrium. Nonethelesshuman activities, which involve large quantities of heavy metals beingemitted into the environment, have dramatically increased naturalconcentrations. Mining activities are well-known for their deleteriouseffects on the environment given the presence of high volumes of tailingsand toxic metal concentrations (Conesa et al., 2006; Garcia et al., 2008;Navarro et al., 2008). The Sierra Minera of Cartagena was one of themost important lead reserves in Europe in the 19th century. The LaUnion mining district has undergone important mining and metallurgicactivities since ancient times, such as extracting and exploiting lead andsilver in Roman times, and it was an important mining centre for morethan 2500 years. Given the long-standing mining activity in the area,high concentrations of heavy metals, especially lead, have been recordedin the soil (Conesa et al., 2006; Martínez García et al., 2001).
Soil disturbances affect the structure communities of soil organisms.By studying these changes, we can assess the deterioration of the soilenvironment. Soil nematodes are interesting for such purposes as theyare abundant and they offer high diversity. Several trophic groups arewell-adapted to a wide range of environmental conditions, play an im-portant role in soil ecological processes, such as primary production, nu-trient cycling, decomposition, and respond rapidly to disturbances. Allthese features make them useful indicators of soil health (Neher,2001; Yeates and Bongers, 1999). The assessment of changes in thestructure and function of nematode communities can be used to evalu-ate disturbances in soil ecosystems (Bongers and Ferris, 1999) or tostudy the effect of heavy metal pollution (Georgieva et al., 2002).Some studies have recently detected that the trophic groups and long-living persistor-type nematodes around the mining area are not soabundant, especially in relation to Pb and Zn pollution (Park et al.,2011; Shao et al., 2008). Other studies have observed that the activitiesand species richness of nematode communities have diminished inpolluted soils in accordance with specific heavy metals and theirconcentrations (Park et al., 2011; Šalamún et al., 2012).
Several indices are used as tools to assess these changes. Ecologicalindices, based on abundance, species richness and equitability amongsttaxa, provide information about the diversity of nematode communities(Höss et al., 2011). Functional indices are useful tools to provide infor-mation on the functioning and condition of the nematode food web(Bongers, 1990; Ferris et al., 2001). They are based on trophic preference(plant feeders, bacterial feeders, fungal feeders, root-fungal feeders,omnivores and predators) and life strategy assessed with a c–p(coloniser–persister) scale from 1 to 5 (from extreme r- to extremeK-strategists, where the coloniser nematodes are considered op-portunist and stress-tolerant, whereas the persisters are sensitive),and the relative weighted abundance of functional guilds (based onfeeding habits and a c–p classification, weighted according to their indi-cator characteristics). The new metabolic footprint concept improvesfunctional indices as they help estimate the biomass and metabolicactivity for each functional group of the foodweb, and also providemet-rics for themagnitudes of the ecosystem functions and services provid-ed by components of the nematode soil food web (Ferris, 2010). This
work aims to determine the lead content and influence of soil parame-ters in the abandoned Cartagena–La Unión mining district (E Spain), toanalyse spatial variability and to investigate their effects on the trophicstructure, c–p scaling and metabolic footprints of soil nematodes underdifferent soil land use and lead pollution conditions. As mining workstopped a long time ago, we hypothesise that the soil nematodes inthe mining area are well-adapted to high concentrations of heavymetals.
2. Material and methods
2.1. Study area and sampling
The Cartagena–La Unión District (100 km2) is located in the south-east of the Iberian Peninsula (Fig. 1) and presents high heterogeneityin geological and mineral paragenesis. At the end of the Miocene, vastpostorogenic magmatic activity took place, which produced importantpolymetallic ore deposits (Garcia et al., 2008). La Unión was an impor-tant mining centre for more than 2500 years, but its activity ceased in1991. The ore deposits of this zone include iron, lead and zinc, whichare the main metal components (Conesa et al., 2006). As a result ofthe long-standingmining activity there, large volumes ofwaste contain-ing high concentrations of heavy metals, especially lead and zinc, havebeen generated and accumulated in this area duringmineral concentra-tion and smelting processes (Benedicto et al., 2008). The mineralogy ofthe Cartagena–La Unión district was performed by X-ray powder dif-fraction (XRD). Quartz (46%) andmuscovite (21%)were themost repre-sentative minerals. They were followed by calcite (11%), feldspars(CaNa 4.5% and K 4%), dolomite (4.5%), kaolinite (3.4%) and, to a lesserextent, gypsum (1%), chlorite (1.7%), goethite (0.6%) hematite (0.4%),smectite (0.3%), paragonite (0.4%), barite (0.1%) and jarosite (0.1%),reflecting the heterogeneity of the materials in the study area. Thestudy area is characterised by a semiarid Mediterranean climate, with200–350 mm of rainfall, which concentrates in spring and autumn.The vegetation coverage in the study area offers broad diversity. Indeed,238 taxa from 49 families of vascular plants have been recordedand correspond to forests of Pinus halepensis, shrub areas containingChamaerops humilis, Pistacea lentiscus. There are also species of muchbotanic interest: endemic plants like Tetraclinis articulata, Cheirolophusmansanetianus, Limonium carthaginense, Teucrium carthaginense,T. freynii, and africanisms such as Periploca angustifolia and Maytenussnegalensis.
The samplingprotocol aims to ensure soil variability in the definitionof three land uses, hence three sampling intensities were used: miningarea (0.5 × 0.5 km), corresponding to recent mining works and largevolumes of waste; natural area (1 × 1 km), covered with forest, shruband grasses, and some ancient mine wastes; agricultural area(2 × 2 km), at a distance from where mining activities took place,where vegetables and fruits are produced. Sampling points were locatedby a global positioning system and orthophotos (1 m/pixel). The sampleswere taken in spring and autumn during the 2010–2011 period. At eachsampling site, 21 subsamples were taken with the Eijkelkamp soil sam-pling kit from the upper 20 cm of soil to obtain three soil subsamplesfrom each location. Subsamples were thoroughly mixed to provide acomposite sample of 3 kg of soil. In all, 193 sample points were defined,with 76, 66 and 51 from the natural, mining and agricultural areas,respectively.
2.2. Soil chemical analyses
The soil samples were firstly sieved by a 2 mm grid and were thenair-dried. Each sample soil texture was determined. After shaking witha dispersing agent, sand (63 μm–2 mm) was separated from clay andsilt with a 63 μm sieve (wet sieving). Clay (b2 μm) and silt (2–63 μm)fractions were determined by the pipette method (sedimentation). Astandard soil analysis was carried out to determine the soil reaction

Fig. 1. General map of the study area and scheme of the sampling design showing the 193 samples in the Cartagena–La Unión mining area (Murcia, E Spain).
520 J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
(pH) in a 1:2.5 soil–water suspension (measured by the glass electrodeCRISON model Microph 2002). Firstly, soil organic matter (SOM) wasanalysed by the Walkley–Black method (Walkley, 1935). Carbonateconcentration was established by a manometric measurement of theCO2 released following acid (HCl) dissolution (Houba et al., 1995). Inaddition, exchangeable potassium (K) and available phosphorous (P)were determined byOlsen's extractionmethod. Ca,Mg, andNa contentswere extracted by the ammonium acetate method, whilst Pb contentswere extracted by DTPA and an aqua regia digestion of the soil fractionin a microwave in accordance with standard ISO 11466. The Pb, Ca, Mgand Na in the soil extracts were determined by optical emission spec-trometry (IPC) with an ICAP-AES plasma spectrometer. The methodvalidation of the analytical procedure was performed with a calcareousloam soil (BCR-141 R), obtained from the European Commission Com-munity Bureau of Reference.
2.3. Nematode analyses
The nematode population was extracted for each composite samplefrom 100 g of fresh soil. The nematode population was extracted foreach composite sample from 100 g of fresh soil. All the nematodeswere counted and expressed as 100 g of dry weight soil. One hundredindividuals from each soil sample were identified mainly at the genuslevel (Andrássy, 2005; Bongers, 1988; Siddiqi, 2001). Different ecophys-iological attributeswere assigned to identify nematode taxa: i): Biomassvalues (Andrassy, 1956; Ferris et al., 2001) were obtained from theNematode–Plant Expert Information System (NEMAPLEX) (Ferris,
1998). ii) Trophic groups, predators, omnivores bacterial, fungal andplant feeders, were assessed (Yeates et al., 1993); the Filenchus genuswas considered a fungal feeder as Okada demonstrated (Okada et al.,2005; Okada and Kadota, 2003) its capacity to survive and reproduceon fungi alone. iii) Coloniser–persister groups: based on the c–p valuewithin a 1–5 range (Bongers, 1990; Bongers and Bongers, 1998).iv): Functional guild: as result of a combination of trophic with c–pgroups (Bongers and Bongers, 1998; Ferris et al., 2001). v): Metabolicfootprints that measure the functional service of nematodes to the soilecosystem (Ferris, 2010; Ferris et al., 2012). They have two components:a production component based on the amount of C used for growth andreproduction; a respiration component based on the C used formetabolicactivity. The enrichment footprint (EFP) corresponds to opportunist nem-atodes, bacterial and fungal feeders with a low c–p value (Bongers andFerris, 1999), and represents the ecosystem nutrient mineralisation ser-vice. The structure footprint (SFP) corresponds to the persister nema-todes with a high c–p value, which belong to higher trophic levelnematodes, including predators of opportunistic species, which performa regulatory function in the food web (Ferris et al., 2001). The metabolicfootprints of plant (PltFP), bacterial (BacFP) and fungal (FunFP) feedersindicate the amount of C and energy entering the soil food web throughtheir respective channels.
We used the terms “total abundance” assessed as the number of in-dividuals, trophic and c–p groups in the sample, and the term “relativeabundance” assessed as a percentage of total abundance in order to pro-vide standardised comparison of the numbers of different features ofthe free leaving nematode communities of organisms (Bush et al.,

521J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
1997). In this sense, different proportions of taxa were established inthe community, assessing genera dominance as eudominant (N10%),dominant (5–10%), and subdominant (b5%); (Šalamún et al., 2012).
2.4. Statistical and geostatistical analyses
A standard statistical analysis (mean,median, quartiles, standard de-viation, etc.) was carried out to describe soil properties, lead contentsand nematofauna in the three land uses. Significant differences betweenmeans (natural, mining and agricultural areas) were assessed throughnon-parametric Kruskal–Wallis tests. The PCA (principal componentanalysis) was used to study and visualise the correlations between soilparameters and to obtain non-correlated factors, which are linear com-binations of these soil chemical analyses. The varimax rotational tech-nique was utilised in the PCA for easier interpretation purposes. Theseanalysis statistics are essential to understand the relationships betweensoil elements, and one or more new variables were created (Factors),each representing a cluster of interrelated variables in the soil data set.
Furthermore, the Canonical Correlation Analysis (CCorA) is one ofthe many methods that allows us to study the relationship betweentwo sets of variables. We used CCorA to study the correlation betweensoil factors and the nematode structure communities, and to extract aset of canonical variables from these tables, which are as best correlatedwith both tables as possible, and are orthogonal to each other. In CCorA,there are two tables and the ultimate intentionwas tomaximise the co-variance between two sets of variables and tominimise their respectivevariance (Kianifard, 1993). Let Y1 and Y2 be (the soil factor, obtained byPCA), and the response variables (Y2) based on the nematode structurecommunities), with variables p and q, respectively:
p ið Þ ¼ cor Yla ið Þ;Y2b ið Þð Þ ¼ cov Yla ið Þ;Y2b ið Þð Þvar Yla ið Þð Þ: var Y2b ið Þð Þ ð1Þ
The CCorA (used considerably in ecology) provided two vectors, a(i)and b(i), which are maximised. Constraints must be introduced so thatthe solution for a(i) and b(i) is unique. The purposes were to maximisethe covariance between Y1a(i) and Y2b(i) and tominimise their respec-tive variance. Three CCorAwere performed to assess the effect of soil pa-rameters on nematode variables: 1) total abundance of individuals andtrophic groups; 2) abundance of the c–p groups; 3) metabolic foot-prints. Based on these analyses, we selected the variables with thehighest correlation value to perform the geostatistical analysis.
The PCA soil factors and the main nematode bioindicators selectedfrom the three CCorA analyseswere used to analyse the spatial structureand to assess the spatial connection between both. Semivariogramswere done to establish the degree of spatial continuity of the soil param-eters and heavy metals amongst the data points, and to determine the
Table 1Statistical summary of the general descriptive soil parameters and Pb contents. Mean values fo
Mean Median SD Min.
S.O.M (%) 2.29 2.00 1.51 0.08pH 7.66 8.10 1.02 3.00CaCO3 (%) 14.7 10.0 12.6 0.0E.C. 1.32 0.36 2.08 0.07A.W. 13.0 13.0 3.65 2.8Clay (%) 14.6 13.2 7.0 2.4Silt (%) 33.6 32.6 10.8 4.1Sand (%) 51.9 52.4 14.9 15.6N (%) 0.11 0.10 0.07 0.01P (mg kg−1) 12.6 4.0 18.8 0.0K (mg kg−1) 287 256 230 3Pb (A. regia) 5202 2209 7033 26Pb (DTPA) 261.9 147.0 327.5 1.8
Total soil samples 193. Value inside parentheses in the natural, mining and agricultural area(P b 0.01) are indicatedbydifferent letters after thenon-parametricKruskal–Wallis test SD, stanconductivity (dS m−1).
range of spatial dependence. Variogram γ was calculated using therelative locations of the samples (Goovaerts, 1997), defined as (1):
y hð Þ ¼12n
Xi¼n
i¼1
Z u1ð Þ−Z uiþh
� �� �2 ð2Þ
where Z(ui) is the Z value at location ui, and Z(ui + h) is the Z value at alocation separated from ui by distance h.
Kriging estimates were calculated as the weighted sums of the adja-cent sampled concentrations using an experimental semivariogram.There are many different kriging algorithms, most of which are reviewedin Goovaerts (1997)with references to soil applications. Soil factors weremapped by ordinary kriging (OK). All the statistical analyseswere carriedout using the SPSS statistical package forWinV19 (SPSS/PC advanced sta-tistics, v.21.9, Chicago: SPSS Inc.), XLSTAT (Addinsoft Version 2012.2.02),ISATIS V10.0 and the Geostatistical Analyst extension for ArcGis 9.3.
3. Results and discussion
3.1. Soil properties and lead contents
The summary statistics of soil parameters and Pb contents in the top-soils are listed in Table 1. Soil pH ranged from 3.0 to 8.7 (amean of 8.1),which is awide-ranging variation, although 80%of the values for the soilreaction were estimated to be between 6.5 and 8.3. Significant differ-ences were observed between the mine and alkaline soils from theagricultural area, where a high percentage of carbonates was recorded(the upper 50% in several samples). The same trend was observed forCaCO3 content, with the highest level found in arable alkaline soils,which provided the conditions to retain the metals (Plassard et al.,2000). The low pH in themining area canmobilise the cationic element,thus affecting the retention of heavy metals in the soil matrix and in-creasing their uptake rate by plants (Gil et al., 2004). The highest electri-cal conductivity (EC) value was recorded in mine soils. EC is controlledby ionic concentration, clay, and soil moisture (Alloway, 1995), and itpartly conditions the mobility of heavy metals in soil solution. Both pHand EC affect edaphicmicroorganisms, and bacterial populations gener-ally prefer alkaline soils, whereas fungi are more tolerant to pH and sa-linity. The mean percentage of organic matter (SOM) was 2.29%, whichranged from low values of 0.10% in mine bare soils from recently aban-doned sites (mean value of 1.56% in the area) to 1.88% in the agriculturalarea and to 8.05% in natural forest areas (mean value of 3.20%) due tothe accumulation of vegetal materials. The available water capacity(AW), affected by the abundance of rock fragments and soil texture,did not significantly differ in the studied areas. However, significant dif-ferences were observed amongst areas for granulometric fraction, and
r the natural, mining and agricultural areas.
Max. Natural (76) Mining (66) Agricultural (51)
8.05 3.20 (b) 1.56 (a) 1.88 (a)8.70 7.88 (b) 7.08 (a) 8.10 (c)62.0 13.5 (b) 8.7 (a) 24.2 (c)18.87 0.97 (a) 2.14 (b) 0.79 (a)23.8 13.1 (a) 12.7 (a) 13.2 (a)41.6 13.1 (a) 11.5 (a) 20.7 (b)64.7 20.9 (b) 16.2 (a) 24.6 (c)93.5 52.7 (b) 60.4 (c) 39.6 (a)0.37 0.15 (c) 0.08 (a) 0. 10 (b)99.0 7.7 (b) 3.1 (a) 32.2 (c)1513 287 (b) 133 (a) 485 (c)37,652 4044 (b) 8520 (c) 2633 (a)1990.0 257.3 (b) 343.9 (b) 162.7 (a)
s indicates the number of soil samples. Significant differences amongst the three areasdarddeviation. S.O.M: soil organicmatter (%). CaCO3: soil carbonates (%). EC: soil electrical
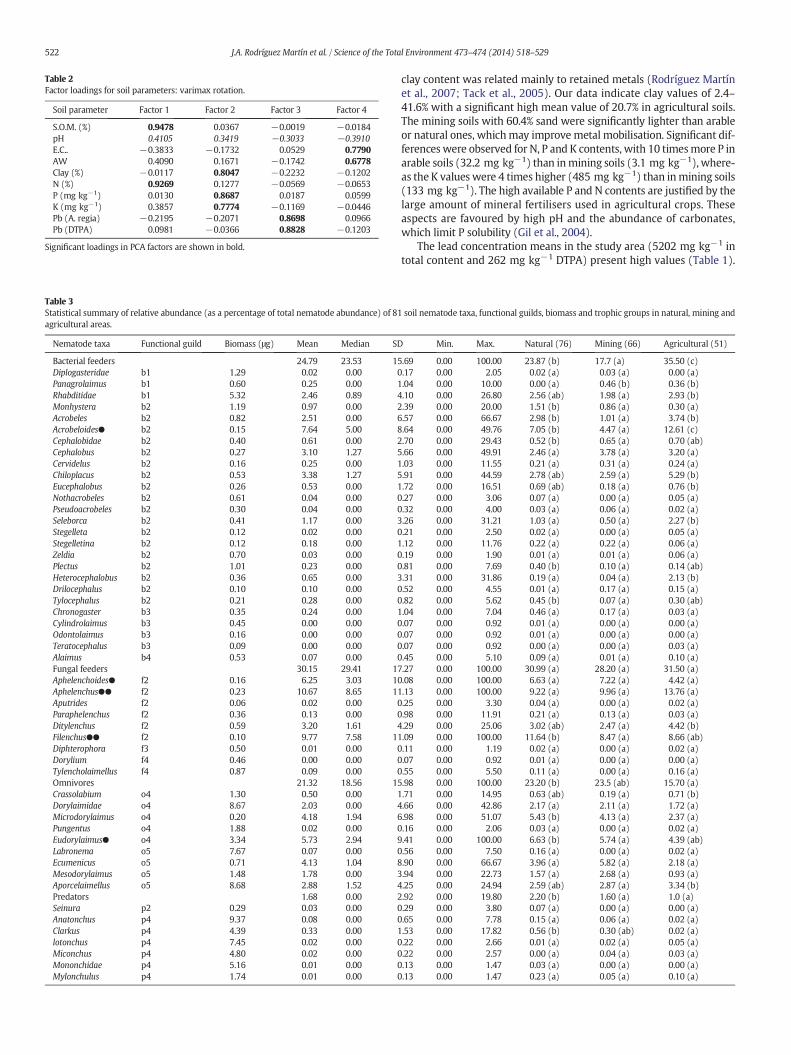
Table 2Factor loadings for soil parameters: varimax rotation.
Soil parameter Factor 1 Factor 2 Factor 3 Factor 4
S.O.M. (%) 0.9478 0.0367 −0.0019 −0.0184pH 0.4105 0.3419 −0.3033 −0.3910E.C.. −0.3833 −0.1732 0.0529 0.7790AW 0.4090 0.1671 −0.1742 0.6778Clay (%) −0.0117 0.8047 −0.2232 −0.1202N (%) 0.9269 0.1277 −0.0569 −0.0653P (mg kg−1) 0.0130 0.8687 0.0187 0.0599K (mg kg−1) 0.3857 0.7774 −0.1169 −0.0446Pb (A. regia) −0.2195 −0.2071 0.8698 0.0966Pb (DTPA) 0.0981 −0.0366 0.8828 −0.1203
Significant loadings in PCA factors are shown in bold.
Table 3Statistical summary of relative abundance (as a percentage of total nematode abundance) of 81agricultural areas.
Nematode taxa Functional guild Biomass (μg) Mean Median SD
Bacterial feeders 24.79 23.53 15Diplogasteridae b1 1.29 0.02 0.00 0Panagrolaimus b1 0.60 0.25 0.00 1Rhabditidae b1 5.32 2.46 0.89 4Monhystera b2 1.19 0.97 0.00 2Acrobeles b2 0.82 2.51 0.00 6Acrobeloides● b2 0.15 7.64 5.00 8Cephalobidae b2 0.40 0.61 0.00 2Cephalobus b2 0.27 3.10 1.27 5Cervidelus b2 0.16 0.25 0.00 1Chiloplacus b2 0.53 3.38 1.27 5Eucephalobus b2 0.26 0.53 0.00 1Nothacrobeles b2 0.61 0.04 0.00 0Pseudoacrobeles b2 0.30 0.04 0.00 0Seleborca b2 0.41 1.17 0.00 3Stegelleta b2 0.12 0.02 0.00 0Stegelletina b2 0.12 0.18 0.00 1Zeldia b2 0.70 0.03 0.00 0Plectus b2 1.01 0.23 0.00 0Heterocephalobus b2 0.36 0.65 0.00 3Drilocephalus b2 0.10 0.10 0.00 0Tylocephalus b2 0.21 0.28 0.00 0Chronogaster b3 0.35 0.24 0.00 1Cylindrolaimus b3 0.45 0.00 0.00 0Odontolaimus b3 0.16 0.00 0.00 0Teratocephalus b3 0.09 0.00 0.00 0Alaimus b4 0.53 0.07 0.00 0Fungal feeders 30.15 29.41 17Aphelenchoides● f2 0.16 6.25 3.03 10Aphelenchus●● f2 0.23 10.67 8.65 11Aputrides f2 0.06 0.02 0.00 0Paraphelenchus f2 0.36 0.13 0.00 0Ditylenchus f2 0.59 3.20 1.61 4Filenchus●● f2 0.10 9.77 7.58 11Diphterophora f3 0.50 0.01 0.00 0Dorylium f4 0.46 0.00 0.00 0Tylencholaimellus f4 0.87 0.09 0.00 0Omnivores 21.32 18.56 15Crassolabium o4 1.30 0.50 0.00 1Dorylaimidae o4 8.67 2.03 0.00 4Microdorylaimus o4 0.20 4.18 1.94 6Pungentus o4 1.88 0.02 0.00 0Eudorylaimus● o4 3.34 5.73 2.94 9Labronema o5 7.67 0.07 0.00 0Ecumenicus o5 0.71 4.13 1.04 8Mesodorylaimus o5 1.48 1.78 0.00 3Aporcelaimellus o5 8.68 2.88 1.52 4Predators 1.68 0.00 2Seinura p2 0.29 0.03 0.00 0Anatonchus p4 9.37 0.08 0.00 0Clarkus p4 4.39 0.33 0.00 1lotonchus p4 7.45 0.02 0.00 0Miconchus p4 4.80 0.02 0.00 0Mononchidae p4 5.16 0.01 0.00 0Mylonchulus p4 1.74 0.01 0.00 0
522 J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
clay content was related mainly to retained metals (Rodríguez Martínet al., 2007; Tack et al., 2005). Our data indicate clay values of 2.4–41.6% with a significant high mean value of 20.7% in agricultural soils.The mining soils with 60.4% sand were significantly lighter than arableor natural ones, whichmay improve metal mobilisation. Significant dif-ferences were observed for N, P and K contents, with 10 timesmore P inarable soils (32.2 mg kg−1) than inmining soils (3.1 mg kg−1), where-as the K values were 4 times higher (485 mg kg−1) than inmining soils(133 mg kg−1). The high available P and N contents are justified by thelarge amount of mineral fertilisers used in agricultural crops. Theseaspects are favoured by high pH and the abundance of carbonates,which limit P solubility (Gil et al., 2004).
The lead concentration means in the study area (5202 mg kg−1 intotal content and 262 mg kg−1 DTPA) present high values (Table 1).
soil nematode taxa, functional guilds, biomass and trophic groups in natural, mining and
Min. Max. Natural (76) Mining (66) Agricultural (51)
.69 0.00 100.00 23.87 (b) 17.7 (a) 35.50 (c)
.17 0.00 2.05 0.02 (a) 0.03 (a) 0.00 (a)
.04 0.00 10.00 0.00 (a) 0.46 (b) 0.36 (b)
.10 0.00 26.80 2.56 (ab) 1.98 (a) 2.93 (b)
.39 0.00 20.00 1.51 (b) 0.86 (a) 0.30 (a)
.57 0.00 66.67 2.98 (b) 1.01 (a) 3.74 (b)
.64 0.00 49.76 7.05 (b) 4.47 (a) 12.61 (c)
.70 0.00 29.43 0.52 (b) 0.65 (a) 0.70 (ab)
.66 0.00 49.91 2.46 (a) 3.78 (a) 3.20 (a)
.03 0.00 11.55 0.21 (a) 0.31 (a) 0.24 (a)
.91 0.00 44.59 2.78 (ab) 2.59 (a) 5.29 (b)
.72 0.00 16.51 0.69 (ab) 0.18 (a) 0.76 (b)
.27 0.00 3.06 0.07 (a) 0.00 (a) 0.05 (a)
.32 0.00 4.00 0.03 (a) 0.06 (a) 0.02 (a)
.26 0.00 31.21 1.03 (a) 0.50 (a) 2.27 (b)
.21 0.00 2.50 0.02 (a) 0.00 (a) 0.05 (a)
.12 0.00 11.76 0.22 (a) 0.22 (a) 0.06 (a)
.19 0.00 1.90 0.01 (a) 0.01 (a) 0.06 (a)
.81 0.00 7.69 0.40 (b) 0.10 (a) 0.14 (ab)
.31 0.00 31.86 0.19 (a) 0.04 (a) 2.13 (b)
.52 0.00 4.55 0.01 (a) 0.17 (a) 0.15 (a)
.82 0.00 5.62 0.45 (b) 0.07 (a) 0.30 (ab)
.04 0.00 7.04 0.46 (a) 0.17 (a) 0.03 (a)
.07 0.00 0.92 0.01 (a) 0.00 (a) 0.00 (a)
.07 0.00 0.92 0.01 (a) 0.00 (a) 0.00 (a)
.07 0.00 0.92 0.00 (a) 0.00 (a) 0.03 (a)
.45 0.00 5.10 0.09 (a) 0.01 (a) 0.10 (a)
.27 0.00 100.00 30.99 (a) 28.20 (a) 31.50 (a)
.08 0.00 100.00 6.63 (a) 7.22 (a) 4.42 (a)
.13 0.00 100.00 9.22 (a) 9.96 (a) 13.76 (a)
.25 0.00 3.30 0.04 (a) 0.00 (a) 0.02 (a)
.98 0.00 11.91 0.21 (a) 0.13 (a) 0.03 (a)
.29 0.00 25.06 3.02 (ab) 2.47 (a) 4.42 (b)
.09 0.00 100.00 11.64 (b) 8.47 (a) 8.66 (ab)
.11 0.00 1.19 0.02 (a) 0.00 (a) 0.02 (a)
.07 0.00 0.92 0.01 (a) 0.00 (a) 0.00 (a)
.55 0.00 5.50 0.11 (a) 0.00 (a) 0.16 (a)
.98 0.00 100.00 23.20 (b) 23.5 (ab) 15.70 (a)
.71 0.00 14.95 0.63 (ab) 0.19 (a) 0.71 (b)
.66 0.00 42.86 2.17 (a) 2.11 (a) 1.72 (a)
.98 0.00 51.07 5.43 (b) 4.13 (a) 2.37 (a)
.16 0.00 2.06 0.03 (a) 0.00 (a) 0.02 (a)
.41 0.00 100.00 6.63 (b) 5.74 (a) 4.39 (ab)
.56 0.00 7.50 0.16 (a) 0.00 (a) 0.02 (a)
.90 0.00 66.67 3.96 (a) 5.82 (a) 2.18 (a)
.94 0.00 22.73 1.57 (a) 2.68 (a) 0.93 (a)
.25 0.00 24.94 2.59 (ab) 2.87 (a) 3.34 (b)
.92 0.00 19.80 2.20 (b) 1.60 (a) 1.0 (a)
.29 0.00 3.80 0.07 (a) 0.00 (a) 0.00 (a)
.65 0.00 7.78 0.15 (a) 0.06 (a) 0.02 (a)
.53 0.00 17.82 0.56 (b) 0.30 (ab) 0.02 (a)
.22 0.00 2.66 0.01 (a) 0.02 (a) 0.05 (a)
.22 0.00 2.57 0.00 (a) 0.04 (a) 0.03 (a)
.13 0.00 1.47 0.03 (a) 0.00 (a) 0.00 (a)
.13 0.00 1.47 0.23 (a) 0.05 (a) 0.10 (a)

523J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
Moreover, 50% of the soil samples were higher than 2200 mg kg−1,with 30,000 mg kg−1 being recorded at some mining area sites. ThesePb contents can be considered to be polluted. Agricultural soil obtainedamean Pb level of 2633 mg kg−1, with 90% of the Pb contents above theupper reference value (137 mg kg−1) for Mediterranean soil (Roca-Perez et al., 2010). In Spain, the mean value of arable soils recordedwas 14 mg kg−1 (Nanos and Rodríguez Martín, 2012). The EuropeanPb mean (Adriano, 2001) for soil content (39 mg kg−1), and mostcontents demonstrated by several authors for soils in different countries(McDermott et al., 2011; Zhang et al., 1998), are lower than thosedescribed in the present study. In Granada (S Spain), Díez et al. (2009) re-corded amean value of 17.1 mg kg−1,with amaximumof 822 mg kg−1,in relation to the mining sediments deriving from the Sierra de Bazamountain range. Nevertheless, the high Pb contents recorded hereinagree with the results obtained by other authors for the La Unión miningdistrict: 4077–8016 mg kg−1 (Álvarez-Rogel et al., 2004); 4900–7900 mg kg−1 (Conesa et al., 2006); and 47,600 mg kg−1 (MartínezGarcía et al., 2001). It is well-known that the soil Pb in this area is relatedhighly with past mining activities. The relative Pb input contribution inthis area is a permanent risk. In line with this, McDermott et al. (2011)showed a high probability of intellectual disability (ID) in childrenwhen Pb was over 200 mg kg−1 (an 11% risk for ID).
The associations of Pb content and soil parameters (PCA) are shownin Table 2. Based on eigenvalues (eigen value N1), four main factors ex-plain 76.87% of the total variance. The first factor (F1), explaining 35% ofthe total variance, strongly and positively related to SOM and N, andpartly related to pH and soil AW. The second factor, explaining 16.93%of the total variance, showed high positive factor loadings on P, K andclay contents. The third factor (F3) accounted for 13.41% of the ex-plained variance, and is grouped, showing a high degree of covarianceto the total (aqua regia) and bioavailable (DTPA) lead contents. Thelast factor (F4), explaining 11.52% of the total variance, was related toAW and EC. However pH, which is strongly related to the retention ofmetals in the soil, presented a clear separation in the PCA, but showedpositive loadings on Factors 1 and 2, and negative loadings on Factors3 and 4. The highest loading on F1 of SOM, N, and partly on pH andAW, can be primarily ascribed to natural influences. These soil parame-ters showing the highest variance were related mainly to soil fertilityand availability of nutrients. Besides, Factor 2 was affected by severalanthropic and natural influences on a local scale. P and K levelsincreased through fertiliser use as part of the composition of manytreatments and amendments in agricultural soils (Gil et al., 2004;Ramos-Miras et al., 2011). However, soil clay content was present inthis grouping (F2), andwas attributed to agricultural location. Fig. 1 de-picts how the agricultural area is located in flat areas, corresponding tothe accumulation of finematerials. The present study indicates that totalPb and DTPA Pb are isolated and are not associated with other soil pa-rameters. Generally, soil Pb content is related to parent materials andis influenced by geochemical processes (Rodríguez Martín et al.,2013b). However it is possible to consider that, in the present study,Pb content can be anthropogenically influenced. Hence, it is feasible tobelieve that Factor 3 is attributed to geological and mining influences.
3.2. Characteristics of the nematode community
The study of the nematode soil food web assemblage is shown inTables 3 and 4. The nematofauna in the study area was highly diverse,although no entomopathogenic nematode populations were detected.Eighty-one taxa from 38 families were identified: bacterial feeders(26), fungal feeders (9), omnivores (9), predators (10) and plantfeeders (27), with 74 and 71 different taxa recorded in the natural andagricultural areas, respectively, but only 63 were found in the miningarea (Table 3). Although this large number of taxa indicates complex re-lations at trophic levels and provides the ecosystem with ecological re-silience (Peterson et al., 1998), the main effect observed was that mineactivities have significantly reduced nematode diversity, abundance and
biomass, which affects community structure (Tables 3 and 4). This nem-atode sensitivity to metals has been previously described for mine soils(Park et al., 2011; Shao et al., 2008).
Bacterial feeders have been generally related to disturbed areas. Inthis study however, where an extremely high mean Pb content wasdetected (5202 mg kg−1), the most abundant and dominant tro-phic groups were found in this order: fungal feeders N bacterialfeeders N omnivores N plant feeders N predators (Tables 3 and 4).These results agree with Pen-Mouratov et al. (2008), who recordedonly 249.05–62.98 mg kg−1 Pb. Furthermore, this result disagreeswith the results obtained by other authors (Park et al., 2011; Šalamúnet al., 2012; Shao et al., 2008), who showed greater abundance of bacte-rial feeders than fungal feeders with Pb contents of 249, 29 and15,000 mg kg−1, respectively.
We observed a differential response of fungal feeder genera toPb content. In agreement with other authors, Aphelenchoides andAphelenchus were the dominant and eudominant genera, respec-tively (6.25–10.67%), and can be considered insensitive and/ortolerant to heavy metal pollution because no significant differenceswere found amongst the areas (Korthals et al., 1996; Pen-Mouratovet al., 2008; Sánchez-Moreno et al., 2006). However, the dominantFilenchus genus (9.77%) was sensitive to mining and agriculturalactivities (8.47–8.66%) as compared to the natural areas (11.64%).The dominant (7.64%) bacterial feeder Acrobeloides, was significantlysensitive to mining activities (4.47%) vs. agricultural (12.63%) and nat-ural areas (7.05%), although it has been previously considered tolerantto metals (Šalamún et al., 2012). Furthermore, the subdominant bacte-rial feeders Chiloplacus (3.38%) and Acrobeles (2.51%) were also highlysensitive to mining activities with 2.59% and 1.01%, respectively, aspreviously reported by Pen-Mouratov et al. (2008). Nevertheless, inour work they were favoured by the agricultural area (5.29% and3.74%, respectively). The higher proportion of fungal feeders as com-pared to bacterial feeders in themining areamight be due to the positiveeffect of Pb content on fungal development, and it can be considered asign of heavy metal pollution, as other studies have suggested (Bongersand Bongers, 1998; Korthals et al., 1996).
Georgieva et al. (2002) showed that the genera with an r-strategycan be stimulated by metals, whereas those with a K-strategy candiminish drastically. In line with this, we also observed that omni-vores (o4–5) and predators (p4–5) were the most sensitive groupsto metal content. Nonetheless, our results show a differential effectfor mine soils in terms of their relative abundance. In general, pred-ators significantly reduced due to mining and agricultural activi-ties, whereas omnivores were affected more by agricultural soils.The o4 omnivores, the subdominant, Microdorilaimus (4.18%) andthe dominant Eudorylaimus (5.73%), were much more sensitive toagricultural soils (2.37 and 4.39%, respectively) than to mine soils(4.13 and 5.74%, respectively), which disagrees with Georgievaet al. (2002). Similarly, the o5 subdominant, Ecumenicus (4.13%),was the most abundant in mine soils (5.82%) and was sensitive to ag-ricultural soils (2.18%), but the differences found were not significant. Allthe predator generawere subdominant (0.01–0.42%), and themost abun-dant genera were: Nygolaimus N Clarkus N Discolaimoides N Discolaimus.Although sonewhat significant differences were observed, they were allless abundant in agricultural soils, with p4 beingmore sensitive tometalsthan p5, which disagrees with Georgieva et al. (2002) The relative abun-dance of plant-feeding nematodes did not significantly differ amongstsoils, with genera being subdominant (4.86–0.01%). Some authors (Pen-Mouratov et al., 2008; Šalamún et al., 2012) have reported that Tylenchus,Tylenchorhynchus and Pratylenchus are more abundant in contaminatedsoils. In agreement with those authors, only Pratylenchus (1.89%) wasfound to significantly abound in areas with highmetal contents, whereasHelicotylenchus (4.86%) was reduced in all soils.
The statistical summary of the nematode community structure, basedon its absolute abundance and metabolic footprint values, is provided inTable 4. The total abundance of individuals, trophic and c–p groups, as
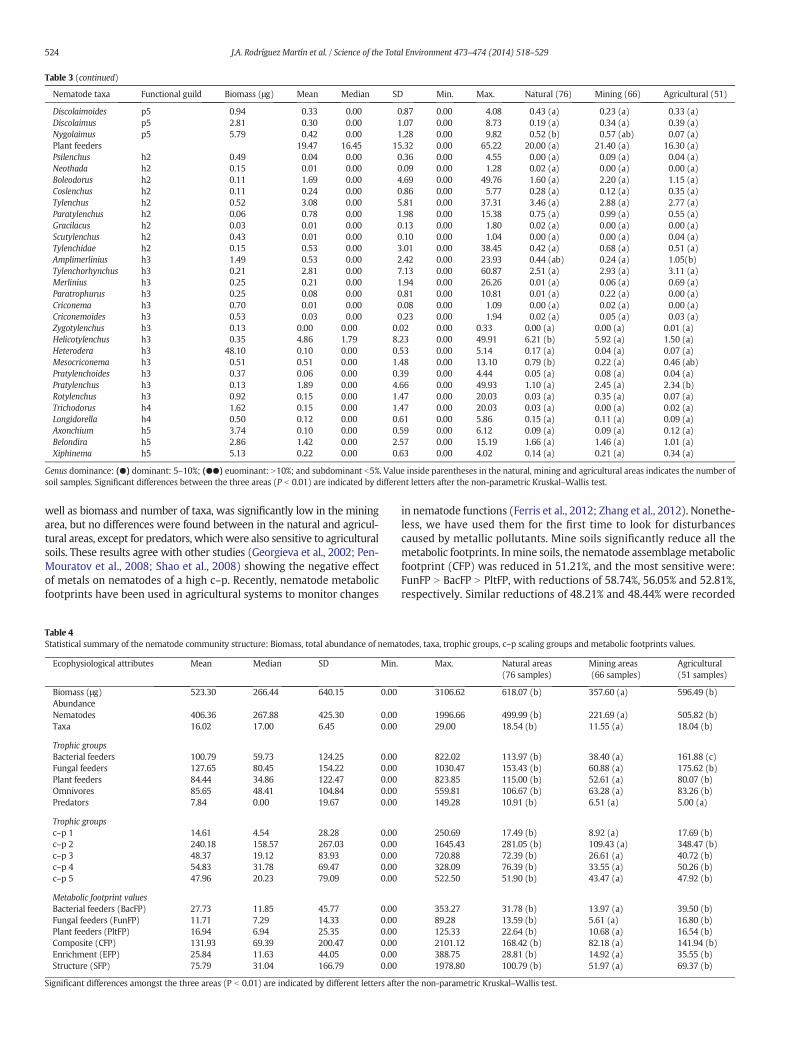
Table 3 (continued)
Nematode taxa Functional guild Biomass (μg) Mean Median SD Min. Max. Natural (76) Mining (66) Agricultural (51)
Discolaimoides p5 0.94 0.33 0.00 0.87 0.00 4.08 0.43 (a) 0.23 (a) 0.33 (a)Discolaimus p5 2.81 0.30 0.00 1.07 0.00 8.73 0.19 (a) 0.34 (a) 0.39 (a)Nygolaimus p5 5.79 0.42 0.00 1.28 0.00 9.82 0.52 (b) 0.57 (ab) 0.07 (a)Plant feeders 19.47 16.45 15.32 0.00 65.22 20.00 (a) 21.40 (a) 16.30 (a)Psilenchus h2 0.49 0.04 0.00 0.36 0.00 4.55 0.00 (a) 0.09 (a) 0.04 (a)Neothada h2 0.15 0.01 0.00 0.09 0.00 1.28 0.02 (a) 0.00 (a) 0.00 (a)Boleodorus h2 0.11 1.69 0.00 4.69 0.00 49.76 1.60 (a) 2.20 (a) 1.15 (a)Coslenchus h2 0.11 0.24 0.00 0.86 0.00 5.77 0.28 (a) 0.12 (a) 0.35 (a)Tylenchus h2 0.52 3.08 0.00 5.81 0.00 37.31 3.46 (a) 2.88 (a) 2.77 (a)Paratylenchus h2 0.06 0.78 0.00 1.98 0.00 15.38 0.75 (a) 0.99 (a) 0.55 (a)Gracilacus h2 0.03 0.01 0.00 0.13 0.00 1.80 0.02 (a) 0.00 (a) 0.00 (a)Scutylenchus h2 0.43 0.01 0.00 0.10 0.00 1.04 0.00 (a) 0.00 (a) 0.04 (a)Tylenchidae h2 0.15 0.53 0.00 3.01 0.00 38.45 0.42 (a) 0.68 (a) 0.51 (a)Amplimerlinius h3 1.49 0.53 0.00 2.42 0.00 23.93 0.44 (ab) 0.24 (a) 1.05(b)Tylenchorhynchus h3 0.21 2.81 0.00 7.13 0.00 60.87 2.51 (a) 2.93 (a) 3.11 (a)Merlinius h3 0.25 0.21 0.00 1.94 0.00 26.26 0.01 (a) 0.06 (a) 0.69 (a)Paratrophurus h3 0.25 0.08 0.00 0.81 0.00 10.81 0.01 (a) 0.22 (a) 0.00 (a)Criconema h3 0.70 0.01 0.00 0.08 0.00 1.09 0.00 (a) 0.02 (a) 0.00 (a)Criconemoides h3 0.53 0.03 0.00 0.23 0.00 1.94 0.02 (a) 0.05 (a) 0.03 (a)Zygotylenchus h3 0.13 0.00 0.00 0.02 0.00 0.33 0.00 (a) 0.00 (a) 0.01 (a)Helicotylenchus h3 0.35 4.86 1.79 8.23 0.00 49.91 6.21 (b) 5.92 (a) 1.50 (a)Heterodera h3 48.10 0.10 0.00 0.53 0.00 5.14 0.17 (a) 0.04 (a) 0.07 (a)Mesocriconema h3 0.51 0.51 0.00 1.48 0.00 13.10 0.79 (b) 0.22 (a) 0.46 (ab)Pratylenchoides h3 0.37 0.06 0.00 0.39 0.00 4.44 0.05 (a) 0.08 (a) 0.04 (a)Pratylenchus h3 0.13 1.89 0.00 4.66 0.00 49.93 1.10 (a) 2.45 (a) 2.34 (b)Rotylenchus h3 0.92 0.15 0.00 1.47 0.00 20.03 0.03 (a) 0.35 (a) 0.07 (a)Trichodorus h4 1.62 0.15 0.00 1.47 0.00 20.03 0.03 (a) 0.00 (a) 0.02 (a)Longidorella h4 0.50 0.12 0.00 0.61 0.00 5.86 0.15 (a) 0.11 (a) 0.09 (a)Axonchium h5 3.74 0.10 0.00 0.59 0.00 6.12 0.09 (a) 0.09 (a) 0.12 (a)Belondira h5 2.86 1.42 0.00 2.57 0.00 15.19 1.66 (a) 1.46 (a) 1.01 (a)Xiphinema h5 5.13 0.22 0.00 0.63 0.00 4.02 0.14 (a) 0.21 (a) 0.34 (a)
Genus dominance: (●) dominant: 5–10%; (●●) euominant: N10%; and subdominant b5%. Value inside parentheses in the natural, mining and agricultural areas indicates the number ofsoil samples. Significant differences between the three areas (P b 0.01) are indicated by different letters after the non-parametric Kruskal–Wallis test.
524 J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
well as biomass and number of taxa, was significantly low in the miningarea, but no differences were found between in the natural and agricul-tural areas, except for predators, whichwere also sensitive to agriculturalsoils. These results agree with other studies (Georgieva et al., 2002; Pen-Mouratov et al., 2008; Shao et al., 2008) showing the negative effectof metals on nematodes of a high c–p. Recently, nematode metabolicfootprints have been used in agricultural systems to monitor changes
Table 4Statistical summary of the nematode community structure: Biomass, total abundance of nema
Ecophysiological attributes Mean Median SD Min.
Biomass (μg) 523.30 266.44 640.15 0.00AbundanceNematodes 406.36 267.88 425.30 0.00Taxa 16.02 17.00 6.45 0.00
Trophic groupsBacterial feeders 100.79 59.73 124.25 0.00Fungal feeders 127.65 80.45 154.22 0.00Plant feeders 84.44 34.86 122.47 0.00Omnivores 85.65 48.41 104.84 0.00Predators 7.84 0.00 19.67 0.00
Trophic groupsc–p 1 14.61 4.54 28.28 0.00c–p 2 240.18 158.57 267.03 0.00c–p 3 48.37 19.12 83.93 0.00c–p 4 54.83 31.78 69.47 0.00c–p 5 47.96 20.23 79.09 0.00
Metabolic footprint valuesBacterial feeders (BacFP) 27.73 11.85 45.77 0.00Fungal feeders (FunFP) 11.71 7.29 14.33 0.00Plant feeders (PltFP) 16.94 6.94 25.35 0.00Composite (CFP) 131.93 69.39 200.47 0.00Enrichment (EFP) 25.84 11.63 44.05 0.00Structure (SFP) 75.79 31.04 166.79 0.00
Significant differences amongst the three areas (P b 0.01) are indicated by different letters afte
in nematode functions (Ferris et al., 2012; Zhang et al., 2012). Nonethe-less, we have used them for the first time to look for disturbancescaused by metallic pollutants. Mine soils significantly reduce all themetabolic footprints. Inmine soils, the nematode assemblagemetabolicfootprint (CFP) was reduced in 51.21%, and the most sensitive were:FunFP N BacFP N PltFP, with reductions of 58.74%, 56.05% and 52.81%,respectively. Similar reductions of 48.21% and 48.44% were recorded
todes, taxa, trophic groups, c–p scaling groups and metabolic footprints values.
Max. Natural areas(76 samples)
Mining areas(66 samples)
Agricultural(51 samples)
3106.62 618.07 (b) 357.60 (a) 596.49 (b)
1996.66 499.99 (b) 221.69 (a) 505.82 (b)29.00 18.54 (b) 11.55 (a) 18.04 (b)
822.02 113.97 (b) 38.40 (a) 161.88 (c)1030.47 153.43 (b) 60.88 (a) 175.62 (b)823.85 115.00 (b) 52.61 (a) 80.07 (b)559.81 106.67 (b) 63.28 (a) 83.26 (b)149.28 10.91 (b) 6.51 (a) 5.00 (a)
250.69 17.49 (b) 8.92 (a) 17.69 (b)1645.43 281.05 (b) 109.43 (a) 348.47 (b)720.88 72.39 (b) 26.61 (a) 40.72 (b)328.09 76.39 (b) 33.55 (a) 50.26 (b)522.50 51.90 (b) 43.47 (a) 47.92 (b)
353.27 31.78 (b) 13.97 (a) 39.50 (b)89.28 13.59 (b) 5.61 (a) 16.80 (b)125.33 22.64 (b) 10.68 (a) 16.54 (b)2101.12 168.42 (b) 82.18 (a) 141.94 (b)388.75 28.81 (b) 14.92 (a) 35.55 (b)1978.80 100.79 (b) 51.97 (a) 69.37 (b)
r the non-parametric Kruskal–Wallis test.

Fig. 2. Ordination diagram based on the CCorA of the soil environmental variables (F1, F2, F3 and F4) and a) data for the absolute abundance of nematodes (TN: total nematodes, B: bac-terial feeders, F: fungal feeders, H: plant feeders, O: omnivores and P: predatory nematodes); b) the c–p values that ranged from coloniser (c–p1) to persistor (c–p5); c) metabolic foot-prints (EFP: enrichment footprint, SFP: structure footprint, CFP: composite footprint, and bacterial, fungal and plant feeders footprints (BacFP, FunFP and PltFP).
Fig. 3. The experimental variogram and spatial models for soil factors (F1, F2, F3 and F4) and metabolic footprints: enrichment footprint (EFP), structure footprint (SFP), and bacterial,fungal and plant feeders footprints (BacFP, FunFP and PltFP).
525J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529

Table 5Semivariogram models for the soil factors and nematode footprint values.
Soil Factors (in PCA analysis) Some indices of nematode community
Nugget(Co)
Sill(Co + C)
Range(Ao)
Nugget/Sill(Co/Co + C)
Nugget(Co)
Sill(Co + C)
Range(Ao)
Nugget/Sill(Co/Co + C)
F1 3.5 7.1 2000 0.49 SFP 2520 8050 2300 0.31F2 1.0 9.8 14,000 0.10 EFP 570 1950 2100 0.29F3 2.5 3.8 4000 0.65 BFP 750 2120 2300 0.35F4 0.85 1.8 800 0.46 FuFP 100 190 4000 0.53
PltFP 225 670 3000 0.34
Co/(Co + C): proportion of spatial structure. SFP, EFP, BacFP, FunFP, PltFP: structure footprint, enrichment footprint, and bacterial, fungal and plant feeders footprints. Range (Ao) inmetres.
526 J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
for EFP, based mainly on bacterial and fungal feeders with c–p1 and c–p2, and SFP, basedmainly on nematodeswith high c–p values. In the ag-ricultural area, BacFP, FunFP and EFP increased within a 23.30–24.30%range. Although available Pb contentwas 2 times lower than in themin-ing area, PlFP and SFP (corresponding to herbivores and omnivores–predators, respectively) were sensitive to agricultural practices, andwere reduced by 26.93% and 31.80%, respectively. Ferris et al. (2012)obtained similar results for BacFP and EFP in fields used for vegetableproduction, but no differences were found for FunFP, probably due tothe unsuitable conditions for fungal growth caused by these agriculturalpractices.
Fig. 4. Grid contour map of the four soil factors (F1, F2, F3 and F4) and the principal metabolicplant feeders footprints (BacFP, FunFP and PltFP).
3.3. Quantifying relationships between nematode community structure andsoil factors
The CCorA resultswere plotted separately for the three ecological at-tributes: total abundance of individuals and trophic groups (Fig. 2a), c–pvalues (Fig. 2b) and metabolic footprints (Fig. 2c). The first analysisproved highly significant with Factors 1 and 2 representing 92% of thetotal variance (Fig. 2a), and it showed that total abundance of nema-todes and trophic groups are negatively influenced by high lead content(Factor 3), and also by EC andwater retention capacity (AW) (Factor 4),which are involved in metal retention. This result is in accordance with
footprints: enrichment footprint (EFP), structure footprint (SFP), and bacterial, fungal and

527J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
other studies into heavy metal effects on nematode communities (Hösset al., 2011; Šalamún et al., 2012; Shao et al., 2008). Bacterial feeders(Bacf) and plant feeders (Pltf) have been considered to be the mostresistant to adverse pollution effects (Šalamún et al., 2012). However,the present study reveals that metal content affected Bacf than Pltfmore negatively. This effect agrees with Bongers and Ferris (1999),who stated that the abundance of plant-feeding nematodes is largelydetermined by community structure, biomass and vigour of plants.Other studies (Park et al., 2011; Zhang et al., 2006) have reported theopposite effect on the proportion of bacteriovorus and plant feeders inthe soil nematode community. This contradiction can be explained bythe different physical and chemical properties of soil, such as soil nutri-ents, SOM or soil texture (Šalamún et al., 2012). Along these lines, weherein show that Bacf was more positively influenced by soil Factor 2(P, K and clay) than by Factor 1 (SOM, N), whereas contrary relation-ships were observed for Pltf. Omnivores (Om) and predators (Pred)(with a K-strategy-type life history) have generally been consideredthe most sensitive to disturbances, including heavy metal pollution(Pen-Mouratov et al., 2008; Šalamún et al., 2012). Yet in the presentstudy, both were influenced more by SOM, N, clay, P and K contents(Factors 1 and 2) than lead content (Factor 3), where predators werethe least influenced, probably due to the highest trophic level. Thepoor sensitivity to lead content of both groups may be because theyhave naturally adapted to high levels of metals in the study area.
The second CCorA (Fig. 2b) explored the soil factors ~c–p grouprelationship as abundance of nematodeswith different life history strat-egies and sensitivities to enrichment and disturbance. In these analyses,Factors 1 and 2 represented 88% of the total variance. The c–p3–5groups (K-strategy-type life history), including omnivores and preda-tors, have been considered to be highly sensitive to environmental dis-turbance (Bongers and Ferris, 1999; Pen-Mouratov et al., 2008; Šalamúnet al., 2012; Shao et al., 2008). According to Park et al. (2011), all the c–pgroupswere negatively affected byhigh Pb content. Yetwe show that c–p3, c–p4 and c–p5 were less negatively related to Pb content (Factor 3)than the c–p1 and c–p2 groups, these being opportunist nematodeswith short life cycles and high fecundity, which are mainly bacterialand fungal feeders that respond positively to enrichment of resourcesand are considered resistant to contamination. The sensitivity of c–p1and 2 to Pb might be due to the toxic effect of Pb on soil microflora(Hu et al., 2007). Our results agree with Sánchez-Moreno et al. (2006),who found that the c–p2 nematode was affected negatively by Pb con-tamination in the Guadiamar river basin (S Spain), andwho also provedwith laboratory tests Pb toxicity against two c–p2 species: Aphelenchusavenae and Cephalobus persegnis (fungal and bacterial feeders, respec-tively). Similarly, c–p1 and 2 were influenced positively by Factors 1and 2 in accordance with their response to enrichment conditions insoil. c–p3 to c–p5 responded in the same way as trophic groups Pltf,Om and Pred, and related more positively to nutrients and organicmatter.
Themetabolic footprint analysis by CCorA (Fig. 3c) can be interpretedin the light of metabolic activity and indicated that this ecosystem's func-tions are associated with different soil parameters and lead pollutionlevels. The first two factors in the multivariate analysis were significant,representing 83% of the total variance and showing that metabolic activ-ity negatively relates to lead content,with FunFP being themost sensitive.Soil Factor 4 (AW, EC)was also negatively related tometabolic footprints,except for BacFP and EFP, which were weakly, but positively, related tosoil Factor 4. All the metabolic footprints positively related to soil Factors1 and 2, but showed different trends. BacFP, FunFP and EFPwere affectedsimilarly by SOM, N (Factor 1) and clay, P, K (Factor 1), which agreeswithprevious studies (Ferris et al., 2012; Zhang et al., 2012). These authorsrecorded enhanced levels of these footprints as a response to organic re-sources after cover crops or residue incorporation treatments in agricul-tural fields. These findings indicated the entry of resources into the soilfood web to represent the ecosystem's nutrient mineralisation service.In our study, SFP and PltFP were more closely related to SOM, N (Factor
1) than to clay, P, K (Factor 2). Both the above-cited authors also recordeda slight increase of PltFP as a response to increased resources, whereasSFP was enhanced by cover crops and residue incorporation, but washighly sensitive to frequent tillage events in agricultural soils. In short,the footprints based on abundance andmetabolic activity aremore sensi-tive for soil disturbances than other nematode indicators (Ferris, 2010).Hence we selected them to study their spatial relationship with soilfactors by geostatistics analysis techniques.
3.4. Geostatistics analysis and spatial relationships
The attributes of semivariograms for soil factors and footprints aresummarised in Table 5. We selected: the Enrichment footprint (EFP),corresponding to the opportunist nematodes that are rapidly respon-sive to resource enrichment and quantify the ecosystem nutrientmineralisation service; the bacterial, fungal and plant feeder footprints(BacFP, FunFP and PltFP) to assess the main channel that contributesto nutrient mineralisation; the structure footprint (SFP), correspondingto the higher trophic level of nematodes, which perform a regulatoryfunction in the food web (Ferris, 2010), to describe the spatial patternsof structure and connections for soil environmental variables. A strongspatial dependence of soil properties can be usually attributed to intrin-sic properties (such as lithology), whilst weak spatial dependence maybe attributed to extrinsic aspects like pollution (Liu et al., 2004;Rodríguez Martín et al., 2013a). The Nugget/Sill ratios range from 0.10for Factor 2 to 0.65 for Factor 3 in soil, and from 0.29 for EFP to 0.53for FunFP in the soil nematodes bioindicators (Table 5). This range isconsidered to be the distance beyond which observations are not spa-tially dependent. This range of influence was greater in F2 than in theother soil factors. For the study area, Factor 2, which mainly representsthe P and K levels in soil and clay content variation, presented a range of14 km with a typical agricultural influence in these arable lands(Rodríguez Martín et al., 2013b). However, F1, F3 and F4 presented ashorter range (2 km, 4 km, and 0.8 km, respectively). Basically, the fac-tors presenting variability that can be attributed to a lithologic compo-nent or edaphic characteristics presented a shorter range in variogramadjustments. Normally, a wide range expresses a major area of influ-ence, which is attributed to intrinsic properties (Goovaerts, 1997; Liuet al., 2004; Rodríguez Martín et al., 2009), such as parent material; ifthe distance amongst samples, considered to be related, is short, it isassociated with extrinsic aspects (e.g., pollution), which are more spo-radic. In the present study, this general aspect of the analysis of these at-tributions contrastedwith the results obtained; Pb (soil Factor 3),whichwas related to human activities and was considered to be a pollutingkind, not only represented greater continuity, but also reflected a largerscale of influence.We can ascribe this spatial continuity to the longmin-ing activity period (over 2500 years).Moreover, the nematode footprintindicators displayed ranges from 2100 m to 4000 m (Fig. 3), and theirdistances were more homogeneous than the distances of the soil diver-sity indicators. This fact indicates that the effect of the environment onthese biological indicators is limited to soil processes, whose influencereaches distances of less than 5 km on a short spatial scale.
The experimental semivariograms (Fig. 3) suggest that the theoreti-cal spherical isotropic model is in reasonable agreement with the data.The theoreticalmodelwas interpolatedwith OK for the selected soil fac-tors and nematode footprint indicators. In general, the areas with highcontents of soil nutrients (SOM, N, P, and K, related to soil factors F1and F2) corresponded to agricultural and forest lands, which presentedabundance and broad diversity of nematodes (Fig. 4). The soil Factor 3map explains wider Pb variation with significant anthropic input, andreflects the long mining activity period in the Sierra Minera district.On a long spatial scale, it is easier to note the natural component orthe lithological domain, and the highest Factor 3 values (total andDTPA Pb contents) reflect the influence of vast mineral wealth associat-ed with accumulations of metals (Pb, Zn and Fe) in the area, as men-tioned in Section 3.1. However, the fact that a large area of high lead

528 J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
content is located in the agricultural area, close to the Mar Menor, isnoteworthy (Fig. 4). This result agrees with Álvarez-Rogel et al. (2004)(Álvarez-Rogel et al., 2004), who previously reported high lead concen-trations (8000 mg/kg), which have been associated with the impacts ofmining activities when polluted sediments are carried away by riversand runoffs. Thus the site can be considered a strongly polluted area.
On the whole, the analysis of the soil factor spatial patterns in thestudy area showed three regions according to land uses. The central re-gion corresponded to the modern mining area, which remains un-changed and is clearly delimited by high Pb contents (Fig. 4), and alsoby the low values of the edaphic parameters included in Factors 1 and2. This result suggests a geographical interaction phenomenon. Thusthe areas with low nutrient contents also present high lead pollution.The heavymetal pollution induced by pastmining activities has had ad-verse effects on the soil environment. The metabolic footprints, used asan indicator of soil health, showed significant spatial differences (main-ly the Enrichment FP). The reduced nematode population in soil con-taminated with high Pb concentrations comes over clearly in Fig. 4,where the metal-polluted plots increased.
4. Conclusions
High lead concentrations have been quantified in the La Union min-ing area due to the long mining activity period, which was expected.However, these high levels of lead have not only been recently recordedfrom mining areas, but have also been detected in the soils of the adja-cent agricultural area. Given their concentration, they can pose a risk ifcrops accumulate lead beyond health safety levels.
Broad nematode diversity is observed in the study area. However,soil disturbances induce changes in their communities. Nematodeabundance, number of taxa, and consequently biomass, as well as thestructure and function attributes of nematode communities, are signifi-cantly reduced in polluted areas. Although these were the main effects,significant differential responses from soil factors were recorded. Inter-estingly nematode functional guilds, b1–2 and f2, have been previouslyrelated to soil disturbances and enrichment conditions, and described asbeing tolerant/resistant to pollution. They were most sensitive to themining area, whilst fungal feeders were more abundant than bacterialfeeders, probably due to the soil conditions which promote mainly fun-gal growth. Likewise, guilds o4–5 and p4–5, described as being highlysensitive to pollution, were more tolerant in our study than expected.The metabolic footprints, used for the first time to assess nematode re-sponses in a highly polluted area, showed a similar response patternwith accurate sensitivity to soil physico-chemical changes.
One interesting approach of this study was to utilise the statistictechniques of multivariate and geostatistic analyses to study the spatialpatterns of nematode community structure and connection with soilfactors. Lead from past mining activities has a serious impact on theabundance and diversity of nematodes of the Cartagena–La Unión dis-trict soils. However, soil organic matter and nutrient (NPK) contentsare actually the most influential factors on soil nematodes. We believethat the soil nematode communities in this study area are well-adapted to high Pb concentrations, and that the effect on their popula-tion levels can be due to the soil conditions in the mining area ratherthan to the toxic effect owing to lead pollution.
Acknowledgements
We gratefully acknowledge the funding provided by the SpanishMinistry of Innovation through Projects CGL2009-14686-C02-01 andCGL2009-14686-C02-02, as well as to the EIADES and CARESOILProgrammes supported by the Madrid Community to promote collabo-rations amongst researchers. R. Campos-Herrera was supported by aMarie Curie International Outgoing Fellowship (FP7-PEOPLE-2009-IOF-252980). The authorswish to thank Francisco Artero,Miguel Juanco
andManuel Fernández for their technical assistance, and they are grate-ful to Dr. Pedro Sánchez-Gómez for botanical advice.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.scitotenv.2013.12.075.
References
Adriano DC. Trace elements in terrestrial environments: biogeochemistry, bioavailability,and risks of metals. New York: Springer Verlag; 2001.
Álvarez-Rogel J, Aparicio MJR, Iniesta MJD, Lozano RA. Metals in soils and above-groundbiomass of plants from a salt marsh polluted by mine wastes in the coast of theMar Menor Lagoon, SE Spain. Fresen. Environ Bull 2004;13:274–8.
Alloway BJ. Heavy metals in soils. Netherlands: Blackie Academic & Professional; 1995.Andrassy I. Die rauminhalts-und gewichtsbestimmung der fadenwürmer (Nematoden).
Acta Zool Hung 1956;2:1–5.Andrássy I. Free-living nematodes of Hungary (Nematoda errantia). Pedozool Hung
2005;3:518.Benedicto J, Martínez-Gómez C, Guerrero J, Jornet A, Rodríguez C. Metal contamination in
Portman Bay (Murcia, SE Spain) 15 years after the cessation of mining activities.Cienc Mar 2008;34:389–98.
Bongers T. De Nematoden van Nederland: Een identificatietabel voor de in Nederlandaangetroffen zoetwater-en bodembewonende nematoden: Koninklijke NederlandseNatuurhistorische Vereiniging; 1988.
Bongers T. The maturity index: an ecological measure of environmental disturbancebased on nematode species composition. Oecologia 1990;83:14–9.
Bongers T, Bongers M. Functional diversity of nematodes. Appl Soil Ecol 1998;10:239–51.Bongers T, Ferris H. Nematode community structure as a bioindicator in environmental
monitoring. Trends Ecol Evol 1999;14:224–8.Bush AO, Lafferty KD, Lotz JM, Shostak AW. Parasitology meets ecology on its own terms:
Margolis et al. revisited. J Parasitol 1997:575–83.Conesa HM, Faz Á, Arnaldos R. Heavy metal accumulation and tolerance in plants from
mine tailings of the semiarid Cartagena–La Unión mining district (SE Spain). SciTotal Environ 2006;366:1–11.
Díez M, Simón M, Martín F, Dorronsoro C, García I, Van Gestel CAM. Ambient trace ele-ment background concentrations in soils and their use in risk assessment. Sci TotalEnviron 2009;407:4622–32.
Ferris H. The role of nematodes in soil fertility [Online]. NEMAPLEX: the Nematode–PlantExpert Information System. A virtual encyclopedia on soil and plant nematodes. Uni-versity of California; 1998.
Ferris H. Form and function: metabolic footprints of nematodes in the soil food web. Eur JSoil Biol 2010;46:97–104.
Ferris H, Bongers T, de Goede RGM. A framework for soil food web diagnostics: extensionof the nematode faunal analysis concept. Appl Soil Ecol 2001;18:13–29.
Ferris H, Sánchez-Moreno S, Brennan EB. Structure, functions and interguild relationshipsof the soil nematode assemblage in organic vegetable production. Appl Soil Ecol2012;61:16–25.
Garcia G, Peñas JM, Manteca JI. Zn mobility and geochemistry in surface sulfide miningsoils from SE Spain. Environ Res 2008;106:333–9.
García Navarro F, Amorós Ortiz-Villajos J, Sánchez Jiménez C, Jiménez Ballesta R. Red soilgeochemistry in a semiarid Mediterranean environment and its suitability forvineyards. Environ Geochem Health 2011;33:279–89.
Georgieva SS, McGrath SP, Hooper DJ, Chambers BS. Nematode communities under stress:the long-term effects of heavy metals in soil treated with sewage sludge. Appl SoilEcol 2002;20:27–42.
Gil C, Boluda R, Ramos J. Determination and evaluation of cadmium, lead and nickel ingreenhouse soils of Almería (Spain). Chemosphere 2004;55:1027–34.
Goovaerts P. Geostatistics for natural resources evaluation. USA: Oxford University Press;1997.
Höss S, Claus E, Von der Ohe PC, Brinke M, Güde H, Heininger P, et al. Nematode species atrisk—a metric to assess pollution in soft sediments of freshwaters. Environ Int2011;37:940–9.
Houba V, Van der Lee J, Novozamsky I. Soil and plant analysis, a series of syllabi part 5B:soil analysis procedures, other procedures. Department of Soil Science and Plant Nu-trition: Agricultural University, Wageningen, The Netherlands; 1995.
Hu Q, Qi H-y, Zeng J-h, Zhang H-x. Bacterial diversity in soils around a lead and zinc mine.J Environ Sci 2007;19:74–9.
Kianifard F. Applied multivariate data analysis, volume ii: categorical and multivariatemethods. Technometrics 1993;35:326–7.
Korthals GW, Bongers T, Kammenga JE, Alexiev AD, Lexmond TM. Long-term effects ofcopper and ph on the nematode community in an agroecosystem. Environ ToxicolChem 1996;15:979–85.
Liu X, Xu J, Zhang M, Zhou B. Effects of land management change on spatial variability oforganic matter and nutrients in paddy field: a case study of Pinghu, China. EnvironManage 2004;34:691–700.
Martínez García MJ, Moreno-Grau S, Martínez García JJ, Moreno J, Bayo J, Guillén Pérez JJ,et al. Distribution of the metals lead, cadmium, copper, and zinc in the top soil ofCartagena, Spain. Water Air Soil Pollut 2001;131:329–47.
McDermott S, Wu J, Cai B, Lawson A, Marjorie Aelion C. Probability of intellectual disabil-ity is associated with soil concentrations of arsenic and lead. Chemosphere 2011;84:31–8.

529J.A. Rodríguez Martín et al. / Science of the Total Environment 473–474 (2014) 518–529
Nanos N, Rodríguez Martín JA. Multiscale analysis of heavy metal contents in soils:spatial variability in the Duero river basin (Spain). Geoderma 2012;189–190:554–62.
Navarro MC, Pérez-Sirvent C, Martínez-Sánchez MJ, Vidal J, Tovar PJ, Bech J. Abandonedmine sites as a source of contamination by heavy metals: a case study in asemi-arid zone. J Geochem Explor 2008;96:183–93.
Neher DA. Role of nematodes in soil health and their use as indicators. J Nematol 2001;33:161–8.
Okada H, Harada H, Kadota I. Fungal-feeding habits of six nematode isolates in the genusFilenchus. Soil Biol Biochem 2005;37:1113–20.
Okada H, Kadota I. Host status of 10 fungal isolates for two nematode species, Filenchusmisellus and Aphelenchus avenae. Soil Biol Biochem 2003;35:1601–7.
Park B-Y, Lee J-K, Ro H-M, Kim YH. Effects of heavy metal contamination from an aban-doned mine on nematode community structure as an indicator of soil ecosystemhealth. Appl Soil Ecol 2011;51:17–24.
Pen-Mouratov S, Shukurov N, Steinberger Y. Influence of industrial heavy metal pollutionon soil free-living nematode population. Environ Pollut 2008;152:172–83.
Peterson G, Allen CR, Holling CS. Ecological resilience, biodiversity, and scale. Ecosystems1998;1:6–18.
Plassard F, Winiarski T, Petit-Ramel M. Retention and distribution of three heavy metalsin a carbonated soil: comparison between batch and unsaturated column studies.J Contam Hydrol 2000;42:99–111.
Ramos-Miras J, Roca-Perez L, Guzmán-Palomino M, Boluda R, Gil C. Background levelsand baseline values of available heavy metals in Mediterranean greenhouse soils(Spain). J Geochem Explor 2011;110:186–92.
Roca-Perez L, Gil C, Cervera M, Gonzálvez A, Ramos-Miras J, Pons V, et al. Selenium andheavy metals content in some Mediterranean soils. J Geochem Explor 2010;107:110–6.
RodríguezMartín J, Vázquez de la Cueva A, Grau Corbí J, López AriasM. Factors controllingthe spatial variability of copper in topsoils of the northeastern region of the IberianPeninsula, Spain. Water Air Soil Pollut 2007;186:311–21.
Rodríguez Martín J, Vazquez de la Cueva A, Grau Corbí J, Martínez Alonso C, López AriasM. Factors controlling the spatial variability of mercury distribution in Spanish top-soil. Soil Sediment Contam 2009;18:30–42.
Rodríguez Martín JA, Carbonell G, Nanos N, Gutiérrez C. Source identification of soil mer-cury in the Spanish Islands. Arch Environ Contam Toxicol 2013a;64:171–9.
Rodríguez Martín JA, Ramos-Miras JJ, Boluda R, Gil C. Spatial relations of heavy metals inarable and greenhouse soils of a Mediterranean environment region (Spain).Geoderma 2013b;200–201:180–8.
Šalamún P, Renčo M, Kucanová E, Brázová T, Papajová I, Miklisová D, et al. Nematodes asbioindicators of soil degradation due to heavy metals. Ecotoxicology 2012;21:2319–30.
Sánchez-Moreno S, Camargo JA, Navas A. Ecotoxicological assessment of the impact ofresidual heavy metals on soil nematodes in the Guadiamar river basin (SouthernSpain). Environ Monit Assess 2006;116:245–62.
Shao Y, Zhang W, Shen J, Zhou L, Xia H, Shu W, et al. Nematodes as indicators of soilrecovery in tailings of a lead/zinc mine. Soil Biol Biochem 2008;40:2040–6.
Siddiqi MR. Tylenchida: parasites of plants and insects: CABI; 2001.Tack FMG, Vanhaesebroeck T, Verloo MG, Van Rompaey K, Van Ranst E. Mercury baseline
levels in Flemish soils (Belgium). Environ Pollut 2005;134:173–9.Walkley A. An examination of methods for determining organic carbon and nitrogen in
soils. J Agric Sci 1935;25:598–609.Yeates G, Bongers T, De Goede R, Freckman D, Georgieva S. Feeding habits in soil nema-
tode families and genera—an outline for soil ecologists. J Nematol 1993;25:315.Yeates GW, Bongers T. Nematode diversity in agroecosystems. Agric Ecosyst Environ
1999;74:113–35.Zhang C, Selinus O, Schedin J. Statistical analyses for heavy metal contents in till and root
samples in an area of southeastern Sweden. Sci Total Environ 1998;212:217–32.Zhang X, Li Q, Zhu A, Liang W, Zhang J, Steinberger Y. Effects of tillage and residue man-
agement on soil nematode communities in North China. Ecol Indic 2012;13:75–81.Zhang XK, Li Q, Wang SB, Jiang Y, Liang W. Effect of zinc addition to soil on nematode
community structure. Bull Environ Contam Toxicol 2006;76:589–94.




















