
Plasmid DNA damage caused by methylated arsenicals, ascorbic acid and human liver ferritin
Upload
independentCategory
view
0download
0
JOURNAL OF MEDICINAL FOODVolume 4 Number 2 2001Mary Ann Liebert Inc
Effect of Ascorbic Acid and Its Hydrophobic DerivativePalmitoyl Ascorbate on the Redox State of Primary
Human Fibroblasts
GENNADY ROSENBLAT1 MARTIN F GRAHAM2 ADI JONAS1 MARK TARSHIS3
SHAY YEHOSHUA SCHUBERT1 MINA TABAK1 and ISHAK NEEMAN1
ABSTRACT
Ascorbic acid (AA) and its derivatives participate in vitro in oxidative-reductive reactions bothas antioxidants and as prooxidants The physiological relevance of these prooxidant effects ofAA and its derivatives remains unclear There is little evidence that AA can initiate formationof reactive oxygen species (ROS) or lipid peroxidation in cells or tissue In order to examine theeffect of AA and its derivative palmitoyl ascorbate on in situ intracellular ROS production andlipid peroxidation 2 9 7 9 -dichlorofluorescin diacetate (DCFH-DA) and cis-parinaric acid wereused as fluorescent probes in cultural neonatal foreskin fibroblasts The results demonstratedthat the effect of AA depended on the in vitro growth conditions AA induced augmentationof the intracellular ROS concentration in newly plated (24 hours) cells However in cells cul-tured for 72 hours AA had a different effect it moderately reduced intracellular ROS concen-tration but stimulated lipid peroxidation in the cytoplasmic membrane Palmitoyl ascorbatedemonstrated significant inhibition of intracellular DCFH-DA oxidation presumably caused byinhibition of reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase
107
INTRODUCTION
THE BALANCE BETWEEN oxidized and reducedcompounds in a cell (redox state) is an im-
portant characteristic of the cellrsquos conditionUnder physiological conditions the cellular re-dox state is characterized by a reducing envi-ronment in the cytosol A shift of the balanceto the oxidation side referred to as oxidativestress is usually associated with production ofhigh reactive oxygen species (ROS) The in-creased ROS generation is implicated in thepathogenesis of a wide variety of disorders1
and in the mechanism of programmed celldeath (apoptosis)2 where ROS may be in-volved through damage to cellular constituentsandor alteration of cell function and the in-termediates in signal transduction3 Oxidativechallenge to the cells comes from both exoge-nous agents including environmental pollu-tants and endogenous generation of ROS (egsuperoxide hydrogen peroxide) by mainly re-duced nicotinamide adenine dinucleotidephosphate (NADPH) oxidase complexes mito-chondria electron transport systems oxidationof purines and hypoxanthine by xanthine oxi-
1Department of Food Engineering and Biotechnology Technion-Israel Institute of Technology 32000 Haifa Israel2Interdepartmental Units The Hebrew University-Hadassah Medical School POB 12272 Jerusalem Israel3Laboratory of Tissue Repair Medical College of Virginia Campus of Virginia Commonwealth University Rich-
mond Virginia 23298
dase4 Antioxidants protect the cells against ex-cessive ROS generation Nevertheless the bio-logical effects of some antioxidants are para-doxically associated with their ability in vitro toundergo redox cycling to generate ROS
Ascorbic acid (AA) is involved in manyphysiological functions in living organismsincluding the regulation of collagen synthe-sis56 This activity may be explained by thecapacity of AA to participate in oxidative-re-ductive reactions both as an antioxidant andas a prooxidant depending on the local envi-ronment in which the molecule is present7
The antioxidant activity of AA and its deriv-atives is a result of its ability to react with avariety of free radicals and ROS AA readilyundergoes oxidation forming an intermediateradical of low reactivity that may account forthe antioxidant effect The prooxidant activityof AA is derived from its ability to reducetransition metals such as Fe13 and Cu12 andalso to reduce O2 The formation of severalROS in the presence of AA may be illustratedby the following reactions8
AscH2 1 Fe13(Cu12) RAsc2 1 Fe12(Cu1) 1 H1
Fe12(Cu1) 1 O2 R O22 1 Fe13(Cu12)
AscH2 1 O22 1 H1 R H2O2 1 Asc2
Fe12(Cu1) 1 H2O2 ROH2 1 OH 1 Fe13(Cu12)
In the reactions above AscH2 is the reducedform of ascorbic acid Asc2 is the semi-oxidizedascorbic acid radical OH2 is the hydroxyl anionand OH is the hydroxyl radical One hypothe-sis that has been proposed regarding the role ofAA in collagen synthesis is that AA stimulateslipid peroxidation with the formation of reactivealdehydes The aldehydes then upregulate ex-pression of the procollagen genes9ndash11 Despite alarge body of literature on the subject the phys-iological relevance of the prooxidative effect ofAA or its derivatives remains unclear A limitednumber of reports have demonstrated that ox-idative damage to DNA lipids and proteins isseen in cells treated with AA712 A major ques-tion still remains whether AA and its derivativesaffect cellular metabolism by augmentation or by
the opposite inhibition of intracellular ROS pro-duction
Our current interest in this subject focuses onthe in situ effect of physiologically active con-centrations of AA (in terms of stimulation ofcollagen synthesis) and of its hydrophobic de-rivative palmitoyl ascorbate (PA) on intracel-lular ROS generation and lipid peroxidation inthe cytoplasm membrane of cultured cells
In the present study we used two fluorescentldquoreporter moleculesrdquo to examine the intracel-lular production of ROS and lipid peroxidationin cultured neonatal foreskin fibroblasts or skinfibroblasts in response to AA and PA The firstmethod was based on the intracellular interac-tion of the nonfluorescent compound 2979-dichlorofluorescin diacetate (DCFH-DA) withROS (particularly hydrogen peroxide H2O2) toproduce the fluorescence product 2979-dichlo-rofluorescein (DCF) The second method mon-itored cis-parinaric acid fluorescence which isdecreased upon reaction with free radicals in alipid environment
MATERIALS AND METHODS
Cell culture
Dermal and foreskin fibroblasts were grownin culture from explants of human adult skinor neonatal foreskin Cells were grown in 75-mm2 plastic flasks (Corning Glass WorksCorning NY) in Dulbeccorsquos modified Eaglemedium (Biological Industries Kibbutz BeitHaemek Israel or Sigma Chemical Co StLouis MO) containing 4500 mgL glucose andsupplemented with 10 fetal calf serum (FCS)(Biological Industries or Bio-Whittaker Wal-tersville MA) in a humidified atmosphere of7 carbon dioxide Once confluent primarycultures were subcultured by trypsinizationand used between passages 3 and 18
Intracellular oxidation of DCFH-DA
Cells were seeded in two- or four-chamber tis-sue culture slides (Nalge Nunc InternationalNaperville IL) at densities of 10000ndash20000 cellsper well and incubated in a humidified atmos-phere for 24 or 72 hours at 37degC according to theprotocol described earlier Prior to the experi-ments with skin fibroblasts the medium was re-
ROSENBLAT ET AL108
moved and replaced with phosphate-bufferedsaline (PBS) supplemented with 5 mM DCFH-DA In experiments with foreskin fibroblasts thecells were adapted to room temperature for 20minutes before addition of DCFH-DA The ex-perimental samples were additionally supple-mented with AA or PA or hydrogen peroxideH2O2 at appropriate concentrations In other ex-periments DCFH-DA was added to the cells af-ter their pretreatment with AA for 2 hours fol-lowed by washing and replacement with PBSAs selected for the study the concentrations ofAA and PA that were optimal for collagen syn-thesis stimulation in cultured fibroblasts con-sisted of 100 mM and 10 mM respectively15
The intracellular formation of DCF was deter-mined by either an Ultima Imaging Cytometer(Biomedical Imaging Inc Okemos MI) or byconfocal microscopy using a Zeiss (Jena Ger-many) LSM 410 confocal laser scanning micro-scope The system was equipped with an air-cooled argon laser The fluorescence wascollected by employing a 63312 C-apochro-mate water immersion lens (Zeiss) Differentialinterference images according to Nomarski werecollected using a transmitted light detector
For the confocal microscopy experiments thecellular fluorescence values of four separate ar-eas on the slide were measured and divided bythe number of cells in each area For the cy-tometer experiments the fluorescence of threecells was measured
Monitoring of lipid peroxidation by cis-parinaricacid fluorescence
The examination of lipid peroxidation by cis-parinaric acid fluorescence was performed bytwo methods In the first the cells were loadedwith 5 mM cis-parinaric acid (Molecular ProbesEugene OR) as described elsewhere16 After15 hours of incubation at 37degC the cells werewashed and then treated with 100 mM AA or10 mM PA in medium for 6 hours In the sec-ond method the cells were first treated with100 mM AA or 10 mM PA for 18 hours afterwhich they were washed and further incubatedwith 5 mM cis-parinaric acid for 15 hours Thefluorescence measurements (as arbitrary unitsper cell) were made by confocal microscopy atan excitation wavelength of 334ndash364 nm and anemission wavelength of 424 nm
Incorporation of radiolabeled palmiticacid into the cells
Foreskin fibroblasts were plated in 24-wellmicroculture plates at a density of 20000 cells per well for 72 hours The medium wasaspirated and replaced with the fresh mediumsupplemented with 100 mM AA or 10 mM PAAfter 18 hours of pretreatment with AA or PAthe cells were washed and then incubated inmedium containing a mixture of 10 mCiml[3H]-palmitic acid (ARC Inc St Louis MO)and nonradiolabeled palmitic acid (as a carrier)at a final concentration of 5 mM At the end ofthe experiment supernatants were aspiratedcells were washed three times with PBS trans-ferred to the scintillation vials and processedfor liquid scintillation counting
Statistics
All values are expressed as mean 6 SD
RESULTS
Effect of ascorbic acid on intracellular DCFH-DAoxidation by cultured skin and foreskin fibroblasts
The rate of DCFH-DA oxidation in cells is de-pendent on the release of the ROS (mainly pro-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 109
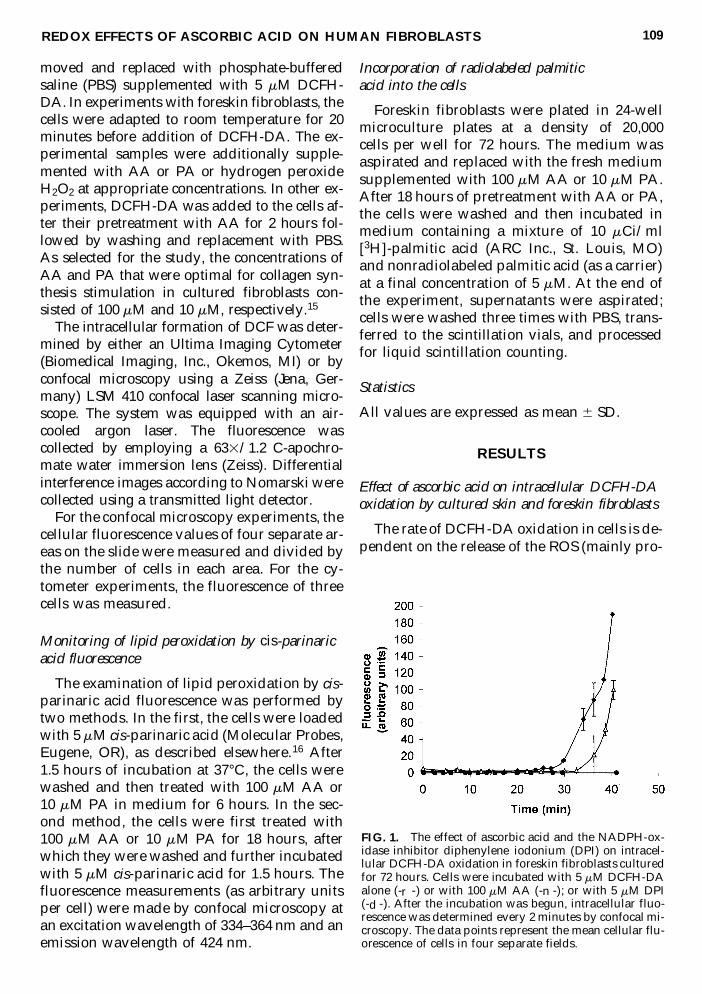
FIG 1 The effect of ascorbic acid and the NADPH-ox-idase inhibitor diphenylene iodonium (DPI) on intracel-lular DCFH-DA oxidation in foreskin fibroblasts culturedfor 72 hours Cells were incubated with 5 mM DCFH-DAalone (-r -) or with 100 mM AA (-n -) or with 5 mM DPI(-d -) After the incubation was begun intracellular fluo-rescence was determined every 2 minutes by confocal mi-croscopy The data points represent the mean cellular flu-orescence of cells in four separate fields
duced by NADPH-oxidase1718 and on the in-teraction of ROS with DCFH-DA leading to theformation of fluorescent DCF19ndash21
The time required for the increase in DCFconcentration to reach a detectable level wasused as a quantitative measure of intracellularROS formation in the following studies In aninitial experiment the level of intracellular ROSproduction in cultured foreskin fibroblasts wasevaluated The NADPH-oxidase enzyme com-plex was demonstrated previously to be themain source of superoxide and superoxide-de-rived ROS in dermal fibroblasts1718 In the cur-rent study DCFH-DA oxidation in foreskinfibroblasts was completely inhibited by 5 mMdiphenylene iodonium (a specific inhibitor ofthe leukocyte NADPH oxidase) thus demon-strating that in foreskin fibroblasts superoxideradicals (or H2O2 formed under superoxidedismutation) generated by NADPH-oxidaseare responsible for intracellular DCFH-DA ox-idation (Fig 1)
In nontreated skin fibroblasts cultured for 24hours after passage detectable DCF fluores-cence was seen after 75 minutes The exposureof the cells to H2O2 or to AA increased the rate
of intracellular DCFH-DA oxidation the effectof H2O2 was much greater than that of AA (Fig2) A representative imaging cytometer photo-graph of experimental human skin fibroblastsexposed for 75 minutes is shown in Fig 3 In-creased fluorescence was seen on microscopicimages when dermal fibroblasts (24 hours af-ter passage) were exposed to H2O2 (averagefluorescence 750 arbitrary units per cell) or toAA (210 arbitrary units per cell) comparedwith control (25 arbitrary units per cell) (Fig3D B versus A) When cells were pretreatedwith AA for 2 hours followed by cell washingbefore the experiment they did not demon-strate a significant difference in intracellularfluorescence (30 arbitrary units per cell) com-pared with control cells (Fig 3C versus A)
The accelerated effects of AA and H2O2 onDCFH-DA oxidation rate were also observedin primary foreskin fibroblasts (cultured for 24hours after passage) adapted to room temper-ature detectable DCF fluorescence in non-treated cells was seen after 32 minutes of incu-bation with DCFH-DA (Fig 4 the data aretypical of the results of three independent ex-periments) Cells incubated with 100 mM AA
ROSENBLAT ET AL110
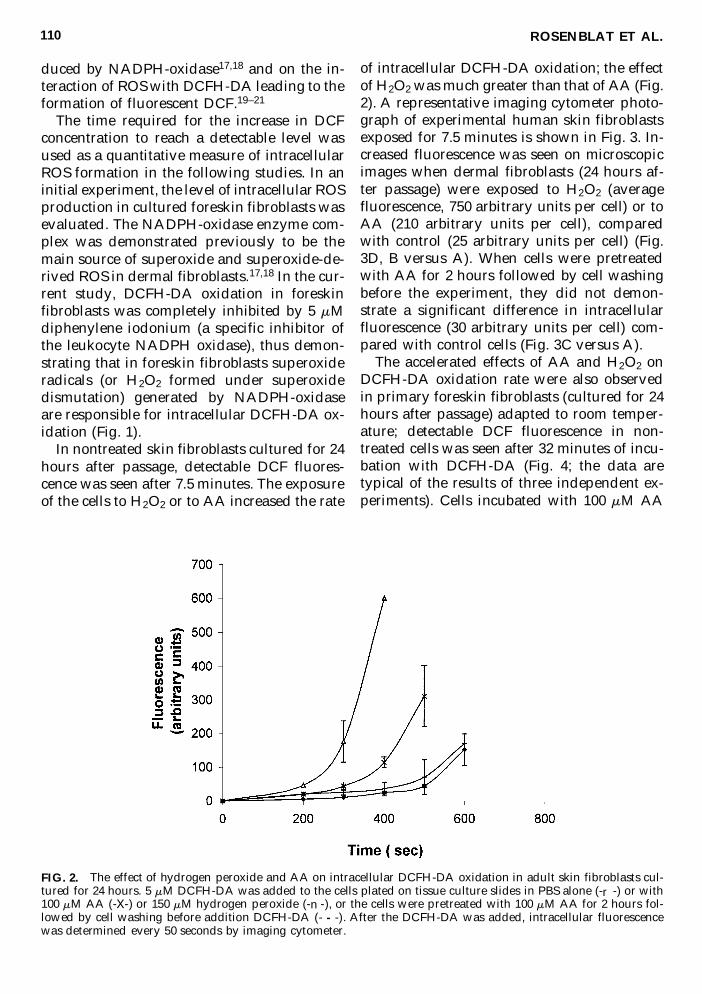
FIG 2 The effect of hydrogen peroxide and AA on intracellular DCFH-DA oxidation in adult skin fibroblasts cul-tured for 24 hours 5 mM DCFH-DA was added to the cells plated on tissue culture slides in PBS alone (-r -) or with100 mM AA (-X-) or 150 mM hydrogen peroxide (- n -) or the cells were pretreated with 100 mM AA for 2 hours fol-lowed by cell washing before addition DCFH-DA (- - -) After the DCFH-DA was added intracellular fluorescencewas determined every 50 seconds by imaging cytometer
or 150 mM identical DCFH-DA oxidation pro-files the increase in intracellular fluorescencewas seen already at 28 minutes (Fig 4) Repre-sentative confocal microscopic images of hu-man foreskin fibroblasts tested under theseconditions are shown in Fig 5 (B C versus D)
When the foreskin fibroblasts were culturedfor 72 hours after passage the control (non-treated) cells developed a detectable fluores-cence after 28 minutes of incubation Howeverin this case AA treatment led to a prolonga-tion in the time required for the increased flu-orescence due to DCFH-DA oxidation 34 min-utes in response to 100 mM AA (Fig 1) Whencells were incubated with PA (3ndash10 mM) therewas a marked prolongation of the time for syn-thesis of the detectable amount of the fluores-
cent compound DCF (Fig 6) This prolongationwas abrogated by the addition of 150 mM H2O2(Fig 6) Representative confocal microscopicimages of this inhibition of fluorescence by AAand PA in cells cultured for 72 hours are shownin Fig 5 E through H Cells treated with AAhad less fluorescence at 38 and 41 minutes thandid control cells at 38 minutes (Fig 5F G ver-sus E) Cells treated with PA had no observablefluorescence (Fig 5H)
Determination of ascorbate-induced lipidperoxidation by parinaric acid fluorescence
In order to determine whether AA affectslipid peroxidation cells were preincubatedwith AA (100 mM) for 18 hours and then with
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 111
FIG 3 Image assay of the intracellular formation and distribution of DCF in adult human dermal fibroblasts ex-posed to AA or hydrogen peroxide Primary human dermal fibroblasts were cultured for 24 hours before the exper-iment Cells were treated with 5 mM DCFH-DA alone (A) with 100 mM AA (B) or with 150 mM hydrogen peroxide(D) or pretreated with 100 mM AA for 2 hours before PBS rinse and addition of 5mM DCFH-DA (C) IntracellularDCF fluorescence was measured 450 seconds after the beginning of the incubation
cis-parinaric acid a fluorescent probe sensitiveto lipid peroxidation Cis-parinaric acid is a flu-orescent structural analog of a polyunsaturatedfatty acid that readily incorporates into mem-
branes Its fluorescence is destroyed on reac-tion with free radicals thus providing a quan-titative measurement of lipid membrane dam-age22ndash24 Cells preincubated with AA for 18hours showed a marked 77 6 2 inhibition offluorescence compared with control indicatinglipid peroxidation induced by the AA (Fig 7Bversus A) The effect of 10 mM PA on the inhi-bition of fluorescence was less pronounced(46 6 2 inhibition data not shown) In a sec-ond experiment parinaric acid was incorpo-rated into fibroblasts for 15 hours before ex-posure to AA for 6 hours In cells exposed toAA parinaric acid fluorescence was decreasedby 28 6 8 (Fig 7C) No change in cis-parinaricacid fluorescence in the control cells was ob-served after 6 hours
In order to investigate whether the inhibitionof fluorescence by AA might have resultedfrom an inhibition of parinaric acid incorpora-tion into cells fibroblasts were incubated withradiolabeled palmitic acid and uptake of ra-dioisotope quantitated in the presence of AAor PA In cells pretreated for 18 hours with 100mM AA or 10 mM PA there was 13 and 22inhibition of the uptake of palmitic acid re-spectively (Table 1) which did not account forthe 77 and 46 inhibition of fluorescence
ROSENBLAT ET AL112
FIG 4 The effect of hydrogen peroxide and AA on in-tracellular DCFH-DA oxidation in newborn foreskinfibroblasts cultured for 24 hours The cells plated on tis-sue culture slides in PBS were incubated with 5 mMDCFH-DA alone (-r -) or with 100 mM AA (-m -) or 150mM hydrogen peroxide (-u -) After the incubation wasbegun intracellular fluorescence was determined every 2minutes by confocal microscopy The fluorescence of fourrepresentative fields was quantitated for each time pointThe data are representative of the results of three inde-pendent experiments
FIG 5 Confocal microscopy of intracellular DCF fluorescence as a measure of intracellular ROS production in new-born foreskin fibroblasts (AndashD) Cells cultured for 24 hours after passage A 50 mM AA for 0 minutes B 50 mM AAfor 33 minutes C 150 mM hydrogen peroxide for 33 minutes D control cells 33 minutes (EndashH) Cells cultured for72 hours after passage E control cells 38 minutes F 50 mM AA for 38 minutes G 50 mM AA for 41 minutes H 3mM PA for 41 minutes
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115

dase4 Antioxidants protect the cells against ex-cessive ROS generation Nevertheless the bio-logical effects of some antioxidants are para-doxically associated with their ability in vitro toundergo redox cycling to generate ROS
Ascorbic acid (AA) is involved in manyphysiological functions in living organismsincluding the regulation of collagen synthe-sis56 This activity may be explained by thecapacity of AA to participate in oxidative-re-ductive reactions both as an antioxidant andas a prooxidant depending on the local envi-ronment in which the molecule is present7
The antioxidant activity of AA and its deriv-atives is a result of its ability to react with avariety of free radicals and ROS AA readilyundergoes oxidation forming an intermediateradical of low reactivity that may account forthe antioxidant effect The prooxidant activityof AA is derived from its ability to reducetransition metals such as Fe13 and Cu12 andalso to reduce O2 The formation of severalROS in the presence of AA may be illustratedby the following reactions8
AscH2 1 Fe13(Cu12) RAsc2 1 Fe12(Cu1) 1 H1
Fe12(Cu1) 1 O2 R O22 1 Fe13(Cu12)
AscH2 1 O22 1 H1 R H2O2 1 Asc2
Fe12(Cu1) 1 H2O2 ROH2 1 OH 1 Fe13(Cu12)
In the reactions above AscH2 is the reducedform of ascorbic acid Asc2 is the semi-oxidizedascorbic acid radical OH2 is the hydroxyl anionand OH is the hydroxyl radical One hypothe-sis that has been proposed regarding the role ofAA in collagen synthesis is that AA stimulateslipid peroxidation with the formation of reactivealdehydes The aldehydes then upregulate ex-pression of the procollagen genes9ndash11 Despite alarge body of literature on the subject the phys-iological relevance of the prooxidative effect ofAA or its derivatives remains unclear A limitednumber of reports have demonstrated that ox-idative damage to DNA lipids and proteins isseen in cells treated with AA712 A major ques-tion still remains whether AA and its derivativesaffect cellular metabolism by augmentation or by
the opposite inhibition of intracellular ROS pro-duction
Our current interest in this subject focuses onthe in situ effect of physiologically active con-centrations of AA (in terms of stimulation ofcollagen synthesis) and of its hydrophobic de-rivative palmitoyl ascorbate (PA) on intracel-lular ROS generation and lipid peroxidation inthe cytoplasm membrane of cultured cells
In the present study we used two fluorescentldquoreporter moleculesrdquo to examine the intracel-lular production of ROS and lipid peroxidationin cultured neonatal foreskin fibroblasts or skinfibroblasts in response to AA and PA The firstmethod was based on the intracellular interac-tion of the nonfluorescent compound 2979-dichlorofluorescin diacetate (DCFH-DA) withROS (particularly hydrogen peroxide H2O2) toproduce the fluorescence product 2979-dichlo-rofluorescein (DCF) The second method mon-itored cis-parinaric acid fluorescence which isdecreased upon reaction with free radicals in alipid environment
MATERIALS AND METHODS
Cell culture
Dermal and foreskin fibroblasts were grownin culture from explants of human adult skinor neonatal foreskin Cells were grown in 75-mm2 plastic flasks (Corning Glass WorksCorning NY) in Dulbeccorsquos modified Eaglemedium (Biological Industries Kibbutz BeitHaemek Israel or Sigma Chemical Co StLouis MO) containing 4500 mgL glucose andsupplemented with 10 fetal calf serum (FCS)(Biological Industries or Bio-Whittaker Wal-tersville MA) in a humidified atmosphere of7 carbon dioxide Once confluent primarycultures were subcultured by trypsinizationand used between passages 3 and 18
Intracellular oxidation of DCFH-DA
Cells were seeded in two- or four-chamber tis-sue culture slides (Nalge Nunc InternationalNaperville IL) at densities of 10000ndash20000 cellsper well and incubated in a humidified atmos-phere for 24 or 72 hours at 37degC according to theprotocol described earlier Prior to the experi-ments with skin fibroblasts the medium was re-
ROSENBLAT ET AL108
moved and replaced with phosphate-bufferedsaline (PBS) supplemented with 5 mM DCFH-DA In experiments with foreskin fibroblasts thecells were adapted to room temperature for 20minutes before addition of DCFH-DA The ex-perimental samples were additionally supple-mented with AA or PA or hydrogen peroxideH2O2 at appropriate concentrations In other ex-periments DCFH-DA was added to the cells af-ter their pretreatment with AA for 2 hours fol-lowed by washing and replacement with PBSAs selected for the study the concentrations ofAA and PA that were optimal for collagen syn-thesis stimulation in cultured fibroblasts con-sisted of 100 mM and 10 mM respectively15
The intracellular formation of DCF was deter-mined by either an Ultima Imaging Cytometer(Biomedical Imaging Inc Okemos MI) or byconfocal microscopy using a Zeiss (Jena Ger-many) LSM 410 confocal laser scanning micro-scope The system was equipped with an air-cooled argon laser The fluorescence wascollected by employing a 63312 C-apochro-mate water immersion lens (Zeiss) Differentialinterference images according to Nomarski werecollected using a transmitted light detector
For the confocal microscopy experiments thecellular fluorescence values of four separate ar-eas on the slide were measured and divided bythe number of cells in each area For the cy-tometer experiments the fluorescence of threecells was measured
Monitoring of lipid peroxidation by cis-parinaricacid fluorescence
The examination of lipid peroxidation by cis-parinaric acid fluorescence was performed bytwo methods In the first the cells were loadedwith 5 mM cis-parinaric acid (Molecular ProbesEugene OR) as described elsewhere16 After15 hours of incubation at 37degC the cells werewashed and then treated with 100 mM AA or10 mM PA in medium for 6 hours In the sec-ond method the cells were first treated with100 mM AA or 10 mM PA for 18 hours afterwhich they were washed and further incubatedwith 5 mM cis-parinaric acid for 15 hours Thefluorescence measurements (as arbitrary unitsper cell) were made by confocal microscopy atan excitation wavelength of 334ndash364 nm and anemission wavelength of 424 nm
Incorporation of radiolabeled palmiticacid into the cells
Foreskin fibroblasts were plated in 24-wellmicroculture plates at a density of 20000 cells per well for 72 hours The medium wasaspirated and replaced with the fresh mediumsupplemented with 100 mM AA or 10 mM PAAfter 18 hours of pretreatment with AA or PAthe cells were washed and then incubated inmedium containing a mixture of 10 mCiml[3H]-palmitic acid (ARC Inc St Louis MO)and nonradiolabeled palmitic acid (as a carrier)at a final concentration of 5 mM At the end ofthe experiment supernatants were aspiratedcells were washed three times with PBS trans-ferred to the scintillation vials and processedfor liquid scintillation counting
Statistics
All values are expressed as mean 6 SD
RESULTS
Effect of ascorbic acid on intracellular DCFH-DAoxidation by cultured skin and foreskin fibroblasts
The rate of DCFH-DA oxidation in cells is de-pendent on the release of the ROS (mainly pro-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 109
FIG 1 The effect of ascorbic acid and the NADPH-ox-idase inhibitor diphenylene iodonium (DPI) on intracel-lular DCFH-DA oxidation in foreskin fibroblasts culturedfor 72 hours Cells were incubated with 5 mM DCFH-DAalone (-r -) or with 100 mM AA (-n -) or with 5 mM DPI(-d -) After the incubation was begun intracellular fluo-rescence was determined every 2 minutes by confocal mi-croscopy The data points represent the mean cellular flu-orescence of cells in four separate fields
duced by NADPH-oxidase1718 and on the in-teraction of ROS with DCFH-DA leading to theformation of fluorescent DCF19ndash21
The time required for the increase in DCFconcentration to reach a detectable level wasused as a quantitative measure of intracellularROS formation in the following studies In aninitial experiment the level of intracellular ROSproduction in cultured foreskin fibroblasts wasevaluated The NADPH-oxidase enzyme com-plex was demonstrated previously to be themain source of superoxide and superoxide-de-rived ROS in dermal fibroblasts1718 In the cur-rent study DCFH-DA oxidation in foreskinfibroblasts was completely inhibited by 5 mMdiphenylene iodonium (a specific inhibitor ofthe leukocyte NADPH oxidase) thus demon-strating that in foreskin fibroblasts superoxideradicals (or H2O2 formed under superoxidedismutation) generated by NADPH-oxidaseare responsible for intracellular DCFH-DA ox-idation (Fig 1)
In nontreated skin fibroblasts cultured for 24hours after passage detectable DCF fluores-cence was seen after 75 minutes The exposureof the cells to H2O2 or to AA increased the rate
of intracellular DCFH-DA oxidation the effectof H2O2 was much greater than that of AA (Fig2) A representative imaging cytometer photo-graph of experimental human skin fibroblastsexposed for 75 minutes is shown in Fig 3 In-creased fluorescence was seen on microscopicimages when dermal fibroblasts (24 hours af-ter passage) were exposed to H2O2 (averagefluorescence 750 arbitrary units per cell) or toAA (210 arbitrary units per cell) comparedwith control (25 arbitrary units per cell) (Fig3D B versus A) When cells were pretreatedwith AA for 2 hours followed by cell washingbefore the experiment they did not demon-strate a significant difference in intracellularfluorescence (30 arbitrary units per cell) com-pared with control cells (Fig 3C versus A)
The accelerated effects of AA and H2O2 onDCFH-DA oxidation rate were also observedin primary foreskin fibroblasts (cultured for 24hours after passage) adapted to room temper-ature detectable DCF fluorescence in non-treated cells was seen after 32 minutes of incu-bation with DCFH-DA (Fig 4 the data aretypical of the results of three independent ex-periments) Cells incubated with 100 mM AA
ROSENBLAT ET AL110
FIG 2 The effect of hydrogen peroxide and AA on intracellular DCFH-DA oxidation in adult skin fibroblasts cul-tured for 24 hours 5 mM DCFH-DA was added to the cells plated on tissue culture slides in PBS alone (-r -) or with100 mM AA (-X-) or 150 mM hydrogen peroxide (- n -) or the cells were pretreated with 100 mM AA for 2 hours fol-lowed by cell washing before addition DCFH-DA (- - -) After the DCFH-DA was added intracellular fluorescencewas determined every 50 seconds by imaging cytometer
or 150 mM identical DCFH-DA oxidation pro-files the increase in intracellular fluorescencewas seen already at 28 minutes (Fig 4) Repre-sentative confocal microscopic images of hu-man foreskin fibroblasts tested under theseconditions are shown in Fig 5 (B C versus D)
When the foreskin fibroblasts were culturedfor 72 hours after passage the control (non-treated) cells developed a detectable fluores-cence after 28 minutes of incubation Howeverin this case AA treatment led to a prolonga-tion in the time required for the increased flu-orescence due to DCFH-DA oxidation 34 min-utes in response to 100 mM AA (Fig 1) Whencells were incubated with PA (3ndash10 mM) therewas a marked prolongation of the time for syn-thesis of the detectable amount of the fluores-
cent compound DCF (Fig 6) This prolongationwas abrogated by the addition of 150 mM H2O2(Fig 6) Representative confocal microscopicimages of this inhibition of fluorescence by AAand PA in cells cultured for 72 hours are shownin Fig 5 E through H Cells treated with AAhad less fluorescence at 38 and 41 minutes thandid control cells at 38 minutes (Fig 5F G ver-sus E) Cells treated with PA had no observablefluorescence (Fig 5H)
Determination of ascorbate-induced lipidperoxidation by parinaric acid fluorescence
In order to determine whether AA affectslipid peroxidation cells were preincubatedwith AA (100 mM) for 18 hours and then with
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 111
FIG 3 Image assay of the intracellular formation and distribution of DCF in adult human dermal fibroblasts ex-posed to AA or hydrogen peroxide Primary human dermal fibroblasts were cultured for 24 hours before the exper-iment Cells were treated with 5 mM DCFH-DA alone (A) with 100 mM AA (B) or with 150 mM hydrogen peroxide(D) or pretreated with 100 mM AA for 2 hours before PBS rinse and addition of 5mM DCFH-DA (C) IntracellularDCF fluorescence was measured 450 seconds after the beginning of the incubation
cis-parinaric acid a fluorescent probe sensitiveto lipid peroxidation Cis-parinaric acid is a flu-orescent structural analog of a polyunsaturatedfatty acid that readily incorporates into mem-
branes Its fluorescence is destroyed on reac-tion with free radicals thus providing a quan-titative measurement of lipid membrane dam-age22ndash24 Cells preincubated with AA for 18hours showed a marked 77 6 2 inhibition offluorescence compared with control indicatinglipid peroxidation induced by the AA (Fig 7Bversus A) The effect of 10 mM PA on the inhi-bition of fluorescence was less pronounced(46 6 2 inhibition data not shown) In a sec-ond experiment parinaric acid was incorpo-rated into fibroblasts for 15 hours before ex-posure to AA for 6 hours In cells exposed toAA parinaric acid fluorescence was decreasedby 28 6 8 (Fig 7C) No change in cis-parinaricacid fluorescence in the control cells was ob-served after 6 hours
In order to investigate whether the inhibitionof fluorescence by AA might have resultedfrom an inhibition of parinaric acid incorpora-tion into cells fibroblasts were incubated withradiolabeled palmitic acid and uptake of ra-dioisotope quantitated in the presence of AAor PA In cells pretreated for 18 hours with 100mM AA or 10 mM PA there was 13 and 22inhibition of the uptake of palmitic acid re-spectively (Table 1) which did not account forthe 77 and 46 inhibition of fluorescence
ROSENBLAT ET AL112
FIG 4 The effect of hydrogen peroxide and AA on in-tracellular DCFH-DA oxidation in newborn foreskinfibroblasts cultured for 24 hours The cells plated on tis-sue culture slides in PBS were incubated with 5 mMDCFH-DA alone (-r -) or with 100 mM AA (-m -) or 150mM hydrogen peroxide (-u -) After the incubation wasbegun intracellular fluorescence was determined every 2minutes by confocal microscopy The fluorescence of fourrepresentative fields was quantitated for each time pointThe data are representative of the results of three inde-pendent experiments
FIG 5 Confocal microscopy of intracellular DCF fluorescence as a measure of intracellular ROS production in new-born foreskin fibroblasts (AndashD) Cells cultured for 24 hours after passage A 50 mM AA for 0 minutes B 50 mM AAfor 33 minutes C 150 mM hydrogen peroxide for 33 minutes D control cells 33 minutes (EndashH) Cells cultured for72 hours after passage E control cells 38 minutes F 50 mM AA for 38 minutes G 50 mM AA for 41 minutes H 3mM PA for 41 minutes
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115
moved and replaced with phosphate-bufferedsaline (PBS) supplemented with 5 mM DCFH-DA In experiments with foreskin fibroblasts thecells were adapted to room temperature for 20minutes before addition of DCFH-DA The ex-perimental samples were additionally supple-mented with AA or PA or hydrogen peroxideH2O2 at appropriate concentrations In other ex-periments DCFH-DA was added to the cells af-ter their pretreatment with AA for 2 hours fol-lowed by washing and replacement with PBSAs selected for the study the concentrations ofAA and PA that were optimal for collagen syn-thesis stimulation in cultured fibroblasts con-sisted of 100 mM and 10 mM respectively15
The intracellular formation of DCF was deter-mined by either an Ultima Imaging Cytometer(Biomedical Imaging Inc Okemos MI) or byconfocal microscopy using a Zeiss (Jena Ger-many) LSM 410 confocal laser scanning micro-scope The system was equipped with an air-cooled argon laser The fluorescence wascollected by employing a 63312 C-apochro-mate water immersion lens (Zeiss) Differentialinterference images according to Nomarski werecollected using a transmitted light detector
For the confocal microscopy experiments thecellular fluorescence values of four separate ar-eas on the slide were measured and divided bythe number of cells in each area For the cy-tometer experiments the fluorescence of threecells was measured
Monitoring of lipid peroxidation by cis-parinaricacid fluorescence
The examination of lipid peroxidation by cis-parinaric acid fluorescence was performed bytwo methods In the first the cells were loadedwith 5 mM cis-parinaric acid (Molecular ProbesEugene OR) as described elsewhere16 After15 hours of incubation at 37degC the cells werewashed and then treated with 100 mM AA or10 mM PA in medium for 6 hours In the sec-ond method the cells were first treated with100 mM AA or 10 mM PA for 18 hours afterwhich they were washed and further incubatedwith 5 mM cis-parinaric acid for 15 hours Thefluorescence measurements (as arbitrary unitsper cell) were made by confocal microscopy atan excitation wavelength of 334ndash364 nm and anemission wavelength of 424 nm
Incorporation of radiolabeled palmiticacid into the cells
Foreskin fibroblasts were plated in 24-wellmicroculture plates at a density of 20000 cells per well for 72 hours The medium wasaspirated and replaced with the fresh mediumsupplemented with 100 mM AA or 10 mM PAAfter 18 hours of pretreatment with AA or PAthe cells were washed and then incubated inmedium containing a mixture of 10 mCiml[3H]-palmitic acid (ARC Inc St Louis MO)and nonradiolabeled palmitic acid (as a carrier)at a final concentration of 5 mM At the end ofthe experiment supernatants were aspiratedcells were washed three times with PBS trans-ferred to the scintillation vials and processedfor liquid scintillation counting
Statistics
All values are expressed as mean 6 SD
RESULTS
Effect of ascorbic acid on intracellular DCFH-DAoxidation by cultured skin and foreskin fibroblasts
The rate of DCFH-DA oxidation in cells is de-pendent on the release of the ROS (mainly pro-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 109
FIG 1 The effect of ascorbic acid and the NADPH-ox-idase inhibitor diphenylene iodonium (DPI) on intracel-lular DCFH-DA oxidation in foreskin fibroblasts culturedfor 72 hours Cells were incubated with 5 mM DCFH-DAalone (-r -) or with 100 mM AA (-n -) or with 5 mM DPI(-d -) After the incubation was begun intracellular fluo-rescence was determined every 2 minutes by confocal mi-croscopy The data points represent the mean cellular flu-orescence of cells in four separate fields
duced by NADPH-oxidase1718 and on the in-teraction of ROS with DCFH-DA leading to theformation of fluorescent DCF19ndash21
The time required for the increase in DCFconcentration to reach a detectable level wasused as a quantitative measure of intracellularROS formation in the following studies In aninitial experiment the level of intracellular ROSproduction in cultured foreskin fibroblasts wasevaluated The NADPH-oxidase enzyme com-plex was demonstrated previously to be themain source of superoxide and superoxide-de-rived ROS in dermal fibroblasts1718 In the cur-rent study DCFH-DA oxidation in foreskinfibroblasts was completely inhibited by 5 mMdiphenylene iodonium (a specific inhibitor ofthe leukocyte NADPH oxidase) thus demon-strating that in foreskin fibroblasts superoxideradicals (or H2O2 formed under superoxidedismutation) generated by NADPH-oxidaseare responsible for intracellular DCFH-DA ox-idation (Fig 1)
In nontreated skin fibroblasts cultured for 24hours after passage detectable DCF fluores-cence was seen after 75 minutes The exposureof the cells to H2O2 or to AA increased the rate
of intracellular DCFH-DA oxidation the effectof H2O2 was much greater than that of AA (Fig2) A representative imaging cytometer photo-graph of experimental human skin fibroblastsexposed for 75 minutes is shown in Fig 3 In-creased fluorescence was seen on microscopicimages when dermal fibroblasts (24 hours af-ter passage) were exposed to H2O2 (averagefluorescence 750 arbitrary units per cell) or toAA (210 arbitrary units per cell) comparedwith control (25 arbitrary units per cell) (Fig3D B versus A) When cells were pretreatedwith AA for 2 hours followed by cell washingbefore the experiment they did not demon-strate a significant difference in intracellularfluorescence (30 arbitrary units per cell) com-pared with control cells (Fig 3C versus A)
The accelerated effects of AA and H2O2 onDCFH-DA oxidation rate were also observedin primary foreskin fibroblasts (cultured for 24hours after passage) adapted to room temper-ature detectable DCF fluorescence in non-treated cells was seen after 32 minutes of incu-bation with DCFH-DA (Fig 4 the data aretypical of the results of three independent ex-periments) Cells incubated with 100 mM AA
ROSENBLAT ET AL110
FIG 2 The effect of hydrogen peroxide and AA on intracellular DCFH-DA oxidation in adult skin fibroblasts cul-tured for 24 hours 5 mM DCFH-DA was added to the cells plated on tissue culture slides in PBS alone (-r -) or with100 mM AA (-X-) or 150 mM hydrogen peroxide (- n -) or the cells were pretreated with 100 mM AA for 2 hours fol-lowed by cell washing before addition DCFH-DA (- - -) After the DCFH-DA was added intracellular fluorescencewas determined every 50 seconds by imaging cytometer
or 150 mM identical DCFH-DA oxidation pro-files the increase in intracellular fluorescencewas seen already at 28 minutes (Fig 4) Repre-sentative confocal microscopic images of hu-man foreskin fibroblasts tested under theseconditions are shown in Fig 5 (B C versus D)
When the foreskin fibroblasts were culturedfor 72 hours after passage the control (non-treated) cells developed a detectable fluores-cence after 28 minutes of incubation Howeverin this case AA treatment led to a prolonga-tion in the time required for the increased flu-orescence due to DCFH-DA oxidation 34 min-utes in response to 100 mM AA (Fig 1) Whencells were incubated with PA (3ndash10 mM) therewas a marked prolongation of the time for syn-thesis of the detectable amount of the fluores-
cent compound DCF (Fig 6) This prolongationwas abrogated by the addition of 150 mM H2O2(Fig 6) Representative confocal microscopicimages of this inhibition of fluorescence by AAand PA in cells cultured for 72 hours are shownin Fig 5 E through H Cells treated with AAhad less fluorescence at 38 and 41 minutes thandid control cells at 38 minutes (Fig 5F G ver-sus E) Cells treated with PA had no observablefluorescence (Fig 5H)
Determination of ascorbate-induced lipidperoxidation by parinaric acid fluorescence
In order to determine whether AA affectslipid peroxidation cells were preincubatedwith AA (100 mM) for 18 hours and then with
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 111
FIG 3 Image assay of the intracellular formation and distribution of DCF in adult human dermal fibroblasts ex-posed to AA or hydrogen peroxide Primary human dermal fibroblasts were cultured for 24 hours before the exper-iment Cells were treated with 5 mM DCFH-DA alone (A) with 100 mM AA (B) or with 150 mM hydrogen peroxide(D) or pretreated with 100 mM AA for 2 hours before PBS rinse and addition of 5mM DCFH-DA (C) IntracellularDCF fluorescence was measured 450 seconds after the beginning of the incubation
cis-parinaric acid a fluorescent probe sensitiveto lipid peroxidation Cis-parinaric acid is a flu-orescent structural analog of a polyunsaturatedfatty acid that readily incorporates into mem-
branes Its fluorescence is destroyed on reac-tion with free radicals thus providing a quan-titative measurement of lipid membrane dam-age22ndash24 Cells preincubated with AA for 18hours showed a marked 77 6 2 inhibition offluorescence compared with control indicatinglipid peroxidation induced by the AA (Fig 7Bversus A) The effect of 10 mM PA on the inhi-bition of fluorescence was less pronounced(46 6 2 inhibition data not shown) In a sec-ond experiment parinaric acid was incorpo-rated into fibroblasts for 15 hours before ex-posure to AA for 6 hours In cells exposed toAA parinaric acid fluorescence was decreasedby 28 6 8 (Fig 7C) No change in cis-parinaricacid fluorescence in the control cells was ob-served after 6 hours
In order to investigate whether the inhibitionof fluorescence by AA might have resultedfrom an inhibition of parinaric acid incorpora-tion into cells fibroblasts were incubated withradiolabeled palmitic acid and uptake of ra-dioisotope quantitated in the presence of AAor PA In cells pretreated for 18 hours with 100mM AA or 10 mM PA there was 13 and 22inhibition of the uptake of palmitic acid re-spectively (Table 1) which did not account forthe 77 and 46 inhibition of fluorescence
ROSENBLAT ET AL112
FIG 4 The effect of hydrogen peroxide and AA on in-tracellular DCFH-DA oxidation in newborn foreskinfibroblasts cultured for 24 hours The cells plated on tis-sue culture slides in PBS were incubated with 5 mMDCFH-DA alone (-r -) or with 100 mM AA (-m -) or 150mM hydrogen peroxide (-u -) After the incubation wasbegun intracellular fluorescence was determined every 2minutes by confocal microscopy The fluorescence of fourrepresentative fields was quantitated for each time pointThe data are representative of the results of three inde-pendent experiments
FIG 5 Confocal microscopy of intracellular DCF fluorescence as a measure of intracellular ROS production in new-born foreskin fibroblasts (AndashD) Cells cultured for 24 hours after passage A 50 mM AA for 0 minutes B 50 mM AAfor 33 minutes C 150 mM hydrogen peroxide for 33 minutes D control cells 33 minutes (EndashH) Cells cultured for72 hours after passage E control cells 38 minutes F 50 mM AA for 38 minutes G 50 mM AA for 41 minutes H 3mM PA for 41 minutes
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115
duced by NADPH-oxidase1718 and on the in-teraction of ROS with DCFH-DA leading to theformation of fluorescent DCF19ndash21
The time required for the increase in DCFconcentration to reach a detectable level wasused as a quantitative measure of intracellularROS formation in the following studies In aninitial experiment the level of intracellular ROSproduction in cultured foreskin fibroblasts wasevaluated The NADPH-oxidase enzyme com-plex was demonstrated previously to be themain source of superoxide and superoxide-de-rived ROS in dermal fibroblasts1718 In the cur-rent study DCFH-DA oxidation in foreskinfibroblasts was completely inhibited by 5 mMdiphenylene iodonium (a specific inhibitor ofthe leukocyte NADPH oxidase) thus demon-strating that in foreskin fibroblasts superoxideradicals (or H2O2 formed under superoxidedismutation) generated by NADPH-oxidaseare responsible for intracellular DCFH-DA ox-idation (Fig 1)
In nontreated skin fibroblasts cultured for 24hours after passage detectable DCF fluores-cence was seen after 75 minutes The exposureof the cells to H2O2 or to AA increased the rate
of intracellular DCFH-DA oxidation the effectof H2O2 was much greater than that of AA (Fig2) A representative imaging cytometer photo-graph of experimental human skin fibroblastsexposed for 75 minutes is shown in Fig 3 In-creased fluorescence was seen on microscopicimages when dermal fibroblasts (24 hours af-ter passage) were exposed to H2O2 (averagefluorescence 750 arbitrary units per cell) or toAA (210 arbitrary units per cell) comparedwith control (25 arbitrary units per cell) (Fig3D B versus A) When cells were pretreatedwith AA for 2 hours followed by cell washingbefore the experiment they did not demon-strate a significant difference in intracellularfluorescence (30 arbitrary units per cell) com-pared with control cells (Fig 3C versus A)
The accelerated effects of AA and H2O2 onDCFH-DA oxidation rate were also observedin primary foreskin fibroblasts (cultured for 24hours after passage) adapted to room temper-ature detectable DCF fluorescence in non-treated cells was seen after 32 minutes of incu-bation with DCFH-DA (Fig 4 the data aretypical of the results of three independent ex-periments) Cells incubated with 100 mM AA
ROSENBLAT ET AL110
FIG 2 The effect of hydrogen peroxide and AA on intracellular DCFH-DA oxidation in adult skin fibroblasts cul-tured for 24 hours 5 mM DCFH-DA was added to the cells plated on tissue culture slides in PBS alone (-r -) or with100 mM AA (-X-) or 150 mM hydrogen peroxide (- n -) or the cells were pretreated with 100 mM AA for 2 hours fol-lowed by cell washing before addition DCFH-DA (- - -) After the DCFH-DA was added intracellular fluorescencewas determined every 50 seconds by imaging cytometer
or 150 mM identical DCFH-DA oxidation pro-files the increase in intracellular fluorescencewas seen already at 28 minutes (Fig 4) Repre-sentative confocal microscopic images of hu-man foreskin fibroblasts tested under theseconditions are shown in Fig 5 (B C versus D)
When the foreskin fibroblasts were culturedfor 72 hours after passage the control (non-treated) cells developed a detectable fluores-cence after 28 minutes of incubation Howeverin this case AA treatment led to a prolonga-tion in the time required for the increased flu-orescence due to DCFH-DA oxidation 34 min-utes in response to 100 mM AA (Fig 1) Whencells were incubated with PA (3ndash10 mM) therewas a marked prolongation of the time for syn-thesis of the detectable amount of the fluores-
cent compound DCF (Fig 6) This prolongationwas abrogated by the addition of 150 mM H2O2(Fig 6) Representative confocal microscopicimages of this inhibition of fluorescence by AAand PA in cells cultured for 72 hours are shownin Fig 5 E through H Cells treated with AAhad less fluorescence at 38 and 41 minutes thandid control cells at 38 minutes (Fig 5F G ver-sus E) Cells treated with PA had no observablefluorescence (Fig 5H)
Determination of ascorbate-induced lipidperoxidation by parinaric acid fluorescence
In order to determine whether AA affectslipid peroxidation cells were preincubatedwith AA (100 mM) for 18 hours and then with
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 111
FIG 3 Image assay of the intracellular formation and distribution of DCF in adult human dermal fibroblasts ex-posed to AA or hydrogen peroxide Primary human dermal fibroblasts were cultured for 24 hours before the exper-iment Cells were treated with 5 mM DCFH-DA alone (A) with 100 mM AA (B) or with 150 mM hydrogen peroxide(D) or pretreated with 100 mM AA for 2 hours before PBS rinse and addition of 5mM DCFH-DA (C) IntracellularDCF fluorescence was measured 450 seconds after the beginning of the incubation
cis-parinaric acid a fluorescent probe sensitiveto lipid peroxidation Cis-parinaric acid is a flu-orescent structural analog of a polyunsaturatedfatty acid that readily incorporates into mem-
branes Its fluorescence is destroyed on reac-tion with free radicals thus providing a quan-titative measurement of lipid membrane dam-age22ndash24 Cells preincubated with AA for 18hours showed a marked 77 6 2 inhibition offluorescence compared with control indicatinglipid peroxidation induced by the AA (Fig 7Bversus A) The effect of 10 mM PA on the inhi-bition of fluorescence was less pronounced(46 6 2 inhibition data not shown) In a sec-ond experiment parinaric acid was incorpo-rated into fibroblasts for 15 hours before ex-posure to AA for 6 hours In cells exposed toAA parinaric acid fluorescence was decreasedby 28 6 8 (Fig 7C) No change in cis-parinaricacid fluorescence in the control cells was ob-served after 6 hours
In order to investigate whether the inhibitionof fluorescence by AA might have resultedfrom an inhibition of parinaric acid incorpora-tion into cells fibroblasts were incubated withradiolabeled palmitic acid and uptake of ra-dioisotope quantitated in the presence of AAor PA In cells pretreated for 18 hours with 100mM AA or 10 mM PA there was 13 and 22inhibition of the uptake of palmitic acid re-spectively (Table 1) which did not account forthe 77 and 46 inhibition of fluorescence
ROSENBLAT ET AL112
FIG 4 The effect of hydrogen peroxide and AA on in-tracellular DCFH-DA oxidation in newborn foreskinfibroblasts cultured for 24 hours The cells plated on tis-sue culture slides in PBS were incubated with 5 mMDCFH-DA alone (-r -) or with 100 mM AA (-m -) or 150mM hydrogen peroxide (-u -) After the incubation wasbegun intracellular fluorescence was determined every 2minutes by confocal microscopy The fluorescence of fourrepresentative fields was quantitated for each time pointThe data are representative of the results of three inde-pendent experiments
FIG 5 Confocal microscopy of intracellular DCF fluorescence as a measure of intracellular ROS production in new-born foreskin fibroblasts (AndashD) Cells cultured for 24 hours after passage A 50 mM AA for 0 minutes B 50 mM AAfor 33 minutes C 150 mM hydrogen peroxide for 33 minutes D control cells 33 minutes (EndashH) Cells cultured for72 hours after passage E control cells 38 minutes F 50 mM AA for 38 minutes G 50 mM AA for 41 minutes H 3mM PA for 41 minutes
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115

or 150 mM identical DCFH-DA oxidation pro-files the increase in intracellular fluorescencewas seen already at 28 minutes (Fig 4) Repre-sentative confocal microscopic images of hu-man foreskin fibroblasts tested under theseconditions are shown in Fig 5 (B C versus D)
When the foreskin fibroblasts were culturedfor 72 hours after passage the control (non-treated) cells developed a detectable fluores-cence after 28 minutes of incubation Howeverin this case AA treatment led to a prolonga-tion in the time required for the increased flu-orescence due to DCFH-DA oxidation 34 min-utes in response to 100 mM AA (Fig 1) Whencells were incubated with PA (3ndash10 mM) therewas a marked prolongation of the time for syn-thesis of the detectable amount of the fluores-
cent compound DCF (Fig 6) This prolongationwas abrogated by the addition of 150 mM H2O2(Fig 6) Representative confocal microscopicimages of this inhibition of fluorescence by AAand PA in cells cultured for 72 hours are shownin Fig 5 E through H Cells treated with AAhad less fluorescence at 38 and 41 minutes thandid control cells at 38 minutes (Fig 5F G ver-sus E) Cells treated with PA had no observablefluorescence (Fig 5H)
Determination of ascorbate-induced lipidperoxidation by parinaric acid fluorescence
In order to determine whether AA affectslipid peroxidation cells were preincubatedwith AA (100 mM) for 18 hours and then with
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 111
FIG 3 Image assay of the intracellular formation and distribution of DCF in adult human dermal fibroblasts ex-posed to AA or hydrogen peroxide Primary human dermal fibroblasts were cultured for 24 hours before the exper-iment Cells were treated with 5 mM DCFH-DA alone (A) with 100 mM AA (B) or with 150 mM hydrogen peroxide(D) or pretreated with 100 mM AA for 2 hours before PBS rinse and addition of 5mM DCFH-DA (C) IntracellularDCF fluorescence was measured 450 seconds after the beginning of the incubation
cis-parinaric acid a fluorescent probe sensitiveto lipid peroxidation Cis-parinaric acid is a flu-orescent structural analog of a polyunsaturatedfatty acid that readily incorporates into mem-
branes Its fluorescence is destroyed on reac-tion with free radicals thus providing a quan-titative measurement of lipid membrane dam-age22ndash24 Cells preincubated with AA for 18hours showed a marked 77 6 2 inhibition offluorescence compared with control indicatinglipid peroxidation induced by the AA (Fig 7Bversus A) The effect of 10 mM PA on the inhi-bition of fluorescence was less pronounced(46 6 2 inhibition data not shown) In a sec-ond experiment parinaric acid was incorpo-rated into fibroblasts for 15 hours before ex-posure to AA for 6 hours In cells exposed toAA parinaric acid fluorescence was decreasedby 28 6 8 (Fig 7C) No change in cis-parinaricacid fluorescence in the control cells was ob-served after 6 hours
In order to investigate whether the inhibitionof fluorescence by AA might have resultedfrom an inhibition of parinaric acid incorpora-tion into cells fibroblasts were incubated withradiolabeled palmitic acid and uptake of ra-dioisotope quantitated in the presence of AAor PA In cells pretreated for 18 hours with 100mM AA or 10 mM PA there was 13 and 22inhibition of the uptake of palmitic acid re-spectively (Table 1) which did not account forthe 77 and 46 inhibition of fluorescence
ROSENBLAT ET AL112
FIG 4 The effect of hydrogen peroxide and AA on in-tracellular DCFH-DA oxidation in newborn foreskinfibroblasts cultured for 24 hours The cells plated on tis-sue culture slides in PBS were incubated with 5 mMDCFH-DA alone (-r -) or with 100 mM AA (-m -) or 150mM hydrogen peroxide (-u -) After the incubation wasbegun intracellular fluorescence was determined every 2minutes by confocal microscopy The fluorescence of fourrepresentative fields was quantitated for each time pointThe data are representative of the results of three inde-pendent experiments
FIG 5 Confocal microscopy of intracellular DCF fluorescence as a measure of intracellular ROS production in new-born foreskin fibroblasts (AndashD) Cells cultured for 24 hours after passage A 50 mM AA for 0 minutes B 50 mM AAfor 33 minutes C 150 mM hydrogen peroxide for 33 minutes D control cells 33 minutes (EndashH) Cells cultured for72 hours after passage E control cells 38 minutes F 50 mM AA for 38 minutes G 50 mM AA for 41 minutes H 3mM PA for 41 minutes
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115

cis-parinaric acid a fluorescent probe sensitiveto lipid peroxidation Cis-parinaric acid is a flu-orescent structural analog of a polyunsaturatedfatty acid that readily incorporates into mem-
branes Its fluorescence is destroyed on reac-tion with free radicals thus providing a quan-titative measurement of lipid membrane dam-age22ndash24 Cells preincubated with AA for 18hours showed a marked 77 6 2 inhibition offluorescence compared with control indicatinglipid peroxidation induced by the AA (Fig 7Bversus A) The effect of 10 mM PA on the inhi-bition of fluorescence was less pronounced(46 6 2 inhibition data not shown) In a sec-ond experiment parinaric acid was incorpo-rated into fibroblasts for 15 hours before ex-posure to AA for 6 hours In cells exposed toAA parinaric acid fluorescence was decreasedby 28 6 8 (Fig 7C) No change in cis-parinaricacid fluorescence in the control cells was ob-served after 6 hours
In order to investigate whether the inhibitionof fluorescence by AA might have resultedfrom an inhibition of parinaric acid incorpora-tion into cells fibroblasts were incubated withradiolabeled palmitic acid and uptake of ra-dioisotope quantitated in the presence of AAor PA In cells pretreated for 18 hours with 100mM AA or 10 mM PA there was 13 and 22inhibition of the uptake of palmitic acid re-spectively (Table 1) which did not account forthe 77 and 46 inhibition of fluorescence
ROSENBLAT ET AL112
FIG 4 The effect of hydrogen peroxide and AA on in-tracellular DCFH-DA oxidation in newborn foreskinfibroblasts cultured for 24 hours The cells plated on tis-sue culture slides in PBS were incubated with 5 mMDCFH-DA alone (-r -) or with 100 mM AA (-m -) or 150mM hydrogen peroxide (-u -) After the incubation wasbegun intracellular fluorescence was determined every 2minutes by confocal microscopy The fluorescence of fourrepresentative fields was quantitated for each time pointThe data are representative of the results of three inde-pendent experiments
FIG 5 Confocal microscopy of intracellular DCF fluorescence as a measure of intracellular ROS production in new-born foreskin fibroblasts (AndashD) Cells cultured for 24 hours after passage A 50 mM AA for 0 minutes B 50 mM AAfor 33 minutes C 150 mM hydrogen peroxide for 33 minutes D control cells 33 minutes (EndashH) Cells cultured for72 hours after passage E control cells 38 minutes F 50 mM AA for 38 minutes G 50 mM AA for 41 minutes H 3mM PA for 41 minutes
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115
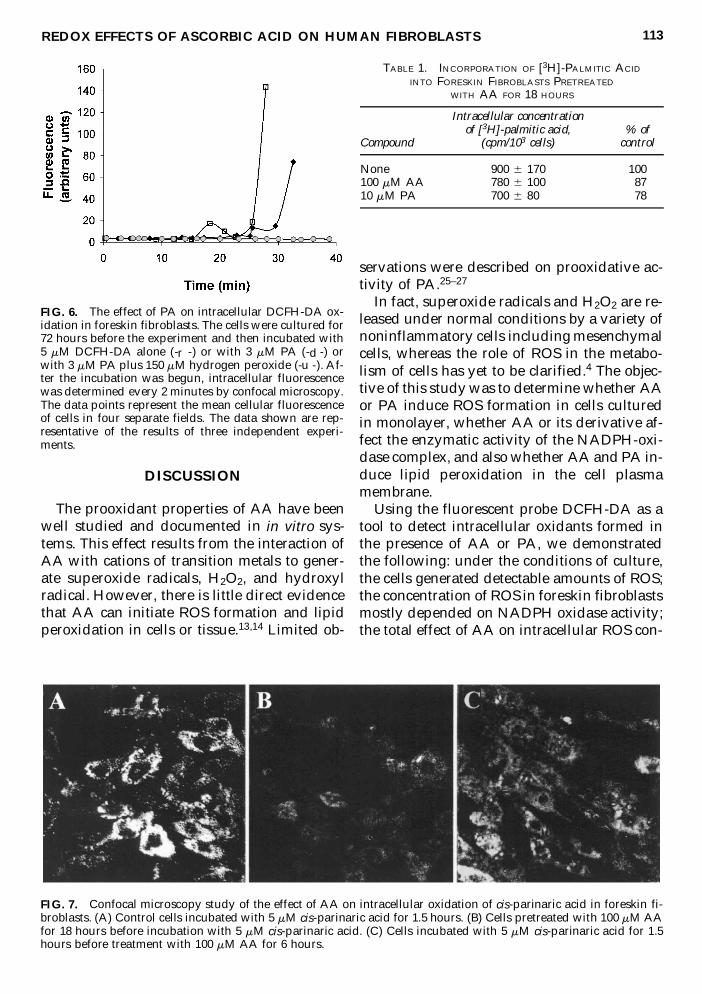
DISCUSSION
The prooxidant properties of AA have beenwell studied and documented in in vitro sys-tems This effect results from the interaction ofAA with cations of transition metals to gener-ate superoxide radicals H2O2 and hydroxylradical However there is little direct evidencethat AA can initiate ROS formation and lipidperoxidation in cells or tissue1314 Limited ob-
servations were described on prooxidative ac-tivity of PA25ndash27
In fact superoxide radicals and H2O2 are re-leased under normal conditions by a variety ofnoninflammatory cells including mesenchymalcells whereas the role of ROS in the metabo-lism of cells has yet to be clarified4 The objec-tive of this study was to determine whether AAor PA induce ROS formation in cells culturedin monolayer whether AA or its derivative af-fect the enzymatic activity of the NADPH-oxi-dase complex and also whether AA and PA in-duce lipid peroxidation in the cell plasmamembrane
Using the fluorescent probe DCFH-DA as atool to detect intracellular oxidants formed inthe presence of AA or PA we demonstratedthe following under the conditions of culturethe cells generated detectable amounts of ROSthe concentration of ROS in foreskin fibroblastsmostly depended on NADPH oxidase activitythe total effect of AA on intracellular ROS con-
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 113
FIG 6 The effect of PA on intracellular DCFH-DA ox-idation in foreskin fibroblasts The cells were cultured for72 hours before the experiment and then incubated with5 mM DCFH-DA alone (-r -) or with 3 mM PA (-d -) orwith 3 mM PA plus 150 mM hydrogen peroxide (-u -) Af-ter the incubation was begun intracellular fluorescencewas determined every 2 minutes by confocal microscopyThe data points represent the mean cellular fluorescenceof cells in four separate fields The data shown are rep-resentative of the results of three independent experi-ments
FIG 7 Confocal microscopy study of the effect of AA on intracellular oxidation of cis-parinaric acid in foreskin fi-broblasts (A) Control cells incubated with 5 mM cis-parinaric acid for 15 hours (B) Cells pretreated with 100 mM AAfor 18 hours before incubation with 5 mM cis-parinaric acid (C) Cells incubated with 5 mM cis-parinaric acid for 15hours before treatment with 100 mM AA for 6 hours
TABLE 1 INCORPORATION OF [3H]-PALMITIC ACID
INTO FORESKIN FIBROBLASTS PRETREATED
WITH AA FOR 18 HOURS
Intracellular concentrationof [3H]-palmitic acid of
Compound (cpm103 cells) control
None 900 6 170 100100 mM AA 780 6 100 8710 mM PA 700 6 800 78
centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115

centration depended on the cell growth condi-tions the intracellular ROS concentration in-creased in newly plated cells (24 hours) but itmoderately diminished in cells cultured for along time (72 hours) in cytoplasm membraneAA and PA caused lipid peroxidation
The reason for the discrepancy between theresponses in cells cultivated for 24 and 72 hoursis not clear It is possible that the newly-platedcells allowed exogenously generated ROS toenter freely into the cell This question was fur-ther investigated by removing the AA from theculture medium before measurement of ROS in24-hour cells In that experiment ROS forma-tion was not augmented by AA pretreatmentsuggesting that in the presence of AA exoge-nously generated ROS (probably H2O2) was theextra source (besides NADPH oxidase) of theintracellular ROS in the 24-hour cells
The reason for the induction of lipid peroxi-dation in membranes with a concomitant inhi-bition of cytosolic ROS formation by AA andPA is obscure A possible explanation is thatthe mechanism for lipid peroxidation by AA infibroblasts is not via ROS formation but by the formation of an ascorbatendashplasma mem-branendashiron complex as has been demonstratedin rat liver microsomes28 It is also possible thatthe decrease in intracellular ROS concentrationmay be explained by an inhibition of NADPHoxidase activity The residual cytosol ROS con-centration (even in the case of simultaneousROS generation by AA) is less than that in thecells without NADPH-oxidase inhibition PAstrongly inhibited intracellular DCFH-DA oxi-dation in cultured fibroblasts probably bystronger inhibition of NADPH oxidase activityA similar effect was described in a study of theeffect 6-O-acylated AA derivatives (but notAA) on NADPH-oxidase activity in polymor-phonuclear neutrophils2930
Further studies will be necessary to moreclearly define the redox properties of PA andthe mechanism of inhibition of NADPH-oxi-dasendashassociated ROS formation by PA
In conclusion the effect of AA or PA on theredox state of the cultured cells depends ontheir growth condition In long-time culturedcells both AA and PA demonstrated a doubleeffect they reduced intracellular ROS genera-tion but stimulated lipid peroxidation in cyto-
plasm membranes Therefore in cultured cellsAA and its derivatives may be involved in theregulation of cell metabolism acting simulta-neously as antioxidants in cytosol and asprooxidants in lipid membranes
ACKNOWLEDGMENTS
This work was supported by TAGRABiotechnology Ltd Israel and NIH GrantDK34151
REFERENCES
1 Halliwell B Free radicals reactive oxygen species andhuman disease Br J Exp Pathol 198970737ndash757
2 Buttke TM Sandstrom PA Oxidative stress as a me-diator of apoptosis Immunol Today 1994157ndash10
3 Sen CK Packer L Antioxidant and redox regulationof gene transcription FASEB J 199610709ndash720
4 Burdon RH Superoxide and hydrogen peroxide in re-lation to mammalian cell proliferation Free Radic BiolMed 199518775ndash794
5 Tsao CS An overview of ascorbic acid chemistry andbiochemistry In Vitamin C in Health and Disease(Packer L Fuchs J eds) Marcel Dekker New York1997 pp 25ndash58
6 Hitomi K Tsukagoshi N Role of ascorbic acid in mod-ulation of gene expression Subcell Biochem 19962541ndash56
7 Halliwell B Whiteman M Antioxidant and prooxi-dant properties of vitamin C In Vitamin C in Healthand Disease (Packer L Fuchs J eds) Marcel DekkerNew York 1997 pp 59ndash74
8 Stadtman ER Ascorbic acid and oxidative inactiva-tion of proteins Am J Clin Nutr 1991541125Sndash1128S
9 Chojkier M Houglum KP Solis-Herruzo J BrennerDA Stimulation of collagen gene expression by ascor-bic acid in cultured skin fibroblasts A role of lipidperoxidation J Biol Chem 198926416957ndash16962
10 Geesin J Hendricks LJ Falkenstein PA Gordon JSBerg RA Regulation of collagen synthesis by ascor-bic acid Characterization of the role of ascorbate de-pendent lipid peroxidation Arch Biochem Biophys1991290127ndash132
11 Geesin J Brown LJ Gordon JS Berg RA Regulationof collagen synthesis in human dermal fibroblasts incontracted collagen gels by ascorbic acid growth fac-tor and inhibitors of lipid peroxidation Exp Cell Res1993206283ndash290
12 Podmore ID Griffiths HR Herbert KE Mistry N Mis-try P Lunec J Vitamin C exhibits pro-oxidant prop-erties Nature 1998392559
13 Carr A Frei B Does vitamin C act as pro-oxidant under physiological conditions FASEB J 1999131007ndash1024
ROSENBLAT ET AL114
14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115

14 Poulsen HE Weimann A Salonen JT Nyyssonen KLoft S Cadet J Douki T Ravanat JL Does vitamin Chave a pro-oxidant effect Nature 1998395231ndash232
15 Rosenblat G Perelman N Katzir E Gal-Or S JonasA Nimni M Sorgente N Neeman I Acylated ascor-bate stimulates collagen synthesis in cultured humanforeskin fibroblasts at lower doses than does ascorbicacid Connect Tissue Res 199837303ndash311
16 Hedley D Chow S Flow cytometric measurement oflipid peroxidation in vital cells using parinaric acidCytometry 199213686ndash692
17 Meier B Radeke HH Selle S Younes M Sies H ReschK Habermehl GG Human fibroblasts release oxygenspecies in response to interleukin-1 or tumour necro-sis factor-a Biochem J 1989263539ndash545
18 Meier B Cross A Hancock JT Kaup FJ Jones OTGIdentification of superoxide-generating NADPH oxi-dase system in human fibroblasts Biochem J1991275241ndash245
19 Rosenkranz AR Schmaldienst S Stuhlmeier K ChenW Knapp W Zlabinger GJ A microplate assay forthe detection of oxidative products using 2979-dichlo-rofluorescin-diacetate J Immunol Methods 199215639ndash45
20 LeBel CP Ischiropoulos H Bondy SC Evaluation ofthe probe 2979-dichlorofluorescin as an indicator ofreactive oxygen species formation and oxidativestress Chem Res Toxicol 19925227ndash231
21 Royall JA Ischiropoulos H Evaluation of 2979-dichlo-rofluorescin and dihydrorhodamine 123 as fluores-cent probes for intracellular H2O2 in cultured en-dothelial cells Arch Biochem Biophys 1993302348ndash355
22 Kuypers FA van den Berg JJ Schalkwijk C Roelof-sen B Op den Kamp JAF Parinaric acid as a sensi-tive fluorescent probe for the determination of lipidperoxidation Biochim Biophys Acta 1987921266ndash274
23 Di Giulio A Saletti A Oratore A Bozzi A Monitor-ing by cis-parinaric fluorescence of free radical in-duced lipid peroxidation in aqueous liposome sus-pensions J Microencapsul 199613435ndash445
24 Hockenbery DM Oltvai ZN Yin X-M Milliman CLKorsmeyer SJ Bcl-2 functions in an antioxidant path-way to prevent apoptosis Cell 199375241ndash251
25 Naofumi M Kiyoka T Masanosuke T Effect of L-ascorbic acid and its palmitoyl derivatives on the per-oxidation of linolenic acid in aqueous solutions con-taining alcohol Bioelectrochem Bioenerg 198718163ndash170
26 Miwa N Yamazaki H Nagaoka Y Kageyama KOnoyama Y Matsui-Yuasa I Otani S Morisawa S Al-tered production of active oxygen species is involvedin enhanced cytotoxic action of acylated derivativesof ascorbate to tumor cells Biochim Biophys Acta1988972144ndash151
27 Tyrsina EG Rossikhina OG Abilev SK Tyrsin YuAInhibition of the bacterial mutagenicity of N-methyl-N9-nitro-N-nitrosoguanidine by ascorbic acid andascorbyl palmitate Mutat Res 199432181ndash87
28 Casalino S Sblano C Landriscina C A possible mech-anism for initiation of lipid peroxidation by ascorbatein rate liver microsomes Int J Biochem Cell Biol199628137ndash149
29 Baader WJ Hatzelmann A Ullrich V The suppres-sion of granulocyte functions by lipophilic antioxi-dants Biochem Pharmacol 1988371089ndash1098
30 Schmid E Figala V Ullrich V Inhibition of NADPH-oxidase activity in human polymorphonuclear neu-trophils by lipophilic ascorbic acid derivatives MolPharmacol 199445815ndash825
Address reprint requests toGennady Rosenblat
Department of Food Engineering and Biotechnology
Technion-Israel Institute of Technology32000 Haifa Israel
E-mail GennadyrPolyhealcoil
REDOX EFFECTS OF ASCORBIC ACID ON HUMAN FIBROBLASTS 115
Copyright © 2022 FDOKUMEN




















