
Effect of anisotonic cell-volume modulation on glutathione-S-conjugate release, t-butylhydroperoxide...
8
bur. J. Biochem. 209,437-444 (1992) (c> FEBS 1992 Effect of anisotonic cell-volume modulation on glutathione-S-conjugate release, t-butylhydroperoxidemetabolism and the pentose-phosphate shunt in perfused rat liver Nirmalendu SAHA', Barbara STOLL ', Floridn LANG' and Dieter HAUSSINGER' ' Medizinische Universitatsklinik Freiburg, Federal Republic of Germany PhysiologisLhes Institut der Universitat Innsbruck, Austria (Rcceivcd May 8iJuly 29, 1992) EJB 92 0635 1. Addition of 1 -chloro-2,4-dinitrobenzene to isolated perfused rat liver results in the rapid formation of its glutathione-S-conjugate [S-(2,4-dinitrophenyl)glutathione], which is released into both, bile and effluent perfusate. Anisotonic perfusion did not affect total S-conjugate formation, but release of the S-conjugate into the perfusate was increased (decreased) following hypertonic (hypotonic) exposure at the expense of excretion into bile. Stimulation of S-conjugate release into the perfusate following hypertonic exposure paralleled the time course of volume-regulatory net K + uptake. 2. Basal steady-state release of oxidized glutathione (GSSG) into bile was 1.30+0.12 nmol . g-i .mi"-' (n = 18) during normotonic (305 mOsmol/l) perfusion and was 3.8 f 0.3 nmol . g ' . min ' in the presence of t-butylhydroperoxide (50 pmol/l). Hypotonic exposure (225 mOsmol/ 1) lowered both, basal and ~butylhydroperoxide(50 pmol/l)-stimulated GSSG release into bile by 35% and 20%, respectively, whereas hypertonic exposure (385 mOsmol/l) increased. Anisotonic exposure was without effect on t-butylhydroperoxide removal by the liver. GSSG release into bile also decreased by 33% upon liver-cell swelling due to addition of glutamine plus glycine (2 mmol/l, each). 3. Hypotonic exposure led to a persistent stimulation 14C02production from [l-'4C]glucose by about 80%, whereas 14C02 production from [6-'4C]glucose increased by only 10%. Conversely, hypertonic exposure inhibited 14C02 production from [l-'4C]glucose by about 40%, whereas I4CO2 production from [6-i4C]glucose was unaffected. The effect of anisotonicity on 14C02 production from [l-i4C]glucose was also observed in presence of t-butylhydroperoxide (50 pmol/l), which in- creased I4CO2 production from [I -'4C]glucose by about 40%. 4. t-Butylhydroperoxide (50 pmol/l) was without significant effect on volume-regulatory K + fluxes following exposure to hypotonic (225 mOsmol/l) or hypertonic (385 mOsmol/l) perfusate. Lactate dehydrogenase release from perfused rat liver under the influence of t-butylhydroperoxide was increased by hypertonic exposure compared to hypotonic perfusions. 5. The data suggest that hypotonic cell swelling stimulates flux through the pentose-phosphate pathway and diminishes loss of GSSG under conditions of mild oxidative stress. Hypotonically swollen cells arc less prone to hydroperoxide-induced lactate dehydrogenase release than hyper- tonically shrunken cells. Hypertonic cell shrinkage stimulates the excretion of glutathione-S-conju- gates into the sinusoidal circulation at the expense of biliary secretion. Liver-cell volume is an important determinant of meta- bolic liver-cell function, and several known, but mechan- istically unclear, hormone and amino acid effects on hepatic metabolism recently found an explanation in hormone-in- duced and amino-acid-induccd cell-volume changes (for a re- view see [I]). Liver-cell volume is also an important regulator oftaurocholate excretion into bile [2]. Canalicular secretion of Corre.spondence to D. 1Iaussinge.r. Medizinische Universitiits- klinik, Hugstetterstrasse 55, W-7800 Freiburg, Federal Republic of Germany Fax: +49 761 2703205. Abbreviations. S-conjugate, S-(2,4-dinitrophenyl)glutathione; GSSG; oxidized glutathione; GSH; reduced glutathione; LD€I, lac- tate dehydrogenase. taurocholate is accomplished by an ATP-dependent transport system [3 - 51. Glutathione-S-conjugates are also transported by ATP-dependent systems across the canalicular membrane [6,7], and the question arises whether excretion of glutathione- S-conjugates into bile is affccted by cell volume. In perfused rat liver steady-state rates of glutathione-S-conjugate for- mation can be achieved by infusion of l-chloro-2,4- dinitrobenzene [8 - 111 which is enzymically conjugated with reduced glutathione (GSH) to form S-(2,4-dinitro- pheny1)glutathione [12]. Steady-state rates of oxidized glutathione (GSSG) formation can be achieved upon infusion of t-butylhydroperoxide, which is a substrate for glutathione peroxidase, but not for catalase [13, 141. GSSG and other
Transcript of Effect of anisotonic cell-volume modulation on glutathione-S-conjugate release, t-butylhydroperoxide...

bur. J . Biochem. 209,437-444 (1992) (c> FEBS 1992
Effect of anisotonic cell-volume modulation on glutathione-S-conjugate release, t-butylhydroperoxide metabolism and the pentose-phosphate shunt in perfused rat liver Nirmalendu SAHA', Barbara STOLL ', Floridn LANG' and Dieter HAUSSINGER' ' Medizinische Universitatsklinik Freiburg, Federal Republic of Germany
PhysiologisLhes Institut der Universitat Innsbruck, Austria
(Rcceivcd May 8iJuly 29, 1992) EJB 92 0635
1. Addition of 1 -chloro-2,4-dinitrobenzene to isolated perfused rat liver results in the rapid formation of its glutathione-S-conjugate [S-(2,4-dinitrophenyl)glutathione], which is released into both, bile and effluent perfusate. Anisotonic perfusion did not affect total S-conjugate formation, but release of the S-conjugate into the perfusate was increased (decreased) following hypertonic (hypotonic) exposure at the expense of excretion into bile. Stimulation of S-conjugate release into the perfusate following hypertonic exposure paralleled the time course of volume-regulatory net K +
uptake. 2. Basal steady-state release of oxidized glutathione (GSSG) into bile was 1.30+0.12 nmol
. g - i .mi"- ' (n = 18) during normotonic (305 mOsmol/l) perfusion and was 3.8 f 0.3 nmol
. g ' . min ' in the presence of t-butylhydroperoxide (50 pmol/l). Hypotonic exposure (225 mOsmol/ 1) lowered both, basal and ~butylhydroperoxide(50 pmol/l)-stimulated GSSG release into bile by 35% and 20%, respectively, whereas hypertonic exposure (385 mOsmol/l) increased. Anisotonic exposure was without effect on t-butylhydroperoxide removal by the liver. GSSG release into bile also decreased by 33% upon liver-cell swelling due to addition of glutamine plus glycine (2 mmol/l, each).
3. Hypotonic exposure led to a persistent stimulation 14C02 production from [l-'4C]glucose by about 80%, whereas 14C02 production from [6-'4C]glucose increased by only 10%. Conversely, hypertonic exposure inhibited 14C02 production from [l-'4C]glucose by about 40%, whereas I4CO2 production from [6-i4C]glucose was unaffected. The effect of anisotonicity on 14C02 production from [l-i4C]glucose was also observed in presence of t-butylhydroperoxide (50 pmol/l), which in- creased I4CO2 production from [I -'4C]glucose by about 40%.
4. t-Butylhydroperoxide (50 pmol/l) was without significant effect on volume-regulatory K + fluxes following exposure to hypotonic (225 mOsmol/l) or hypertonic (385 mOsmol/l) perfusate. Lactate dehydrogenase release from perfused rat liver under the influence of t-butylhydroperoxide was increased by hypertonic exposure compared to hypotonic perfusions.
5. The data suggest that hypotonic cell swelling stimulates flux through the pentose-phosphate pathway and diminishes loss of GSSG under conditions of mild oxidative stress. Hypotonically swollen cells arc less prone to hydroperoxide-induced lactate dehydrogenase release than hyper- tonically shrunken cells. Hypertonic cell shrinkage stimulates the excretion of glutathione-S-conju- gates into the sinusoidal circulation at the expense of biliary secretion.
Liver-cell volume is an important determinant of meta- bolic liver-cell function, and several known, but mechan- istically unclear, hormone and amino acid effects on hepatic metabolism recently found an explanation in hormone-in- duced and amino-acid-induccd cell-volume changes (for a re- view see [I]). Liver-cell volume is also an important regulator oftaurocholate excretion into bile [2]. Canalicular secretion of
Corre.spondence to D. 1Iaussinge.r. Medizinische Universitiits- klinik, Hugstetterstrasse 55, W-7800 Freiburg, Federal Republic of Germany
Fax: +49 761 2703205. Abbreviations. S-conjugate, S-(2,4-dinitrophenyl)glutathione;
GSSG; oxidized glutathione; GSH; reduced glutathione; LD€I, lac- tate dehydrogenase.
taurocholate is accomplished by an ATP-dependent transport system [3 - 51. Glutathione-S-conjugates are also transported by ATP-dependent systems across the canalicular membrane [6,7], and the question arises whether excretion of glutathione- S-conjugates into bile is affccted by cell volume. In perfused rat liver steady-state rates of glutathione-S-conjugate for- mation can be achieved by infusion of l-chloro-2,4- dinitrobenzene [8 - 111 which is enzymically conjugated with reduced glutathione (GSH) to form S-(2,4-dinitro- pheny1)glutathione [12]. Steady-state rates of oxidized glutathione (GSSG) formation can be achieved upon infusion of t-butylhydroperoxide, which is a substrate for glutathione peroxidase, but not for catalase [13, 141. GSSG and other

438
glutathione-S-conjugates compete for excretion into bile [I 51, but GSSG excretion into bile is apparently not saturable [13, 161. A close linear relationship was obtained between the intraccllular GSSG concentration and its excretion into bile under different conditions of cxperimental oxidative stress [16], buggesting that GSSG release into bile can be used as an indicator for oxidative stress in perfused rat liver [13]. t-Butylhydroperoxide leads to an oxidation of celldar NADPH [13] and stimulates flux through the pentose-phos- phate pathway [14, 171, thereby providing NADPH for glutathione-reductase-catalyzed regeneration of GSH. We re- port on the effect of anisotonic exposure on glutathione-S- conjugate release, glucose oxidation and hydroperoxide tox- icity.
MATERIALS AND METHODS
Liver perfusion
Livers from male Wistar rats (200-250 g), fed ad libitum on stock diet (Altromin), were perfused as described pre- viously [18] in a non-recirculating manner with the bicarbon- ate-buffered Krebs-Henseleit saline plus L-lactate (2.1 mmul/l) and pyruvate (0.3 mmol/l) In experiments on 14C02 pro- duction from 14C-labeled glucose, the influent perfusate also contained glucose (5 mmol/l). The perfusate was gassed with 0 2 / C 0 2 (95:5, by vol.); the temperature was 37°C. The perfusate osmolarity was 305 mOsmol/l (normotonic con- dition). Anisotonic-perfusion conditions were achieved by corresponding changes of the NaCl concentration in influent. t-Butylhydroperoxide and 1 -chloro-2,4-dinitrobenzene were added to influent perfusate by use of precision micropumps.
Determinations
Bile was sampled at 1-min intervals. Bile flow was deter- mined from the secreted bile, assuming a specific density of bile of 1 g/ml. GSSG in bile was measured according to [19]. Effluent perfusate and bile were assayed for the S-conjugate of l-chloro-2,4-dinitrobenzene [(S-(2,4-dinitrophenyl)gluta- thione] by absorbance at 334 nm according to [12]. Lactate dehydrogenase (LDH) activity and lactate and pyruvate in effluent perfusate were determined as described in [20]. t- Butylhydroperoxide in the perfusate was measured spectrophotometrically according to [14].
I4CO2 production from [l -14C]glucose or [6-' 4C]glucose was determined as described previously [21]. Perfusate con- taining [l-14C]glucose or [6-14C]glucose (120 - 180 kBq/l) was pooled over I-min periods and collected in stoppered conical flasks. 14C02 was trapped in phenylethylamine and radioac- tivity was determined by scintillation spectrometry. Rates of 14C02 production were calculated on the basis of the specific radioactivity of glucose in influent perfusate.
Anisotonicity-induced and amino acid-induced changes in cell volunie were determined as described recently in detail [22, 231 as changes in the intracellular water space. The latter was determined from the [3H]inulin and [14C]urea spaces in perfused rat liver [22, 231.
The effluent K + concentration was continuously moni- tored with a K +-sensitive electrode (Radiometer, Munich), net K + uptake or release was determined by planimetry of areas under curve and calibration was made by infusion of known amounts of potassium chloride. When present, a base- line drift was taken into account.
A
128, L e
LO 65 , 90 Pertusion time irninl
5 5'
Lo 65 90 Perfusion time h i n l
Fig.l. Effect of hypertonic (A) and hypotonic (B) exposure on glutathione-S-conjugate formation from I-chloro-2,4-dinitrobenzene (30 pmol/l) and K + concentration in effluent. Infusion of 1-chloro-2,4- dinitrobenzene (CDNB) was started after 41 rnin of perfusion: the release of the S-conjugate (b, e) into bile (0) and perfusate ( 0 ) is shown. Total S-conjugate release reflects the sum of release into bile and perfusate (a, d). Hypertonic (385 mOsmol/l) and hypotonic (225 mOsmol/l) exposure was performed during the times indicated by adding or removing 40 mmol/l NaCl from the perfusion medium, respectively. Data are given as mean +. SEM and are from four different pcrfusion expcnmcnts for Fig. 1A and B, respectively. (c. f, A representative K + tracing from one of these experiments is shown.
Statistics
mean & SEM. Data from different perfusion experiments are given as
Materials
[l-'4C]Glucose and [6-14C]glucose were from Amersham Buchler (Braunschweig). L-Lactic acid was from Roth (Karlsruhe). l-Chloro-2,4-dinitrobenzene, rhein, t-butylhy- droperoxide, glutathione reductase and glutathione peroxi- dase were from Sigma (Munich). All othcr chemicals were from Merck (Darmstadt).
RESULTS
Effect of anisotonic exposure on glutathione-S-conjugate excretion
Addition of 30 pmol/l l-chloro-2,4-dinitrobenzene to in- fluent perfusate of perfused rat liver leads to the rapid forma- tion of the corresponding glutathione-S-conjugate, which is readily distinguishable spectrophotometrically from the unconjugated parent compound [9]. Total S-conjugate release from the liver approached a steady-state rate after about 15 min of infusion, which was about 80% of the rate of 1- chloro-2,4-dinitrobenzene infusion (Fig. 1 A, B). After 15 min, about 20% of the S-conjugate was released into the perfusate, the remainder into bile (Fig. lA, B). Steady-state total

439
S-conjugate release was not significantly affected when the perfusate osmolarity was increased from 305 mOsmol/l to 385 mOsmol/l by addition of 40 mmol/l NaCl to influent perfusate (Fig. 1 A). Hypertonic exposure, however, resulted in a marked stimulation of S-conjugate release into the perfusate, whereas excretion into bile decreased (Fig. IA). The time- course of hypertonicity-stimulated S-conjugate release into the perfusate roughly resembled that of the accompanying volume-regulatory net K + uptake, which occurs following hyperosmotic exposure (Fig. 1A) and which has been charac- terized recently in detail [24]. Following restoration of the normotonic condition, biliary excretion of the S-conjugate was transiently increased, whereas a slight decrease in ex- cretion into the perfusate was observed. Thereafter, total S- conjugate release dccreased probably due to the depletion of cellular glutathione during the course of the experiments. When, on the other hand, livers were exposed to hypotonic perfusion fluid (225 mOsmol/l), S-conjugate release into bile transiently increased (Fig. 1 B), whereas the release into the perfusate slightly decreased. When the normotonic condition was restored, i.e. the perfusate osmolarity raised from 225 mOsmol/l to 305 mOsmol/l, the volume-regulatory net K ' uptake mas also accompanied by a stimulation of S- conjugate relcase into the perfusatc. Thesc data suggest that cell-volume-regulatory increase affects the partitioning be- tween biliary and sinusoidal excretion of the S-conjugate, but has no effect on the rate of total S-conjugate formation.
In another series of experiments, livers were exposed to either hypotonic (225 mOsmol/l) or hypertonic (385 mOsmol/l) perfusion fluid, and after completion of volume-regulatory ions fluxes, i.e. 15 min after the onset of anisotonic exposure, l-chloro-2,4-dinitrobenzene (30 pmol/l) was infused. In these experiments. total (bile plus perfusate) S-conjugate formation reached a steady state within 10-15 min. Total S-conjugatc formation was 80f5% ( n = 4) and 8 2 5 3 % (n = 5) of the added l-chloro-2,4-dinitrobenzene dose in hypotonic and hypertonic perfusions, respectively. Whereas S-conjugate re- lease into the perfusate reached a steady state within 15 min in hypotonic perfusions, it continued to increase in hypertonic perfusions. After a 30-min infusion period 22 f 4% (n = 4) of total S-conjugate was released into the perfusate in hypotonic perfusions, whereas it was 40*8% (n = 5) in hypertonic perfusions. These data show that S-conjugate excretion into the perfusate is increased at the expense or biliary excretion in hypertonically shrunken livers.
Effect of anisotonic exposure on GSSG release into bile and 14C02 production from [l-14C]glucose and [6-L4C]glucose
As shown in Fig. 2 , during normotonic control perfusions, GSSG release into bile was approximately 1 nmol . g - l .min-l , Exposure to hypotonic perfusion fluid (225 mOsmol/l), decreased GSSG release into bile by about 35*2O/0 (n = 4), whereas GSSG release was increased by 375 1% (n = 3) during hypertonic perfusion (385 mOsmol/l). GSSG release into the perfusate was not detectable. The effect of anisotonicity on biliary GSSG excretion was reversible upon restoration of normotonic perfusion conditions (Fig. 2). It should be noted that the effect of anisotonicity on GSSG excretion into bile persisted throughout anisotonic exposure and was also observed after completion of volume-regulatory K f fluxes, which were largely terminated after about 10 min (compare K + movements depicted in Fig. 1). After completion of volume-regulatory K + fluxes, bile flow was not significantly different in normotonic and hypotonic perfusions
b 2.01 2.01
1 70 100 0 70 100
I2 ol* PerfuLion time iminl Perfusion time imin)
Fig. 2. Effect of (A) hypertonic (385 mOsmol/l) and (B) hypotonic (225 mOsmol/l) exposure on GSSG release into bile. Livers were per- fused with normotonic (305 mOsmol/l) media containing lactate (2.1 mmol/l) and pyruvate (0.3 mmol/l) and hypertonic and hypotonic perfusion conditions were introduced during the time periods indi- cated due to addition or removal of corresponding amounts of NaCl to or from thc perfusion medium, rcspcctively. Data are gvcn as mean f SEM and are from three or four different experiments for each condition.
(1.26 _+ 0.08 pi . g-' .min-l vs. 1.30f0.11 pl . g-' . min-', respectively), whereas bile flow decreased by 9 +4% (n = 3) following hypotonic exposure.
In normotonic (305 mOsmol/l) perfusions 14C02 pro- duction from [l-'4C]glucose ( 5 mmol/l) was 13.0k 2.0 nmol . g- . min-' (n = 10). Hypotonic exposure stimu- lated I4CO2 production from [l-'4C]glucose by 83 k 13% (n = 4), whereas hypertonic exposure inhibited by 3557% (n = 4; Fig. 3A). On the other hand, I4CO2 production from [6-'4C]glucosewas8.9~2.1 nmo1.g-' .min-l(n = 4)during normotonic (305 mOsmol/l) control perfusions and was largely unaffected by anisotonic exposure (Fig. 3A, Table 1). The effects of anisotonicity on I4CO2 production from [l- ''C]glucose/[6-14C]glucose persisted throughout hypotonic exposure and were fully reversible upon restoration of normotonic perfusion conditions (Fig. 3A). Whereas the lac- tate-plus-pyruvate balance across the liver was close to zero during normotonic perfusions, there was a net uptake (release) of lactate plus pyruvate during hypotonic (hypertonic) per- fusions (Fig. 3B). These data suggest that hypotonic cell swell- ing augments the net flux of three-carbon precursors into glucose (i.e. inhibits glycogenolysis and glycolysis as reported earlier [25], in addition to a stimulation of gluconeogenesis), whereas simultaneously glucose flux through the pentose phosphate pathway is increased. Conversely, hypertonic cell shrinkage apparently inhibits glucose flux through the pen- lose-phosphate pathway.
Effect of glutamine and glycine on GSSG release into bile and on 14C02 production from [l-14C]glucose
When liver-cell swelling was induced during normotonic perfusions by addition of glutamine plus glycine (2 mmol/l each), steady-state GSSG relcase into bile also decreased by 33 & 3% (n = 3), as observed following hypotonic cell swell- ing. The decrease of GSSG release into bile following addition of glutamine and glycine was accompanied by a decrease in bile flow by 1052% (a = 7). Addition of glutamine and glycine (2 mmol/l, each) also stimulated 14C02 production from [I- ' 4C]glucose (Table 1). Accordingly, both, hypo- tonicity-induced and amino-acid-induced cell swelling exerted similar effects on GSSG release into bile and 14C0, pro-

440
H Y P O T O N I C
A
H Y P E R T O N I C
[1-l4C1 GLUCOSE or 16-l'Cl GLUCOSE
- . 40 7 0 100 130
PER FUSION TIME Win)
40 70 100 130
Perfusion T i m e ( m i n )
Fig. 3. Effect of anisotonic exposure on I4C0, in emucnt derived from [l-14Cjglucose or [6-'4Cjglucosc (A) and lactate-plus-pyruvate balance across the liver (R). Livers were perluscd with normotonic (305 mOsmol/l) media containing lactate (2.1 mmoljl), pyruvate (0.3 mmoljl) and [l-'4CJglucose (a) or [6-'4C]glucose (0; 5 mmol/l). Hypertonic (385 mOsmol/l) and hypotonic (225 mOsmol/l) pcrfusion conditions were introduced during the times indicated due to addition or removal of corresponding amounts of NaCl to or from the per- fusion medium, respcctively. Lactate plus pyruvatc balance is the effluent-minus-influent concentration difference for lactate plus pyruvate. Accordingly positive values indicate net releasc of lactate plus pyruvatc from thc liver. Data arc given as mean SEM and are from 4 different perfusion experiments for each condition.
duction from [I-'4C]gIucose (Table I), suggesting a role of cell-volume changes in mediating these metabolic responses.
Effect of anisotonicity on the metabolism of t-butylhydroperoxide
Up to portal t-butylhydroperoxide additions of 2 pmol . g- ' . min- ', single-pass uptake of t-butylhydro-
peroxidc by the liver was more than 95% and was not affected during anisotonic exposure (not shown). GSSG release into bile was stimulated by infusion of t-butylhydroperoxide (50 pmol/l; Fig. 4), in line with previous reports [13]. Hyper- tonic (385 mOsmol/l) exposure transiently decreased GSSG release into bile, in parallel with a decrease in bile flow (Fig. 4A). Conversely, a transient increase of both, bile flow and GSSG releasc into bile was observed following hypotonic exposure (Fig. 4B). However, after completion of volume- regulatory K + fluxes, bile flow returned to the normotonic base line despite continuing anisotonicity. However, the steady-state release of GSSG into bile was significantly elev- ated by 16 & 1 % (n = 4) throughout hypertonic exposure and was decreased by 20+2% (n = 4) throughout hypotonic exposure. t-Butylhydroperoxide (50 pmol/l) slightly, but sig- nificantly increased I4CO2 production from [l-14C]glucose in line with previous reports on a stimulation of glucose oxi- dation via the pentose-phosphate pathway [14, 17, 27, 281. Further exposure to hypotonic (hypertonic) media persistently increased (decreased) I4CO2 release from added [l- ''C]glucose (Fig. 4), as also observed in the absence of t-butyl- hydroperoxide (compare Fig. 3A).
Effect of anisotonicity on t-butylhydroperoxide-induced cell injury
Sustained oxidative stress is known to impair cell viability, frequently assessed as LDH release from the cells. As shown in Fig. 4, LDH release into effluent perfusate was markedly stimulated by t-butylhydroperoxide (50 pmoljl) when the livers were transiently exposed to hypertonic media (Fig. 4A), but not to hypotonic media (Fig.4B). As a control, in the absence of the hydroperoxide, LDH activity in effluent perfusate was 1.9 f 0.2 U/1 (n = 3) after 30 min of hypertonic exposure (not shown), indicating that stimulation of LDH release during hypertonic exposure in Fig. 4A was dependent on the presence of the hydroperoxide. It should be noted that stimulation of LDH release in Fig. 4A was accompanied by a net K + uptake by the liver due to a volume-regulatory increase following hypertonic exposure. This suggests that despite stimulation of LDH release, the cells were still able to maintain their K balance. In addition, volume-regulatory net K f up- take (release) following hypertonic (hypotonic) exposure was not affected by t-butylhydroperoxide (50 pmoljl): in the ab- sence and presence of t-butylhydroperoxide. net K + uptake following hypertonic (385 mOsmol/l) exposure was 12.9 + 0.4 pmol/g (n = 4) and 12.7 f 0.3 pmol/g ( n = 3), respectively, and net K ' release following hypotonic (225 mOsmol/l) ex- posure was 12.7k0.5 pmol/g (n = 8) and 12.9k0.1 pmol/ g (n = 4), respectively. As shown in Fig. 4, addition of t- butylhydroperoxide (50 pmol/l) led to a small net K' release from the liver of 0.8+0.1 pnol/g (n = 5), which was com- pleted within 8 min. Nct K f release under the influence of t- butylhydroperoxide has also been observed by others [13, 281 and is due to an opening of Ba2+-sensitive K + channels (C. Hallbrucker and D. Haussinger, unpublished result).
r-Butylhydroperoxide(0.2 mmol/l)-induced LDH release into the perfusate was also significantly enhanced when the hydroperoxide was added 15 min after onset of hypertonic (385 mOsmol/l) perfusion. Aftcr a 40-min exposure to t-butyl- hydroperoxide (0.2 mmol/l), LDH activity in effluent pcrfusate was 69+4 (n = 5), 121f5 (n = 3) and 204t39 (n = 4) U/l during hypotonic (225 mOsmol/l), normotonic (305 mOsmol/l) and hypertonic (385 mOsmol/l) perfusions, respectively. As a control, after a 60-min hypertonic perfusion

44 1
Table 1. Effect of anisotonic exposure and amino acid addition on steady-state GSSG release into bile, l4COZ production from 14C-labeled glucose and cell volume. Livers were perfused with a medium containing [l -'4C]glucose or [6-'4C]glucose ( 5 mmol/l), lactate (2.1 mmol/l) and pyruvate (0.3 mmol/l). Steady-state GSSG release under control conditions was 1.30&0.12 nmol. g- ' 'min-' (n = 18) and was set to 100% in the individual perfusion experiment. Likewise, I4CO2 production from [l-'4C]glucose and [6-14C]glucose (5 mmol/l) was 13.0 f 2.0 nmol . g- ' . min-' (n = 10) and 8.9 f 2.1 nmol g- . min ~ ' (n = 4) in normotonic control perfusions and was set to 100% in the individual perfusion experiment. Data on anisotonicity-induced and amino acid-induced changes in GSSG release into bile and 14C02 production from ''C-labeled glucose are gven as percentages. Hypotonic and hypertonic exposure was performed by adding or removing corresponding amounts of NaCl in influent perfusate. Cell volume was determined as intracellular water space, as described in [22]. In normotonic control perfusions, cell volume was 559 7 pl/g (n = 88) [23] and was set to 100% in each perfusion experiment. Glulamine and glycine were added at 2 mmol/l each. Data on cell volume are taken from previous studies [22, 231, in which identical perfusion conditions werc used, except that glucose was absent in influent perfusate; however, as shown previously. glucose ( 5 mmolil) is without effect on cell volume [26]. Xcgativc values indicate decrease, positive valucs increase of the respective parameters.
Condition Change (n) in
GSSG release cell volume 14C02 production from
[~-'~C]glucose [6-' 4C]glucose
Yo
Hypotonic (225 mosmol) - 35 f 2 (4) + 18.9 k 2.6 (4) +83 f 13 (4) + 1 0 f S ( 4 ) Hypertonic (385 mosmol) +37 f 1(3) -12.1 * 0.2 (3) -35* 7 (4) + 3 Ik 4 (4) Glutamine/glycine (2 mmol/l each) -33 f 3 (3) +12.1 f 1.4 (3) +45 f 19 (3) + 4 f 1 (3)
without t-butylhydroperoxide, LDH activity in effluent perfusate was only 3.5 0.5 Ujl (n = 3), suggesting that hyper- tonic exposure per se had little effect on LDH release from the liver. Although hydroperoxide-induced LDH release was much more pronounced in hypertonic than in hypotonic per- fusions, neither the time course nor the total amount of GSSG released into bile and perfusate wcre significantly different in hypotonic and hypertonic perfusions, respectively. Aftcr a 40-min exposure to t-butylhydroperoxide (0.2 mmol/l), GSSG release from the liver was 2 7 k 1 nmol ' . min-' (n = 3) and 24+4 nmol . g p l . minpl (n = 3) in hypertonic and hypotonic perfusions, respectively; values similar to those re- ported in [14]. Interestingly, LDH release under the influence of t-butylhydroperoxide (0.2 mmol/l) was not accompanied by a detectable net K + release from the livers (not shown), suggesting that LDH release was not due to a general increase in membrane permeability.
DISCUSSION
Anisotonicity and 14C02 production from I l-'4C]glucose/[6-'4C]glucose
During metabolism of [l-14C]glucose, labeled C 0 2 is liber- ated by decarboxylation reactions in the pentose-phosphate cycle, as well as in the citric-acid cycle. In the latter case, 14C02 is produced from 14C-labeled pyruvate derived from glycolysis, which in our experiments was markedly diluted by added unlabeled lactate (2.1 mmol/l) and pyruvate (0.3 mmol/l) in order to minimize I4CO2 production via the citric-acid cycle or utilization of l4C-labc1ed pyruvate for other reactions. In addition, the contribution of the citric-acid cycle to COz, production from [l-'4C]glucose was estimated from ' 'C02 production from [6-14C]glucose, which will not pick up C 0 2 production via the pentose-phosphate cycle. Hypotonic ex- posure only slightly increased l4CO, production from [6- 14C]glucose, but markedly stimulated 14C02 production from [l-14C]glucose (Fig. 3, Table 1). These findings suggest a stimulation of flux through the pentose-phosphate pathway
following hypotonic cell swelling. Conversely, hypertonic cell shrinkage reduced 14C02 production from [l-14C]glucose, but had no effect on 14C02 production from [6-'4C]glucose, pointing to a diminished flux through the pentose-phosphate cycle following hypertonic cell shrinkage. These effects of anisotonicity may add to other recently described effects of cell volume on hepatic carbohydrate metabolism, such as inhi- bition of glycogenolysis and stimulation of glycogen synthesis
Stimulation of flux through the pentose-phosphate shunt by hypotonic cell swelling could either reflect a direct effect on the shunt or an effect secondary to an increased NADPH consumption by other cell-volume-sensitive reactions. In this respect, lipogenesis and acetyl-CoA carboxylase flux were shown to be activated following hypotonic cell swelling 131, 321. Furthermore, the NADPH system may be linked via the action of transhydrogenase to cellular NADH systems. Hypotonic cell swelling shifts both the cytosolic and the mito- chondrial NADH/NAD + systems to more oxidized states, as evidenced in experiments using lactatelpyruvate and acetoacetate/3-hydroxybutyrate as redox-indicator metab- olite couples (not shown). T h s hypotonicity-induced oxi- dation is probably due to a stimulation of the respiratory chain resulting from mitochondria1 swelling (for a review see 1341). In order to obtain more insight into an anisotonicity modulation of flux through transhydrogenase reaction. exper- iments with the transhydrogenasc inhibitor rhein (66 pmol/l) were performed, however, this compound itself produced marked side effects: GSSG release into bile and 14C02 pro- duction from [l-14C]glucose wcre stimulated by rhein (not shown). Accordingly, this issue remains unsettled.
[25, 29 -331.
Anisotonicity and glutathione turnover
We have recently shown that hypotonic cell swelling stimu- lates the release of GSH into the perfusate (see Fig. 1 in [35]); as shown here, this is accompanied by a decreased GSSG rclease into bile (Fig. 2). Also when t-butylhydroperoxide (50 pmol/l) was present, GSSG release into bile decreased following hypotonic cell swelling and increased following hy-
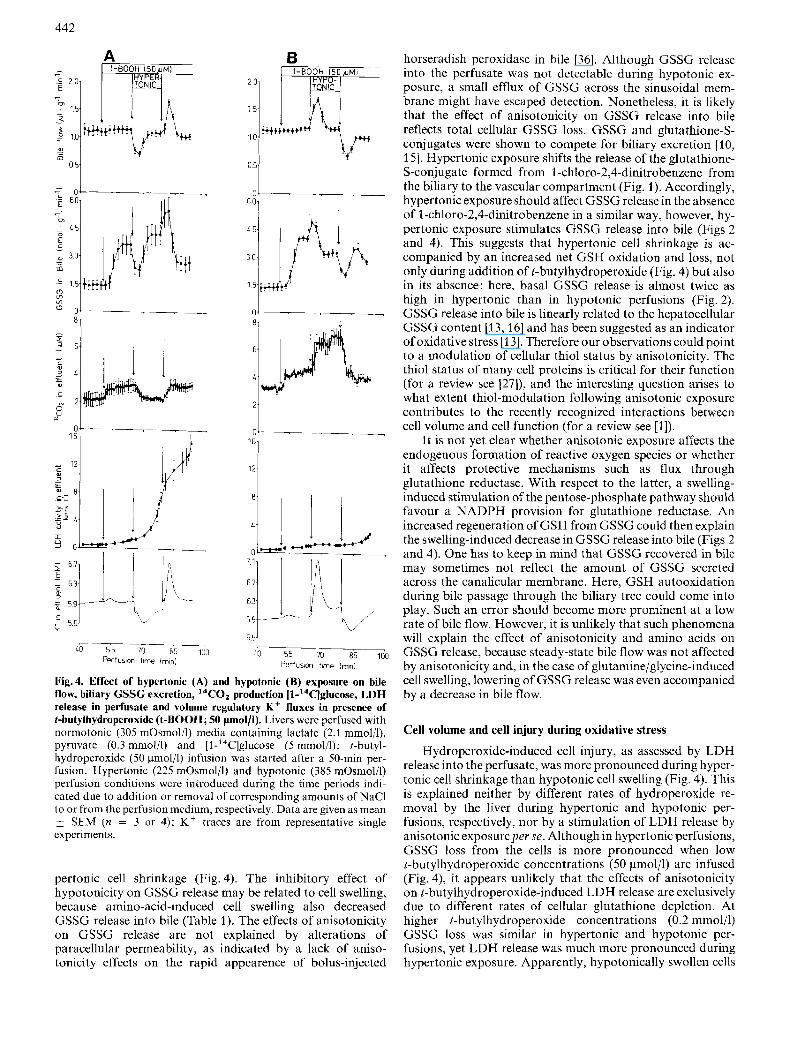
442
n - n1 I 1
21
26.
12-
:I 1 ...1./ 0-
I I 'I 1-2 I I 0-
-1
60 55 70 85 1!10 Perfusion time imin]
-- :n 55 'JO 85 loo Perfusion tome (mm)
Mg.4. Effect of hypertonic (A) and hypotonic (B) exposure on bile flow, biliary GSSC excretion, I4CO2 production [l-'4C]glucose, LDH release in perfusate and volume regulatory K + fluxes in presence of t-butylhydroperoxide (t-BOOII; 50 pmoI/l). Livers were perfused with normotonic (305 mOsmol/l) media containing lactate (2.1 mmolil), pyruvate (0.3 mmol/l) and [l-'4C]glucose ( 5 mmol/l); I-hutyl- hydroperoxide (50 ~mol/ l ) infusion was started after a 50-min per- fusion. Hypertonic (225 mOsmol/l) and hypotonic (385 mOsmol/l) perfusion conditioiis wcre introduced during the time periods indi- cated due to addition or removal of corresponding amounts of NaCl lo or from the perfusion mcdium, rcspectively. Data are gven as mean
SEM (n = 3 or 4); K + traces are from representative single experiments.
pertonic cell shrinkage (Fig. 4). The inhibitory effect of hypotonicity on GSSG release may be related to cell swelling, because amino-acid-induced cell swelling also decreased GSSG release into bile (Table 1). The effects of anisotonicity on GSSG release are not explained by alterations of paracellular permeability, as indicated by a lack of aniso- tonicity effects on the rapid appearence of bolus-injected
horseradish peroxidase in bile [36]. Although GSSG release into the perfusate was not detectable during hypotonic ex- posure, a small efflux of GSSG across the sinusoidal mem- brane might have escaped detection. Nonetheless, it is likely that the effect of anisotonicity on GSSG release into bile reflects total cellular GSSG loss. GSSG and g1utathione-S- conjugates were shown to compete for biliary excretion [lo, 151. Hypertonic exposure shifts the release of the glutathione- S-conjugate formed from I -chloro-2,4-dinitrobenzene from the biliary to the vascular compartment (Fig. 1). Accordingly, hypertonic exposure should affect GSSG relcase in the absence of 1 -chloro-2,4-dinitrobenzene in a similar way, however, hy- pertonic exposure stimulates GSSG release into bile (Figs 2 and 4). This suggests that hypertonic cell shrinkage is ac- companied by an increased net GSH oxidation and loss, not only during addition of t-butylhydroperoxide (Fig. 4) but also in its absence: here. basal GSSG release is almost twice as high in hypertonic than in hypotonic perfusions (Fig. 2). GSSG release into bile is linearly related to thc hepatocellular GSSG content [13,16] and has been suggested as an indicator of oxidative stress [13]. Therefore our observations could point to a modulation of cellular thiol status by anisotonicity. The thiol status o f many cell proteins is critical for their function (for a review see [27]), and the interesting question arises to what extent thiol-modulation following anisotonic exposure contributes to the recently recognized interactions between cell volume and cell function (for a review see [I]).
It is not yet clear whether anisotonic exposure affects the endogenous formation of reactive oxygen species or whether it affects protective mechanisms such as flux through glutathione reductase. With respect to the latter, a swelling- induced stimulation of the pentose-phosphate pathway should Favour a NADPH provision for glutathione reductase. An increased regeneration of GSH from GSSG could then explain the swelling-induced decrease in GSSG release into bile (Figs 2 and 4). One has to keep in mind that GSSG rccovercd in bilc may sometimes not reflect the amount of GSSG secreted across the canalicular membrane. Here, GSH autooxidation during bilc passage through the biliary tree could come into play. Such an error should become more prominent at a low rate of bile flow. However, it is unlikely that such phenomena will explain the effect of anisotonicity and amino acids on GSSG release, because steady-state bile flow was not affected by anisotonicity and, in the case of glutamine/glycine-induced cell swelling, lowering of GSSG release was even accompanicd by a decreasc in bile flow.
Cell volume and cell injury during oxidative stress
Hydroperoxide-induced cell injury, as assessed by LDH release into the perfusatc, was more pronounced during hyper- tonic cell shrinkage than hypotonic cell swelling (Fig. 4). This is explained neither by different rates of hydroperoxide re- moval by the liver during hypertonic and hypotonic per- fusions, respectively, nor by a stimulation of LDH release by anisotonic exposure per se. Although in hypertonic perfusions, GSSG loss from the cells is more pronounced when low l-butylhydroperoxide concentrations (50 pmol/l) are infused (Fig. 4), it appears unlikely that the effects of anisotonicity on t-butylhydroperoxide-induced LDH release are exclusively due to different rates of cellular glutathione depletion. At higher t-butylhydroperoxide concentrations (0.2 mmolil) GSSG loss was similar in hypertonic and hypotonic per- fusions, yet LDH release was much more pronounced during hypertonic exposure. Apparently, hypotonically swollen cells

443
are less susceptible to hydroperoxide-induced membrane dam- age. The reason for this is unclear, but may relate to different degrees of membrane stretching and multiple effects of aniso- tonicity on the cytoskeleton (C. Stournaras, B. Stoll and D. Haussinger, unpublished observation) and hepatocellular metabolism (for a review see [l]). t-Butylhydroperoxide per- turbs cellular CaZ+ homeostasis and leads to blebbing of the hepatocyte surface (for a review see [37]). Theoretically enhanced bleb formation, bleb dissociation and rupture in hypertonic perfusions could contribute to increased LDH re- lease (Fig. 4), however, anisotonic exposure of rat hepatocytes was shown to be without effect on bleb formation during both, control conditions and chemical hypoxia [38]. Cell swelling due to impairment of Na+/K+-ATPase, in response to hypoxia and various toxins, was reported to precede cell death [38 -421, and the beneficial effect of hypertonic exposure on ischemia/reperfusion injury in kidney and heart was attributed to a reversal of cell swelling with improvement of vascular perfusion [41, 431. In hepatocytes, however, cell swelling did not promote the onset of cell death following chemical hypoxia; cell swelling was not a prerequisite for bleb forma- tion, and the protective effmt of mannitol was due to antioxi- dant activity but not due to its osmotic properties [38]. How- ever, as shown in this study, hydroperoxide-induced LDH leakage is considerably less from hypotonically swollen cells than from hypertonically shrunken cells. This suggests that cell swelling may exert beneficial effccts during early stages of oxidative cell injury probably due to swelling-induced alter- ations in hepatic metabolism (for a review see [l]).
Dr Nirmalendu Saha is recipient of a fellowship from the Deutsche Akademischer Austuuschdienst. This study was supported by Deutsche For,schungsgemrinsrhujt through Srinde~orsc.hungshereic.E2 154 and the Gottfried- Wilhelm-Leihniz-Prize, the Fond7 der Chemischen lndustrie and the Schilling Stiftung.
REFERENCES 1 . Haussinger, D. & Lang, F. (1991) Cell volume in the regulation of
hepatic function: a mechanism for metabolic control, Biochinz. Bioplzps. Acta 1071, 331 -350.
2. Hallbrucker, C.. Lang, I?’., Gerok, W. & Haussinger D. (1992) Cell swelling increases bile flow and taurocholate excretion into bile in isolated perfused rat liver, Biochem. J. 281, 593 - 595.
3. Adachi, Y., Kobayashi, H., Kurumi, Y . , Shouji, M., Kitano, M. & Yamamoto, T. (2991) ATP-dependent taurocholate trans- port by rat liver canalicular membrane vesicles, Hepatolog-y 14,
4. Miiller, M., Ishikawa, T., Berger, U., Klunemann, C., Lucka, L., Schrcyer, A., Kannicht, C., Reutter, W., Kurz, G. & Keppler, D. (199 1) ATP-dependent transport of taurocholate across the hepatocyte canalicular membrane mediated by a 110-kDa gly- coprotein binding ATP and bile salt, J. B i d . Chem. 266,
5. Nishida, T., Gatmaitan, Z., Che, M. & Arias, I. M. (1991) Rat liver canalicular membrane vesicles contain an ATP-dependent bile acid transport system, Proc. Natl Acad. Sci. U S A 88,
6. Kohayashi, K., Sogame, Y., Hayashi, K., Nicotera, P. & Orrenius, S. (1988) ATP stimulates the uptake of S- dinitrophenylglutathione by rat liver plasma membrane ves- icles, FEBS Left . 240, 55 - 58.
7. Akerboom, T. P. M., Narayanaswami, V., Kunst, M. & Sies. H. (1 99 1) ATP-dependent S-(2,4-dinitrophenyl)glutathionc trans- port in canalicular plasma membrane vesicles from rat liver, J. B i d . Chrm. 266, 13147-13152.
8. Wahlllnder. A. & Sies, H. (1979) Clulathione-S-conjugate forma- tion from l -chloro-2,4-dinitrobenzene and biliary S-conjugate
655 - 659.
18 920 - 18 926.
6590 - 6594.
excretion in the perfused rat liver, Eur. J . Biochem. 96, 441 - 446.
9. Akerboom, T. P. M. & Sies, H. (1989) Transport of glutathione, glutathione disulfide, and glutathione conjugates across the hepatocyte plasma membrane, Methods Enzymol. 173, 523 - 534.
10. Sies, H. (1988) Intracellular effects of glutathione conjugates and their transport from the cell, in Glutathione conjugation (Sies, H. & Ketterer, R., eds) pp. 175 - 292, Academic Press, London.
11. Hinchman, C. A., Matsumoto, H., Simmons, T. W. & Ballatori, N. (1991) Intrahepatic conversion of a glutathione conjugate and its mercapturic acid, .I. Biol. Chem. 266, 22179-22 185.
12. Habig, W., Pahst, M. J. & Jakoby. W. B. (1974) Glutathione S- Transferases, J . Biol. Chem. 249> 7130-7139.
13. Sies, H. (1985) Hydroperoxides and thiol oxidants in the study of oxidative stress in intact cells, in Oxidative stress (Sies, H., ed.) pp. 73 -90. Academic Press, Loadon.
14. Sies, H. & Summer, K.-H. (1975) Hydroperoxide-metabolizing systems in rat liver, Eur. J . Biochem. 57, 503 - 512.
15. Akerboom, T. P. M., Bilzer. M. & Sies, H. (1982) Competition between transport of glutathionc disulfidc (GSSG) and glutathione S-conjugates from perfused rat liver into bile, FEBS
16. Akerboorn, T. P. M., Bilzer, M. & Sies, H. (1982) The relationship of biliary glutathione disulfide efflux and intracellular glutathionc disulfide content in perfused rat liver. .I. Bid. Chem.
17. Brigelius, R. (1983) Glutathione oxidation and activation of pen- tose-phosphate cycle during hydroperoxide metabolism, Hoppe-Se-vler’s Z . Physiol. Chem. 364, 989 - 996.
18. Sies, H. (1978) The use ofperrusion ofliver and other organs for the study of microsomal electron transport and cytochrome P450 systems, Methods Enzvmul. 52, 48 - 59.
19. Akerboom, T. P. M. & Sies. H. (1981) Assay of glutathione, glutathione disulfide and glutathone mixed disulfides in bio- logical samples, Methods Enzymol. 77, 373 - 382.
20. Bergmeyer, H. U., ed. (1974) Methoden der enzymutischen Ana- lyse, 3rd edn, Verlag Chemie, Weinheim.
21. Stoll, B. & Hiussinger, D. (1989) Functional hepatocyte hetero-
Lpft . 140, 73 - 76.
257,4248 -4252.
22.
23.
24.
25.
26.
27.
28.
29.
30.
. .
geneity : vascular-2-oxoglutarate is almost exclusively taken up by perivenous glutamine-synthetase containing hepatocytes, Eur. 1. Biochem. 181,709-716.
Vom Dahl, S., Hallbrucker, C., Lang, F., Gerok, W. & Haussinger, D. (1991) A non-invasive technique for cell volume determination in perfused rat liver, B i d Chem. Hoppe-Seyler
Vom Dahl, S., Hallbrucker, C., Lang, F. & Hiussinger, D. (1991) Regulation of cell volume by hormones, Biochem. J . 280,105 - 109.
Haussinger, D., Stehle, ‘I. & Lang, F. (1990) Volume regulation in liver: further characterization by inhibitors and ionic substi- tutions, Hepatology I I , 243 - 254.
Lang, F., Stehle, T. & Haussinger, D. (1989) Water, K + , H’, lactate and glucose fluxes during cell volume regulation in perfused rat liver, Pflugers Arch. Physiol. 413.209 - 216.
Wettstein, M., vom Dahl, S., Lang, F., Gerok, W. & Haussinger, D. (1990) Cell volume regulatory responses of isolated perfused rat liver. Bid. Chem. Hoppe-Seyler 371, 493 - 501.
Rrigelius, R. (1983) Mixed disullides: biological functions and increase in oxidative stress, in Oxidative stress (Sies, H., ed.) pp. 243 -272, Academic Press, London.
Sies, H., Gerstenecker, C., Summer, K. H., Menzel, H. & Flohe, L. (1974) Glutathione-dependent hydroperoxide metabolism and associated metabolic transitions in hemoglobin-free per- fused rat liver, in Glutathione (Floht., L., Beniihr, C.. Sies, H. & Waller, H. D., eds) pp. 261 -276, G. Thieme Publ., Stuttgart.
Graf, J., Haddad, P., Hiussinger, D. & Lang F. (1988) Cell volume regulation in livcr, Renal Physiol. Biochem. 3 -5,202 - 220.
Vom Dahl. S., Hallbrucker, C., Lang, F. & Haussinger, D. (1991) Role of eicosanoids, inositol phosphates and extracellular Ca’ +
i n cell volume regulation in rat liver, Eur. J . Biochcm. 198. 73- 83.
372.411-418.

444
31. Baquet, A., Lavoinne, A. & Hue, L. (1991) Comparison of the effects ofvarious amino acids on glycogen synthesis, lipogenesis and ketogenesis in isolated rat hepatocytes, Biochem. J . 273,
32. Baquet, A., Maisin, L. & Hue, L. (1991) Swelling of rat hepatocytes activates acetyl-CoA carboxylase in parallel to gly- cogen synthase, Biochem. J . 278, 887 - 890.
33. Baquet, A., Hue, L., Meijer, A. J., van Woerkom, G. M. & Plomp, P. J . A. M. (1990) Swelling of hepatocytes stimulates glycogen synthesis, .J. Bid. Chem. 265, 955-959.
34. Halestrap, A. P. (1989) The regulation of the matrix volume of mammalian mitochondria in vivo and in virro and its role in the control of mitochondrial metabolism, Biochim. Bioph-vs. Acta
35. Hiiussinger, D., Lang, F., Bauers, K. & Gerok, W. (1990) Control or hepatic nitrogen metabolism and glutathione release by cell volume-regulatory mechanisms, Eur. J . Biochem. 193. 891 - 898.
36. Haussinger, D., Hallbrucker, C., Saha, N., Lang, F. & Gerok, W. (1992) Ccll volume and bile acid excretion, Biochem. J . , in the prcss.
37. Smith, M. T., Evans, C., Thor, H. & Orrenius, S. (1985) Quinonc- induced oxidative injury to cells and tissucs, in Oxidative stress (Sies, H., ed.) pp. 91 -113, Acadcmic Press, London.
57 - 62.
973, 355-382.
38. Gores, G. J., Flarsheim, C. H., Dawson, T. L., Nieminen, A. L., Herman, B. & Lemasters, J. J. (1989) Swelling, reductive stress and ccll dcath during chcmical hypoxia in hepatocytcs, Am. J. Physiol. 257, C341- (2354.
39. Willerson, J. T., Scales, F., Mukherjee, A., Platt, M., Templeton, G. H., Fink, G. S. & Buja, L. M. (1977) Abnormal myocardial fluid rctcntion as an carly manifestation of ischemic injury, Am. J . Pathol. 87, 159-181.
40. Laiho, K. U. & Trump, B. F. (1974) Relationship of ionic. water and ccll volumc changcs in ccllular injury of Ehrlich ascites tumor cells, Lab. Invest. 31, 207-215.
41. Flores, J . , DiBona, D. R., Beck, C. H. & Leaf, A. (1972) The role of cell swelling in ischemic renal damage and the protective effect of hypertonic solute, J . Clin. Invest. 51, 118-126.
42. Lemastcrs, J. J., DiGuiscppi, J. , Nieminen, A. L. & Herman, B. (1 987) Blebbing, free Ca2+ and mitochondria1 membrane potcntial prcceding cell death in hepatocytes, Nature 325, 78 - 81.
43. Lucas, S. K., Gordner, ?'. J . , Flaherty, J. T., Bulkley, B. H., Elmcr, E. B. & Gott, V. L. (1980) Beneficial effects of mannilol administration during reperfusion after ischemic arrest, Circu- lation 62, 134 - 141.




















