Early hominid bone tools from Drimolen, South Africa
15
Early hominid bone tools from Drimolen, South Africa Lucinda Backwell a, * , Francesco d’Errico b, c a Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africa b Institut de Pre ´histoire et de Ge´ologie du Quaternaire, CNRS UMR 5199 PACEA, Universite´ Bordeaux I, Avenue des Faculte ´s, F-33405 Talence, France c Institute for Human Evolution, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africa article info Article history: Received 30 January 2008 Received in revised form 24 May 2008 Accepted 28 May 2008 Keywords: Bone tools Early hominid behaviour Drimolen South Africa abstract The earliest use of bone tools is a topic of ongoing debate that concerns the criteria used to identify utilised or minimally modified bone tools, and if verified, the implications for hominid adaptation and cognition. Here we present the first description of 22 possible bone tools from the early hominid site of Drimolen (Gauteng Province, South Africa), dated w1.5–2 Mya. We compare the results of a taphonomic, morphometric and microscopic analysis of these pieces with those obtained from the study of faunal assemblages modified by a variety of non-human agents, and experimentally modified bones. None of the naturally modified assemblages contained pieces bearing the wear pattern observed on specimens from Drimolen and on bones experimentally used in digging activities. Fourteen pieces from Drimolen bear a pattern comparable to one previously described on early hominid bone tools from Sterkfontein and Swartkrans. This suggests that Drimolen bone tools were involved in a similar, if not exactly the same, task. Other common features include favoured bone types, fracture patterns, and the length and position of the worn area. Larger bone tools known from Swartkrans are absent at Drimolen, perhaps due to less availability of large mammal bones. The association of a high number of Paranthropus remains with bone tools at Drimolen, and the exceedingly low number of stone tools at the site supports the hypothesis that Paranthropus robustus used these bone tools. Based on implement-assisted termite for- aging strategies amongst chimpanzees, we have inferred similar social and cultural behaviours for early hominids. Gorillas were recently proposed as a model for P. robustus social structure due to the high degree of sexual dimorphism observed. According to female aggregation practices present in both models, one can speculate that if P. robustus was the user of the bone tools, the foraging activity in which they were used may have been conducted mainly by females. Ó 2008 Elsevier Ltd. All rights reserved. 1. Introduction The oldest use of bone implements, whether intentionally shaped or not, is a controversial issue. Claims that bone tools were in use since the dawn of humanity were first made by Schmidtgen (1929), Breuil (1932, 1938) and Bastin (1932), and culminated in Dart’s (1957) Osteodontokeratic culture of Australopithecus. Since then the earliest use of bone tools has been the subject of much debate, concerning the criteria used to identify intentionally used or minimally modified bone tools, and if verified, the implications of such innovation for hominid adaptation and cognition. Tapho- nomic studies have revealed that natural processes can produce pseudo bone artefacts. Vascular grooves have been shown to mimic cut marks (Shipman and Rose, 1984) and engravings (d’Errico and Villa, 1997), and some types of wear on teeth have been misinterpreted as carved notches (Gautier, 1986). Other mimics include natural breakage and wear of deer antler (Olsen, 1989) and elephant tusk tips (Haynes, 1991; Villa and d’Errico, 2001), gnawing or digestion by carnivores, rodents or herbivores (Pei, 1938; Sutcliffe, 1973, 1977; Binford, 1981; Villa and Bartram, 1996; d’Errico and Villa, 1997), fracture for marrow extraction by homi- nids and carnivores (Bunn, 1981, 1983; Gifford-Gonzalez, 1989; Backwell and d’Errico, 2004), trampling (Haynes, 1988; Olsen and Shipman, 1988), root etching (Binford, 1981), weathering (Brain, 1967), and the abrasive action of sediment (Brain, 1981; Lyman, 1994). In light of these findings it has become widely accepted that in order to distinguish between pseudo and true tools, it is neces- sary to adopt an interdisciplinary approach that combines analysis of bones modified by known agents, taphonomic analysis of the fossil assemblages from which the purported bone tools derive, microscopic studies of possible traces of manufacture and use, experimental replication of purported tools, and quantification of wear patterns. Employing a comparative, contextual approach has enabled authors such as Klein (1975), Shipman and Phillips (1976), Brain (1981) and Maguire et al. (1980) to contest Dart’s * Corresponding author. Tel.: þ27 11 717 6672; fax: þ27 11 717 6694. E-mail addresses: [email protected] (L. Backwell), [email protected] bordeaux1.fr (F. d’Errico). Contents lists available at ScienceDirect Journal of Archaeological Science journal homepage: http://www.elsevier.com/locate/jas 0305-4403/$ – see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.jas.2008.05.017 Journal of Archaeological Science 35 (2008) 2880–2894
Transcript of Early hominid bone tools from Drimolen, South Africa

lable at ScienceDirect
Journal of Archaeological Science 35 (2008) 2880–2894
Contents lists avai
Journal of Archaeological Science
journal homepage: ht tp: / /www.elsevier .com/locate/ jas
Early hominid bone tools from Drimolen, South Africa
Lucinda Backwell a,*, Francesco d’Errico b,c
a Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africab Institut de Prehistoire et de Geologie du Quaternaire, CNRS UMR 5199 PACEA, Universite Bordeaux I, Avenue des Facultes, F-33405 Talence, Francec Institute for Human Evolution, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africa
a r t i c l e i n f o
Article history:Received 30 January 2008Received in revised form 24 May 2008Accepted 28 May 2008
Keywords:Bone toolsEarly hominid behaviourDrimolenSouth Africa
* Corresponding author. Tel.: þ27 11 717 6672; fax:E-mail addresses: [email protected] (L.
bordeaux1.fr (F. d’Errico).
0305-4403/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.jas.2008.05.017
a b s t r a c t
The earliest use of bone tools is a topic of ongoing debate that concerns the criteria used to identifyutilised or minimally modified bone tools, and if verified, the implications for hominid adaptation andcognition. Here we present the first description of 22 possible bone tools from the early hominid site ofDrimolen (Gauteng Province, South Africa), dated w1.5–2 Mya. We compare the results of a taphonomic,morphometric and microscopic analysis of these pieces with those obtained from the study of faunalassemblages modified by a variety of non-human agents, and experimentally modified bones. None ofthe naturally modified assemblages contained pieces bearing the wear pattern observed on specimensfrom Drimolen and on bones experimentally used in digging activities. Fourteen pieces from Drimolenbear a pattern comparable to one previously described on early hominid bone tools from Sterkfonteinand Swartkrans. This suggests that Drimolen bone tools were involved in a similar, if not exactly thesame, task. Other common features include favoured bone types, fracture patterns, and the length andposition of the worn area. Larger bone tools known from Swartkrans are absent at Drimolen, perhaps dueto less availability of large mammal bones. The association of a high number of Paranthropus remainswith bone tools at Drimolen, and the exceedingly low number of stone tools at the site supports thehypothesis that Paranthropus robustus used these bone tools. Based on implement-assisted termite for-aging strategies amongst chimpanzees, we have inferred similar social and cultural behaviours for earlyhominids. Gorillas were recently proposed as a model for P. robustus social structure due to the highdegree of sexual dimorphism observed. According to female aggregation practices present in bothmodels, one can speculate that if P. robustus was the user of the bone tools, the foraging activity in whichthey were used may have been conducted mainly by females.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
The oldest use of bone implements, whether intentionallyshaped or not, is a controversial issue. Claims that bone tools werein use since the dawn of humanity were first made by Schmidtgen(1929), Breuil (1932, 1938) and Bastin (1932), and culminated inDart’s (1957) Osteodontokeratic culture of Australopithecus. Sincethen the earliest use of bone tools has been the subject of muchdebate, concerning the criteria used to identify intentionally usedor minimally modified bone tools, and if verified, the implicationsof such innovation for hominid adaptation and cognition. Tapho-nomic studies have revealed that natural processes can producepseudo bone artefacts. Vascular grooves have been shown to mimiccut marks (Shipman and Rose, 1984) and engravings (d’Errico andVilla, 1997), and some types of wear on teeth have been
þ27 11 717 6694.Backwell), [email protected]
All rights reserved.
misinterpreted as carved notches (Gautier, 1986). Other mimicsinclude natural breakage and wear of deer antler (Olsen, 1989) andelephant tusk tips (Haynes, 1991; Villa and d’Errico, 2001), gnawingor digestion by carnivores, rodents or herbivores (Pei, 1938;Sutcliffe, 1973, 1977; Binford, 1981; Villa and Bartram, 1996;d’Errico and Villa, 1997), fracture for marrow extraction by homi-nids and carnivores (Bunn, 1981, 1983; Gifford-Gonzalez, 1989;Backwell and d’Errico, 2004), trampling (Haynes, 1988; Olsen andShipman, 1988), root etching (Binford, 1981), weathering (Brain,1967), and the abrasive action of sediment (Brain, 1981; Lyman,1994). In light of these findings it has become widely accepted thatin order to distinguish between pseudo and true tools, it is neces-sary to adopt an interdisciplinary approach that combines analysisof bones modified by known agents, taphonomic analysis of thefossil assemblages from which the purported bone tools derive,microscopic studies of possible traces of manufacture and use,experimental replication of purported tools, and quantification ofwear patterns. Employing a comparative, contextual approach hasenabled authors such as Klein (1975), Shipman and Phillips (1976),Brain (1981) and Maguire et al. (1980) to contest Dart’s

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2881
‘Osteodontokeratic’ hypothesis. This has created scepticism as towhether early hominids truly used bone tools, but at the same timehas provided a frame of reference for assessing new potentialevidence.
Robinson (1959) proposed that a metapodial shaft fragmentwith a smoothed pointed end from Sterkfontein Member 5 West(w1.7–1.4 Ma) was a tool. Nearly 30 years later Brain described 68similarly modified bones and horn cores from Members 1–3 atSwartkrans, dated w1.8–1 Ma (Brain et al., 1988; Brain, 1989).Comparative scanning electron microscope inspection of replicas ofthe worn area on archaeological specimens and experimental shaftfragments used to extract tubers from the ground and work skins,suggested to Brain and Shipman (1993, 2004) that the wear pat-terns on the smoothed tips of the Swartkrans specimens matchedthose produced experimentally. Reanalysis of these pieces con-firmed the anthropogenic origin of the use-wear (Backwell, 2000;Backwell and d’Errico, 2001; d’Errico et al., 2001). The analysis wasbased on (1) comparison with a large reference collection of faunamodified by 10 non-human agents, (2) morphometric and fractureanalysis of bone flakes from Members 1–3 at Swartkrans, (3)experimental use of shaft fragments in a variety of tasks, (4)microscopic analysis of natural, experimental and archaeologicalwear patterns, and (5) quantification of striation width and orien-tation on archaeological and experimental specimens. The break-age patterns and size of the bone tools, compared with theremainder of the faunal remains, indicated that early hominidsselected weathered, elongated and robust bone fragments for useas tools. The wear pattern on the Swartkrans tools did not matchthose produced experimentally to extract tubers or work skins. Itfitted more closely with one created experimentally when exca-vating termite mounds in the area (Fig. 1), which suggested this was
Fig. 1. Transparent resin replicas of wear patterns on Swartkrans, Drimolen and experimeMember 2 (SKX 1142). (b) Newly identified bone tool (DN 414) from the early hominid site osmoothed by use-wear. (c) Experimental bone tool used to dig in a termite mound. (d) Bonethe orientation and width of the striations in (a, b and c). Scales¼ 1 mm.
the most common function for the Swartkrans bone tools. Evidenceof possible intentional shaping through grinding is also identifiedby d’Errico and Backwell (2003) on the tips of six horn cores and anulna in the Swartkrans collection. These pieces bear flat facets closeto the tip that are covered by parallel spindle-shaped striationsoblique to the tool main axis, comparable to bone experimentallyground on the surfaces of termite mounds.
In another paper Backwell and d’Errico (2003) described 16 newbone tools from Swartkrans and used the enlarged collection tosearch for patterns of variation between Members. No significantdifferences were observed between Members 1–3 in the type andsize of the bone fragments used as tools, as well as in the length andtype of the wear pattern, demonstrating that no major changesoccurred through time in the bone type favoured, and the motion inwhich the tools were used. In addition, these authors showed thatthe bone tools do not share the same spatial distribution as thestone tools, suggesting the two types of artefacts did not enter thecave at the same time and may have been used in different activ-ities, at different times, and by different members of a hominidgroup, or different hominid taxa. Members 1–3 at Swartkranscontain the remains of the robust australopithecine, Paranthropusrobustus. Members 1 and 2 have yielded the remains of Homoerectus (Brain et al., 1988). The absence of this taxon in Member 3,from where most of the bone tools derive, suggests, but does notprove, that these implements were used by P. robustus (Backwelland d’Errico, 2003).
The discovery of bone tools similar to those found at Swartkransand Sterkfontein was announced for the nearby early hominid siteof Drimolen by Keyser in 2000 (Keyser, 2000a; d’Errico et al., 2001).The aim of this paper is to provide a description of the Drimolenspecimens and assess their artefactual nature. We also intend to
ntal bone tool tips photographed in transmitted light. (a) Bone tool from Swartkransf Drimolen showing a rounded tip covered with longitudinal striations and flake scarstool used in Brain’s experiment to dig up Scilla marginata bulbs. Note the similarity in
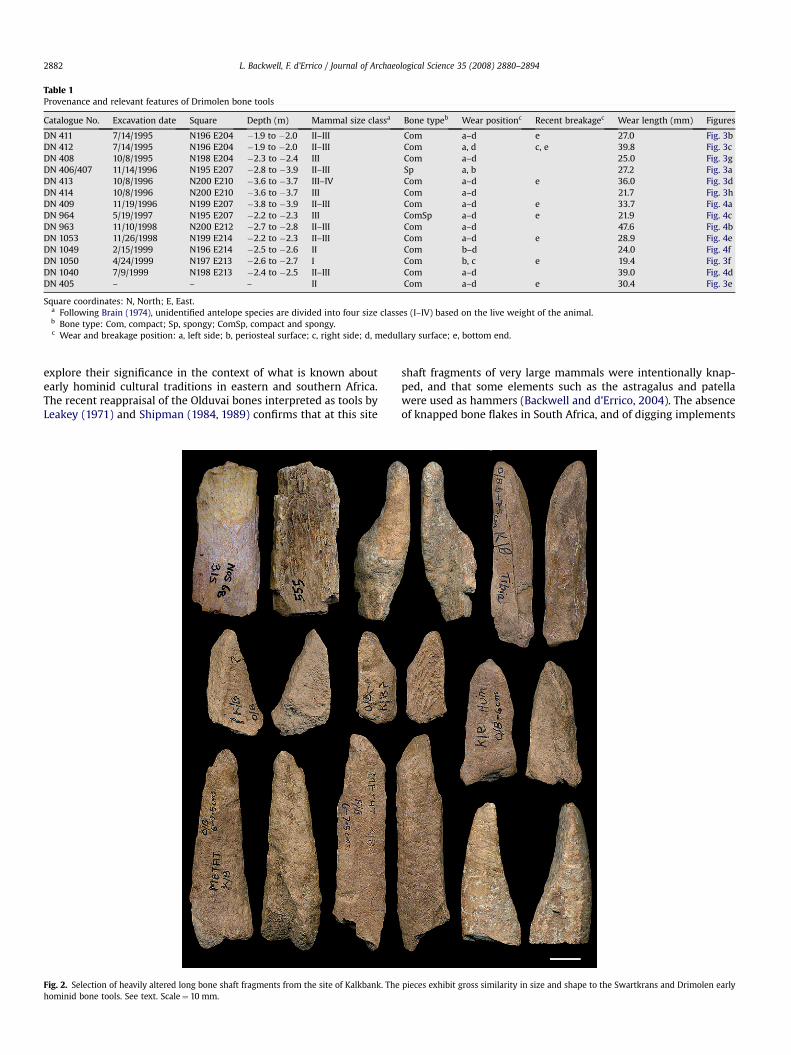
Table 1Provenance and relevant features of Drimolen bone tools
Catalogue No. Excavation date Square Depth (m) Mammal size classa Bone typeb Wear positionc Recent breakagec Wear length (mm) Figures
DN 411 7/14/1995 N196 E204 �1.9 to �2.0 II–III Com a–d e 27.0 Fig. 3bDN 412 7/14/1995 N196 E204 �1.9 to �2.0 II–III Com a, d c, e 39.8 Fig. 3cDN 408 10/8/1995 N198 E204 �2.3 to �2.4 III Com a–d 25.0 Fig. 3gDN 406/407 11/14/1996 N195 E207 �2.8 to �3.9 II–III Sp a, b 27.2 Fig. 3aDN 413 10/8/1996 N200 E210 �3.6 to �3.7 III–IV Com a–d e 36.0 Fig. 3dDN 414 10/8/1996 N200 E210 �3.6 to �3.7 III Com a–d 21.7 Fig. 3hDN 409 11/19/1996 N199 E207 �3.8 to �3.9 II–III Com a–d e 33.7 Fig. 4aDN 964 5/19/1997 N195 E207 �2.2 to �2.3 III ComSp a–d e 21.9 Fig. 4cDN 963 11/10/1998 N200 E212 �2.7 to �2.8 II–III Com a–d 47.6 Fig. 4bDN 1053 11/26/1998 N199 E214 �2.2 to �2.3 II–III Com a–d e 28.9 Fig. 4eDN 1049 2/15/1999 N196 E214 �2.5 to �2.6 II Com b–d 24.0 Fig. 4fDN 1050 4/24/1999 N197 E213 �2.6 to �2.7 I Com b, c e 19.4 Fig. 3fDN 1040 7/9/1999 N198 E213 �2.4 to �2.5 II–III Com a–d 39.0 Fig. 4dDN 405 – – – II Com a–d e 30.4 Fig. 3e
Square coordinates: N, North; E, East.a Following Brain (1974), unidentified antelope species are divided into four size classes (I–IV) based on the live weight of the animal.b Bone type: Com, compact; Sp, spongy; ComSp, compact and spongy.c Wear and breakage position: a, left side; b, periosteal surface; c, right side; d, medullary surface; e, bottom end.
L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942882
explore their significance in the context of what is known aboutearly hominid cultural traditions in eastern and southern Africa.The recent reappraisal of the Olduvai bones interpreted as tools byLeakey (1971) and Shipman (1984, 1989) confirms that at this site
Fig. 2. Selection of heavily altered long bone shaft fragments from the site of Kalkbank. Thehominid bone tools. See text. Scale¼ 10 mm.
shaft fragments of very large mammals were intentionally knap-ped, and that some elements such as the astragalus and patellawere used as hammers (Backwell and d’Errico, 2004). The absenceof knapped bone flakes in South Africa, and of digging implements
pieces exhibit gross similarity in size and shape to the Swartkrans and Drimolen early

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2883
in East Africa, lends support to the hypothesis that two distinctbone tool traditions existed in Africa between 1 and 2 Ma (Backwelland d’Errico, 2004), either as variations of a single speciesbehaviour, or due to manufacture by different hominid taxa.Considering that only a single piece is reported from Sterkfontein,this hypothesis is based on the findings from only two sites,Swartkrans and Olduvai. The existence of an additional South
Fig. 3. Bone tools from Drimolen. When applicable, the periosteal aspect is on the lef
African Plio-Pleistocene site yielding the same type of bone tools asthose from Swartkrans would demonstrate that the Swartkransbone tools are not an isolated case, and that the digging activity inwhich they were used was relatively widespread and well estab-lished in the region. Comparison of bone tools between sites canreveal variation in bone type choice, applied motion and toolfunction. Drimolen has yielded an abundance of Paranthropus
t and the medullary aspect is on the right. Lines indicate breaks. Scale¼ 10 mm.

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942884
remains and only two possible stone tools, an association that mayhelp elucidate the early hominid user of the bone tools.
1.1. Archaeological context
Drimolen is a former cave within the dolomites of the MonteChristo Formation of the Malmani Subgroup of the ChunniespoortGroup (Palmer, 1991). The site, located 7 km North of Sterkfontein,was discovered by Keyser in 1992 (Keyser, 2000b; Keyser et al.,2000). Most of the ceiling of the cave has been removed byweathering and erosion. The large mammal fossil-bearing depositsconsist of in situ cave sediment and collapsed blocks of dolomite,chert, large boulders of Blocky Breccia and cave siltstone, togethertermed Collapsed sinkhole Fill. Miners in search of limestonereportedly dumped some of the decalcified waste in the centre of
Fig. 4. Bone tools from Drimolen. The delineated area in (d) marks a weathered
the site. Although large numbers of fossils, including hominids,have been recovered from the dump, the majority of hominidremains and bone tools have been found in the decalcified Col-lapsed sinkhole Fill. However, one specimen (DN 406, Fig. 3a) hascalcified breccia adhering to it, suggesting that all of the bone toolsderived from in situ calcified cave sediment (Keyser, personalcommunication).
The age of the cave fill is estimated to be 1.5 to 2 Ma, basedmostly on faunal correlation with Member 1 at Swartkrans (Brain,1993). Excavation of this site has yielded 79 hominid specimens ofwhich only two are firmly attributed to Homo sp. Interestingly,nearly half of the P. robustus specimens, which constitute theremainder of the hominid collection, are assigned to juveniles(Keyser et al., 2000). Other large mammal taxa identified (Keyseret al., 2000; Table 1) include cercopithecines, carnivores and
surface altered by porcupine gnawing. Lines indicate breaks. Scale¼ 10 mm.

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2885
bovids. Noteworthy are the absence of suids and equids, and theabundance of small- to medium-sized bovids. Twenty-three bonefragments were earmarked in the field as possible bone tools. Ofthese, 22 were made available for analysis and form the subject ofthis paper.
Fig. 5. Pseudo bone tools from Drimolen. (a) DN 1038 showing a smoothly rounded end witshowing smooth, rounded tips; (e, f) DN 1039 and (g, h) DN 1042 showing smooth, rounded e(b–d, f, and h).
1.2. Comparative collections
Human and non-human modification of bone is the topic ofa large and constantly growing literature (see for example Brain,1981; Olsen and Shipman, 1988; White, 1992; Fisher, 1995; Hannus
hout any evidence of striations. (b) Close up of the same tip. (c) DN 765 and (d) DN 72nds without multiple fine parallel striations. Scale¼ 5 mm in (a, e, and g) and 10 mm in

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942886
et al., 1997; Plummer and Stanford, 2000; Selvaggio and Wilder,2001; Njau and Blumenschine, 2006) which has defined the criteriato be used in the identification of agents responsible for bonealteration. Experimental replication has also refined the criteriaused to identify shaping techniques and bone tool function(LeMoine, 1997; Scheinsohn, 1997; Choyke and Bartosiewicz, 2001).We have previously examined 35 reference collections of bone fromboth modern and fossil contexts, modified by known non-humanagents (hyaena, dog, leopard, cheetah, porcupine, river gravel,spring water, plain flooding, trampling) in search for mimics ofmodifications recorded on the Swartkrans specimens interpreted
Fig. 6. (a) Newly identified Drimolen bone tool (DN 413) showing a smooth, rounded tip coshowing multiple, fine, individual striations oriented longitudinally. Scales¼ 1 mm.
as bone tools (Backwell and d’Errico, 2001, 2004). All bone fromthese collections was analysed at macroscopic and microscopicscale using optical and scanning electron microscopy. Our researchon the recognition of shaping techniques includes the reproductionand microscopic analysis of marks produced with different tech-niques and motions (d’Errico et al., 1984; d’Errico and Backwell,2003; Backwell and d’Errico, 2004). We have also created andmicroscopically analysed a reference collection comprising bonesused in a variety of tasks (Backwell and d’Errico, 2004), experi-mentally manipulated and transported (d’Errico, 1993a,b). In theframework of this study we have increased our comparative
vered with use-wear. Scale¼ 5 mm. (b–d) Close up of wear pattern on the same object

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2887
collection to include the bone assemblage from the site of Kalk-bank, excavated by Mason in 1954 (Mason, 1958). Located 64 kmnorth-west of Polokwane (formerly Pietersburg), Limpopo Prov-ince, South Africa, Kalkbank is an open air site comprised ofa number of layers of calcrete, indicative of former pan surfaces.Several layers of hardpan calcrete interspersed with layers of siltysand and sandy clay overlie a 0.3 m fossil horizon, which sits atop
Fig. 7. Selection of newly identified bone tools from Drimolen showing wear pattern consisti(c, d) DN 409; (e, f) DN 412. Scale¼ 1 mm in (a, b, and d–f) and 5 mm in (c).
clayey sand containing artefacts (Netterberg, 1974). It has yieldeda bone assemblage composed of 3619 fragments, described by Dartand Kitching (1958) as containing Osteodontokeratic tools, andlithics attributed by Mason (1958) to the Middle Stone Age. Theinterest of this assemblage lies in the large number of elongatedshaft fragments with pointed or rounded tips, and an overall grossmorphology that is reminiscent of the Swartkrans bone tools
ng of parallel or sub-parallel individual striations oriented longitudinally. (a, b) DN 963;

Fig. 8. Newly identified bone tool specimen DN 414 from Drimolen showing (a) wear pattern on the periosteal aspect of the specimen, and (b) resin replica of the same areaobserved close up in transmitted light. (c, d) Medullary aspect of the same specimen showing fine individual sub-parallel striations oriented longitudinally. Scales¼ 1 mm.
Table 2Orientation of striations on each aspect of the Drimolen bone tools
CatalogueNo.
Periosteala Right Medullary Left
Long. Obl. Trans. Long. Obl. Trans. Long. Obl. Trans. Long. Obl. Trans.
DN 411 1 1 1 1 1 1 1 1DN 412 1 1 1DN 408 1 1 1 1 1 1 1 1DN 406 1 1 1 1DN 413 1 1 1 1 1 1DN 414 1 1 1 1 1 1 1 1DN 409 1 1 1 1 1 1 1 1 1DN 964 1 1 1 1 1 1 1DN 963 1 1 1 1 1 1 1DN 1053 1 1 1 1 1 1 1DN 1049 1 1 1 1 1DN 1050 1 1 1 1DN 1040 1 1 1 1 1 1 1 1 1 1 1 1DN 405 1 1 1 1
Right and Left represent the sides of a bone flake when viewing the periostealsurface. Our sequence of analysis begins on the periosteal aspect, and is followed bythe right side, medullar and left, as the flake is rotated counter-clockwise.Striation orientation: Long, longitudinal; Obl, oblique; Trans, transverse.
a In the few instances for which the anatomical orientation is unknown, thebroadest convex aspect is taken as the periosteal surface.
L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942888
(Fig. 2). These pieces were interpreted by Dart and Kitching asevidence that long bones were longitudinally broken to be used astools. However, these authors mention that 38.7% of the postcranialbone fragments show porcupine gnawing. This, together with thepredominance of weathered fractures (Brain, 1981) points toporcupines as the main modifying agents and accumulators of theKalkbank faunal assemblage. In our analysis of this materialwe have focused on shaft fragments morphologically anddimensionally similar to those identified as bone tools fromSwartkrans and Drimolen, and analysed them with the same pro-cedures described below for the Drimolen site’s purported bonetools.
2. Method
Drimolen specimens selected as potential tools were analysedwith an optical microscope in search of the localised wear patternrecorded on the tips of Swartkrans, Sterkfontein and experimentalbone tools. On the specimens retained as tools we recorded a rangeof variables related to mammal size class, type of bone used,
L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2889
fracture patterns, shape and size of the bone flake, includingcompact bone thickness and the length and position of the wornarea. The orientation of the striations (longitudinal, oblique,transverse) on the worn end was recorded for each aspect of thepiece (periosteal, right, medulla, left). The same variables wererecorded on the 84 specimens from Swartkrans, and the singlepiece from Sterkfontein.
The tips of 11 specimens from Drimolen and 14 from Kalkbank,selected for their overall similarity to Swartkrans bone tools, were
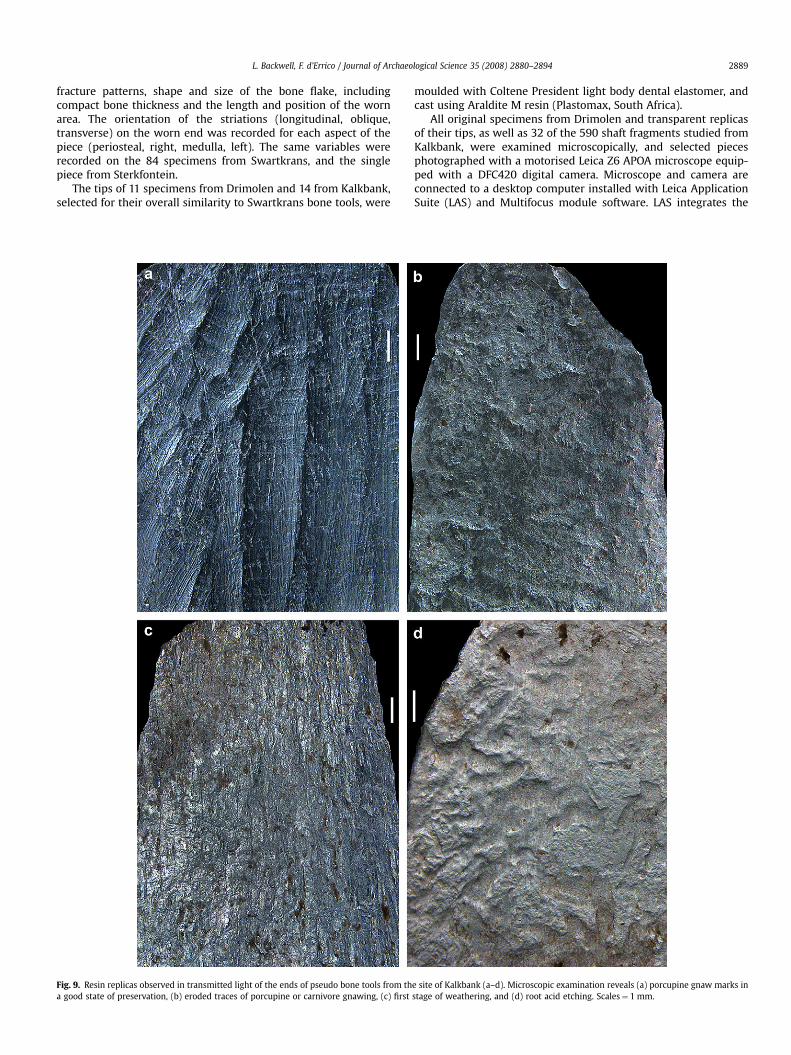
Fig. 9. Resin replicas observed in transmitted light of the ends of pseudo bone tools from tha good state of preservation, (b) eroded traces of porcupine or carnivore gnawing, (c) first
moulded with Coltene President light body dental elastomer, andcast using Araldite M resin (Plastomax, South Africa).
All original specimens from Drimolen and transparent replicasof their tips, as well as 32 of the 590 shaft fragments studied fromKalkbank, were examined microscopically, and selected piecesphotographed with a motorised Leica Z6 APOA microscope equip-ped with a DFC420 digital camera. Microscope and camera areconnected to a desktop computer installed with Leica ApplicationSuite (LAS) and Multifocus module software. LAS integrates the
e site of Kalkbank (a–d). Microscopic examination reveals (a) porcupine gnaw marks instage of weathering, and (d) root acid etching. Scales¼ 1 mm.

Fig. 10. (a) Tip of original Drimolen bone tool (DN 1040). (b) Transparent resin replica of the same area viewed under transmitted light, revealing an unusually high proportion oftransverse and oblique striations. Scales¼ 1 mm.
L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942890
Leica automated microscope, digital camera and software intoa common environment. The Multifocus module is designed toacquire extended depth of field images from the microscope.Exposure, gain, shading and all other camera parameters areindividually set to optimise the quality of image acquisition. Oncedigital images have been collected at different Z-positions, adaptedalgorithms combine them into a single, sharp, composite imagewith a significantly extended depth of field.
Fig. 11. Frequency of mammal size classes present in the (a) Drimolen and (b)Swartkrans bone tool collections. Both sites show a high representation of medium- tolarge-sized mammals. Size class III–IV mammals are comparatively under-representedat Drimolen.
3. Results
3.1. Assessing the artefactual nature of the Drimolen collection
Microscopic inspection of the Drimolen specimens reveals thatonly 14 (Figs. 3 and 4, Table 1) of the 22 pieces analysed bear thefeatures characteristic of the Swartkrans bone tools (Fig. 1a),namely a single smoothed end covered with individual striationsconfined to an average of ca. 30 mm from the tip. On the Swartkransspecimens most of these striations are oriented parallel or sub-parallel to the main axis of the object, and decrease in number awayfrom the tip. They generally vary in width from 5 to 40 mm, and can
Fig. 12. Frequency distribution of compact bone thickness recorded in tools from (a)Drimolen and (b) Swartkrans. The Swartkrans distribution includes pieces from verylarge mammals, absent at Drimolen.
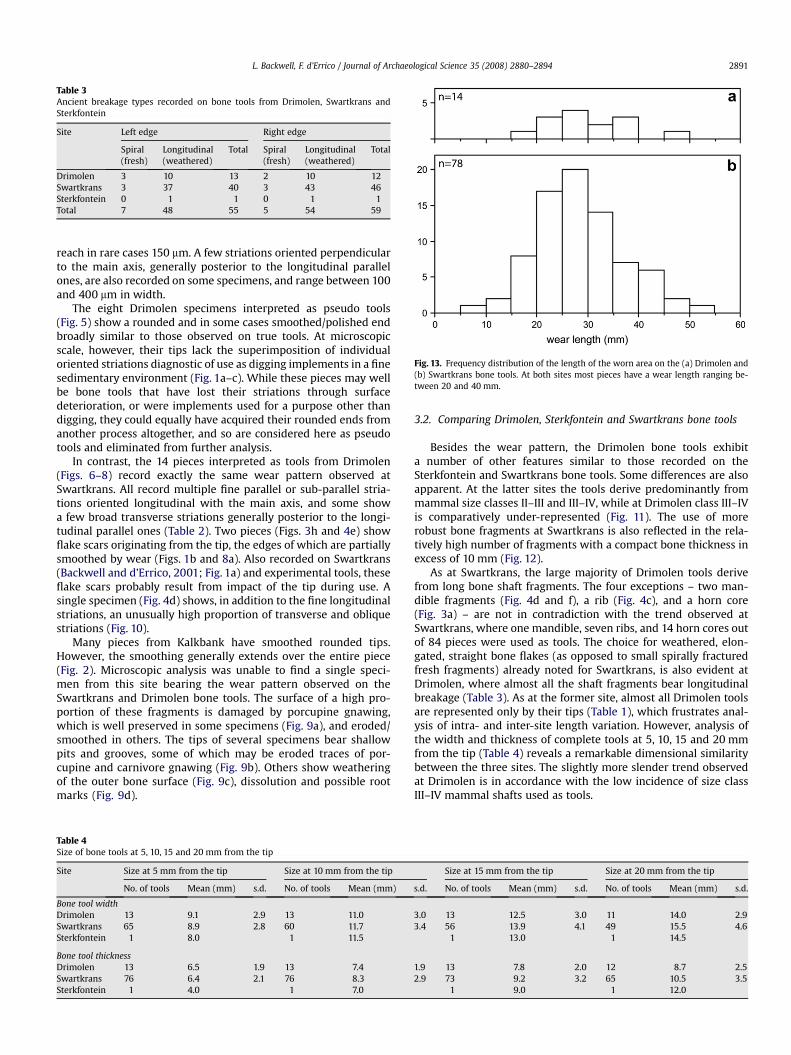
Table 3Ancient breakage types recorded on bone tools from Drimolen, Swartkrans andSterkfontein
Site Left edge Right edge
Spiral(fresh)
Longitudinal(weathered)
Total Spiral(fresh)
Longitudinal(weathered)
Total
Drimolen 3 10 13 2 10 12Swartkrans 3 37 40 3 43 46Sterkfontein 0 1 1 0 1 1Total 7 48 55 5 54 59
Fig. 13. Frequency distribution of the length of the worn area on the (a) Drimolen and(b) Swartkrans bone tools. At both sites most pieces have a wear length ranging be-tween 20 and 40 mm.
L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2891
reach in rare cases 150 mm. A few striations oriented perpendicularto the main axis, generally posterior to the longitudinal parallelones, are also recorded on some specimens, and range between 100and 400 mm in width.
The eight Drimolen specimens interpreted as pseudo tools(Fig. 5) show a rounded and in some cases smoothed/polished endbroadly similar to those observed on true tools. At microscopicscale, however, their tips lack the superimposition of individualoriented striations diagnostic of use as digging implements in a finesedimentary environment (Fig. 1a–c). While these pieces may wellbe bone tools that have lost their striations through surfacedeterioration, or were implements used for a purpose other thandigging, they could equally have acquired their rounded ends fromanother process altogether, and so are considered here as pseudotools and eliminated from further analysis.
In contrast, the 14 pieces interpreted as tools from Drimolen(Figs. 6–8) record exactly the same wear pattern observed atSwartkrans. All record multiple fine parallel or sub-parallel stria-tions oriented longitudinal with the main axis, and some showa few broad transverse striations generally posterior to the longi-tudinal parallel ones (Table 2). Two pieces (Figs. 3h and 4e) showflake scars originating from the tip, the edges of which are partiallysmoothed by wear (Figs. 1b and 8a). Also recorded on Swartkrans(Backwell and d’Errico, 2001; Fig. 1a) and experimental tools, theseflake scars probably result from impact of the tip during use. Asingle specimen (Fig. 4d) shows, in addition to the fine longitudinalstriations, an unusually high proportion of transverse and obliquestriations (Fig. 10).
Many pieces from Kalkbank have smoothed rounded tips.However, the smoothing generally extends over the entire piece(Fig. 2). Microscopic analysis was unable to find a single speci-men from this site bearing the wear pattern observed on theSwartkrans and Drimolen bone tools. The surface of a high pro-portion of these fragments is damaged by porcupine gnawing,which is well preserved in some specimens (Fig. 9a), and eroded/smoothed in others. The tips of several specimens bear shallowpits and grooves, some of which may be eroded traces of por-cupine and carnivore gnawing (Fig. 9b). Others show weatheringof the outer bone surface (Fig. 9c), dissolution and possible rootmarks (Fig. 9d).
Table 4Size of bone tools at 5, 10, 15 and 20 mm from the tip
Site Size at 5 mm from the tip Size at 10 mm from the tip
No. of tools Mean (mm) s.d. No. of tools Mean (mm)
Bone tool widthDrimolen 13 9.1 2.9 13 11.0Swartkrans 65 8.9 2.8 60 11.7Sterkfontein 1 8.0 1 11.5
Bone tool thicknessDrimolen 13 6.5 1.9 13 7.4Swartkrans 76 6.4 2.1 76 8.3Sterkfontein 1 4.0 1 7.0
3.2. Comparing Drimolen, Sterkfontein and Swartkrans bone tools
Besides the wear pattern, the Drimolen bone tools exhibita number of other features similar to those recorded on theSterkfontein and Swartkrans bone tools. Some differences are alsoapparent. At the latter sites the tools derive predominantly frommammal size classes II–III and III–IV, while at Drimolen class III–IVis comparatively under-represented (Fig. 11). The use of morerobust bone fragments at Swartkrans is also reflected in the rela-tively high number of fragments with a compact bone thickness inexcess of 10 mm (Fig. 12).
As at Swartkrans, the large majority of Drimolen tools derivefrom long bone shaft fragments. The four exceptions – two man-dible fragments (Fig. 4d and f), a rib (Fig. 4c), and a horn core(Fig. 3a) – are not in contradiction with the trend observed atSwartkrans, where one mandible, seven ribs, and 14 horn cores outof 84 pieces were used as tools. The choice for weathered, elon-gated, straight bone flakes (as opposed to small spirally fracturedfresh fragments) already noted for Swartkrans, is also evident atDrimolen, where almost all the shaft fragments bear longitudinalbreakage (Table 3). As at the former site, almost all Drimolen toolsare represented only by their tips (Table 1), which frustrates anal-ysis of intra- and inter-site length variation. However, analysis ofthe width and thickness of complete tools at 5, 10, 15 and 20 mmfrom the tip (Table 4) reveals a remarkable dimensional similaritybetween the three sites. The slightly more slender trend observedat Drimolen is in accordance with the low incidence of size classIII–IV mammal shafts used as tools.
Size at 15 mm from the tip Size at 20 mm from the tip
s.d. No. of tools Mean (mm) s.d. No. of tools Mean (mm) s.d.
3.0 13 12.5 3.0 11 14.0 2.93.4 56 13.9 4.1 49 15.5 4.6
1 13.0 1 14.5
1.9 13 7.8 2.0 12 8.7 2.52.9 73 9.2 3.2 65 10.5 3.5
1 9.0 1 12.0

Table 5Number of bone tools recording longitudinal, oblique and transverse striations on the four aspects
Site No. of tools Periosteala Right Medullary Left
Long. Obl. Trans. Long. Obl. Trans. Long. Obl. Trans. Long. Obl. Trans.
Drimolen 14 12 (86) 11 (79) 3 (21) 12 (86) 5 (36) 4 (29) 12 (86) 5 (36) 3 (21) 12 (86) 7 (50) 6 (43)Swartkrans 81 70 (86) 38 (47) 16 (20) 56 (69) 23 (28) 6 (7) 64 (79) 22 (27) 8 (10) 55 (68) 20 (26) 4 (5)Sterkfontein 1 1 0 0 1 0 0 1 0 0 1 0 0
Right and left represent the sides of a bone flake when viewing the periosteal surface. Our sequence of analysis begins on the periosteal aspect, and is followed by the right side,medullar and left, as the flake is rotated counter-clockwise.Striation orientation: Long, longitudinal; Obl, oblique; Trans, transverse.Numbers in parentheses are the percentage (%) of bone tools that record the different striation orientations on the four aspects.
a In the few instances for which the anatomical orientation is unknown, the broadest convex aspect is taken as the periosteal surface.
L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942892
The length of the worn area on the Drimolen specimens fallswell within the range of that observed at Swartkrans (Fig. 13) withthe majority of the wear ranging between 20 and 40 mm. The lackof pieces with a very short or long worn area at Drimolen is mostlikely a function of the small sample size. At all three sites theoccurrence and location of differently oriented striations shows anabundance of longitudinal relative to oblique, and rarity of trans-verse striations (Table 5).
Microscopic inspection of the worn tips has highlighted anotherfeature that parallels what we have observed at Swartkrans. Theonly horn core from Drimolen records an area close to the tipcovered with parallel striations perpendicular to the main axis(Fig. 14). These closely match those described on six horn coresfrom Swartkrans (d’Errico and Backwell, 2003), interpreted aspossible evidence of grinding, for the purpose of tapering orre-sharpening the functional area of the tool.
Finally, among the pieces interpreted as bone tools, two havepunctures probably produced by small carnivores (Fig. 4a and f) andone piece (Fig. 4d) shows at the end opposite to the worn tip,damage consisting of shallow grooves reminiscent of porcupinegnawing. Carnivore damage is recorded on one of the bone toolsfrom Swartkrans (d’Errico and Backwell, 2003: p. 1565, Fig. 3),where it is quite clearly covered with use-wear.
4. Discussion
The wear pattern found on the tips of bone fragments from thesites of Swartkrans, Sterkfontein and Drimolen has so far not beenfound in any comparative collections that we have studied. Kalk-bank is amongst the collections in which the proportion of elon-gated bone fragments with rounded tips is high. Still, a closescrutiny reveals no match with the bones interpreted as early
Fig. 14. Newly identified bone tool (DN 406) from Drimolen, representing the only horn corecords possible evidence of grinding (right). Scales¼ 1 mm.
hominid tools. Considering the close similarity in location, extentand microscopic features between the wear on the Swartkrans,Sterkfontein and Drimolen specimens and that produced experi-mentally in digging activities, an anthropogenic origin for theobserved modifications is retained as the best-fit hypothesis. Inorder to strengthen this interpretation, detailed inspection of fau-nal remains and morphometric analysis of shaft fragments fromDrimolen will be conducted as soon as the material becomesavailable for study. Functional interpretation of the tools from thesethree sites may also be refined. Quantification of the wear patternhas shown termite foraging to be the closest match. However, thisinterpretation must be verified by enlarging the variety of tasks inwhich similar bone tools may be used experimentally, and byrefining the methods used to quantify the resulting surface modi-fication. This is the goal of our ongoing research project (Backwelland d’Errico, 2005). For the purposes of this paper we observea striking similarity between bone tools from southern Africanhominid sites, which suggests that they were involved in a similar,if not exactly the same, task. The only possible difference betweenthe bone tools from Drimolen and those from Swartkrans – rarity ofvery robust pieces at the former site – may be due either to thesmall size of the sample, or to different availability of weatheredshaft fragments, or to a preference for slightly smaller fragments atDrimolen. The second hypothesis is partially supported by therarity of very large mammals reported for this site (Keyser et al.,2000). The third hypothesis is almost contradicted by the mor-phometry of the tool tips, showing, in spite of the use of thinnerblanks at Drimolen, a strong similarity between sites. Thesehypotheses may be tested in future, when results of taphonomicanalysis of the faunal assemblage from this site become available.More detailed contextual information may also elucidate why, inthree instances, bone tools have been found in very close proximity,
re in the collection. It shows a smooth rounded end (left), which at microscopic scale

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–2894 2893
and why others cluster in a 4 m2 area, within a 50 cm depth of eachother (Table 1). Spatial analysis of hominid and other mammalremains, bone and possible stone tools, is also required to helpaddress the question of who used the bone tools. Lockwood et al.(2007) recently proposed that the high degree of sexualdimorphism observed in P. robustus from Swartkrans, Drimolen andKromdraai, all situated in the Cradle of Humankind, South Africa,may reflect social structures comparable to that of gorillas, and thatstable groups of females formed in response to the distribution offood sources. Female aggregation is also known amongst chim-panzees, where in some populations tools are used to forage fortermites (Whiten et al., 1999). Based on implement-assisted termiteforaging strategies amongst chimpanzees, we have inferred similarcultural behaviours for early hominids (d’Errico et al., 2001). In lightof the female aggregation models mentioned, we can speculate thatif P. robustus was the user of the bone tools, then the foragingactivity in which they were used may have been conducted mainlyby females.
5. Conclusion
Although the presence of bone tools at Drimolen has beenmentioned before in the literature, a detailed assessment of thismaterial was needed. Our study has reduced from 23 to 14 thenumber of pieces bearing modifications similar to those recordedon bone tools from other southern Africa early hominid sites, andconfirmed – by comparing the Drimolen specimens to an expandedcomparative collection – that the wear pattern recorded on the tipsmost likely results from use in digging, which we posit was asso-ciated with termite foraging. This finding demonstrates that the useof bone tools at early hominid sites was apparently widespread, andconsidering available information on fauna, geochronology and siteformation processes at Swartkrans and Drimolen (Vrba, 1982;Brain, 1993; Watson, 1993; Turner, 1997; Keyser et al., 2000; Curnoeet al., 2001; de Ruiter, 2003), lasted unchanged for at least 300,000years, and possibly as long as one million years. The association ofa high number of Paranthropus remains with bone tools and thevirtual absence of stone tools at Drimolen supports the hypothesis,first proposed by Brain and Shipman (1993) and followed by us(2001, 2003), that P. robustus may have used these bone tools.
Acknowledgements
We thank Andre Keyser and Colin Menter for inviting us todescribe the material, and for polishing the final version of themanuscript. We are grateful to Dominique Armand for assistancewith bone identification, Cynthia Kemp for editing a draft of thismanuscript, and two anonymous reviewers for their helpful com-ments. This research was funded by the National Research Foun-dation (NRF) in South Africa, University of the WitwatersrandResearch Council (URC), Palaeontological Scientific Trust (PAST),Cultural Service of the French Embassy in South Africa, FrenchMinistry of Research and Education, and Origin of Man, Languageand Languages (OMLL) Programme of the European ScienceFoundation.
References
Backwell, L.R., 2000. A Critical Assessment of Southern African Early Hominid BoneTools. MSc. dissertation, University of the Witwatersrand, South Africa.
Backwell, L.R., d’Errico, F., 2001. Evidence of termite foraging by Swartkrans earlyhominids. Proceedings of the National Academy of Sciences of the United Statesof America 98 (4), 1358–1363.
Backwell, L.R., d’Errico, F., 2003. Additional evidence on the early hominid bonetools from Swartkrans with reference to spatial distribution of lithic and organicartefacts. South African Journal of Science 99, 259–267.
Backwell, L.R., d’Errico, F., 2004. A reassessment of the Olduvai Gorge ‘bone tools’.Palaeontologia Africana 40, 95–158.
Backwell, L.R., d’Errico, F., 2005. The origin of bone tool technology and theidentification of early hominid cultural traditions. In: d’Errico, F., Backwell, L.R.(Eds.), From Tools to Symbols. From Early Hominids to Modern Humans. WitsUniversity Press, Johannesburg, pp. 238–275.
Bastin, A.H., 1932. Nouvelle contribution a l’etude de l’utilisation de l’os au Paleo-lithique inferieur. 1. L’epieu acheuleen de Saint-Mard (Aisne). 2. L’humerusdebite de Saratov (U.R.S.S.). Bulletin de la Societe Prehistorique Française 29,458–465.
Binford, L.R., 1981. Bones, Ancient Men and Modern Myths. Academic Press, NewYork.
Brain, C.K., 1967. Bone weathering and the problem of bone pseudo-tools. SouthAfrican Journal of Science 63, 97–99.
Brain, C.K., 1974. Some suggested procedures in the analysis of bone accumulationsfrom southern African Quaternary sites. Annals of the Transvaal Museum 29 (1),1–8.
Brain, C.K., 1981. The Hunters or The Hunted? An Introduction to African CaveTaphonomy. University of Chicago Press.
Brain, C.K., 1989. The evidence of bone modification by early hominids in southernAfrica. In: Bonnichsen, R., Sorg, M.H. (Eds.), Bone Modification. University ofMaine Centre for the Study of the First Americans. Thompson-Shore Inc., Orono,United States of America, pp. 291–297.
Brain, C.K., 1993. Swartkrans. A Cave’s Chronicle of Early Man. In: Transvaal MuseumMonograph, No. 8 Pretoria.
Brain, C.K., Shipman, P., 1993. The Swartkrans bone tools. In: Brain, C.K. (Ed.),Swartkrans: a Cave’s Chronicle of Early Man. Palaeontology Scientific Trust, pp.195–215.
Brain, C.K., Shipman, P., 2004. The Swartkrans bone tools. In: Brain, C.K. (Ed.),Swartkrans, a Cave’s Chronicle of Early ManTransvaal Museum Monograph,second ed., No. 8, pp. 195–215.
Brain, C.K., Churcher, C.S., Clark, J.D., Grine, F.E., Shipman, P., Susman, R.L., Turner, A.,Watson, V., 1988. New evidence of early hominids, their culture and environ-ment from the Swartkrans cave, South Africa. South African Journal of Science84, 828–835.
Breuil, H., 1932. Pointe d’epieu en os du Mousterien de la caverne du Castillo(Santander). L’Anthropologie 42, 679–680.
Breuil, H., 1938. Bone and antler industry of the Chou-Koutien Sinanthropus site.Paleontologia Sinica n.s. 6 Pekin.
Bunn, H.T., 1981. Archaeological evidence for meat-eating by Plio-Pleistocenehominids from Koobi Fora and Olduvai Gorge. Nature 291, 574–577.
Bunn, H.T., 1983. Comparative analysis of modern bone assemblages from a Sanhunter-gatherer camp in the Kalahari desert, Botswana, and from a spottedhyena den near Nairobi, Kenya. In: Clutton-Brock, J., Grigson, C. (Eds.), Animalsand Archaeology: 1. Hunters and their Prey. British Archaeological ReportsInternational Series 163, pp. 143–148.
Choyke, A.M., Bartosiewicz, L., 2001. Crafting Bone: Skeletal Technologies ThroughTime and Space. British Archaeological Reports International Series 937, Oxford.
Curnoe, D., Grun, R., Taylor, L., Thackeray, F., 2001. Direct ESR dating of a Pliocenehominin from Swartkrans. Journal of Human Evolution 40, 379–391.
Dart, R.A., 1957. The Osteodontokeratic Culture of Australopithecus prometheus.Transvaal Museum Memoirs, No. 10, Pretoria.
Dart, R., Kitching, J., 1958. Bone tools at the Kalkbank Middle Stone Age site and theMakapansgat australopithecine locality, central Transvaal. Part 2. The Osteo-dontokeratic contribution. Archaeological Bulletin 13 (51), 94–116.
d’Errico, F., 1993a. Identification des traces de manipulation, suspension, polissagesur des objets d’art mobilier en os, bois de cervides, ivoire. In: Traces et fonc-tion: les Gestes Retrouves. ERAUL, Liege, pp. 177–188. 50.
d’Errico, F., 1993b. Criteria for identifying utilised bone: the case of the Cantabrian‘Tensors’. Current Anthropology 34 (3), 298–311.
d’Errico, F., Villa, P., 1997. Holes and grooves: the contribution of microscopy andtaphonomy to the problem of art origins. Journal of Human Evolution 33,1–31.
d’Errico, F., Backwell, L.R., 2003. Possible evidence of bone tool shaping from theearly hominid site of Swartkrans, South Africa. Journal of Archaeological Science30, 1559–1576.
d’Errico, F., Giacobini, G., Puech, P.-F., 1984. Varnish replicas. A new method forstudying worked bone surfaces, OSSA. International Journal of Skeletal Research9/10, 29–51.
d’Errico, F., Backwell, L.R., Berger, L.R., 2001. Bone tool use in termite foraging byearly hominids and its impact on understanding early hominid behaviour.South African Journal of Science 97, 71–75.
Fisher, J.W., 1995. Bone surface modifications in zooarchaeology. Journal ofArchaeological Method and Theory 2 (1), 7–68.
Gautier, A., 1986. Une histoire de dents: les soi-disant incisives travaillees duPaleolithique moyen de Sclayn. Helinium 26, 177–181.
Gifford-Gonzalez, D., 1989. Ethnographic analogues for interpreting modifiedbones: some cases from East Africa. In: Bonnichsen, R., Sorg, M.H. (Eds.), BoneModification. University of Maine Centre for the Study of the First Americans.Thompson-Shore Inc., Orono, United States of America, pp. 179–246.
Hannus, L.A., Rossum, L., Winham, R.P., 1997. Proceedings of the 1993 Bone Modi-fication Conference, Hot Springs, South Dakota. In: Occasional Publication, No. 1.Archeology Laboratory, Augustana College, Sioux Falls, USA.
Haynes, G., 1988. Longitudinal studies of African elephant death and bone deposits.Journal of Archaeological Science 15, 131–157.
Haynes, G., 1991. Mammoths, Mastodonts, and Elephants: Biology, Behaviour andthe Fossil Record. Cambridge University Press.
Keyser, A., May 2000a. New finds in South Africa, National Geographic, pp. 77–83.

L. Backwell, F. d’Errico / Journal of Archaeological Science 35 (2008) 2880–28942894
Keyser, A., 2000b. The Drimolen skull: the most complete australopithecinecranium and mandible to date. South African Journal of Science 96,189–193.
Keyser, A., Menter, C.G., Moggi-Cecchi, J., Pickering, T.R., Berger, L.R., 2000. Drimo-len: a new hominid-bearing site in Gauteng, South Africa. South African Journalof Science 96, 193–197.
Klein, R.G., 1975. Palaeoanthropological implications of the non-archaeologicalbone assemblage from Swartklip I, south-western Cape Province, South Africa.Quaternary Research 5, 275–288.
Leakey, M.D., 1971. Olduvai Gorge. Vol. 3. Excavations in Beds I and II. CambridgeUniversity Press.
LeMoine, G., 1997. Use Wear Analysis on Bone and Antler Tools of the MackenzieInuit. British Archaeological Reports International Series 679, Oxford.
Lockwood, C., Menter, C., Keyser, A., Moggi-Cecchi, J., 2007. Extended male growthin a fossil hominin species. Science 318, 1443–1446.
Lyman, R.L., 1994. Vertebrate Taphonomy. Cambridge University Press.Maguire, J.M., Pemberton, D., Collett, M.H., 1980. The Makapansgat Limeworks Grey
breccia: hominids, hyaenas, hystricids or hillwash? Palaeontologia Africana 23,75–98.
Mason, R., 1958. Bone tools at the Kalkbank Middle Stone Age site and the Maka-pansgat australopithecine locality, central Transvaal. Part 1. The Kalkbank site.Archaeological Bulletin 13 (51), 85–93.
Netterberg, F., 1974. Observations on calcretes at some archaeological and palae-ontological sites in southern Africa. The South African Archaeological SocietyGoodwin Series 2, 20–24.
Njau, J.K., Blumenschine, R., 2006. A diagnosis of crocodile feeding traces on largermammal bone, with fossil examples from the Plio-Pleistocene Olduvai Basin,Tanzania. Journal of Human Evolution 50, 142–162.
Olsen, S.L., 1989. On distinguishing natural from cultural damage on archaeologicalantler. Journal of Archaeological Science 16, 125–135.
Olsen, S.L., Shipman, P., 1988. Surface modification of bone: Trampling versusbutchery. Journal of Archaeological Science 15, 535–553.
Palmer, A.N., 1991. Origin and morphology of limestone caves. Geological Society ofAmerica Bulletin 103, 1–21.
Pei, W.C., 1938. Le role des animaux et des causes naturelles dans la cassure des os.Palaeontologia Sinica 7, 1–16.
Plummer, T.W., Stanford, C.B., 2000. Analysis of a bone assemblage made bychimpanzees at Gombe National Park, Tanzania. Journal of Human Evolution 39,345–365.
de Ruiter, D.J., 2003. A revised faunal list for the Australopithecine-bearingMembers of Swartkrans. Annals of the Transvaal Museum 39, 29–41.
Robinson, J.T., 1959. A bone implement from Sterkfontein. Nature 184, 583–585.
Scheinsohn, V.G., 1997. Use-wear patterns on bark removers. In: Hannus, L.A.,Rossum, L., Winham, R.P. (Eds.), Proceedings of the 1993 Bone ModificationConference, Hot Springs, South Dakota. Occasional Publication No. 1. Archeol-ogy Laboratory, Augustana College, Sioux Falls, USA, pp. 265–276.
Schmidtgen, O., 1929. Knochenartefakte aus den Mosbacher Sanden. Jahrbucher desNassauischen Vereins fur Naturkunde, Wiesbaden. 80, 1–6.
Selvaggio, M.M., Wilder, J., 2001. Identifying the involvement of multiple carnivoretaxa with archaeological bone assemblages. Journal of Archaeological Science28, 465–470.
Shipman, P., 1984. The earliest tools: re-assessing the evidence from Olduvai Gorge.Anthroquest 29, 9–10.
Shipman, P., 1989. Altered bones from Olduvai Gorge, Tanzania: techniques, prob-lems, and implications of their recognition. In: Bonnichsen, R., Sorg, M.H. (Eds.),Bone Modification. University of Maine Centre for the Study of the FirstAmericans. Thompson-Shore Inc., Orono, United States of America, pp. 317–334.
Shipman, P., Phillips, J., 1976. On scavenging by hominids and other carnivores.Current Anthropology 17 (1), 170–172.
Shipman, P., Rose, J., 1984. Cutmark mimics on modern and fossil bovid bones.Current Anthropology 25 (1), 116–117.
Sutcliffe, A.J., 1973. Similarity of bones and antlers gnawed by deer to humanartefacts. Nature 246, 428–430.
Sutcliffe, A.J., 1977. Further notes on bones and antler chewed by deer and otherungulates. Deer 4 (2), 73–82.
Turner, A., 1997. Further remains of Carnivora (Mammalia) from the Sterkfonteinhominid site. Palaeontologia Africana 34, 115–126.
Villa, P., Bartram, L., 1996. Flaked bone from a hyena den. Paleo 8, 143–159.Villa, P., d’Errico, F., 2001. Bone and ivory points in the Lower and Middle Paleolithic
of Europe. Journal of Human Evolution 41, 69–112.Vrba, E.S., 1982. Biostratigraphy and chronology, based particularly on Bovidae, of
southern hominid-associated assemblages: Makapansgat, Sterkfontein, Taung,Kromdraai, Swartkrans; also Elandsfontein (Saldanha), Broken Hill (nowKabwe) and Cave of Hearths. In: De Lumley, M.A. (Ed.), Homo erectus et la Placede l’Homme de Tautavel parmi les Hominides Fossiles. Premiere CongresInternational de Paleontologie Humaine, Vol. 2. UNESCO Colloque Internationaldu Centre National de la Recherche Scientifique, France, pp. 707–752.
Watson, V., 1993. In: Brain, C.K. (Ed.), Swartkrans. A Cave’s Chronicle of Early Man.Transvaal Museum Monograph, No. 8, pp. 35–73. Pretoria.
White, T.D., 1992. Prehistoric Cannibalism at Mancos 5MTUMR-2346. PrincetonUniversity Press, New Jersey. United States of America.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C.E.G., Wrangham, R.W., Boesch, C., 1999. Cultures in chimpanzees. Nature 399,682–685.