
The Functional Morphology of the Hominid Foot.
69
The Functional Morphology of the Hominoid Foot Francesca Campbell BSc Registration number 120189663 UCard number 001561981 A report submitted in part fulfilment of the examination requirements for the award of an MSc in Palaeoanthropology awarded by the University of Sheffield, 2013, supervised by Dr. Kevin Kuykendall.
Transcript of The Functional Morphology of the Hominid Foot.

The Functional Morphology of the
Hominoid Foot
Francesca Campbell BSc
Registration number 120189663
UCard number 001561981
A report submitted in part fulfilment of the examination requirements for the award of an MSc
in Palaeoanthropology awarded by the University of Sheffield, 2013, supervised by Dr. Kevin
Kuykendall.

2
Certificate of Originality
This is to certify that I am responsible for the work submitted in this thesis, that the original
work is my own, except as specified in the acknowledgments and in references, and that neither
the thesis nor the original work contained therein has been previously submitted to any other
institution for a degree.
Name: Francesca Campbell
Signed:
Date:

3
Acknowledgements
The author wishes to thank;
Dr. Kevin Kuykendall for supervising this study.
Inbal Livne at The Powell-Cotton Museum for help and advice during data collection.
Prof. Tim Birkhead at The Alfred Denny Museum for help and advice during data collection.
Mark Edwards for advice, assistance and patience.

4
Abbreviations
ABBREVIATION FULL DESCRIPTION
G. gorilla Gorilla gorilla Where G. gorilla is used, this refers to the sub species Gorilla
gorillla gorilla, the western lowland gorilla. G.g.diehli, or the
upland gorilla, is not used in this study.
H. erectus Homo erectus 1.8 - 0.2 mya
H. ergaster Homo ergaster 1.9 - 1.4 mya
H. habilis Homo habilis 2.3 - 1.4 mya
H.
heidelbergensis
Homo
heidelbergensis
0.6 - 0.35
H.
neanderthalensi
s
Homo
neanderthalensis
0.35 - 0.03 mya
H. sapiens Homo sapiens Modern humans, sometimes refered to a sub species - H. sapiens
sapiens
MT Metatarsal Metatarsal bones, numbered from medial to lateral, I - V
OH 8 Olduvai Hominid
8
H. habilis assemblage, c. 18 mya, discovered in Olduvai Gorge,
Tanzania

5
P. troglodtyes Pan troglodytes Where P. troglodytes is used, this refers to the chimpanzee species,
not to be confused with 'pygmy' chimpanzees, bonobos (P.
pygmaeus). Since this study uses research cited before the
recognition of the bonobo as a separate species, where the common
name 'chimpanzee' is used, specimens may be P. troglodtyes or P.
pygmaeus. All of the species used in this study's data collection are
P. troglodytes as specified.
PCA Principle
Component
Analysis
Data reduction technique
R R Statistics programming language developed by R Development and
Licensed by GNU General Public License
SPSS Statistical
Product and
Service Solutions
IBM SPSS statistics package
STW 573 Sterkfontein 573 The catalogue number of an Australopithecus fossil from South
Africa. Dating is difficult but most tenchniques range in the 3-4mya
bracket.

6
Contents
Page
1. Abstract 7
2. Introduction 9
3. Aims and Objectives 36
4. Materials and Method 37
5. Results 47
6. Discussion 54
7. Conclusion 59
8. Further Research 60
9. References 61
10. Appendices 68

7
1. Abstract
The functional morphology of the hominoid foot is reviewed with a particular emphasis on the
comparison of the species Gorilla gorilla, Pan troglodytes and Homo sapiens. Collated
measurements from specimens at the University of Sheffield and the Powell-Cotton Museum are
compared using linear divisions of the foot. Linear divisions are compared using descriptive
statistics using SPSS, average relative lengths using R, and finally analysed by PCA using SPSS.
Attention is given to the second toe due to the unique morphology of the adducted H. sapiens
hallux. From the comparative model of extant primates the foot bones of extinct homind bones
are analysed, with special attention paid to the most complete foot specimen; Homo ergaster
'Narikotome Boy', Little Foot (STW 573) and OH 8 Homo habilis. This work supports the
previous comparative work carried out by Adolph Schultz in the early 20th Century, the work
of John Napier, John Robinson in the 1960's and Will Harcourt-Smith in the late 20th and early
21st Century. The wide range of human variation is demonstrated and the results are discussed
in the context of there being a greater diversity in hominoid locomotion that the 'less upright -
more upright' model favoured for many years {fig1}. Recent developments in human walking
style are discussed in the context of a closer link between extinct species than previously
thought.

8
Francesca Campbell
Figure 1. the 'less upright to more upright' model of evolution

9
2. Introduction
2.1 Introduction
The evolution of mammals has created many diverse forms of locomotion, from brachiating
monkeys, swimming whales and bipedal humans. With the exception of the head, the axial
skeleton stays relatively conserved with the majority of evolutionary changes being driven by
locomotive style and behaviours, leading to diverse changes in the limbs (including the tail) and
head/brain. In the great apes, the comparative anatomy and serial homology of limbs has been
documented since Victorian times, but gained significant impact with the evolutionary studies
of Ernst Hæckel . More recent work with genetic material has led to an increased sophistication
of the homology theory; as OJ Lewis states
“homology has come to mean not merely similarity of position in a body; that is of
anatomical relationships, but also derivation by a similar pathway of development,
controlled by genealogically related genes” [Lewis, 1989, 27].
Early students of anatomy (skeletal, osteological and musculature) always considered the
similarities from a human point of view, determining the standard anatomical position in an
upright, standing pose {Fig 2}.

10
Figure 2. Leonardo Da Vinci’s Vitruve Luc Viatour, c. 1490, showing standard
anatomical position; erect, with forearm supinated. Image from
wikimediacommons.com

11
This view is seriously compromised by the development of the bipedal upper limb being
supinated in natural pose; in quadrupeds the upper or front limb is pronated and this is the
‘natural’ position for human limbs when assuming a position with all four limbs on the ground.
Despite the apparent differences in the function of the lower limb and foot in terrestrial
mammals due to body type and size, foot shape and function, arrangement of structures and
loading conditions, the study of comparative morphology of the foot shows many common
features of organization, structure and function.
Synthesis is clouded by the considerable variation in scale and proportions in distal limb
structure among quadrupeds, knuckle walkers and obligate bipeds. The posterior to anterior
mass loading mechanics of locomotion vary considerably due to mass and body shape, as do the
changes of distribution of mass during locomotion. Because of the wide variety of body shape,
size and mass, organization and morphology of the foot bones may vary between species.
Obligate bipedal locomotion and its temporal position in the hominid lineage is one of the most
significant events in hominid evolutionary history [Chaplin et al. 1994; Schmitt 2003; Harcourt-
Smith and Aiello, 2004]. There is considerable literature on the subject of the evolution of
hominid bipedalism, ranging from the hypothetical locomotion of last common ancestors
[Byrnes and Spence, 2011; Cartmill, 1974; Harcourt-Smith and Aiello, 2004; Jungers, 1982.,
Richmond and Strait, 2000; Richmond and Strait, 2001; Robinson and Robinson, 1972; Schmitt,
2003; Stern Jr and Susman, 1983], to ecological pressures which may have led to the positive
selection for bipedalism [Hunt, 1994; Krebs and Davies, 1978; Taylor and van Schaik, 2007] and
the degree of bipedalism exhibited by extinct hominids [Asfaw et al., 1999; Brunet et al., 2002;
Day and Wood, 1968; Gebo, 1992; Köhler and Moyà-Solà; 1997, Leakey, 1973; McHenry, 1994;
Stern Jr and Susman, 1983; White et al., 1995].

12
The foot of extant primates is extremely specialised [Ali and Meier, 2008; Cartmill, 1872] ;
evidence shows that the lower limb carries the majority of the stress of locomotion even in
arboreal great apes [Schultz, 1969]. Humans, as the only extant hominid obligate biped, show a
range of unique adaptions in both anatomy and pedal function that differs from both extant apes
and extinct hominids. In obligate bipedal locomotion the lower limbs must take the forces
generated by locomotion and the entire body weight through only one foot at a time as the
movement is transferred from swing to stance phases [Lovejoy, 1988; Schmitt, 2003]. During toe
off the morphology of the foot must be robust enough to accommodate these forces and as a
result the hallux is much larger relative to the other four toes in humans than other primates
[Schultz, 1969].
2.2 Osteology of the human foot
In humans and great apes there are twenty six bones in the foot, divided into three separate
groups. The tarsals are the robust bones of the ankle, the metatarsals the smaller bones of the
instep (forming the arch of the foot in humans) and the phalanges make up the toes (seven, five
and fourteen bones consecutively). The phalanges form a homologous structure with the hand
with five proximal phalanges, four middle phalanges (the hallux having no morphologically
distinct intermediate phalanx) and five distal phalanges {fig 3}.
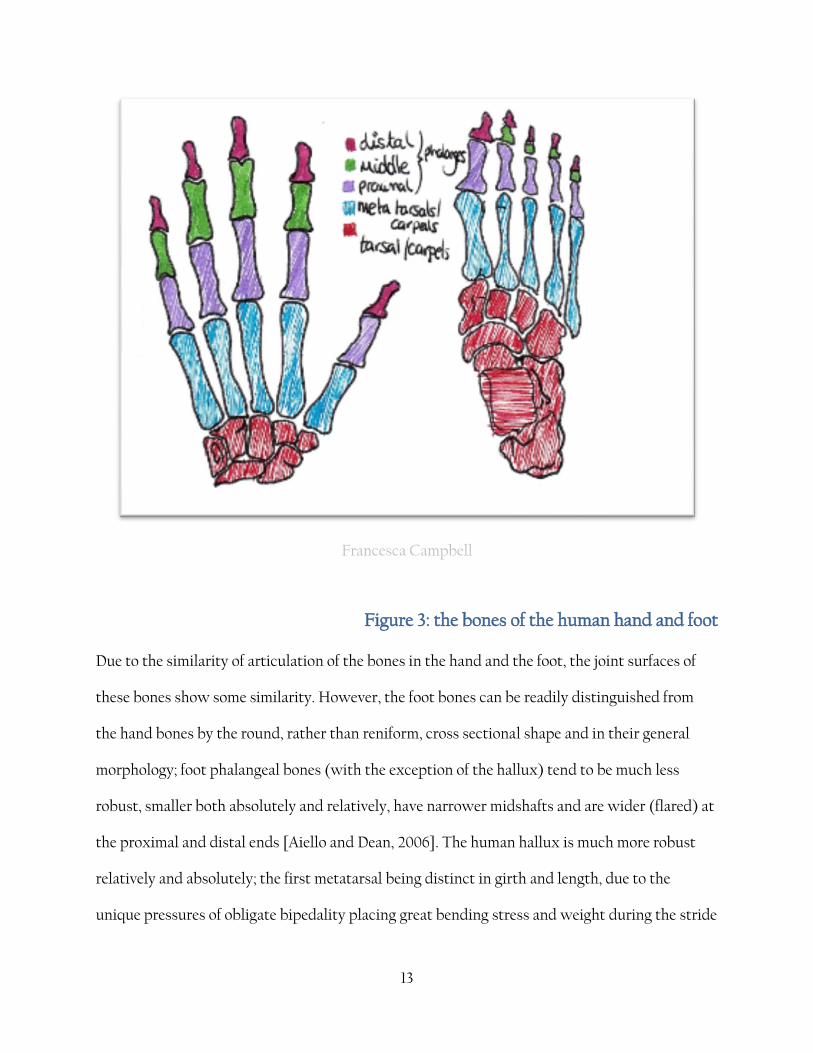
13
Francesca Campbell
Figure 3: the bones of the human hand and foot
Due to the similarity of articulation of the bones in the hand and the foot, the joint surfaces of
these bones show some similarity. However, the foot bones can be readily distinguished from
the hand bones by the round, rather than reniform, cross sectional shape and in their general
morphology; foot phalangeal bones (with the exception of the hallux) tend to be much less
robust, smaller both absolutely and relatively, have narrower midshafts and are wider (flared) at
the proximal and distal ends [Aiello and Dean, 2006]. The human hallux is much more robust
relatively and absolutely; the first metatarsal being distinct in girth and length, due to the
unique pressures of obligate bipedality placing great bending stress and weight during the stride

14
cycle, and distinct in shape; the medial head has an articular facet to allow dorsiflexion of the toe
during the toe-off phase.
The seven tarsal bones are the calcaneus, talus, navicular, three cuneiforms and the cuboid. The
calcaneus is the largest bone in the foot and forms the heel; it articulates with the talus, the
cuboid and the navicular. Identification and siding of the calcaneus and talus is not relatively
difficult since the bones are so distinctive and articulate in a regular fashion. The calcaneus has
its calcaneal tuberosity posterior and the non-articulating portion is inferior. Siding is not
difficult since the 'heel' portion is so distinctive and the position of the sustenaculum tali
indicates the side. For the talus, the saddle shaped facet is superior and the anterior point forms
the talar head. The large malleolar articulation with the fibula is on the lateral side. The talus is
the medium by which the foot is linked to the lower leg. In humans, the talus does not have a
muscle attached to it. The navicular is named for a ‘little ship’ (Latin) and bears a
correspondence to the scaphoid (‘a little boat’ in Greek) in the hand and from this the shape is
easily identified. It has a pronounced concave surface which articulates with the talus. The
convex side articulates with all three cuneiforms. The navicular tubercle is on the medial side.
The three cuneiform bones, medial, intermediate and lateral, are wedge shaped bones that form
the transverse arch of the human foot. The largest, the medial cuneiform, has a reniform facet
where it articulates with the talus. Therefore the longest surface is distal. The second largest is
the lateral cuneiform and the smallest is the intermediate. The second cuneiform (aka the
intermediate) articulates with both cuneiforms on either side. The broadest surface of the
cuneiform is dorsal and the proximal surface is the most concave as it articulates with the
navicular. The third cuneiform (lateral) is intermediate in size and articulates with MTII, III and
IV distally . The cuboid forms the top of the arch in the longitudinal arch of the human foot . The

15
cuboid articulates with the navicular and the third cuneiform and is identified by the cuboid
tuberosity. The non- articular portion lies supero-laterally and the articulting facet lies on the
medial side. Shape and angle of articular surfaces of the tarso-metatarsal joint have been used to
infer bipedality [Harcourt-Smith and Aiello, 2004; Sarmiento and Marcus, 2000] and the
articular facets of the navicular have been used in a similar manner [Sarmiento and Marcus,
2000]. Round articular facets on the medial cuneiform allow a wide degree of abduction in the
primate hallux. The human medial cuneiform has a compressed articular facet which holds the
hallux into an adducted position, parallel to the other rays with extremely limited, if any, lateral
movement [Aeillo and Dean, 2006] {Figure 4}.

16
Francesca Campbell
Figure 4. The bones of the human foot. Redrawn from Aiello and Dean [2006, 408]
Compared to Pan and Gorilla the human calcaneus is relatively robust and large. In humans the
calcaneal tuberosity is the first part of the body to make contact with the ground during the
stride cycle {fig 6} and as such is larger and more robust in order to provide stability. The
calcaneus serves as an attachment point for the ligaments forming the arches of the foot; these in
turn act as a spring or shock absorber [Aeillo and Dean, 2006]. The metatarsals, like the
metacarpals, are all tubular bones with the distal articular heads forming rounded stubs and the
proximal heads forming square stubs. The metatarsals are concave in lateral view and the bases

17
of MTII-V articulate with each neighbouring pair. Each of the tarsals in the distal row articulate
with at least one MT base and it is from the combination of the convex profile and the articular
facets on the base that the MT can be identified, sided and ordered. The first MT is more
massive in humans than in Gorilla or Pan but even in these two species the first MT is still
distinctive by its robusticity. Sesamoid bones present in the tendons of the muscle flexor
hallucis brevis articulate with the MTI head; in apes (especially larger species such as Pongo and
Gorilla) these sesamoid bones tend to be larger and so by extension create larger articular
facets. MTII is, in humans, the longest MT. It has a very narrow shaft in humans, Gorilla and Pan
and its two lateral facets articulate with the lateral cuneiform and MTIII. It also articulates
proximally with the intemediate cuneiform and with the medial cuneiform on the medial edge.
The base of MTII is also less platform-like than MTIII and MTIV. For this reason it may be
confused with the styloid process of MTV, especially in fragmentary assemblages; however the
articular morphology is distinctive enough for this not to be an issue for this study. MTIIIs
medial facets are similar in position to MTII but are much smaller and the base of MTIII is much
squarer due to the slight styloid projection on the MTII. MTIII also has a large tubercule on the
lateral edge. MTIV has a slightly more robust shaft than the previous two MT and it has
articulating factets for the MTIII and MTV, medially and laterally respectively. MTV is
extremely distinctive due to the protruding styloid process on the lateral side. The base of the
MTV articulates with the cuboid and forms a 'footprint' shape [White and Folkens, 2005].
As discussed previously, positioning the phalanges is difficult and so the method employed here
must be treated with a degree of consideration. The phalanges for the hallux are more
distinguishable and by comparison can be sided. The hallux lacks an intermediate phalanx and
therefore the distal phalanx is larger and more robust than the other distal phalanges. The

18
remaining bones can be sorted into proximal, intermediate and distal phalanxes (3 for each ray,
12 in total) and can be sorted by comparing the associated MT head and the size compared to
other rays. Distal phalanxes II - V are almost impossible to order due to their relatively uniform
size and shape.
It has been suggested that differences in the human and ape foot are a direct result of the
remodelling of the human foot around the subtalar axis of the primitive prehensile primate foot
as a direct result of a more bipedal stance [Keith, 1929; Lewis, 1981]. It is around the subtalar
axis that the foot everts and inverts[Aeillo and Dean, 2004]. The subtalar axis differs from
humans, Pan and Gorilla in its gradient. It is steeper dorsoplanarly in humans than in apes due
to the relatively large curve of the articular surfaces in the human foot. Because of this sharp
gradient, lateral movement is severely limited in humans while apes still maintain a certain
degree of flexibility at the subtalar joint.
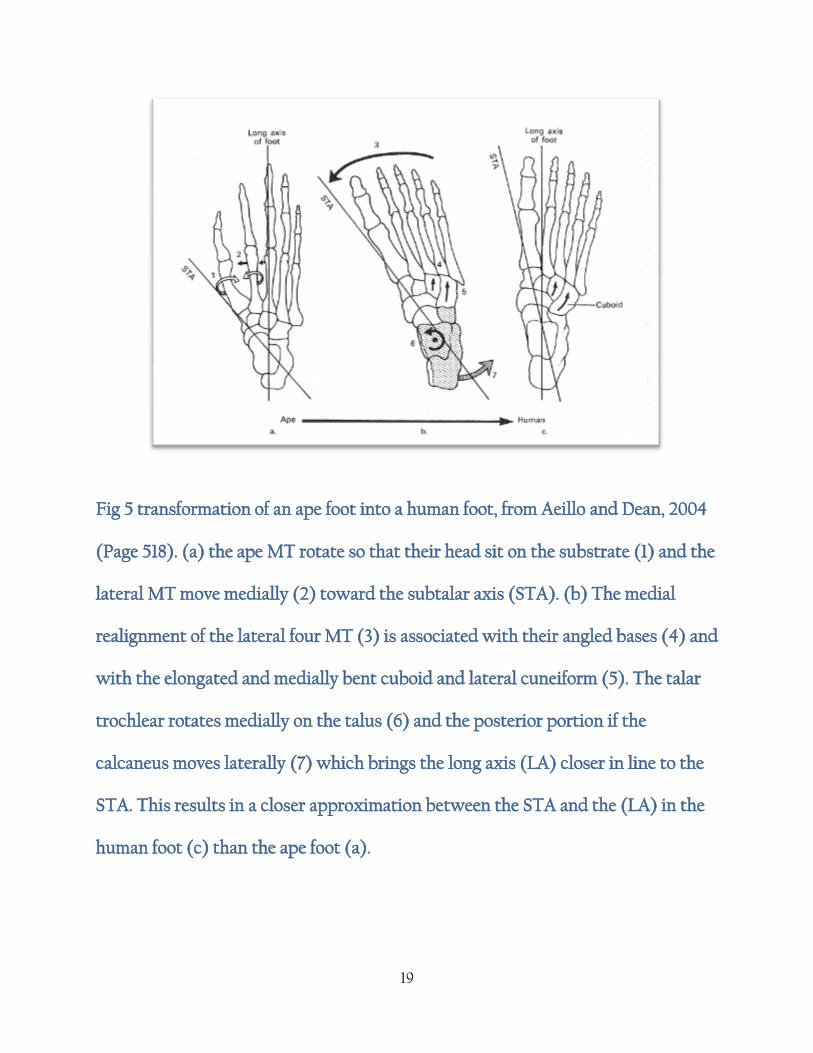
19
Fig 5 transformation of an ape foot into a human foot, from Aeillo and Dean, 2004
(Page 518). (a) the ape MT rotate so that their head sit on the substrate (1) and the
lateral MT move medially (2) toward the subtalar axis (STA). (b) The medial
realignment of the lateral four MT (3) is associated with their angled bases (4) and
with the elongated and medially bent cuboid and lateral cuneiform (5). The talar
trochlear rotates medially on the talus (6) and the posterior portion if the
calcaneus moves laterally (7) which brings the long axis (LA) closer in line to the
STA. This results in a closer approximation between the STA and the (LA) in the
human foot (c) than the ape foot (a).

20
For this reason it may be prudent to consider the functional divisions of the foot in three
segments, taking into account the lever arm portions of the foot as explained by Schultz [1963]
{fig 6}.
Fig 6 Bone and lever lengths of the human foot. After [Schultz, 1963]
The forefoot – the most distal portion of the foot, providing leverage and load transfer
properties.
The midfoot – the intermediate portion of the foot, providing force transmission through the
transverse tarsal joint from the hindfoot (subtalar joint) to the forefoot (metatarso and inter-
phalangeal joints).
The hindfoot – The most proximal part of the foot, containing the most robust structures for
transmitting force between the proximal and distal aspects of the foot during locomotion and
load bearing in standing phase. This is also the area the foot articulates with the bones of the leg,
forming the ankle joint.
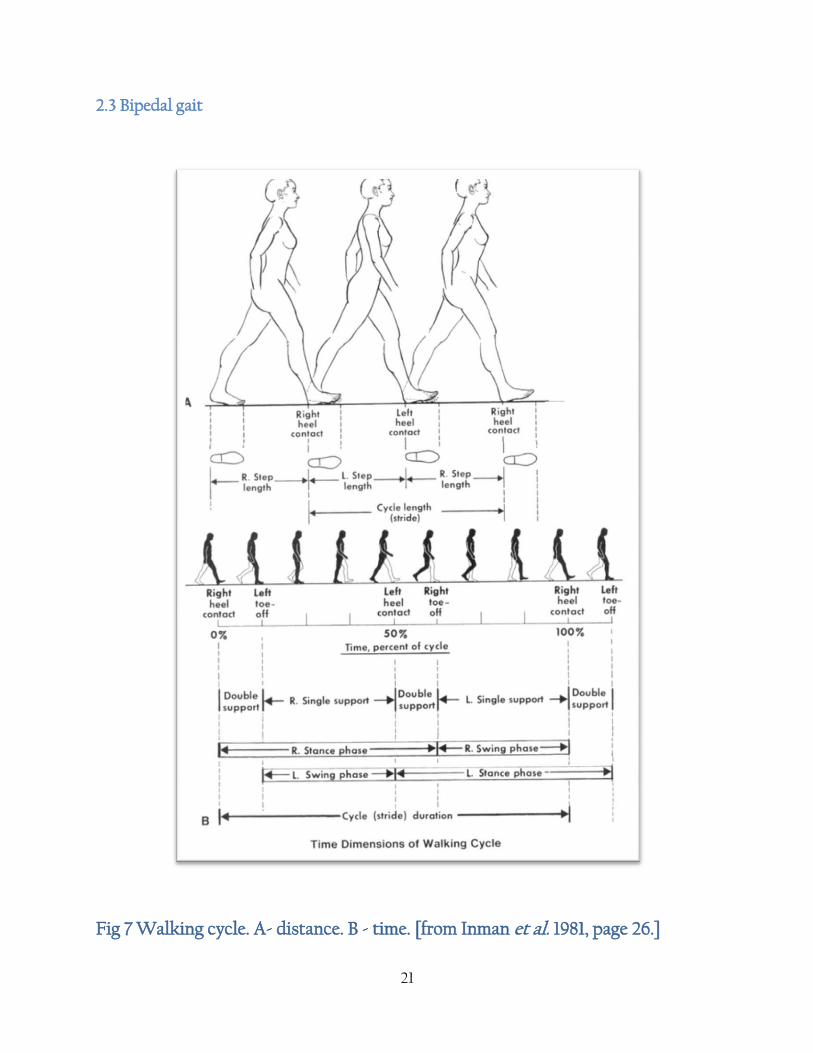
21
2.3 Bipedal gait
Fig 7 Walking cycle. A- distance. B - time. [from Inman et al. 1981, page 26.]

22
Phases of the gait cycle from the foot striking the ground to the same foot striking ground again.
Initial contact with substrate - extended knee, ankle in neutral anatomical position. Heel
contacts the substrate first.
Loading response - during loading knee flexes, ankle plantarflexes and gluteal muscles
remain active to maintain hip stability.
Mistance - begins with toe off. By midstance the knee is extended and the ankle is in
neutral anatomical position.
Terminal stance - Centre of gravity is maintained over supporting foot.
Pre-swing - pushes centre of gravity over supporting foot and brings opposing foot
forward to heel strike. Knee flexes and ankle plantar flexes. Toes dorsiflex at
metatarsaophalangeal joints.
Subtalar joint assumes near neutral anatomical position.
2.4 The ape gait
Walking gait differs in apes due to the morphological and musculature differences in the ape hip
and lower limb. Neither the hip joint or ankle joint are fully extended during walking and the
knee and ankle joints both stay in front of the hip during the stance phase. As a result ape stride
is shorter than human stride length and greater lateral placement is required to maintain the
centre of gravity. The swing phase is slightly different in apes too, as the musculature in the hip
does not allow for full abductor capabilities in the ape, and so the entire trunk is shifted across
the lateral plane. This style of walking requires more energy to maintain than full bipedality in
humans, and this may be why the arboreal origins theory of bipedality may have gained favour,

23
since arboreal movements more closely mimic human bipedality in kinematics and energy
expenditure. This theory may be quiet attractive to those seeking an answer to why our
ancestors stood tall and our cousins did not [Aeillo and Dean, 2004].
2.5 The extant Hominoid foot
There are a number of differences between the ape and human foot, the greatest being the
opposability of the hallux in apes, a feature which is lost in humans. These differences are due to
differing style of locomotion and lower limb adaptations as a result of bipedal stance. The fully
adducted hallux reduces grasping ability in the human toes, and in general the other toes are
relatively shorter in length than in other primates. Walking bipedally with abducted hallux
restricts efficient bipedalism and so relative toe length is likely an adaptation of obligate
biedalism . In human walking, the lateral posterior aspect of the calcaneus comes into contact
with the ground (the heel). The foot then levers forward onto the lateral metatarsal (MV, or the
little toe) and the weight is then transferred medially across the foot, to the ball of the foot at the
end of MI. {fig7} The entire weight of the body is then placed on the hallux as the opposing leg
swings forward. The last part of the foot to leave the ground is the hallux during toe-off, just
preceding or in conjunction with the heel strike of the opposing foot. There is some debate,
however, about how much a difference wearing shoes makes to this arrangement in modern
human locomotion. Studies have shown that in some individuals the weight is borne on all five
metatarsal head simultaneously instead of the rolling weight transfer from lateral to medial. This
seems especially true with the common place wearing of heeled shoes (not just tall stilettos
favoured majorly by women, but even heels with a 2.5cm lift may have some effect) [Frey,
1993].Variations in the kinematics of walking and running are well documented in sports
science and athletic development but the features described above are the most common

24
trademarks of human bipedal walking and in these features the differences in the ape foot are
highlighted. For example, in P. troglodytes there is no medial transfer; the weight being
transferred from posterior to anterior in a uniform line. The lateral border of the foot comes into
contact with the ground at the same time as heel strike and the medial border follows soon after
as the foot is placed flat (or as flat as the opposable hallux will allow) on the ground. The second
and third toes are the last remaining contact with the ground, rather than the first toe in
humans. The ape footprint also exhibits a phenomenom called the ‘midtarsal break’ due to the
increased mobility of the calcaneocuboid joint in the ape foot. As the calcaneus leaves the
ground, the foot pivots at the calcaneocuboid joint and part of the lateral side of the foot comes
in contact to the ground. In humans this would be an impossibility due to the relatively high
arch of the foot and the mobility in the calcaneocuboid joint to facilitate bipedal load bearing.
2.6 The proportions of the foot
Aside from the adducted hallux in apes, a major difference between the great ape and modern
human foot is the proportion of the length of the phalanges. Humans have much shorter third
toes in relation to the other toes and the total length of the foot than great apes. In Gorilla, the
third toe represents 33% of the total length of the foot and chimpanzees it is 35% of the total
length, while in humans it is only 18% [Keith, 1929] (Schultz [1969] reports 32% for gorillas and
19% for humans.) Shorter lateral toes appears to be a response to a more terrestrial form of
locomotion, since predominantly terrestrial primates such as geladas also exhibit shorter lateral
toes relatively. The sequence of toe length also differs from humans to apes. Total toe length is
sequential in humans, from I to V. In apes, the sequence goes from III being the longest, IV, II, V
and finally I the shortest [Schultz, 1969]. Lever length in humans is also longer in relation to
trunk length than apes, despite having shorter foot length in total. Metatarsal robusticity differs

25
between humans and apes; as may be expected due to the loading forces {fig 6}; MI in humans is
very robust, followed by MV due to the large proportion of weight bearing during the lateral roll
on to the ball of the foot during walking phases. In apes, MV is the least robust due to the
majority of the weight being placed more medially towards MI during locomotion.
Figure. Representation of the average relative lengths of the functional divisions of
the foot, proportional [tarsal + MIII length = 100]. (after Schultz, 1963, reproduced
in Schultz, 1969 and Aiello and Dean, 2006)
2. 7 Fossil foot bones
Whilst early Miocene foot bones of primates have been known for a few decades, the primitive
condition in the human lineage is still under much scrutiny. Gorilla and Pan share features
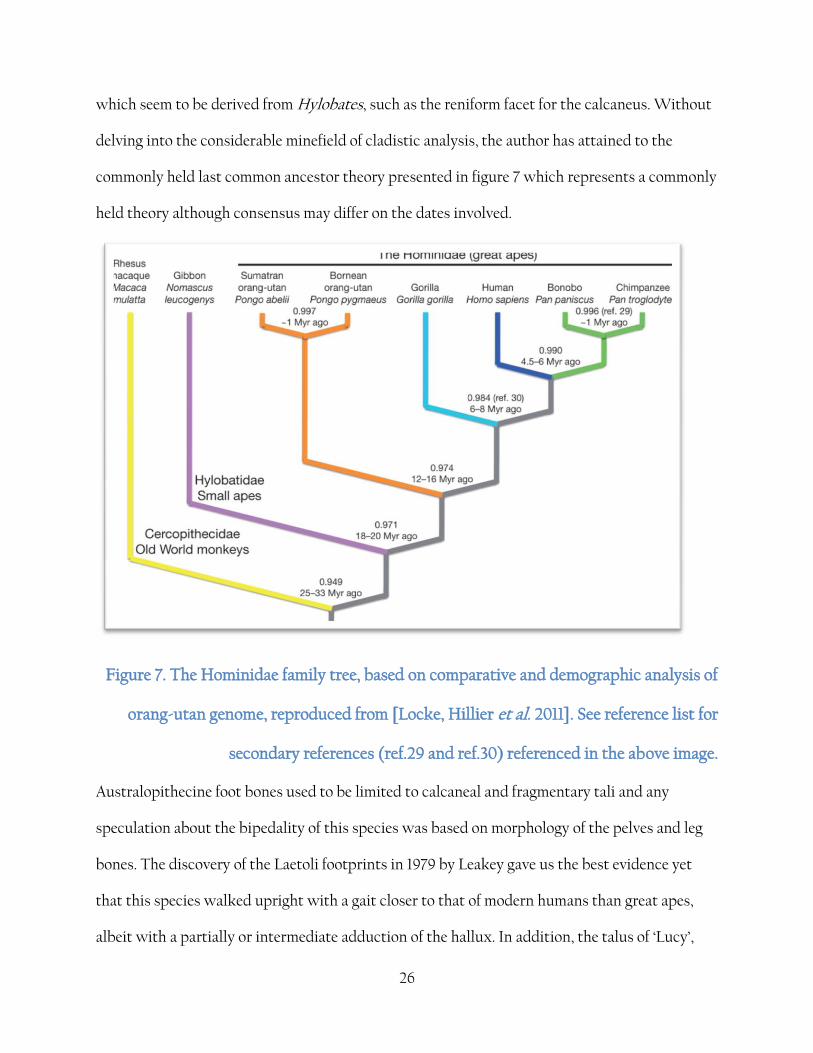
26
which seem to be derived from Hylobates, such as the reniform facet for the calcaneus. Without
delving into the considerable minefield of cladistic analysis, the author has attained to the
commonly held last common ancestor theory presented in figure 7 which represents a commonly
held theory although consensus may differ on the dates involved.
Figure 7. The Hominidae family tree, based on comparative and demographic analysis of
orang-utan genome, reproduced from [Locke, Hillier et al. 2011]. See reference list for
secondary references (ref.29 and ref.30) referenced in the above image.
Australopithecine foot bones used to be limited to calcaneal and fragmentary tali and any
speculation about the bipedality of this species was based on morphology of the pelves and leg
bones. The discovery of the Laetoli footprints in 1979 by Leakey gave us the best evidence yet
that this species walked upright with a gait closer to that of modern humans than great apes,
albeit with a partially or intermediate adduction of the hallux. In addition, the talus of ‘Lucy’,

27
regardless of its fragmentary nature, seemed to suggest a relationship with the leg bones more
similar to that of the great apes [Lewis, 1989]. In 1996 a 3.2 million year old complete MIV fossil
was discovered in Hadar, Ethiopia by William Kimbel and the shape and robusticity of this bone
seemed comparable to that of modern humans; the orientation of both ends of the bone reflect
the longitudinal arch of the foot and the flat tarsal facets suggest that the midfoot was lacking a
midtarsal break which is so indicative of the morphology of the ape foot. This view was
challenged in 2012 by Yohannes Haile-Selassie when pedal elements from a 3.4 million year old
fossil also from Ethiopia seemed to suggest a species which had morphology more similar to the
primitive form of Ardipithecus ramidus, with a more abducted hallux and the possibility of
opposability in the morphology of the pedal elements, leading to the possibility that a mosaic of
different features across a number of contemporaneous species existed.
Francesca Campbell
Figure 8. The bones of the Burtele BRT-VP - 2/736 foot, drawn to a G. gorilla
footprint, from Haile-Sellaise, [2011]

28
The temporally earlier Ar. ramidus material also exhibits a mosaic of primitive and derived
features with evidence pointing to a fully abductable hallux like the primitive state, but little
lateral midfoot flexibility as shown in the more derived forms, with little evidence of a midtarsal
break.
The Homo habilis foot is best represented by an almost complete foot fossil (OH 8). The tarsals
and metatarsals have unfortunate omissions, with the metatarsal heads broken away and the
calcaneal tuber eroded, but the subsequent discovery of more material from the same site has
revealed a complete ankle joint [Lewis, 1989]. Although the reconstruction of the soft tissues of
the articulations of the OH foot have been debated, OH 8 is remarkably Pan like in proportion
and morphology.
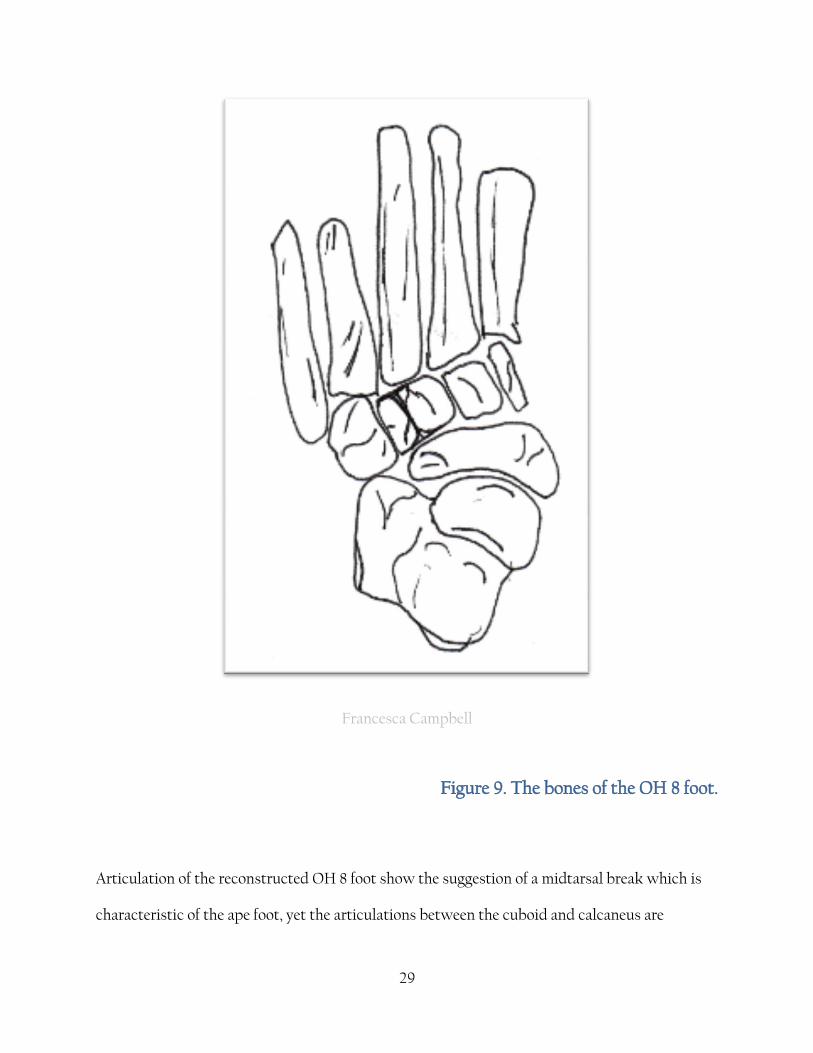
29
Francesca Campbell
Figure 9. The bones of the OH 8 foot.
Articulation of the reconstructed OH 8 foot show the suggestion of a midtarsal break which is
characteristic of the ape foot, yet the articulations between the cuboid and calcaneus are

30
reminiscent of the ‘close-packed’ situation developed in the human foot to give stability during
load bearing [Lewis, 1989]. This evidence has led to the theory that the flexibility of the Pan and
Gorilla foot is a derived feature . Lewis [1989] and Kidd [1999] both challenge the conventional
view that the human foot is derived from a Gorilla type foot and instead propose a more vice
versa model where the ape foot is more derived from a human type foot. As Harcourt-Smith
explains
"here the first ray adducts to become in line with the functional axis of the foot, and the
foot everts so that the sole is flat on the ground" [Harcourt-Smith and Aiello, 2004, page 406-7].
Kidd concludes that the lateral side of the foot evolved into its modern state before the medial
side, based on the morphology of the OH 8 foot [Kidd, 1999]. However, he uses just one
specimen in his analyses and discounts (or ignores) some of the evidence from the tarsal bones .
The degree of variation cannot possibly be inferred from one specimen although it may help to
support other evidence. The most recent discovery of the seemingly bipedal Sahelanthropus
tchadensis from around 7mya [Brunet et al., 2002] would seem to disprove Kidd's conclusions.
2.8 The extinct hominid foot
Whilst the bipedality of species closer temporally to H. sapiens is well known [Smith and
Aiello, 2004; Aiello and Dean, 2004; Trinkaus, 1983; Lorenzo et al., 1999.] a great majority of this
work is inferred from the post cranial skeleton. For instance, AL 288-1 "Lucy", is suspected to be
bipedal from the morphology of the hip joint and no foot bones are associated with this find
[Jungers, 1988] . Later species such as H. hedelbergensis and H.neanderthalensis have differing
pedal morphology (see 2.6.) their species are clearly bipedal [Aeillo and Dean, 2004; Lorenzo et
al 1999;]. Similarly, although few foot bones are known from H. ergaster its bipedality is inferred

31
from its post-cranial morphology and cranial position. In some cases the body proportions may
be used to infer bipedality although it is imporatnt to consider that there may be a wide range of
variation in these proportions since there may also be significant differences and mosaic of
features in the early hominids due to a range of locomotive style and a mosiac of features as
result. When Day and Napier documented the OH 8 H. habilis foot {fig 9} in 1964 they argued
that the morphology of the bones reflected a fully obligate bipedal stance [Day and Napier,
1964.]. This theory was supported by Richard Leakey and Bernard Wood [Leakey et al., 1964, M.
H. Day and B. A. Wood, 1968]. The OH 8 foot seemed to Day and Napier to have a non-
opposable hallux and contained strong longitudinal arches. However, collaborative research in
the early 1990’s seemed to support the evidence that the OH 8 foot showed arboreal adaptation
[Kidd et al., 1996; Kidd and Oxnard, 2005; Bernhard Zipfel & Robert S. Kidd, 2008] In
particular, the features found show modern adapatations on the bones of the lateral side of the
foot such as the cuboid and a mosaic of features on the medial side; talus and navicular are
primitive in morphology while the cuboid shows modern features. Some researchers have used
this mosiac of morphololgic features to demonstrate an intermediate phase between the mid-
tarsal break seen in P. troglodtyes and the absence of such a feature in modern humans. This
argument has been supported by later finds such as STW 573 'Little Foot' {fig 10}, whose
hindfoot bones show a similar arrangement to the OH 8 material. STW 573 also provided
supporting evidence for the argument for an adducted hallux in OH 8, a view which was
contested until the later material was discovered. Oxnard [1972] concluded that the OH 8 tali
was significantly different from both H. sapiens and ape tali.
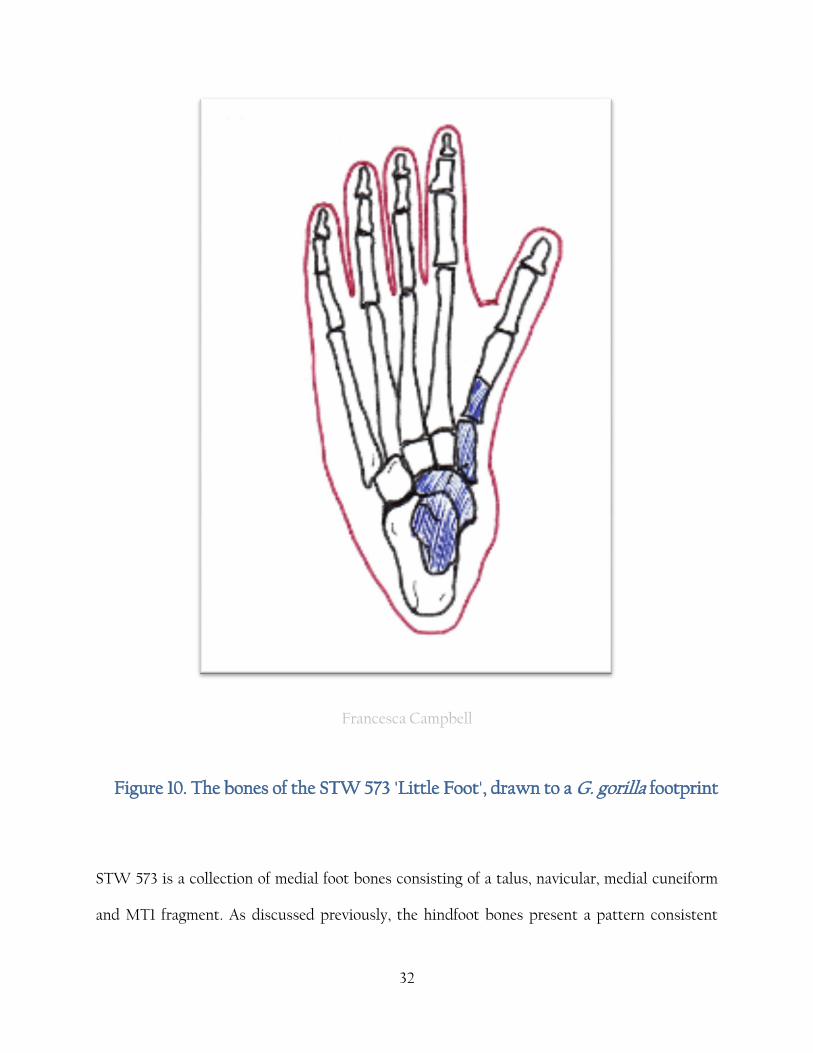
32
Francesca Campbell
Figure 10. The bones of the STW 573 'Little Foot', drawn to a G. gorilla footprint
STW 573 is a collection of medial foot bones consisting of a talus, navicular, medial cuneiform
and MT1 fragment. As discussed previously, the hindfoot bones present a pattern consistent

33
with the OH 8 material, and this find refutes the divergent hallux theory almost conclusively
[Zipfel and Kidd, 2006; Zipfel, DeSilva and Kidd, 2009]. Zipfel et al., [2011] describes the
findings as
“(they) collectively lead to an intregrative model of pedal evolution of caudo-cranial,
disto-proximal changes”.
However, the specimen does seem to have a hallux capable of a great degree of grasping
capability and a mobile mid-tarsal break. The species to which STW 573 is assigned is hotly
contested although it does appear to be conclusively places in the Australopithecus genus, and
conclusive evidence is clouded by the temporally contemporaneous A. afaerensis foot bones
found in Ethiopia [Walter. 1994]. The two models have been resolved into the following
conslusion; A. afarensis has a modern talus, unopposable hallux and strong longitudianal arches
but possess a primitive navicular and an ape like talocrural joint. 1
A. sediba findings [Zipfel et al, 2011] reinforce the semi-arboreal model of pedal morphology
described by Zipfel, Kidd, Oxnard and DeSilva, collectively. Morton’s [ref] 1935 hypothetical
prehuman model of the foot, which shows a divergent hallux alongside a substantial calcaneus,
would appear to be incorrect; however, the reconstruction of the STW 573 specimen by Clarke
and Tobias [1995] would appear to be in direct contrast to this view; indeed, this reconstruction
almost exactly replicates the hypothetical model proposed by Morton [Morton, 1935; Clarke and
Tobias, 1995].
Despite the comparative lack of extinct specimens to work with, Morton's work was based on a
great deal of comparative data and his conclusion is that the prehominid foot would be
somewhere intermediate between that of Gorilla and H. sapiens due to the terrestrial nature of

34
Gorilla, as opposed to the more arboreal Pan. There are features of the Gorilla foot that may
support this argument; a more robust heel (as seen in the OH 8 specimen), MTII to MTV
relatively smaller in size and a less abducted hallux and no longitudinal arch present. Indeed,
many reconstructions of hominid fossils are drawn to a Gorilla footprint/outline. Morton also
outlined how the prehuman foot would have differed from the Gorilla morphology - a relatively
long opposable hallux, an even larger heel and shorter toes than the Gorilla. Harcourt-Smith
[2004] notes the similarity between Morton's reconstruction and Clarke and Tobias' 1995
reconstruction of the STW 573 foot. Morton's work underwent something of a revisionst phase
in the 1990's when the theory that Gorilla and Pan feet were distinctly modified to cope with
increased body weight, especially in the case of male Gorilla which may be up to 60% larger than
the female.
2.9 H. neanderthalensis foot morphology
The most complete pedal elements of H. neanderthalensis are La Ferrassie 1 and the morphology
of the feet and the rest of the post-cranial skeleton are fully adapted to bipedal locomotion
[Trinkaus, 1983]. H. neanderthalensis feel are similar to H. sapiens in the gross morphology, yet
the H. neanderthalensis show a greater degree of robusticity and the musculature attachment
sites are much more strongly developed. [Aeillo and Dean, 2004]. H. neanderthalensis also sho a
unique pattern in that the proximal phalanges are wider than they are high while the opposite is
true for H. sapiens. The proximal phalanx is relatively short compared to total foot length, and is
reflected in the serial homology of the hand. The hallux shows a small degree of adduction but
does not appear to be opposable [Trinkaus, 1983]. In other areas, the H. neanderthalensis foot is

35
indistinguishable from H. sapiens. Highly characteristic, however, is the talus neck in H.
neanderthalensis , which is relatively short, although this appears to be a secondary adaptation
to a high muscle mass and does not appear to have much functional significance. [Aeillo and
Dean. 2004].

36
3. Aims and Objectives
It is the aim of this research to analyse the linear dimensions of the foot of modern humans and
great apes (Gorilla gorilla and Pan troglodytes), and compare those analyses to published data
on fossil hominids. The research attempts to synthesise comparative data by analysing
measurements on non-articulated remains to reflect the nature of the fossil record. Increased
knowledge on the subject of structure of the hominid foot and its relationship to locomotor style
is important in our understanding of human evolution and bipedality.

37
4. Materials and Methods
4.1 Introduction
In humans, the bones of the foot are serial homologies of the bones in the hand and are usually
described in a similar way to the hand bones - tarsal bones (calcaneus, talus, lateral cuneiform,
medial cuneiform and intermediate cuneiform, navicular and the cuboid), the metatarsals
(numbered I to V from medial to lateral) and the phalanges (two for ray I and three each for rays
II to V). Occasionally sesamoid bones may be present and are more common in primates
[Nystrom, 2011]. In common terms the foot is divided into four sections; the heel, the ankle, the
arch of the foot and the toes.
The heel is defined as the prominence at the end of the foot, the most posterior aspect of the
foot. The bony aspect of the calcaneus, the calcaneal tuberosity, forms the posterior projection of
the heel and lies posterior to the articulation of the talocrural region of the ankle.
The ankle is defined as the talocrural region and forms the articulation between the bones of the
foot and the distal end of the tibia and fibula. The ankle is formed of three articulated hinge
joints; the talocrural joint, the subtalar joint and the inferior tibialfibular joint. These joints are
primarily associated with plantarflexion and dorsiflexion of the foot in humans since the
'locked' ankle configuration allows for only a little movement mediolaterally (inversion and
eversion) and in circumvention. Thus the ankle joint is involved in lower limb stability.
Dorsiflexion is the movement of the dorsum of the foot towards the anterior surface of the leg.
This movement is essential in order to have the heel strike the ground first during heel strike
and to clear the foot from the ground during the swing phase of walking. Plantar flexion occurs

38
when the toes are in contact with the substrate and the heel is raised. This movement provides
propulsion and force to lift the limb off the substrate and start it swinging forward during toe-
off phase of walking.
The longitudinal arch of the foot can be broken down into several smaller arches; the medial
arch, the lateral arch and the fundamental arch. The medial arch is formed higher on the foot,
more dorsally, and is mostly responsible for the 'missing' part of the medial side of the foot in a
typical human footprint. The medial arch is made up of the calcaneus, talus, navicular, the
medial, intermediate and lateral cuneiforms and MTI-III. The most dorsal aspect of the
longitudinal arch is at the articular surface of the talus. The bony arch is maintained by strong
plantar calcaneonavicular ligament ('spring' ligament and the tibialis posterior tendon
supporting the arch inferiorly as well as the plantar aponeurosis and the intrinsic and extrinsic
muscles and tendons of the sole of the foot. The major function of the arch is elasticity and to act
as a shock absorber when bearing the entire weight of the bipedal skeleton, especially whilst
running or jumping where a great deal of pressure is placed on the relatively small foot bones.

39
Francesca Campbell
Figure 11. Representation of the direction of the medial and lateral arches of the
foot.

40
The lateral arch also consists of the calcaneus, the cuboid and MT IV and V. It is elevated
dorsally although is not as high as the medial arch. The major function of this arch is strength,
especially in dorsiflexion and plantarflexion of the foot and as such is supported by the tendons
of the extensor muscles as well as the long plantar and the intrinsic muscles such as flexor
digitorum brevis.
The fundamental longitudinal arch is made up from aspects from both the medial and lateral
longitudinal arches and consists of the calcaneus, cuboid, lateral cuneiform and MTIII. This is
described as the fundamental arch since the removal of all the other bones in the foot would
nevertheless leave this arch in place, due in part to the shape and articulations of the bony
structures if the arch.
It is convenient to categorise the bony divisions of the foot (hindfoot, midfoot, and forefoot) but
it is necessary for this research to define the functional divisions of the foot. Whilst the ankle
and heel may be clearly defined as the talus and calcaneus, the author has chosen to include
within this functional division the cuboid and navicular due to the way the bones articulate and
the biomechanics of walking {Fig 7}. It is most important to take into account the plantigrade
walking style of Pan and Gorilla and may reflect primitive condition and walking style of
hominids that may not have had a fully formed longitudinal arch. For this reason, the G. gorilla
dimensions are used as a hypothetical 'primitive' model.

41

42
Francesca Campbell
Figure 12 A, B, C - breakdown of the divisions of the foot and where linear
measurements were made. Displayed on a G. gorilla foot for clarity regarding
measurement of the divergent hallux.
The foot was divided into three sections, labelled A, B and C.
Section A is defined as the hind division of the foot (Hindfoot). It is measured via the following
method.
A = {Aa + Ab}
where Aa = maximum calcaneal length
Ab = length of navicular measured at a 15% from the centre of the 'boat' shape [fig 12 A,
13]
Section B is defined as the mid division of the foot (Midfoot). It is measured via the following
method.
B = {Ba + Bb}
where Ba = maximum length of intermediate cuneiform
Bb = maximum length of MTII

43
Section C is defined as the foremost division of the foot (Forefoot). It is measured via the
following method.
C = {Ca + Cb + Cc}
where Ca = maximum length of proximal phalanx II
Cb = maximum length of middle phalange. Due to the inconsistencies of distinguishing
middle phalanges, the second longest phalanges were chosen for G. gorilla and P. troglodytes
and the third largest for H. sapiens, based on Schultz average relative lengths.
Cc = maximum length of distal phalange. As before, due to the inconsistencies of
distinguishing distal phalanges, the an average of the distal phalanges was taken for G. gorilla
and P. troglodytes and for H. sapiens.
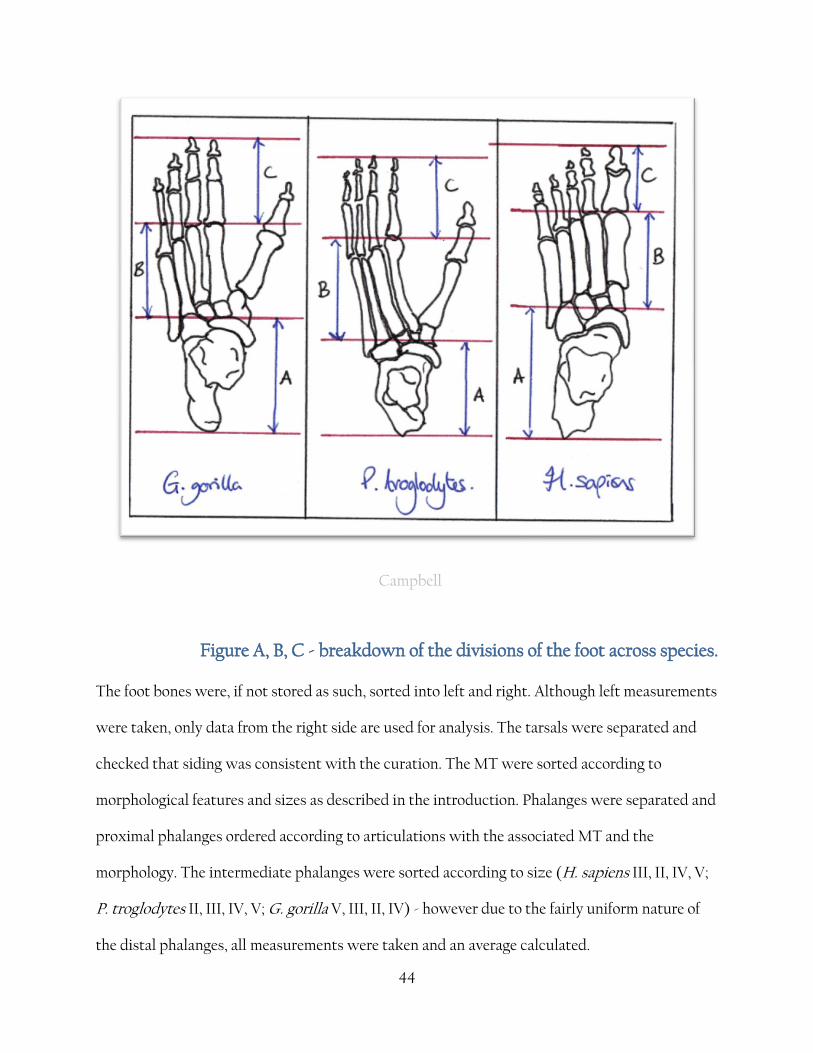
44
Campbell
Figure A, B, C - breakdown of the divisions of the foot across species.
The foot bones were, if not stored as such, sorted into left and right. Although left measurements
were taken, only data from the right side are used for analysis. The tarsals were separated and
checked that siding was consistent with the curation. The MT were sorted according to
morphological features and sizes as described in the introduction. Phalanges were separated and
proximal phalanges ordered according to articulations with the associated MT and the
morphology. The intermediate phalanges were sorted according to size (H. sapiens III, II, IV, V;
P. troglodytes II, III, IV, V; G. gorilla V, III, II, IV) - however due to the fairly uniform nature of
the distal phalanges, all measurements were taken and an average calculated.

45
A pilot study for this project was undertaken in May 2013 to iron out any discrepancies with
using disarticulated remains, to develop a uniform method of collecting data, and to enhance a
method of measurement that was consistent across species. During the pilot study for this
project, five genera of primates were used to collect data; Pongo, Hylobates, Gorilla, Pan and
Homo. These species were chosen to give a cross section of size, sexual dimporhism and
locomotive style. However, when taking the number of measurements required into account,
along with the desire to gain a representative sample (n=30), it was subsequently it was decided
by the author to focus on three major species; Gorilla, Pan and Homo, and compare these species
to extinct hominids. Some data from the pilot study is used during analysis for comparison.
A sample of modern human specimens and modern primate specimens was measured to the
nearest 0.1mm using digital calipers. Individual bones and functional divisions of the foot were
measured and the combined distances compared. Dimensions compared were length of
individual bones and length of functional divisions of the foot {Fig12, 13}.
Material was from the skeletal collections housed at; The University of Sheffield; The University
of Lincoln; The Alfred Denny Museum, Sheffield; The Western Bank Museum, Sheffield; and the
Powell-Cotton Museum, Quex Park, Kent. In addition, cast material from The University of
Sheffield and published data were used for extinct hominid data collection. Measurements from
both feet were taken where possible, but for consistency comparative data was used from the
right foot where possible. Figure 12 summarises the measurements taken and indices will be
calculated from these measurements. Body size was not taken into account although records
exist. Body size may also be estimated using published formulae. Data collection pro-forma may
be found at Appendix A.

46
4.2 Materials
Digital calipers were Wilko make, model D406/07 (resolution 0.001mm, accuracy 0.02mm) and
were thoroughly cleaned and calibrated prior to use using a micrometer and gauge blocks, with
kind permission of the laboratory staff at The University of Lincoln. Both inside and outside
jaws were calibrated although only outside jaws were used to take measurements.
Measurements taken at the Powell-Cotton Museum were all disarticulated specimens. Infant,
juvenile, ‘young adult’ and ‘old adult’ specimens were discounted, only those categorised as
‘adult’ were used.
4.3 Hominid measurments
Where possible, measurements were taken from published data for pedal elements in the fossil
record. Some measurements were taken from cast collection at The University of Sheffield
where disarticulated remains were used where possible to keep with the experimental design.

47
5. Results
Data from 30 specimens (for each species) were collected. Data was shown to be normal as
displayed in the table {1}
G. gorilla P. troglodytes H. sapiens
A -
Hindfoot
0.08 0.07 0.94
B - Midfoot 0.20 0.2 0.13
C -
Forefoot
0.20 0.08 0.06
Table 1 Kolmogorov-Smirnov test significance test results N = 30
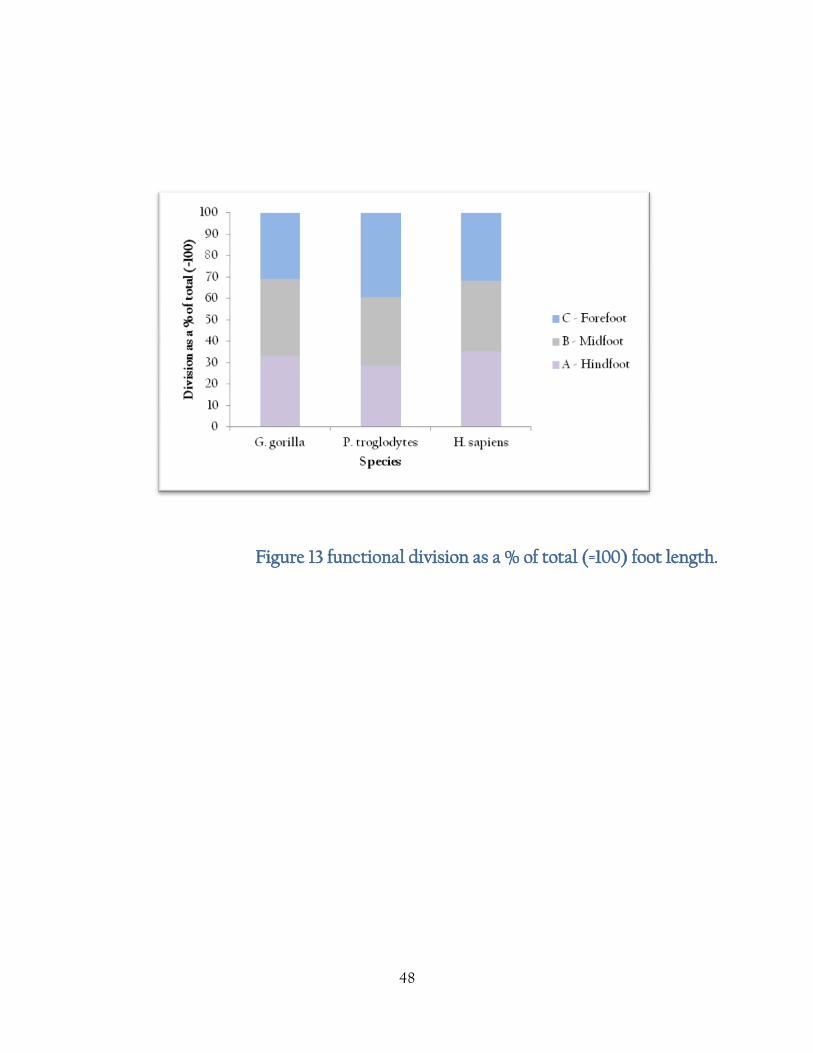
48
Figure 13 functional division as a % of total (=100) foot length.
49
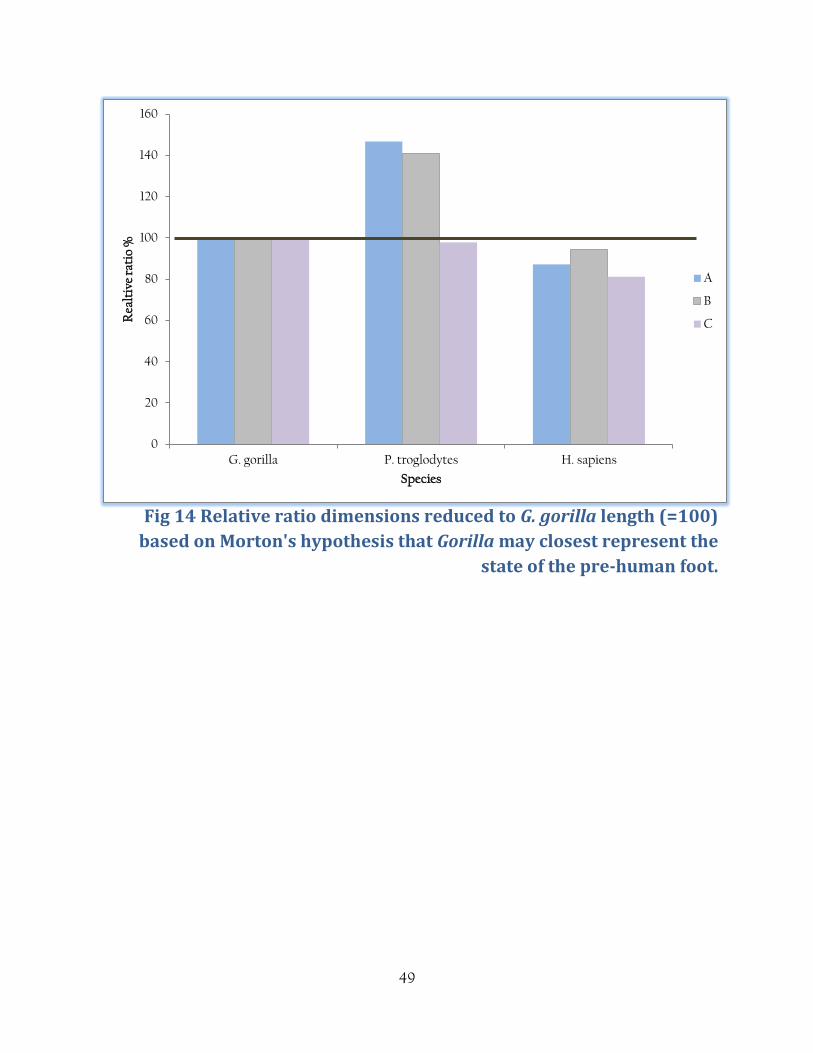
Fig 14 Relative ratio dimensions reduced to G. gorilla length (=100)
based on Morton's hypothesis that Gorilla may closest represent the
state of the pre-human foot.
0
20
40
60
80
100
120
140
160
G. gorilla P. troglodytes H. sapiens
Rea
ltiv
e ra
tio
%
Species
A
B
C
50
51
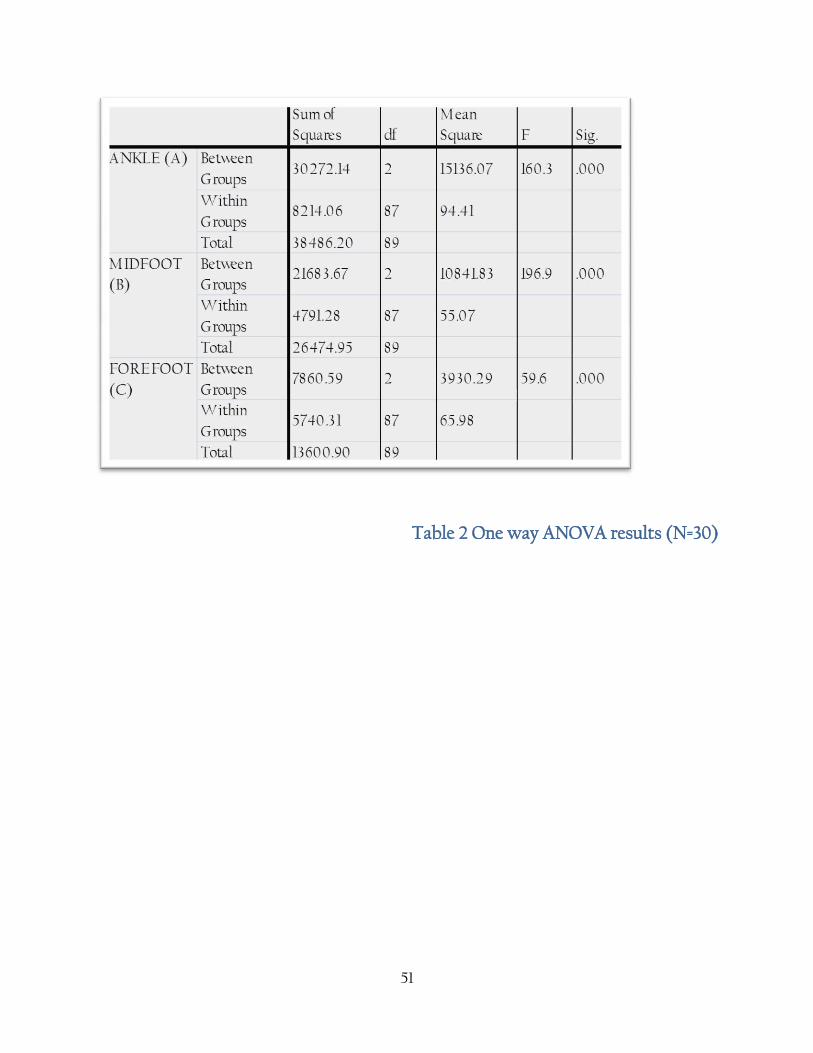
Table 2 One way ANOVA results (N=30)
52
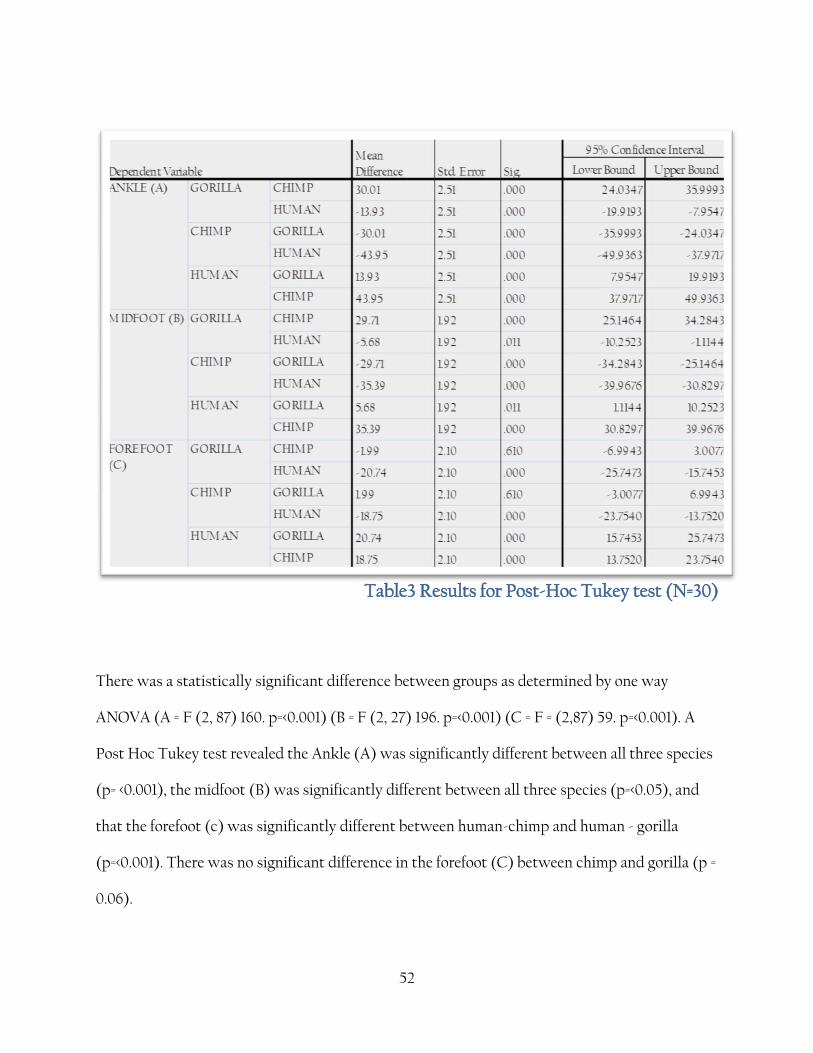
Table3 Results for Post-Hoc Tukey test (N=30)
There was a statistically significant difference between groups as determined by one way
ANOVA (A = F (2, 87) 160. p=<0.001) (B = F (2, 27) 196. p=<0.001) (C = F = (2,87) 59. p=<0.001). A
Post Hoc Tukey test revealed the Ankle (A) was significantly different between all three species
(p= <0.001), the midfoot (B) was significantly different between all three species (p=<0.05), and
that the forefoot (c) was significantly different between human-chimp and human - gorilla
(p=<0.001). There was no significant difference in the forefoot (C) between chimp and gorilla (p =
0.06).
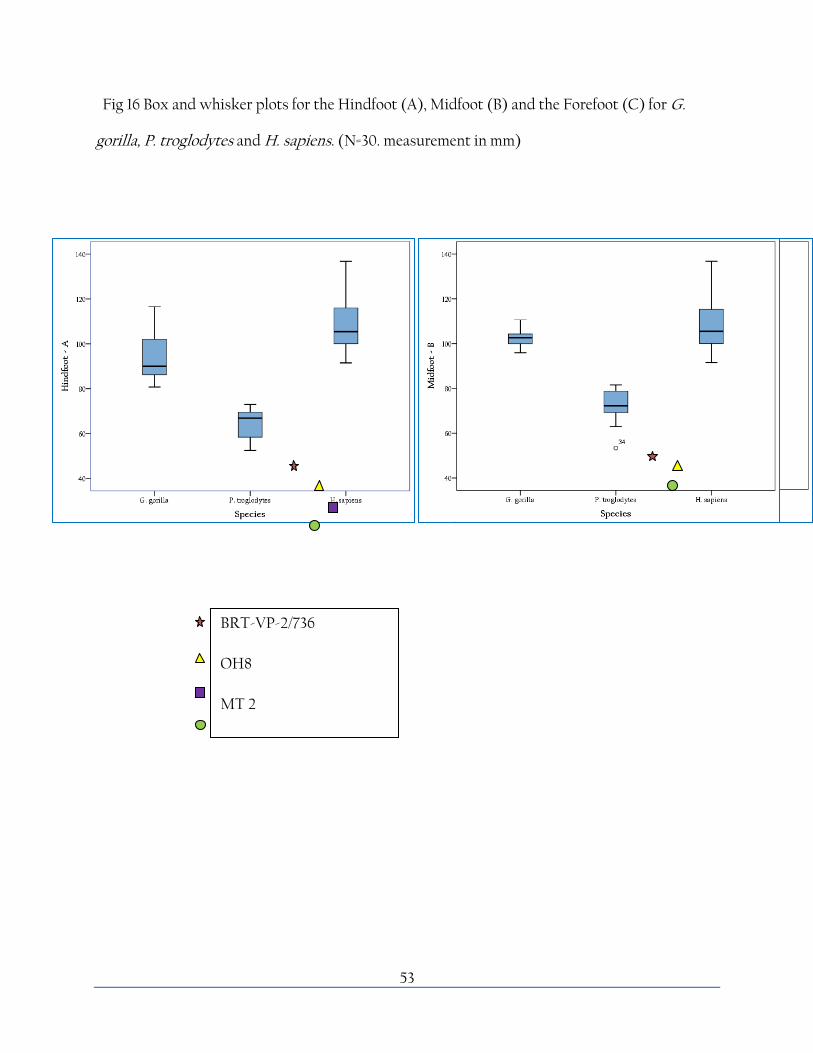
53
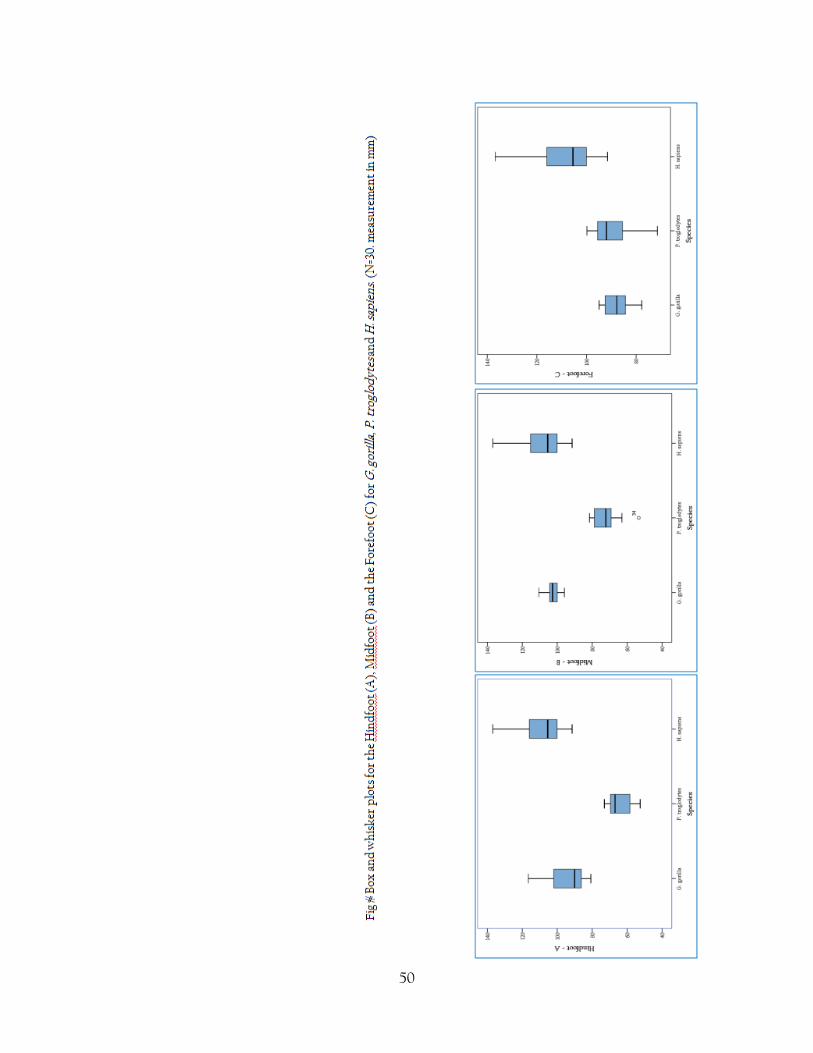
Fig 16 Box and whisker plots for the Hindfoot (A), Midfoot (B) and the Forefoot (C) for G.
gorilla, P. troglodytes and H. sapiens. (N=30. measurement in mm)
BRT-VP-2/736
OH8
MT 2
STW 573

54
6. Discussion
Data from 30 specimens for each species was taken {table 1}. Although this data was shown to be
normal using the Kolmogorov-Smirnov significance test, ideally a larger data set would be
collected and collated and this would have the effect of creating a better data set for PCA to be
more effective.
As expected, the {fig 14} bar charts show that G. gorilla have functional divisions at fairly
uniform lengths. P. troglodytes show relatively larger toe length which may be a response to a
more arboreal lifestyle than Gorilla. H. sapiens show a relatively large hindfoot length due to the
kinematics of heel strike in bipedal walking. Interestingly, the Burtele foot bones show a ratio
more closely aligned with the Pan arrangement than in Gorilla which may reflect the arboreal
nature of this species.
{Fig 14} shows a similar situation when displayed as a % of the Gorilla foot, where Gorilla is
100%. Pan show relative dimensions at between 90% and 140% of the Gorilla measurements
reflecting the longer toes in Gorilla and Pan. In H. sapiens functional divisions are consistently
smaller than in Gorilla, which may be a reflection of the morphological changes in Gorilla due to
increased body weight.

55
Fig 16 shows box and whisker plots for the median, interquartile range and outliers for the three
species studied. These box and whisker charts show the massive range of human variation
compared to the relatively small variation demonstrated by G. gorilla, even when considering
the sexual dimorphism of Gorilla it is very noticeably exactly how much variation there is in a
such a small human sample. The range of human variation is not something that is very well
documented in literature and a massive study would be required to assess this fully, but a larger
data set than 30 specimens would help to clarify this study in a greater context. Specimens from
a more international range to help assess the range of geographical and regional variation may
also shed some light on exactly how much the range of human variation differs. In fact, at the
time of writing, a pre-released paper by Jeremy DeSilva [June 2013] seems to show that around 1
-1 3 modern humans show evidence of a midtarsal break, research which has gone against many
previous comparative studies. In fact, the phenomenon has been document by podiatrists who
did not realise the link between this feature in feet and our evolutionary history. DeSilva plans to
assess this intriguing data further by undertaking an ambitious project involving MRI scans and
3D printing techniques to visualise the walking mechanism of these so called 'floppy feet'. Most
previous studies have shown human feet to be much more rigid with the bones in a 'close
packed' position, with the bones connecting to form the arch and the spring mechanism of the
foot. The connection to the A. sediba fossils cannot be underestimated and these findings may
suggest and even closer link from us to the A. sediba material than anyone previously dare guess.
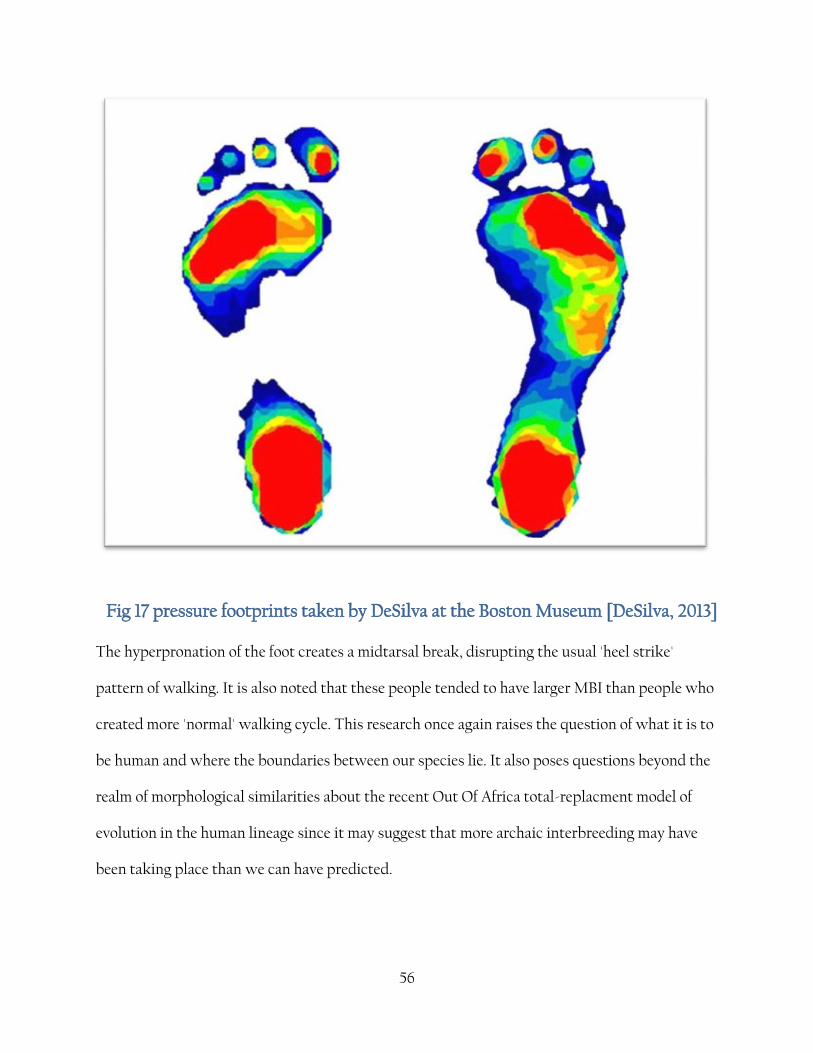
56
Fig 17 pressure footprints taken by DeSilva at the Boston Museum [DeSilva, 2013]
The hyperpronation of the foot creates a midtarsal break, disrupting the usual 'heel strike'
pattern of walking. It is also noted that these people tended to have larger MBI than people who
created more 'normal' walking cycle. This research once again raises the question of what it is to
be human and where the boundaries between our species lie. It also poses questions beyond the
realm of morphological similarities about the recent Out Of Africa total-replacment model of
evolution in the human lineage since it may suggest that more archaic interbreeding may have
been taking place than we can have predicted.

57
Table 3 shows the results of the one way ANOVA, showing that despite quite a large range of
overlap between the species there is none the less a significant difference between the three
species in all three divisions of the foot; however the post-hoc Tukey shows that there was no
significant difference between the chimp and gorilla forefoot; P. trogloytes having a wider range
of variation than Gorilla in this division.
Fig 17 plots selected hominid specimens against the three species surveyed. All of these data are
outside the range for human variation, especially in the hindfoot region, as may be expected due
to the highly developed human foot. However, the hindfoot measurements are also well out of
the range for Gorilla range, refuting the theory that Gorilla may be more similar to the primitive
style. However, these species also vary from each other; since we cannot know the range of
variation exhibited in these species from a handful of fossils, it is impossible to say which of
these differences are significant; however it may be reflective of the range of locomotive styles
and a mosiac of morphological features across contemporaneous species. The midfoot in humans
is highly developed which may account for the wide difference between humans and fossil
homindis. Again, the situation between extinct hominids and Gorilla is repeated, with Gorilla
showing relatively large midfoot measurements but with a narrow range of variation. It is
important to consider that OH 8 is a juvenile specimen and may not represent the fully
developed ratio of linear measurements. Forefoot measurements are difficult to assess in fossil
hominids as the distal phalanges are often lost, and the Burtele foot displays out of the charted
range, well below all three species surveyed.
Since only phalanges are available for A. afarensis, it is difficult to chart this species against the
parameters of this study. However morphology of these bones reflects a mosaic of features. The

58
general sizes and relative dimensions are more ape-like than human which highlights the
importance of supporting the study here with morphological features also present.

59
7. Conclusion
This study supports the conclusions of the work of other researchers that both primates and
humans have separately evolved a unique mosaic of features derived from an unknown collection
of primitive features. Since the primitive condition is as yet unknown, and the hypothetical
model which has been prevalent for the past century has been disproved, it is suggested that
further work must be done to fully understand the wide range of locomotive styles present in
the hominid family tree. Tarsal bones of African Miocene apes show specializations
characteristic of hominoid evolution and provide reasonable precursors for the morphology of
Pan and Gorilla. The OH 8 foot shows many ape-like features, with many close
resemblances to Pan. The relative lengths support this conclusion, and although fairly clearly
from a bipedal primate, it lacked important functional specializations found in the human foot.

60
8. Further Research
This research may be continued to its maximum efficiency via the following methods
Widening the number of specimens and the range of data collected
Extending the study to include a wider range of locomotive styles, including more
terrestrial and habitual bipeds such as Hylobates and Gelada.
Assess the range of human variation across time and geographical location
DeSilva's work on the mid-tarsal break in humans may also inform many aspects of this
study further
Develop better understanding of the long term effect of shoe wearing in the modern
human lineage.

61
9. References
Aeillo, L. and Dean, C. [2006]. An Introduction to Human Evolutionary Anatomy. London:
Elsevier
Ali, F. and Meier, R. [2008]. Positive selection in ASPM is correlated with cerebral cortex
evolution across primates but not with whole-brain size. Molecular Biological Evolution, 25:
2247-2250
Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S. and Suwa, G. [1999]. Australopithecus
garhi: a new species of early hominid from Ethiopia. Science. 284: 629-635
Berger, L. R., De Ruiter, D. J., Churchill, S. E., Schmid, P., Carlson, K. J., Dirks, P. H. G. M. and
Kibii, J. M. [2010]. Australopithecus sediba: A new species of Homo-like australopith from
South Africa. Science. 328: 195-204.
Berillon, G. [1999]. Geometric pattern of the hominoid hallucal tarsometatarsal complex.
Quantifying the degree of hallux abduction in early hominids. Comptes Rendus de l'Académie
des Sciences-Series IIA-Earth and Planetary Science. 328: 627-633
Brunet, M., Guy, F., Pilbeam, D., Mackaye, H. T., Likius, A., Ahounta, D., Beauvilain, A., Blondel,
C., Bocherens, H. and Boisserie, J.-R. [2002]. A new hominid from the Upper Miocene of Chad,
Central Africa. Nature. 418, 145-151

62
Byrnes, G. and Spence, A. J. [2011[. Ecological and Biomechanical Insights into the Evolution of
Gliding in Mammals. Integrative and Comparative Biology. 51: 991-1001
Cartmill, M. [1872]. Arboreal adaptations and the origin of the order Primates. The functional
and evolutionary biology of Primates. 97-122
Cartmill, M. [1974]. Pads and claws in arboreal locomotion. Academic Press: New York
Chaplin, G., Jablonski, N. G. and Cable, N. T. [1994]. Physiology, thermoregulation and
bipedalism. Journal of Human Evolution. 27: 497-510
Clark, W., [1935] Man's Place Among the Primates. Man, 1-6.
Clarke, R. J. and Tobias, P. V., [1995]. Sterkfontein Member 2 foot bones of the oldest South
African hominid. Science. 269: 521-524
Day, M. H. and Wood, B. A. [1968]. Functional affinities of the Olduvai Hominid 8 talus. Man. 3:
440-455
Frey, Carol, et al. [1993]American Orthopaedic Foot and Ankle Society women's shoe
survey." Foot & Ankle International 14 (2): 78-81.
Gebo, D. L. [1992]. Plantigrady and foot adaptation in African apes: implications for hominid
origins. American Journal of Physical Anthropology. 89: 29-58
Gruss, L. T. and Schmitt, D. [2004]. Bipedalism in Homo ergaster: An experimental study of the
effects of tibial proportions on locomotor biomechanics. From Biped to Strider. Springer
Harcourt-Smith, W. E. H. and Aiello, L. C. [1999]. An investigation into the degree of hallux
abduction of the OH 8 foot. Journal of Anatomy. 168: 403-416

63
Harcourt-Smith, W. E. H. and Aiello, L. C. [2004]. Fossils, feet and the evolution of human
bipedal locomotion. Journal of Anatomy. 204: 403-416
Hunt, K. D. [1994]. The evolution of human bipedality: ecology and functional morphology.
Journal of Human Evolution. 26: 183-202
Inman, V. T., Ralston. H. and Todd, F. [1981] Human Walking. Baltimore: Williams and
Wilkins.
Jungers, W. L. [1982]. Lucy's limbs: skeletal allometry and locomotion in Australopithecus
afarensis. Nature. 297: 676-678
Jungers, W.L. [1988]. Lucy's length: Stature reconstruction in Australopithecus afarensis
(A.L.288-1) with implications for other small-bodied hominids. American Journal of Physical
Anthropology . 76: (2): 227–231
Kidd R.S and Oxnard C.E. [2005]. Little Foot and Big Thoughts – a Re-evaluation of the Stw573
Foot from Sterkfontein, South Africa. Journal of Comparative Human Biology. 55:(3) 189-212
Kidd, R. [1999]. Evolution of the rearfoot. A model of adaptation with evidence from the fossil
record. Journal of the American Podiatric Medical Association. 89: 2-17.
Kidd, R. S., O'Higgins, P. and Oxnard, C. E. [1996]. The OH 8 foot: a reappraisal of the functional
morphology of the hindfoot utilizing a multivariate analysis. Journal of Human Evolution. 31:
269-291
Kivell, T. L., Kibii, J. M., Churchill, S. E., Schmid, P. and Berger, L. R. [2011]. Australopithecus
sediba hand demonstrates mosaic evolution of locomotor and manipulative abilities. Science.
333: 1411-1417

64
Köhler, M. and Moyà-Solà, S. [1997]. Ape-like or hominid-like? The positional behavior of
Oreopithecus bambolii reconsidered. Proceedings of the National Academy of Sciences. 94:
11747-11750
Krebs, J. R. and Davies, N. B. (eds.) [1978]. Behavioural Ecology. Oxford: Blackwell Publishing
Latimer, B.; Lovejoy, C. O. [1989]. "The calcaneus of Australopithecus afarensis and its
implications for the evolution of bipedality". American Journal of Physical Anthropology. 78 (3):
369–386
Leakey, R. E. F. [1973]. Evidence for an advanced Plio-Pleistocene hominid from east Rudolf,
Kenya. Nature. 242: 447
Lewis, O. J. [1980a]. The joints of the evolving foot. Part I. The ankle joint. Journal of anatomy.
130: 527
Lewis, O. J. [1980b]. The joints of the evolving foot. Part II. The intrinsic joints. Journal of
Anatomy. 130: 833
Lewis, O. J. [1980c]. The joints of the evolving foot. Part III. The fossil evidence. Journal of
Anatomy. 131: 275
Lewis, O.J. [1989]. Mortuary Functional Morphology of the Evolving Hand and Foot. Oxford:
Clarendon Press
Locke, D., Hillier, L., et al. [2011]. Comparative and demographic analysis of orang-utan genomes.
Nature. 469: 529-533 Contains references #29 - Yu, N. et al. [2003] Low nucleotide diversity
in chimpanzees and bonobos. Genetics. 164: 1511–1518 # 30 Chen, F. C. & Li, W. H. [2001]
Genomic divergences between humans and other hominoids and the effective population size of

65
the common ancestor of humans and chimpanzees. American Journal Human Genetics. 68: 444–
456
Lovejoy, C. O. [1988]. Evolution of human walking. Sci Am. 259: 82-89
McHenry, H. M. [1994]. Behavioral ecological implications of early hominid body size. Journal of
Human Evolution. 27: 77-87
Miller, R. A. [1932]. Evolution of the pectoral girdle and fore limb in the primates. American
Journal of Physical Anthropology. 17: 1-56
Morris AG. [2010]. New hominin fossils from Malapa: The unveiling of Australopithecus sediba .
South African Journal of Science. 106(3/4): 9-10
Morton, D. J. [1935]. The Human Foot, Columbia University Press
Napier, J. R. D. [1958]. The Forelimb Bones of Proconsul africanus. Proceedings of the Fifteenth
International Congress for the Zoological Society of London. 432-434
Richmond, B. G. and Strait, D. S. [2000]. Evidence that humans evolved from a knuckle-walking
ancestor. Nature. 404: 382-385
Richmond, B. G. and Strait, D. S. [2001]. Reply: “Did our ancestors knuckle-walk?” Nature: 410:
326-326
Robinson, J. T. and Robinson, J. T. [1972]. Early hominid posture and locomotion. University of
Chicago Press, Chicago

66
Sarmiento, E. E. and Marcus, L. F. [2000]. The Os Navicular of humans, great apes, OH 8, Hadar,
and Oreopithecus: function, phylogeny, and multivariate analyses. American Museum Novitates.
1-38
Schmitt, D. [2003]. Insights into the evolution of human bipedalism from experimental studies
of humans and other primates. Journal of Experimental Biology. 206: 1437-1448
Schultz, A. H. [1969]. The Life of Primates, Weidenfeld and Nicolson, London
Selassie, Y. H. and Woldegabriel, G. [2009]. Ardipithecus kadabba: late miocene evidence from
the Middle Awash, Ethiopia. University of California Press
Stern JR, J. T. and Susman, R. L. [1983]. The locomotor anatomy of Australopithecus afarensis.
American Journal of Physical Anthropology. 60: 279-317
Taylor, A. B. and Van Schaik, C. P. [2007]. Variation in brain size and ecology in Pongo. Journal
of Human Evolution, 52, 59-71
Vitruvian Man Image, [c.1490], Leonard Da Vinci Wikimedia Commons [online]. Available at: <
http://commons.wikimedia.org/wiki/File:Da_Vinci_Vitruve_Luc_Viatour.jpg> [Accessed 21st June
2012]
Ward, C. V., Kimbel, W. H., and Johanson, D. C., [2011] Complete Fourth Metatarsal and
Arches in the Foot of Australopithecus afarensis. Science. 331 (6018) 750-753
Washburn, S. L. [1967]. Behaviour and the origin of man. Proceedings of the Royal
Anthropological Institute of Great Britain and Ireland, 21-27

67
White, T. D., Suwa, G. and Asfaw, B. [1995]. Australopithecus ramidus, a new species of early
hominid from Aramis, Ethiopia. Nature. 375: 88-88
Haile-Selassie , Y., Saylor, B. Z., Deino, A., Levin, N. E., Alene, M., Latimer, B.M.[2012] A new
hominin foot from Ethiopia shows multiple Pliocene bipedal adaptations. Nature 483, 565–569
Zipfel, B., and Kidd., R. S. [2008] Size and shape of a human foot bone from Klasies River main
site, South Africa. Palaeontology Africa 43: 51–56
Zipfel, B., DeSilva, J. M. and Kidd., R. S. [2009]. Earliest Complete Hominin Fifth Metatarsal—
Implications for the Evolution of the Lateral Column of the Foot. American Journal of Physical
Anthropology 140:532–545
Zipfel, B., DeSilva, J. M. and Kidd., R. S., Carlson, J.K., Churchill, S., Berger, L.R. [2011] The Foot
and Ankle of Australopithecus sediba. Science 333 9

68
10 - Appendices Appendix A - proforma for data collection
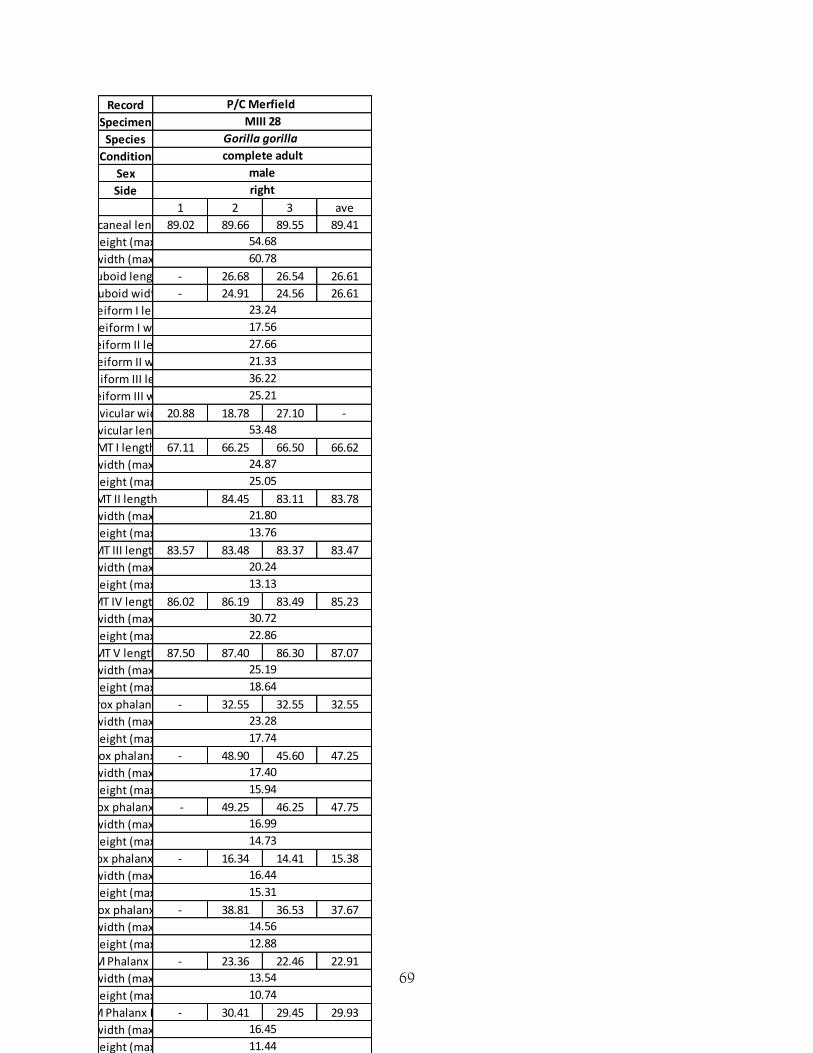
69
Record
Specimen
Species
Condition
Sex
Side
1 2 3 ave
Calcaneal length 89.02 89.66 89.55 89.41
height (max)
width (max)
Cuboid length - 26.68 26.54 26.61
Cuboid width - 24.91 24.56 26.61
Cuneiform I length
Cuneiform I width
Cuneiform II length
Cuneiform II width
Cuneiform III length
Cuneiform III width
Navicular width20.88 18.78 27.10 -
Navicular length
MT I length 67.11 66.25 66.50 66.62
width (max)
height (max)
MT II length 84.45 83.11 83.78
width (max)
height (max)
MT III length 83.57 83.48 83.37 83.47
width (max)
height (max)
MT IV length 86.02 86.19 83.49 85.23
width (max)
height (max)
MT V length 87.50 87.40 86.30 87.07
width (max)
height (max)
Prox phalanx I - 32.55 32.55 32.55
width (max)
height (max)
Prox phalanx II - 48.90 45.60 47.25
width (max)
height (max)
Prox phalanx III - 49.25 46.25 47.75
width (max)
height (max)
Prox phalanx IV - 16.34 14.41 15.38
width (max)
height (max)
Prox phalanx V - 38.81 36.53 37.67
width (max)
height (max)
M Phalanx II - 23.36 22.46 22.91
width (max)
height (max)
M Phalanx III - 30.41 29.45 29.93
width (max)
height (max)
M Phalanx IV - 28.49 27.45 27.97
width (max)
height (max)
M Phalanx V - 21.68 21.45 21.57
width (max)
height (max)
Dist phalanx I - 21.31 21.55 21.43
width (max)
height (max)
Dist phalanx II - 16.90 16.24 16.57
width (max)
height (max)
Dist phalanx III - 18.10 18.50 18.30
width (max)
height (max)
Dist phalanx IV - 16.64 16.45 16.55
width (max)
height (max)
Dist phalanx V - 13.91 13.56 13.74
width (max)
height (max)
P/C Merfield
MIII 28
Gorilla gorilla
complete adult
male
right
36.22
25.21
53.48
24.87
25.05
21.80
54.68
60.78
23.24
17.56
27.66
21.33
18.64
23.28
17.74
17.40
15.94
16.99
13.76
20.24
13.13
30.72
22.86
25.19
10.74
16.45
11.44
13.49
11.39
12.07
14.73
16.44
15.31
14.56
12.88
13.54
8.23
11.04
8.59
8.90
6.83
9.09
17.55
9.18
11.97
9.30
11.76




















