
Consistent protection from pancreatitis in canine pancreas allografts treated with 5-fluorouracil

DNA-PKcs deficiency sensitizes the human hepatoma HepG2 cellsto cisplatin and 5-fluorouracil through suppression of the PI3K/Akt/NF-jB pathway
Yuan Fang • Zongtao Chai • Dansong Wang •
Tiantao Kuang • Wenchuan Wu • Wenhui Lou
Received: 11 July 2014 / Accepted: 17 October 2014
� Springer Science+Business Media New York 2014
Abstract The aim of the present study was to investigate
the effects of DNA-PKcs deficiency on the chemosensi-
tivity of human hepatoma HepG2 cells to cisplatin (CDDP)
and 5-fluorouracil (5-Fu), and to explore the underlying
molecular mechanism. After transfection with DNA-PKcs
siRNA or control siRNA, HepG2 cells were exposed to
combination treatment of CDDP and 5-Fu. The cell via-
bility, DNA damage, cell apoptosis, intracellular reactive
oxygen species and glutathione (GSH) level, expression of
apoptosis related proteins, activity of phosphatidylinositol
3-kinase/protein kinase B (PI3K/AKT) pathway, and
nuclear factor-jB (NF-jB) pathways were assessed. The
combination of CDDP and 5-Fu had a synergistic cytotoxic
effect in HepG2 cells in terms of the cell viability, DNA
damage, apoptosis, and oxidative stress level. DNA-PKcs
siRNA could sensitize the HepG2 cells to the combined
treatment. DNA-PKcs suppression further reduced the Akt
phosphorylation level and Bcl-2 expression in HepG2 cells
exposed to CDDP and 5-Fu, but enhanced the expression of
pro-apoptotic proteins p53 and caspase-3. Moreover,
CDDP could inhibit the transcriptional activity of NF-jB
through degradation of IkB-a, while 5-Fu alone seemed in
some extent increases the NF-jB activity. The combined
treatment with CDDP and 5-Fu resulted in significantly
decrease of the transcriptional activity of NF-jB, which
was further aggravated by DNA-PKcs siRNA treatment. In
conclusion, DNA-PKcs suppression had complementary
effects in combination with CDDP and 5-Fu treatment in
HepG2 cells, which was associated with suppression of
NF-jB signaling pathway cascade, activation of caspase-3
and p53, as well as down-regulation of Bcl-2 and GSH.
Keywords DNA-PKcs � HepG2 cells � Apoptosis �Oxidative stress � NF-jB � PI3K/AKT pathway � Bcl-2 �GSH
Introduction
Hepatocellular carcinoma (HCC) is one of the most com-
mon malignancies worldwide and the third leading cause of
cancer-induced mortality. Hepatocellular carcinoma has an
increasing incidence in recent years, especially in the
developing Asian countries [1]. Clinically, most patients
are not suitable for surgical resection due to the advanced
stage at presentation, extensive disease, or poor liver
function [2]. Therefore, the systemic chemotherapy is a
widely used treatment option for HCC, and current che-
motherapy agents include fluorouracil (5-Fu), cisplatin
(CDDP), doxorubicin, mitomycin C, and so on [3, 4].
However, the effectiveness of chemotherapy using
individual antitumor agent is practically unsatisfactory
because of the low response rate usually below 20 % and
the severe chemoresistance [5, 6]. Recently, several groups
have used combination of CDDP and 5-Fu for the
advanced HCC, indicating favorable results with an overall
20–30 % response rate [7]. It was reported that CDDP and
5-Fu were synergistic in vitro [8], and CDDP could
Yuan Fang and Zongtao Chai have contributed equally to this work.
Y. Fang � D. Wang � T. Kuang � W. Wu � W. Lou (&)
Department of General Surgery, Zhongshan Hospital, Fudan
University, Shanghai 200032, People’s Republic of China
e-mail: [email protected]
Z. Chai
Key Laboratory of Carcinogenesis and Cancer Invasion,
Ministry of Education, Zhongshan Hospital, Liver Cancer
Institute, Fudan University, Shanghai 200032
People’s Republic of China
123
Mol Cell Biochem
DOI 10.1007/s11010-014-2253-6

enhance the cell growth inhibition by 5-Fu through
increasing the intracellular folate pool and activating a Fas-
mediated receptor signaling pathway [9]. Practically, the
combination of 5-Fu and CDDP has become a conventional
chemotherapy strategy for the advanced HCC [10, 11].
However, the antitumor efficiency of the combined treat-
ment still needs to be elevated, and the underlying
molecular mechanisms should be better understood.
The PI3K/Akt pathway is frequently activated in various
malignancies, which consequently activates the prosurvival
pathways and enhances the chemoresistance through NF-
jB activation and p53 inhibition [12, 13]. Arenobufagin
induces apoptosis and autophagy in the HCC cells through
inhibition of PI3K/Akt pathway [14]. The combination of
5-Fu and wogonin could inhibit cyclooxygenase-2
expression and increase the chemosensitivity through reg-
ulation on the PI3K/Akt signaling pathway [15]. CDDP
also exerts the anticancer effects through inhibition of NF-
jB transcriptional activity [16]. The interactions between
PI3K/Akt and NF-jB signaling pathways have become
research focus to explore effective complementary
approaches for cancer therapy [17, 18].
DNA-dependent protein kinase catalytic subunit (DNA-
PKcs) is the key component of DNA-PK and mediates
DNA double-strand break (DSB) repair in response to
DNA damage. Suppression of DNA-PKcs through DNA-
PKcs inhibitor (NU7441) resulted in defects of DNA-PKcs
autophosphorylation at Ser 2056, sensitizing the chronic
lymphocytic leukemia cells to mitoxantrone treatment [19].
The ATM-mediated DNA-PKcs activation is involved in
sustained survival of breast cancer MCF-7 cells under
genotoxic stress induced by CDDP treatment [20]. Panta
et al. also proved that DNA-PKcs activation could induce
the IKK/NF-jB activity and promote cell survival in
murine embryo fibroblasts upon anthracycline doxorubicin
treatment [21]. Thus, we hypothesized that DNA-PKcs
suppression might have an adjuvant therapeutic signifi-
cance in improving the chemotherapy efficiency for HCC
with the existing agents CDDP and 5-Fu.
Materials and methods
Chemicals and reagents
Cisplatin (CDDP) and 5-fluorouracil (5-Fu) were pur-
chased from Selleck Chem (Houston, Texas, USA). Dul-
becco’s modified Eagle’s medium (DMEM), fetal calf
serum (FBS) were obtained from Invitrogen (Paisley, UK).
Cell counting kit-8 (CCK-8) was from Dojindo (Kuma-
moto, Japan). Mammalian protein extraction reagent (M-
PER), nuclear and cytoplasmic extraction reagent kit (NE-
PER), and bicinchoninic acid (BCA) protein assay kit were
from Thermo Fisher Scientific Inc (Waltham, MA, USA).
Nitrocellulose membrane was purchased from Millipore
(Darmstadt, Germany). DCFH-DA (20,70-dichlorofluores-
cein diacetate) was from Sigma (Saint Louis, MO, USA).
GSH assay kit was purchased from Jiancheng (Nanjing,
China). Chemiluminescent EMSA Kit and Biotin-labeled
NF-jB probes were from Beyotime (Wuhan, China). NF-
jB p65, phospho-IKKa/b, and IKKa/b, p-IjBa and IjBa,
p-Akt (Ser473), and Akt, p53, Caspase-3, and Bcl-2 anti-
bodies were from Cell Signaling (Beverly, MA, USA);
DNA-PKcs antibody was from Santa Cruz (Dallas, Texas,
USA). GAPDH antibody was from Multisciences Bio-
technology (Hangzhou, China); anti-Rabbit IgG (H ? L)/
HRP and anti-Mouse IgG (H ? L)/HRP were from Ding-
guo Biotechnology (Beijing, China). All other analytical
grade reagents were obtained from Sigma (Saint Louis,
MO, USA) unless specified otherwise. Absorbance was
recorded using a Multiskan Mk3 plate reader (Thermo
Electron Corporation, Waltham, MA, USA).
Cell culture and treatments
Human hepatoma HepG2 cells were obtained from the
American Type Culture Collection (ATCC, Manassas, VA)
and cultured in DMEM (high glucose concentration) sup-
plemented with 10 % FBS, 0.33 % sodium bicarbonate,
and antibiotics (100 units/ml penicillin and 0.1 mg/ml
streptomycin) at 37 �C in a 95:5 air/CO2 water-saturated
atmosphere. The cells were passaged every 3 days and
used for experiments when in the exponential growth
phase. CDDP and 5-Fu were dissolved in dimethyl sulf-
oxide (DMSO), and controls were treated with the vehicles
only. HepG2 cells were treated with different concentra-
tions of CDDP (0, 1, 2, 5, 10, 20, and 50 mg/L) or/and 5-Fu
(0, 5, 10, 20, 50, 100, and 200 mg/L) for 24 h. All mea-
surements were performed in triplicate.
RNA interference
The DNA-PKcs siRNA (h) (containing a pool of two
target-specific 19–25 nt siRNAs designed to knockdown
the expression of human DNA-PKcs) and a control siR-
NA were purchased from Santa Cruz (Dallas, Texas,
USA). Cells were seeded overnight and then transfected
with siRNA for 48 h using Lipofectamine-2000 (Invitro-
gen, Paisley, UK) according to the manufacturer’s
instruction. The negative control cells (HepG2-Sc) were
transfected with a non-targeting siRNA (control siRNA)
with the same transfection procedure. Cells were then
treated with CDDP and/or 5-Fu for 24 h, and harvested
for following analysis.
Mol Cell Biochem
123

Cell viability
HepG2 cells were seeded in 96-well plates at a density of
3,000 cells per well. Exponentially growing cells were
allowed to attach overnight before being exposed to vari-
ous concentrations of CDDP and 5-Fu, and incubated for
another 24 h. Cell viability was measured using the cell
counting kit-8 (CCK-8) assay according to the manufac-
turer’s instruction. Absorbance was measured with a
spectrophotometric plate reader at 450 nm.
Comet assay
DNA single-strand breaks (SSB) were evaluated with the
alkaline single-cell gel electrophoresis [22]. Briefly, normal
melting point agarose (NMA, 1 %) was prepared on the
frosted microscope slides. Then cells were mixed with low
melting point agarose (LMA, 1 %) and were dripped onto the
pre-coated NMA layers. The third LMA layers were coated
on the cell mixture layer. After lysis for 1 h, the cell-con-
taining slides were soaked into the electrophoresis solution
(300 mM NaOH, 1 mM Na2 EDTA, pH [ 13) for 40 min,
and then subjected to electrophoresis (25 V, *300 mA) for
20 min and subsequent neutralization with Tris–HCl
(400 mM, pH 7.5). Finally, cells were stained with PI (5 mg/
L), and the tail moment (% Tail DNA) was evaluated under a
fluorescence microscope (Olympus BX-51, Japan).
Apoptosis analysis
The apoptosis of HepG2 cells were measured by Hoechst
33342 staining and flow cytometry using Annexin V-FITC/
PI assay according to the operating instructions. After
treatment, approximate 106 cells were harvested and washed
twice with cold PBS solution. Then cells were stained with
1 mg/L of Hoechst 33342 for 15 min at 37 �C. Cells with
apoptotic nuclei (i.e., condensed or fragmented) were
counted in comparison with total population (n = 300 cells).
For the flow cytometry analysis, cells were suspended with
400 lL binding buffer, incubated with 5 lL of Annexin
V-FITC at 37 �C for 15 min and 10 lL of PI at 37 �C for 5
min in the dark. The stained cells were immediately analyzed
by flow cytometry (Becton–Dickinson, San Jose, USA).
Determination of reactive oxygen species (ROS)
and glutathione (GSH)
The intracellular ROS level was assessed by the fluorescent
probe DCFH-DA according to the manufacturer’s instruc-
tion. The fluorescence intensity was observed under a
fluorescence microscope (Olympus BX-51, Tokyo, Japan)
and analyzed by Image-pro plus 6.0 software. GSH activity
was detected by the dithio-bis-nitrobenzoic acid (DTNB)
assay using colorimetry at 420 nm. Absorbance was
recorded using a Multiskan Mk3 plate reader. All experi-
ments were carried out at least three times.
Western blotting
HepG2 cells after various treatments were lysed to obtain dif-
ferent cell extracts using the NE-PER kit or M-PER kit. Protein
concentrations were determined by the BCA protein assay.
Equal amounts of protein samples were separated by sodium-
dodecylsulfate-polyacrylamide gel electrophoresis, and trans-
ferred onto nitrocellulose membranes using the Trans-Blot SD
Electrophoretic Transfer Cell (Bio-Rad Hercules, CA, USA).
The membranes were then blocked and incubated with primary
antibodies and corresponding second antibodies according to
the manual. Finally, blots were visualized through enhanced
chemiluminescence and quantized using the Chemi-Imager
digital imaging system (Alpha Innotech, San Leandro, USA).
GAPDH was used as the internal inference, and each experi-
ment was repeated at least three times.
Electrophoretic mobility-shift assay (EMSA)
Nuclear extracts were prepared with the NE-PER Kit, and
the DNA–protein binding reactions were conducted using
the chemiluminescent EMSA Kit according to the manu-
facturer’s instructions. NF-jB consensus oligo sequences
were as follows: 50-AGT TGA GGG GAC TTT CCC AGG
C-30 and 30-TCA ACT CCC CTG AAA GGG TCC G-50.Sample response system (a total volume of 10 lL) inclu-
ded 5 lL of nuclease-free water, 2 lL of EMSA/Gel-Shift
binding buffer (59), 2 lL of purified nuclear protein, and
0.5 lL of biotin-labeled NF-jB probe. The reaction mix-
tures were loaded on 4 % non-denaturing polyacrylamide
gels, which were then blotted onto nylon membranes and
subjected to chemiluminescence detection.
Statistical analyses
All experiments were performed in triplicate, and data
were expressed as mean ± SD. All data were subjected to
Analysis of Variance (ANOVA) and Tukey’s post hoc test
for comparisons. A p value of \0.05 was considered to
indicate statistical significance.
Results
DNA-PKcs suppression inhibited cell survival
of HepG2 exposed to CDDP and 5-Fu
HepG2 cells were exposed to different concentrations of
CDDP or/and 5-Fu, and CCK-8 assay was conducted to
Mol Cell Biochem
123

Fig. 1 DNA-PKcs suppression sensitizes HepG2 cells to CDDP and
5-Fu. HepG2 cells were treated with different concentrations of
CDDP and 5-Fu for 24 h, and the cell survival rates were measured
with CCK-8 assay. a Cells were treated with various concentrations of
CDDP (0, 1, 2, 5, 10, 20, and 50 mg/L). b Cells were treated with
different concentrations of 5-Fu (0, 5, 10, 20, 50, 100, and 200 mg/L).
c HepG2 cells were transfected with DNA-PKcs siRNA, and then
exposed to 2 mg/L CDDP or/and 10 mg/L 5-Fu. Cont HepG2 cells
treated with vehicles only. Sc HepG2 cells transfected with the
negative control siRNA. Si HepG2 cells transfected with the DNA-
PKcs siRNA. C-2 HepG2 cells were exposed to 2 mg/L CDDP for
24 h. F-10 HepG2 cells were exposed to 10 mg/L 5-Fu for 24 h.
C ? Sc Cells transfected with the control siRNA were treated with
2 mg/L CDDP. C ? Si Cells transfected with the DNA-PKcs siRNA
were treated with 2 mg/L CDDP. F ? Sc Cells transfected with the
control siRNA were treated with 10 mg/L 5-Fu. F ? Si Cells
transfected with the DNA-PKcs siRNA were treated with 10 mg/L
5-Fu. C ? F HepG2 cells were treated with 2 mg/L CDDP and
10 mg/L 5-Fu for 24 h. C ? F?Sc HepG2 cells were transfected with
the control siRNA, and then treated with 2 mg/L CDDP and 10 mg/L
5-Fu for 24 h. C ? F?Si HepG2 cells were transfected with the
DNA-PKcs siRNA, and then treated with 2 mg/L CDDP and 10 mg/L
5-Fu for 24 h. *p \ 0.05, **p \ 0.01 versus Cont group; #p \ 0.05
versus C-2 or F-10 group; ^p \ 0.05 versus C ? F?Sc group
Mol Cell Biochem
123

determine the cell viability. As shown in Fig. 1a and b,
CDDP and 5-Fu induced decrease of cell viability in a
concentration-dependent manner. The cell viability rates
were approximately 78.6 and 80.3 % under treatment of
2 mg/L CDDP and 10 mg/L 5-Fu, respectively. Compared
with the individual treatment group, the combined treat-
ment group with CDDP and 5-Fu showed a more severe
inhibition on cell viability with an average survival rate
about 65.2 %. In addition, transfection of DNA-PKcs
siRNA or control siRNA alone had no obvious inhibiting
effect on the cell viability. However, DNA-PKcs siRNA
(not control siRNA) resulted in further decrease of cell
survival (with an elevated efficiency by 20–30 %) induced
by CDDP and/or 5-Fu treatment, as shown in the C ? Si,
F ? Si, and C ? F?Si groups (Fig. 1c). In the following
experiments, we mainly focused on the effects of DNA-
PKcs siRNA in the combined treatment of CDDP and 5-Fu
to HepG2 cells.
DNA-PKcs suppression increased CDDP and 5-Fu
induced apoptosis and DNA damage
Hoechst 33342 staining and Annexin V-FITC/PI double
staining assay were used to measure the cell apoptosis
ratio. As shown in Fig. 2, the cell apoptosis ratio of 2 mg/L
CDDP and 10 mg/L 5-Fu groups were significantly higher
than that of control groups (p \ 0.01). The combination of
CDDP and 5-Fu treatment had a synergistic effect on the
cell apoptosis (p \ 0.01), and DNA-PKcs siRNA further
increased the apoptosis ratio (p \ 0.01). Similarly, the
results of comet assay showed that CDDP and 5-Fu could
induce DNA damage, combined treatment with CDDP and
Fig. 2 DNA-PKcs suppression increased the cell apoptosis induced
by CDDP and 5-Fu. a Hoechst 33342 staining was used to measure
the cell apoptosis ratio. Cells with apoptotic nuclei (i.e., condensed or
fragmented) were counted in comparison with total population
(n = 300 cells). b Annexin V-FITC/PI double staining assay was
used to measure the cell apoptosis ratio. Data were expressed as
Mean ± SD from three independent flow cytometry experiments.
c The represented figures of annexin V-FITC/PI double-staining
assay. Cont HepG2 cells treated with vehicles only. Sc HepG2 cells
transfected with the negative control siRNA. Si HepG2 cells
transfected with the DNA-PKcs siRNA. C-2 HepG2 cells were
exposed to 2 mg/L CDDP for 24 h. F-10 HepG2 cells were exposed
to 10 mg/L 5-Fu for 24 h. **p \ 0.01 versus Cont group; ##p \ 0.01
versus C-2 or F-10 group; ^p \ 0.05, ^^p \ 0.01 versus C ? F?Sc
group
Mol Cell Biochem
123
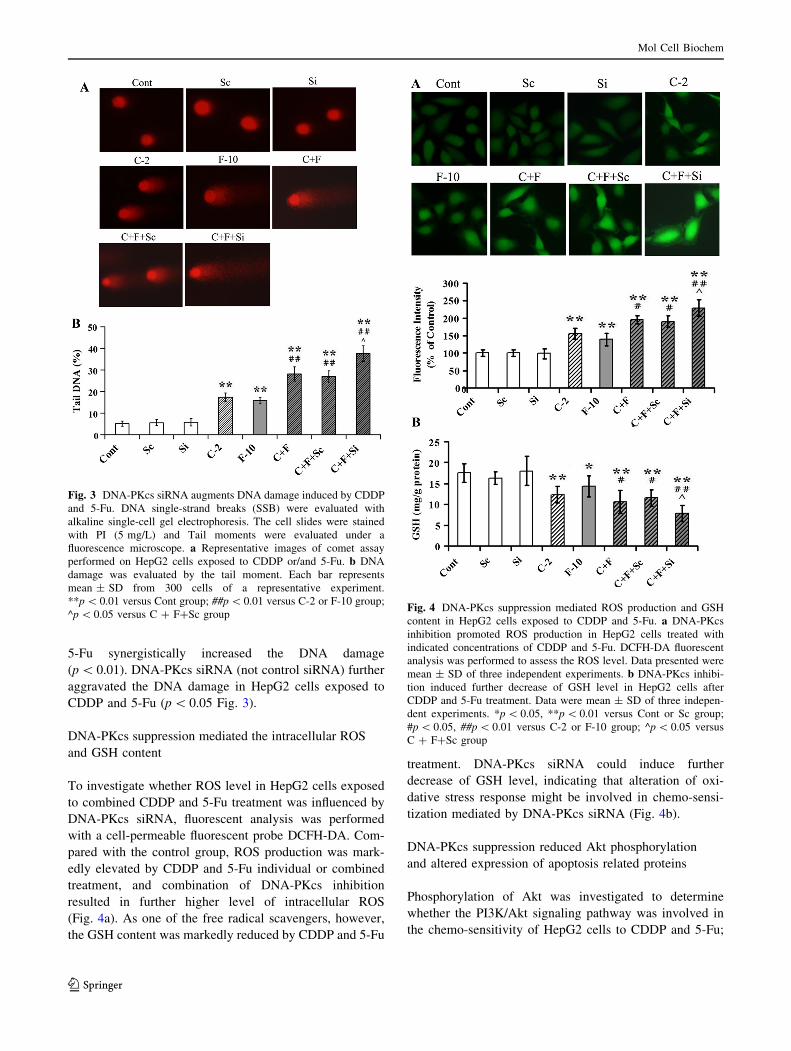
5-Fu synergistically increased the DNA damage
(p \ 0.01). DNA-PKcs siRNA (not control siRNA) further
aggravated the DNA damage in HepG2 cells exposed to
CDDP and 5-Fu (p \ 0.05 Fig. 3).
DNA-PKcs suppression mediated the intracellular ROS
and GSH content
To investigate whether ROS level in HepG2 cells exposed
to combined CDDP and 5-Fu treatment was influenced by
DNA-PKcs siRNA, fluorescent analysis was performed
with a cell-permeable fluorescent probe DCFH-DA. Com-
pared with the control group, ROS production was mark-
edly elevated by CDDP and 5-Fu individual or combined
treatment, and combination of DNA-PKcs inhibition
resulted in further higher level of intracellular ROS
(Fig. 4a). As one of the free radical scavengers, however,
the GSH content was markedly reduced by CDDP and 5-Fu
treatment. DNA-PKcs siRNA could induce further
decrease of GSH level, indicating that alteration of oxi-
dative stress response might be involved in chemo-sensi-
tization mediated by DNA-PKcs siRNA (Fig. 4b).
DNA-PKcs suppression reduced Akt phosphorylation
and altered expression of apoptosis related proteins
Phosphorylation of Akt was investigated to determine
whether the PI3K/Akt signaling pathway was involved in
the chemo-sensitivity of HepG2 cells to CDDP and 5-Fu;
Fig. 3 DNA-PKcs siRNA augments DNA damage induced by CDDP
and 5-Fu. DNA single-strand breaks (SSB) were evaluated with
alkaline single-cell gel electrophoresis. The cell slides were stained
with PI (5 mg/L) and Tail moments were evaluated under a
fluorescence microscope. a Representative images of comet assay
performed on HepG2 cells exposed to CDDP or/and 5-Fu. b DNA
damage was evaluated by the tail moment. Each bar represents
mean ± SD from 300 cells of a representative experiment.
**p \ 0.01 versus Cont group; ##p \ 0.01 versus C-2 or F-10 group;
^p \ 0.05 versus C ? F?Sc groupFig. 4 DNA-PKcs suppression mediated ROS production and GSH
content in HepG2 cells exposed to CDDP and 5-Fu. a DNA-PKcs
inhibition promoted ROS production in HepG2 cells treated with
indicated concentrations of CDDP and 5-Fu. DCFH-DA fluorescent
analysis was performed to assess the ROS level. Data presented were
mean ± SD of three independent experiments. b DNA-PKcs inhibi-
tion induced further decrease of GSH level in HepG2 cells after
CDDP and 5-Fu treatment. Data were mean ± SD of three indepen-
dent experiments. *p \ 0.05, **p \ 0.01 versus Cont or Sc group;
#p \ 0.05, ##p \ 0.01 versus C-2 or F-10 group; ^p \ 0.05 versus
C ? F?Sc group
Mol Cell Biochem
123

Western blot analysis showed that CDDP and 5-Fu indi-
vidual or combined treatment reduced the phosphorylation
level of Akt in HepG2 cells after 24 h exposure, which was
most evident in the combined treatment group under
depression of DNA-PKcs, although no obvious change was
found in the total amount of Akt protein. At the same time,
the expression of anti-apoptotic protein Bcl-2 was signifi-
cantly down-regulated, while the pro-apoptotic factors p53
and caspase-3 were obviously elevated in the combined
treatment group (Fig. 5).
DNA-PKcs suppression modulated the activity of NF-
jB signaling pathway
As a primary transcription factor, NF-jB activation could
lead to fairly rapid changes in the cell growth-related gene
expressions when respond to harmful cellular stimuli [23].
In the present study, we measured the transcriptional
activity of NF-jB using the chemiluminescent EMSA
assay. In HepG2 cells, CDDP individual treatment inhib-
ited the transcriptional activity of NF-jB, while 5-Fu
induced slightly increase of NF-jB activity. In the com-
bined treatment group, the transcriptional activity of NF-
jB obviously reduced, which was consistent with the
protein expression of NF-jB in the nuclei (Fig. 6b).
Moreover, DNA-PKcs suppression further depressed the
NF-jB transcriptional activity in the combined treatment
group (Fig. 6a).
The phosphorylation of IjB kinases (IKK-a/b) and IjB-
a is prerequisite for the activation of NF-jB [23]. As
shown in Fig. 6b, the phosphorylated IKK-a/b and IjB-aobviously reduced in the individual CDDP-treated HepG2
group, but not in the individual 5-Fu-treated group. The
combined treatment with CDDP and 5-Fu significantly
reduced the phosphorylation level of IKK-a/b and IjB-a,
which was lower than both these two individual treatment
groups. DNA-PKcs suppression further reduced the phos-
phorylation of IKK-a/b and IjB-a in the HepG2 cells
exposed to CDDP and 5-Fu treatment.
Discussion
Numerous experimental and clinical studies have revealed
that combination of CDDP and 5-Fu has a pronounced
Fig. 5 Influence of DNA-PKcs suppression on Akt phosphorylation
and expression of apoptosis related proteins. HepG2 cells were
exposed to 2 mg/L CDDP or/and 10 mg/L 5-Fu for 24 h. Protein
expressions were analyzed by Western blot. GAPDH was used as the
internal reference. Cont HepG2 cells were treated with vehicles only.
Sc HepG2 cells were transfected with the negative control siRNA. Si
HepG2 cells were transfected with the DNA-PKcs siRNA. C-2
HepG2 cells were treated with 2 mg/L of CDDP for 24 h. F-10
HepG2 cells were treated with 10 mg/L of 5-Fu for 24 h
Fig. 6 DNA-PKcs suppression reduced the activity of NF-jB
signaling pathways in HepG2 cells exposed to CDDP and 5-Fu.
HepG2 cells were exposed to 2 mg/L of CDDP or/and 10 mg/L of
5-Fu for 24 h. a NF-jB transcriptional activity was assayed by EMSA
assay. b Proteins expression was analyzed by Western blot. GAPDH
was used as the internal reference. Presented images were from the
representative results of three independent experiments. Cont HepG2
cells were treated with vehicles only. Sc HepG2 cells were transfected
with the negative control siRNA. Si HepG2 cells were transfected
with the DNA-PKcs siRNA. C-2 HepG2 cells were treated with 2 mg/
L of CDDP for 24 h. F-10 HepG2 cells were treated with 10 mg/L of
5-Fu for 24 h
Mol Cell Biochem
123

antitumor activity in various types of human tumors, but
the academia has not form consensus about the synergistic
mechanism at present [7–9]. In this study, HepG2 cells
were transfected with DNA-PKcs siRNA or negative con-
trol siRNA, and then exposed to different concentrations of
CDDP and 5-Fu. The cell viability, apoptosis, oxidative
stress response, and DNA damage were measured and
compared, indicating that CDDP and 5-Fu individual
treatment had toxic effects in the HepG2 cells, and com-
binatory treatment with CDDP and 5-Fu had a synergistic
effect. Compared with the individual treatment group, the
combined treatment group showed even lower cell survival
rate, elevated apoptosis ratio and DNA damage, increased
ROS production, and reduced intracellular GSH level.
Combination of DNA-PKcs siRNA obviously augmented
the cytotoxicity of CDDP and 5-Fu in HepG2 cells.
Reactive oxygen species originated from the aerobic
cells are well recognized to be involved in various cell
processes such as cell proliferation, apoptosis, and DNA
damage [24, 25]. We described in the present study that
CDDP and 5-Fu treatment resulted in a higher level of ROS
production and DNA damage in HepG2 cells as compared
with the CDDP/5-Fu individual treatment group, which
was further aggravated by combination of DNA-PKcs
siRNA. Meanwhile, the antioxidant glutathione reduced
most significantly in HepG2 cells exposed to the combined
treatment group. Glutathione plays a vital role in protecting
cells from oxidative stress and controlling apoptotic gene
expression, membrane transportation, and chemotherapy
resistance [26–28]. The Bcl-2 expression is also closely
related to GSH in that Bcl-2 modulates the GSH production
and nucleus relocalization under oxidative stress [29]. In
the present study, DNA-PKcs suppression in HepG2 cells
exposed to CDDP and 5-Fu treatment resulted in Bcl-2 and
GSH reduction, accompanying by increased expression of
p53 and caspase-3, indicating activation of the mitochon-
drial apoptotic pathways.
Overexpression of DNA-PKcs is frequently found in
various cancers, which is closely associated with the tumor
cell growth, poor prognosis, and clinical therapeutic out-
come [30–32]. Depletion of DNA-PKcs through shRNA
transfection resulted in inhibition of cell proliferation and
increased apoptosis in human cervical cancer HeLa cell
line exposed to cisplatin [33]. Moreover, besides the
capability of the non-homologous-end-joining repair for
DNA double strands breaks (DSBs), DNA-dependent pro-
tein kinase (DNA-PK) is involved in processing the oxi-
datively induced non-DSB clustered DNA lesions, which is
important for the therapeutic efficiency of cancer treat-
ments based on the induction of oxidative stress [34].
Mutant forms of DNA-PKcs at serine/threonine residues in
the 2,609–2,647 cluster or specific inhibitor KU57788
could completely block or markedly reduce the efficiency
of end joining [35]. In the present study, DNA-PKcs defi-
ciency through small interfering RNA transfection signifi-
cantly sensitized the HepG2 cells to CDDP and 5-Fu in
terms of the cell viability, apoptosis, oxidative stress, and
DNA damage.
DNA-PKcs is a member of the phosphatidylinositol
3-kinase (PI3K) family, and the PI3K/AKT signaling
pathway is implicated in essential cellular functions such
as survival, proliferation, migration, and differentiation.
Wang et al. confirmed that suppression of the PI3K/AKT
signaling can effectively sensitize osteosarcoma cells to
cisplatin [36]. Our results also showed that combination of
CDDP and 5-Fu inhibited the activity of PI3K/Akt path-
way; DNA-PKcs siRNA could effectively suppress the
expression of DNA-PKcs and further reduced the activity
of PI3K/Akt signaling, which may be associated with
sensitization of HepG2 cells to combination treatment of
CDDP and 5-Fu.
The PI3K/Akt/NF-jB signaling pathway is known to be
involved in stimulation of tumor cell survival, invasive
behavior, and chemosensitivity in various malignancies
[37]. Two serine threonine IjB kinases (IKK-a/-b) are
responsible for phosphorylation and degradation of IjB
molecules, which may result in release of NF-jB from its
binding with IjB. Free NF-jB is then translocated into the
nuclei and exerts transcriptional activity to regulate phys-
iological processes such as cell proliferation and cell death
[23]. CDDP is a potent inhibitor to prevent NF-jB acti-
vation, while 5-Fu could induce NF-jB activation in many
cancers. Hartojo et al. reported that the combination of a
specific NF-jB inhibitor curcumin with 5-Fu and CDDP
could increase apoptosis and enhance both 5-FU- and
CDDP-mediated chemosensitivity in esophageal adeno-
carcinoma [38]. Interferon-gamma could also sensitize
hepatitis B virus (HBV)-expressing HCC cells to 5-Fu
through inhibition of HBV-mediated NF-jB activation
[39]. The Western blotting results of the present study
showed that CDDP led to reduced nuclear translocation of
NF-jB/p65 protein, NF-jB transcriptional activity, and
phosphorylation levels of IKK-a/-b. However, 5-Fu medi-
ated an opposite protein expression profile of NF-jB sig-
naling pathway, as well as the NF-jB transcriptional
activity. Moreover, the activation of NF-jB from 5-Fu
treatment was inversed by the concomitant treatment of
CDDP and further suppressed by combination of DNA-
PKcs siRNA, which may make HepG2 cells more vulner-
able to chemotherapeutic agents.
Taken together, combination with CDDP and 5-Fu had
synergistic effects on HepG2 cells in inducing proliferation
inhibition, ROS production, DNA damage, and cell apop-
tosis. DNA-PKcs suppression could augment the cytotox-
icity induced by CDDP and 5-Fu, which was associated
with blocking of PI3K/Akt/NF-jB signaling pathway,
Mol Cell Biochem
123

increased expression of caspase-3 and p53, and down-
regulation of Bcl-2 and GSH.
Acknowledgments The study was supported by Special Research
Fund for public welfare from Ministry of Health of China
(201202007)
Conflict of interest The authors declare that there are no conflicts
of interest.
References
1. Mendizabal M, Reddy KR (2009) Current management of
hepatocellular carcinoma. Med Clin North Am 93(4):885–900
2. Llovet JM, Burroughs A, Bruix J (2003) Hepatocellular carci-
noma. Lancet 362:1907–1917
3. Lin DY, Lin SM, Liaw YF (1997) Non-surgical treatment of
hepatocellular carcinoma. J Gastroenterol Hepatol 12(9–10):
S319–S328
4. Ueno H, Okada S, Okusaka T, Ikeda M, Kuriyama H (2002)
Phase I and pharmacokinetic study of 5-fluorouracil administered
by 5-day continuous infusion in patients with hepatocellular
carcinoma. Cancer Chemother Pharmacol 49:155–160
5. Stehlin JS Jr, de Ipolyi PD, Greeff PJ, McGaff CJ Jr, Davis BR,
McNary L (1988) Treatment of cancer of the liver. Twenty years’
experience with infusion and resection in 414 patients. Ann Surg
208(1):23–35
6. Falkson G, Ryan LM, Johnson LA, Simson IW, Coetzer BJ,
Carbone PP, Creech RH, Schutt AJ (1987) A random phase II
study of mitoxantrone and cisplatin in patients with hepatocel-
lular carcinoma. An ECOG study. Cancer 60(9):2141–2145
7. Ellis PA, Norman A, Hill A, O’Brien ME, Nicolson M, Hickish
T, Cunningham D (1995) Epirubicin, cisplatin and infusional
5-fluorouracil (5-FU) (ECF) in hepatobiliary tumours. Eur J
Cancer 31A(10):1594–1598
8. Kogure T, Ueno Y, Iwasaki T, Shimosegawa T (2004) The effi-
cacy of the combination therapy of 5-fluorouracil, cisplatin and
leucovorin for hepatocellular carcinoma and its predictable fac-
tors. Cancer Chemother Pharmacol 53:296–304
9. Kim R, Tanabe K, Inoue H, Toge T (2002) Mechanism(s) of
antitumor action in protracted infusion of low dose 5-fluorouracil
and cisplatin in gastric carcinoma. Int J Oncol 20(3):549–555
10. Woo HY, Bae SH, Park JY, Han KH, Chun HJ, Choi BG, Im HU,
Choi JY, Yoon SK, Cheong JY (2010) A randomized compara-
tive study of high-dose and low-dose hepatic arterial infusion
chemotherapy for intractable, advanced hepatocellular carci-
noma. Cancer Chemother Pharmacol 65(2):373–382
11. Yoshikawa M, Ono N, Yodono H, Ichida T, Nakamura H (2008)
Phase II study of hepatic arterial infusion of a fine-powder for-
mulation of cisplatin for advanced hepatocellular carcinoma.
Hepatol Res 38(5):474–483
12. Grandage VL, Gale RE, Linch DC, Khwaja A (2005) PI3-kinase/
Akt is constitutively active in primary acute myeloid leukaemia
cells andregulates survival and chemoresistance via NF-kappaB,
Mapkinase and p53 pathways. Leukemia 19(4):586–594
13. Pellegrino R, Calvisi DF, Neumann O, Kolluru V, Wesely J,
Chen X, Wang C, Wuestefeld T, Ladu S, Elgohary N, Bermejo
JL, Radlwimmer B, Zornig M, Zender L, Dombrowski F, Evert
M, Schirmacher P, Longerich T (2014) EEF1A2 inactivates p53
via PI3K/AKT/mTOR-dependent stabilization of MDM4 in
hepatocellular carcinoma. Hepatology 59(5):1886–1899
14. Zhang DM, Liu JS, Deng LJ, Chen MF, Yiu A, Cao HH, Tian HY,
Fung KP, Kurihara H, Pan JX (2013) Arenobufagin, a natural
bufadienolide from toad venom, induces apoptosis and autophagy
in human hepatocellular carcinoma cells through inhibition of
PI3K/Akt/mTOR pathway. Carcinogenesis 34(6):1331–1342
15. Zhao L, Sha YY, Zhao Q, Yao J, Zhu BB, Lu ZJ, You QD, Guo
QL (2013) Enhanced 5-fluorouracil cytotoxicity in high COX-2
expressing hepatocellular carcinoma cells by wogonin via the
PI3K/Akt pathway. Biochem Cell Biol 91(4):221–229
16. Nagai H, Sumino Y (2008) Therapeutic strategy of advanced
hepatocellular carcinoma by using combined intra-arterial che-
motherapy. Recent Pat Anticancer Drug Discov 3(3):220–226
17. Kavitha K, Kowshik J, Kishore TK, Baba AB (1830) Nagini S
(2013) Astaxanthin inhibits NF-jB and Wnt/b-catenin signaling
pathways via inactivation of Erk/MAPK and PI3K/Akt to induce
intrinsic apoptosis in a hamster model of oral cancer. Biochim
Biophys Acta 10:4433–4444
18. Thevenod F, Lee WK (2013) Cadmium and cellular signaling
cascades: interactions between cell death and survival pathways.
Arch Toxicol 87(10):1743–1786
19. Elliott SL, Crawford C, Mulligan E, Summerfield G, Newton P,
Wallis J, Mainou-Fowler T, Evans P, Bedwell C, Durkacz BW,
Willmore E (2011) Mitoxantrone in combination with an inhib-
itor of DNA-dependent protein kinase: a potential therapy for
high risk B-cell chronic lymphocytic leukaemia. Br J Haematol
152(1):61–71
20. Yoon JH, Ahn SG, Lee BH, Jung SH, Oh SH (2012) Role of
autophagy in chemoresistance: regulation of the ATM-mediated
DNA-damage signaling pathway through activation of DNA-
PKcs and PARP-1. Biochem Pharmacol 83(6):747–757
21. Panta GR, Kaur S, Cavin LG, Cortes ML, Mercurio F, Lothstein
L, Sweatman TW, Israel M, Arsura M (2004) ATM and the
catalytic subunit of DNA-dependent protein kinase activate NF-
kappaB through a common MEK/extracellular signal-regulated
kinase/p90 (rsk) signaling pathway in response to distinct forms
of DNA damage. Mol Cell Biol 24(5):1823–1835
22. An J, Zou W, Chen C, Zhong YF, Yu ZQ, Wang QJ (2013) The
cytological effects of HBCDs on human hepatocyte L02 and the
potential molecular mechanism. J Environ Sci Health Part A
48(11):1333–1342
23. Birbach A, Gold P, Binder BR, Hofer E, de Martin R, Schmid JA
(2002) Signaling molecules of the NF-kappa B pathway shuttle
constitutively between cytoplasm and nucleus. J Biol Chem
277:10842–10851
24. Buttke TM, Sandstrom PA (1994) Oxidative stress as a mediator
of apoptosis. Immunol Today 15:7–10
25. Fleury C, Mignotte B, Vayssiere JL (2002) Mitochondrial reac-
tive oxygen species in cell death signaling. Biochimie
84:131–141
26. Anderson ME (1998) Glutathione: an overview of biosynthesis
and modulation. Chem Biol Interact 111–2:1–14
27. Hammond CL, Lee TK, Ballatori N (2001) Novel roles for glu-
tathione in gene expression, cell death, and membrane transport
of organic solutes. J Hepatol 34:946–954
28. Chen X, Carystinos GD, Batist G (1998) Potential for selective
modulation of glutathione in cancer chemotherapy. Chem Biol
Interact 111–2:263–275
29. Franco R, Cidlowski JA (2012) Glutathione efflux and cell death.
Antioxid Redox Signal 17(12):1694–1713
30. Beskow C, Skikuniene J, Holgersson A, Nilsson B, Lewensohn
R, Kanter L, Viktorsson K (2009) Radioresistant cervical cancer
shows upregulation of the NHEJ proteins DNA-PKcs, Ku70 and
Ku86. Br J Cancer 101:816–821
31. Xing J, Wu X, Vaporciyan A, Spitz MJG (2008) Prognostic
significance of ataxia-telangiectasia mutated, DNA-dependent
protein kinase catalytic subunit, and Ku heterodimeric regulatory
complex 86-kD subunit expression in patients with nonsmall cell
lung cancer. Cancer 112:2756–2764
Mol Cell Biochem
123

32. Willmore E, Elliott S, Mainou-Fowler T, Summerfield G, Jackson
G, O’Neill F, Lowe C, Carter A, Harris R, Pettitt A, Cano-
Soumillac C, Griffin RJ, Cowell IG, Austin CA, Durkacz BW
(2008) DNA-dependent protein kinase is a therapeutic target and
an indicator of poor prognosis in B-cell chronic lymphocytic
leukemia. Clin Cancer Res 14:3984–3992
33. Tian X, Chen G, Xing H, Weng D, Guo Y, Ma D (2007) The
relationship between the down-regulation of DNA-PKcs or Ku70
and the chemosensitization in human cervical carcinoma cell line
HeLa. Oncol Rep 18(4):927–932
34. Peddi P, Loftin CW, Dickey JS, Hair JM, Burns KJ, Aziz K,
Francisco DC, Panayiotidis MI, Sedelnikova OA, Bonner WM,
Winters TA, Georgakilas AG (2010) DNA-PKcs deficiency leads
to persistence of oxidatively induced clustered DNA lesions in
human tumor cells. Free Radic Biol Med 48(10):1435–1443
35. Povirk LF, Zhou RZ, Ramsden DA, Lees-Miller SP, Valerie K
(2007) Phosphorylation in the serine/threonine 2609–2647 cluster
promotes but is not essential for DNA-dependent protein kinase-
mediated nonhomologous end joining in human whole-cell
extracts. Nucleic Acids Res 35(12):3869–3878
36. Wang K, Zhuang Y, Liu C, Li Y (2012) Inhibition of c-Met
activation sensitizes osteosarcoma cells to cisplatin via suppres-
sion of the PI3K-Akt signaling. Arch Biochem Biophys
526(1):38–43
37. Azijli K, Weyhenmeyer B, Peters GJ, de Jong S, Kruyt FA (2013)
Non-canonical kinase signaling by the death ligand TRAIL in
cancer cells: discord in the death receptor family. Cell Death
Differ 20(7):858–868
38. Hartojo W, Silvers AL, Thomas DG, Seder CW, Lin L, Rao H,
Wang Z, Greenson JK, Giordano TJ, Orringer MB, Rehemtulla
A, Bhojani MS, Beer DG, Chang AC (2010) Curcumin promotes
apoptosis, increases chemosensitivity, and inhibits nuclear factor
kappaB in esophageal adenocarcinoma. Transl Oncol
3(2):99–108
39. Chung C, Park SG, Park YM, Joh JW, Jung G (2007) Interferon-
gamma sensitizes hepatitis B virus-expressing hepatocarcinoma
cells to 5-fluorouracil through inhibition of hepatitis B virus-
mediated nuclear factor-kappaB activation. Cancer Sci
98(11):1758–1766
Mol Cell Biochem
123
Copyright © 2022 FDOKUMEN




















