
Distribution, classification, and development ofDrosophila glial cells in the late embryonic and...
24
Roux's Arch Dev Biol (1995) 204:284-307 © Springer-Verlag 1995 Kei Ito - Joachim Urban • Gerhard Martin Technau Distribution, classification, and development of Drosophila glial cells in the late embryonic and early larval ventral nerve cord Received: 22 June 1994 / Accepted in revised form: 10 October 1994 Abstract To facilitate the investigation of glial develop- ment in Drosophila, we present a detailed description of the Drosophila glial cells in the ventral nerve cord. A GAL4 enhancer-trap screen for glial-specific expression was performed. Using UAS-lacZ and UAS-kinesin-lacZ as reporter constructs, we describe the distribution and morphology of the identified glial cells in the fully dif- ferentiated ventral nerve cord of first-instar larvae just after hatching. The three-dimensional structure of the glial network was reconstructed using a computer. Using the strains with consistent GAL4 expression during late embryogenesis, we traced back the development of the identified cells to provide a glial map at embryonic stage 16. We identify typically 60 (54-64) glial cells per ab- dominal neuromere both in embryos and early larvae. They are divided into six subtypes under three catego- ries: surface-associated glia (16-18 subperineurial glial cells and 6-8 channel glial cells), cortex-associated glia (6-8 cell body glial cells), and neuropile-associated glia (8-10 nerve root glial cells, 14-16 interface glial cells, and 3-4 midline glial cells). The proposed glial classifi- cation system is discussed in comparison with previous insect glial classifications. Key words Drosophila • Central nervous system • Glia GAL4 enhancer trap • Classification Abbreviations %NAP Percent neuromere antero-posterior %NML Percent neuromere medio-lateral • %NVD Percent neuromere ventro-dorsal - ac Anterior commissure • act Cell body fibre tracts to the anterior commissure - aCC anterior corner cell AEL After egg laying - A-SPG A subperineurial glial cell • B-SPG B subperineurial glial cell • cbfCell body fibres - CBG Cell body glia (glial cell) • CG Channel glia (glial cell) - CNS Central nervous system - D Dorsal • DAB 3,3'-diaminobenzidine • DL- SPG Dorsal lateral subperineurial glial cell -dn Dorsal nerve (transverse nerve) • DV channel Dorsoventral channel • D-CG K. Ito 1 • J. Urban • G.M. Technau (tS~) lnstitut ftir Genetik, Universit~t Mainz, Saarstr. 21, D-55122 Mainz, Germany Present address: 1 ERATO Project, Mitsubishi Kasei Institute of Life Sciences, Minami-Ooya 11, Machida, 194 Tokyo, JAPAN Dorsal channel glial cell • D-IG Dorsal interface glial cell (cluster) D-MG Dorsal midline glial cell (cluster) - DRG Dorsal roof glia EG Exit glia (glial cell) - FETi Fast extensor tibiae. GDA glutardialdehyde • GFAP Glial fibrillary acid protein • G1Gl Glial glia • IG Interface glia (glial cell) • ISNG Intersegmental nerve root glia (glial cell) • ISNR Intersegmental nerve root • L Lateral • L1 First instar larvae just after hatching • D-SPG Lateral dorsal subperineurial glial cell • LG longitudinal glia (glial cell) • LV-SPG Lateral ventral subperineurial glial cell • L-CBG Lateral cell body glial cell. L-IG Lateral interface glial cell (cluster) • L-ISNG Lat- eral intersegmental nerve root glial cell • L-SNG Lateral segmental nerve root glial cell • M medial • MD-SPG Medial dorsal subperineurial glial cell - MG midline glia (glial cell) (cluster) • MM medialmost • MM-CBG medialmost cell body glial cell • MNB Median neuroblast • MV-SPG Medial ventral subperineurial glial cell. m-cbfCell body fibre tracts from the midline neurons • M-CBG Medial cell body glial cell. M-CG Medial channel glial cell (cluster) • M-ISNG Medial intersegmental nerve root glial cell • M-SNG Medial segmental nerve root glial cell • NG Nerve root glia (glial cell) • nho Neurohemal organ - NPG Neuropilar glia (glial cell) • PBS phosphate-buffered saline • PBT PBS with Triton-X .pc Posterior commissure • pct Cell body fibre tracts to the posterior commissure • PEM PIPES-EGTA-Mg SO 4 buffer. PG Peripheral glia (glial cell) • PNG Perineurial glia (glial cell). RT room temperature • SaGl Satellite glia • SBC Segment boundary cell SNG Segmental nerve root glia (glial cell) • SNR Segmental nerve root • SPG Subperinenrial glia (glial cell) • TpGl Transport glia TrGl tracheal glia • V Ventral • VL-CBG Ventral lateral cell body glial cell. VL-SPG Ventral lateral subperineurial glial cell • VNC Ventral nerve cord • vt Vertical cell body fibre tracts to the dorsal neuropile • VUM Ventral unpaired median - V-CG Ventral channel glial cell • V-IG Ventral interface glial cell • V-MG Ventral midline glial cell (cluster) Introduction Neurons of the central nervous system (CNS) are sup- ported by glial cells in various ways. Vertebrate astrocyte glia transport and provide nutrients to neurons, and oligo- dendrocyte glia form insulation layers around axons. Dur- ing neurogenesis glial cells also provide a scaffold for the correct migration of neurons and growth cones (Singer et al. 1979; Hatten 1990; Gray and Sanes 1992) as well as guidance cues for growing axons in insects (Bastiani and Goodman 1986; Jacobs and Goodman 1989a, b; Kl~imbt
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Distribution, classification, and development ofDrosophila glial cells in the late embryonic and...

Roux's Arch Dev Biol (1995) 204:284-307 © Springer-Verlag 1995
Kei Ito - Joach im U r b a n • G e r h a r d M a r t i n T e c h n a u
Distribution, classification, and development of Drosophila glial cells in the late embryonic and early larval ventral nerve cord
Received: 22 June 1994 / Accepted in revised form: 10 October 1994
Abstract To faci l i ta te the i nves t iga t ion of g l ia l deve lop- m e n t in Drosophila, we presen t a de ta i led desc r ip t ion o f the Drosophila gl ia l cel ls in the ven t ra l ne rve cord. A G A L 4 e n h a n c e r - t r a p sc reen for g l i a l - spec i f i c express ion was pe r fo rmed . U s i n g U A S - l a c Z a n d U A S - k i n e s i n - l a c Z as repor ter cons t ruc ts , we desc r ibe the d i s t r ibu t ion and m o r p h o l o g y of the iden t i f i ed gl ia l cel ls in the fu l ly dif- fe ren t ia ted ven t ra l n e rv e cord of f i r s t - i n s t a r l a rvae j u s t after ha tch ing . Th e t h r e e - d i m e n s i o n a l s t ruc ture of the gl ia l n e t w o r k was r econs t ruc t ed u s i n g a computer . U s i n g the s t ra ins wi th cons i s t en t G A L 4 express ion d u r i n g late e m b r y o g e n e s i s , we t raced b a c k the d e v e l o p m e n t o f the iden t i f i ed cells to p rov ide a gl ia l m a p at e m b r y o n i c s tage 16. We iden t i fy typ ica l ly 60 ( 5 4 - 6 4 ) gl ial cel ls per ab- d o m i n a l n e u r o m e r e bo th in e m b r y o s a n d ear ly larvae. T h e y are d iv ided in to six sub types u n d e r three ca tego- ries: s u r f a c e - a s s o c i a t e d g l ia ( 1 6 - 1 8 subpe r ineu r i a l gl ial cel ls and 6 - 8 c h a n n e l gl ial cells) , c o r t e x - a s s o c i a t e d g l ia ( 6 - 8 cel l b o d y gl ia l cells) , and n e u r o p i l e - a s s o c i a t e d g l ia ( 8 - 1 0 ne rve root gl ia l cel ls , 1 4 - 1 6 in te r face gl ia l cells , and 3 - 4 m i d l i n e gl ia l cells). Th e p r o p o s e d gl ia l c lassif i - ca t ion sys t em is d i scussed in c o m p a r i s o n wi th p rev ious insec t g l ia l c lass i f ica t ions .
K e y w o r d s Drosophila • Cent ra l ne rvous sys t em • G l i a G A L 4 e n h a n c e r trap • C lass i f i ca t ion
Abbreviations %NAP Percent neuromere antero-posterior %NML Percent neuromere medio-lateral • %NVD Percent neuromere ventro-dorsal - ac Anterior commissure • act Cell body fibre tracts to the anterior commissure - aCC anterior corner cell AEL After egg laying - A-SPG A subperineurial glial cell • B-SPG B subperineurial glial cell • cbfCell body fibres - CBG Cell body glia (glial cell) • CG Channel glia (glial cell) - CNS Central nervous system - D Dorsal • DAB 3,3'-diaminobenzidine • DL- SPG Dorsal lateral subperineurial glial cell -dn Dorsal nerve (transverse nerve) • DV channel Dorsoventral channel • D-CG
K. Ito 1 • J. Urban • G.M. Technau (tS~) lnstitut ftir Genetik, Universit~t Mainz, Saarstr. 21, D-55122 Mainz, Germany
Present address: 1 ERATO Project, Mitsubishi Kasei Institute of Life Sciences, Minami-Ooya 11, Machida, 194 Tokyo, JAPAN
Dorsal channel glial cell • D-IG Dorsal interface glial cell (cluster) D-MG Dorsal midline glial cell (cluster) - DRG Dorsal roof glia EG Exit glia (glial cell) - FETi Fast extensor tibiae. GDA glutardialdehyde • GFAP Glial fibrillary acid protein • G1Gl Glial glia • IG Interface glia (glial cell) • ISNG Intersegmental nerve root glia (glial cell) • ISNR Intersegmental nerve root • L Lateral • L1 First instar larvae just after hatching • D-SPG Lateral dorsal subperineurial glial cell • LG longitudinal glia (glial cell) • LV-SPG Lateral ventral subperineurial glial cell • L-CBG Lateral cell body glial cell. L-IG Lateral interface glial cell (cluster) • L-ISNG Lat- eral intersegmental nerve root glial cell • L-SNG Lateral segmental nerve root glial cell • M medial • MD-SPG Medial dorsal subperineurial glial cell - MG midline glia (glial cell) (cluster) • MM medialmost • MM-CBG medialmost cell body glial cell • MNB Median neuroblast • MV-SPG Medial ventral subperineurial glial cell. m-cbfCell body fibre tracts from the midline neurons • M-CBG Medial cell body glial cell. M-CG Medial channel glial cell (cluster) • M-ISNG Medial intersegmental nerve root glial cell • M-SNG Medial segmental nerve root glial cell • NG Nerve root glia (glial cell) • nho Neurohemal organ - NPG Neuropilar glia (glial cell) • PBS phosphate-buffered saline • PBT PBS with Triton-X .pc Posterior commissure • pct Cell body fibre tracts to the posterior commissure • PEM PIPES-EGTA-Mg SO 4 buffer. PG Peripheral glia (glial cell) • PNG Perineurial glia (glial cell). RT room temperature • SaGl Satellite glia • SBC Segment boundary cell SNG Segmental nerve root glia (glial cell) • SNR Segmental nerve root • SPG Subperinenrial glia (glial cell) • TpGl Transport glia TrGl tracheal glia • V Ventral • VL-CBG Ventral lateral cell body glial cell. VL-SPG Ventral lateral subperineurial glial cell • VNC Ventral nerve cord • vt Vertical cell body fibre tracts to the dorsal neuropile • VUM Ventral unpaired median - V-CG Ventral channel glial cell • V-IG Ventral interface glial cell • V-MG Ventral midline glial cell (cluster)
Introduction
N e u r o n s of the centra l ne rvous sys t em (CNS) are sup- por ted by gl ial cells in var ious ways. Vertebrate as t rocyte gl ia t ranspor t and p rov ide nu t r ien t s to neu rons , and o l igo- dendrocy te g l ia fo rm insu l a t i on layers a round axons. Dur - ing neu rogenes i s gl ia l cells also p rov ide a scaffold for the correct m ig ra t i on of n e u r o n s and g rowth cones (S inger et al. 1979; Ha t t en 1990; Gray and Sanes 1992) as wel l as gu idance cues for g rowing axons in insects (Bas t ian i and G o o d m a n 1986; Jacobs and G o o d m a n 1989a, b; Kl~imbt

et al. 1991; Jacobs 1993; Seeger et al. 1993). It is also likely that glial cells participate in controlling neuronal proliferation (Ebens et al. 1993) and in the maintenance of mature CNS structure (Buchanan and Benzer 1993).
In spite of their importance in neurogenesis and CNS function, relatively little is known about the mechanisms of glial determination, development and glia-neuron in- teractions. Because of the vast array of genetic, molecu- lar and cellular experimental tools available, the fruit fly Drosophila melanogaster offers an attractive model sys- tem to study these mechanisms.
For the interpretation of mutant phenotypes and gene expression patterns, a detailed knowledge of the types, distribution and development of glial cells in wild-type Drosophila is required. Although several glial cells have been identified in the Drosophila early embryonic CNS (Jacobs and Goodman 1989a,b; Kl~imbt and Goodman 1991), as well as in the peripheral and postembryonic ner- vous system (Winberg et al. 1992; Giangrande et al. 1993; Nelson and Laughon 1993; Choi and Benzer 1994; Gian- grande 1994; Prokop and Technau 1994), researchers of- ten have to rely on the analogy with larger insects (mostly adults) such as the housefly (Sohal et al. 1972; Strausfeld 1976), locust (Hoyle 1986), and moth (Cantera 1993).
In this study, we describe the number, distribution and morphology of Drosophila glial cells in more detail by observing GAL4 enhancer-trap strains that label certain subsets of CNS cells. For the following reasons, we se- lected the first instar larva just after hatching (termed here as L1) for identifying glia (see Fig. 1): First, it represents the end of embryonic development; the state of differenti- ation reached by each cell makes it easy to clearly distin- guish glia and neurons. Second, since there is no cell pro- liferation in the ventral nervous system of early larvae (Truman and Bate 1988; Ito and Hotta 1992), the number of cells should be stable. Third, the morphology and dis- tribution of glial cells in the functional larval CNS pro- vide a better classification of glial cell types; in late em- bryos, many neurons and glia show immature morpholo- gy and there are also some neuroblasts still dividing at this stage (Prokop and Technau 1991).
We identified 60 glial cells per abdominal neuromere at L1; the number may vary between segments and indi- viduals from 54 to 64. Using strains with consistent GAL4 expression during late embryogenesis, we traced back the identified glia through the late embryonic stag- es. The resulting glial map of the stage-16 embryonic ventral nerve cord (VNC, stages according to Campos- Ortega and Hartenstein 1985), which can easily be stained en masse as whole mounts, will be useful for characterization of gene expression and large-scale screenings for glial mutants.
Based on our description of identified glial cells in the Drosophila embryonic and early larval VNC, we pro- pose a glial classification system with six subtypes under three categories. For the late larval and adult VNC and brain, the system can accommodate further cell types without altering its framework. In comparison, previous glial descriptions for various insects are discussed.
285
Materials and methods
Screening, fly strains, and markers
Enhancer-trap strains were generated by crossing a pGawB strain (Brand and Perrimon 1993) with a Pity+; A2-3] strain (Robertson et al. 1988). To reveal the GAL4 expression pattern, male GAL4 flies were crossed with female flies carrying the UAS-lacZ detec- tor construct (lacZ gene under the control of the GAL4 yeast UAS (upstream activation sequence); Brand and Perrimon 1993) or with flies carrying the UAS-kinesin-lacZ construct (courtesy of Ed Gi- niger; see also Giniger et al. 1993 for the kinesin-lacZ gene con- struct). The UAS-IacZ construct labelled nuclei and the surround- ing cytoplasm; sometimes large protrusions as well as axons were also labelled. The UAS-kinesin-lacZ construct labelled the cyto- plasm of more distal parts of the protrusions, while the nuclei re- mained unstained (for comparison of staining by the two reporters, see Fig. 5D, F and Fig. 9A,B).
We generated about 1,600 fly strains carrying single GAL4 en- hancer-trap insertions. The first screening was performed for the embryonic GAL4 expression, and we chose 90 strains that label various subsets of embryonic CNS cells. The second screening was carried out by staining the CNS dissected from newly hatched larvae. Nineteen strains were found to be useful for glial identifi- cation. Among these six labelled only glia, while the others also labelled subsets of neurons (Table 1). To confirm the consistency of the GAL4 expression pattern during late embryogenesis, we also stained the CNS dissected from late embryos (mid stage 17; 18-22 h after egg laying: AEL) and compared them with the larval (24 h AEL) and late stage-16 (15-16 h AEL) CNS.
Most of our strains also showed GAL4 expression in structures other than the CNS, such as salivary glands, oenocytes, epidermis and gut. Since we dissected out the larval CNS by cutting the pe- ripheral nerves, glia in the peripheral nervous system (PNS) were not investigated (exit glia and peripheral glia in Klfimbt and Good- man 1991; dorsal roof glia in Nelson and Laughon 1993).
We also used several markers other than the GAL4 strains. The P-lacZ enhancer-trap strains AA142, M84, 3-109, and P I O1 were kindly provided by C. Kl~mbt. The transformant strains sim-lacZ and slit-lacZ were gifts from S. Crews. The anti-prospero anti- body was kindly provided by E. Spana and C. Q. Doe, and anti-re- po 4(z3 by D. Halter and A. Travers.
Staining of whole-mount embryos
Collected embryos were devitellinized with Clorox (50%, 4 rain), fixed for 30 rain with heptane saturated with phosphate-buffered saline (PBS) containing 10% formaldehyde, and devitellinized with methanol/heptane mixture. After five washes with PBT (PBS with 0.5% Triton-X) embryos were pre-incubated for 2 h at room temperature (RT) with 10% lamb serum in PBT. Incubation with the anti-fl galactosidase (]3 Gal) rabbit polyclonal primary anti- body (Cappel; 1:3000- 1:7000 in 10% lamb serum/PBT) and bi- otin-conjugated secondary antibody (Dianova; 1:500 in 10% lamb serum/PBT) were both performed at 4°C over night. Labeling was revealed using a Vectastain Elite ABC kit with 0.5-1 mg/ml DAB (3,3'-diaminobenzidine) and 0.05% H202. In some cases 0.06% of NiC12 was added to enhance the signal contrast. Stained specimens were dehydrated, cleared, mounted in araldite (Serva; 46% CY 212, 46% HY 964, 5% phtalic acid dibuthyl ester, 3% DY 964), and sucked into capillaries for observation (Prokop and Technau 1993).
Photographs were taken with an Axiophot microscope (Carl Zeiss) with x63 Nomarski optics and x2.00ptovar. In some cases, several photographs with slightly different focal planes were digi- tally montaged with Adobe Photoshop 2.0 (Adobe) on a Macin- tosh computer (Apple).

286
Staining of dissected larval and late embryonic CNS
Flies were allowed to lay eggs for 3 h. After 22-24 h at 25°C, larvae already hatched were removed and, 3 h later, newly hatched larvae were collected, so they represented a population 0 to 3 h after hatch- ing. The CNS were dissected in PBS with watchmakers' forceps and transferred with a 200-gl pipette into a 1.5-ml microfuge tube, which contained about 50 gl of the fixative solution. Mid to late stage-17 embryonic CNS (stages according to Campos-Ortega and Hartenstein 1985) were dissected from embryos that were devitel- linized and fixed as described. After 30-50 specimens were pooled, they were further treated with fresh fixative. Either 4% formalde- hyde in PEM (0.1 MPIPES, 2 mMEGTA, 1 mM MgSO 4, pH 7.0) (30-40 min at RT) or 1% glutardialdehyde (GDA) in PBS (10 min at RT) was used as fixative. For anti-fl GaI polyclonal antibody staining, using GDA resulted in better conservation of the speci- mens and lower background. Some monoclonal antibodies, howev- er, showed no staining when fixed with GDA.
The CNS were treated with 0.3% H202 in methanol for 30 min at RT to inactivate the endogeneous peroxidase, followed by anti- body staining as described. Because of the small size and fragile surface of the dissected CNS, the amount of Triton-X detergent in PBT was reduced to 0.01-0.005% or PBS was used instead.
Staining of larval CNS sections
The fixation and antibody staining were done as described earlier; GDA fixation was preferred. The H202 added to the DAB was re- duced to 0.01% in order to avoid Oz bubbles emerging within the specimens. NiC12 enhancement was avoided, since the resulting blue-black staining tends to be lost during the subsequent GDA post-fixation (6% GDA in PBS for 2 h at RT). Specimens were dehydrated with a series of ethanol (30%, 50%, 70%, 90%, and two times 100%), cleared with xylene, and incubated in 50% ara- ldite in xylene over night at RT. To allow gradual evaporation of the xylene, the microfuge tubes were kept open in a draft chamber.
The specimens were further incubated in 100% araldite for 6-8 h at RT, placed in moulds with fresh araldite, and polymerized for 12 h at 45°C and 36 h at 65°C, followed by 1-~tm semi-thin sec- tioning and counter staining with Richardson's medium (1% tolui- dine blue and 1% azur II in 1% Borax) for 15 s to 3 min at 50°C.
Three-dimensional reconstruction
For the stereoscopic 3-D reconstruction (Figs. 7, 8), the contours of the nervous system and the labelled cells were traced from pho- tographs of 1-gm serial sections onto transparent films, and scanned into a Macintosh computer. About 30 sections were need- ed to reconstruct two neuromeres. The scanned data were three-dimensionally aligned and rendered with StrataVision 2.6 (Strata Inc.) using ray tracing algorithm. Two pictures with six de- grees of viewing-angle distance were generated and combined to achieve the stereoscopic effect.
The schematic 3-D reconstruction (Fig. 10) was drawn by hand with the aid of the Serial Section Reconstruction System (SSRS; Eutectics Inc.).
Results
The UAS-IacZ and UAS-kinesin-lacZ reporter con- structs revealed the cytoplasmic structure of the GAL4-expressing cells. This enabled us to identify glial cells by their non-neuronal cell morphology. The en- hancer-trap strains labelled various overlapping subsets of glial cells; we found no single strain that labels all the glia reported in this study (Table 1). By comparing la- belled cells in different strains, we deduced the consen- sus distribution pattern of 60 glial cells per neuromere
Table 1 Summary of the staining patterns used in this study (gray boxes indicate that the marker labels all or a subset of cells of the subtype). The first 23 are enhancer-trap strains. The expression of the genes sire, and slit were revealed with p r o m o t e r - l a c Z constructs (slit: Rothberg et al. 1988; sire: Nambu et al. 1991; strains courtesy of S. Crews). The en, pros, and repo expression were studied with antibodies (anti-en 4D9: Patel et al. 1989; anti-pJvs: courtesy of E. Spana and C. Q. Doe; anti-repo 4a3: courtesy of D. Halter and A. Travers)
Mz 97
Mz 143 Mz1251 Mz 317 Mz 512 Mz 820 Mz1067 Mz 229 Mz 376 Mz1127 Mz 709 Mz 840 Mz 770 Mz1457 Mz 360 Mz 419 Mz 520 Mz 685 Mz1299
M 8 4 P 101 3-109
A A 142 sire sli t
pros en
rcpo
neurons glia
surface ~ neuropile
PNG
!--
NPG

Table 2 Unique names of identified glial cells and com- parison with previous descrip- tions (the names of the A and B glia were changed to A- and B-SPG, since they were found to be members of the SPG sub- type. The VUM support glia was found to be the medialmost member of the CBG. As for the nerve root glia (NG) subtypes, we took the names ISNG and SNG. Characters "M" and "L" were used to indicate the posi- tions. IG and LG define differ- ent sets of cells; IG is defined by position and LG by origin - see text. Although the lateral, ventral and a part of dorsal IG are identical to the LG1-6, we could not establish the one-to-one relationship. The origin of IG other than the LG -unnamed cells in Goodman and Doe 1993 - is unknown. We did not find cells equivalent to the MGP. The MGA and MGM form one cluster - MG cluster - at stage 16 and then randomly separate into two clusters. It seems there is no one-to-one relationship be- tween the MGAIMGM and D-MGIV-MG)
our nomenclature
- - - - - - 1 SNG 1 h :?::I
SPG
CG
M -SNG2 L -SNG
D -IG cluster (3-5) L -IG cluster (2)
A -SPG B -SPG
MD-SPG LD -SPG DL -SPG VL -SPG LV -SPG MV-SPG
D -CG
M -CG cluster (2)
V -CG
MG D -MG cluster (1-2) V -MG cluster (1-2)
(Fig. 2 and Table 2). The number and position of labelled glial cells showed some variability between segments and individuals (54-64 cells per neuromere). To describe the positions of cells in the ventral nerve cord, we intro- duced a metameric Cartesian coordinate system (%NAP, %NML, and%NVD, see Fig. 1).
Unique names were given to single cells that are easi- ly identifiable. Other cells were identified only as groups of cells (see Fig. 2 for detail). We classified the identi- fied cells in two levels. The individual cells that share common morphological, positional and molecular fea- tures were classified into six "subtypes". These fell into three "categories" according to their association with the basic regions of the CNS: the surface, the cortex and the neuropile.
I: Surface-associated glia
The "surface-associated glia" category represents the cells that are closely associated with the CNS surface. The surface is covered by the perineurium, which con- sists of the acellular neural lamella (Scharrer 1939) and the surface-associated glial cells underneath (Fig. 3). In the embryonic and L1 VNC we defined two subtypes of this glial category. The "subperineurial glia" (SPG) lie beneath the outer surface of the VNC, and the "channel glia" (CG) lie along the dorsoventral channels.
A -SPG B -SPG
MD-SPG LD -SPG DL -SPG VL -SPG LV -SPG MV-SPG
D -CG M -CGl M -CG2 V -CG
Stage 16
MM-CBG VUM support cell M -CBG - VL -CBG I -
previous nomenclature
Klambt and Goodman and Goodman (1991)
Stage 15
L -CBG - M 4SNG / SBC
M -SNG2 L -SNG
D -IG cluster (3-5) LG1-6 + 2-4 unnamed
Subperineurial glia (SPG)
The distribution pattern of the SPG is metameric but shows slight variability (Fig. 4A,B at L1, C and D at late stage 16). The nuclei and the cell bodies of SPG are round and flat (Fig. 3, also compare Fig. 4D and F). Cy- toplasmic extensions of the SPG sometimes protrude a short distance into the underlying cortex and fill the space between the outermost neuronal cell bodies (e.g. Fig. 3C). In a typical case we found eight SPG (some- times nine) per abdominal hemineuromere (Fig. 2 for summary).
The ventral surface has two SPG per hemineuromere (black arrowheads in Fig. 4B and D). The medial one (medial ventral SPG, MV-SPG) lies at 30-SO%NML and around SO%NAP. In the thoracic segments it often lies slightly more medially than in the abdomen. The lat- eral one (lateral ventral SPG, LV-SPG) lies at 60-90%NML and near the anterior border of the neuro- mere (0-lO%NAP), hence antero-lateral to the medial ventral SPG. Both ventral SPG remain at the same posi- tion between stage 16 and L1 (compare Fig. 4B and D).
Along the lateral surface, two lateral SPG lie at about 30 and 80%NVD (ventral lateral, VL-SPG and dorsal lateral, DL-SPG, Fig. 4E and F). Sometimes there are two ventral lateral SPG instead of one, resulting in a to- tal of three lateral SPG. The exit point of the peripheral nerve in the abdominal neuromeres is slightly above the dorsal lateral SPG (see Fig. 4A and C). Anti-prospero

288
A Supraoesophageal Ganglion (brain)
~ i ! i ~ */.NAP ~ ~ . , / B o ] w i g ' s nerve 0l~ll00
~ ~ - 2 - o e s o p h a g u s [ ' ' [ %NVD . . . . . . . 100
. . . . . . . . . . . Ai / T1 ¢ I I __ A1 A2 A3 A4 A5 A6 A7 A8/9 ] T2 T3
Thorax Abdomen i ~ Ventral Nerve Cord (VNC) = Suboesophageal Ganglion
E oesophagus foramen
C
dorsal nerve
dorsoventral channel
Fig. 1A-C Schematic drawings of the first-instar larval central nervous system (CNS). Lateral (A), horizontal (B) and perspective (C) views of the larval CNS just after hatching. A,B The metameric Cartesian coordinate system (%NAP percent neuromere an- tero-posterior - indicates positions along the longitudinal axis, O%NAP corresponds to the anterior segment border, IO0%NAP posterior segmental border, %NML percent neuromere medio-lat- eral - indicates positions along the mediolateral axis, O%NML cor- responds to the midline, IO0%NML lateralmost surface of the CNS, %NVD percent neuromere ventro~torsal - indicates positions along the vertical axis, O%NVD is the ventral surface, IO0%NVD dorsal surface). The segment borders are marked by the dorsoven- tral channel (DV channel), a duct-like structure penetrating the nervous system vertically on the midline. The inner surface of the channel is contiguous with the outer ventral nerve cord (VNC) sur- face (Fig. 3A,H). In early embryos the channels first appear as the space between a pair of longitudinal connectives and the neigh- bouring neuromeres (Figs. 6 K and 14E). C Shows the neuropile of the T3 neuromere and the abdominal ventral nerve cord. One larval abdominal neuromere (A1-A6) just after hatching is 11-13 gm thick, 70-80 gm wide and 35-40 gm high. Caudal neuromeres (A7 and A8/9) are narrower and distorted. The thoracic neuromeres are thicker, narrower and higher than the abdominal ones. With numer- ous intrinsic axon fibres associated with the commissures and lon- gitudinal tracts, the larval neuromere is much larger and more com- plex than in early embryos. The midline region between the com- missure neuropile is almost devoid of cell bodies (see also Fig. 3). Each neuromere carries three nerves: a pair of peripheral nerves and a neurohemal organ. The neurohemal organ connects to the
a n t i b o d y (cour tesy o f E. Sparta and C. Q. Doe) labels the la teral S P G at the pe r iphery of the lateral c lus ter o f the prospero-positive cells (Fig. 6 o f Do e et al. 1991). The la teral S P G show ver t ica l ly e l o n g a t ed b e l t - l i k e m o r p h o l - ogy at s tage 14. Th i s sugges ts that the la teral S P G are
dorsal VNC at the segment border. The abdominal neurohemal or- gan forms a V-shaped bifurcation to send a pair of dorsal nerves (Hertweck 1931; "transverse nerves" in Gorczyca et al. 1994) to both sides of the body wall, where they are called segment bound- ary nerves (Bodmer and Jan 1987). The bifurcation occurs just above the VNC in the embryo. In larvae, especially in late stages, the stalk between the VNC and the bifurcation point becomes elon- gated, running above the dorsal midline (see Fig. 1 of White and Kankel 1978 for late larvae). The thoracic segments lack the lateral projections from the neurohemal organ (see also Fig. 3 of N~ssel et al. 1988 for Calliphora). The peripheral nerve consists of two sepa- rate fibre bundles until embryonic stage 15-16 (see Fig. l lB,C); they fuse during stage 16-17. The level where the nerves leave the VNC is relatively ventral in thoracic segments and more dorsal in the abdomen (A). The peripheral nerve has two nerve roots. The in- tersegmental nerve root (ISNR) crosses the segment border and forms two branches. The anterior branch enters the neuropile of the anterior segment in the dorsalmost region (90%NVD) at about 50-70%NAP (see also Fig. 3D). The posterior branch enters the neuropile slightly more ventrally (75%NVD) and posteriorly (65-80%NML; see also Fig. 3E). Both branches are associated with the mediolateral fibre tracts in the dorsalmost neuropile. The segmental nerve root (SNR) forms many small branches that enter the ventralmost region of the neuropile (50-60% NVD) at various antero-posterior levels and invade the ventralmost neuropile. In mid-stage embryos, all the nerve roots run perpendicularly to the body axis. As the nervous system contracts, most of them are skewed to pass through the cortex obliquely. This distortion of the nerve roots does not affect the cortex structure
the equ iva l en t s o f the "be l t g l ia" (Doe et al. 1991). In the late s t a g e - 1 6 and la rva l CNS, anti-pros does no t l abe l the la teral S P G (Fig. 16C and D).
The dorsal surface has typ ica l ly four S P G per hemi - n e u r o m e r e in the a b d o m e n and three in the thorax. The

Late Stage 16 ~ , , ~M~SNG
, j ~ .~ "!.: ~ M-SNGI* L-IS /
I I A-SPG
0 ' ~ MV-SpG LV~PG PG
0'~:~-M"CG1 1 %3v2d Surface-associated Glia
Subperineurial Glia (SPG) 16-18 A-SPG, B-SPG, MD-SPG, LD-SPG DL-SPG, VL-SPG*, LV-SPG, MV-SPG
0 Dorsoventral Channel Glia (CG) 7 - 8 D-CG, M-CG cluster (M-CG1, M-CG2), V-CG
~ ortex-associated Glia Cellbody Glia (CBG) 6 - 8
MM-CBG, M-CBG, VL-CBG*, L-CBG
Fig. 2, Summary of glial cells identified in this study. A consensus distribution of cells in one abdominal neuromere is shown. Left and right panels show the late stage-16 embryonic and the first-instar larval CNS, respectively. The top and bottom panels show the hori- zontal view of the dorsal and ventral half of a neuromere (anterior to the top). The middle panels show the frontal view (dorsal to the top). The "unique name" of each identified cell is shown in the fig- ure and in the table below. (The exit glia, EG, are excluded from the table because they are peripheral nervous system glia.) The naming scheme is as follows. The characters after the hyphen indi- cate the subtype the cell belongs to. The characters before the hy- phen indicate the position of the cell (D dorsal, V ventral, L lateral, M medial, MM medialmost). In case of combined characters such as DL, the second character indicates the region in the neuromere while the first character shows the precise position within this re- gion. Suffix numbers are added to identify cells in the same area (M~2G1, M-CG2, M-SNG1 and M-SNG2). Although the A- and B-SPG should be addressed as anterior and posterior MMD-SPG according to our scheme, we took the name A- and B- in favour of compatibility with previous descriptions (see Table 2). In the table, the word "cluster" is added for cells that are only identified as groups. The medial CG are singly identifiable at late stage 16 but not at L1. Cells in the dorsal IG and lateral IG clusters are not iden- tifiable even at stage 16 but can be identified singly until stage 14. The MG can be identified singly until stage 14, they form one clus- ter at stage 16 and then randomly separate into dorsal and ventral clusters. Cells with asterisks showed higher variability than the others. There are sometimes two ventral lateral SPG instead of one. The ventrolateral CBG and one of the medial SNG are sometimes missing. The distribution and number of cells in the dorsal IG clus- ter are highly variable
1st Instar (L1) M.s.,
289
L-ISNG
Neuropile-associated Glia Nerve Root Gila
Intersegmental: M-ISNG, LqSNG ~ , Segmental: M-SNGI*, M-SNG2*, L-SNG (SNG) O Interface Glia (IG)
D-IG cluster*, L-IG cluster, V-IG • Midline Gila (MG)
D-MG cluster, V-MG cluster (MG cluster)
L-CBG
(ISNG) 4 4 - 6 14-16
3 - 4
Total 5 4 - 6 4
lateral dorsal SPG (LD-SPG, 8 0 - 9 0 % N M L ) lies near, but slightly above, the exit point o f the peripheral nerve (Figs. 4A,C and l lC). The medial dorsal SPG ( M D - S P G , 3 0 - 6 0 % N M L ) lies at the centre between the intersegmentai nerve roots (Figs. 4A and 5D,E), slightly antero-lateral to the medial intersegmental nerve root gli- al cell (M- ISNG, see later). Their positions do not change during late embryogenesis . The medialmost re- gion has two glial cells in the abdomen ( A - S P G and B - S P G , 20 -30%NML) . The A - S P G and B - S P G in the abdominal segments lie in tandem above the neuropile (Fig. 4A). They are identical to the "A and B glia" de- scribed by Kl~imbt and Goodman (1991, Fig 5A,B). The thoracic neuromeres have only one glial cell in this area; they lack the B - S P G . The thoracic A - S P G has the same position and morphology as its abdominal counterparts al though it belongs to a different cell lineage ( "A-B- l ike glia" in Udolph et al. 1993). The A - S P G lies near the an- terior segment border and sends a process to the dorso- ventral channel (thin arrows in Fig. 4A). At stage 15 the A - S P G lies posterolateral to the channel (* in Fig. 5A); it migrates slightly anteriorly during stage 16 (Fig. 5B). The cytoplasmic staining at this stage (Fig. 4C) shows a wing- l ike structure formed by the A - S P G and the dorsal channel glial cells (D-CG) . The B - S P G lies at about the centre of the segment ( 5 0 - 6 0 % N A R Fig. 5A,B), sending

Fig. 3 A - H The anatomy of the wild-type Drosophila larval ven- tral nerve cord (VNC), showing eight of the t7 serial sections (1 gm) from the first abdominal (A1) neuromere of a mid sec- ond-instar larva (30 h after larval hatching) stained with toluidine blue. Since this stage is slightly before the onset of postembryonic cell proliferation (Truman and Bate 1988), the number of glial and neuronal cells should be the same as at L1. The size of the nervous system, however, has increased by 50-60% during the 30 h of post- embryonic development. The acellular neural lamella is lightly stained along the outer VNC surface and the DV channels; it is thin at the lateral surface but thick at the dorsal midline and along the channel. Glial cells are stained darker than the neuronal cells. Un- like the ventral and lateral cortex, the dorsal cortex above the neu- ropile (15-40% NML) is very thin (about one cell diameter); few
neurons lie in this region. The medialmost region of the dorsal cor- tex (0-15%NML) is thicker and accommodates two rows (one row in each hemineuromere) of big neuronal cell bodies, among which are the aCC (anterior Corner Cell) motoneurons. The cortex/neuro- pile interface in this region forms a V-shaped "valley". The medial dorsal IG lie along the slope of this valley, while the dorsal CG and MG reside in it. Large bundles of cell body fibres (cbf) enter the neuropile at the ventrolateral region of the cortex/neuropile inter- face (ac anterior commissure, pc posterior commissure, act cbf tracts to the ac, pct cbf tracts to the pc, vt vertical cbf tracts to the dorsal neuropile, m-cbf cbf tracts from the midline neurons, ISNR intersegmental nerve root, SNR segmental nerve root, see Fig. 2 and Abbreviations list for glial abbreviations)

291
Fig. 4A-F Subperineurial glia (anterior to the left, bar 50 gm). A-D Strain 347,.97 labelled with UAS-IacZ (Mz97/lacZ). A Dorsal surface at L1 (montage of 3 pictures). White arrowheads indicate labelled SPG of one segment (A-, B-, MD- and LD-SPG). Thin black arrows point to the connections between the A-SPG and the dorsal CG. The channels are interconnected by thin glial processes running on the midline. Thick arrows indicate the intersegmental nerve root glial ceils (ISNG). B Ventral surface of the same speci- men (montage of 2 pictures), showing the two ventral SPG per hemineuromere (LV, MV). C Dorsal surface at late stage 16 (mon- tage of 4 pictures). Note the wing-like structure between the A-SPG (A) and the dorsal CG (White arrowheads indicate a clus- ter of IG labelled transiently in some segments). D Ventral surface of the same specimen (montage of 4 pictures). E Lateral surface of the L1 VNC of Mz143/kinesin-lacZ, showing lateral SPG and their fiat processes (DL, VL). F Lateral surface of the stage-16 VNC of M84, showing the flat SPG nuclei
processes in three directions: medially to the midline, an- teriorly and posteriorly (Fig. 4C). They form a meshwork of glial processes that covers the medialmost region of the dorsal VNC surface (Fig. 5C,D,F).
To see the extent of the thin glial protrusions, which cannot be observed in whole-mount preparations, we
made a three-dimensional computer reconstruction from the serial sections of L1 neuromeres of the strain Mz1251 (Fig. 7). The strain labels a large subset of SPG as well as channel glia and nerve root glia. From the re- construction it is clear that the larval SPG protrusions cover most of the VNC outer surface area.
Channel glia (CG)
The CG lie along the dorsoventral channels (DV chan- nels). The cell bodies lie close to and send processes along the channel surface. Their nuclei are smaller and more spherical compared to the flat nuclei of the SPG. We found three groups of channel glial cells: dorsal (D-CG: 2 cells), medial (M-CG: 3-4 cells), and ventral (V-CG: 2 cells).
The dorsal CG lie at the dorsal end of the channel above the neuropile (one per hemineuromere; Fig. 6A, B, G, H, L). The cells send processes ventrally along the channel, while receiving processes from the A-SPG (Fig. 4A,C). The position of the dorsal CG cell body is

292
Fig. 5A-F Subperineurial glia (anterior to the left, bar 50 gm). A,B Dorsal surface of M84 abdominal VNC at stage 15 (A) and early stage 17 (B). A and B indicate A- and B-SPG, respectively. Asterisks indicate the DV channels. Note the more random SPG distribution and the more anterior position of the A-SPG at stage 17. C Dorsal surface of Mz1251/lacZ at late stage 16, showing the meshwork structure made by the B-SPG. Note that the structure exists only in the abdominal neuromeres and disappears at the tho- rax/abdomen border (T/A). CG, channel glia. D The same strain (Mz1251/lacZ) at L1. The distorted mesh structure is still visible. The medial ISNG lies just lateral to the B-SPG. The medial dorsal SPG (MD) lie anterolateral to the medial ISNG. E Dorsal surface of Mz143/lacZ VNC at L1, showing flat processes made by the medial dorsal SPG. F The same area of Mz1251/kinesin-lacZ (montage of 2 pictures). The thick B-SPG (B) processes cover the midline region. The thinner processes from the medial dorsal SPG (MD) are weakly stained. Note that D and F show the same GAL4 strain crossed with different detector strains
restricted between 95 and 100%NVD. The ventral CG lies at the other end of the channel, but the position may vary between 0 and 40%NVD (Fig. 6B, C, H, I, L). The two ventral CG of one segment do not necessarily lie at the same dorsoventral level; often one lies at the ventral end and the other more dorsally.
The dorsal and ventral CG send processes to connect with each other. They form a sheath structure that covers the inner surface of the DV channels. The structure is es- tablished already at late stage 16, when the formation of the channel is completed (compare Fig. 6B and H). Al- though the dorsal and ventral CG lie along the midline, they do not originate from the midline progenitors (see Bossing and Technau 1994); during stage 14 they mi- grate from a lateral region towards the midline (Fig. 6J and K). It is likely that the cells are associated with the posterior border of segments. They express engrailed and occupy the posteriormost positions among the en-posit ive cells (Fig. 16A,B).
The medial CG lie and send processes only along the channel at the level of the neuropile (50-95% NVD). Their cell bodies lie just beneath the ventral side of the cortex/neuropile interface (Fig. 6D,E, see also Fig. 3A, H). Unlike the dorsal and ventral CG, sometimes the me- dial CG cell bodies lie slightly away from the channel surface (Fig. 6D). Even in these cases, they always send their processes to and along the channel surface and not to the neighbouring neuropile or other CNS structures. The medial CG are not found among the en-posit ive cells.

Fig. 6A-L Dorsoventral channel glia (anterior to the left, bar 50 ~tm). A-C L1 VNC of Mz820/lacZ; A Horizontal view of the dorsal CG. B Lateral view of the dorsal and ventral CG (D and V, nho neurohemal organ). Montage of 2 pictures. C Horizontal view of the ventral CG (montage of 2 pictures). D-F Medial channel glia: D Lateral view of the L1 VNC of Mz709/lacZ showing the medial CG (M). E Horizontal view of MzTO9/lacZ at the level of the ventralmost neuropile. The medial CG are identifiable only as groups of 3-4 cells (arrows). F Stage 17 VNC (18 h AEL) stained with anti-repo (courtesy of D. Halter and A. Travers). The anterior M-CG1 (1) and the posterior M-CG2 (2) are identifiable at this
stage. G-I Stage-16 VNC of Mz820/lacZ, showing the views cor- responding to the left panels of L1 VNC (A-C). J - K Formation of the DV channel (Mz820/lacZ). At late stage 14 (J) the dorsal half of the channel is already formed (asterisks). Since the midline re- gion of the ventral cortex is formed later than the other cortex re- gions, the ventral half of the channel remains much wider, forming a downward funnel-Iike opening. At early stage 14, the channel appears as a wide opening between the immature commissures and connectives (K). The CG lie at the medial border of this open- ing. L Frontal optical section of Mz820/lacZ L1 VNC, showing dorsal (D) and ventral (V) CG (montage of 2 pictures)

294
In mid stage-17 embryos, the medial CG appear to occupy the four corners of the square flanked by the lon- gitudinal connectives and the posterior and anterior com- missures (Fig. 6F). The anterior and posterior medial CG can be singly identified at this stage (M-CG1 and M-CG2, respectively, see Fig. 2). This distinction is no longer possible after the VNC condensation; we identi- fied only the medial CG cluster at L1 (Fig, 2 and Table 2). Although we could not trace back the medial CG in earlier stages, it is very unlikely that they originate from the midline progenitors. None of the midline-specific markers label any of the channel glia (see also Bossing and Technau 1994).
II: Cortex-associated glia
The "cortex-associated glia" are the cells that lie among the neuronal cell bodies in the cortex. Those that are as- sociated with axonal tracts (e.g. nerve roots) are exclud- ed from this category. Although several subtypes of glia in this category have been described in the adult nervous systems of other insects (Hoyle 1986; Cantera 1993), we found only one subtype in the Drosophila embryonic and early larval VNC: the "cell body gtia" (CBG).
Cell body glia (CBG)
The CBG are scattered among neuronal cell bodies in the lateral and ventral cortex between 10 and 75%NVD (Figs. 8,9,10). There is no CBG in the dorsal cortex. In the abdominal and thoracic segments there are typically 3-4 and 4-5 CBG per hemineuromere, respectively. The positions of the CBG cell bodies in the cortex vary sig- nificantly among individuals as well as between neigh- bouring segments. The nuclei and cell bodies of the CBG have quite irregular shapes (Fig. 3D, G, H). Cytoplasmic protrusions fill the space between neighbouring neuronal cell bodies. Their distal ends are either sharp or with blebs (Fig. 10). Some protrusions are up to 30 g m long, connecting the CBG cell body with the VNC surface and the cortex/neuropile interface (Figs. 8, 9A and 10). In some cases CBG processes are associated with bundles of cell body fibres (Fig. 3A,D).
The medialmost CBG (MM-CBG) lies close to the midline at the centre of the segment (10-20%NML and 50%NAR Figs. 3C and D, 9, 10). The thoracic hemineu- romere has two medialmost CBG instead of one in the abdomen (MM-CBG1 and MM-CBG2; Fig. 9C). The medialmost CBG flank the VUM (ventral unpaired me- dian) neuron cluster on the midline (Fig. 9B) and, in em- bryos, they appear to ensheath the VUM neuron cluster (Fig. 9C). The medialmost CBG, therefore, is identical to the VUM-suppor t cell (Kl~imbt and Goodman 1991; Menne and Klfimbt 1994).
The medial CBG (M-CBG) lies in the ventral cortex lateral to the medialmost CBG (Figs. 3F, 9A, 10). The
position of the medial CBG can vary from 20 to 80%NML. The ventrolateral CBG (VL-CBG) and lateral CBG (L-CBG) lie in the lateral cortex (Fig. 3F-H, 9A, 10). The ventrolateral CBG is often missing.
The same number of CBG are observed at stage 15, when we first detect the CBG in the Mz840 strain. The ventrolateral and lateral CBG lie more close to the ven- trolateral VNC surface than at L1 (Fig. 9C).
III: Neuropile-associated glia
The "neuropile-associated glia" category covers the glial cells that associate with the axonal structures: the nerve roots and the neuropile that includes the connectives and commissures. We classified three subtypes in this catego- ry: the "nerve root glia" (NG) that is further subdivided into the "intersegmental nerve root glia" (ISNG) and the "segmental nerve root glia" (SNG), the "interface glia" (IG), and the "midline gila" (MG),
Nerve root glia (NG: ISNG and SNG)
The nerve root glial cells lie along the nerve roots. The dorsally-running intersegmental nerve root has two ISNG at its medial and lateral ends (Fig. 11A). The me- dial ISNG, or M- ISNG, (ISG1 of Klfimbt and Goodman 1991; SBC of Goodman and Doe 1993), lies at the medi-
Fig. 7 Stereoscopic views of a computer reconstruction from LI VNC of Mz1251/kinesin-lacZ, which labels a subset of SPG, CG, and medial and lateral ISNG (see Fig. 5D,F for the whole-mount staining pattern). Sections from T3 and AI neuromeres are recon- structed. The two membranes represent the outer surface of the VNC and the cortex/neuropile interface, respectively. The white structures show the glial processes labelled by kinesin-lacZ. Mar- bles represent the positions of glial nuclei, coded by colours ac- cording to the glial subtype. The top and middle panels show an oblique view, with and without the glial processes. The bottom panels show the frontal view. The fiat processes from the subset of SPG shown here cover most of the VNC surface. The protrusions are limited only to the surface region. The borders between the processes of neighbouring SPG were hardly identifiable. The neu- rohemal organs (structure above the dorsal midline) are also cov- ered with glial processes, which may derive from either the B-SPG or the dorsal CG. Due to the relatively low resolution along the longitudinal axis, the duct-like structure of the channel is not clear (columns on the midline). Below the SPG processes, the glial protrusions of ISNG are also visible. The medial ISNG lie above the neuropile. The lateral ISNG lie just near the exit points of the peripheral nerves (50%NVD in T3 and 80% in A1)
Fig. 8 Stereoscopic views of a computer reconstruction from L1 VNC of Mz840/kinesin-lacZ, which labels CBG, IG, and MG (see Fig. 9 for the whole-mount staining pattern). Sections from AI and A2 neuromeres are reconstructed. The IG and MG cover a sig- nificant portion of the cortex/neuropile interface, though their cov- erage is not as complete as that of SPG along the CNS surface (Fig. 7). The shape of IG is relatively simple compared to that of the MG; unlike the MG, the IG do not penetrate deep into the neu- ropile (see bottom panel). The CBG have extremely irregular shape and long processes (see also Fig. 10). The flat ends of the CBG processes contact a small part of the VNC surface

295
t" ' -

296
al end of the anterior branch of the intersegmental nerve root (see Fig. 1C), above the lateral neuropile (20-40%NML) at about 50-70%NAE In this region the dorsal cortex is very thin; the medial ISNG contacts both the perineurium and the cortex/neuropile interface (Fig. 3C). The position of the medial ISNG is slightly lateral to the B-SPG (Figs. 4A,C, 5B,C). The medial ISNG sends a process laterally along the nerve root, showing a characteristic triangular morphology (Figs. 5C, 11A, 12). In early embryos the cells lie more posteriorly near the segment border (Fig. 12 A,B), and were hence referred to as the segment boundary cells (SBC; Bastiani and Goodman 1986; Jacobs and Goodman 1989a,b; Goodman and Doe 1993).
The lateral ISNG, or L-ISNG, (ISG2 of Klfimbt and Goodman 1991) lies near the exit point of the nerve root (Fig. 11A). In the L1 the cell sometimes lies just outside of the VNC surface (Fig. 12D, see also arrows in Fig. 4A).
The ventrally-running segmental nerve root has two medial SNG, or M-SNG (SG1 and 2 of Kl~imbt and Goodman 1991), and one lateral SNG, or L-SNG (SG3 of Kl~imbt and Goodman 1991). The two medial SNG lie in tandem along the longitudinal axis (M-SNG1 and M-SNG2), covering several branches of the nerve root (Fig. l lB). Unlike the medial ISNG, which lie at the junction between the nerve root and the connectives, the medial SNG are slightly detached from the neuropile proper.
The lateral SNG lies at the point where the interseg- mental and segmental nerve roots meet to form a single peripheral nerve. As a consequence, the lateral SNG and the lateral ISNG lie in close proximity; sometimes one of them appears to be absent (compare nerves A4 and A5 in Fig. 11C).
Interface glia (IG)
Interface glia are the glial cells that lie at the cortex/neu- ropile interface and send processes along it (Strausfeld 1976; Meyer et al. 1987; "neuropile cover glia" of Can- tera 1993). The embryonic and early larval Drosophila VNC has 7-8 IG per hemineuromere. The "longitudinal glia" (Jacobs and Goodman 1989a,b; Klfimbt and Good- man 1991; Goodman and Doe 1993) are a subset of the IG. In this study we define the longitudinal glia as the progeny of the longitudinal glioblast (Jacobs et al. 1989; Doe 1992) and the interface glia as the generic name of the glial subtype.
The IG nuclei are slightly ellipsoidal. In early embryos the cell bodies are a bit elongated along the longitudinal axis (Jacobs and Goodman 1989b); in larvae the cells have flat cytoplasmic extensions perpendicular to this ax- is, forming a cage-like structure that surrounds the neuro- pile (bottom panel of Fig. 8). The processes only occa- sionally invade the neuropile. The IG cover the ventrolat- eral region of the interface less densely; many bundles of cell body fibres enter the neuropile through this region.
Fig. 9A-C Cell body glia (anterior to the left, bar 50 gm). A L1 VNC of Mz840/kinesin-lacZ, showing the ventral half of the cor- tex that contain 2-3 of the total 3-4 CBG per hemineuromere (MM, M and VL; montage of 3 pictures). B CBG cell bodies la- belled with IacZ at L1 (VUM: ventral unpaired median neurons, montage of 3 pictures). C Mz840/lacZ at late stage 16. Note that the abdominal neuromere (At) contains only one medialmost CBG (MM), while thoracic segments (T3) have two (1 and 2; montage of 3 pictures)

Fig. 10 Schematic 3-D reconstruction of the cell body glia (CBG) in an L1 abdominal neuromere. For clarity, three CBG in the left hemineuromere were omitted
The dorsal interface has up to five dorsal IG (D-IG). Three or four of them lie at the medialmost area of the dorsal interface (0-20%NML, arrowheads in Fig. 13) and separate the neuropile from the pair of rows of big neuronal cell bodies (Fig. 3B-D, F-H). Another one or two dorsal IG lie at the more lateral region of the dorsal interface (204O%NML), which is beneath the thinnest part of the dorsal cortex (Figs. 3F, G and 13C). Their po- sitions vary significantly in the L1 VNC.
The lateral interface (50% NML) has two lateral IG (L-IG; Fig. 13A,D). They form two parallel flat process- es dorso-ventrally along the lateral interface (Figs. 3A, B, H and 13D).
The ventral interface has one ventral IG (V-IG) that lies at the ventralmost region of the interface (15-25%NML) near the centrue of the segment (50% Fig. 11A-C Nerve root glia (anterior to the left, bar 50 pm). A
N ~ P ) , just lateral to the ventral midline glial cells Late stage-15 CNS of Mz1067/lacZ at 90%NVD. The characteris- tic obliquely elongated shape of the medial ISNG (M-ISNG) can
(Fig. 13B). In some cases there are ventra1 IG per be seen (see also Fig. 5C,D). B The same preparation at hemisegment (arrowheads in Fig. 13B), lying along the 60%NVD, a pair of medial SNG (M-SNG1 and M-SNG2, 1 and longitudinal axis. The ventral IG sends processes both 2) can be seen along the SNR. The ISNR and SNR meet, but do
laterally and medially. ~h~ medial process runs towards not fuse, at the exit point from the CNS. A peripheral nerve at this stage consists of two twisted bundles. C Peripheral nerves at early
the glial but does not connect to stage 17 (M&'/lacZ). The two bundles within peripheral nerves them (Fig. 3F). It runs along the inner surface of the neu- fuse until this stage. Both the lateral ISNG and lateral SNG are la- ropile between the anterior and posterior commissures belled along the nerve root A5, while only the lateral ISNG is re- and seems to contact the medial dorsal IG (Fig. 13E,F). vealed along A4. The medialmost exit glial cell (EG) lies along the
The lateral and ventral IG show an interesting migra- peripheral nerve. Note the thin dorsal nerves (dn) running from the neurohemal organ to the lateral body wall
tion during late embryogenesis. At stage 15 all the IG are aligned dorsally in two rows above the longitudinal con- nectives (see Fig. 3 of Menne and Klambt 1994, in pile, passing by the cluster of midline glial cells which the IG are collectively referred to as "longitudinal (Fig. 13E,G). The lateral and ventral IG occupy their fi- glia"). At early stage 16 two cells in the lateral row begin nal positions at early stage 17 (Fig. 13F). The migrating to move laterally, and one in the medial row moves ven- cells leave processes behind and thus maintain a connec- trally along the medial border of the connectives neuro- tion with the remaining dorsal IG (Fig. 13E,F).

298
The longitudinal glioblast (Jacobs et al. 1989) and its progeny (longitudinal glia: LG) express prospero. At stage 14 anti-pros antibody labels six (occasionally five) of the seven or eight IG (per hemineuromere) above the longitudinal connectives. In the late stage-16 and L1 VNC, the two lateral IG, one ventral IG, and two to three of the five dorsal IG are likely to be the longitudinal glia, since they are pros positive (Fig. 16C and D).
Midline glia (MG)
The MG associate with the anterior and posterior com- missure neuropile. They derive from the mesectodermal midline progenitors (Kl~imbt and Goodman 1991; Boss- ing and Technau 1994).
The larval VNC has either three or four MG per neu- romere. They are arranged above and below the neuro- pile at about 50% NAP (Fig. 14B). At embryonic stage 16 the dorsal and ventral MG are still more closely asso- ciated (Fig. 14C). The MG cell bodies are relatively large and round. Their cytoplasmic extensions are re- stricted to the midline, covering the medial part of the commissure neuropile. Fine extensions are also observed within the neuropile (Fig. 14A,C).
Since previous reports described more MG per neuro- mere than we found (four to six in Fredieu and Maho- wald 1989; six in Kl~imbt and Goodman 1991), we care- fully studied MG numbers by using various markers (Fig. 15). They revealed either three or four MG in most of the abdominal segments both at late stage 16 and L1. The percentage of segments with more than four MG was higher in the thorax than in the abdomen.
The AA142 labels up to six cells at stage 13. The pos- terior cells (MGP according to Kl~imbt and Goodman 1991) then migrate ventrally (arrowheads in Fig. 14D) to join the median neuroblast (MNB) progeny (see Bossing and Technau 1994). The staining becomes faint as they move to the ventral cortex and, at late stage 16, they are no longer recognizable. Since other strains and markers labelled no glial cells in this region, it is rather unlikely that the migrated MGP represent glia.
At stage 14 the MG lie on either side of the midline (Fig. 14E). As the longitudinal connectives increase their
Fig. 12A-D Late embryonic development of glial cells (SPG, CG, ISNG, SNG and IG) in the dorsal region of the VNC (Mz317/IacZ, anterior to the left, bar 50 gm). A Stage 14 (mon- tage of 4 pictures). The dorsal CG lie at the posterolateral side of the channels, which are still relatively wide at this stage (aster- isks). A cluster of exit glia (EG) lies near the exit point of the pe- ripheral nerve; they migrate out to the periphery. B Stage 15 (mon- tage of 5 pictures). The wing-like structure of the A-SPG at the dorsal channel opening is apparent already at this stage. The tran- sient labelling of about six IG can be seen; two of them lie just medial to the intersegmental nerve roots with a cluster of four be- tween them. C Late stage 16 (montage of 3 pictures). Note the wing-like structure of the A-SPG and the meshwork structure of the B-SPG. D L1 (montage of 2 pictures). Despite the 50% con- densation of the VNC, the labelled cells have essentially the same distribution as in panel C (the SNG are out of focus)

299
Fig. 13A-G Interface glia (A-D and G: anterior to the left, E and F dorsal to the top, bar 50 ~tm). A L1 dorsal neuropile of Mzl127/lacZ (montage of 3 pictures). About three medial dorsal IG (per hemineuromere) lie at the medial region of the dorsal cor- tex/neuropile interface (D-IG; see also Fig. 3). Some of them lie adjacent to the dorsal MG. Arrows point to the thin IG processes perpendicular to the longitudinal axis. The lateral dorsal IG above the neuropile are out of focus in this composite picture. B Ventral interface of the same specimen (montage of 2 pictures). The ven- tral IG lies lateral to the ventral MG, at the centre of the neuro- mere (50%NAP). Thin IG processes run medio-laterally. In some neuromeres two IG can be seen (arrowheads 1 and 2). C A lateral view of L1 Mzl127/lacZ at 25%NML, showing lateral dorsal IG (D) and the ventral IG (V). D A lateral view of L1 Mz709/lacZ at
50%NML, showing lateral IG (L). E A frontal view of T1 neuro- mere of Mz97/lacZ at early stage 16. Because of the 90-degree bend of the anteriormost VNC (see Fig. 1B), the horizontal view of the T1 is topologically equivalent to the frontal view of more posterior neuromeres. The ventral IG (lO lie between the medial surfaces of the neuropile. The lateral IG (L) are still above the neuropile. F A frontal view of T1 of Mzl127/lacZ at late stage 16. The migration of the ventral IG (V) and lateral IG (L) is slightly advanced compared to panel E. They send processes to contact the dorsal IG (D). Note the spatial relationship between the ventral IG and the MG (arrows). G A horizontal view of abdominal neuro- meres of stage-16 Mzl127/lacZ, showing the migrating ventral IG (V) just adjacent to the MG
width, the M G are a l igned on the mid l ine (Fig. 14F). The M G cel l bod ies r ema in be tween the longi tud ina l connec- tives unti l s tage 16 (Fig. 14C,D). Dur ing condensa t ion o f the V N C and the further growth o f the neuropi le at s tage 17, they move apar t f rom each other to f inal ly occupy pos i t ions above and be low the neuropi le , whi le keep ing
their cy top lasmic extens ions around the commissu re s (Fig. 14A,B). It seems that cel ls choose their f inal dor- sa l /ventral pos i t ions rather r andomly ; more than 60% of the inves t iga ted neuromeres showed a symmet r i ca l distr i- but ion be tween the two clusters (see Fig. 15).
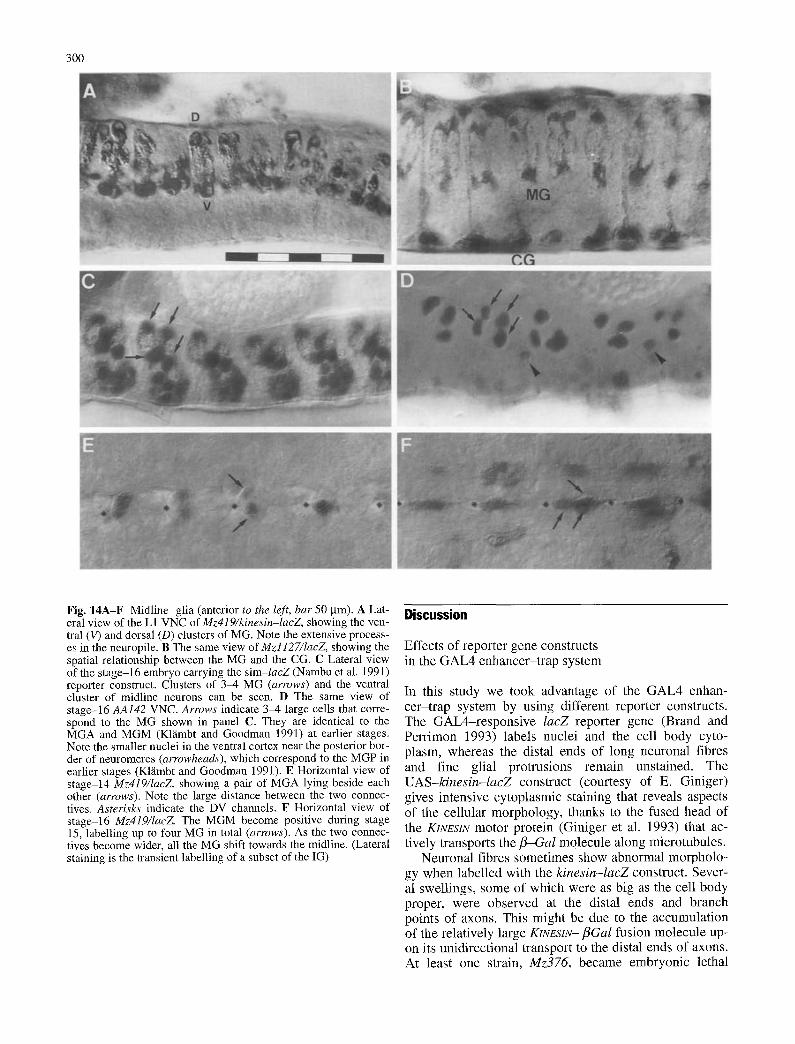
300
Fig. 14A-F Midline glia (anterior to the left, bar 50 gm). A Lat- eral view of the L1 VNC of Mz419/kinesin-lacZ, showing the ven- tral (V) and dorsal (D) clusters of MG. Note the extensive process- es in the neuropile. B The same view of Mzl127/lacZ, showing the spatial relationship between the MG and the CG. C Lateral view of the stage-16 embryo carrying the sim-lacZ (Nambu et al. 1991) reporter construct. Clusters of 3-4 MG (arrows) and the ventral cluster of midline neurons can be seen. D The same view of stage-16 AA142 VNC. Arrows indicate 3-4 large cells that corre- spond to the MG shown in panel C. They are identical to the MGA and MGM (Klfimbt and Goodman 1991) at earlier stages. Note the smaller nuclei in the ventral cortex near the posterior bor- der of neuromeres (arrowheads), which correspond to the MGP in earlier stages (Kl~nbt and Goodman 1991). E Horizontal view of stage-14 Mz419/IacZ, showing a pair of MGA lying beside each other (arrows). Note the large distance between the two connec- tives. Asterisks indicate the DV channels. F Horizontal view of stage-16 Mz419/lacZ The MGM become positive during stage 15, labelling up to four MG in total (arrows). As the two connec- tives become wider, all the MG shift towards the midline. (Lateral staining is the transient labelling of a subset of the IG)
Discussion
Effects of reporter gene constructs in the G A L 4 enhancer- t rap system
In this study we took advantage of the G A L 4 enhan- cer- t rap system by using different reporter constructs. The GAL4- respons ive lacZ reporter gene (Brand and Perr imon 1993) labels nuclei and the cell body cyto- plasm, whereas the distal ends of long neuronal fibres and fine glial protrusions remain unstained. The UAS-kinesin- lacZ construct (courtesy of E. Giniger) gives intensive cytoplasmic staining that reveals aspects of the cellular morphology, thanks to the fused head of the KINESI~V motor protein (Giniger et al. 1993) that ac- tively transports the f l-GaI molecule along microtubules.
Neuronal fibres sometimes show abnormal morpholo- gy when labelled with the kinesin-lacZ construct. Sever- al swellings, some of which were as big as the cell body proper, were observed at the distal ends and branch points o f axons. This might be due to the accumulat ion o f the relatively large KINESIN-- flGal fusion molecule up- on its unidirectional transport to the distal ends of axons. At least one strain, Mz376, became embryonic lethal

Thorax AA142
, St. 1 6 (N=18, n=53) 7 6 6
5 5
4 4 3 3
01~ ....... 4.0 0 ' 10 20 30 40 50 60 % 10
sim-lacZ N St. 16 (N=29, n=87)
5
; 4.25 16 20 30 40 50 66 %
L1 ( N = 3 9 , n = 1 1 5 )
20 30 40 56 %
o' : L1 (,=28,,=8o) 5
4 ;-;;-; ; .............
; I 3.79 10 20 30 45 56 %
Mz 685
n.d.
7 6 3
4 3 'p 1 o 3.84
10 20 30 40 50 %
Mz 419 ' L St. 16 (N=21,.=59) 7 L1 (N=38, n=114)
/
6 ~ 6 5 5
4 ~!i!iii!ii!i!!i;ili;iii;iiii;i!i;iiiiii;iiii!::!il 4
2
I
o 3.05 16 3.57 10 20 30 40 50 66 % 10 20 30 40 50 %
Fig. 15 Frequency distribution of the number of midline glial cells (MG) per neuromere. The numbers of MG counted from the thoracic and abdominal segments were pooled. N and n indicate the number of specimens and the neuromeres counted, respective- ly. The number at the bottom-right of each panel indicates the av- erage MG number per neuromere. In the embryonic Mz685 the in- spection of MG was not possible because of dense epidermal staining. AA142 is a P-lacZ enhancer-trap strain (KlSxnbt and Goodman 1991), and sim-lacZ is a transformant strain with lacZ under the single minded promoter (Nambu et al. 1991). The GAL4 strains (Mz685 and Mz419) revealed fewer cell numbers more of- ten than the other two markers (see discussion). For L1, cells in the dorsal and ventral clusters were counted separately; the black bar indicates the percentage of neuromeres with three or more MG in either of the two clusters. Less than 40% of the investigated neuromeres had two cells in both positions ("2+2 cells", gray bar). The "2+1" combination was as common as the "2+2". The "3+1" or "4+0" combination occurred in 5-25% of the investigated neu- romeres ("others", black bar). The probability of having more cells in the dorsal and less in the ventral position was essentially the same as vice versa
when crossed with the UAS-kines in- lacZ strain, al- though it was viable when crossed with the UAS- lacZ flies. However, we found no abnormalit ies in other types o f cells labelled with the kinesin-IacZ, such as glia, epi- dermis, muscles, intestine and salivary gland cells. The aspects o f glial morpho logy revealed with the kine- sin-lacZ construct were also confirmed with other cyto-
Abdomen 67 ~ St. 16(N=lS, n=142)
4 ~ ii!iii~iiiii!i~iiii!ii!il;!i;i!i!ili;iliJ 3 iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiSiiiiiiiiiiiililiJliiii
"0 ................... 3 .56
L1 (N=39, n=312) 6 o the rs 2+2 c e l l s
5
4 3
2
1 0
301
52 10 20 30 40 50 60 % 10 20 30 40 50 %
St. 16 (N=29, n=229)
3.76
67 = , =
5
4 3
; 3.41 10 20 30 40 50 60 % 10 20 30 40 50 %
n.d.
7 L1 (N=33, n=263) 6
5 I 4
1 o 3.31
10 25 30 40 50
7 St. 16 (N=24, n=188) 7 L1 (.=3e, .=304) 6 J 6
° iiiiiiiiiiiii!jiiiiiiiii!ii: !iiiii . . . . . . . . . . . . . . . . .
4 4 ~ili;ii:i~it
10 20 30 40 50 60 % 10 20 30 40 50 %
plasmic markers as well as with semi-thin sections (com- pare Fig. 14A, B and C, for example).
Al though transient expression was observed in some cells, the perdurance of ]3 Gal expressed in the G A L 4 lines was generally rather long. In the Mz360 strain, for example, lacZ was detectable even at L1, which is more than 12 h after the G A L 4 m R N A expression ends at stage 14. This feature was vital for our purpose to follow the fate and migrat ion o f labelled cells during late em- bryogenesis. The long perdurance may be due to the de- lay in the two-s tep gene activation mechanism; the G A L 4 protein may well continue to activate the UAS even after the decline o f the enhancer activity. The UAS-kinesin- lacZ construct showed shorter perdurance than the UAS-IacZ; the active transport o f the protein may remove the marker quickly f rom the cell body.
As is shown in Fig. 15 for the midline glia, G A L 4 strains tend to reveal fewer cells as compared to the con- ventional P-lacZ enhancer- t rap strains and other mark- ers. It seems that the G A L 4 transcription factor - UAS promoter combinat ion can sometimes fail to activate the reporter gene; in this case the cells exist but some of them lack the reporter protein.
Only a few of our G A L 4 strains show staining pat- terns in early embryos. This may also be due to the de- lay o f lacZ expression by the two-s t ep promoter interac- tion.
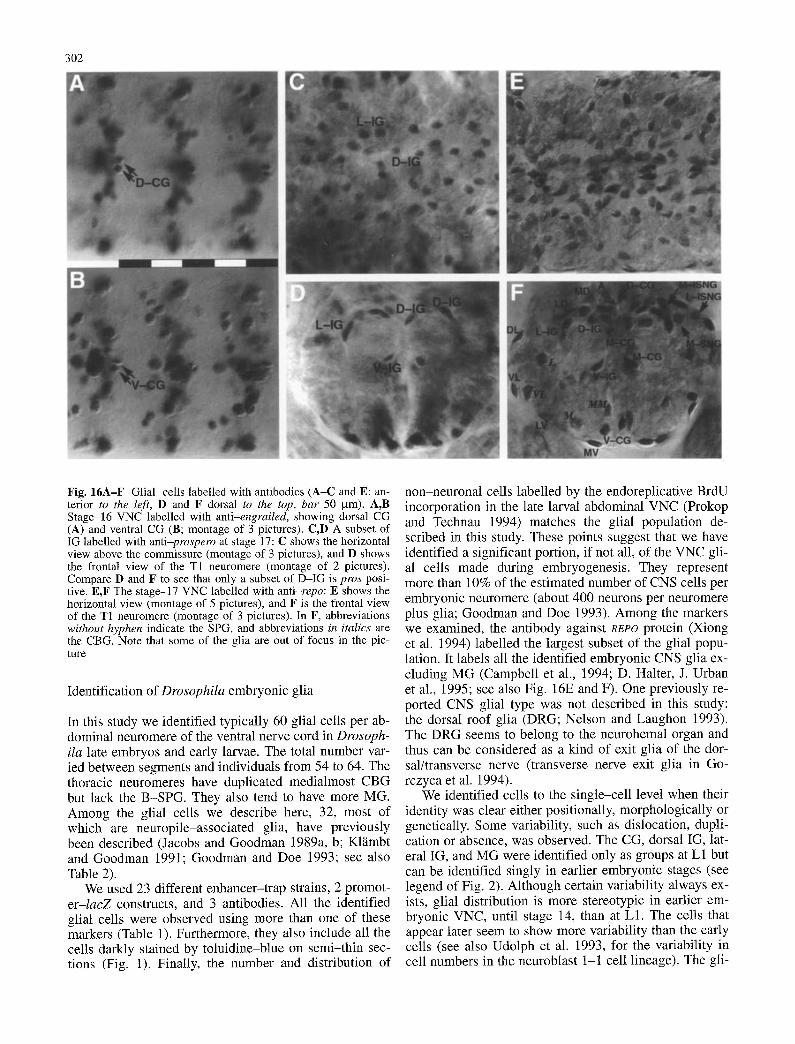
302
Fig. 16A-F Glial cells labelled with antibodies (A-C and E: an- terior to the left, D and F dorsal to the top, bar 50 gm). A,B Stage-16 VNC labelled with anti-engrailed, showing dorsal CG (A) and ventral CG (B; montage of 3 pictures). C,D A subset of IG labelled with anti-prospero at stage 17: C shows the horizontal view above the commissure (montage of 3 pictures), and D shows the frontal view of the T1 neuromere (montage of 2 pictures). Compare D and F to see that only a subset of D-IG is pros posi- tive. E,F The stage-17 VNC labelled with anti-repo: E shows the horizontal view (montage of 5 pictures), and F is the frontal view of the T1 neuromere (montage of 3 pictures). In F, abbreviations without hyphen indicate the SPG, and abbreviations in italics are the CBG. Note that some of the glia are out of focus in the pic- ture
Identification of Drosophila embryonic glia
In this study we identified typically 60 glial cells per ab- dominal neuromere of the ventral nerve cord in Drosoph- ila late embryos and early larvae. The total number var- ied between segments and individuals from 54 to 64. The thoracic neuromeres have duplicated medialmost CBG but lack the B-SPG. They also tend to have more MG. Among the glial cells we describe here, 32, most of which are neuropile-associated glia, have previously been described (Jacobs and Goodman 1989a, b; Kl~imbt and Goodman 1991; Goodman and Doe 1993; see also Table 2).
We used 23 different enhancer-trap strains, 2 promot- er- lacZ constructs, and 3 antibodies. All the identified glial cells were observed using more than one of these markers (Table 1). Furthermore, they also include all the cells darkly stained by toluidine-blue on semi-thin sec- tions (Fig. 1). Finally, the number and distribution of
non-neuronal cells labelled by the endoreplicative BrdU incorporation in the late larval abdominal VNC (Prokop and Technau 1994) matches the glial population de- scribed in this study. These points suggest that we have identified a significant portion, if not all, of the VNC gli- al cells made during embryogenesis. They represent more than 10% of the estimated number of CNS cells per embryonic neuromere (about 400 neurons per neuromere plus glia; Goodman and Doe 1993). Among the markers we examined, the antibody against REPO protein (Xiong et al. 1994) labelled the largest subset of the glial popu- lation. It labels all the identified embryonic CNS glia ex- cluding MG (Campbell et al., 1994; D. Halter, J. Urban et al., 1995; see also Fig. 16E and F). One previously re- ported CNS glial type was not described in this study: the dorsal roof glia (DRG; Nelson and Laughon 1993). The DRG seems to belong to the neurohemal organ and thus can be considered as a kind of exit glia of the dor- sal/transverse nerve (transverse nerve exit glia in Go- rczyca et al. 1994).
We identified cells to the single-cell level when their identity was clear either positionally, morphologically or genetically. Some variability, such as dislocation, dupli- cation or absence, was observed. The CG, dorsal IG, lat- eral IG, and MG were identified only as groups at L1 but can be identified singly in earlier embryonic stages (see legend of Fig. 2). Although certain variability always ex- ists, glial distribution is more stereotypic in earlier em- bryonic VNC, until stage 14, than at L1. The cells that appear later seem to show more variability than the early cells (see also Udolph et al. 1993, for the variability in cell numbers in the neuroblast 1-1 cell lineage). The gli-

al distribution in the imaginal wing also shows high vari- ability (Giangrande et al. 1993). On the other hand, cer- tain glial cells show significantly lower variability than others even in later stages (such as medialmost CBG), suggesting the importance of the consistent position and number for their function.
Classification of insect glia
We gave "unique names" to the identified cells according to their positions and subtypes (Fig. 2). The names are specific for Drosophila embryonic and early larval VNC; they are not directly applicable to other CNS regions, stages, and species because of the different number and distribution of glial cells in these cases. The "subtypes" and "categories", on the other hand, should easily be ap- plicable to other CNS regions and stages, as well as to other insect and arthropod species (Table 3). Without al- tering the framework, subtypes and categories can ac- commodate more cell types than proposed in this study, such as further glia in the late larval and adult CNS. Among the gtial cells in the Drosophila optic lobe (Win- berg et al. 1992), for example, it is likely that the satellite glia belongs to the cortex-associate glia while the others are various types of neuropile-associated glia (see Table 1 for the VNC staining pattern of the 3-109 strain, which was used to reveal the optic lobe glia).
The three different glial categories can clearly be ad- dressed in the lateral and ventral region of the VNC, where prominent cortex separates the neuropile and the CNS surface. The difference between the surface-associ- ated and neuropile-associated glia is less obvious in the dorsal VNC, where the cortex is very thin and glial cells (A- and B-SPG, medial ISNG, and dorsal IG) contact both the neuropile and the CNS surface.
There have been debates and confusion about the def- inition of glia. For example, the parts of the tracheal sys- tem that invade the CNS have been assigned to the glial population ("tracheal glia", Hoyle 1986). The tracheal branches in the CNS, however, are structurally and func- tionally contiguous with the outside tracheal system. In analogy to the capillary cells in the vertebrate brain, we excluded the tracheal cells from the glial population.
The classification of the outermost cell layer of the CNS, the perineurial glia (PNG), is also a matter of de- bate. It has been assumed to be of mesodermal origin (Bauer 1904; Scharrer 1939) and was reported to be ab- sent in twist mutant embryos that lack the mesodermal cells ("sheath cells", Edwards et al. 1993). The ul- tra-structural characteristics of these cells are rather dif- ferent from the other glial cells (Hoyle 1986). Strausfeld (1976), Hoyle (1986) and Edwards et al. (1993), there- fore, excluded these cells from the glial population. Wig- glesworth (1959), Carlson and Saint Marie (1990) and Cantera (1993), on the other hand, included them in their glial description. We incorporate the perineurial glia in the glial population regardless of their origin and feature, since they form a distinct non-neuronal part of the insect
303
CNS and probably function with other glial cells to sup- port neurons.
However, none of our enhancer-trap strains labelled cells that are likely to be the PNG. Using electron mi- croscopy, the Drosophila perineurial layer becomes de- tectable from stage 17 onwards (Tepass and Hartenstein 1994). In the blowfly Calliphora etythrocephala the layer is formed even later (Lane and Swales 1978). The ultrastructural characteristics of glial cells along the embryonic VNC outer surface resemble the definition of subperineurial glia by Hoyle (1986) more than that of perineurium cells (J. Roger Jacobs, personal commu- nication). It is possible that PNG show only immature morphology in late embryonic stages and at L1, making it impossible to distinguish them from the SPG at a light microscopic level. In this case, some of the cells we classified as the SPG might actually be the PNG. Another possibility is that we missed the PNG because we performed the first screening using whole-mount embryos, which can detect GAL4 expression in the CNS only until late stage 16. In the third-instar larvae and imago, markers such as the M84 enhancer-trap strain (Kl~imbt and Goodman 1991) and the anti-repo antibody (courtesy of D. Halter and A. Travers) la- belled at least two types of surface-associated glia (un- published observation). The smaller, more abundant cells, which often lie above the SPG, may correspond to the PNG.
Comparison of previous classifications
Table 3 summarizes some of the previous glial classifica- tions since the first description of insect glia by Ram6n y Cajal and S~inchez y Sfinchez (1915). While the glial classifications in other insects are mainly based on the observation of larval and/or adult CNS, most of the Dro- sophila CNS glial cells known so far have been identi- fied in the embryo. The diversity of glial types seems to be higher in the adult CNS than in larvae (Cantera 1993), which have similar sets of glia to embryos.
Hoyle (1986) and Cantera (1993) described several cells we did not find in the Drosophila embryonic VNC. In the cortex region of large insects, large motoneurons have more than one glial cell dedicated to their cell body: satellite glia (SAG1, Hoyle) or cortical type 2 (C2, Cantera). The lack of this glial subtype in the early larval Drosophila CNS may be due to its much smaller size. The FETi (fast extensor tibiae) motoneuron of the adult locust is 95 gm in diameter, surrounded by 30 gm of gli- al coat (Hoyle 1986). In comparison, the whole single neuromere of first-instar larval Drosophila is only 80 gm wide and 13 gm thick. The same dimensional dif- ference may explain the lack of the glial glia (G1G1, Hoyle 1986), cells that contact only other glial cells, and the transport glia (TpG1, Hoyle 1986), a subtype of sur- face-associated glia.
The moth neuropile cover glia type 1 (NC 1) of Can- tera (1993) correspond to our interface glia. For the

304
Table 3 Summary of the glial classification and comparison with previous studies. The left part shows our proposal of the glial classification. The subtypes written in italics represent the cells we did not identify in this study. Based on the study of the adult CNS, one or more subtypes may be added in the cortex-as- sociated glial category (shown here as a dash). The right part compares some of the previous classifications fitted into our
classification hierarchy. Names written in italics indicate glial types without corresponding cells found in our study (see discus- sion). Wigglesworth (1959), Nordlander and Edwards (1969), Sohal et al. (1972), and Stransfeld (1976) classified glia into a few large classes. Meyer et al.'s classification (1987) is unique in that it is based on antibody staining patterns (antibody names shown in parentheses). Hoyle defined subtypes in more detail.
Our propgsal (1995, Drosophila embryo and early larva, VNC, enhancer trap, promoter-lacZ, antibody)
Glial Category Glial Subtype CNS glia Surface-
associated Glia
Cortex- associated Glia
Neuropile- associated Glia
PNS glia Nerve- associated Glia
Sensory Organ Glia
Ramdn y Cajal and S~inchez y Sfinchez (1915, Musca adult, retina and brain, Golgi / silver nitrate)
Wigglesworth (1959, Rhodnius late larva, VNC, ethyl ganate)
Nordlander and Ed- wards (1969, Danaus, larva and adult, brain, toluidine blue)
perineurial glia ( P N G ) - - type i glial I
subperineurial glia (SPG)
channel glia /CG} cell body glia (CBG)
nerve root glia (NG)
interface glia (IG)
midline glia (NG} neuropilar glia (NPG)
epithelial cells (?)
neuroglia cells radiate glia, displaced epithelial cells
perineurial ~lia exit glia (EG) peripheral glia (PG)
type iii
type ii
type iv
Tracheae tracheae
glial V
glial III
glial II
glial IV
Sohal et al. (1972, Musca, adult, brain, EM)
m
type I
type II
type III
adult-specific NC 2, 3 and 4, and NC5 that exists both in Manduca larvae and imago, we found no corresponding cells in Drosophila larvae.
In the adult CNS, neuropilar glia (NPG) have been described that reside within the neuropile (Sohal et al. 1972; Strausfeld 1976; Hoyle 1986; Meyer et al. 1987; Cantera 1993). We found no strains and markers that la- bel such glia. Semi-thin sections revealed no NPG cell bodies in early larval VNC (Fig. 3), although we found a few glial nuclei in the brain neuropile. The existence of the NPG in the embryonic and early larval Drosophila VNC, therefore, is rather unlikely, as is also the case in Manduca larvae (Cantera 1993). Using Manduca pupae, Tolbert and Oland (1989, 1990) showed that glial cells in the antennal lobe remain at the cortex/neuropile interface until early stages of metamorphosis, when they migrate inward to delineate the antennal lobe glomeruli. This suggests that the embryonic interface glia later differen- tiate into further sub-subtypes, some of which represent neuropilar glia.
The MG have not been described in other insects, al- though it is unlikely that the VNC of other insects do not have MG. The CG has not been described, possibly be- cause insects with separate ganglia, as well as Drosophi-
la early embryos (before stage 15), do not have the ap- parent channel structure. The DV channel is equivalent to the empty space flanked by the parallel longitudinal connectives and neighbouring neuromeres in primitive insects. This relationship suggests that Drosophila chan- nel glia may correspond to the surface-associated glia along the connectives in these insects.
It is possible that there is essentially no difference be- tween the exit glia (EG) and the peripheral glia (PG) classified by Kl~imbt and Goodman (1991). During stage 14 and 15 the exit glial cells lie at the exit point of the peripheral nerve, while the peripheral glial cells lie out- side the CNS (Fig. 12A). Before stage 14, however, most peripheral glial cells lie within the CNS, and all the exit glial cells migrate out of the CNS after stage 15. The two terms may only describe a transient situation. The mono- clonal antibody 5B12 shows no difference between exit glia and peripheral glia (Meyer et al. 1987; Klfimbt and Goodman 1991). The outward migration of PNS glial cells seems to be rather common; it is also observed for glial cells along nerves in the developing wing and optic stalk (Giangrande et al. 1993; Giangrande 1994; Choi and Benzer 1994). These glial cells however, do not originate from the CNS. Similarly, some of the embryon-

Cantera made a two-level hierarchical system, using descriptive subtype names followed by a numerical suffix for sub-subtypes. Edwards et al. deal only with glia along the CNS surface. The classification of Klfimbt and Goodman is unique in that it is based on singly identified glial cells. Their nomenclature is slightly modified by Goodman and Doe (1993, see Table 2). The names and genus of insects listed here are: Musca domestica:
305
Common House-fly (Diptera), Rhodnius prolixus: Wheel Bug (Hemiptera), Danaus plexippus plexippus: Monarch Butterfly (Lepidoptera), Acheta domesticus: House Cricket (Orthoptera), Schistocerca americana: Locust (Orthoptera), Manduca sexta: Hawk Moth (Lepidoptera), and Drosophila melanogaster: Fruit-fly (Diptera)
Strausfeld (1976, Musca adult, brain, Golgi / silver nitrate)
Meyer et al. (1987, Acheta adult, VNC, antibody)
' Hoyle (1986, Schistocerca adult, VNC, EM)
Cantera (1993, Manduca larva and adult, VNC, toluidine blue)
Edwards et al. (1993, Drosophila embryo, VNC, EM)
Kl~mbt and Good- man (1991, Drosophila embryo, VNC, enhancer trap)
perineural cells perineurial glia perineurium cells ( P n ) perineurial type 1 (P1) perineurial - - (3F6) sheath cells (SC)
- - perineurial type 2 (P2) barrier glia class 1 (perineural- perikaryal glia)
cortical glia (1D7, 1E7, 6D7)
class 2
subperineurial glia (SuPnGI) transport glia ('I'pGO
cell body glia (CbGI)
satellite glia (SAGO ~lial ~lia (GIGI I
axon-hilloek glia (AxHGI)
- - tract glia (5B12)
cortical type 1 (Cl)
cortical type 2 (C2)
neuropile cover type 6 (NC6)
neuropile cover type 1, 2, 3, 4, 5 (NC1-5 )
class 2 (interface glia)
interface glia (5B12)
A glia B glia
VUM support cell
nerve root glia (ISG, SG)
longitudinal glia (LG)
- - - - midline glia (MG) class 3 neuropile glia dark neuropilar glia (DNpGI) neuropile glia (N1, N4) - - (neuropilar glia) (3G6, 8BI1) light neuropilar glia (LNpGI) tract glia (N2, N3) class 4
- - lemnoblasts peripheral glia exit glia (EG) (5B12) peripheral glia (PG)
tracheae tracheal glia (TrGI) - -
ic PG also seem to emerge outside the CNS, although their origin is still unknown (D. Halter, J. Urban et al., 1995).
Structural comparison between insect and vertebrate glia
Four major classes of glia exist in the vertebrate CNS. The ependymal cells cover the outer surface of the ner- vous system and are quite similar to the surface-associ- ated glial category in insects.
The astrocyte glia have irregular-shaped cell bodies with many branched processes, which contact both with neurons and capillary vessels. They are characterized by the existence of glial fibrillary acid protein (GFAP, Eng and DeArmond 1982). Although insect glia lack interme- diate filaments like the GFAP (Carlson and SaintMarie 1990), the CBG shares many characteristics with the as- trocyte glia.
With regard to the distribution and morphology, the neuropile-associated giia are comparable to the oligo- dendrocytes, although insect neurons lack the myelin sheath. The separation of neuronal cell bodies in the cor- tex and the dendritic structure in the neuropile may ex-
plain the separate d is t r ibut ion of a s t rocy t e - l i ke and oli- g o d e n d r o c y t e - l i k e g l ia in the insect CNS.
Mic rog l i a are the smal les t ver tebra te glia, which func- t ion as phagocytes . They are cons ide red to be o f meso - dermal origin, and hence are often exc luded f rom the ca tegory o f neurogl ia . A l though cell death is a ubiqui- tous p h e n o m e n o n in the embryon ic Drosoph i la CNS, no phagocy tes were obse rved in the CNS p rope r (Abrams et al. 1993). Whe the r cel ls such as C B G or SPG funct ion as mic rog l i a remains to be invest igated.
The comprehens ive documenta t ion o f ident i f ied gl ia l cel ls made in the Drosoph i la embryon ic CNS prov ides us with an impor tan t bas is for s tudying the or ig in and deve lopmen t o f gl ia as wel l as g i l a - n e u r o n in teract ions during embryon ic neurogenes i s by means o f mutants , mo lecu la r genet ic and cel lu lar exper imenta l tools. It a lso serves as a start ing poin t to invest igate the pos t embryon- ic deve lopmen t of the Drosoph i la glia.
Acknowledgements We thank A. Brand (pGawB, UAS-lacZ), S. Crews (sim-lacZ and slit-lacZ), E. Giniger (UAS-kinesin-lacZ), D. Halter and A. Travers (anti-repo 4cG), C. Kl~mbt (AA142, M84, PIO1, 3-109), and E. Spana and C. Q. Doe (anti-pros) for provid- ing us with fly stocks and antibodies. We are grateful to A. Brand for critically reading the manuscript, and to R. Cantera, V. Harten-

306
stein, A. Prokop, J. R. Jacobs, and the members of our research group for helpful discussions. This study was supported by post~ doctoral fellowships from the Yamada Foundation and the EMBO to K.I. and by the DFG grant Te130/4-1 and the EEC grant SCI- CT92-0790 to J.U. and G.M.T.
References
Abrams JM, White K, Fessler LI, Steller H (1993) Programmed cell death during Drosophila embryogenesis. Development 117:29-43
Bastiani MJ, Goodman CS (1986) Guidance of neuronal growth cones in the grasshopper embryo. III. Recognition of specific glial pathways. J Neurosci 6:3542-3551
Bauer V (1904) Zur inneren Metamorphose des Zentralnervensys- terns der Insekten. Zool Jahrb Abt Anat Ontog Tiere 20:123-150
Bodmer R, Jan YN (1987) Morphological differentiation of the embryonic peripheral neurons in Drosophila. Roux's Arch Dev Biol 196:69-77
Bossing T, Technau GM (1994) The fate of the CNS midline pro- genitors in Drosophila as revealed by a new method for single cell labelling. Development 120:1895-1906
Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cell fates and generating dominant pheno- types. Development 118:401-415
Buchanan RL, Benzer S (1993) Defective glia in the Drosophila brain degeneration mutant drop-dead. Neuron 10:839-850
Campbell G, G6ring H, Lin T, Spana E, Andersson S, Doe CQ, Tomlinson A (1994) RK2, a glial-specific homeodomain protein required for embryonic nerve cord condensation and viability in Drosophila. Development 120:2957-2966
Campos-Ortega JA, Hartenstein V (1985) The embryonic develop- ment of Drosophila melanogaster. Springer, Berlin Heidelberg New York
Cantera R (1993) Glial cells in adult and developing prothoracic ganglion of the hawk moth Manduca sexta. Cell Tissue Res 272:93-108
Carlson SD, SaintMarie RL (1990) Structure and function of in- sect glia. Annu Rev Entomol 35:597-621
Choi K-W, Benzer S (1994) Migration of glia along photorecep- tor axons in the developing Drosophila eye. Neuron 12:423-431
Doe CQ (1992) Molecular markers for identified neuroblasts and ganglion mother cells in the Drosophila central nervous sys- tem. Development 116:855-863
Doe CQ, Chu-LaGraff Q, Wright DM, Scott MP (1991) The pros- pero gene specifies cell fates in the Drosophila central nervous system. Cell 65:451-464
Ebens AJ, Garren H, Cheyette BNR, Zipursky SL (1993) The Dro- sophila anachronism locus: a glycoprotein secreted by glia in- hibits neuroblast proliferation. Cell 74:15-27
Edwards JS, Swales LS, Bate M (1993) The differentiation be- tween neuroglia and connective tissue sheath in insect ganglia revisited: the neural lamella and perineurial sheath cells are absent in a mesodermless mutant of Drosophila. J Comp Neu- rol 333:301-308
Eng LF, DeArmond SJ (1982) Immunocytochemical studies of as- trocytes in normal development and disease. Adv Cell Neuro- biol 3:145-171
Fredieu JR, Mahowald AP (1989) Glial interactions with neurons during Drosophila embryogenesis. Development 106:739-748
Giangrande A (1994) Glia in the fly wing are clonally related to epithelial cells and use the nerve as a pathway for migration. Development 120:523-534
Giangrande A, Murray MA, Palka J (1993) Development and or- ganization of glial cells in the peripheral nervous system of Drosophila melanogaster. Development 117:895-904
Giniger E, Jan LY, Jan YN (1993) Specifying the path of the inter- segmental nerve of the Drosophila embryo: a role for Delta and Notch. Development 117:431-440
Goodman CS, Doe CQ (1993) Embryonic development of the Drosophila central nervous system. In: Bate M, Martinez- Arias A (eds) The development of Drosophila melanogaster, vol II. Cold Spring Harbor Laboratory Press, Cold Spring Har- bor, pp 1131-1206
Gorczyca MG, Phillis RW, Budnik V (1994) The role of tinman, a mesodermal cell fate gene, in axon pathfinding during the de- velopment of the transverse nerve in Drosophila. Development 120:2143-2152
Gray GE, Sanes JR (1992) Lineage of radial glia in the chicken optic tectum. Development 114:271-283
Halter DA, Urban J, Rickert C, Ner SS, Ito K, Travers AA, Technan GM (1995) The homeobox gene repo is required for the differentiation and maintenance of glia function in the embryonic nervous system of Drosophila melanogaster. Development in press
Hatten ME (1990) Riding the glial monorail: a common mecha- nism for glial-guided neuronal migration in different regions of the developing mammalian brain. Trends Neurosci 13:179-184
Hertweck H (1931) Anatomie und Variabilit~it des Nervensystems und der Sinnesorgane yon Drosophila melanogaster (Meigen). Z Wiss Zool Abt A 139:559-663
Hoyle G (1986) Glial cells of an insect ganglion. J Comp Neurol 246:85-103
Ito K, Hotta Y (1992) Proliferation pattern of postembryonic neu- roblasts in the brain of Drosophila meIanogaster. Dev Biol 149:134-148
Jacobs JR (1993) Perturbed glial scaffold formation precedes axon tract malformation in Drosophila mutants. J Neurobiol 24:611-626
Jacobs JR, Goodman CS (1989a) Embryonic development of axon pathways in the Drosophila CNS. I. A glial scaffold appears before the first growth cones. J Neurosci 9:2402-2411
Jacobs JR, Goodman CS (1989b) Embryonic development of axon pathways in the Drosophila CNS. II. Behaviour of pioneer growth cones. J Neurosci 9:2412-2422
Jacobs JR, Hiromi Y, Patel NH, Goodman CS (1989) Lineage, migration, and morphogenesis of longitudinal glia in the Dro- sophila CNS as revealed by a molecular lineage marker. Neu- ron 2:1625-1631
Klfimbt C, Goodman CS (1991) The diversity and pattern of glia during axon pathway formation in the Drosophila embryo. Glia 4:205-213
Klfimbt C, Jacobs JR, Goodman CS (1991) The midline of the Drosphila central nervous system: A model for the genetic analysis of cell fate, cell migration, and growth cone guidance. Cell 64:801-815
Lane NJ, Swales LS (1978) Changes in the blood-brain barrier of the central nervous system in the blowfly during development, with special reference to the formation and disaggregation of gap and tight junctions. 1. Larval development. Dev Biol 62:389-414
Menne TV, Klfimbt C (1994) The formation of commissures in the Drosophila CNS depends on the midline cells and on the Notch gene. Development 120:123-133
Meyer MR, Reddy GR, Edwards JS (1987) Immunological probes reveal spatial and developmental diversity in insect neuroglia. J Neurosci 7:512-521
Nambu JR, Lewis JO, Wharton KA, Crews ST (1991) The Dro- sophila single-minded gene encodes a helix-loop-helix protein that acts as a master regulator of CNS midline development. Cell 67:1157-1167
N~issel DR, Ohlsson LG, Cantera R (1988) Metamorphosis of identified neurons innervating thoracic neurohemal organs in the blowfly: Transformation of cholecystokioninlike immuno- reactive neurons. J Comp Neurol 267:343-356
Nelson HB, Laughon A (1993) Drosophila glial architecture and development: analysis using a collection of new cell-specific markers. Roux's Arch Dev Biol 202:341-354
Nordlander RH, Edwards JS (1969) Postembryonic brain develop- ment in the monarch butterfly Danaus plexippus plexippus, L.: I. Cellular events during brain morphogenesis. Roux's Arch Dev Biol 162:197-217

Patel NH, Martin-Blanco E, Coleman KG, Poole S J, Ellis MC, Kornberg TB, Goodman CS (1989) Expression of engrailed proteins in arthropods, annelids, and chordates. Cell 58:955-968
Prokop A, Technau GM (1991) The origin of postembryonic neu- roblasts in the ventral nerve cord of Drosophila melanogaster. Development 111:79-88
Prokop A, Technau GM (1993) Cell transplantation. In: Hartley DA (ed) Cellular interactions in development: a practical ap- proach. Oxford University Press, Oxford New York Tokyo, pp 33-57
Prokop A, Technau GM (1994) BrdU incorporation reveals DNA replication in non dividing glial cells in the larval abdominal CNS of Drosophila. Roux's Arch Dev Biol 204:54-61
Ramon y Cajal S, Sfinchez y Sfinchez D (1915) Contribuction al conocrimiento de los centros nerviosos de los insectos. Parte I. Retina y centros opticos. Trab Lab Invertebr Biol Univ Madrid 13:1-168
Robertson HM, Preston CR, Phillis RW, Johnson-Schlitz D, Benz WK, Engels WR (1988) A stable genomic source of P element transposase in Drosophila melanogaster. Genetics 118:461-470
Rothberg MJ, Hartley DA, Walther Z, Artavanis Tsakonas S (1988) slit: An EGF-homologous locus of D. melanogaster in- volved in the development of the embryonic central nervous system. Cell 55:1047-1059
Scharrer BC (1939) The differentiation between neuroglia and connective tissue sheath in the cockroach Periplaneta ameri- cana. J Comp Neurol 70:77-88
Seeger M, Tear G, Ferres-Marco D, Goodman CS (1993) Muta- tions affecting growth cone guidance in Drosophila: genes necessary for guidance toward or away from the midline. Neu- ron 10:409-426
Singer M, Norlander RH, Egar M (1979) Axonal guidance during embryogenesis and regeneration in the spinal cord of the newt:
307
the blueprint hypothesis of neural pathway patterning. J Comp Neurol 185:1-22
Sohal RS, Sharma SR Couch EF (1972) Fine structure of the neu- ral sheath, glia and neurons in the brain of the housefly, Musca domestica. Z Zellforsch 135:449-459
Strausfeld NJ (1976) Atlas of an insect brain. Springer, Berlin Heidelberg New York
Tepass U, Hartenstein V (1994) The development of cellular junc- tions in the Drosophila embryo. Dev Biol 161:563-596
Tolbert LR Oland LA (1989) A role for glia in the development of organized neuropilar structures. Trends Neurosci 12:70-75
Tolbert LR Oland LA (1990) Glial cells form bounderies for de- veloping insect olfactory glomeruli. Exp Neurol 109:19-28
Truman JW, Bate M (1988) Spatial and temporal patterns of neu- rogenesis in the central nervous system of Drosophila melano- gaster. Dev Biol 125:145-157
Udolph G, Prokop A, Bossing T, Technau GM (1993) A common precursor for glia and neurons in the embryonic CNS of Dro- sophila gives rise to segment-specific lineage variants. Devel- opment 118:765-775
White K, Kankel DR (1978) Patterns of cell division and cell movement in the formation of the imaginal nervous system in Drosophila melanogaster. Dev Biol 65:296-321
Wigglesworth VB (1959) The histology of the nervous system of an insect, Rhodnius prolixus (Hemiptera). II. The central gan- glia. Quart J Micr Sci 100:299-313
Winberg ML, Perez SE, Steller H (1992) Generation and early dif- ferentiation of glial cells in the first optic ganglion of Dro- sophila melanogaster. Development 115:903-911
Xiong W, Okano H, Patel NH, Blendy JA, Montell C (1994) repo encodes a glial-specific homeo domain protein required in the Drosophila nervous system. Gene Dev 8:981-994




















