
Coherent structures and larval transport in the California ...
145
UNIVERSITY OF CALIFORNIA SANTA CRUZ Coherent structures and larval transport in the California Current system A dissertation submitted in partial satisfaction of the requirements for the degree of DOCTOR OF PHILOSOPHY in EARTH SCIENCES by Cheryl S. Harrison December 2012 The Dissertation of Cheryl S. Harrison is approved: Professor Eli A. Silver, Chair Professor David A. Siegel Research Scientist Adina Payton Professor Pete Raimondi Tyrus Miller Vice Provost and Dean of Graduate Studies
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Coherent structures and larval transport in the California ...

UNIVERSITY OF CALIFORNIA SANTA CRUZ
Coherent structures and larval transport in the California Current system
A dissertation submitted in partial satisfaction of the requirements for the degree of
DOCTOR OF PHILOSOPHY
in EARTH SCIENCES
by Cheryl S. Harrison
December 2012
The Dissertation of Cheryl S. Harrison is approved: Professor Eli A. Silver, Chair Professor David A. Siegel Research Scientist Adina Payton Professor Pete Raimondi
Tyrus Miller Vice Provost and Dean of Graduate Studies

Copyright © by Cheryl S. Harrison
2012

iii
Contents
List of Figures.............................................................................................................. v
Abstract....................................................................................................................... vi
Acknowledgements .................................................................................................... ix
Chapter 1 Introduction .............................................................................................. 1 Bibliography.......................................................................................................................10
Chapter 2 LCS in the CCS—sensitivities and limitations..................................... 14
Chapter 3 The role of filamentation and eddy-eddy interactions in marine larval accumulation and transport..................................................................................... 37
Abstract ..............................................................................................................................37 3.1 Introduction .............................................................................................................38 3.2 Lagrangian Coherent Structures (LCS)................................................................43 3.3 Models.......................................................................................................................47 3.4 Results.......................................................................................................................50
3.4.1 LCS, filamentation and larval density patterns ..................................................50 3.4.2 Filaments and fronts ...........................................................................................54 3.4.3 Eddy-eddy interactions, source regions and larval age distributions .................57
3.5 Discussion .................................................................................................................66 3.5.1 Application of LCS.............................................................................................66 3.5.2 Comparisons with observations and prior studies ..............................................68 3.5.3 Potential Roles of Larval Behavior ....................................................................72
3.6 Summary ..................................................................................................................73 3.7 Acknowledgements ..................................................................................................74 3.8 Bibliography.............................................................................................................75
Chapter 4 The tattered curtain hypothesis revisited: coastal jets control larval settlement patterns.................................................................................................... 83
Abstract ..............................................................................................................................83 4.1 Introduction .............................................................................................................84 4.2 Background theory ..................................................................................................89
4.2.1 Eulerian dynamics: upwelling jets and offshore transport .................................89 4.2.2 Lagrangian dynamics: a kinematic model of the upwelling jet..........................92
4.3 Biophysical Model....................................................................................................94 4.4 Modeling Results......................................................................................................96
4.4.1 Modeled patterns of coastal retention and settlement ........................................96

iv
4.4.2 Statistical Correlation Between Wind and Settlement .....................................101 4.5 Conclusions.............................................................................................................104 4.6 Discussion ...............................................................................................................104 4.7 Bibliography...........................................................................................................108
Chapter 5 Conclusions and directions for future research................................. 124 5.1 Conclusions.............................................................................................................124 5.2 Directions for future research ..............................................................................126
5.2.1 Quantifying biological patchiness. ...................................................................126 5.2.2 LCS and predators in the CCS..........................................................................129
5.3 Bibliography...........................................................................................................132
Appendix: Glossary ................................................................................................ 134
List of Supplemental Movies.................................................................................. 135 Movie 2.1 Daily LCS visualization for Fall 2005. .....................................................135 Movie 2.2 Comparison of Eulerian and Lagrangian metrics for 2002...................135 Movie 3.1 What happens to particles near LCS?.....................................................135 Movie 3.2 Larval density field....................................................................................135 Movie 4.1 Particles in the CCS-in-a-box model........................................................136 Movie 5.1 LCS and sea lion tracks ............................................................................136

v
List of Figures
Figure 1.1 Satellite observations of the California Current system (CCS)................... 1 Figure 1.2 Stagnation points and their manifolds ......................................................... 4 Figure 1.3 FTLE fields and LCS in the CCS................................................................ 5 Figure 3.1 Schematics of fluid/particle behavior near Lagrangian coherent structures (LCS)........................................................................................................................... 44 Figure 3.2 Comparison of Eulerian and Lagrangian surface fields. ........................... 48 Figure 3.3 North transect. ........................................................................................... 51 Figure 3.4 South transect. ........................................................................................... 53 Figure 3.5 Filament and frontal statistics.................................................................... 55 Figure 3.6 Packet formation........................................................................................ 57 Figure 3.7 Packet age class and source region distribution. ....................................... 60 Figure 3.8 Density distributions by age class on model day 177................................ 61 Figure 3.9 Source and age distributions...................................................................... 64 Figure 3.10 Particle density and packet statistics. ...................................................... 65 Figure 3.11 Effect of swimming on density patterns.................................................. 67 Figure 4.1 Features of upwelling dynamics in Eastern Boundary Currents (EBCs). 113 Figure 4.2 Gaussian jet. ............................................................................................ 114 Figure 4.3 Alongshore jet surface fields. .................................................................. 115 Figure 4.4 Coastal jet transects. ................................................................................ 116 Figure 4.5 Settlement patterns and wind for Run 111. ............................................. 117 Figure 4.6 Jet retention patterns................................................................................ 118 Figure 4.7 Settlement patterns and wind for Run 141. ............................................. 119 Figure 4.8 Settlement patterns and wind for Run 113. ............................................. 120 Figure 4.9 Statistics of wind-settlement correlation for Run 111............................. 121 Figure 4.10 Ensemble wind-settlement correlation statistics. .................................. 122 Figure 4.11 Scatterplot of integrated wind product W20 vs. number of settling particles. .................................................................................................................... 123

vi
Abstract
Coherent structures and larval transport in the California Current system
Cheryl S. Harrison
In the California Current system (CCS), coherent structures such as jets and eddies
strongly control the biological response to coastal upwelling. One of the outstanding
problems in marine ecology is to understand the mechanisms of larval transport.
Details of transport dynamics from nearshore to offshore, and subsequent delivery of
coastally spawned propagules back to favorable settlement areas, strongly control
marine population dynamics. Recent developments in applied dynamical systems
allow the identification of coherent structure boundaries by numerical calculation of
Lagrangian coherent structures (LCS), finite-time analogs of stable and unstable
manifolds of hyperbolic fixed points. These material lines divide the flow into regions
of disparate transport fate and illuminate the skeleton of filamentation and mixing.
Using altimetric observations of the CCS, we show that LCS are less sensitive to
noise and under-resolution than Eulerian metrics, and can map events undetectable by
analysis of the frozen-time velocity field. However, small-scale dynamics such as
lobes are not well resolved by altimetric observations. In an ocean circulation model
of an idealized CCS, LCS are used to track filamentation and eddy-eddy interactions,
important larval transport pathways from the shelf offshore. Filaments and packets
caused by eddy-eddy interaction are stable to horizontal swimming perturbations, as
predicted by the structural stability of hyperbolic manifolds. Retention in the

vii
upwelling jet is high and strongly patterns larval settlement. Settlement events are
best correlated with integrated upwelling intensity and not upwelling relaxation
events as commonly assumed. These studies demonstrate that coherent structures play
a large role in coastal ecosystems, and that techniques and theorems from dynamical
systems can provide insight into dominant transport pathways controlling coastal
ecosystems.

viii
Dedicated to Ralph
And all the other midwives of scientific revolution
Chase a deer and end up everywhere
-Rumi

ix
Acknowledgements
This work, and my subsequent academic trajectory, would have been in no way
possible without the academic assistance, logistical support and general sensei-like
qualities of Prof. David A. Siegel. He has been an example of how to forge your own
path and not just get away with it, but be wildly successful and have fun in the
process. His commitment and respect towards my academic vision, general academic
jedi training, and especially his ability to perturb trajectories toward more favorable
outcomes have been invaluable, to put it mildly.
I would like to thank the senior faculty of the UCSC Earth and Planetary Science
Dept, especially Eli Silver, for personifying that really the most fun you could ever
have is to do and teach science. This and the familial, supportive environment they
created enabled me to see that there may be a place for me in academia. I would also
like to thank the members of my committee, especially Adina Paytan, without whose
support and encouragement I would not be here. Gary Glatzmaier advised the first
chapter of this work, and his example of high integrity in both scientific practice and
in all academic endeavors has been immensely valuable.
The majority of acknowledgement for this work belongs to my family. For my entire
life, I have been nurtured with the crazy idea that I could do anything I wanted to.
Believing this and thus supported, so I now stand

1
Chapter 1 Introduction
Transport in the California Current system (CCS) is inherently chaotic (Fig. 1.1).
Instabilities generated in the upwelling jet (Brink 1987, Barth 1989, Durski and Allen
2005) coalesce into filaments and eddies that remain semi-coherent for weeks,
penetrating 100’s of kilometers offshore (Brink and Cowles 1991, Marchesiello et al.
2003, Capet et al. 2008, Ch. 3, Ch. 4). Large eddies (up 100-200 km in diameter)
form off instabilities in the main branch of the CCS as it migrates offshore at the end
of the upwelling season (Chelton et al. 2007, Ch 2), and are constrained by the beta
effect to propagate westward for months to years, transporting biologically important
materials far into the quiescent ocean interior, and forming important foraging habitat
for birds and marine predators (Capet et al. 2008, Ainly et al. 2009). Continuously
forced by ever-changing coastal winds (Burk and Thompson 1996), poleward
propagating coastally trapped waves (Brink 1991) and the ubiquitous interseasonal
variability of ENSO and PDO (Checkley and Barth 2009), the resulting quasi-
turbulent equilibrium state of the CCS is not quite turbulent enough to render
statistical methods applicable (Lekien and Coulliette 2003, Pasquero et al. 2007).
Figure 1.1 Satellite observations of the California Current system (CCS) (left) NOAA AVHRR satellite image of sea surface temperature on 10/9/92. (right) Ocean color (chlorophyll) near Monterey Bay.

2

3
There is a great amount of dynamical systems formalism for dealing with chaotic
systems; indeed this is one of the great mathematical achievements of the last century
(Abraham and Marsden 1978, Abraham and Shaw 1992, Wiggins 1992,
Guckenheimer and Holmes 1997). Dynamical systems focuses on the trajectories, i.e.
solutions of differential equations, instead of focusing on the forces, the terms in the
differential equations, or on the velocity field itself. The paradigm here is that much
of the behavior of the solutions is universal; there are a limited number of structures
that are combined to generate many different transport patterns—kind of a periodic
table of solution patterns with well-defined evolutions (i.e. bifurcations; Okubo
1970). There is a natural application of these methods to ocean transport (Wiggins
2005, Samelson and Wiggins 2006), which is governed by the non-linear Navier-
Stokes (NS) equations (e.g., McWilliams 2006). These equations are inherently
analytically intractable, and looking at the NS equations tells one very little about the
emergent behavior of the system.
This point is highlighted by the large number of new ocean dynamics discovered
since the advent of remote sensing by satellite (Ch 2 and references therein), and the
application of regional eddy-resolving ocean models (Marchesiello et al. 2004; Ch 3
& 4). How we think about the CCS and eastern boundary currents (EBCs) in general
has changed immensely in a very short time, with one of the great realizations being
that these systems are much more dynamic and unpredictable than we previously

4
thought (Hickey 1998, Hill et al. 1998, Marchesiello 2004). Here the endeavor is to
add to this knowledge base by applying recent results from dynamical systems.
Figure 1.2 Stagnation points and their manifolds Centers or elliptical (E) and saddle or hyperbolic (H) stagnation points organize transport in oceanic flows. Trajectories orbit elliptical points (dashed line). Near saddle points trajectories move on hyperbolic trajectories (black lines). Special trajectories form the inset or stable manifold (red) and outset or unstable manifold (blue) of the hyperbolic fixed point. These separatrices act as transport barriers and divide the flow into regions experiencing distinct transport fates in both forward and backward time.
Structures governing chaotic transport are organized around fixed or stagnation points
in the flow and their associated manifolds (Fig. 1.2). For time invariant systems there
is no motion at fixed points, i.e. the velocity vanishes, and fixed points can be found
by zeros of the velocity field and classified by eigenvalues of the Jacobian matrix
(Okubo 1970). In 2D incompressible ocean dynamics there are two types of fixed

5
points: centers and saddles. Near centers or elliptical stagnation points trajectories
move on closed elliptical orbits; these commonly occur in the centers of eddies. Near
saddles trajectories are hyperbolic in shape (Fig. 1.2). Saddles are stable in one
direction, with flow moving in toward the fixed point (red lines in Fig. 1.2) and
unstable in another direction, with flow moving away from the fixed point (blue
lines). These special trajectories are called the stable and unstable manifolds, or the
inset and outset of the hyperbolic fixed point (Abraham and Shaw 1992). Nearby
trajectories move asymptotically toward unstable manifolds, and we call these
attractors; likewise trajectories move asymptotically away from stable manifolds, i.e.
they are repelling.
The insets and outsets of saddle points extend out into the flow, often forming a
complex tangle of intersections as they wrap around eddies and other more exotic
structures (Fig. 1.2 & 1.3). Because they are trajectories, solutions to the NS
equations, existence and uniqueness of such solutions insures that they are also
transport boundaries, i.e. material cannot cross over them (see Shadden et al. 2005 for
exceptions). In time varying systems, these manifolds move with the fluid
Figure 1.3 FTLE fields and LCS in the CCS Shown are backward (left) and forward (middle) finite-time Lyapunov exponent (FTLE, days-1) maps computed from observations of sea surface height in the California Current system (CCS). (right) Lagrangian coherent structures (LCS) are identified by maxima or ridges of the forward FTLE (red) and backward FTLE (blue) and combined to make a map of coherent structure boundaries. Axes are degrees N (vertical) and degrees E (horizontal).

6

7
(supplemental movies 2.1, 2.2, 3.1), acting as a skeleton around which turbulent
transport is organized (Samelson and Wiggins 2006, Shadden et al. 2005, Mathur et
al. 2007).
Over the last decade, arguably the greatest advance to come out of applied dynamical
systems is the development of techniques to detect and track hyperbolic manifolds in
realistic velocity data (Haller 2002, Shadden et al. 2005). This innovation has resulted
in a large number of applications, from flow dynamics in the heart to the break up of
the atmospheric polar vortex associated with the ozone hole (reviewed in Shadden
2012 and Samelson 2013). Traditionally, hyperbolic manifolds were tediously
determined by identifying hyperbolic fixed points in the flow and seeding a dense
cloud of particles that would then stretch out and trace the unstable manifold of the
fixed point (e.g., supplemental movie 3.1). But this is a laborious process, involving
tracking of fixed points through time, which is quite difficult due to the complexity of
interactions (supplemental movie 3.1). These problems are compounded further by
the fact that zeros of the frozen time velocity field do not necessarily correspond to
fixed points in the time varying dynamics (Shadden et al. 2005, Mancho et al. 2006).
In order automate the detection of coherent structures, methods were pioneered using
various metrics on dense particle grids or groups of particles (Boffetta et al 2001,
Haller 2002, d’Ovidio et al. 2004), enabled by the exponential increase in computing
power. This culminated in the seminal work of Shadden et al. (2005), which defines
finite-time analogs of stable and unstable hyperbolic manifolds as Lagrangian

8
coherent structures (LCS), the ridges of the finite-time Lyapunov exponent (FTLE)
scalar field (Fig. 1.3). Details of this methodology are summarized in Ch. 2.
A central problem in marine ecology is the transport and dispersal of marine
propagules (larvae, eggs, spores), an inherently Lagrangian problem. Many species,
both pelagic (open ocean) and benthic (sea floor dwelling) spawn their young in the
productive coastal oceans of Eastern Boundary Currents (EBCs) such as the CCS
(e.g., Parrish et al. 1981, Rodríguez et al. 1999). These propagules are released into
the water column at some depth and must grow and often feed in the plankton,
drifting and swimming in the ocean currents until reaching maturity. In the case of
benthic species, residence time in the pelagic stage, called pelagic larval duration
(PLD), allow larvae to gain competency to recruit to suitable habitat and thus begin
the next stage of their lives, a strategy going back to the Cambrian (Valentine 2002).
PLD varies greatly between species, from a few days (abalone) up to many months
(some rockfish species). Likewise, swimming and other locomotive ability also varies
for benthic larvae, with a number of diverse strategies employed (e.g., Shanks 1985,
Morse 1991). But to leading order larvae are often considered passive or buoyant
floaters (Metaxas 2011), making the dispersal of these planktonic organisms a natural
application problem for chaotic Lagrangian transport techniques.
The main questions of this work is then, are dynamical systems techniques that detect
the separatrices bounding jets and eddies useful for determining Lagrangian (and thus

9
biological) transport patterns in the fecund and relentlessly productive CCS? Can the
wealth of mathematical formalism, intuition and theorems from the dynamical
systems study of coherent structures, lobe transport and other theorems tell us
anything useful about how transport works in the coastal ocean that can enable us to
better detect, model and manage our coastal ecosystems? This is the task at hand, and
is by no means complete.
This has been a challenging project. The inherent interdisciplinary nature of the
problem and the lack of existing literature and tradition have required forging my
own path, an inherently risky undertaking, but one with many rewards. The central
joy of science is the process of discovery: of finding new ideas and phenomena that
make sense of existing observations and expand our conceptual understanding of
important problems. The intersection between oceanography, dynamical systems and
marine ecology is an especially rich emerging field, and I am very happy to be on its
forefront.
This dissertation is organized as follows. Chapter 2 is a reproduction of a manuscript
published in GAFD (Harrison and Glatzmaier 2012) that applies LCS to
observational data in the CCS (geostrophic velocity fields calculated from satellite
observations of sea surface height) and empirically assesses the sensitivity to noise
and resolution, testing the structural stability of these features. Chapter 3 is a
manuscript currently in review on LCS and eddy interactions controlling nearshore to

10
offshore larval transport and packet formation in an idealized ocean model of the
Central California coast (Mitarai et al. 2008, Siegel et al. 2008). Chapter 4 is a
manuscript revisiting and revising a famous hypothesis on coherent structures
controlling settlement patterns in the same model (Roughgarden et al. 1991); here we
find that retention in the upwelling jet largely controls settlement patterns, and
similarity to other studies suggests this is a widespread phenomenon. Finally, Chapter
5 offers overall conclusions and outlines directions for future work, including
presentation of some work in progress.
Bibliography
R. H. Abraham and J. E. Marsden. Foundations of Mechanics. AMS Chelsea Publishing, 1978.
R. Abraham and C. D. Shaw. Dynamics–the geometry of behavior. Addison Wesley Publishing Company, 1992.
D. Ainley, K. Dugger, R. Ford, S. Pierce, D. Reese, R. Brodeur, C. Tynan, and J. Barth. Association of predators and prey at frontal features in the California Current: competition, facilitation, and co-occurrence. Marine Ecology Progress Series, 389:271–294, 2009.
J. A. Barth. Stability of a coastal upwelling front 2. Model results and comparison with observations. Journal of Geophysical Research, 94(C8):10857–10, 1989.
K. H. Brink. Upwelling fronts: implications and unknowns. South African Journal of Marine Science, 5(1):3–9, 1987.
K. Brink. Coastal-trapped waves and wind-driven currents over the continental shelf. Annual review of fluid mechanics, 23(1):389–412, 1991.
K. H. Brink and T. J. Cowles. The coastal transition zone program. Journal of Geophysical Research, 96(C8):14637–14, 1991.

11
G. Boffetta, G. Lacorata, G. Redaelli, and A. Vulpiani. Detecting barriers to transport: a review of different techniques. Physica D: Nonlinear Phenomena, 159(1-2):58 – 70, 2001.
S. D. Burk and W. T. Thompson. The summertime low-level jet and marine boundary layer structure along the California coast. Monthly Weather Review, 124(4):668–686, 2012/01/24 1996.
X. Capet, F. Colas, J. C. McWilliams, P. Penven, and P. Marchesiello. Eddies in eastern boundary subtropical upwelling systems, Vol. 177, pages 131–147. AGU, Washington, DC, 2008.
D. M. Checkley Jr and J. A. Barth. Patterns and processes in the California Current System. Progress in Oceanography, 83(1-4):49–64, 2009.
D. B. Chelton, M. G. Schlax, R. M. Samelson, and R. A. de Szoeke. Global observations of large oceanic eddies. Geophys. Res. Lett, 34(15):L15606, 2007.
F. d’Ovidio, V. Fernandez, E. Hernandez-Garcia, and C. Lopez. Mixing structures in the Mediterranean Sea from finite-size Lyapunov exponents. Geophysical Research Letters, 31(17), 2004.
S. M. Durski and J. S. Allen. Finite-amplitude evolution of instabilities associated with the coastal upwelling front. Journal of physical oceanography, 35(9):1606–1628, 2005.
J. Guckenheimer and P. Holmes. Nonlinear oscillations, dynamical systems, and bifurcations of vector fields. Springer-Verlag, 1997.
G. Haller. Lagrangian coherent structures from approximate velocity data. Physics of Fluids, 14(6):1851– 1861, 2002.
C. S. Harrison and G. A. Glatzmaier. Lagrangian coherent structures in the California Current system – sensitivities and limitations. Geophysical & Astrophysical Fluid Dynamics, 106(1):22–44, 2012.
B. M. Hickey. Coastal oceanography of western North America from the tip of Baja California to Vancouver Island. In A. Robinson and K. Brink, editors, The Sea—The Global Coastal Ocean, Regional Studies and Synthesis, volume 11, pages 345–393. John Wiley and Sons, Inc., New York, 1998.
A. E. Hill, B. M. Hickey, F. A. Shillington, P. T. Strub, K. H. Brink, E. D. Barton, and A. C. Thomas. Eastern ocean boundaries coastal segment. In A. Robinson and K. Brink, editors, The Sea—The Global Coastal Ocean, Regional Studies and Synthesis, volume 11, pages 29–67. John Wiley & Sons Inc, New York, 1998.

12
F. Lekien and C. Coulliette. Chaotic stirring in quasi-turbulent flows. Philosophical Transactions of the Royal Society a-Mathematical Physical and Engineering Sciences, 365(1861):3061–3084, 2007.
A. Mancho, D. Small, and S. Wiggins. A tutorial on dynamical systems concepts applied to Lagrangian transport in oceanic flows defined as finite-time data sets: Theoretical and computational issues. Physics Reports, 437(3-4):55–124, 2006.
P. Marchesiello, J. C. McWilliams, and A. Shchepetkin. Equilibrium structure and dynamics of the California current system. Journal of Physical Oceanography, 33(4):753–783, 2003.
M. Mathur, G. Haller, T. Peacock, J. Ruppert-Felsot, and H. Swinney. Uncovering the Lagrangian skeleton of turbulence. Physical review letters, 98(14):144502, 2007.
J. McWilliams. Fundamentals of geophysical fluid dynamics. Cambridge Univ Pr, 2006.
A. Metaxas. Behaviour in flow: perspectives on the distribution and dispersion of meroplanktonic larvae in the water column. Canadian Journal of Fisheries and Aquatic Sciences, 58(1):86–98, 2001.
S. Mitarai, D. Siegel, and K. Winters. A numerical study of stochastic larval settlement in the California Current system. Journal of Marine Systems, 69(3-4):295–309, 2008.
A. N. C. Morse. How do planktonic larvae know where to settle? American Scientist, 79(2):pp. 154–167, 1991.
A. Okubo. Horizontal dispersion of floatable particles in the vicinity of velocity singularities such as convergences. Deep-Sea Research, 17:445–454, 1970.
R. Parrish, C. Nelson, and A. Bakun. Transport mechanisms and reproductive success of fishes in the California Current. Biol. Oceanogr, 1(2):175–203, 1981.
C. Pasquero, A. Bracco, A. Provenzale, and J. B. Weiss. Particle motion in a sea of eddies. In A. Griffa, A. Kirwan, A. Mariano, T. Ozgokmen, and T. Rossby, editors, Lagrangian analysis and prediction of coastal and ocean dynamics, page 89. Cambridge University Press, Cambridge ; New York, NY, 2007.
J. Rodríguez, S. Hernández-Léon, and E. Barton. Mesoscale distribution of fish larvae in relation to an upwelling filament off Northwest Africa. Deep Sea Research Part I: Oceanographic Research Papers, 46(11):1969–1984, 1999.

13
J. Roughgarden, J. Pennington, D. Stoner, S. Alexander, and K. Miller. Collisions of upwelling fronts with the intertidal zone: the cause of recruitment pulses in barnacle populations of Central California. Acta Oecologica, 12(1):35–51, 1991.
R. M. Samelson and S. Wiggins. Lagrangian transport in geophysical jets and waves: the dynamical systems approach. Springer, New York, N.Y., 2006.
R. M. Samelson. Lagrangian motion, coherent structures, and lines of persistent material strain. Annual Review of Marine Science, 5(1), 2013.
A. Shanks. Behavioral basis of internal-wave-induced shoreward transport of megalopae of the crab Pachygrapsus crassipes. Marine Ecology Progress Series, 24:289–295, 1985.
S. C. Shadden, F. Lekien, and J. E. Marsden. Definition and properties of Lagrangian coherent structures from finite-time Lyapunov exponents in two-dimensional aperiodic flows. Physica D, 212(3-4):271–304, 2005.
S. C. Shadden. Lagrangian coherent structures. In R. Grigoriev and H. G. Schuster, editors, Transport and Mixing in Laminar Flows: From Microfluidics to Oceanic Currents, Annual Reviews of Nonlinear Dynamics and Complexity, Chapter 3. Wiley, 2012.
D. A. Siegel, S. Mitarai, C. J. Costello, S. D. Gaines, B. E. Kendall, R. R. Warner, and K. B. Winters. The stochastic nature of larval connectivity among nearshore marine populations. Proceedings of the National Academy of Sciences, 105(26):8974, 2008.
J. W. Valentine. Prelude to the Cambrian explosion. Annual Review of Earth and Planetary Sciences, 30(1):285–306, 2002.
S. Wiggins. Chaotic transport in dynamical systems. Springer-Verlag, 1992.
S. Wiggins. The dynamical systems approach to lagrangian transport in oceanic flows. Annual Review of Fluid Mechanics, 37:295–328, 2005.

14
Chapter 2 LCS in the CCS—sensitivities and limitations
Geophysical and Astrophysical Fluid DynamicsVol. 106, No. 1, February 2012, 22–44
Lagrangian coherent structures in the California CurrentSystem – sensitivities and limitations
CHERYL S. HARRISON* and GARY A. GLATZMAIER
Earth and Planetary Sciences Department, University of California,1156 High Street, Santa Cruz, CA 95064, USA
(Received 28 May 2010; in final form 27 August 2010; first published online 13 December 2010)
We report on numerical experiments to test the sensitivity of Lagrangian coherent structures(LCSs), found by identifying ridges of the finite-time Lyapunov exponent (FTLE), to errors intwo systems representing the California Current System (CCS). First, we consider a syntheticmesoscale eddy field generated from Fourier filtering satellite altimetry observations of theCCS. Second, we consider the full observational satellite altimetry field in the same region.LCS are found to be relatively insensitive to both sparse spatial and temporal resolution and tothe velocity field interpolation method. Strongly attracting and repelling LCS are robust toperturbations of the velocity field of over 20% of the maximum regional velocity. Contours ofthe Okubo–Weiss (OW) parameter are found to be consistent with LCS in large mature eddiesin the unperturbed systems. The OW parameter is unable to identify eddies at the uncertaintylevel expected for altimetry observations of the CCS. At this expected error level, the FTLEmethod is reliable for locating boundaries of large eddies and strong jets. Small LCS featuressuch as lobes are not well resolved even at low error levels, suggesting that reliabledetermination of lobe dynamics from altimetry will be problematic.
Keywords: Lagrangian coherent structures; Lyapunov exponents; mesoscale eddies; oceantransport; California Current
1. Introduction
Transport at the ocean surface has important implications for biological processes atmany trophic levels and is governed by coherent structures of varying scale (cf. Martin2003, Wiggins 2005). Many regions of the coastal and open ocean transport aredominated by meandering jets, mesoscale ocean eddies and eddy dipoles (Simpson andLynn 1990, Beron-Vera et al. 2008). Theoretical studies, numerical models and in situmeasurements show these eddies have often isolated water masses in their interiors forlong periods of time, indicating transport barriers exist on their edges. Eddies can carrythis isolated interior water with them as they travel, leading to such phenomenaas nutrient transport into depleted oligotrophic waters which triggers biologicalproductivity, retention of planktonic organisms in eddy interiors and transport of
*Corresponding author. Email: [email protected]
Geophysical and Astrophysical Fluid DynamicsISSN 0309-1929 print/ISSN 1029-0419 online ! 2012 Taylor & Francis
http://dx.doi.org/10.1080/03091929.2010.532793
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

15
coastal waters over long distances out to the open sea (e.g. Bracco et al. 2000, Pasqueroet al. 2007, Rovegno et al. 2009).
In contrast, eddy edges are regions of rapid transport and mixing where tracergradients are intensified (Tang and Boozer 1996, Abraham and Bowen 2002, d’Ovidioet al. 2009). These boundaries have been associated with vertical transport (Capet et al.2008, Calil and Richards 2010), chlorophyll concentration (Lehahn et al. 2007) and seabird foraging (Kai et al. 2009). Globally, predators are observed to preferentially swimand forage along sea surface temperature gradients associated with coherent structureedges (e.g. Worm et al. 2005). On a larger scale, there is interest in parameterizingstatistics of mixing excited by ocean eddies for inclusion in global ocean circulationmodels (e.g. Ponte et al. 2007, Waugh and Abraham 2008).
Though of great interest, identification of eddy boundaries in both model andobservational fields is not trivial. Often a snapshot of the velocity field is analyzedusing classical dynamical system techniques as in the Okubo–Weiss (OW) partition(Okubo 1970, Weiss 1991). When 3D model fields are analyzed, the vertical componentof the vorticity is often employed for coherent structure identification (e.g. Calil andRichards 2010, Koszalka et al. 2010). But what is of primary interest – the transport ofwater parcels in and out of an eddy or across a coherent structure – is fundamentally aLagrangian problem. This requires consideration of the full time-varying dynamics andintegration of particle trajectories to adequately capture a feat that is only now tractablewith increased computational resources.
Recent dynamical system techniques (e.g. Haller and Yuan 2000, d’Ovidio et al. 2004,Mancho et al. 2004, Shadden et al. 2005, Lekien et al. 2007, Rypina et al. 2007) haveprovided tools for mapping the wealth of information that the Lagrangian approachprovides, including the identification of transport barriers, transport mechanisms, andregions of rapid dispersion (cf. Wiggins 2003). These tools have been applied togeophysical systems as diverse as the Earth’s mantle (Farnetani and Samuel 2003), theozone hole (Rypina et al. 2007), and harmful algal blooms in the coastal ocean(Olascoaga et al. 2008). Here we focus on identification of coherent structures in theCalifornia Current System (CCS) using the Lagrangian coherent structure (LCS)technique, which uses maxima of the finite-time Lyapunov exponent (FTLE) field(Shadden et al. 2005). We calculate LCS in systems derived from satellite altimetryobservations. Currently, there is much interest in both the oceanography and marineecology communities to fully explore the application of LCS. The need for a study ofthe limitations of this approach is the motivation of this work.
Finite Lyapunov exponents have not been previously studied in the CCS except onvery regional scales (Coulliette et al. 2007, Ramp et al. 2009, Shadden et al. 2009), wherethey have been used to study transport associated with the Monterey Bay eddy, and onvery large scales (Beron-Vera et al. 2008, Waugh and Abraham 2008, Rossi et al. 2009),where they have been applied to studies of global mixing and eddy visualization. Sinceeddies and other coherent structures are similar throughout all Eastern Boundaryupwelling systems and other regions in the global ocean (Chelton et al. 2007), we believethe results here will be widely applicable.
To assess the efficacy of the dynamical systems approach in studying the generaltransport characteristics of the CCS we ask the question, how well does the FLTEmethod work given the available data and its inherent uncertainty? There is sometheoretical (Haller 2002) and experimental (Shadden et al. 2009) evidence that LCSderived from approximate, discrete (i.e. real-world) data are robust to errors in the
LCS in the CCS 23
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

16
velocity field. Our goal is to experimentally test the extent of this stability for LCSderived from satellite altimetry observations, which are known to have high levels ofuncertainty, especially in regions of low variability such as the CCS (e.g. Pascual et al.2009). To this end, we first construct a synthetic model that captures the dynamics ofthe largest eddies in the CCS but can represent them with finer spatial and temporalresolution than the observations. In contrast to other studies (Olcay et al. 2010,Poje et al. 2010), we are not asking how small perturbations affect LCS location, sincewe know there are always errors in the observations and thus the LCS are inherentlyapproximate, but how large the errors have to be before the overall topological structureof the system is obscured. That is, we are testing when LCS break down and howreliable they are in observational oceanography. We do this with the motivation that wewould like to apply the LCS method of identifying ocean transport processes inobservational data in future studies.
The remainder of this article is organized as follows. In section 2 we provide somebackground on LCSs and the FTLE. In section 3 we present the data, models andmethods employed in this study. This includes explanation of how the synthetic field isconstructed. Section 4 contains results of this study. First we compare the LCS and OWmethods, then report results of testing the effects of noise and resolution in both the fullobservational and synthetic models, and finally show that the maximum FTLE dependson the initial spacing of the Lagrangian particles. We conclude with a summary anddiscussion of the implications of these results in section 5.
2. Lagrangian coherent structures
2.1. Background
In time-invariant systems distinct dynamical regions can be divided up by findingseparatrices, trajectories that divide one flow structure from another. For theapplication to mesoscale ocean dynamics, a well-studied example is the boundarybetween an eddy and a jet (e.g. Samelson 1992). This system can be visualized byexamining the structure of an analytical kinematic model representing cold and warmcore eddies straddling a western boundary current (such as the Gulf Stream) after itseparates from the coast. These kinematics can be described by the streamfunction(Samelson and Wiggins 2006)
! "by# A sin$x" !t% sin$ y%, $1%
where x and y are the along-jet and across-jet horizontal coordinates, respectively.From (1) the velocity field can be extracted by taking the partial derivatives
_x ! u ! " @ @y
! b" A sin$x" !t% cos$ y%, $2%
_y ! v ! @
@x! A cos$x" !t% sin$ y%, $3%
where the dot represents a derivative in time.Consider initially the stationary system (!! 0), i.e. no phase velocity. From
inspection of the velocity field (figure 1) we can see that there are two recirculation
24 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

17
regions (eddies) separated by a meandering jet. Near the centers of the eddies fluidrecirculates in elliptical orbits around the fixed point where the velocity vanishes, whilefluid near the center of the jet is transported downstream. Two questions emerge:(1) what divides these two regions and (2) how do we find this separatrix? This is themotivation for the identification of hyperbolic material curves, also called LCSs,which divide the dynamics into distinct regions and provide a framework, or skeleton(Mathur et al. 2007), for understanding transport.
In addition to the elliptical fixed points in the center of the eddies there are hyperbolicfixed points, or saddle points, on the boundary between the jet and the eddy. Near thesehyperbolic points, some particles that begin close together separate at an initiallyexponential rate; whereas other particles that are initially far apart move closer to eachother at an exponential rate, causing water parcels to be stretched along the eddy-jetboundary (figure 1). Between these trajectories, there exists a trajectory on whichparticles arrive at (depart from) the hyperbolic fixed point in the limit of infinite-time.This is called the stable (unstable) manifold, hyperbolic material curve or repelling(attracting) LCS of that fixed point. Nearby trajectories diverge away from(converge to) it as they near the saddle point. (Note the unfortunate definition ofrepelling LCS as stable; this is historical.)
In the time-invariant case, LCS correspond to streamlines. For time-varying(non-autonomous) systems fixed points and hyperbolic material curves may stillexist but cannot be found by classifying the fixed points in a snapshot of the flow(Shadden et al. 2005, Wiggins 2005, Samelson and Wiggins 2006). The time-varyingtopology can be close to the frozen-time topology if the system is evolving slowlyenough, as is often found to be the case in oceanographic systems (Haller and Poje1998). Still, in the extension to time-varying systems instantaneous streamlines do notcorrespond to transport barriers, necessitating the use of other methods in theirdetermination (Boffetta et al. 2001, Haller 2001, Abraham and Bowen 2002,
Figure 1. Meandering jet kinematic model (1) with b! 0.25m/s, A! 0.5m/s and !! 0.04m/s plotted fort! 11 days. Domain size is 2!" 100 km by !" 100 km. With these parameters the eddies are moving to theright. Here we plot the coincidence of particles (white dots) moving with the flow forward in time, and theattracting LCS, the ridge of the FTLE field (gray field), calculated from backwards integration of particletrajectories. Particles visualized by white dots were initialized in a horizontal line near the bottom boundary att! 0, and have been swept up and concentrated along the edge of the eddy.
LCS in the CCS 25
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

18
d’Ovidio et al. 2004, Mancho et al. 2004, Shadden et al. 2005). LCS in time-varyingsystems can be visualized using maxima of the direct or FTLE field, though othermethods (notably the finite size Lyapunov exponent) exist and have been widely applied(cf. d’Ovidio et al. 2009).
2.2. Definition of the FTLE
The velocity field
dx
dt! v"x, t# "4#
can be integrated to generate Lagrangian particle trajectories x(x0, t). If we wish tofollow the evolution of the difference between some initially close particles, we can trackthe relative dispersion !x(t)! x(a, t)$ x(b, t) as the particle positions evolve in time.The evolution of !x is governed to first order by
!x"t2# ! D!x"t1#, "5#
where !x(t1) is the initial separation between the particles at time t1 and D is thedeformation gradient tensor (cf. Ottino 1989) and Dij! (@xi(t2)/@xj(t1)).Particles straddling a repelling LCS separate at an exponential rate as they approacha saddle point. This allows one to define the LCS as maximal curves of the localmaximum rate of average exponential particle separation (Haller 2001, Shadden et al.2005), i.e.
! ! 1
jt2 $ t1jln""max#1=2, "6#
where ! is the FTLE and "max is the maximum eigenvalue of (DTD). Note that ! is ascalar field and depends on space, the initial time t1 and the integration time jt2$ t1j.Forward integration is used to determine repelling LCS, and backward integration isused to determine attracting LCS.
Practically, D is calculated by finite differencing. Given velocities on a regular 2Dgrid at a sequence of discrete times, we interpolate (4) in space and time to advect tracerparticles. For every snapshot of the FTLE field we start (at time t1) a new set ofuniformly distributed tracer particles that are advected to time t2 to numericallycompute a deformation tensor D at time t1, defined on the cartesian grid (x, y) as
D"x, y, t1, t2# !
@x2@x1
@x2@y1
@y2@x1
@y2@y1
0
BBB@
1
CCCA: "7#
The partial derivatives in (7) are computed as ratios of the separation betweentwo neighboring tracer particles at time t2 over their separation at time t1. Forexample, consider a given grid location (x1, y1) on the ocean surface and its
26 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

19
four nearest neighbors at time t1 (figure 2). We make the followingapproximation:
D!x, y, t1, t2" #
xE2 $ xW2xE1 $ xW1
xN2 $ xS2yN1 $ yS1
yE2 $ yW2xE1 $ xW1
yN2 $ yS2yN1 $ yS1
0
BBBB@
1
CCCCA: !8"
3. Data, models and methods
3.1. Data and construction of the synthetic field
The time averaged structure of the CCS is characterized by an equatorward jet thatintensifies seasonally in response to alongshore wind stress at the ocean surface (Hickey1998) (figure 3(a)). The variability of the CCS is often conceptualized as a mesoscaleeddy field superimposed on this average structure, with maximum eddy kinetic energyin the late summer and early fall (e.g. Marchesiello et al. 2003, Veneziani et al. 2009).Examination of the altimetry standard deviation (figure 3(b)) reveals the scope of theeddy field, which extends from NW to SE (parallel to the coastline) from roughly 32% to43%N and 229% to 238%E. The peak in this standard deviation (9 cm) is almost half ofthe time-averaged rise in the sea surface height (&20 cm) across the intense part ofthe mesoscale eddy region. Thus, rather than being a small perturbation on theclimatological mean, the mesoscale eddy field dominates the dynamics (and ofparticular interest to this study, the transport dynamics) in the CCS.
To capture the dynamics of this eddy field in a synthetic model in which the temporaland spatial resolution of the velocities can be varied, we Fourier filter remotely sensedaltimetric sea surface height data. By retaining only the larger dominant modes in theresulting empirical dispersion relation, we construct a synthetic time-dependent eddyfield of horizontal flows that has the same spatial structure, eddy propagation speedsand decorrelation timescale as the available mesoscale observations of the CCS.
(x2W , y2
W )
(x2N ,y2
N )
(x1W ,y1
W )
(x2,y2)
(x1N ,y1
N )
(x1S , y1
S )
(x1E ,y1
E )
(x2E ,y2
E )
(x2S ,y2
S )(x1 ,y1 )
Figure 2. Schematic representation of particle grid evolution from model time t1 to t2.
LCS in the CCS 27
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

20
Satellite altimetry is taken from the Archiving, Validation and Interpretation ofSatellite Oceanographic Data (AVISO) updated absolute dynamic topography (ADT)product, which represents sea surface height relative to a reference geoid. This productis the most accurate data currently available from remotely sensed altimetric sea surfaceheight off the US West Coast, merging signals from multiple satellites duringpost-processing to increase accuracy (Dibarboure et al. 2009, Pascual et al. 2009). WeFourier analyze the AVISO data within an offshore region extending from 32–47.5!Nand 225–235!E (boxes in figure 3) to minimize inclusion of inaccurate data near thecoastline. This region covers a large extent of the mesoscale eddy field off the CentralCalifornia coast. ADT weekly output is analyzed from 1992–2008, the total time theupdated product was available from AVISO at the time of this study.
The discrete Fourier transform (DFT) of the altimetric data is calculated over theocean surface in latitude, longitude, and through time. The synthetic model isconstructed by examination of the dispersion relation (figure 4), and restriction of theinverse transform to the nine most dominant modes in the mesoscale range,corresponding to wavelengths ranging from 260 to 419 km and with periods rangingfrom 7 to 10 months. These modes account for only about 30% of the power in thisrange but when inverted to physical space result in a synthetic sea surface height fieldthat exhibits the same size ("2!) and westward phase propagation velocity ("2 cm/s) ofthe larger eddies seen in the full observational field. The decorrelation time scale,calculated as the first zero crossing of temporal autocorrelation of the synthetic seasurface height, is "10 days over the entire spatial domain, compared to spatiallyvarying 10–15 day decorrelation time scale for the observational field. Note that,
5
7.5
10
10
12.5
12.5
12.5
12.5
15
15
15
1517.5
17.5
17.5
20
20
20
22.5
22.5
25
25
27.5
27.5
30
30
32.5
32.5
35
3537.5
40
40
Mean ADT (cm)1992 2008
Longitude
Latit
ude
226 228 230 232 234 236 238 240 242
32
34
36
38
40
42
44
46
48
50
4
45
55
5
5
5
5
5
5
5 5
5
55
66
6
6
6
6
6
6
6
6
6
6
6 6
6
6
7
7
7
7
7
7
8
8
8
8
9
ADT Standard Deviation (cm)1992 2008
Longitude226 228 230 232 234 236 238 240 242
32
34
36
38
40
42
44
46
48
50
(a) (b)
Figure 3. The 16-year average (a) and standard deviation (b) of ADT, a proxy for sea surface height, takenfrom AVISO satellite altimetry data, with the Fourier analyzed region denoted by the box. The CCS iscommonly characterized by an eddy field (b) superimposed on an equatorward jet (a). The strength and extentof this eddy field can be visualized by examination of areas of maximal standard deviation. Note that thechange in ADT across the intense part of the eddy field is only "20 cm, whereas the standard deviation has amaximum of 9 cm and so represents a significant departure from the mean.
28 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

21
however, a more practical time scale for the LCS is the average time for the phase ofthese eddies to propagate a distance equal to their size, which is roughly 70 days.
For both the reduced synthetic model and the full observational altimetry field,geostrophic velocities are calculated in the usual manner:
u ! " g
fRe
@h
@!, v ! g
fRe cos!
@h
@", #9$
where u and v are the zonal and meridional velocities, respectively, g is the gravitationalacceleration, f is the Coriolis parameter, " is longitude, ! is latitude, Re is the Earth’sradius, and h is the sea surface height. That is, because of the geostrophicapproximation, velocities at the ocean surface are simply defined as being directedperpendicular to the local gradient of the sea surface height, with amplitudesproportional to the amplitude of this gradient. Since the maxima of the sea surfaceheight are smaller for the synthetic field (owing to the reduction in power spectrum), wescale the synthetic velocity field to have the same maximal value as the full AVISOobservational field. This ensures that the timescales of the particle dynamics and FTLEevolution are consistent across the two models.
3.2. LCS calculation
We use the FTLE method (section 2.2) for this study because it has been shown bothanalytically and experimentally that if the ridges of the FTLE field are sharp and wellbehaved then the flux across them is negligible (Shadden et al. 2005). Further, it hasbeen shown that hyperbolic material curves found by any method are robust to errors inobservational or model velocity fields if they are strongly attracting or repelling or existfor a sufficient amount of time. In such a case, errors in the velocity field can even be
1 1.2 1.4 1.6 1.8 20.012
0.014
0.016
0.018
0.02
0.022
0.024
0.026
0.028
(oscillations year–1)
K (
rad
km–1
)
log
10P
ower
1.45
1.4
1.35
1.3
1.25
1.2
1.15
Figure 4. Dominant frequencies (#) and wavenumbers (K) in the dispersion relation for the observationalfield, here restricted to the mesoscale range. The nine modes from the Fourier analysis with the most energy(squares) are retained and inverted to create a synthetic field capturing the dynamics of eddies in the CCS.Here K ! #k2x % k2y$
1=2. Color figures are available in the electronic version.
LCS in the CCS 29
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

22
large if they are short lived (Haller 2002). This is due to the fact that though the particletrajectories will in general diverge exponentially from the true trajectories near arepelling hyperbolic material curve (i.e. sensitive dependence on initial conditions), theLCS are not expected to be perturbed to the same degree, because errors in the particletrajectories spread along the LCS. This is explored experimentally in section 4.3.
In this study, each temporal realization of the FTLE (initialized at time t1) isdetermined by initializing particles on a regular, dense grid and integrating theirtrajectories forward to time t2 using a fourth-order Runge–Kutta integration algorithm.Initial particle separations of 0.01!–0.10! resolution in both latitude and longitude wereexplored to visualize the LCS (figure 5), with 0.05 degrees chosen for most of oursimulations as a balance between computational speed and clarity of visualization.
Whereas the maximum value of the FTLE depends on the initial separation of theseeded particles used to calculate the trajectories (section 4.4), there is little effect on thelocations of the LCS. We interpolate velocities in space and time by both cubic andlinear methods to test the dependence of the sensitivity of the LCS on interpolationscheme. The FTLE is calculated every 1–7 days, since it was found for the systemsstudied here that the LCS change very little in this amount of time.
For each temporal snapshot of the forward or backward FTLE, particle trajectoriesare integrated for 24 days or longer. The ridges of the FTLE field lengthen and attainsharp maxima in more regions when using longer integration times, revealing morestructure in the LCS (figure 5) and identifying chaotic regions by eventually ‘‘filling’’them with LCS. We found that after 24–36 days the LCS capture the majority of theeddy boundaries for high activity regions and cover the study region enough to test theirsensitivity. To visualize the LCS at a given t1, maxima of forward and backward FTLEmaps (figure 11 in section 4.3) are combined (figure 6).
One difficulty in calculating the FTLE (or any Lagrangian measurement) in a finitedomain is what to do for particles that leave the model boundary during the integrationtime (Tang et al. 2010). Here we follow the method of Shadden et al. (2009), andcalculate the FTLE field for particles exiting the model boundary, and their neighbors,at the time the particles leave instead of at t2. This fix is very inaccurate for quickly
Figure 5. Forward FTLE for (a) 0.1! and (b) 0.025! initial particle separation and 24 day trajectoryintegration time for the synthetic model (section 3.1), and (c) 0.025! particle separation and 48 dayintegration. Finer FTLE resolution affects the width of the LCS and the maximum FTLE value (notevariation in gray scale limits). Longer integration time ‘‘grows’’ the LCS.
30 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

23
exiting particles, where the FTLE does not have time to converge and develop the sharpmaxima needed to identify the LCS. Therefore, for this study we only calculate the LCSwell inside the model boundary (1–2! in all directions) to minimize the number ofparticles exiting during each integration. Because the maximum FTLE amplitudedecreases with time (section 4.4) care must be taken when identifying the ridges if theFTLE are calculated this way.
4. Results
4.1. LCSs and OW contours
Here we test the use of LCSs as a diagnostic tool for measuring how well the interiors ofeddies are isolated from their surroundings. We also compare the use of LCS with themore commonly and historically used Eulerian time-invariant diagnostics, such as theOW parameter (Okubo 1970, Weiss 1991).
The OW parameter measures the relative strengths of vorticity and shear and isdefined as
OW " s2 # !2,
Figure 6. Lagrangian vs. Eulerian determination of coherent structures in the observational field forOctober 19, 2005. Attracting (blue) and repelling (red) LCS are shown on the left. Short black lines on the leftfigure are trajectories for $1 day, with a black circle indicating the final particle position. In the right figurecontours of sea surface height (SSH), geostrophic velocity vectors, and the OW parameter (color field, unitsare %10#10 s#2) are plotted for the same day. The SSH contour interval is 3 cm, the maximum fluid velocity is35 cm/s. Dark blue regions indicate elliptical trajectories (high vorticity), yellow and orange regions indicateregions of high shear near instantaneous saddle points. The zero contour of the OW parameter is indicated bya white line. See text for interpretation. Color figures are available in the electronic version.
LCS in the CCS 31
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

24
where
s2 ! "@u=@x# @v=@y$2 % "@u=@y% @v=@x$2
is the sum of the squares of the normal and shearing deformation (Okubo 1970), and
!2 ! "@v=@x# @u=@y$2
is the square of the vertical vorticity. OW can be used to ‘‘roughly’’ divide theinstantaneous velocity field into regions dominated by vorticity (OW5 0), such as theinteriors of eddies, and regions dominated by shear (OW4 0). This is equivalent toclassifying the eigenvalues of the Jacobian of the velocity field, a technique used toclassify fixed points of steady systems as elliptical (centers) or hyperbolic (saddles) intime-invariant systems (Okubo 1970).
It has recently been shown that for a persistent standing eddy in the WesternMediterranean there is a strong correlation between LCS and contours of the zerolevel of the OW parameter (d’Ovidio et al. 2009), whereas previous work inturbulence simulations has found that the OW zero contours generally plot inside theeddy boundaries defined by the LCS (Haller and Yuan 2000). Because eddies in theCCS represent a unique dynamical system relative to these two studies, we seek tovalidate the common use of the OW parameter in determination of eddy boundaries(e.g. Isern-Fontanet et al. 2003, Chelton et al. 2007, Rovegno et al. 2009, Nencioliet al. 2010).
In the CCS, eddy phase propagation velocities (&2 cm/s) are an order of magnitudesmaller than fluid velocities along eddy boundaries (&10–50 cm/s), especially for largeeddies. This velocity scale separation arguably holds for most large eddies in theglobal ocean (Chelton et al. 2007), indicating that the instantaneous velocity fieldcould represent the Lagrangian dynamics well. Some dynamical systems studies testthe limit of fast eddy propagation speeds using a kinematic model and suggest thatthis order of magnitude difference is small enough that the Eulerian and Lagrangiantopologies remain consistent (e.g. Haller and Poje 1998). In contrast, a more recentstudy using observational data has suggested that the OW parameter is not adequatefor eddy identification (Beron-Vera et al. 2008). So how correlated are LCS and OWcontours in the CCS? Here we begin to explore this question by assuming (for now)that geostrophic velocities taken from altimetry exactly represent the dynamics ofthe CCS.
Figure 6 compares Eulerian and Lagrangian diagnostics for a large anti-cycloniceddy (37'N, 231.5'E) soon after it detaches from a large meander in the main branch ofthe California current and begins its 20 plus week journey westward to our domainboundary. By inspection of both the velocity field and the LCS ‘‘map’’, we can see thatthe large eddy is accompanied to the south by a smaller cyclonic eddy and by a smallcyclonic/anti-cyclonic eddy pair to the northwest. This coupling of eddies in the form ofeddy dipoles is common and widely studied in physical and biological oceanography(e.g. Simpson and Lynn 1990, Beron-Vera et al. 2008).
By inspection of the LCS map, we see that there is extensive entrainment of thesurrounding ocean into the larger eddy, with a direct transport pathway or ‘‘mixingchannel’’ from the equatorward jet (visible in the north east of the figure), consistentwith previous kinematic studies of eddy–jet interaction (Haller and Poje 1998). There isalso a smaller eddy, centered at 39'N and 232'E, in the process of being absorbed into
32 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

25
the large eddy (movie 1).y The attracting LCS in the center of this large eddy representregions of exponential particle separation in backward time and thus particles straddlingthem were recently widely separated and outside of the eddy. This pathway ofentrainment crosses the zero contour of the OW parameter, indicating it is not a robustbarrier in this instance.
Later in its evolution the mixing channel is broken and there are no longer LCS in thelarge eddy’s interior (movie 1), indicating that entrainment largely stops after formationand the large eddy is perhaps immune to perturbation by smaller scale features. Thisobservation is consistent with the theory that high potential vorticity gradients at eddyedges make them impermeable to the surrounding fluid except during formation,dissolution and when the eddies are highly perturbed (cf. Pasquero et al. 2007).Contrary to Beron-Vera et al. (2008), we find that for these large, mature eddies the OWpartition and the LCS are consistent, validating the use of the OW parameter in thiscase. This agreement between the OW partition and LCS is seen for large well-formededdies in other years as well (movie 2)y.
Due to the filtering technique, eddies in the synthetic model are not generated bypinching off from a jet-like structure, but always appear well-developed. These largeeddies also do not interact strongly, but instead propagate westward at similar speeds.Thus the dynamics of the synthetic model can be thought of as ‘‘all large eddies all thetime’’. Similar to what is found for the large, well-developed eddies in the observationalsystem, the OW partition for the synthetic model is consistent with the eddy boundariesdetermined by the LCS (figure 7a) and is discussed further in section 4.2.1.
4.2. Noise sensitivity
Though the ability of satellite altimetry to accurately represent ocean dynamics hasvastly improved over the last decade (cf. Le Traon et al. 2009), there is still significantuncertainty resulting from poor satellite coverage in space and time. This uncertainty,often called the mapping error, can dominate the signal in regions of low variability,such as the central North Pacific, leading to large errors in altimetry derived velocityfields (cf. Pascual et al. 2007). Such velocity fields are often used without considerationof the large errors involved. Recently, Pascual et al. (2009) compared global altimetry-derived surface velocities with in-situ drifter velocities and found average differences of!10 cm/s in the CCS region. West and North of the main jet in the region, theCalifornia Current (where velocities are upwards of 50 cm/s), lies a low variabilityregion where average velocities are often less than 10 cm/s (figure 7). In this region theuncertainty is often as large as the recorded observations, necessitating carefulconsideration of error effects.
In this section we examine the effects of errors in the altimetry field and show thatLCS identification of transport barriers is less sensitive than the more commonly usedOW parameter (section 4.1). Our error field is constructed using normally distributedrandom noise added to both the synthetic model and observational sea surface heightfields, with a unique random noise distribution added each week. Four different testswere conducted with the RMS sea surface height noise being 1, 2, 3, and 4 cm,respectively. Each centimeter of added noise corresponds to an additional perturbation
ySee supplemental material in electronic version.
LCS in the CCS 33
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

26
in the derived velocity of about !5 cm/s, resulting in velocity perturbations ranging onaverage from 5 to 20 cm/s.
Satellite mapping errors in the CCS can be larger than 3 cm (Pascual et al. 2006), butbecause they are spatially correlated velocity, errors will not be as large as those in ourtest simulations for the same SSH perturbation. Our approach to represent these errorsresults in artificially large SSH gradients which lead to velocity errors could beunrealistic. For this reason, we choose to emphasize the average velocity error in orderto compare our results with the observed velocity errors reported in the literature.
4.2.1. Noise in the synthetic field. The synthetic CCS model is homogeneous enoughto determine a threshold for LCS identification, but also contains enough variability to
Figure 7. Effect of noise on the synthetic field. Model week 600 velocities are shown in (c), where the color-field represents the magnitude of the velocity field and the arrows indicate velocity direction and magnitude.Attracting (blue) and repelling (red) LCS are plotted for the unperturbed field (a) and four levels of noise(b–f). The dashed lines in a, b and d are the OW contours, plotted at "20% of the first standard deviation forthat week. See section 4.2.1. Color figures are available in the electronic version.
34 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

27
examine the effects of noise on regions of differing signal strength. Figure 7 depicts thevelocity field and LCS for week 600 of the synthetic model. During this time the eddyfield exhibits a weaker circulation in the north, with maximum velocities less than15 cm/s, and a stronger region to the south, with maximum velocities of 35 cm/s. Highermaximum velocities correspond to faster relative dispersion, consequently LCS in thesouthern portion of the domain are more intense, develop in shorter integration timeand are theoretically more resistant to perturbation (section 3.2).
For the unperturbed synthetic field (figure 7(a)) we see that the OW parameter iseffective at finding the intense eddy cores corresponding to low mixing. Eddy cores arethe regions where there are no strong LCS and thus where particles do not disperse ineither forward or backward time for an appreciable period. Here we have plotted theOW contour at 20% of the first standard deviation in the negative direction orOWc!"0.2!, where sigma is the standard deviation taken over the entire field for alltime, as is commonly done in application of the OW parameter in automated eddyidentification routines (e.g. Rovegno et al. 2009, Nencioli et al. 2010). We note that thechoice of contour level for the OW parameter is somewhat arbitrary since there is strongspatial and temporal variation in the eddy field. Though the LCS and the OW partitionare consistent, the LCS cross the OW contours in many locations, indicating thatthough the OW contours are effective at eddy identification, they cannot be consideredrobust transport barriers since flow is in general tangental to LCS curves.
At the 1 cm rms SSH (5 cm/s velocity) noise level, both the OW and LCS methods areeffective in eddy identification in the high velocity region, whereas in the weak eddyregion the signal is dominated by noise and both methods fail to identify any coherentstructures (figure 7(b)). At the 2 cm rms (10 cm/s) noise level the OW contours are notable to identify eddies even in the strongest regions, whereas the LCS method isvery effective at identifying regions of low dispersion for the stronger eddy cores(figure 7(d)). Note that this noise level corresponds to roughly 30% of the maximumvelocity and so represents a large perturbation on the field. OW contours are omitted atthe higher noise levels in figure 7 as they become even more noise dominated.
Regions of low mixing, such as the large eddy center at (40#N, 230#E), are stillevident even at the 4 cm rms (20 cm/s) noise level (figure 7(f)), a perturbation that is onaverage greater than 50% of the maximum velocity. From these observations it is clearthat, if the motivation of eddy identification is to determine regions of coherent andisolated water parcels free of entrainment from the surrounding fluid, the LCS methodis more effective than the OW parameter. It is also clear that the LCS method is robustto uncertainties of 20% or more of the maximum regional velocity.
4.2.2. Noise in the observational field. In this section we report that strong LCS in theobservational velocity field are also able to withstand a 20–30% perturbation. Figure 8shows the perturbed LCS for October 19th, 2005, as in figure 6. There are two eddydipole pairs evident at this time. The largest, centered at 37#N and 231.5#E, hasmaximum velocities of 35 cm/s between the two eddies. To the northwest, centered at40#N and 230#E, is a smaller and weaker eddy pair with maximum velocities of less than25 cm/s. The LCS structures for the weaker eddy pair are still clearly evident at the 1 cm(5 cm/s) noise level (figure 8(d)), which represents a perturbation of over 25% of themaximum velocity for this dipole. At the 2 cm (10 cm/s) noise level (figure 8(e)), thesmall eddy pair is no longer identifiable, whereas the largest eddy is clearly evident. This
LCS in the CCS 35
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

28
noise level represents an approximately 30% perturbation on the large eddy pair. TheLCS mapping also shows the major jet of the CCS, the California Current, in theeastern portion of figure 8. This jet (figure 8(c)) has maximum velocities upwards of40 cm/s, and is still evident in the LCS plots at the 3 cm (15 cm/s) noise level,representing an uncertainty of well over 30% (figure 8(d)).
Notably, the OW contours are not able to identify the jet structure of the CCS, one ofthe main features in the dynamics in this system. The LCS method allows visualizationof these jet structures and continuity of eddies over time, including their formationand dissolution, eddy-eddy interaction and entrainment pathways (movies 1 and 2).As found for the synthetic model, OW contours are somewhat effective at the 1 cmnoise level but ineffective at eddy identification at higher levels of perturbation
Figure 8. Noise in the observational field. OW contours for two noise levels (a), (b) for the same day andscaling as in figure 6, geostrophic velocities (c), and attracting (blue) and repelling (red) LCS for three noiselevels (d)–(f) for altimetry observations of the CCS on October 19, 2005. See section 4.2.2. Color figures areavailable in the electronic version.
36 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

29
(figures 8(a) and (b)). Since average velocity errors for altimetric observations of theCCS have been found to be on the order of 10 cm/s (Pascual et al. 2009), consistent withthe 2 cm perturbation level used in this study, this raises the question of how reliable theOW contours are for eddy identification using altimetric observations in the CCSwithout smoothing of the velocity field (Chelton et al. 2007). It should be noted thateven with the noisy fields, we found that the FTLE maxima are insensitive to theinterpolation scheme (cubic or linear), which we explore more in the next section.
4.3. Resolution sensitivity
To test the effects of resolution on LCS identification for CCS-sized eddies, weconstruct a high-resolution synthetic model. To this end, an inverse DFT isused to obtain a sea surface height field with three times better resolution than theAVISO data, or !0.1" (10 km) in both the latitudinal and longitudinal directions.From this higher resolution field, velocities are calculated using (9) and the FTLE fieldscomputed.
The velocities of the triple-resolution synthetic model are sub-sampled at increasinglypoorer spatial resolution and the LCS recalculated. We compare two synthetic cases,the triple resolution model described above to one with 2.3 times worse resolution thanthe best AVISO observations, i.e. !0.7" (70 km). Even at this coarse resolution the LCSare remarkably unperturbed, regardless of the interpolation scheme used. Thisinsensitivity is illustrated in figure 9, which depicts a typical temporal snapshot of theforward FTLE for the synthetic field with both the highest resolution, interpolated witha cubic scheme, and the lowest resolution, interpolated with the less accurate linearscheme. Here we see that, although the LCS are less smooth in some regions for thepoorer spatial resolution, they still retain their continuity. Remarkably, within a smallfraction of a degree, the LCS are in the same location in both realizations.
Figure 9. Typical snapshot of the forward finite-time Lyapunov (FFTLE) field for the synthetic model fieldsampled at 0.1" (a) and 0.7" (b) velocity resolution. The absolute difference between the final particle positions(after 24-day integration) is shown in (c) for the two realizations of the FTLE shown in (a) and (b). Althoughthe maximum difference in the particle positions is !500 km or !5", the LCS are relatively similar (less than0.5" different) between the two realizations.
LCS in the CCS 37
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

30
This insensitivity is more robust for the strongest LCS (those with the highest FTLEvalues), in accordance with theoretical results (Haller 2001).
Comparing the difference in particle positions at the final integration time for the twovelocity fields (figure 9(c)), we note that the error is greatest along the LCS. This is notsurprising since the forward-time LCS depicted here are repelling; thus along these theparticle trajectories exhibit sensitive dependence on initial conditions and relativedispersion is maximized. Whereas the maximum difference in the final particle positionsis !500 km (which is roughly 5" along a great circle), the error in the LCS is an order ofmagnitude less, around 0.5".
To give a clear picture of the sharpness of the FTLE maxima and their insensitivity toresolution, figure 10 illustrates a typical cross section of the FTLE fields describedabove. The locations of the velocity grid for both velocity resolutions are plotted (at thebottom) in order to compare them with the separation size between the sharp peaks ofthe FTLE field representing the LCS. With the exception of a tail of the poorlydeveloped LCS located at 37"N, all FTLE peaks produced with the 0.7" resolution areonly slightly perturbed in their locations relative to the 0.1" resolution. Also shown infigure 10 are the effects of the interpolation scheme. Although linear interpolation andcubic interpolation differ greatly for small values of the FTLE, at the maxima thedifference is relatively small.
Since LCS in the synthetic field are insensitive to resolutions worse than the bestavailable for the observational data, we now turn to analyzing the AVISO ADT field(section 3.1). Geostrophic velocities are calculated using the highest ADT resolutionavailable, 1/3", then sub-sampled to produce a 2/3" (!70 km) velocity field. Figure 11shows a detail of a representative section of the forward and backward LCS for bothresolutions. Whereas there is some effect on the LCS, the strongest structures (indicatedin black) are surprisingly robust to under-resolution. Notably, there is only a smallperturbation to both the attracting and repelling LCS for the small southern eddy,which has a diameter comparable to the grid spacing in the coarser resolution.
34 N 35 N 36 N 37 N 38 N 39 N 40 N 41 N 42 N 43 N 44 N 45 N
0
0.05
0.1
0.15
0.2
Latitude
FF
TLE
(da
ys1 )
0.1° FFTLE0.7° linear0.7° cubic0.7° grid0.1° grid
Figure 10. Transect of the forward finite-time Lyapunov exponent (FFTLE) fields shown in figure 9 along230"E. Also shown here is the effect of the interpolation scheme for the 0.7" velocity resolution synthetic fieldand the spacing of the velocity grid for the two resolutions (dots). Though the error between realizations isrelatively large in regions of lower FTLE, differences in the locations of FTLE maxima (the LCS) are minimalat poor resolution for both the cubic and linear interpolation algorithms.
38 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

31
In contrast, the smallest eddy centered at 39!N has completely vanished at the coarseresolution. Tests of the sensitivity of the LCS to interpolation scheme were repeated onthe observational field with the same results as in the synthetic field. Even in the 2/3!
field, the sharpest peaks of the FTLE fields indicating well-developed LCS were notsensitive to the interpolation scheme used.
Similar to the findings for spatial resolution, LCS were found to be insensitive tocoarse temporal resolution in both the observational and synthetic fields. Littledifference was found for large-scale structures at 14 day and finer temporal resolutionfor both the observational and synthetic fields (not shown), with the synthetic fieldinsensitive to even coarser sub-sampling than 14 days. This is surprising since thedecorrelation timescale of the SSH is "10 days for both fields. We do note that the eddyturnover time ("10 days for a particle to circle a 200 km eddy) is much less than the timefor the same eddy to propagate one eddy diameter ("70 days), i.e. the field is notchanging rapidly, at least for larger eddies. Therefore coarser temporal resolution thanthe decorrelation and eddy turnover times imply is needed to ‘‘break’’ the LCS.
Figure 11. Detail of backward FTLE (BFTLE) and FFTLE fields for 1/3! and 2/3! observation-basedvelocity fields. Surprisingly, most LCS are visible at the 2/3! velocity resolution, even the small eddy (centeredat 36.5!N) which is less than 1! in diameter. Note that the smallest eddy visible in the BFTLE field, centeredat 39!N, is not resolved at the coarser resolution.
LCS in the CCS 39
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

32
4.4. Statistics of the FTLE
It is known that Lyapunov exponents change with particle integration time(cf. Abraham and Bowen 2002, Bracco et al. 2004). In the classical definition of theLyapunov exponent, a positive value in the limit t!1 is an indication that a system(or a region of a system) is chaotic. This convergence can be clearly seen in the temporalevolution of the pdf of the forward FTLE fields through narrowing of the pdf over time(figure 12). We report here that the pdf is also dependent on the initial particle densityused to compute the FTLE. Whereas the temporal evolution of the spatially averagedFTLE is not dependent on the initial particle density, the maximum FTLE is (figure 13).Here we have calculated these statistics using the synthetic model described in section3.1 in the domain shown in figure 5, but we note that these results hold for all threesystems described in this article, including the kinematic model presented in section 2.
5. Summary and discussion
Assuming perfect observational data, the LCS method resolves many features of theCCS, such as the main jet of the California Current and eddy–eddy interactions, thatthe OW parameter does not. Given ideal observations, we do find for large well-developed westward propagating eddies in the CCS the OW parameter is capable ofeddy identification and is fairly congruent with transport barriers defined by the LCS.
Figure 12. Probability density function (pdf) of the FFTLE field for the domain shown in figure 5 using fourdifferent integration times t! jt2" t1j. The dashed line is the pdf for a lower initial particle density,corresponding to a particle separation of 0.05#, the solid for a higher initial particle density with a separationof 0.025#. The pdf narrows over time as the finite Lyapunov exponent converges to the classical Lyapunovexponent. The maximum value of the FTLE is reduced for larger initial particle separation for all integrationtimes.
40 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

33
This similarity breaks down when eddies are forming, are small, or interact stronglywith other features. We have also found that maximum FTLE values are affected by theinitial particle density.
We find that LCS are robust to fairly sparse spatial resolution, and to perturbationsto the model velocity field up to 20–30% or more of the maximum fluid velocity of thecoherent feature to be resolved. Owing to the level of errors in the best satellite altimetryproducts currently available, we conclude that the LCS method is robust only for verylarge and strong features in the CCS, and should be used with caution in regions of lowSSH variability and for small scale features. This suggests that altimetry observationsare not fine enough in space and time to resolve lobe dynamics (e.g. Samelson andWiggins 2006, Mendoza et al. 2010), at least in the CCS. In addition, we have foundthat the OW method of eddy identification is much more sensitive to noisy data thanthe FTLE method, and is unable to identify coherent features at uncertainty levelsexpected for altimetric observations of the CCS. Thus, this research highlights the needfor more adequate altimetry observations in low SSH variability regions in order toidentify mesoscale structures using any method.
Our finding that minima of the FTLE field are sensitive to velocity errors hasimportant implications for use of the recently proposed method of finding KAM-liketori (another type of coherent structure, e.g. Rypina et al. (2009)) in non-analyticalsystems. In this method coincident minima of both the forward and backwardFTLE are identified as jets or closed trajectories (Beron-Vera et al. 2010). Since theFTLE field is more sensitive at minima than maxima, the determination of theseminima may not be well constrained for noisy and/or poorly resolved velocity fields,and may be especially vulnerable for models using satellite data where the errors arerelatively large.
Figure 13. The spatially averaged, hFFTLEi (lower lines), and the maximum forward FTLE (upper lines)for the domain shown in figure 5, shown for both 0.025! and 0.05! initial grid spacings. The horizontal axis isthe integration time t" jt2# t1j for one realization of the FTLE field. Note the coincidence of the averageFTLE for both grid spacings.
LCS in the CCS 41
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

34
We have not considered the additional limitations of assuming geostrophy and 2Ddynamics for systems derived from altimetry, though these simplifications likely add tothe uncertainties. We also do not explore the limit of how fast flow patterns definedinstantaneously as eddies can propagate before transport boundaries are conceptuallyincongruent relative to those those found in steady state dynamics. We do, however,note that eddy phase velocities are in general observed to be an order of magnitude lessthan the fluid velocities in the global ocean (Chelton et al. 2007); therefore these resultsshould be applicable to many ocean regions.
Finally, because of the limits in the accuracy of altimetry measurements near thecoastline, we have excluded this dynamically and ecologically important region from ourdomain. Current efforts exist to improve remote observations of coastal waters boththrough advancements in altimetric processing and the construction and real-timeoperation of HF radar arrays. These new products will enable application of dynamicalsystem techniques to coastal regions using high-quality observations of surface dynamics.
Acknowledgements
We would like to thank C.A. Edwards and M. Mangel for their support. We would alsolike to thank A. Pascual for discussion of her results. This work was partially supportedby the Center for Stock Assessment Research (CSTAR) a partnership between NOAAFisheries Santa Cruz Laboratory and UCSC. Satellite altimetry products wereproduced by SSALTO/DUACS and distributed by AVISO, with support from CNES(http://www.aviso.oceanobs.com/duacs/).
References
Abraham, E.R. and Bowen, M.M., Chaotic stirring by a mesoscale surface-ocean flow. Chaos 2002, 12,373–381.
Beron-Vera, F.J., Olascoaga, M.J., Brown, M.G., Kocak, H. and Rypina, I.I., Invariant-tori-like Lagrangiancoherent structures in geophysical flows. Chaos 2010, 20, 017514.
Beron-Vera, F.J., Olascoaga, M.J. and Goni, G.J., Oceanic mesoscale eddies as revealed by Lagrangiancoherent structures. Geophys. Res. Lett. 2008, 35, 7.
Boffetta, G., Lacorata, G., Radaelli, G. and Vulpiani, A., Detecting barriers to transport: a review ofdifferent techniques. Physica D 2001, 159, 58–70.
Bracco, A., Provenzale, A. and Scheuring, I., Mesoscale vortices and the paradox of the plankton. P. Roy.Soc. B – Biol. Sci. 2000, 267, 1795–1800.
Bracco, A., von Hardenberg, J., Provenzale, A., Weiss, J. and McWilliams, J.C., Dispersion and mixing inquasigeostrophic turbulence. Phys. Rev. Lett. 2004, 92, 4.
Calil, P.H.R. and Richards, K.J., Transient upwelling hot spots in the oligotrophic North Pacific.J. Geo-phys. Res. 2010, 115, C02003.
Capet, X., McWilliams, J., Molemaker, M. and Shchepetkin, A., Mesoscale to submesoscale transition in theCalifornia Current system. Part II: frontal processes. J. Phys. Oceanogr. 2008, 38, 44–64.
Chelton, D.B., Schlax, M.G., Samelson, R.M. and de Szoeke, R.A., Global observations of large oceaniceddies. Geophys. Res. Lett. 2007, 34, L15606.
Coulliette, C., Lekien, F., Paduan, J.D., Haller, G. and Marsden, J.E., Optimal pollution mitigation inMonterey Bay based on coastal radar data and nonlinear dynamics. Environ. Sci. Technol. 2007, 41,6562–6572.
Dibarboure, G., Lauret, O., Mertz, F., Rosmorduc, V. and Maheu, C., SSALTO/DUACS User Handbook:(M)SLA and (M)ADT Near-Real Time and Delayed Time Products 2009. (Ramonville St Agne, France:AVISO).
42 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

35
d’Ovidio, F., Fernandez, V., Hernandez-Garcia, E. and Lopez, C., Mixing structures in the MediterraneanSea from finite-size Lyapunov exponents. Geophys. Res. Lett. 2004, 31, 1L17203.
d’Ovidio, F., Isern-Fontanet, J., Lopez, C., Hernandez-Garcia, E. and Garcia-Ladon, E., Comparisonbetween Eulerian diagnostics and finite-size Lyapunov exponents computed from altimetry in theAlgerian basin. Deep-Sea Res. Pt. I 2009, 56, 15–31.
Farnetani, C.G. and Samuel, H., Lagrangian structures and stirring in the Earth’s mantle. Earth Planet. Sci.Lett. 2003, 206, 335–348.
Haller, G., Distinguished material surfaces and coherent structures in three-dimensional fluid flows. PhysicaD 2001, 149, 248–277.
Haller, G., Lagrangian coherent structures from approximate velocity data. Phys. Fluids 2002, 14, 1851–1861.Haller, G. and Poje, A.C., Finite time transport in aperiodic flows. Physica D 1998, 119, 352–380.Haller, G. and Yuan, G., Lagrangian coherent structures and mixing in two-dimensional turbulence.
Physica D 2000, 147, 352–370.Hickey, B.M., Coastal oceanography of western north America from the tip of Baja California to Vancouver
Island. In The Sea – The Global Coastal Ocean, Regional Studies and Synthesis, edited by A. Robinsonand K. Brink, Vol. 11, chap. 12, pp. 345–393, 1998. (John Wiley and Sons, Inc.: New York).
Isern-Fontanet, J., Garcıa-Ladona, E. and Font, J., Identification of marine eddies from altimetric maps.J. Atmos. Ocean Technol. 2003, 20, 772–778.
Kai, E.T., Rossi, V., Sudre, J., Weimerskirch, H., Lopez, C., Hernandez-Garcia, E., Marsac, F. andGarcon, V., Top marine predators track Lagrangian coherent structures. Proc. Natl. Acad. Sci. USA2009, 106, 8245–8250.
Koszalka, I., Ceballos, L. and Bracco, A., Vertical mixing and coherent anticyclones in the ocean: the role ofstratification. Nonlinear Proc. Geoph. 2010, 17, 37–47.
Le Traon, P.Y., Larnicol, G., Guinehut, S., Pouliquen, S., Bentamy, A., Roemmich, D., Donlon, C.,Roquet, H., Jacobs, G., Griffin, D., Bonjean, F., Hoepffner, N. and Breivik, L.A., Data assembly andprocessing for operational oceanography: 10 years of achievements. Oceanography 2009, 22, 56.
Lehahn, Y., d’Ovidio, F., Levy, M. and Heifetz, E., Stirring of the northeast Atlantic spring bloom: aLagrangian analysis based on multisatellite data. J. Geophys. Res. 2007, 112, 15.
Lekien, F., Shadden, S.C. and Marsden, J.E., Lagrangian coherent structures in n-dimensional systems.J. Math. Phys. 2007, 48, 19.
Mancho, A.M., Small, D. and Wiggins, S., Computation of hyperbolic trajectories and their stable andunstable manifolds for oceanographic flows represented as data sets. Nonlinear Proc. Geoph. 2004, 11,17–33.
Marchesiello, P., McWilliams, J.C. and Shchepetkin, A., Equilibrium structure and dynamics of theCalifornia Current System. J. Phys. Oceanogr. 2003, 33, 753–783.
Martin, A., Phytoplankton patchiness: the role of lateral stirring and mixing. Prog. Oceanogr. 2003, 57,125–174.
Mathur, M., Haller, G., Peacock, T., Ruppert-Felsot, J.E. and Swinney, H.L., Uncovering the Lagrangianskeleton of turbulence. Phys. Rev. Lett. 2007, 98, 144502.
Mendoza, C., Mancho, A. and Rio, M., The turnstile mechanism across the Kuroshio current: analysis ofdynamics in altimeter velocity fields. Nonlinear Proc. Geoph. 2010, 17, 103–111.
Nencioli, F., Dong, C., Dickey, T., Washburn, L. and McWilliams, J.C., A vector geometry based eddydetection algorithm and its application to a high-resolution numerical model product and high-frequency radar surface velocities in the Southern California Bight. J. Atmos. Ocean Tech. 2010, 27,564–574.
Okubo, A., Horizontal dispersion of oatable particles in the vicinity of velocity singularities such asconvergences. Deep-Sea Res. 1970, 17, 445–454.
Olascoaga, M.J., Beron-Vera, F.J., Brand, L.E. and Kocak, H., Tracing the early development of harmfulalgal blooms on the West Florida Shelf with the aid of Lagrangian coherent structures. J. Geophys. Res.2008, 113, c12014.
Olcay, A.B., Pottebaum, T.S. and Krueger, P.S., Sensitivity of Lagrangian coherent structure identification toflow field resolution and random errors. Chaos 2010, 20, 017506.
Ottino, J.M., The Kinematics of Mixing: Stretching, Chaos, and Transport, 1989. (Cambridge; New York:Cambridge University Press).
Pascual, A., Boone, C., Larnicol, G. and Traon, P.Y.L., On the quality of real-time altimeter gridded fields:comparison with in situ data. J. Atmos. Ocean Tech. 2009, 26, 556–569.
Pascual, A., Faugere, Y., Larnicol, G. and Le Traon, P.Y., Improved description of the ocean mesoscalevariability by combining four satellite altimeters. Geophys. Res. Lett. 2006, 33, 611.
Pascual, A., Pujol, M.I., Larnicol, G., Le Traon, P.Y. and Rio, M.H., Mesoscale mapping capabilities ofmultisatellite altimeter missions: first results with real data in the Mediterranean Sea. J. Marine Syst.2007, 65, 190–211.
Pasquero, C., Bracco, A., Provenzale, A. and Weiss, J.B., Particle motion in a sea of eddies. In LagrangianAnalysis and Prediction of Coastal and Ocean Dynamics, edited by A. Griffa, A.D. Kirwan, A.J. Mariano,T. Ozgokmen and T. Rossby, p. 89, 2007. (Cambridge University Press: Cambridge; New York).
LCS in the CCS 43
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

36
Poje, A.C., Haza, A.C., Ozgokmen, T.M., Magaldi, M.G. and Garraffo, Z.D., Resolution dependent relativedispersion statistics in a hierarchy of ocean models. Ocean Model. 2010, 31, 36–50.
Ponte, R.M., Wunsch, C. and Stammer, D., Spatial mapping of time-variable errors in Jason-1 and TOPEX/Poseidon sea surface height measurements. J. Atmos. Ocean Technol. 2007, 24, 1078–1085.
Ramp, S.R., Davis, R.E., Leonard, N.E., Shulman, I., Chao, Y., Robinson, A.R., Marsden, J.,Lermusiaux, P.F.J., Fratantoni, D.M., Paduan, J.D., Chavez, F.P., Bahr, F.L., Liang, S., Leslie, W.and Li, Z., Preparing to predict: the second Autonomous Ocean Sampling Network (AOSN-II)experiment in the Monterey Bay. Deep-Sea Res. Pt. II 2009, 56, 68–86.
Rossi, V., Lopez, C., Hernandez-Garcia, E., Sudre, J., Garcon, V. and Morel, Y., Surface mixing andbiological activity in the four eastern boundary upwelling systems. Nonlinear Proc. Geoph. 2009, 16,557–568.
Rovegno, P.S., Edwards, C.A. and Bruland, K.W., Observations of a Kenai Eddy and a Sitka Eddy in theNorthern Gulf of Alaska. J. Geophys. Res. 2009, 114, C11012.
Rypina, I.I., Brown, M.G., Beron-Vera, F.J., Kocak, H., Olascoaga, M.J. and Udovydchenkov, I.A., On theLagrangian dynamics of atmospheric zonal jets and the permeability of the stratospheric polar vortex.J. Atmos. Sci. 2007, 64, 3595.
Rypina, I.I., Brown, M.G. and Kocak, H., Transport in an idealized three-gyre system with application to theadriatic sea. J. Phys. Oceanogr. 2009, 39, 675–690.
Samelson, R.M., Fluid exchange across a meandering jet. J. Phys. Oceanogr. 1992, 22, 431–440.Samelson, R.M. and Wiggins, S., Lagrangian Transport in Geophysical Jets and Waves: The Dynamical
Systems Approach, 2006. (New York: Springer).Shadden, S.C., Lekien, F. and Marsden, J.E., Definition and properties of Lagrangian coherent structures
from finite-time Lyapunov exponents in two-dimensional aperiodic flows. Physica D 2005, 212, 271–304.Shadden, S.C., Lekien, F., Paduan, J.D., Chavez, F.P. and Marsden, J.E., The correlation between surface
drifters and coherent structures based on high-frequency radar data in Monterey Bay. Deep-Sea Res.Pt. II 2009, 56, 161–172.
Simpson, J.J. and Lynn, R.J., A mesoscale eddy dipole in the offshore California Current. J. Geophys. Res.1990, 95, 13009.
Tang, W., Chan, P. and Haller, G., Accurate extraction of Lagrangian coherent structures over finite domainswith application to flight data analysis over Hong Kong International Airport. Chaos 2010, 20, 017502.
Tang, X.Z. and Boozer, A.H., Finite time Lyapunov exponent and advection–diffusion equation. Physica D1996, 95, 283–305.
Veneziani, M., Edwards, C.A., Doyle, J.D. and Foley, D., A central California coastal ocean modeling study:1. Forward model and the inuence of realistic versus climatological forcing. J. Geophys. Res. 2009, 114,16.
Waugh, D.W. and Abraham, E.R., Stirring in the global surface ocean. Geophys Res. Lett. 2008, 35, 5.Weiss, J.B., The dynamics of enstrophy transfer in two-dimensional hydrodynamics. Physica D 1991, 48,
273–294.Wiggins, S., Introduction to Applied Nonlinear Dynamical Systems and Chaos, 2003. (New York: Springer).Wiggins, S., The dynamical systems approach to Lagrangian transport in oceanic flows. Annu. Rev. Fluid
Mech. 2005, 37, 295–328.Worm, B., Sandow, M., Oschlies, A., Lotze, H. and Myers, R., Global patterns of predator diversity in the
open oceans. Science 2005, 309, 1365.
44 C. S. Harrison and G. A. Glatzmaier
Dow
nloa
ded
by [C
hery
l Har
rison
] at 1
3:39
01
Sept
embe
r 201
2

37
Chapter 3 The role of filamentation and eddy-eddy interactions in marine larval accumulation and transport
Cheryl S. Harrison1*, David A. Siegel2 and Satoshi Mitarai3
1Earth and Planetary Sciences Department, University of California, Santa Cruz, CA 95064, USA [email protected] 2Earth Research Institute and Department of Geography, University of California, Santa Barbara, California, 93106 USA 3Marine Biophysics Unit, Okinawa Institute of Science and Technology, Okinawa 904-0495, Japan
Publication status: in review in MEPS
Abstract
A coupled particle-tracking/ocean circulation model of an idealized eastern boundary
upwelling current is used to explore the mesoscale pathways of larval transport. We
find that simulated larvae are often organized into filaments found between mesoscale
eddies that correspond to attracting Lagrangian coherent structures (LCS). LCS are
material curves that map filamentation and transport boundaries, often corresponding
to the locations of sea surface temperature fronts. Filamentation and eddy-eddy
interactions aggregate larvae from many source regions and release times into small,
highly dense packets that can be transported back to the shelf. Larval densities in
these packets can be up to two orders of magnitude greater than initial release
densities near the coast and are robust to strong levels of random “swimming”

38
perturbations. This study suggests that coherent flow structures play an important role
in pelagic transport of marine larvae.
3.1 Introduction
Eastern boundary currents (EBCs) such as the California Current System (CCS)
contain many of the most productive fisheries in the world (e.g., Ryther 1969,
Chassot et al. 2010), and coastal waters in EBCs are important habitats and spawning
grounds for many species of fishes and invertebrates (Parrish et al. 1981, Love et al.
2002). The elevated rate of productivity in EBCs is due to coastal upwelling, where
equatorward coastal winds drive ocean surface transport offshore, bringing nutrient-
rich water to the surface, thus driving primary productivity. Coastal upwelling also
generates mesoscale coherent structures, notably upwelling filaments, jets and eddies
(Flament et al. 1985, Brink and Cowles 1991, Haynes et al. 1993). These structures
organize the transport of upwelled waters containing biologically important material
such as nutrients, plankton and marine propagules (e.g., larvae, eggs, spores; Parrish
et al. 1981, Larson et al. 1994, Rodriguez et al. 1999).
Ocean transport of propagules connects populations of many coastal marine species
(Levin 2006, Cowen and Sponaugle 2009). Consequently, understanding marine
propagule transport is important for a wide range of scientific and management
problems, from assessment of recruitment dynamics and population genetics to
fisheries management and marine conservation planning (e.g., Warner and Cowen

39
2002, Palumbi 2003, Botsford et al. 2009, Costello et al. 2010). Marine propagules
themselves are typically small (a few mm’s to a few cm’s) and require days to months
in the plankton to develop competency for their next life stage (Strathmann 1985,
Metaxis and Saunders 2009). While larvae of many species are capable of a variety
of behaviors that can alter their dispersal patterns (Shanks 1985, Sponaugle et al.
2002), they are also strongly affected by prevailing currents, where they are dispersed
over spatial scales from 100’s of m to 100’s of km (e.g., Jackson and Strathmann
1981, Kinlan and Gaines 2003, Siegel et al. 2003, Shanks 2009). Current variability
in EBCs is typically much stronger than the mean flow (Marchecello et al. 2003,
Harrison and Glatzmaier 2012) and depends on topographic, remote and seasonal
forcing (Haynes et al 1993, Hill et al. 1998, Marchecello et. al 2003, Marchecello and
Estrade 2009), making the effect of eddies on dispersal both important and highly
variable in space and time (Largier 2003, Mitarai et al. 2008, Siegel et al. 2008).
Larval transport has been commonly modeled as a diffusive process (e.g., Okubo
1980, Jackson and Strathmann 1981, Largier 2003), sometimes with the addition of a
simplistic mean flow advection (Roberts 1997, Gaylord and Gaines 2000, Siegel et al.
2003, Shanks 2009). This led to the idea of a Gaussian dispersion kernel to quantify
larval transport (e.g., Largier 2003, Siegel et al. 2003), a concept that has been used
extensively to inform ecosystem management practices (see Botsford et al. 2009 for a
review). However, observations of larval settlement across many oceanographic
regimes show that large, local, intermittent settlement events are common, and that

40
single events can dominate local recruitment for a year or more (Ebert and Russell
1998, Warner et al. 2000). This suggests the Gaussian kernel approach does not
satisfactorily represent larval transport processes on timescales relevant to marine
populations (Mitarai et al. 2008, Siegel et al. 2008), and that resolving recruitment
events driven by ocean variability is crucial to advancing understanding of dispersal
and connectivity (Warner et al. 2000, Largier 2003, Botsford 2009, Mitarai et al.
2009).
Recent approaches have utilized ocean circulation model flow fields, remote sensing
observations, as well as a growing number of in situ observations to characterize
larval transport and recruitment patterns more realistically than ever before (e.g.,
Bjorksted et al. 2002, Cowen et al. 2006, Aiken et al. 2007, Broitman et al. 2008,
Mitarai et al. 2009, Watson et al. 2010, Aiken et al. 2011, Drake et al. 2011, Kim and
Barth 2011, Woodson et al. 2012). These observations and models illustrate the large
role that coherent flow structures (such as filaments and eddies) play in the transport
of plankton, larvae, and other propagules. In particular, several recent studies have
highlighted the delivery of discrete “packets” of larvae back to the nearshore as an
important settlement process (e.g., Mitarai et al. 2008, Siegel et al. 2008), consistent
with observations of episodic recruitment (Warner et al. 2000). The temporally
intermittent and spatially heterogeneous patterns of larval settlement and connectivity
seen in these studies are thought to have important consequences for metapopulation
dynamics, species coexistence and optimal fisheries management (e.g., Berkley et al.

41
2010, Costello et al. 2010, Aiken and Navarette 2011, Bode et al. 2011, Watson et al.
2011a, 2011b, 2012). It has been hypothesized that the complex eddying dynamics of
the coastal ocean are driving packet formation and subsequent transport (Siegel et al.
2008); however, the physical details remain largely unclear.
The goal of this study is to explore the processes that control the generation and
movement of filaments and larval packets. One of the hallmarks of ocean turbulence
is eddy-eddy interaction and merger (McWilliams 1984, Bracco et al. 2004), moving
energy from small to large scales and resulting in eddies up to 200 kilometers in
diameter that can persist for weeks to years (Marchecello et al. 2003, Marchecello
and Estrade 2009, Harrison and Glatzmaier 2012). During eddy merger smaller
eddies are entrained and filamented along the edges of larger eddies (McWilliams
1984, Harrison and Glatzmaier 2012). Here we find eddy interaction results in
creation of dense, often long filamental packets of larvae that can bring together
larvae of many ages and source regions into a small area.
Our modeling system was created to replicate mesoscale dynamics off the Central
California coast (Mitarai et al. 2008, Siegel et al. 2008), but it captures much of the
dynamics common to all eastern boundary upwelling currents. The modeling
framework consists of a three-dimensional ocean model, a particle-tracking model to
keep track of simulated larvae, and an analytical tool from dynamical systems theory:
Lagrangian coherent structures (LCS). LCS are used as a metric to map the attractors

42
of the filamentation process, to visualize transport boundaries and eddy interactions,
and provide a framework for understanding the kinematics of ocean transport
(d’Ovidio et al. 2004, Wiggins 2005, Samelson and Wiggins 2006, Beron-Vera et al.
2008, Shadden 2012). Our hypothesis is that attracting LCS (ALCS) will mark the
dominant transport pathways of marine propagules. That is, larvae will be
preferentially concentrated in “larval superhighways” mapped by ALCS, as opposed
to within eddy interiors (where there are generally few LCS), or more regularly in
some sort of average, diffusive regime. To the best of our knowledge, the utility of
LCS have yet to be explored in the context of marine propagule transport, although
LCS have been the focus of many recent studies of pollutant and biological transport
in marine systems (e.g., Lehahn et al. 2007, 2011; Olascoaga et al. 2008; Shadden et
al. 2009; Rossi et al. 2009; Tew Kai et al. 2009; d’Ovidio et al. 2010; Calil and
Richards 2010, Katija et al. 2011, Huhn et al. 2012).
The paper is organized as follows. Section 3.2 introduces LCS and highlights
concepts relevant to the findings here. Section 3.3 outlines the ocean and particle
tracking model used in this study. Section 3.4.1 reports our findings on the connection
between filamentation and LCS, 3.4.2 explores the relationship between filaments,
LCS and fronts, and 3.4.3 contains results on how the filamentation and eddying
processes affect age class and source region accumulation, highlighting the role of
eddy-eddy interactions in concentrating larvae. Section 3.5 is a discussion of our
findings, including material demonstrating robustness of filaments and packets to

43
swimming, and outlines directions for future study and application. The paper is
summarized in Sec. 3.6.
3.2 Lagrangian Coherent Structures (LCS)
Our goal in this section is to provide a brief conceptual introduction to LCS and to
make a few specific points relevant to the methods and results presented here. We
refer the interested reader to the excellent reviews by Wiggins (2005) and Shadden
(2012), the textbook by Samelson and Wiggins (2006) and an online tutorial
(Shadden 2005) for more detailed information and examples.
Lagrangian coherent structures (LCS) can be thought of as material lines on the ocean
surface that act as transport barriers and map regions of active filamentation,
effectively allowing us to visualize the skeleton of turbulent transport (e.g., Haller
2002; d’Ovidio et al. 2004; Shadden et al. 2005, 2012; Mathur 2007; Beron-Vera et
al. 2008). Broadly, particles on a surface will move toward and along an attracting
LCS (ALCS) and away from and along a repelling LCS (RLCS). LCS are also ideally
particle trajectories, so a particle will remain on an LCS once it encounters it
(supplemental movie 3.1).
Trajectories of water parcels near attracting and repelling LCS associated with a
saddle (or stagnation) point are illustrated in Fig. 3.1a. Arrows indicate the
instantaneous direction of movement of water parcel trajectories within in the fluid.

44
Figure 3.1 Schematics of fluid/particle behavior near Lagrangian coherent structures (LCS). Attracting (blue) and repelling (red) LCS often intersect at saddle points. Fluid initially surrounding the saddle point at t1 (gray circle, left) stretches out along the attracting LCS, forming a long, thin filament by time t3. Nearby fluid parcels (black) move along hyperbolic trajectories (solid black lines), remaining relatively unstretched through time. Attracting and repelling LCS surrounding an eddy (right) are commonly nearly coincident, often forming a complex tangle of intersections (see also Fig. 3.2a and supplemental movie 3.1). The blue lines represent ALCS, trajectories separating fluid parcels (on either side),
that at this moment are close to each other but in the recent past were much farther
apart. That is, there is confluence of fluid particle trajectories near the ALCS (which
is not necessarily a horizontal convergence or downwelling). The red lines in Fig.
3.1a represent RLCS, trajectories separating fluid parcels that will soon separate, that
is experience diffluence (and not necessarily divergence or upwelling). Near
Lagrangian saddle points—the intersection of repelling and attracting LCS—fluid
stretches out in time along the ALCS into a long, thin filament (Fig. 3.1a, gray, see
also supplemental movie 3.1). In this way the gradient of a passive tracer field can be
!"#"
$$%&'()*+(
,-./01&'()*+(
23(24(25(

45
intensified by the filamentation process without any horizontal convergence or
associated downwelling. For buoyant particles (as used here) or similarly any
planktonic material that maintains its position in the water column, any horizontal
convergence (i.e., downwelling) will intensify the process of filamentation and lead to
stronger gradients in tracer fields (e.g., Genin et al. 2005, Bakun 2010).
How are LCS calculated? Fluid parcels nearby but not on a LCS (Fig. 3.1a, black
ovals, also see supplemental movie 3.1) move along and asymptotically toward the
ALCS and away from the RLCS on hyperbolic trajectories (black lines). Along these
trajectories neighboring fluid parcels are strained differing amounts by the flow field.
These differences in stretching rates allow the detection of LCS by calculating
Lyapunov exponents, which quantify the rate that neighboring particles separate.
Here we use the finite-time Lyapunov exponent (FTLE) (Haller 2002, Shadden et al.
2005), although other methods have been developed (d’Ovidio et al. 2004, Mancho et
al, 2006). In essence, the FTLE is a scalar field that measures the time-averaged local
exponential rate of particle separation. The FTLE is calculated by seeding the fluid
with a dense grid of passive tracer particles, integrating their trajectories, and tracking
the separation of initially nearby particles through time (the relative dispersion).
Particles on either side of an LCS will diverge at an exponential rate as they approach
the saddle point, i.e.,
Δx(x,T) = Δx(t0) exp(λT),

46
where Δx(x,T) is the spatial separation between initially nearby particles at time T, T
= t-t0 is the particle trajectory integration time, and Δx(t0) the initial separation. The
local finite-time Lyapunov exponent is found by solving for the exponent λ, which is
a function of the initial particle location x(t0), T, and the starting time t0,
λ(x(t0),T,t0) = 1/|T| ln(||Δx(x,T)/Δx(t0)||),
where the norm of the deformation gradient ||Δx(x,T)/Δx(t0)|| is found by identifying
the square root of the maximal singular value (i.e., the maximum stretching factor) at
each initial particle location (Haller 2002, Shadden et al. 2005). LCS are identified
from FTLE scalar fields as high values or ridges. Repelling LCS are found by
evolving particle trajectories forward in time (T > 0) while attracting LCS by
evolving particle trajectories backward in time (T < 0). Longer integration times
effectively grow the LCS away from the saddle points, heightening and thinning the
LCS (e.g., Fig. 2.5). Here we use an initial particle separation of 500 m and an
integration time of 24 days, more than long enough to grow the LCS so that the
relevant flow features are visualized.
The implication of LCS being material curves is that fluid moving along an LCS will
stay on this curve and not pass across it (supplemental movie 3.1). Thus, ideally LCS
act as transport barriers for passive particles and tracers advected by the large-scale
flow (Shadden et al. 2005). LCS can map long-lived fronts separating ocean water
masses with different characteristics (e.g., temperature, salinity, oxygen). While

47
temperature or density fronts imply a transport barrier is present, the converse is not
necessarily true, and LCS may map transport barriers that are not associated with
fronts. However for ocean flows in near-geostrophic balance, LCS and fronts will
likely coincide.
Repelling LCS are often found nearly coincident with attracting LCS, especially
around eddies (Figs. 3.1b, 3.2a). This coincidence should be expected. For time-
invariant systems, attracting LCS associated with one saddle point often form the
repelling LCS of another saddle point (heteroclinic trajectories), or even circle back
and attach to same saddle point (homoclinic trajectories). Thus an LCS can be both
attracting and repelling at the same time, with respect to different source material
(Samelson and Wiggins 2006). However, this connectivity breaks up with
perturbation (Wiggins 2005, Samelson and Wiggins 2006). In general we expect
attracting and repelling LCS on the ocean surface should in many cases be close to
each other, but not coincide exactly (Fig. 3.1b, 3.2a). This will become important in
the statistical analysis to follow.
3.3 Models
The CCS-in-a-box model, implemented in the Regional Ocean Modeling System
(ROMS) (Shchepetkin and McWilliams 2005), was designed to replicate the
mesoscale eddy dynamics of central California coastal circulation in a simplified

48
Figure 3.2 Comparison of Eulerian and Lagrangian surface fields. Here we show (a) Lagrangian coherent structures (LCS), (b) larval density (larvae/km2), (c) sea surface temperature (SST, color) with sea surface height (SSH, contours), and (d) surface horizontal velocity (arrows) and near-surface (color, second model level ~5 m depth, positive upwelling) vertical velocity for day 37 in model run 111. In (a) blue lines represent attracting LCS, red lines repelling LCS. Larval density (b) is binned to 1 km squares and shown in a log scale as larvae are highly concentrated in narrow regions; simulated larvae were released inshore of the dashed line. The SSH interval is 1 cm. Larvae are primarily located in filaments that correspond to attracting LCS, SST fronts and enhanced vertical velocities on either side of the filament. Transects along bold black lines are shown in Fig. 3.3 (north transect) and Fig. 3.4 (south transect).

49
domain (Mitarai et al. 2008, Siegel et al. 2008). The three-dimensional model domain
features a north/south periodic (wrap-around) boundary, an open western boundary,
and a narrow shelf (~10km) along a straight eastern boundary coastline. A constant
alongshore pressure gradient is imposed within the model which creates a poleward
undercurrent. The model is forced by a variable wind stress that replicates the
temporal wind variability off Central California during summer upwelling (a typical
July). A total of 28 runs with different wind stress time courses (with identical wind
stress statistics) were created to represent interannual variability in upwelling forcing.
Implementation details for the CCS-in-a-box model can be found in Mitarai et al.
(2008) and Siegel et al. (2008).
Marine propagules are modeled as quasi-Lagrangian tracer ocean particles originating
from the shelf region but are constrained to remain on the top ocean model layer.
These modeled propagules are meant to represent larvae that prefer to stay near the
surface, a common strategy (e.g. Love et al. 2002). This simplifies the transport
dynamics into a horizontal, two-dimensional, time-dependent problem. For both the
larval transport and LCS calculations, tracer particle positions are integrated in time
using a fourth-order Runge-Kutta scheme with the ROMS-simulated surface
velocities (saved every 6 hrs) interpolated to the particle locations each integration
time step. Larvae (i.e., tracer particles) are released daily from day 1 to day 170
within the shelf zone (<10 km offshore) on a regular 2 km-grid and tracked from the
time of release until the end of each 180-day run; effectively we are assuming all

50
shelf areas are larval source regions and that larvae are continuously released.
Because larval mortality is not modeled, the number of larvae (and the range of ages
since release of those larvae) increases with time. Each tracer particle is considered to
potentially represent a large number of actual larvae because individuals of many
species release thousands to millions of larvae at one time (e.g., Love et al. 2002;
Mitarai et al. 2008, 2009). Here we will use larva, particle and propagule
interchangeably with the understanding that we mean modeled, surface-following
particle for each of these terms.
3.4 Results
3.4.1 LCS, filamentation and larval density patterns
Flow features produced in the model (Fig. 3.2) broadly resemble those observed, both
off California (Flament et al. 1985, Brink and Cowles 1991) and in other eastern
boundary currents (Haynes et al. 1993, Hill et al. 1998). Instabilities in the upwelling
jet produce eddies and upwelling filaments, which are fast, relatively narrow (on the
order of 10’s of km wide) features with coherent flow reaching up to ~100 meters
deep into the water column that can quickly move material offshore (Flament et al.
1985). As these upwelling filaments and other flow features interact with the eddy
field offshore, they are stirred and filamented further, resulting in a patchy
distribution of larvae.

51
Figure 3.3 North transect. Transects of model surface fields across a filament spiraling into an eddy (upper bold line in Fig. 3.2b). Larval abundance (a) is high in narrow regions corresponding to attracting Lagrangian coherent structures (LCS, light gray vertical bars). LCS are found by identifying maxima of the finite-time Lyapunov exponent (FTLE, c). The sea surface temperature (SST) gradient (dashed line in b) is high where LCS separate cold and warm water. Vertical velocities (d) from the second model level (∼5m depth) are strongest on either side of the strongest temperature fronts (here at 180 km alongshore) with positive vertical velocity (upwelling) on the warmer side of the front as the ageostrophic circulation attempts to stratify the front.
Similarly patchy density distributions are seen throughout this model run
(supplemental movie 3.2) as well as all other runs. Regions of high larval density are
observed to correspond with attracting LCS, but only sometimes with repelling LCS
(Fig. 3.2a & 3.2b). Coincidence of filaments, LCS and surface variables is illustrated
in Figs. 3.3 & 3.4, where transects are taken across two regions (bold vertical lines in
Fig. 3.2b). Fig. 3.3 transects a filament in three places as it spirals into a cyclonic

52
eddy. Fig. 3.4 transects two filaments, one filament is crossed twice as it is swept
shoreward around an elongated eddy. Transects of the forward and backward FTLE
fields are plotted in Figs. 3.3c and 3.4c; attracting LCS are marked with light gray
bars and plotted on other panels. Regions of high density are found on or near ALCS
(Figs. 3.3a & 3.4a). Conversely not all ALCS correspond to regions of high larval
abundance, as they are not connected to a region where particles were released (cf.
Figs. 3.4a & 3.4c). Regions of high larval concentration only sometimes coincide
with RLCS, high values of the forward FTLE field (red in Fig. 3.2a, short-dashed
lines in Figs. 3.3c & 3.4c), with coincidence mainly occurring when the RLCS are
close to ALCS (e.g., Fig. 3.4c at 50 km).
To assess the statistical relationship between LCS and filaments, statistics of the
forward (FFTLE) and backward (BFTLE) FTLE fields throughout the model domain
were compared to the values of these fields interpolated to particle positions. We
asked the following: 1) How often are larvae on an LCS? and 2) Which type of LCS
do they end up on? Complementary cumulative distribution functions for backward
and forward FTLE values are shown in Fig. 3.5a for a single day (day 37, also shown
in Fig. 3.2) and in Fig. 3.5b for a 100-day ensemble. Cumulative distribution
functions for the FTLE values are calculated for the entire domain (gray lines in Figs.
3.5a & b) and for where particles are located (black lines). Pairwise comparisons of
cumulative distribution functions (BFTLE vs. BFTLE at particle positions, FFTLE
vs. FFTLE at particle positions, and BFTLE at particle positions vs. FFTLE at

53
Figure 3.4 South transect. Transects of model surface fields across two filaments (lower bold line in Fig. 3.2b). Model fields are as in Fig. 3.3. Note that not every attracting LCS (gray bars) corresponds to a filament containing coastal material (a), and that some regions of larval filamentation do not correspond with temperature fronts but are still marked by LCS (here at 50 km alongshore).
particle positions) were done using a Kolmogorov–Smirnov test. All pairs of
distributions were significantly different (p < 0.001), as expected given the large
number of data points (N > 104 for all distributions).
Particles were determined to be on or sufficiently near an LCS when the FTLE values
at their positions were above 0.2 day−1 (shown in Fig. 3.3c and Fig. 3.4c as a
horizontal dashed line). This threshold is determined experimentally and depends on
the ocean dynamics and the FTLE integration time. FTLE field statistics at the

54
particle positions were compared to FTLE statistics for the entire domain. Larvae
were more likely on an LCS (40-60%) than if they were randomly distributed (~25%
of all larvae expected for day 37; Fig. 3.5a). Furthermore for day 37, larvae were
much more likely to be on attracting LCS (~60% of all larvae) than on repelling LCS
(~40% of all larvae) consistent with the arguments presented in Sec. 3.2. Extending
the analysis to a 100-day ensemble (Fig. 3.5b), larvae were more likely on LCS (25-
40%) than if they were randomly distributed (~20%), and again more likely on
attracting LCS (~40% of all larvae), than on repelling LCS (~25% of all larvae). This
suggests filamentation mapped by LCS plays an important role in transport of
material originating over the shelf in EBCs.
3.4.2 Filaments and fronts
In the CCS-in-a-box model, larval filaments marked by LCS are often found between
cyclonic (cold-core) and anti-cyclonic (warm-core) eddies (Figs. 3.2-3.4). Hence
larval filaments are also associated with high sea surface temperature (SST) gradients,
consistent with larval surveys in the CCS (e.g., Larson et al. 1994). However, the
transects in Fig. 3.4 show some cases where LCS and high particle densities are not
coincident with a SST front (e.g., at 50 km alongshore in Fig. 3.4). To test the
relationship of LCS and SST fronts we asked: If a particle is on an LCS, how much
more likely is it to be on a front? Fig. 3.5c compares the complementary cumulative
distribution of the horizontal (surface) SST gradient magnitude |∇hSST| with the
distribution at all particle positions and for particles near LCS (FTLE > 0.2 day−1)

55
over the 100-day ensemble. Particles were more likely to be on higher values of
|∇hSST| than if randomly distributed (e.g. 40% above 0.1oC/km, ~25% expected), and
particles on or near LCS were even more likely to be on high values (50-60% above
0.1oC/km, twice as many as expected; p < 0.001 for all comparisons). This suggests
larvae originating over the shelf and those on LCS in particular are more likely to be
on fronts.
Regions of high SST gradients are observed to be associated with locally enhanced
vertical velocities on the second model layer (∼5 m) on either side of the front (and
thus LCS, Figs 3.2d, 3.3d, 3.4d), consistent with a secondary ageostrophic circulation
attempting to stratify the front (e.g., Capet et al. 2008; Klein and Lapeyre, 2009).
Frontal vertical velocities are difficult to correlate statistically with SST fronts (and
by extension LCS and filaments) since they are maximal at either side of the front and
not along it (Capet et al. 2008). Possible implications are discussed in Sec. 3.5.2.
Figure 3.5 Filament and frontal statistics. Comparison of backward (BFTLE) and forward (FFTLE) finite-time Lyapunov exponent statistics for (a) the day shown in previous figures, and (b) a 100-day ensemble. Also shown are the SST gradient statistics (hSST, in oC/km) for the 100-day ensemble (c). Cumulative distribution functions have been summed from the highest to lowest values. LCS are here defined by values of the FTLE fields greater than 0.2 days-1 (vertical lines). In general particles are more often on LCS than if they are randomly distributed (gray lines), and especially more likely to be on attracting LCS (black solid lines) than repelling LCS (black dashed lines). Likewise particles are more likely to be on temperature fronts (high hSST) than if they were randomly distributed (c), and even more likely when they are on attracting LCS (ALCS) or repelling LCS (RLCS).

56

57
3.4.3 Eddy-eddy interactions, source regions and larval age distributions
The ages of larvae along filaments are generally older the farther out to sea they are
from their initial release location, as has been observed in situ (Rodriguez et al.
1999). In Fig. 3.3 each of the three filament crossings are at a different oceanographic
(or integrated path) distance from the shelf. Larvae are shown in two age classes, 0-20
days old and 20 days and older (Fig. 3.3a). Along the filament, a higher abundance of
older larvae is found in regions that have a greater oceanographic distance from
shore. This age pattern is also seen in the southern transect (Fig. 3.4a), where both
filaments are farther in a path sense from the source region than in the eddy transect.
Here larvae are older, with the age gradient of larvae along the twice-crossed filament
increasing with oceanographic distance from shore.
In contrast to these simple examples, the interaction of larval filaments and eddies can
result in complex density and age structure patterns. As an example of this process, a
time series of eddy interaction, merging, and larval particle packet formation is shown
in Fig. 3.6. Here a 25 km diameter cyclonic eddy is formed initially near the shelf
Figure 3.6 Packet formation. The interaction of anticylonic eddies over model days 91-103 leads to the formation of a dense accumulation of larvae in a narrow filamental packet. Top row: density of all age classes (particles/km2, shown in log scale). Middle row: attracting (blue) and repelling (red) LCS. Bottom row: sea surface height (SSH) with 0.5 cm contour interval. Note this is a subregion of the model domain shown in other figures. Numbers in the lower left panel refer to eddies mentioned in the text. A transect of larval density is shown in Fig. 3.7 along the black line in the upper right panel.
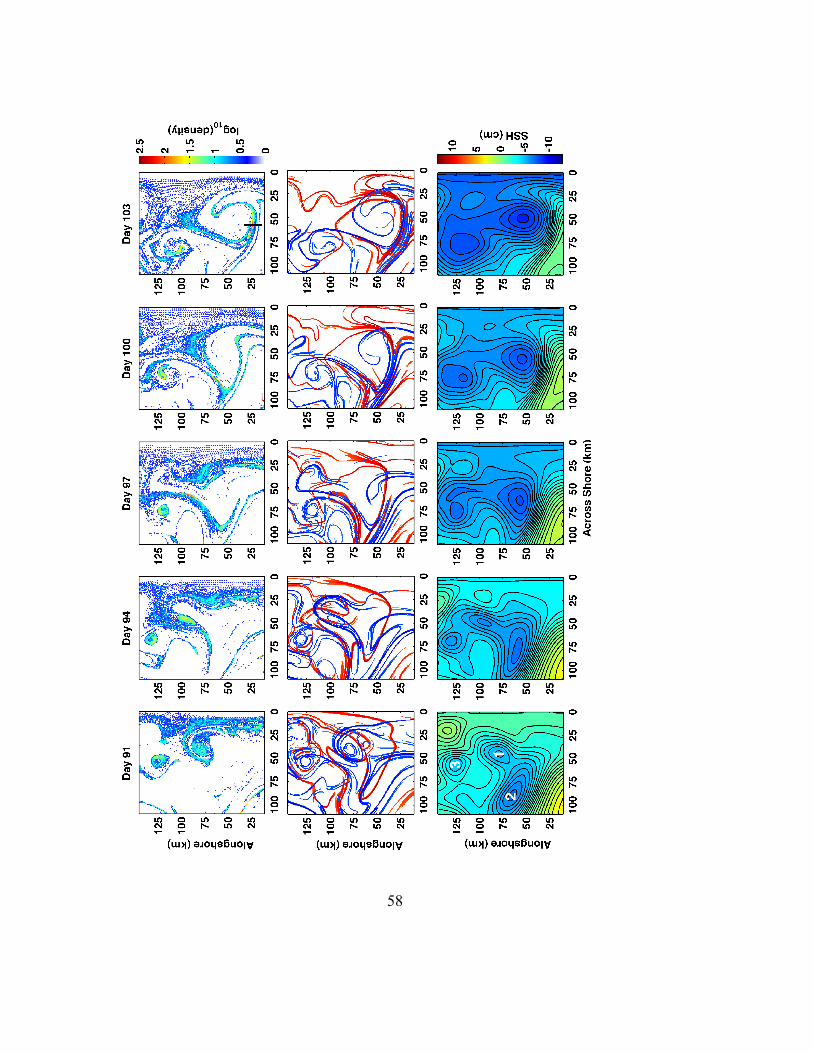
58

59
with a mass of simulated larvae spiraling into its center (day 91, eddy 1). This small
eddy interacts with a similarly sized eddy (eddy 2), and a larger cyclonic eddy (eddy
3) over a ten-day period (days 94-103), which can be visualized by the particle
density field, the LCS locations and the sea surface height (SSH; Fig. 3.6). During the
interaction, both particle density and LCS distributions show that the contents of eddy
1 are compressed and filamented around the periphery of the larger eddy, resulting in
a thin, dense packet of material that was formerly the core of eddy 1 being transported
back towards the coast along the larger eddy’s exterior. In contrast, the SSH
signatures of the two small eddies appear to merge during the interaction (Fig. 3.6,
day 97), illustrating that what is observed in the Eulerian frame (SSH) does not
necessarily determine the transport fate of larval particles (the time-integrated
Lagrangian flow), as has often been noted in the dynamical systems literature (i.e.,
Shadden et al. 2005). The packet has a maximal density of over 90 particles/km2,
roughly two orders of magnitude greater than the initial particle release density (<1
larvae/km2). Note that many LCS are swept together and combined as the packet is
formed, and the resulting large number of collinear attracting LCS are coincident with
striations of dense material (Fig. 3.6, day 103).
Packets formed by eddy interactions can result in a wide range of larval age classes
from many source locations being concentrated in a very small spatial area. The dense
packet shown in Fig. 3.6 (black line on day 103) contains larvae ranging from less
than 20 to more than 60 days old (Fig. 3.7a). The material in the densest portion of

60
Figure 3.7 Packet age class and source region distribution. Transect of larval density (a) from 56 km offshore on model day 103 (along the vertical black line in the upper right panel in Fig. 3.6) with age class makeup (in days, grayscale). Ages range from less to 20 days to more than 60 days within a narrow spatial region. Larvae in the densest part of the larval packet (22-23 km alongshore, 92 particles) come from a wide range of source locations (b), where source regions have been unwrapped to account for transport across the periodic boundary in the ocean circulation model. Bin size for this histogram in (b) is 10 km; the vertical axis is the number of particles from each source region.

61
the packet (22-23 km alongshore in Fig. 3.7a), representing one square km bin in Fig.
3.6, came from sources up to 50 km south and 220 km north of the packet location on
day 103 (Fig. 3.7b), where the source regions have been unwrapped to account for the
periodic boundary in the ocean circulation model. Notably some of this material came
from south of its current location, having traveled against the average southward
alongshore flow.
Filamentation can lead to a quick trip offshore, decreasing transit time away from the
shelf. For example, on day 177 particles ranging from 7-29 days old are found over
150 km offshore concentrated in a filament that wraps around the periodic model
boundary (Fig. 3.8a, denoted by X’s). Conversely, filaments often bend back toward
the coast or get folded into a persistent retention zone (i.e., an eddy), resulting in
older larvae being transported back towards the shelf. Inshore of the filament
described above are many regions of older larvae (30-120+ days old, Fig. 3.8b-e); we
will focus on two of these areas, marked in boxes in Fig. 3.8f. Histograms of source
Figure 3.8 Density distributions by age class on model day 177. Shown are ages 7-29 days (a), 30-59 days (b), 60-89 days (c), 90-119 days (d), 120 days and over (e), and all age classes (e). Larval density (particles/km2) is shown on log scale to facilitate visualization. Note the feature centered at 100 km offshore and 175 km alongshore (dashed box in f) has integrated particles from many different age classes (b-e), and that some particles are near the coast (<50 km) even after 120 days (e, solid box in f). In contrast young larvae (a) have reached upwards of 150 km offshore in a fast moving filament (denoted by X’s) that wraps around the periodic model boundary. See supplemental movie 3.2 for visualization of density through the model run. Age and source distributions for the boxed regions are shown in Fig. 3.9.

62

63
locations and age distributions for the two boxed regions are shown in Fig. 3.9.
For the coherent structure on day 177 (dashed box in Fig. 3.8f), again a large number
of larvae with a diverse range of ages have been concentrated into a small region (see
supplemental movie 3.2 for animation). There are over 15,000 particles in this
structure, 16% of the total number of particles in the entire domain at this time.
Within the coherent structure particles range from 40 to 170 days old and are from
source regions spanning 150 to over 800 km alongshore (Fig. 3.9a-b). Note that in
Fig. 3.9a the tails of the distribution are not completely shown and some larvae
originate from source locations more than 1200 km alongshore. These particles with
distant sources have crossed the periodic domain four times, representing distant
poleward sources. Some larvae older than 120 days are within 50 km of the coast
(solid box in Fig. 3.8f). This packet contains particles ranging from 20 to 170 days
old and from -10 to 500 km alongshore (Fig. 3.9c-d). Thus eddy interaction may
result in transport of material back to the shelf (and for some larvae back to their
source locations) long after release.
In general, while the filamentation and eddy merging process is complex and takes
variable forms, dense packets resulting from eddy-eddy interaction and filamentation
are found throughout this model run (see supplemental movies), and all other model
runs. To assess the average statistics of packet formation and their associated larval
age and source distributions, we define packets as 1 km square regions. Note this is

64
Figure 3.9 Source and age distributions. Histograms of source locations (a,c) and age distributions (b,d) for the two boxed regions in Fig. 3.8f. Here in both examples larvae from a wide range of sources and release times are found in the same region. The coherent structure (a-b, dashed box in 3.8f) has integrated larvae originating from 150 to over 800 km alongshore (a), and contains particles ranging from 40 to 170 days old. A coastal packet (c-d, solid box in 3.8f) has particles that came from -10 to 600 km alongshore, and are less than 30 to over 175 days old. Source locations have been unwrapped as in Fig. 3.7, with a bin size of 50 km. Age bins are 10 days wide. N is the total number of particles in the histogram. Note the difference in vertical scales for each plot.
much smaller than the features described above. Figure 3.10 shows the
complementary cumulative distribution for density in these 1 km bins interpolated to
particle positions for all of model run 111. In most cases particles are in packets many
times denser than their release densities (<1 km-2; i.e. P = 0.54 for density greater
than 5 km-2). Average age and source histograms were determined for very dense

65
packets (density >50 km-2, P = 0.06, gray shading in Fig. 3.10, N = 8140 packets) by
Figure 3.10 Particle density and packet statistics. Complementary cumulative distribution of density at particle locations shows that over the model run 6% (P = 0.059) of particles are found in packets (shaded gray, defined by density > 50 in a 1 km2 bin). Combined histograms (with source and age averages removed for each packet) show a wide range of source regions (inset top) and ages (inset bottom) for larvae in packets. Horizontal axes on histograms show a portion of the long tails for each distribution.
subtracting out the averages for each packet and combining (Fig. 3.10 inset, N =
7x105 particles in total). Standard deviations are 73 km for source regions and 14
days for ages in packets, with long tails for both histograms as seen in previous
examples. As in Fig. 3.8, larger packets are made of many of these smaller dense
packets, which could result in a wider range of age and source regions for larger

66
features (as in Fig. 3.9). These results suggest that on average dense regions of larvae
will contain larvae from a wide range of source locations and release times. Since
eddy interaction processes are similar throughout the global ocean (McWilliams
1984, Bracco et al. 2004), similar packets are expected be found across a wide range
of marine ecosystems.
3.5 Discussion
3.5.1 Application of LCS
It may be possible to use LCS operationally for tracking regions of dense planktonic
accumulation. Given availability of high-quality ocean surface velocity fields (i.e. HF
radar, operational data assimilative coastal ocean models), attracting LCS can be
assessed for connection to spawning regions and tracked through time (i.e. Shadden et
al. 2009). However, instead of just a diagnostic metric it may prove more powerful to
use LCS as a conceptual tool to understand the kinematic framework that larval
transport dynamics rests in, as LCS make the connection between filamentation,
transport boundaries and frontal processes. Perhaps more importantly, theorems from
dynamical systems can help predict the sensitivity of ecological processes associated
with LCS.
For example, while the effects of swimming are difficult to model, involving species-
specific assumptions about cues, swimming duration, speed and direction (Metaxas

67
Figure 3.11 Effect of swimming on density patterns. Density distributions are shown as in Fig. 3.8f with addition of random-walk type swimming at two levels: (a) 1 cm/s and (b) 10 cm/s standard deviation. As expected given the structural stability of LCS, dense packets
and Saunders 2009), we can predict that LCS will still delineate important transport
pathways for slow swimmers. Since LCS are structurally stable (Haller 2002,
Harrison and Glatzmaier 2012), i.e. largely insensitive to perturbations, it follows that
up to a certain swimming speed in any direction, or for short, strong bursts of
swimming, larvae will effectively remain on LCS. To provide a rough demonstration
of this stability, Fig. 3.11 shows the effect of normally distributed, random-walk
horizontal “swimming” on the density patterns shown in Fig. 3.8f. The random-walk
perturbations were added with a standard deviation σ of 1 cm/s and 10 cm/s. For both
cases the horizontal swimming direction and magnitude was randomized at each
particle time step. For a perturbation of σ = 1 cm/s density patterns remain close to
their original distributions (compare Figs. 3.8f & 3.11a). At a perturbation level of σ

68
= 10 cm/s (much higher than the expected swimming speeds of most early stage
larvae; e.g., Love et al. 2002) packets and filaments are still in evidence; maximum
concentrations have been reduced, but are still an order of magnitude higher than the
release density (Fig. 3.11b). Thus we expect filaments and eddies are robust
aggregators of planktonic and nektonic organisms under a wide range of behavioral
strategies.
3.5.2 Comparisons with observations and prior studies
The present analyses linking processes of filamentation and larval distributions
should be widely applicable as upwelling dynamics are similar across all EBCs (e.g.,
Haynes et al. 1993, Hill et al. 1998). For example, neritic larvae and coastal
planktonic species are persistently found in upwelling filaments and their associated
eddies in the Canary EBC (Rodriguez et al. 1999, Bécognée et al. 2009), and this is
thought to be an important larval transport pathway from the mainland to offshore
islands. In the CCS, the transport of propagules in filaments is consistent with the
observed higher abundance of larvae at fronts (Larson et al. 1994, Bjorkstedt et al.
2002), though to our knowledge these two phenomena have not been previously
connected. Packet formation by eddy-eddy interaction, filamentation and transport
back to the shelf may explain the delivery of discrete settlement pulses of larvae that
contain a wide range of age classes and species, and also contribute to shorter than
expected dispersal distances (Warner et al. 2000, Warner and Cowen 2002, Shanks

69
2009). These observations may be explained to leading order by ocean transport
alone, without invoking complex larval behavior or life histories.
Larval concentration in filaments may explain regions of predictably higher
settlement downstream from headlands (Ebert and Russell 1988), as well as
preferential spawning areas observed at submerged promontories (Karnauskas et al.
2011). Instead of adding more variability, a complex coastline will often organize the
flow into more regular and predictable patterns. When an alongshore current reaches
a coastal headland it is directed offshore, often forming a filament or eddy (e.g.,
Haynes et al. 1993), and so can regularly generate high larval concentration, making
these areas advantageous spawning grounds (Karnauskas et al. 2011). If this
concentrated material were directed back to shore it would result in a large, perhaps
quick settlement pulse over a narrow coastal area, as seen in modeling studies
downstream from promontories (Kim and Barth 2011). Such settlement pulses are
expected to be correlated with frontal arrival, as has been observed in the CCS
(Woodson et al. 2012).
Recent modeling studies suggest packet delivery back to the shore is a common
phenomenon. Packet delivery will manifest in connectivity diagrams (Cowen et al.
2006, Siegel et al. 2008) as a linear feature, indicating a wide range of source regions
were deposited at one arrival location (as in Fig. 3.7). This pattern is prominent in
many connectivity studies (Aiken et al. 2011, Drake et al. 2011, Kim and Barth

70
2011). However, it is unclear from these studies if linear connectivity patterns are due
to single events or accrue over longer timescales. Since packet formation appears to
be a consequence of the eddy-eddy interaction and filamentation processes, we expect
it will occur across all ocean regions and likely generates the patterns seen in these
studies.
Biophysical and trophic interactions along coastal upwelling filaments and fronts
could influence larval survivability. Primary production is observed to be higher in
upwelling filaments, as nutrient-rich water is carried offshore (Brink and Cowles
1991, Hill et al. 1998), and also along fronts, where both planktonic and fish biomass
can be enhanced (e.g., Landry et al. 2012). In addition, frontal vertical velocities due
to submesoscale instabilities have been shown in coupled physical-ecosystem models
to induce nutrient injection and increase productivity, but also induce export materials
from the euphotic zone (see Lévy 2008 and Klein and Lapeyre 2009 for reviews).
Further seabirds, fish and other marine animals are known to use fronts for foraging
and efficient transport (e.g., Fiedler and Bernard 1987, Tew Kai et al. 2009, Munk et
al. 2010), indicating increased predation susceptibility for larvae at fronts. Thus larval
survivability on filaments will be controlled by a combination of enhanced fitness due
to food availability, higher mortality due to vertical export, or locally enhanced
predation.

71
Many EBCs have wider shelves than the California coastline, such as those off Africa
and the Iberian Peninsula. For these systems, upwelling can be centered at the shelf
break creating a kinematic boundary between a poleward shelf flow and the
equatorward upwelling jet (Peliz et al. 2002). This on-shelf/shelf break flow
partitioning appears to depend on the details of both the pycnocline and wind forcing
(Estrade et al. 2008), and has been observed to act as a retention mechanism for river
outflow and very nearshore species (Santos et al. 2004). Similarly, observations (Hill
et al. 1998) and more realistic models of the CCS often exhibit poleward flow close to
the coast (e.g., Drake and Edwards 2011). We speculate that retention within this
poleward flow, similar to the studies off the Iberian Peninsula, may limit the offshore
extent of many intertidal and very nearshore spawners (Shanks and Sherman 2009),
and perhaps contribute to the shorter than expected dispersal distances observed for
some species (Shanks 2009). To our knowledge it remains unclear what the temporal
dynamics, response to wind variation, and three-dimensional kinematics are for this
type of retention mechanism. However the dynamical systems approach to identifying
and classifying kinematic boundaries should be useful for diagnosing larval retention
in these systems.
The breakdown of fronts by small eddies and meanders is readily seen in satellite
observations (Flament et al. 1985); in ROMS these frontal instabilities are known to
intensify with increasing model resolution (Capet et al. 2008). Such instabilities
would likely lead to more mixing along filaments (Badin et al. 2011), but to our

72
knowledge has yet to be studied in the context of marine ecology. However, in situ
measurements suggest that frontal accumulation of planktonic organisms in filaments
is robust in the extension from model to reality (Larson et al. 1994, Rodriguez et al.
1999).
3.5.3 Potential Roles of Larval Behavior
Many species’ larvae and eggs are found preferentially near the ocean surface (e.g.,
Shanks 1985, Rodriguez et al. 1999, Bjorkstedt et al. 2002, Love et al. 2002, Metaxas
and Saunders 2009). Our particle model formulation assumes propagules swim or
float to overcome than the modest model vertical velocities (<5 m/d; Figs. 3.2-3.4),
i.e. prefer to be in the productive euphotic zone. If particles were free to move
vertically this could result in a lesser degree of density enhancement; instead of
aggregating at the surface, non-buoyant particles will be pushed down by surface
convergence and downwelling (e.g., Bakun 2010). Though modeled vertical
velocities here are instantaneously small, the accumulated effect over time can be
large and will increase density patchiness (Zhong et al. 2011). Our analysis of
transport boundaries and particle filimentation also applies to any floating material
originating over the shelf (pollutants, macrophytes, etc.), and aggregations of these
materials can form pelagic habitats for a variety of marine organisms (Shanks 1985,
Dempster and Taquet 2004), indicating a possible secondary effect on larval
distribtutions.

73
Previous results show ontogenetic vertical migration does not have strong effects on
the larval settlement statistics in the CCS-in-a-box model (Mitarai et al. 2008, Siegel
et al. 2008). Upwelling filaments are generally coherent up to ~100 m depth (e.g.,
Brink and Cowles 1991), so it follows that with even a moderate change in vertical
position larvae will remain within these flow structures. In contrast, regions with
generally high vertical velocity shear, for example estuaries (North et al. 2008) or
systems with bifurcations in major currents, such as off the north coast of Norway
(Fiksen et al. 2007), will be more sensitive to larval behavior. A common feature of
EBCs is vertical shear between the coastal equatorward upwelling jet and the
poleward undercurrent (Hill et al 1998), and larval transport in this region is known to
be affected by diurnal vertical migration (Aiken and Navarette 2011). It still remains
largely unclear where, when and to what extent vertical swimming will be important
given realistic flow fields.
3.6 Summary
Larvae released above the shelf in an idealized EBC are often transported in filaments
that coincide with LCS. Larvae are more likely to be found on an attracting LCS
(~40% over a 100-day ensemble) than on repelling LCS (~25%), and more likely on
LCS than if they were randomly distributed (~20% expected). Larvae on LCS are also
more likely to be on SST fronts, although this is not always the case. Eddy-eddy
interaction and filamentation processes result in the creation of dense packets, which

74
bring larvae of many age classes and source regions together into a small spatial
region. Results are consistent with observations of larval distributions and settlement
patterns in EBCs and other systems. Packet formation as a manifestation of eddy
interaction and filamentation processes should occur globally. These packets
sometimes get transported back to the coast, and the associated pattern of
connectivity is seen in other studies. LCS are insensitive to a moderate degree of
perturbation (e.g., swimming) and so are expected to be robust in mapping transport
pathways under a variety of larval behavioral strategies.
3.7 Acknowledgements
Input from G. Glatzmaier and P. Raimondi were helpful in focusing this project, as
well as overall support and writing assistance from R. Warner, K. Brink, A. Payton
and E. Silver. Four anonymous reviewers supplied thoughtful critiques that greatly
improved the manuscript. This project was supported in part by NSF grant OCE-
030844 as part of the Flow, Fish and Fishing Biocomplexity in the Environment
project, by NSF grant OCE-1155813, by the Center for Stock Assessment Research
(CSTAR), a partnership between NOAA Fisheries Santa Cruz Laboratory and UCSC,
and by IGPP Santa Cruz.

75
3.8 Bibliography
Aiken CM, Navarrete CM, Castillo M, Castilla JC (2007) Along-shore larval dispersal kernels in a numerical ocean model of the central Chilean coast. Mar Ecol Prog Ser 339:13
Aiken CM, Navarrete SA (2011) Environmental fluctuations and asymmetrical dispersal: generalized stability theory for studying metapopulation persistence and marine protected areas. Mar Ecol Prog Ser 428:77– 88
Aiken CA, Navarrete SA, and Pelegrí JL (2011) Potential changes in larval dispersal and alongshore connectivity on the central Chilean coast due to an altered wind climate. J Geophys Res 116(G4):G04026
Badin G, Tandon A, Mahadevan A (2011) Lateral mixing in the pycnocline by baroclinic mixed layer eddies. J Phys Oceanogr 41(11):2080–2101
Bakun A (2010) Linking climate to population variability in marine ecosystems characterized by non-simple dynamics: conceptual templates and schematic constructs. J Mar Syst 79:361–373
Bécognée P, Moyano M, Almeida C, Rodríguez J, Fraile-Nuez E, Hernández-Guerra A, Hernández-León S (2009) Mesoscale distribution of clupeoid larvae in an upwelling filament trapped by a quasi-permanent cyclonic eddy off Northwest Africa. Deep-Sea Res Part I 56(3):330–343
Berkley HA, Kendall BE, Mitarai S, Siegel DA (2010) Turbulent dispersal promotes species coexistence. Ecol Lett 13(3):360–371
Beron-Vera FJ, Olascoaga MJ, Goni GJ (2008) Oceanic mesoscale eddies as revealed by Lagrangian coherent structures. Geophys Res Lett 35(12):7
Bjorkstedt EP, Rosenfeld LK, Grantham BA, Shkedy Y, Roughgarden J (2002) Distributions of larval rockfishes Sebastes spp. across nearshore fronts in a coastal upwelling region. Mar Ecol Prog Ser 242:215–228
Bode M, Bode L, Armsworth PR (2011) Different dispersal abilities allow reef fish to coexist. Proc Nat Acad Sci 108(39):16317–16321,
Botsford LW, Brumbaugh DR, Grimes C, Kellner JB, Largier J, O’Farrell MR, Ralston S, Soulanille E, Wespestad V (2009) Connectivity, sustainability, and yield: bridging the gap between conventional fisheries management and marine protected areas. Rev Fish Biol Fish 19(1):69–95

76
Bracco A, von Hardenberg J, Provenzale A, Weiss JB, McWilliams JC (2004) Dispersion and mixing in quasigeostrophic turbulence. Phys Rev Lett 92(8):4
Brink KH, Cowles TJ (1991) The coastal transition zone program. J Geophys Res 96(C8):14637–14647
Broitman BR, Blanchette CA, Menge BA, Lubchenco J, Krenz C, Foley M, Raimondi PT, Lohse D, Gaines SD (2008) Spatial and temporal patterns of invertebrate recruitment along the west coast of the United States. Ecol Monogr 78(3):403–421
Calil PHR, Richards KJ (2010) Transient upwelling hot spots in the oligotrophic North Pacific. J Geophys Res 115(C2):C02003
Capet X, McWilliams JC, Molemaker MJ, Shchepetkin AF (2008) Mesoscale to submesoscale transition in the California Current system. Part II: Frontal processes. J Phys Oceanogr 38(1):44–64
Chassot E, Bonhommeau S, Dulvy NK, Mélin F, Watson R, Gascuel D, Le Pape O (2010) Global marine primary production constrains fisheries catches. Ecol Lett 13(4):495–505
Costello C, Rassweiler A, Siegel DA, De Leo G, Micheli F, Rosenberg A (2010) The value of spatial information in MPA network design. PNAS 107(43):18294–18299
Cowen RK, Paris CB, Srinivasan A (2006) Scaling of connectivity in marine populations. Science 311(5760):522–527
Cowen RK, Sponaugle S (2009) Larval dispersal and marine population connectivity. Annu Rev Mar Sci 1:443–466
Dempster T, Taquet M (2004) Fish aggregation device (FAD) research: gaps in current knowledge and future directions for ecological studies. Rev Fish Bio Fish 14(1):21–42
d’Ovidio F, De Monte S, Alvain S, Dandonneau Y, Lévy M (2010) Fluid dynamical niches of phytoplankton types. PNAS 107(43):18366
d’Ovidio F, Fernández V, Hernández-García E, López C (2004) Mixing structures in the Mediterranean Sea from finite-size Lyapunov exponents. Geophys Res Lett 31(17)
Drake PT, Edwards CA, Barth JA (2011) Dispersion and connectivity estimates along the U.S. west coast from a realistic numerical model. J Mar Res, 69(1):1–37

77
Ebert TA, Russell MP (1988) Latitudinal variation in size structure of the west coast purple sea urchin: a correlation with headlands. Limnol Oceanogr 33(2):286–294
Estrade P, Marchesiello P, Verdiere D, Colin A, Roy C (2008) Cross-shelf structure of coastal upwelling: A two dimensional extension of Ekman’s theory and a mechanism for inner shelf upwelling shut down. J Mar Res 66(5):589–616
Fiedler PC, Bernard HJ (1987) Tuna aggregation and feeding near fronts observed in satellite imagery. Cont Shelf Res 7(8):871–881
Fiksen Ø, Jørgensen C, Kristiansen T, Vikebø F, Huse G (2007) Linking behavioural ecology and oceanography: larval behaviour determines growth, mortality and dispersal. Mar Ecol Prog Ser 347:195–205
Flament P, Armi L, Washburn L (1985) The evolving structure of an upwelling filament. J Geophys Res 90(11):765–11
Gaylord B, Gaines SD (2000) Temperature or transport? range limits in marine species mediated solely by flow. Am Nat 155(6):769
Genin A, Jaffe J, Reef R, Richter C, Franks P (2005) Swimming against the flow: a mechanism of zooplankton aggregation. Science 308(5723):860
Haller G (2002) Lagrangian coherent structures from approximate velocity data. Phys Fluids 14(6):1851–1861
Harrison CS, Glatzmaier GA (2012) Lagrangian coherent structures in the California Current System–sensitivities and limitations. Geophys Astrophys Fluid Dyn 106(1):22–44
Haynes R, Barton E, Pilling I (1993) Development, persistence, and variability of upwelling filaments off the Atlantic Coast of the Iberian Peninsula. J Geophys Res 98(C12):22681–22692
Hedgecock D (1994) Does variance in reproductive success limit effective population sizes of marine organisms? In: Beaumont AR (ed) Genetics and evolution of aquatic organisms. Chapman and Hall, London, UK
Hill AE, Hickey BM, Shillington FA, Strub PT, Brink KH, Barton ED, Thomas AC. (1998) Eastern ocean boundaries coastal segment. In: Robinson AR, Brink, KH (eds). The Sea, Vol. 11. John Wiley & Sons Inc, London
Huhn F, von Kameke A, Pérez-Muñuzuri V, Olascoaga MJ, and Beron-Vera FJ (2012) The impact of advective transport by the South Indian Ocean Countercurrent on the Madagascar plankton bloom. Geophys Res Lett 39(L06602).

78
Jackson GA, Strathmann RR (1981) Larval mortality from offshore mixing as a link between precompetent and competent periods of development. Am Nat 118(1):16-26
Karnauskas M, Chérubin LM, Paris CB (2011) Adaptive significance of the formation of multi-species fish spawning aggregations near submerged capes. PLoS ONE 6(7):e22067
Katija K, Beaulieu WT, Regula C, Colin SP, Costello JH, Dabiri JO (2011) Quantification of flows generated by the hydromedusa Aequorea victoria: a Lagrangian coherent structure analysis. Mar Ecol Prog Ser, 435:111–123
Kim S, Barth JA (2011) Connectivity and larval dispersal along the Oregon coast estimated by numerical simulations. J Geophys Res 116(C6)
Kinlan B, Gaines S (2003) Propagule dispersal in marine and terrestrial environments: a community perspective. Ecology 84(8):2007–2020
Klein P, Lapeyre G (2009) The oceanic vertical pump induced by mesoscale and submesoscale turbulence. Annu Rev Mar Sci 1:351–375
Landry MR, Ohman MD, Goericke R, Stukel MR, Barbeau KA, Bundy R, Kahru M (2012). Pelagic community responses to a deep-water front in the California Current Ecosystem: overview of the A-Front study. J Plank Res, in press.
Largier JL (2003) Considerations in estimating larval dispersal distances from oceanographic data. Ecol App 13(sp1):71–89
Larson RJ, Lenarz WH, Ralston S (1994) The distribution of pelagic juvenile rockfish of the genus Sebastes in the upwelling region off central California. Calif Coop Ocean Fish Invest Rep 35:175– 221
Lehahn Y, d’Ovidio F, Lévy M, Heifetz E (2007) Stirring of the northeast Atlantic spring bloom: A Lagrangian analysis based on multisatellite data. J Geophys Res 112(C8):C08005
Lehahn Y, d’Ovidio F, Lévy M, Amitai Y, Heifetz E (2011) Long range transport of a quasi isolated chlorophyll patch by an Agulhas ring. Geophys Res Lett 38(16):L16610
Levin L (2006) Recent progress in understanding larval dispersal: new directions and digressions. Inta Comp Bio 46(3):282
Lévy M (2008) The modulation of biological production by oceanic mesoscale turbulence. In: Weiss JB, Provenzale A (eds). Transport and Mixing in Geophysical Flows, Springer

79
Love M, Yoklavich M, Thorsteinson L (2002) The rockfishes of the northeast Pacific. Univ of Cal Press
Mancho A, Small D, Wiggins S (2006) A tutorial on dynamical systems concepts applied to Lagrangian transport in oceanic flows defined as finite-time data sets: Theoretical and computational issues. Phys Rep, 437(3-4):55–124
Marchesiello P, McWilliams JC, Shchepetkin AF (2003) Equilibrium structure and dynamics of the California Current System. J Phys Ocean, 33(4):753–783
Marchesiello P, Estrade P (2009) Eddy activity and mixing in upwelling systems: a comparative study of Northwest Africa and California regions. Int J Earth Sci 98(2):299–308
Mathur M, Haller G, Peacock T, Ruppert-Felsot J, Swinney H (2007) Uncovering the Lagrangian skeleton of turbulence. Phys Rev Lett 98(14):144502
McWilliams JC (1984) Emergence of isolated coherent vortices in turbulent flow. J Fluid Mech 146:21–43
Metaxas A, Saunders M (2009) Quantifying the “bio-” components in biophysical models of larval transport in marine benthic invertebrates: advances and pitfalls. Biol Bull 216(3):257
Mitarai S, Siegel DA, Winters KB (2008) A numerical study of stochastic larval settlement in the California Current system. J Mar Syst 69:295–309
Mitarai S, Siegel DA, Watson JR, Dong C, McWilliams JC (2009) Quantifying connectivity in the coastal ocean with application to the Southern California Bight. J Geophys Res 114
Morgan SG, Fisher JL, Miller SH, McAfee ST, and Largier JL (2009) Nearshore larval retention in a region of strong upwelling and recruitment limitation. Ecology, 90(12):3489–3502
Munk P, Hansen MM, Maes GE, Nielsen TG, Castonguay M, Riemann L, Sparholt H, Als TD, Aarestrup K, Andersen NG, et al. (2010) Oceanic fronts in the Sargasso Sea control the early life and drift of Atlantic eels. Proc R Soc B 227(1700):3593–3599
North EW, Schlag Z, Hood RR, Li M, Zhong L, Gross T, Kennedy VS (2008) Vertical swimming behavior influences the dispersal of simulated oyster larvae in a coupled particle-tracking and hydrodynamic model of Chesapeake Bay. Mar Ecol Prog Ser 359(99):115

80
Okubo A (1980) Diffusion and ecological problems: mathematical models. Springer-Verlag, Berlin
Olascoaga MJ, Beron-Vera FJ, Brand LE, Koçak H (2008) Tracing the early development of harmful algal blooms on the West Florida Shelf with the aid of Lagrangian coherent structures. J Geophys Res 113(c12):c12014
Palumbi S (2003) Population genetics, demographic connectivity, and the design of marine reserves. Ecol App 13(sp1):146–158
Parrish RH, Nelson CS, Bakun A (1981) Transport mechanisms and reproductive success of fishes in the California Current. Biol Oceanogr 1(2):175–203
Peliz Á, Rosa T, Santos A, Pissarra J (2002) Fronts, jets, and counter-flows in the Western Iberian upwelling system. J Mar Sys 35(1-2):61–77
Roberts C (1997) Connectivity and management of Caribbean coral reefs. Science, 278(5342):1454–1457
Rodriguez JM, Hernández-León S, Barton ED (1999) Mesoscale distribution of fish larvae in relation to an upwelling filament off Northwest Africa. Deep-Sea Res Part I 46(11):1969–1984
Rossi V, López C, Hernández-García E, Sudre J, Garçon V, Morel Y (2009) Surface mixing and biological activity in the four Eastern Boundary Upwelling Systems. Nonl Proc Geophys 16(4):557–568
Ryther JH (1969) Photosynthesis and fish production in the sea. Science 166(3901):72
Samelson RM, Wiggins S (2006) Lagrangian transport in geophysical jets and waves: the dynamical systems approach. Springer, New York, N.Y.
Santos AMP, Peliz A, Dubert J, Oliveira PB, Angélico M, Ré P (2004) Impact of a winter upwelling event on the distribution and transport of sardine (Sardina pilchardus) eggs and larvae off Western Iberia: a retention mechanism. Cont Shelf Res 24(2):149–165
Shadden SC (2005) LCS tutorial. Accessed 22-Mar-2012. http://mmae.iit.edu/shadden/LCS-tutorial/
Shadden SC (2012) Lagrangian coherent structures. In: Grigoriev R, Schuster HG (eds). Transport and Mixing in Laminar Flows: From Microfluidics to Oceanic Currents, Wiley

81
Shadden SC, Lekien F, Marsden JE (2005) Definition and properties of Lagrangian coherent structures from finite-time Lyapunov exponents in two-dimensional aperiodic flows. Phys D 212(3- 4):271–304
Shadden SC, Lekien F, Paduan JD, Chavez FP, Marsden JE (2009) The correlation between surface drifters and coherent structures based on high-frequency radar data in Monterey Bay. Deep-Sea Res Part II 56(3-5):161–172
Shanks AL (1985) Behavioral basis of internal-wave-induced shoreward transport of megalopae of the crab Pachygrapsus crassipes. Mar Ecol Prog Ser, 24:289–295
Shanks AL (2009) Pelagic larval duration and dispersal distance revisited. The Bio Bull 216(3):373
Shanks AL, Shearman RK (2009) Paradigm lost? Cross-shelf distributions of intertidal invertebrate larvae are unaffected by upwelling or downwelling. Mar Ecol Prog Ser 385:189–204
Shchepetkin AF, McWilliams JC (2005) The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Model 9(4):347–404
Siegel DA, Kinlan BP, Gaylord B, Gaines SD (2003) Lagrangian descriptions of marine larval dispersion. Mar Ecol Prog Ser 260:83–96
Siegel DA, Mitarai S, Costello CJ, Gaines SD, Kendall BE, Warner RR, Winters KB (2008) The stochastic nature of larval connectivity among nearshore marine populations. PNAS 105(26):8974
Sponaugle S, Cowen R, Shanks A, Morgan W, Leis J, Pineda J, Boehlert G, Kingsford M, Lindeman K, Grimes C, et al. (2002) Predicting self-recruitment in marine populations: biophysical correlates and mechanisms. Bull Mar Sci, 70(Sup 1):341–375
Strathmann R (1985) Feeding and nonfeeding larval development and life-history evolution in marine invertebrates. Ann Rev Ecol Sys 16:339–361
Tew Kai E, Rossi V, Sudre J, Weimerskirch H, López C, Hernández-García E, Marsac F, Garçon V (2009) Top marine predators track Lagrangian coherent structures. PNAS 106(20):8245–8250
Warner R, Cowen R (2002) Local retention of production in marine populations: evidence, mechanisms, and consequences. Bull Mar Sci 70(1):245–249

82
Warner R, Swearer S, Caselle J (2000) Larval accumulation and retention: implications for the design of marine reserves and essential habitat. Bull Mar Sci 66(3):821–830
Watson J, Mitarai S, Siegel DA, Caselle JE, Dong C, and McWilliams JC (2010) Realized and potential larval connectivity in the Southern California Bight. Mar Ecol Prog Ser, 401:31–48
Watson J, Hayes CG, Raimondi PT, Mitarai S, Dong C, McWilliams JC, Blanchette CA, Caselle JE, Siegel DA (2011a) Currents connecting communities: Nearshore community similarity and ocean circulation. Ecology, 92, 1193–1200
Watson J, Siegel DA, Kendall BE, Mitarai S, Rassweiler A, Gaines SD, (2011b) Identifying critical regions in small-world marine metapopulations, Proc Nat Acad Sci USA, 108:E907-E913.
Watson J, Kendall BE, Siegel DA, Mitarai S (2012) Changing seascapes, stochastic connectivity and marine metapopulation dynamics. Amer Nat 180:99-112
Wiggins S (2005) The dynamical systems approach to Lagrangian transport in oceanic flows. Annu Rev Fluid Mech 37:295–328
Woodson CB, McManus MA, Tyburczy JA, Barth J, Washburn L, Caselle JE, Carr MH, Malone DP, Raimondi PT, Menge BA, Palumbi SR (2012) Coastal fronts set recruitment and connectivity patterns across multiple taxa. Lim Ocean, 57(2):582–596
Y. Zhong, A. Bracco, and T. A. Villareal. Pattern formation at the ocean surface: Sargassum distribution and the role of the eddy field. Lim Ocean: Fluid Enviro, 2:12–27, 2012.

83
Chapter 4 The tattered curtain hypothesis revisited: coastal jets control larval settlement patterns
Cheryl S. Harrison1 and David A. Siegel2
1 Earth and Planetary Sciences Department, University of California, Santa Cruz, CA
95064, USA [email protected]
2 Earth Research Institute and Department of Geography, University of California,
Santa Barbara, California, 93106 USA
Target journal: ASLO Fluids and Environments
Abstract
The details of upwelling dynamics and retention of coastal waters is important for
understanding dispersal of coastally originating material and thus dynamics of marine
ecosystems. Motivated by observations of larval recruitment patterns, a hypothesis
was put forward in the 1980’s describing the upwelling front as a “tattered curtain,”
retaining coastally released larvae, but broken up by filaments and eddies. This
transport barrier was expected to respond to local wind forcing by moving offshore in
complex meanders, moving back to the coast under upwelling relaxation. Here we
revisit and revise this hypothesis using an idealized ocean model of an eastern
boundary upwelling current driven by realistically temporally varying winds. In the

84
model, coastal retention patterns are strongly controlled by the upwelling jet,
resulting in significant along-shore coherence in settlement patterns when the jet is
near the coast. Modeled particle density is highest near the core of the jet, bounded by
high velocity shear lines on either side, consistent with dynamical systems theory of
jet cores as retentive barriers. Using an ensemble of model runs, we show empirically
that potential settlement is highest after periods of extended moderate upwelling.
Extended strong upwelling “tatters” the upwelling jet, driving material far offshore in
complex patterns. Settlement is best explained by 20-day integrated alongshore wind
(r = 0.62). Settlement is only marginally explained by upwelling wind relaxation
events (r = 0.33). Retention in coastal jets should be a widespread phenomenon as
indicated by numerous studies consistent with the results here.
4.1 Introduction
One of the central problems in marine ecology is the temporal and spatial dynamics
affecting transport of coastally spawned species. Coastal upwelling ecosystems,
though representing only a small extent of the ocean surface, contain a
disproportionate number of the ocean’s fisheries (Ryther 1969). Along the coast,
recently upwelled water brings nutrients to the surface where they can be utilized by
phytoplankton, providing energy to coastal ecosystems. Many coastal species are
benthic – remaining near the sea floor or in intertidal areas for their adult lives. Many
benthic species have a pelagic (open ocean) larval or juvenile stage. A common

85
reproductive strategy is to spawn thousands to millions of larvae that drift or swim
with the currents, feeding in the plankton for days to months until gaining
competency to recruit back to suitable habitat, forming the next generation. This
spawning strategy is utilized across diverse species, from tiny barnacles up to large
groundfish: animals inhabiting the kelp beds, rocky substrate, and intertidal habitats
all along the coastal shelf and slope. Many commercially exploited benthic species
renew their populations very slowly, and so are overfished to the point of being
threatened or endangered. Understanding the lifecycle dynamics of these fisheries,
especially the poorly understood pelagic stage, is important for appropriate
management. Retention over the shelf during the larval competency window insures
that spawn is not swept far offshore away from suitable habitat, and a growing
number of observations suggest that this limited dispersal due to retention is much
more common than previously thought (Warner and Cowen 2002, Shanks and
Sherman 2009). However, the degree to which oceanography determines settlement
patterns remains poorly understood.
For benthic species, distribution of spawn near suitable habitat after some residence
time in the plankton, often called pelagic larval duration (PLD), determines the
success of animals recruiting to the adult stage, and thus the strength of the
population for that spawning season. To explain the observed pulses of recruitment of
barnacle larvae along the coast, a hypothesis was put forward in the 1980’s that
linked dynamics of the upwelling front to settlement pulses (Roughgarden et al. 1988,

86
1991). In this paradigm, the front between recently upwelled water near the coast and
warmer water offshore was considered a convergence zone where planktonic
organisms were aggregated. The distance of this upwelling front from the shore was
thought to be determined largely by the strength of the upwelling winds, carrying
larvae far from shore when upwelling was strong and back towards shore during
relaxation, resulting in a large settlement pulse when the upwelling front collides with
the coast. This hypothesis was thought to be especially relevant for intertidal species
along the Central California coast: in this region the shelf is narrow, the upwelling
front close to shore and many species spawn during the spring to fall upwelling
season. The details of this hypothesis are best appreciated in the original text
(Roughgarden et al. 1991):
“We imagine, somewhat simplistically, the upwelling front as a ‘tattered
curtain’ hanging before an open window. It is tattered because it is punctured
by offshore squirts such as those typical of Point Arena, Point Reyes, and
Point Sur. It does not move as rigid iron curtain in response to the upwelling
strength, but instead is pushed and pulled locally, billowing in toward shore in
some spots while remaining far from shore at others. Thus, during a short
period of relaxation, small parts of the upwelling front may touch the coast
leading to local recruitment. But during an extended relaxation, long sections
of the front collide with the shore leading to nearly synchronous recruitment
across long stretches of the coast. Moreover, the correlation (of say, 1 day lag)

87
between upwelling-causing winds and recruitment is fussed by the distance of
the front from shore: if the front is far from shore, as in the spring, and
extended relaxation is needed to produce a recruitment pulse, but if the front
has gradually moved closer to shore, as in the summer, a short relaxation may
effect a recruitment pulse. Furthermore, when eddies and meanders disrupt a
simple linear geometry for the front, mixed signals result. Because of annual
variability in the strength of the California Current, in the abruptness of the
relaxation events during Summer, and in the prominence of eddies and
meanders in the California Current, the temporal patterns of larval recruitment
in consecutive years are not exact replicates. We term this overall hypothesis
the ‘tattered curtain’ hypothesis.”
In the decades since the tattered curtain hypothesis was first put forward scores of
studies have demonstrated a relationship between upwelling winds, larval distribution
and settlement events for a wide range of coastal species and in regions far beyond
the California coast (cf. Shanks and Sherman 2009). Subsequently, this hypothesis
has become generally accepted as the prevailing wisdom. However, recently the
upwelling relaxation implies settlement paradigm as well as “supply side” ecology in
general (the idea that ocean transport determines larval delivery) has come under
strong scrutiny. Limited dispersal is much more common than previously assumed
(Spongule et al. 2002, Swearer et al. 2002, Warner and Cowen. 2002), and
observations along the California coast in particular demonstrate that many species

88
recruit under strong upwelling conditions (Morgan et al. 2009, Shanks and Sherman
2009). A hypothesis often invoked to explain these observations, especially along the
California coast, is that larval behavior keeps spawn close to the coast, limiting
offshore transport (e.g., Batchelder et al. 2002). However, testing of this alternate
hypothesis requires confidence in the null hypothesis that without larval behavior
upwelling winds imply offshore transport and that upwelling relaxation, with a few
days lag, indeed should result in settlement pulses. Systematic testing of this
upwelling relaxation paradigm has yet to be done from an ocean modeling
perspective and is the goal of this study.
This paper is organized as follows. Section 4.2 outlines background theory on both
the Eulerian and Lagrangian dynamics of upwelling jets. Section 4.3 describes the
biophysical model used. Section 4.4 contains modeling results on patterns of coastal
retention and settlement as well as the statistical correlation between upwelling winds
and settlement patterns. Section 4.5 summarizes our conclusions. Finally, section 4.6
contains discussion of the results, including evidence from other studies that the
mechanisms seen here set settlement patterns in a number of regions.

89
4.2 Background theory
4.2.1 Eulerian dynamics: upwelling jets and offshore transport
In the coastal ocean, jets are a common occurrence, with the most dominant jets along
western boundary currents (e.g., Gulf Stream, Kuroshio). Eastern boundary currents
(EBCs), such as the California Current system (CCS), feature upwelling jets (Fig.
4.1), a geostrophic response to the upwelling of the pycnocline associated with the
“peeling back” of the surface ocean by offshore Ekman transport (Brink 1983, Hill et
al. 1998). Here there is a change in density moving offshore from colder, denser,
recently upwelled water near the coast to warmer, lighter water offshore, i.e. the
upwelling front. This density gradient results in a change in sea surface height and
thus a pressure force towards the coast. When the flow is in geostrophic balance the
Coriolis force balances this pressure gradient, and flow is along lines of constant sea
surface height moving towards the equator, resulting in an equatorward upwelling jet.
In theory the strength of this jet is directly proportional to the density gradient across
the upwelling front, and so in highly stratified regions the jet is fast and relatively
narrow.
In a tattered curtain, a breeze will preferentially flow through rips in the curtain,
unimpeded by the curtain’s weight. Similarly, swift offshore transport in coastal
upwelling systems will preferentially occur in regions where the upwelling jet is
already weakened by instabilities, the openings in the jet. In the case of the tattered

90
curtain, resistance is applied by the weight of the curtain: the heavier the curtain the
more resistance to the wind. In the case of coastal upwelling, resistance is applied by
coastal velocity shear, with more resistance for stronger shear, i.e. a faster alongshore
jet. To see this relationship, consider idealized Ekman mass transport in an upwelling
system. Ekman transport is calculated from the alongshore momentum balance by
assuming that horizontal velocity can be decomposed into a geostrophic and Ekman
component, i.e. u = uG + uE, and that all (non-linear) advection terms are negligible
(e.g., Gill 1982, Brink 1983). In upwelling systems this results in the alongshore
momentum balance
fuE = Τy/ρ,
where f is the Coriolis parameter, uE offshore (upwelling) velocity integrated over the
depth in which the winds act, Τy the wind stress at the ocean surface in the alongshore
direction, and ρ density. In the presence of a steady upwelling jet, offshore advection
of alongshore velocity becomes important and the momentum balance must be
modified to include nonlinear coupling between the Ekman and background
(geostrophic) modes, i.e.
uE δvG/δx + fuE = Τy/ρ,
where δvG/δx is the depth integrated cross-shelf shear of the alongshore velocity. This
results in a reduced upwelling velocity
uE = Τy/ρ(f + δvG/δx).

91
When δvG/δx is positive (an on the inshore flank of the upwelling jet), the strength of
the offshore transport is reduced by the magnitude of the velocity shear (Brink 1987,
Woodson in revision).
Weak points in the upwelling jet (where δvG/δx is small and thus uE stronger) can
occur through spontaneous instability, meanders instigated by local topography or
remotely forced by coastally trapped waves, or by interactions with the energetic
offshore eddy field generated by previous instabilities (Strub et al. 1991, Barth 1994,
Durski and Allen 2005). Most of these generation mechanisms are inherently
complex and involve nonlinear interactions (with the important exception of
interactions with coastal topography) and so exhibit a large degree of variability and
unpredictability (e.g., Durski and Allen 2005). Instabilities commonly result in eddies
and meanders in the coastal jet, as well as upwelling filaments, which often terminate
in eddies (Fig. 4.1), and squirts, which are wider offshore excursions of the upwelling
jet featuring fast cross-shelf velocities (Strub et al. 1991, Haynes et al. 1993,
Marchesiello et al. 2003, Durski and Allen 2005, Bécongée et al. 2009). All of these
features have strong effects on larval distribution patterns (Harrison et al. in review)
Despite the complexity of coastal circulation, simplified analytical results suggest the
distance of the upwelling front from shore is linearly proportional to the integrated
upwelling wind intensity, and this result has been somewhat borne out in observations
(Austin and Barth 2002), as well as in observed and modeled larval distributions and
settlement patterns (Roughgarden et al. 1988, 1991; Kim and Barth 2011). These

92
studies suggest a “settlement index” analogous to an upwelling index may be
tractable. Here we use a model of an idealized upwelling system to examine the
feasibility of finding such an index.
4.2.2 Lagrangian dynamics: a kinematic model of the upwelling jet
To understand how jets impact coastal Lagrangian transport, consider the idealized jet
(e.g., Kundu 1990):
where y is along the jet, x across the jet, A is the maximum jet velocity centered at
x=0 and the width of the jet is a function of the constant L (Fig. 4.2). This system
represents an idealized stationary Gaussian jet which is not accelerating in time in the
along jet-direction. Note the maximum across-jet velocity gradient (dv/dx)max occurs
at
€
x = ± 22 L (marked as vertical lines in Fig. 4.2), corresponding to the highest
velocity shear. This system can be integrated directly to provide the trajectories given
an initial position (x0,y0):
Here particles in the jet move linearly in time with a speed that is a function of their
initial position across the jet x0. It should be clear that particles within the center of
the jet move faster than those near the edge of the jet, in accordance with the shear of
the velocity field adjacent to the jet core. Effectively these lines of maximum shear
!
dx /dt = u = 0dy /dt = v = Aexp("x 2 /L2)
!
x(t) = x0y(t) = y0 + At exp("x0
2 /L2)

93
are a momentum or kinetic energy barrier. To move across the maximal shear zones
material must acquire or lose a large amount of kinetic energy, analogous to stepping
on the accelerator when changing from a slow lane to a fast lane on a busy freeway.
The amount of relative dispersion, i.e. separation between neighboring particle
trajectories, can be calculated as
,
which is maximized in the same location as the velocity shear, at
€
x0 = ± 22 L . Thus
blobs of material deposited at the shear zones will stretch out over time as material
closer to the jet center is transported faster than material towards the flanks of the jet.
The relative dispersion has a minimum at x0 = 0, and so material near the jet center
will remain coherent, staying together as it moves along within the jet core.
In studies motivated by dynamical systems results concerning the stability of
invariant tori, trajectories near jet cores have been shown to be remarkably stable to
perturbations in analytical (e.g., Samelson 1992, Rypina et al. 2007), experimental
(e.g., Beron-Vera et al. 2010), and observational studies (Rypina et al. 2011). These
jet cores, where the flow is shearless, remain a significant transport barrier when
regions on either side of the jet become more turbulent due to wave breaking or other
disturbances. Thus jets in general are expected to be very robust transport barriers
both because of the change in momentum needed to move in and out of them and also
their stability under perturbation.
!
dy /dx0 = "(2Ax0t L2)exp("x0
2 /L2)

94
4.3 Biophysical Model
The ocean model used in this study was formulated in the Regional Ocean Modeling
System (ROMS), a finite-difference, hydrostatic primitive equation community ocean
model that uses sigma coordinates in the vertical direction and a split-explicit time
stepping routine to represent the barotropic and baroclinic modes (Shchepetkin and
McWilliams 2005). The three-dimensional model setup was designed to simulate
mesoscale processes affecting Lagrangian transport along the central California Coast
(see Mitarai et al. 2008 and Siegel et al. 2008 for more details). Bathymetry and
forcing were adapted from observations at CalCOFI line 70 (California Oceanic
Cooperative Fisheries Investigations; e.g., Chelton 1984; Lynn and Simpson, 1987)
off the coast of Big Sur, CA during a typical July, the height of the upwelling season
for this region (Spring – Fall). The model features a straight eastern coast with a free-
slip boundary condition. Minimum depth is set to 10 m and maximum depth to 500
m, with a narrow shelf (~ 10 km) as typical for this region. There is no alongshore
variation in bathymetry. Horizontal resolution is 2 km and 20 sigma levels are used in
the vertical, with increased vertical resolution near the top and bottom of the domain
and near the coast. The north-south boundary is periodic necessitating use of an f-
plane for the Coriolis parameter.
The model is initialized using CalCOFI climatological data and nudged at the open
western boundary toward the climatology, which creates a realistic average vertical

95
temperature profile after the two year spin-up (Fig. 1 in Mitarai et al. 2008). A
climatological alongshore pressure gradient is estimated using mean dynamic height
differences between CalCOFI hydrographic sections off Point Arena and Point
Conception (lines 60 and 80; Lynn and Simpson 1987) and imposed within the model
as a body force that decreases with depth. Surface wind stress is modeled as a
statistically stationary Gaussian random process with magnitudes and decorrelation
timescales taken from buoy wind data (National Data Buoy Center stations 46028,
46012, and 46042) and spatial wind observations for the region. The wind field is
assumed to vary on spatial scales much larger than the model domain while
magnitude decreases toward the shore (i.e. positive wind stress curl), consistent with
observations along the California coast (Picket and Paduan 2003, Capet et al 2008).
This resulting wind product allows an ensemble of runs forced by time varying wind
fields that capture the statistics and temporal patterns driving upwelling dynamics
along the Central California coast (Mitarai et al. 2008).
Each of the 28 model ensemble runs lasts 180 days. Particles are released on a regular
2-km grid over the shelf each day for 170 days, constrained to the surface, and
tracked with a second order Runga-Kutta integration algorithm using ROMS model
surface velocities saved every 6 hrs (much shorter than the Eulerian decorrelation
timescale). For the analysis we focus on particles with 20-40 day residence time, or
pelagic larval duration (PLD), a common PLD for temperate fishes. When these 20-
40 PLD particles are found over the shelf, i.e. less than 10 km from shore, we

96
determine that they have “settled,” by which we mean potential settlement, as we are
not modeling spatial fecundity, life histories or mortality in the plankton. While the
total number of particles increases with time, the number of 20-40 PLD particles is
constant after day 40, as particles age in and out of this age class at constant rates.
4.4 Modeling Results
4.4.1 Modeled patterns of coastal retention and settlement
Inspection of model fields over the ensemble of runs shows that material released
over the shelf is often retained in the core of the upwelling jet. To illustrate this
retention pattern, we first present detailed results from a single model run (Run 111).
Fig. 4.3 shows surface fields of sea surface temperature (SST) and sea surface height
(SSH); surface velocity magnitude and direction, including a close up of the coastal
jet; density of particles of all ages (binned to a 1km2 grid and shown in log scale); and
density of 20-40 day pelagic larval duration (PLD) particles. The upwelling jet is
linear for a significant alongshore extent (over 100 km), but breaks up in a fast
offshore squirt and recirculation region towards the north of the domain (at ~225 km
along shore). Note this squirt occurs even under wind forcing that is coherent
alongshore and is not generated by coastal topography (Strub et al. 1991). Velocity
across the upwelling jet is roughly Gaussian as in the kinematic model (Fig. 4.3c).
Modeled density distributions are high within the core of the jet for larvae of all ages
(Fig. 4.3d). The upwelling jet retains 20-40 PLD larvae over the shelf, within the

97
settlement zone (dashed line in Fig. 4.3e), so that settlement is coherent for a wide
alongshore region.
To further illustrate the spatial relationship between modeled velocity, temperature
and particle density fields, Fig. 4.4 transects the model surface fields shown in Fig.
4.3, including the velocity field, velocity gradient, surface temperature gradient and
particle densities. The core of the jet is marked by the zero crossing of the velocity
gradient (Fig. 4.4b), and corresponds to the maximum SST gradient as predicted by
geostrophic balance (Fig. 4.4c). This jet core is also coincident with the region of
locally highest retention, and maximal velocity shear lines bound this high density
region as predicted by the kinematic model (Fig. 4.4d). This pattern of retention and
correlation in model variables is seen across the ensemble of 28 runs.
Visualization of settlement patterns through time for Run 111 is shown in a
Hovmöller plot (Fig. 4.5a). Here the number of 20-40 PLD particles is summed
between 0-10 km offshore for every 1 km along shore each day and visualized with
time increasing to the right. Settlement is strongly coherent along the coast, with
moderate levels tending to occur simultaneously (blue), with more sparse high
magnitude events interspersed (green to red). Settlement often ceases coherently all
across the domain (e.g., at day 140) often coincident with very strong upwelling
events (Fig. 4.5b). Coherent settlement patterns are broken up by regions of low or no
settlement propagating poleward through time (yellow arrows), while dense packets

98
move equatorward (black arrows). For example, the emerging jet instability at ~200
km alongshore on day 125 (Fig. 4.3d-e) appears in the settlement diagram as a region
of no settlement propagating poleward from day 110 to day 135. Here a meander
becomes unstable, eventually joining with the large squirt to the north. During this
same period a dense packet moves south within the jet during a time of overall high
settlement.
To visualize the flow features causing these patterns, Fig. 4.6 plots particle
distributions along with model SST and SSH surface fields for three days of the
model run shown in previous figures. Particles are colored blue for all particle ages,
black for 20-40 PLD and green for settling particles, i.e. for 20-40 PLD within 10 km
of the coast. On day 80 (Fig. 4.6a,d) there is retention in the coastal jet interrupted by
meanders around smaller eddies (~10 km diameter), and broken by a wide squirt
centered at 125 km alongshore. From day 80 to day 85 (Fig. 4.6b,e) eddies and their
associated breaks in retention have moved northward, while the squirt has narrowed.
The effects of these structures are visible in Fig. 4.5 as poleward propagating regions
of no settlement (yellow arrows). This poleward propagation of eddies is seen
throughout the ensemble of runs, and is consistent with coastally trapped wave theory
and observations (Brink 1991). A range of propagation speeds are seen over the
ensemble, likely due to the wide spectrum of wind forcing frequencies exciting these
features.

99
After extended strong upwelling, patterns of coastal retention move offshore (Fig.
4.6c), coincident with cooler temperatures along the coast (Fig. 4.6f). After the jet has
moved far offshore and broken up (supplemental movie 4.1), it takes a period of
weaker upwelling before the jet reforms and settlement resumes. By day 125 the same
pattern of a coastal jet broken up by a large squirt (Fig. 4.3) seen earlier in the run
(Fig. 4.6), emerges and continues for some time. These patterns of semi-coherence in
settlement are similar throughout all model runs (cf. Fig. 4.7), and are consistent with
the tattered curtain hypothesis, with the exception that retention is caused by stability
of the jet core and not by accumulation at the upwelling front due to downwelling.
Does upwelling relaxation imply delivery of material back to the coast? We have seen
that settlement often stops abruptly after strong upwelling events, but the relationship
of downwelling events and settlement is more complicated. The three largest
settlement events in model Run 111 occur during various wind conditions (Fig. 4.5b).
The first, centered around day 40, peaks after an upwelling relaxation event on day 42
but is significantly strong before this event and continues after relaxation ends. The
second and largest event, centered at day 87, occurs immediately after a large wind
relaxation, with a four-day lag between the peak relaxation and peak settlement times,
in the manner of upwelling-relaxation-implies-settlement paradigm. The next
settlement event (days 110-140) is broad in time and occurs during a time of
intermittent upwelling favorable winds. Notably there is only a small pulse of
settlement after the two wind relaxation events between day 100 and 110. Note that

100
settlement is lowest after prolonged strong upwelling winds, i.e. from days 86-103,
with a few days lag between the initiation of strong upwelling and decrease in
settlement. These model observations suggest an integrated effect of strong upwelling
in breaking patterns of coastal retention, i.e. complete tattering of the curtain.
Another example of settlement patterns is shown in Fig. 4.7 (Run 141). As in Fig. 4.6,
the spatial coherence of settlement is evident, again broken up by poleward
propagating features and dense packets traveling equatorward within the jet. There
are two large settlement events; the first occurs about 5 days after downwelling winds
relax (day 40), the second during a period of persistent moderate upwelling. Again
settlement ceases after strong upwelling events. Upwelling relaxation around day 130
consists of two nearby periods of downwelling favorable winds without any large
settlement events afterward. This relaxation event occurs after a long period of
upwelling favorable winds, when extended upwelling has transported most 20-40
PLD particles far offshore, so that when winds reverse they are not available to be
moved back to shore (supplemental movie 4.1).
As an endmember case, Run 113 exhibits the highest level of coastal retention of the
28 model runs (Fig. 4.8). Here settlement is high for a 40-day period peaking after a
period of moderate upwelling (Fig. 4.8b), and the settlement pattern is coherent along
a significant stretch of the coast (Fig. 4.8a). Inspection of the particle distribution
demonstrates that in this case the upwelling jet, though complex in shape, remains

101
close to the coast for an extended period of time. Again, upwelling relaxation events
have only a moderate correlation with settlement intensity. Wind relaxation on day
80, after a 20 day period of high intensity upwelling, results in only a small amount of
settlement. A subsequent relaxation event at day 100, after another 20-day period of
strong upwelling, has no associated settlement. A third, more pronounced relaxation
event occurs after day 160 and at this time settlement patterns follow the wind pattern
with a 2-5 day lag as seen in other runs, but this is much smaller settlement event than
when the jet is in place.
4.4.2 Statistical Correlation Between Wind and Settlement
The above results suggest that periods of moderate upwelling will be accompanied by
high settlement events, and that extended strong upwelling will completely tatter the
jet, moving particles of 20-40 PLD far offshore, decreasing settlement even if the
winds relax. But how much upwelling is needed to tatter the curtain, and can we
make any predictions about settlement of 20-40 PLD particles based on wind
patterns?
First, let us consider the statistical relationship between alongshore wind and
settlement for the test case Run 111 shown in Figs 3-6. The cross correlation peaks at
r = 0.44 for a 3-day lag (Fig. 4.9a), consistent with the large pulse of settlement after
the upwelling relaxation around day 84. However, there is also a larger broad window
of correlation for over 20 days lag time, consistent with the hypothesis that upwelling

102
intensity over some period of time is important in determining retention (or lack of
retention). To test this hypothesis we take the integrated alongshore wind, integrating
from the day in question backwards in time, and correlate this integrated wind
product with the settlement timeseries. In other words, we construct the wind product
, where v is the alongshore wind speed (far from the coastal
boundary layer), t the model day and T the integration window size. This results in a
peak correlation of r = 0.77 for T = 20 days (Fig. 4.9b), much higher than the lagged
wind result. As in the case of the lagged correlation there is a wide range of averaging
times with significant correlation.
To demonstrate the correlation dependence on the averaging window, Fig. 4.9 shows
the effect of different values of T on the integrated wind product. Here both the wind
product and the settlement timeseries have been normalized for the correlation
calculation. For the shorter averaging time (T = 5 days, Fig. 4.9c) the integrated wind
product predicts the short settlement pulse following the relaxation event and some of
the variability in the second settlement event, but generally does not do well in
predicting times of low or no settlement, for example from day 40 to 80 and from day
140 onward. However, the 20 day averaged wind product (W20), corresponding to the
maximum correlation (Fig. 4.9b), does very well in predicting these periods of low
settlement, as well as the broader, lower settlement window from days 116 -140 (Fig.
4.9d). The result that W20 correlates with settlement patterns is consistent with the
!
WT (t) = v(t)dtt"T
t#

103
observations in the previous section: that retention is low after long periods of strong
upwelling, and retention high during times of moderate sustained upwelling (as
opposed to just during relaxation events).
The combined statistical correlation of the lagged wind and retention is similar for the
ensemble of 28 runs (Fig. 4.10), again with a low correlation in the lagged result (r =
0.33 for a 2 day lag) and a peak correlation for a 20-day integration window. Here the
max correlation coefficient is r = 0.62, less than the single test run case, but with a
similar shape of correlation as a function of T. Note that the peak in the ensemble
case is sharper (compare Fig. 4.10b and 4.9b). If we examine a scatter plot of the
number of settlers and the integrated wind product W20 (Fig. 4.11), we see that for all
runs, very high levels of upwelling (W20 << 0) result in low settlement, and also that
there is always settlement under low integrated upwelling (W20 > -50).
Because of the significant correlation level for a wide range of integrated wind
conditions, as well as the high variability in the amount of settlement for the most
significantly correlated wind product W20 (Fig. 4.11), we determine that settlement is
only partially predictable by an upwelling wind product. However, we can predict the
endmember behavior for this simplified EBC: 1) during low integrated upwelling
winds some amount of potential settlement (with a high degree of variability) is
expected, 2) during extended heavy upwelling settlement will be low, 3) it is not
upwelling relaxation that results in settlement but moderate, sustained upwelling that

104
results in a highly retentive jet that entrains coastal material and acts as a significant
barrier to cross-shelf transport.
4.5 Conclusions
Larval settlement patterns in an ocean model of the central California coast are
largely controlled by retention in the upwelling jet. This jet remains close to the coast
over wide stretches, broken up by large squirts accommodating fast offshore
transport. Retention within the core of the jet is predicted by dynamical systems
theory, and regions of high retention are bounded by high velocity shear at the flanks
of the jet. Squirts, filaments and meanders caused by coastally trapped eddies
propagate poleward, modulating settlement patterns, while dense packets move
equatorward within the jet. High settlement events only weakly correlate with
upwelling wind relaxation (r = 0.33) and are better explained by a 20-day integrated
wind product (r = 0.62). Settlement is low after strong extended upwelling completely
tatters the jet, moving particles of settling age far offshore in complex patterns, and it
takes some time for the jet to reform and affect settlement patterns. For moderate
extended upwelling, coastal retention is high but highly variable in magnitude. This
follows from the inherent nonlinearity of upwelling jet dynamics.
4.6 Discussion
Here modeled retention indeed possesses an inherent stochasticistity due to nonlinear
dynamics of the upwelling jet and its interaction with offshore features, as well as the

105
prevalence of packets of dense material that appear to be folded into the jet
intermittently (Siegel et al. 2008, Harrison et al., in review). This indicates that
determining detailed settlement patterns in similarly turbulent systems (e.g., Mitarai
et al 2009) will require a “weather prediction” approach involving regional data
assimilative modeling (e.g., Roughan et al. 2011), or a significantly sized dataset of
coastal remote sensing observations that can be used to perform Lagrangian tracking
experiments and estimate local settlement events.
Some systems may be more predictable. The winds used to force our model feature
regular strong upwelling, much higher the other regions of the CCS (Picket and
Paduan 2003). Upwelling regions with consistent, weaker upwelling winds exhibit
more linear, predictable coastal jets (such as on well-studied Oregon shelf; Austin and
Barth 2002, Kim and Barth 2011). Our finding that moderate upwelling is associated
with high retention suggests settlement would be strongly modulated by the
upwelling jet in these regions, and may be more deterministic than stochastic for
these more regular flows (Kim and Barth 2011). A long-term study of barnacle and
mussel recruitment along the US West Coast (Broitman et al. 2008) is consistent with
this idea. In that study larval recruitment is largely synchronous along the central
Oregon coast, where the upwelling jet is a semi-permanent feature, and stochastic in
more energetic regions. Modeling results for the region (Kim and Barth, 2011) show
similar results to those here: that upwelling relaxation is poorly correlated with
settlement events and that an integrated wind product better explains settlement

106
events. Kim and Barth (2011) found a peak correlation for an 8-day integrated wind
product and settlement of 15-35 PLD particles (for a single model run), indicating
that optimally correlated wind products will need to be tuned to the region and
pelagic larval duration of interest.
Similar to Oregon, the Peru-Chile upwelling system features a straight N/S coastline
and a series of nearshore coastal jets, both equatorward at the surface and poleward
below. Here upwelling winds are persistently strong in some regions for most of the
year. A modeling study of this system (Aiken et al., 2011) showed that increased
upwelling winds associated with global warming decreased retention for neutrally
buoyant larvae, consistent with the results here. Aiken et al. (2011) also found an
increase in settlement for vertically migrating larvae, which were retained in the
global-warming-intensified poleward undercurrent until settling age. Thus even under
persistently strong upwelling conditions larval behavior coupled with retention in
coastal jets (in this case the undercurrent) may increase larval settlement (Morgan et
al. 2009, Shanks and Sherman 2009).
In upwelling systems with wider shelves, as in the Iberian, Canary and Beneguela
regions, upwelling can be centered far offshore at the shelf break (Estrade et al.
2008). This creates a series of geostropicly balanced alongshore jets, poleward
onshore of the upwelling center and equatorward offshore. Hence where larvae are
retained may depend on their release location: larvae released very close to shore

107
could get caught in the poleward jet (along with river output), while larvae spawned
on the outer shelf or slope could be retained in the more offshore equatorward jet.
This could lead to very strong retention for very nearshore spawning species in the
poleward coastal jet, as has been seen in the Iberian upwelling system in both
observational and modeling studies (Santos et al. 2004, Domingues et al. 2012).
These very nearshore jets are strongly coherent along the coast, reverse with
variability of the large-scale wind forcing, and again exhibit a high degree of
predictably (Domingues et al. 2012). Similarly but for different dynamical reasons,
the CCS also features coastal poleward flows during upwelling relaxation and low
upwelling times of year (Roughan et al. 2006, Drake and Edwards 2011), and flow
reversals of this very nearshore jet have been observed to affect intertidal settlement
(Dudas et al. 2009). These studies suggest highly retentive very nearshore jets will
exert an important hydrodynamic control on the settlement of very nearshore
spawning species.
High shear areas on the flanks of the upwelling jet were found to sometimes coincide
with collinear attracting and repelling Lagrangian coherent structures (LCS; Beron-
Vera et al. 2010). This follows from the high relative dispersion in both backward and
forward time for high velocity shear areas; such features are found farther offshore
along the main jet of the CCS (Harrison and Glatzmaier 2012) and so were expected
for this region. However, here collinear LCS were not found to be reliable markers of
the high density region boundaries, perhaps due to the short timescales that the jet

108
edges are stable. Instantaneous high velocity shear was a better marker for the jet
boundary, and much easier to calculate. Likewise, tori identified by low values of the
finite-time Lyapunov exponents, indicating minima of relative dispersion (Beron-
Vera et al. 2010, Rypina et al. 2011) were sometimes clear at the jet core and
sometimes not; this may be a function of the Lagrangian particle integration time
(Beron-Vera et al. 2010), or the sensitivity of this method (Harrison and Glatzmaier
2012). Another possible explanation is that because the jet is highly interactive with
coastal eddies (Fig. 4.6, Durski and Allen 2005), there is a significant amount of
exchange in and out of the jet (Samelson 1992). Lobe exchange here is highly
“messy,” most likely due to the broad spectrum of frequencies forcing this system.
Further investigation is needed to determine how useful these dynamical system
techniques will be given realistic flows.
4.7 Bibliography
C. M. Aiken, S. A. Navarrete, and J. L. Pelegrí. Potential changes in larval dispersal and alongshore connectivity on the central Chilean coast due to an altered wind climate. Journal of Geophysical Research, 116(G4):G04026, 2011.
J. A. Austin and J. A. Barth. Variation in the position of the upwelling front on the Oregon shelf. J. Geophys. Res, 107:3180, 2002.
J. A. Barth. Short-wave length instabilities on coastal jets and fronts. J. Geophys. Res, 99:16095–16115, 1994.
H. Batchelder, C. Edwards, and T. Powell. Individual-based models of copepod populations in coastal upwelling regions. Progress in Oceanography, 53(2-4):307–334, 2002.

109
P. Bécognée, M. Moyano, C. Almeida, J. M. Rodríguez, E. Fraile-Nuez, A. Hernández-Guerra, and S. Hernández-León. Mesoscale distribution of clupeoid larvae in an upwelling filament trapped by a quasi-permanent cyclonic eddy off Northwest Africa. Deep Sea Research Part I: Oceanographic Research Papers, 56(3):330–343, 2009.
F. J. Beron-Vera, M. J. Olascoaga, M. G. Brown, H. Koçak, and I. I. Rypina. Invariant-tori-like Lagrangian coherent structures in geophysical flows. Chaos, 20(1):017514–017514, 2010.
K. H. Brink. The near-surface dynamics of coastal upwelling. Progress In Oceanography, 12(3):223 – 257, 1983.
K. H. Brink. Upwelling fronts: implications and unknowns. South African Journal of Marine Science, 5(1):3–9, 1987.
K. H. Brink. Coastal-trapped waves and wind-driven currents over the continental shelf. Annual review of fluid mechanics, 23(1):389–412, 1991.
B. R. Broitman, C. A. Blanchette, B. A. Menge, J. Lubchenco, C. Krenz, M. Foley, P. T. Raimondi, D. Lohse, and S. D. Gaines. Spatial and temporal patterns of invertebrate recruitment along the west coast of the United States. Ecological Monographs, 78(3):403–421, 2008.
X. J. Capet, P. Marchesiello, and J. C. McWilliams. Upwelling response to coastal wind profiles. Geophysical research letters, 31(13):L13311, 2004.
D. B. Chelton. Seasonal variability of alongshore geostrophic velocity off central California. Journal of Geophysical Research, 1984.
C. P. Domingues, R. Nolasco, J. Dubert, and H. Queiroga. Model-derived dispersal pathways from multiple source populations explain variability of invertebrate larval supply. PLoS ONE, 7(4):e35794, 04 2012.
P. T. Drake, C. A. Edwards, and J. A. Barth. Dispersion and connectivity estimates along the U.S. West Coast from a realistic numerical model. Journal of Marine Research, 69(1):1–37, 2011.
S. E. Dudas, B. A. Grantham, A. R. Kirincich, B. A. Menge, J. Lubchenco, and J. A. Barth. Current reversals as determinants of intertidal recruitment on the central Oregon coast. ICES Journal of Marine Science, 66(2):396, 2009.
S. M. Durski and J. S. Allen. Finite-amplitude evolution of instabilities associated with the coastal upwelling front. Journal of physical oceanography, 35(9):1606–1628, 2005.

110
P. Estrade, P. Marchesiello, D. Verdiere, A. C. Colin, and C. Roy. Cross-shelf structure of coastal upwelling: A two dimensional extension of Ekman’s theory and a mechanism for inner shelf upwelling shut down. Journal of Marine Research, 66(5):589–616, 2008.
A. E. Gill. Atmosphere-Ocean dynamics. Academic Press, 1982.
C. S. Harrison, D. A. Siegel and S. Mitarai. The role of filamentation and eddy-eddy interactions in marine larval accumulation and transport. MEPS, in review.
R. Haynes, E. D. Barton, and I. Pilling. Development, persistence, and variability of upwelling filaments off the Atlantic coast of the Iberian Peninsula. J Geophys Res, 98(C12):22681–22692, 1993.
A. E. Hill, B. M. Hickey, F. A. Shillington, P. T. Strub, K. H. Brink, E. D. Barton, and A. C. Thomas. Eastern ocean boundaries coastal segment, Vol. 11 of The Sea, pages 29–67. John Wiley & Sons Inc, London, 1998.
S. Kim and J. A. Barth. Connectivity and larval dispersal along the Oregon coast estimated by numerical simulations. J. Geophys. Res., 116(C6), 06 2011.
P. K. Kundu. Fluid Mechanics. Academic Press, 1990.
R. J. Lynn and J. J. Simpson. The California Current system: The seasonal variability of its physical characteristics. J. Geophys. Res, 92(12):947–12, 1987.
P. Marchesiello, J. C. McWilliams, and A. Shchepetkin. Equilibrium structure and dynamics of the California Current system. Journal of Physical Oceanography, 33(4):753–783, 2003.
S. Mitarai, D. A. Siegel, and K. B. Winters. A numerical study of stochastic larval settlement in the California Current system. J. Marine Syst., 69(3-4):295–309, 2008.
S. Mitarai, D. A. Siegel, J. R. Watson, C. Dong, and J. C. McWilliams. Quantifying connectivity in the coastal ocean with application to the Southern California Bight. J. Geophys. Res, 114, 2009.
S. Morgan, J. Fisher, S. Miller, S. McAfee, and J. Largier. Nearshore larval retention in a region of strong upwelling and recruitment limitation. Ecology, 90(12):3489–3502, 2009.
M. Pickett and J. Paduan. Ekman transport and pumping in the California Current based on the U.S. Navy’s high-resolution atmospheric model (COAMPS). Journal of Geophysical Research (Oceans), 108:25–1, 2003.

111
M. Roughan, N. Garfield, J. Largier, E. Dever, C. Dorman, D. Peterson, and J. Dorman. Transport and retention in an upwelling region: The role of across-shelf structure. Deep Sea Research Part II: Topical Studies in Oceanography, 53(25-26):2931–2955, 2006.
M. Roughan, H. Macdonald, M. Baird, and T. Glasby. Modelling coastal connectivity in a western boundary current: Seasonal and inter-annual variability. Deep Sea Research Part II: Topical Studies in Oceanography, 58(5):628–644, 2011.
J. Roughgarden, S. Gaines, and H. Possingham. Recruitment dynamics in complex life cycles. Science, 241(4872):1460–1466, 1988.
J. Roughgarden, J. Pennington, D. Stoner, S. Alexander, and K. Miller. Collisions of upwelling fronts with the intertidal zone: the cause of recruitment pulses in barnacle populations of central California. Acta Oecologica, 12(1):35–51, 1991.
I. I. Rypina, M. G. Brown, F. J. Beron-Vera, H. Koçak, M. J. Olascoaga, and I. Udovydchenkov. On the Lagrangian dynamics of atmospheric zonal jets and the permeability of the stratospheric polar vortex. Journal of Atmospheric Sciences, 64:3595, 2007.
I. I. Rypina, L. J. Pratt, and M. S. Lozier. Near-surface transport pathways in the North Atlantic Ocean: Looking for throughput from the subtropical to the subpolar gyre. Journal of Physical Oceanography, 41(5):911–925, 2011.
J. Ryther. Photosynthesis and fish production in the sea. Science, 166(3901):72, 1969.
R. M. Samelson. Fluid exchange across a meandering jet. Journal of Physical Oceanography, 22(4):431– 444, 2012/05/03 1992.
A. Santos, A. Peliz, J. Dubert, P. Oliveira, M. Angélico, and P. Ré. Impact of a winter upwelling event on the distribution and transport of sardine (Sardina pilchardus) eggs and larvae off western Iberia: a retention mechanism. Continental Shelf Research, 24(2):149–165, 2004.
A. Shanks and R. Shearman. Paradigm lost? Cross-shelf distributions of intertidal invertebrate larvae are unaffected by upwelling or downwelling. Marine Ecology Progress Series, 385:189–204, 2009.
A. Shchepetkin and J. McWilliams. The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Modelling, 9(4):347–404, 2005.
D. A. Siegel, S. Mitarai, C. J. Costello, S. D. Gaines, B. E. Kendall, R. R. Warner, and K. B. Winters. The stochastic nature of larval connectivity among nearshore

112
marine populations. Proceedings of the National Academy of Sciences, 105(26):8974, 2008.
S. Sponaugle, R. Cowen, A. Shanks, S. Morgan, J. Leis, J. Pineda, G. Boehlert, M. Kingsford, K. Lindeman, C. Grimes, et al. Predicting self-recruitment in marine populations: biophysical correlates and mechanisms. Bulletin of Marine Science, 70(Supplement 1):341–375, 2002.
P. Strub, P. Kosro, A. Huyer, and CTZ Collaborators. The nature of the cold filaments in the California Current System. Journal of Geophysical Research, 96(C8):14743–14, 1991.
S. Swearer, J. Shima, M. Hellberg, S. Thorrold, G. Jones, D. Robertson, S. Morgan, K. Selkoe, G. Ruiz, and R. Warner. Evidence of self-recruitment in demersal marine populations. Bulletin of Marine Science, 70(Supplement 1):251–271, 2002.
R. Warner and R. Cowen. Local retention of production in marine populations: evidence, mechanisms, and consequences. Bulletin of Marine Science, 70(1):245–249, 2002.
Woodson, C.B. Spatio-temporal variability in cross-shelf exchange across Monterey Bay, CA. J Phys Oceanogr, in revision.

113
Figure 4.1 Features of upwelling dynamics in Eastern Boundary Currents (EBCs). Driven by persistent equatorward winds, EBCs often feature a surface equatorward upwelling jet modulated by instabilities such as filaments. A poleward undercurrent commonly lies beneath the surface jet, and this undercurrent can move from the shelf slope towards the surface during winter or upwelling relaxation events. (From Hill et al. 1998)
!"#$%&'#"()*+(,-.#%%&/0(1&21,%$34/((&/5(*2&/6(7889:(;<#(=#$(>?(77(
;<#(;$@#2#"(+,2A$&/(BC-4A<#D&D(
!/((A<#((789EFD(($((-$2$"&0G((#G#20#"((A4((#H-%$&/((A<#((4ID#2>#"((%$2>$%("&DA2&I,34/(($/"((D#@%#G#/A((-$@#2/D((4J((A<#((+$%&K42/&$((14$DA(LM4,0<0$2A#/(7899(=1&#/1#:(7887(NO)P?(!/(A<&D(D1#/$2&4(A<#(,-.#%%&/0(K24/A(%&G&AD(4JD<42#(A2$/D-42A(4K(/#2&31(LD<#%KP(D-#1&#D($/"(%$2>$#?(N%%()$DA#2/(*4,/"$2C(+,22#/AD(L)*+DP(D<$2#(D&G&%$2(1<$2$1A#2&D31D5(• (N(D,2K$1#(14$DA$%(#Q,$A42.$2"(,-.#%%&/0(R#A(• (S&A<($/($DD41&$A#"(,-.#%%&/0(K24/A(• (*246#/(,-(IC(DQ,&2AD($/"(T%$G#/AD(• (U2&>#/(IC(3G#V>$2C&/0(.&/"D(• (BC-4A<#D&D5(.&/"D(G4>#(K24/A(4/W4JD<42#:(14/A24%%&/0(D#@%#G#/A(
X#>#2(A#DA#"(&/($(41#$/(G4"#%(

114
Figure 4.2 Gaussian jet. Velocity profile for an idealized Gaussian jet. High velocity shear lines are marked in blue.
!"#$%&'()*+&#$,+-(.'&/")$
%0"$)1)#"2$
0')$#0"$)+3-4+($
'(.$#0"$2'5/2-2$6"3+7/#1$8&'./"(#$.69.5$+77-&)$'#$5$:$;<=>9>?$%0")"$'&"$"@"746"31$2+2"(#-2$A'&&/"&)B$C0"&"$#0"$70'(8"$/($2+2"(#-2$("".".$#+$2+6"$/($+&$+-#$+D$#0"$E"#$/)$8&"'#")#?$F+#"$#0")"$A'&&/"&)$'&"$+($"/#0"&$)/."$+D$#0"$-*C"33/(8$D&+(#$G5$:$HIB$(+#$'3+(8$/#?$$$
!
˙ x = u = 0˙ y = v = Aexp(" x 2 L2)
# $ %
!
x(t) = x0y(t) = y0 + Atexp(" x 2 L2)
# $ %
%0"$&"3'46"$./)*"&)/+(B$$$$$$$ $ $$$$$$$$$$$$$$$$$$$$$$$$$$$$$ $$$$$$$$$$$$$$$$B$/)$'3)+$2'5/2/J".$'#$5$:$;<=>9>B$'(.$."*"(.)$"5*+("(4'331$+($#0"$/(/4'3$7+(./4+($5H?$%0"$K%<L$2"#&/7$C/33$*/7M$-*$#0/)$)0"'&N#1*"$<OP$A+-(.'&1$')$'$2'5/2'3$7-&6"$/($A+#0$D+&C'&.$'(.$A'7MC'&.$42"?$
!
"y "x0 = (#2Ax0t L2)exp(#x0
2 /L2)
Q7"'(/7$E"#)$#"(.$#+$A"$R'-))/'($/($G"?8?$S-(.-I?$T($#0"$7')"$+D$'$7+')#'3$-*C"33/(8$E"#B$8"+)#&+*0/7$A'3'(7"$*&"./7#)$#0"$2'5/2-2$6"3+7/#1$C/33$7+/(7/."$C/#0$#0"$2'5/2-2$PP%$G'(.$#0-)$PPUI$8&'./"(#B$'3+(8$#0"$-*C"33/(8$D&+(#?$,-#$C0"&"$'&"$#0"$#&'()*+&#$A+-(.'&/")V$
R'-))/'($!"#$C/#0$W$:$<$:$X?$

115
Figure 4.3 Alongshore jet surface fields. Model surface fields for Run 111 day 125. Material released on the shelf is retained in the coastal upwelling jet. The model coast is on the right, poleward is up. (a) Sea surface temperature (SST) in color and sea surface height (SSH) contours, shown with 0.5 cm interval. (b) Surface velocity magnitude and direction. (c) Close-up of the upwelling jet velocity field. (d) Particle density for all residence time, binned to 1km2 and shown in log scale. (e) Particle density for 20-40 day residence time. Particles were released inshore of the vertical dashed line in d & e, <10 km from the model coast. A transect along the horizontal line at 50 km along shore is shown in Fig. 4.4.

116
Figure 4.4 Coastal jet transects. Transects across the upwelling jet for model day 125 of Run 111 (Fig. 4.3), where the jet is found running parallel to the coast. Panels are alongshore velocity v (a), cross-shore gradient of alongshore velocity dv/dx (b), across-shore sea surface temperature (SST) gradient (c) and particle density (d). Grey solid lines identify maximum cross-shore velocity shear. Maximum jet velocity (the jet core) is denoted by a dashed grey line (identified by zero crossing of dv/dx). Dense material is found between the high shear regions and is higher towards the center of the jet.
30 20 1010
5
0
5
x (km)
v (c
m/s
)
30 20 100
2
4
6
8
10
x (km)
dens
ity (N
/km
2 )
20 40 PDAll
30 20 101.5
1
0.5
0
0.5
x (km)
dv/d
x (c
m/s
/km
)
Day 125 y = 50 km
30 20 100.1
0.05
0
0.05
0.1
x (km)
dsst
/dx
(o C/k
m)
!" #"
$" %"

117
Figure 4.5 Settlement patterns and wind for Run 111. (a) Hovmöller plot of settlement density, defined as particles with 20-40 day residence time summed over the shelf (< 10 km), through the model run (horizontal axis). (b) Alongshore wind (dashed line) and number of particles retained (solid line) through the model run. Negative (equatorward) wind is upwelling favorable. Black arrows indicate packets of dense material moving within the jet, yellow arrows breaks in settlement patterns caused by poleward propagating coherent structures that break up and modulate the jet (Fig. 4.6).
a!
b!

118
Figure 4.6 Jet retention patterns. Particles and SST/SSH are shown for days 80 (a,d), 85 (b,e) and 100 (c,f) in model Run 111. Particles are colored by residence time and position: blue particles are all releases; black are 20-40 day olds; green are 20-40 day olds and within 10 km of the coast, i.e. settling particles. SSH contour interval is 0.5 cm (black lines in d-f). Material within the coastal jet moves equatorward (black arrow in panel a), while meanders in the coastal jet propagate poleward (yellow arrow in panel a). During a period of moderate upwelling (a,d), offshore transport occurs mainly in an offshore squirt while nearby the jet remains close to the coast. After a long period of heavy upwelling, retention patterns have moved offshore (c,f). See supplemental movie 4.1 for particle visualization through time.

119
Figure 4.7 Settlement patterns and wind for Run 141. Model fields shown as in Fig. 4.5. See text for interpretation.

120
Figure 4.8 Settlement patterns and wind for Run 113. Model fields shown as in Figs 4.5 & 4.7. Note change in scale for number of retained particles from previous figures (b; max is over 4000 on day 141). See text for interpretation

121
Figure 4.9 Statistics of wind-settlement correlation for Run 111. (a) Cross-correlation between number of settlers and alongshore wind peaks at r = 0.44 for a 3-day lag. (b) Correlation is higher for an integrated wind product, peaking at r = 0.77 for a 20 day integration window, with a wide range of similarly correlated window sizes. Some settlement events are better predicted by a short integration time (c), while long periods of low settlement are better predicted by longer wind integration (d).

122
Figure 4.10 Ensemble wind-settlement correlation statistics. (a) Cross correlation between wind and settlement peaks at r = 0.33 for a 3-day lag. (b) Correlation between settlement and integrated alongshore wind WT, where T is the integration window size, peaks at r = 0.62 for T = 20 days, within a wide range of similarly correlated T values.

123
Figure 4.11 Scatterplot of integrated wind product W20 vs. number of settling particles. Settlement is predictably low under extended strong upwelling conditions (W20 << 0) Under moderate levels of integrated upwelling (W20 > -50) settlement is always present but highly variable in magnitude. The outlier case Run 113 is plotted in red.

124
Chapter 5 Conclusions and directions for future research
5.1 Conclusions
It is now clear that coherent structures are ubiquitous in the California Current system
and exert strong controls on larval dispersal and settlement patterns. Methods and
theorems from dynamical systems can lead to important conceptual insights in how
these systems work and their stability under perturbation.
Chapter 2 shows that Lagrangian and Eulerian metrics agree for large, well developed
eddies in the CCS, with Lagrangian metrics less sensitive to model resolution and
noise. Coherent structure interactions (such as eddy-eddy merging) can be illuminated
by LCS maps. Small coherent features, such as lobes, are not well resolved by
altimetry due to spatial and temporal limitations. This, and the lack of adequate
coastal coverage, makes altimetric measurements somewhat limited for studying
ecosystem processes, except those associated with large coherent structures. (Sec.
5.2.2)
Chapter 3 shows filamentation, mapped by attracting LCS, is a major transport
pathway from the shelf offshore. Filaments also correspond to fronts and localized
upwelling and downwelling, indicating that details of frontal submesoscale processes
will be important in patterning ecosystem response in upwelling systems. Eddy-eddy

125
interaction and merging drives formation of dense packets that remain coherent for
long periods. These packets sweep together larvae of many source regions and release
times, and delivery back to the shore can generate quick pulses of high connectivity.
These findings are consistent with both larval surveys and other modeling studies.
Chapter 4 revisits Roughgarden’s tattered curtain hypothesis, which predicts that
during upwelling relaxation collisions of the upwelling front with the coast cause
pulses of larval settlement. Here we find that stability of the jet core causes retention,
and integrated upwelling favorable winds and not upwelling relaxation events are best
correlated with settlement patterns. Low integrated upwelling results in a persistent
upwelling jet, while high integrated upwelling tatters this curtain completely,
destroying retention and limiting settlement. The prevalence of coastal jets and
evidence of their effects on settlement patterns is widespread, indicating this is an
important process in many systems.
What are some of the questions remaining? First, it is important to quantify how
coherent structures impede mixing and pattern coastal ecosystem processes. While
this seems to be a very simple question, the answer is a bit illusive. What metrics do
we use? How do we define ecosystem patchiness and what are the important
processes generating this patchiness? What are the implications of patchiness for
patterning ecosystems? The next section outlines some preliminary studies related to
these problems.

126
5.2 Directions for future research
5.2.1 Quantifying biological patchiness.
We have seen in Chapters 3 and 4 that retention in the upwelling jet, eddy-eddy
interaction, and filamentation can lead to the formation of dense packets of material.
The questions we seek to answer here are: 1) How do we define how patchy the
resulting density distribution is? 2) How is this patchiness affected by particle/larval
behavior?
Mixing or patchiness metrics have a long tradition in fisheries ecology (cf. Bez 2000),
biological oceanography (Abraham 1998), engineering fluids (Dankiwerts 1952), and
dynamical systems studies (Grassberger and Procaccia 1983), with a concomitant
array of applications. Metrics range from simple concentration variance based metrics
(Bez 2000, Stremler 2008), to Fourier based (Abraham 1998, Mahadevan and
Campbell 2002, Mathew et al. 2005, Shaw et al. 2004) and fractal metrics
(Pierrehumbert 1991). The adage here is that a mixing measure “should be selected
according to the specific application and it is futile to devise a single measure to
cover all contingencies” (Ottino 1989). In some systems, the various metrics may be
related (Phelps and Tucker 2006), and at least in the engineering fluids studies some
effort has been made to define a standard of how homogeneity of a mixture should be
defined quantitatively (Boss 1986).

127
Figure 5.1 Various mixing measures for Run 111 (a) The concentration variance based mixing efficiency Φ is a single number measuring the variability of the larval density distribution from a well mixed (Φ = 1) to an unmixed (Φ = 0) state. Here Φ(t) is shown for all ages (black) and 20-40 day olds (gray). (b) The spatial density diagram (SDD) shows the percentage of area covered by the percentage of larvae for the least mixed (day 117) and best mixed (day 179) days as determined by Φ for 20-40 day olds. In all cases, larvae cover less than 30% of the ocean surface, and coverage is less for 20-40 day olds on the least mixed day (grey solid line) than on the best mixed day (grey dashed line). (c) The fractal dimension measures the dimensionality of the density distribution from linear (slope = 1) to 2D (slope = 2). At small scales, fractal dimension is low due to filamental (linear) features (Fig. 5.2d). (d) Fractal dimension D changes in time and appears similar to the variance based mixing metric Φ(t) for 20-40 day old larvae (a).

128
Figure 5.2 Larval density patterns on well mixed and poorly mixed days. Plots of concentration density (particles/km2) binned to one square kilometer for larvae of all age classes (a,c) and at settling age (b,d) assuming a 20-40 day pelagic larval duration. Density is shown for both the most well-mixed day (day 117: a,b) and least mixed day (day 179: c,d) determined by the mixing efficiency Φ for 20-40 day olds (Fig. 5.1a). The least mixed day (d) features a dense packet of larvae ~150km offshore along a long filament.
In larval ecology there is an old meme of the well-mixed-larval-pool, a scenario
where larvae released from the coast are transported offshore and homogenized until
being transported back to shore by downwelling or some other process (e.g., Pineda
2000). Given the insights on packet formation and retention gained in Chapters 3 and

129
4 it is natural to ask: how well mixed is the larval pool, and how do we quantify this?
In this way we can compare the mixing ability of systems across space, time and for
various larval behaviors.
Figure 5.1 shows the application of mixing measures to the same run shown in
Chapters 3 and 4 (Run 111). Here we assess the degree of mixing through time for all
releases and 20-40 day old larvae. Time series of variance based (Fig. 5.1a) and
fractal metrics (Fig. 5.1c-d) show surprising agreement. On poorly mixed days
variance is high, mixing low, and dimensionality more 1D than 2D, which follows
from the linearity of filaments caused by aggregation processes (Fig. 5.2). At no time
do larvae cover more than a small fraction of the total area (Fig. 5.1b), with 20-40 day
olds coving less than 5% of the coastal area on the most poorly mixed day in the
model run. These metrics can be applied to model experiments with larval swimming
(Fig. 3.11) to quantify the effect of swimming on patchiness (not shown).
5.2.2 LCS and predators in the CCS
Recent studies have highlighted the correlation of LCS and predator foraging tracks,
beginning with the work of Tew Kai et al. (2009), which showed that frigate birds
track both attracting and repelling LCS in the Mozambique Channel. Here I show
Figure 5.3 LCS and sea lion tracks Two foraging excursions (green squares) for one individual California sea lion (circles) are located at LCS (left) on the edge of an eddy and (right) on the edge of jet-like coherent structure. See supplemental movie 5.1 for animation.

130

131
some preliminary results on predator tracks in the CCS and discuss some possible
mechanisms of biological enhancement at LCS.
Maps of California sea lion tracks and LCS off the California coast are shown in Fig.
5.3. Here one individual makes two offshore trips over a period of a few weeks (green
squares; see supplemental movie 5.1 for animation). Each trip coincides with foraging
behavior near a coherent structure boundary: the first trip at the edge of an eddy and
coincident with a repelling LCS (red, Fig. 5.3 left panel), the second trip at a jet
boundary coincident with collinear repelling and attracting LCS (red and blue lines,
Fig. 5.3. right panel). Similar results are seen for other individual sea lions
(supplemental movie 5.1), as well as for albatross populations nesting in both in the
southern CCS and near Hawaii (not shown).
What is going on at LCS that makes them desirable foraging areas? The foraging
locations in Fig. 5.3 are far enough offshore (~200 km) that there is may be little
connection to enhancement and filamentation from upwelling sources. This suggests
enhancement of forage species either because of local primary production
enhancement (Capet et al. 2008, Calil and Richards 2010, Ch 3) and an energetic
cascade up to higher trophic levels, patchiness production from eddy-eddy interaction
(Ch 3), or some combination of both. Further study is needed to determine the details.

132
5.3 Bibliography
E. R. Abraham. The generation of plankton patchiness by turbulent stirring. Nature, 391(6667):577–580, 1998.
J. Boss. Evolution of the homogeneity degree of a mixture. Bulk Solids Handling, 6(6):1207–1215, 1986.
P. Calil and K. Richards. Transient upwelling hot spots in the oligotrophic North Pacific. Journal of Geophysical Research, 115(C2):C02003, 2010.
X. Capet, J. McWilliams, M. Molemaker, and A. Shchepetkin. Mesoscale to submesoscale transition in the California Current system. Part II: Frontal processes. Journal of Physical Oceanography, 38(1):44–64, 2008.
P. Danckwerts. The definition and measurement of some characteristics of mixtures. Applied Scientific Research, 3(4):279–296, 1952.
P. Grassberger and I. Procaccia. Measuring the strangeness of strange attractors. Physica D: Nonlinear Phenomena, 9(1-2):189 – 208, 1983.
A. Mahadevan and J. Campbell. Biogeochemical patchiness at the sea surface. Geophys. Res. Lett, 29(19):1926, 2002.
G. Mathew, I. Mezic, and L. Petzold. A multiscale measure for mixing. Physica D: Nonlinear Phenomena, 211(1-2):23–46, 2005.
J. M. Ottino. The kinematics of mixing: stretching, chaos, and transport. Cambridge University Press, Cambridge ; New York, NY, 1989.
J.H. Phelps and C.L. Tucker III. Lagrangian particle calculations of distributive mixing: limitations and applications. Chemical engineering science, 61(20):6826–6836, 2006.
R. Pierrehumbert. Large-scale horizontal mixing in planetary atmospheres. Physics of Fluids A: Fluid Dynamics, 3:1250, 1991.
J. Pineda. Linking larval settlement to larval transport: assumptions, potentials, and pitfalls. Oceanography of the Eastern Pacific, 1:84–105, 2000.

133
T. Shaw, J. Thiffeault, and C. Doering. Stirring up trouble: Multi-scale mixing measures for steady scalar sources. Physica D: Nonlinear Phenomena, 231(2):143–164, 2007.
M. Stremler. Mixing Measures. In: Encyclopedia of microfluidics and nanofluidics Springer Verlag, 2008.
E. Tew Kai, V. Rossi, J. Sudre, H. Weimerskirch, C. Lopez, E. Hernandez-Garcia, F. Marsac, and V. Garcon. Top marine predators track Lagrangian coherent structures. PNAS, 106(20):8245–8250, 2009.

134
Appendix: Glossary
FTLE = finite-time Lyapunov exponent
FFTLE = forward FTLE
BFTLE = backward FTLE
LCS = Lagrangian coherent structure
ALCS = attracting LCS
RLCS = repelling LCS
CCS = California Current system
EBC = eastern boundary current

135
List of Supplemental Movies
Movie 2.1 Daily LCS visualization for Fall 2005. Red curves are repelling (forward-time) LCS, and blue curves are attracting (backward-time) LCS. The 0.2 sigma contour of the Okubo-Weiss parameter is superimposed on the last frame (black). Rapid entrainment in the large eddy stops after it detaches from the main jet, here indicated by the disappearance of the spiraling attracting (blue) LCS in the eddy interior. For the mature eddy the LCS and the OW contours are consistent. See sections 2.4.1 & 2.4.2.
Movie 2.2 Comparison of Eulerian and Lagrangian metrics for 2002. The left frame contains contours of ADT (sea surface height from reference geoid) and the geostrophic velocity field. The right frame shows attracting (blue) and repelling (red) LCS and a contour of the Okubo-Weiss parameter (black). Here we use the 0.5 sigma Okubo-Weiss contour to aid in LCS visualization. See sections 2.4.1 & 2.4.2.
Movie 3.1 What happens to particles near LCS? Attracting and repelling Lagrangian coherent structures (LCS) are mapped by identifying regions in the fluid where particles initially close to one another are maximally stretched out in both backward and forward time. To motivate this conceptually, here we show the evolution of particle trajectories on and near attracting (blue) and repelling (red) LCS for the model run shown in other figures. On model day 33 blue particles are initialized in a dense 100 by 100 grid (this appears as a single dot) on four saddle points and integrated forward in time. The blue particles stretch out in forward time along the attracting LCS, also shown in blue in the background field. Similarly, red particles are initialized at the end of the movie (day 46) and integrated backward in time, where they stretch out along the repelling LCS (red curves); viewed in forward time these red particles converge onto the saddle points where they were initialized. Particles on both the forward and backward LCS stay on LCS as they move with the fluid. Particles in green are initialized nearby the LCS at the same initial densities as the groups of red and blue particles; these move along and towards the blue attracting LCS, but do not stretch out to the same degree as the red and blue particle sets.
Movie 3.2 Larval density field Time series of larval density (all age classes) for the model run shown in figures. Particle density is binned to 1 km2 and shown in a log scale to facilitate visualization of the wide range of densities.

136
Movie 4.1 Particles in the CCS-in-a-box model Particles are colored based on age: blue for all ages, black for 20-40 days old (i.e. potential settlers) and green for settlers (20-40 day olds that are within 10 km of the coast).
Movie 5.1 LCS and sea lion tracks Lagrangian coherent structures (LCS) calculated from altimetric (AVISO) data are plotted with California sea lion tracks for the beginning of 2006. LCS are shown daily with sea lion positions (black and green circles) for the entire day.




















