
Distinct involvement of Cdc42 and RhoA GTPases in actin organization and cell shape in untransformed...
9
Distinct involvement of Cdc42 and RhoA GTPases in actin organization and cell shape in untransformed and Dbl oncogene transformed NIH3T3 cells Cristina Olivo 1,4 , Cristina Vanni 2,4 , Patrizia Mancini 3 , Lorenzo Silengo 1 , Maria Rosaria Torrisi 3 , Guido Tarone 1 , Paola Defilippi* ,1 and Alessandra Eva 2 1 Dipartimento di Genetica, Biologia e Biochimica, Universita ` di Torino, Via Santena 5bis, 10126, Torino, Italy; 2 Laboratorio di Biologia Molecolare, Istituto G Gaslini, 16148 Genova Quarto, Italy; 3 Dipartimento di Medicina Sperimentale e Patologia, Universita ` di Roma ‘La Sapienza’, 00161 Roma, Italy The Dbl oncogene is a putative exchange factor for the small GTPases RhoA and Cdc42, which are involved in actin polymerization into stress fibers and filopodia, respectively. We report here that, upon adhesion to fibronectin, Dbl-transformed NIH3T3 cells display a contracted, polygonal shape with a high number of short stress fibers. In contrast, untransformed NIH3T3 cells acquire the characteristic fibroblast morphology and organize a regular mesh of long stress fibers. We show that in Dbl-transformed and in untransformed NIH3T3 cells the dierent shape and actin cytoskeleton organiza- tion observed in the early steps of adhesion involves activation of distinct GTPases. Upon adhesion to fibronectin, cell morphology of Dbl-transformed NIH3T3 cells depends on activation of RhoA and not of Cdc42. In contrast Cdc42 activation is necessary to untransfected NIH3T3 cells to acquire their fibroblast shape. In both Dbl-transformed and in untransformed NIH3T3 cells a basal Rac activation is necessary to support stress fiber organization, while constitutive Rac activation promotes rues and lamellipodia formation. As a consequence of RhoA activation, Dbl-transformed cells show high activity of ROCK-a and CRIK kinases, two known RhoA eectors. In addition Dbl-transformed and NIH3T3 cells expressing the constitutive active form of RhoA are less motile on fibronectin than cells expressing constitutive active Cdc42. We conclude that in NIH3T3 cells in response to fibronectin the expression of the Dbl oncogene leads to a predominant activation of RhoA which both supports the peculiar cell shape and actin cytoskeleton organization in stress fibers and regulates cell motility. Oncogene (2000) 19, 1428 – 1436. Keywords: Cdc42; RhoA; Dbl oncogene; actin cytos- keleton organization Introduction Guanine nucleotide exchange factor (GEF) for the Rho family of GTPases are comprised in a large family of proteins, all characterized by a region of sequence similarity consisting of the catalytic Dbl homology domain (DH) in tandem with a pleckstrin homology domain (PH) (Whitehead et al., 1997). The Dbl oncogene is the putative GEF for Cdc42 and RhoA (Hart et al., 1991, 1994). It has been shown that Dbl, as well as other Dbl family members, can activate JNK suggesting that they may regulate gene transcription (Coso et al., 1995; Minden et al., 1995). It has also been shown that Dbl, Vav and Ost oncogenes can activate NF-kB (Montaner et al., 1998) by the regulation of specific GTPases. Finally, it has been demonstrated that Dbl family proteins can potently activate transcription from the cyclin D1 promoter and that this activation correlates directly with the transforming activity of Dbl family oncogenic members (Westwick et al., 1997). All these data suggest that Rho GEFs regulate transcription through activation of multiple signaling pathways (Coso et al., 1995; Minden et al., 1995; Zhang et al., 1995; Hill et al., 1995). Cell adhesion to matrix proteins impose a cytoskeleton remodeling which define cell morphology on the adhesive substrate. The first step of this process consists in cell spreading on the matrix protein and organization of actin structures, which leads to formation of focal adhesions and actin stress fibers (Defilippi et al., 1999). The GTP-binding proteins Cdc42, Rac and RhoA are essential regulators of the dynamics of actin cytoskele- ton: Cdc42 regulates the formation of filopodia, Rac induces the formation of lamellipodia and membrane rues and RhoA participates in the formation of actin stress fibers and focal adhesions (Nobes and Hall, 1995; Hall, 1998). Moreover Cdc42 and Rac have been shown to be involved in the early phases of cell-matrix adhesion (Price et al., 1998; Clark et al., 1998). Several lines of evidence suggest that RhoA is also regulated by integrin- mediated cell adhesion to ECM, since integrin stimula- tion enhances stress fibers and focal adhesions (Barry et al., 1997; Ren et al., 1999). We have previously reported that Dbl oncogene controls actin cytoskeleton organization as a conse- quence of cell adhesion to specific extracellular sub- strates and that actin stress fibers are formed in Dbl- transformed cells upon adhesion on fibronectin (Defi- lippi et al., 1997). Other Dbl-like proteins have been shown to react to adhesion by inducing specific cellular responses. Tiam1 acts on Rac GTPase inducing a similar phenotype as constitutive activated Rac, including membrane ruing (Michiels et al., 1995). Moreover Tiam1 and Vav induce cellular responses that are dependent on adhesion to specific extracellular matrix proteins (Sander et al., 1998; Miranti et al., 1998; Yron et al., 1999). Regulation of actin cytoskeleton organization in response to cell adhesion to specific extracellular Oncogene (2000) 19, 1428 – 1436 ª 2000 Macmillan Publishers Ltd All rights reserved 0950 – 9232/00 $15.00 www.nature.com/onc *Correspondence: P Defilippi 4 The first two authors, contributed equally to this work Received 2 August 1999; revised 29 December 1999; accepted 13 January 2000
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Distinct involvement of Cdc42 and RhoA GTPases in actin organization and cell shape in untransformed...

Distinct involvement of Cdc42 and RhoA GTPases in actin organizationand cell shape in untransformed and Dbl oncogene transformed NIH3T3cells
Cristina Olivo1,4, Cristina Vanni2,4, Patrizia Mancini3, Lorenzo Silengo1, Maria Rosaria Torrisi3,Guido Tarone1, Paola De®lippi*,1 and Alessandra Eva2
1Dipartimento di Genetica, Biologia e Biochimica, UniversitaÁ di Torino, Via Santena 5bis, 10126, Torino, Italy; 2Laboratorio diBiologia Molecolare, Istituto G Gaslini, 16148 Genova Quarto, Italy; 3Dipartimento di Medicina Sperimentale e Patologia,UniversitaÁ di Roma `La Sapienza', 00161 Roma, Italy
The Dbl oncogene is a putative exchange factor for thesmall GTPases RhoA and Cdc42, which are involved inactin polymerization into stress ®bers and ®lopodia,respectively. We report here that, upon adhesion to®bronectin, Dbl-transformed NIH3T3 cells display acontracted, polygonal shape with a high number of shortstress ®bers. In contrast, untransformed NIH3T3 cellsacquire the characteristic ®broblast morphology andorganize a regular mesh of long stress ®bers. We showthat in Dbl-transformed and in untransformed NIH3T3cells the di�erent shape and actin cytoskeleton organiza-tion observed in the early steps of adhesion involvesactivation of distinct GTPases. Upon adhesion to®bronectin, cell morphology of Dbl-transformed NIH3T3cells depends on activation of RhoA and not of Cdc42. Incontrast Cdc42 activation is necessary to untransfectedNIH3T3 cells to acquire their ®broblast shape. In bothDbl-transformed and in untransformed NIH3T3 cells abasal Rac activation is necessary to support stress ®berorganization, while constitutive Rac activation promotesru�es and lamellipodia formation. As a consequence ofRhoA activation, Dbl-transformed cells show highactivity of ROCK-a and CRIK kinases, two knownRhoA e�ectors. In addition Dbl-transformed andNIH3T3 cells expressing the constitutive active form ofRhoA are less motile on ®bronectin than cells expressingconstitutive active Cdc42. We conclude that in NIH3T3cells in response to ®bronectin the expression of the Dbloncogene leads to a predominant activation of RhoAwhich both supports the peculiar cell shape and actincytoskeleton organization in stress ®bers and regulatescell motility. Oncogene (2000) 19, 1428 ± 1436.
Keywords: Cdc42; RhoA; Dbl oncogene; actin cytos-keleton organization
Introduction
Guanine nucleotide exchange factor (GEF) for the Rhofamily of GTPases are comprised in a large family ofproteins, all characterized by a region of sequencesimilarity consisting of the catalytic Dbl homologydomain (DH) in tandem with a pleckstrin homology
domain (PH) (Whitehead et al., 1997). The Dbl oncogeneis the putative GEF for Cdc42 and RhoA (Hart et al.,1991, 1994). It has been shown that Dbl, as well as otherDbl family members, can activate JNK suggesting thatthey may regulate gene transcription (Coso et al., 1995;Minden et al., 1995). It has also been shown that Dbl,Vav andOst oncogenes can activate NF-kB (Montaner etal., 1998) by the regulation of speci®c GTPases. Finally, ithas been demonstrated that Dbl family proteins canpotently activate transcription from the cyclin D1promoter and that this activation correlates directly withthe transforming activity of Dbl family oncogenicmembers (Westwick et al., 1997). All these data suggestthat Rho GEFs regulate transcription through activationofmultiple signaling pathways (Coso et al., 1995;Mindenet al., 1995; Zhang et al., 1995; Hill et al., 1995).
Cell adhesion to matrix proteins impose a cytoskeletonremodeling which de®ne cell morphology on theadhesive substrate. The ®rst step of this process consistsin cell spreading on the matrix protein and organizationof actin structures, which leads to formation of focaladhesions and actin stress ®bers (De®lippi et al., 1999).The GTP-binding proteins Cdc42, Rac and RhoA areessential regulators of the dynamics of actin cytoskele-ton: Cdc42 regulates the formation of ®lopodia, Racinduces the formation of lamellipodia and membraneru�es and RhoA participates in the formation of actinstress ®bers and focal adhesions (Nobes and Hall, 1995;Hall, 1998). Moreover Cdc42 and Rac have been shownto be involved in the early phases of cell-matrix adhesion(Price et al., 1998; Clark et al., 1998). Several lines ofevidence suggest that RhoA is also regulated by integrin-mediated cell adhesion to ECM, since integrin stimula-tion enhances stress ®bers and focal adhesions (Barry etal., 1997; Ren et al., 1999).
We have previously reported that Dbl oncogenecontrols actin cytoskeleton organization as a conse-quence of cell adhesion to speci®c extracellular sub-strates and that actin stress ®bers are formed in Dbl-transformed cells upon adhesion on ®bronectin (De®-lippi et al., 1997). Other Dbl-like proteins have beenshown to react to adhesion by inducing speci®c cellularresponses. Tiam1 acts on Rac GTPase inducing a similarphenotype as constitutive activated Rac, includingmembrane ru�ing (Michiels et al., 1995). MoreoverTiam1 and Vav induce cellular responses that aredependent on adhesion to speci®c extracellular matrixproteins (Sander et al., 1998; Miranti et al., 1998; Yron etal., 1999). Regulation of actin cytoskeleton organizationin response to cell adhesion to speci®c extracellular
Oncogene (2000) 19, 1428 ± 1436ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
*Correspondence: P De®lippi4The ®rst two authors, contributed equally to this workReceived 2 August 1999; revised 29 December 1999; accepted 13January 2000

proteins seems to be a characteristic distinctive of Dblfamily proteins as it has been shown that otheroncogenes, such as Ras and Src, induce a generaldisorganization of actin cytoskeleton that is not modi®edby adhesion on di�erent substrates. (Dartsch et al., 1994;Tarone et al., 1985; Felice et al, 1990).
We thus decided to investigate the pathways leadingto cytoskeleton organization in Dbl-transformed cellsas a consequence of cell adhesion. Here we analysedGTPase role in mediating actin stress ®ber organiza-tion and cell morphology in Dbl transformed cellsupon plating on ®bronectin. We demonstrate that themorphological di�erences observed between untrans-formed and Dbl-transformed cells in response to®bronectin are due to distinct GTPase activation.
Results
Dbl expression induces ROCK-a and CRIK kinaseactivity
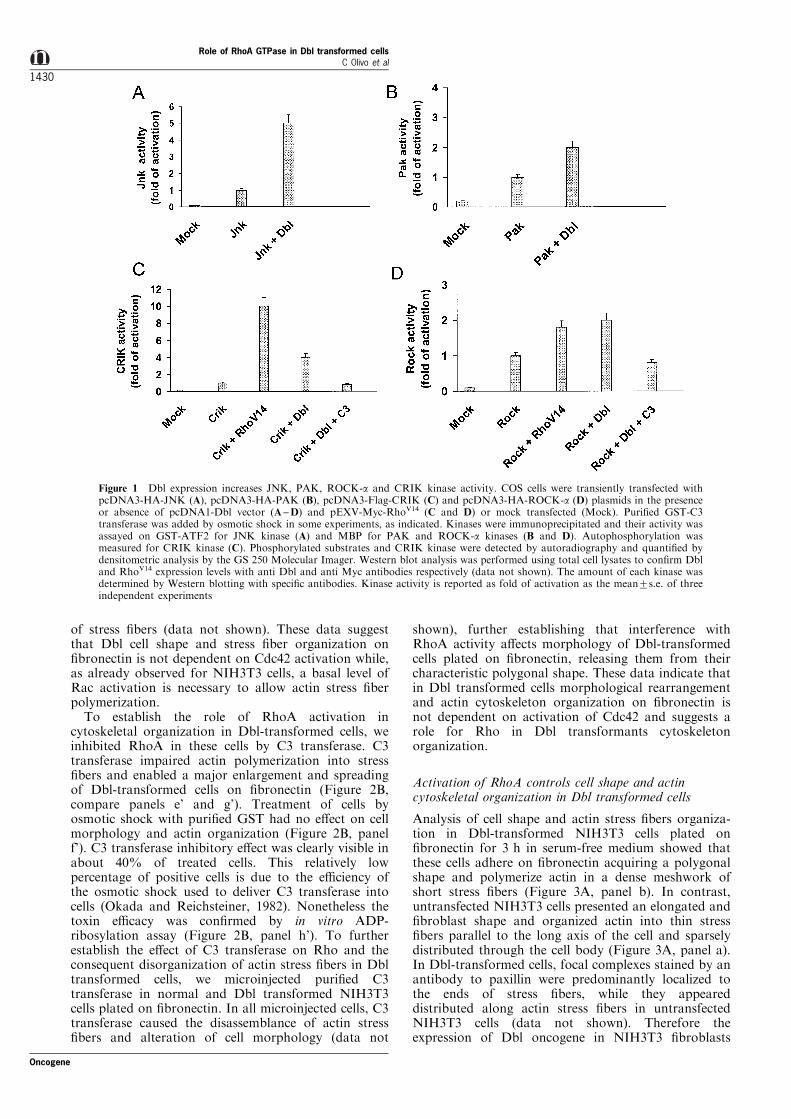
Rho family of GTPases exert their e�ect throughactivation of serine-threonine kinases, such as JNKand PAK-1 kinases, which are targets of active Cdc42and Rac, as well as the RhoA-dependent kinases ROCK-a and the recently identi®ed CRIK (Aspentrom, 1999).Therefore, in order to investigate which GTPases areactivated `in vivo' in cultured cells, COS cells weretransiently transfected with plasmids coding for each ofthese kinases in the absence or presence of Dbl oncogeneand harvested 24 h after incubation in serum-freemedium. As shown in Figure 1, we observed thatexpression of Dbl increased the activity of JNK andPAK (Figure 1A,B), as previously demonstrated (Cosoet al., 1995; Minden et al., 1995; Bagrodia et al., 1998), aswell as of ROCK-a and CRIK kinases (Figure 1C,D),suggesting that Dbl oncogene triggers both Cdc42 andRhoA pathways. Densitometric analysis shows that Dblexpression increases ROCK-a activity by twofold andCRIK activity by fourfold. Positive controls wererepresented by cells transfected with the constitutiveactive form of RhoA, RhoVal14, which increases bothCRIK and ROCK-a activity by tenfold and almosttwofold respectively (Figure 1C,D). To demonstrate thatthe increased ROCK-a and CRIK kinase activityobserved in Dbl-transformed cells was dependent onactivation of endogenous RhoA, kinase assays wereperformed in cells treated with C3 transferase, aClostridium Botulinum toxin which inactivates RhoAby ADP-ribosylation (Chardin et al., 1989). The reducedactivity of CRIK and ROCK-a kinases observed in Dblexpressing cells treated with C3 transferase (Figure 1C,D) indicates the speci®c requirements for RhoA in thisDbl-mediated activation. The same extent of kinaseactivation was observed in NIH3T3 cells (data notshown). Thus, Dbl oncogene `in vivo' activates Cdc42and RhoA as indicated by stimulation of catalyticactivity of the serine-threonine kinases which aresubstrates of Rho GTPases.
Cdc42 activation is essential for NIH3T3 fibroblasticshape on fibronectin
The results of the kinase assays indicated that inNIH3T3 and in COS cells Dbl activates both Cdc42-
and RhoA-associated signaling pathways. We havepreviously shown that Dbl a�ects actin cytoskeletonorganization in response to cell adhesion to speci®cextracellular proteins (De®lippi et al., 1997). We thusset up a series of experiments in order to establish theinvolvement of speci®c Rho GTPases in the acquisitionof the morphologic characteristics displayed by Dbl-transformed NIH3T3 cells in response to adhesion to®bronectin.
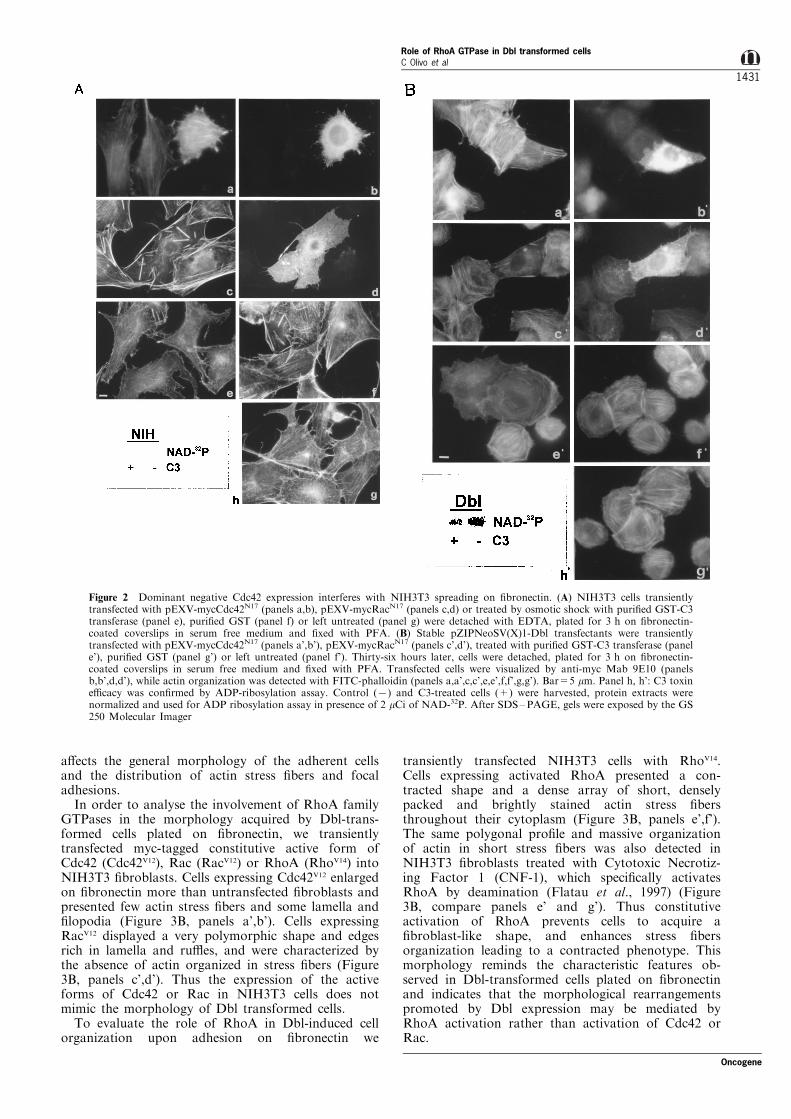
We ®rst analysed which GTPases are involved in theacquisition of the ®broblast shape in untransfectedNIH3T3 ®broblasts. Cells were transiently transfectedwith the dominant negative forms of Cdc42 (Cdc42N17)and Rac (RacN17) or treated with C3 transferase toinhibit RhoA, detached with EDTA, plated for 3 honto ®bronectin in serum free medium and co-stainedwith phalloidin for F-actin and anti-myc antibody9E10 to visualize positive transfected cells. NIH3T3cells expressing Cdc42N17 did not elongate such asnormal ®broblasts, remained roughly polygonal andproduced several tiny membrane protrusions (Figure2A, panels a,b). The dominant negative action ofCdc42 was evident within 1 ± 24 h of adhesion (datanot shown). This observation suggests that interferencewith activation of Cdc42 reduces the ability of NIH3T3cells to polarize and enlarge in response to ®bronectin.We also observe that at 3 h of adhesion, even in theabsence of Cdc42 activation, cells organized actin instress ®bers, suggesting that other GTPases maypromote this process (Figure 2A, panel a,b). Theseresults were further con®rmed by the analysis ofNIH3T3 cells stable expressing Cdc42N17, which donot acquire a ®broblast shape on ®bronectin and havea low degree of spreading similarly to what can beobserved in transient transfectants (data not shown).NIH3T3 cells transfected with RacN17 (Figure 2A,panels c,d) or treated with C3 transferase (Figure 2A,panel e) spread quite regularly on ®bronectin, suggest-ing that interfering with Rac or RhoA activity does notmodify cell shape. Indeed, expression of dominantnegative Rac a�ects stress ®bers organization, decreas-ing the number of actin ®bers and promotingformation of few short thick actin cables. These dataindicate that Cdc42 activation is required in the earlyphases of NIH3T3 cell spreading on matrix proteins inorder to acquire a ®broblast shape. In addition a basallevel of Rac activation is required to achieve normalorganization of actin stress ®bers.
Morphology and actin cytoskeleton organization ofDbl-transformed cells are not dependent on Cdc42activation
To evaluate involvement of Rho GTPases in cellorganization observed in Dbl-transformed cells, stableDbl transfectants were transiently transfected with thedominant negative form of Cdc42 (Cdc42N17). Expres-sion of Cdc42N17 did not modify cell morphology on®bronectin (Figure 2B, panels a',b') suggesting thatCdc42 activation is not involved in Dbl-transformedcell shape organization during adhesion to ®bronectin.Transient expression of RacN17 in Dbl-transformedcells, however, decreased the organization of actinstress ®bers (Figure 2B, panels c',d'). Moreoverexpression of constitutive active RacV12 modi®es Dblmorphology, leading to extended lamella and absence
Oncogene
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1429
of stress ®bers (data not shown). These data suggestthat Dbl cell shape and stress ®ber organization on®bronectin is not dependent on Cdc42 activation while,as already observed for NIH3T3 cells, a basal level ofRac activation is necessary to allow actin stress ®berpolymerization.
To establish the role of RhoA activation incytoskeletal organization in Dbl-transformed cells, weinhibited RhoA in these cells by C3 transferase. C3transferase impaired actin polymerization into stress®bers and enabled a major enlargement and spreadingof Dbl-transformed cells on ®bronectin (Figure 2B,compare panels e' and g'). Treatment of cells byosmotic shock with puri®ed GST had no e�ect on cellmorphology and actin organization (Figure 2B, panelf'). C3 transferase inhibitory e�ect was clearly visible inabout 40% of treated cells. This relatively lowpercentage of positive cells is due to the e�ciency ofthe osmotic shock used to deliver C3 transferase intocells (Okada and Reichsteiner, 1982). Nonetheless thetoxin e�cacy was con®rmed by in vitro ADP-ribosylation assay (Figure 2B, panel h'). To furtherestablish the e�ect of C3 transferase on Rho and theconsequent disorganization of actin stress ®bers in Dbltransformed cells, we microinjected puri®ed C3transferase in normal and Dbl transformed NIH3T3cells plated on ®bronectin. In all microinjected cells, C3transferase caused the disassemblance of actin stress®bers and alteration of cell morphology (data not
shown), further establishing that interference withRhoA activity a�ects morphology of Dbl-transformedcells plated on ®bronectin, releasing them from theircharacteristic polygonal shape. These data indicate thatin Dbl transformed cells morphological rearrangementand actin cytoskeleton organization on ®bronectin isnot dependent on activation of Cdc42 and suggests arole for Rho in Dbl transformants cytoskeletonorganization.
Activation of RhoA controls cell shape and actincytoskeletal organization in Dbl transformed cells
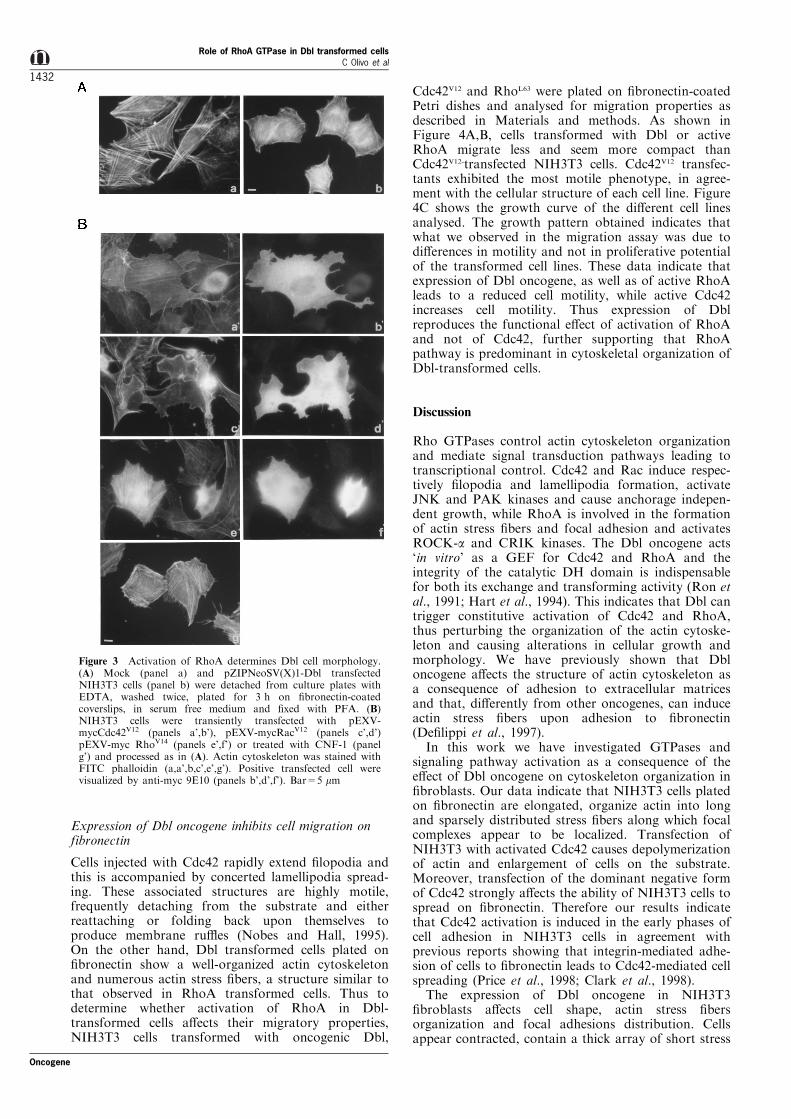
Analysis of cell shape and actin stress ®bers organiza-tion in Dbl-transformed NIH3T3 cells plated on®bronectin for 3 h in serum-free medium showed thatthese cells adhere on ®bronectin acquiring a polygonalshape and polymerize actin in a dense meshwork ofshort stress ®bers (Figure 3A, panel b). In contrast,untransfected NIH3T3 cells presented an elongated and®broblast shape and organized actin into thin stress®bers parallel to the long axis of the cell and sparselydistributed through the cell body (Figure 3A, panel a).In Dbl-transformed cells, focal complexes stained by anantibody to paxillin were predominantly localized tothe ends of stress ®bers, while they appeareddistributed along actin stress ®bers in untransfectedNIH3T3 cells (data not shown). Therefore theexpression of Dbl oncogene in NIH3T3 ®broblasts
Figure 1 Dbl expression increases JNK, PAK, ROCK-a and CRIK kinase activity. COS cells were transiently transfected withpcDNA3-HA-JNK (A), pcDNA3-HA-PAK (B), pcDNA3-Flag-CRIK (C) and pcDNA3-HA-ROCK-a (D) plasmids in the presenceor absence of pcDNA1-Dbl vector (A±D) and pEXV-Myc-RhoV14 (C and D) or mock transfected (Mock). Puri®ed GST-C3transferase was added by osmotic shock in some experiments, as indicated. Kinases were immunoprecipitated and their activity wasassayed on GST-ATF2 for JNK kinase (A) and MBP for PAK and ROCK-a kinases (B and D). Autophosphorylation wasmeasured for CRIK kinase (C). Phosphorylated substrates and CRIK kinase were detected by autoradiography and quanti®ed bydensitometric analysis by the GS 250 Molecular Imager. Western blot analysis was performed using total cell lysates to con®rm Dbland RhoV14 expression levels with anti Dbl and anti Myc antibodies respectively (data not shown). The amount of each kinase wasdetermined by Western blotting with speci®c antibodies. Kinase activity is reported as fold of activation as the mean+s.e. of threeindependent experiments
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1430
Oncogene
a�ects the general morphology of the adherent cellsand the distribution of actin stress ®bers and focaladhesions.
In order to analyse the involvement of RhoA familyGTPases in the morphology acquired by Dbl-trans-formed cells plated on ®bronectin, we transientlytransfected myc-tagged constitutive active form ofCdc42 (Cdc42V12), Rac (RacV12) or RhoA (RhoV14) intoNIH3T3 ®broblasts. Cells expressing Cdc42V12 enlargedon ®bronectin more than untransfected ®broblasts andpresented few actin stress ®bers and some lamella and®lopodia (Figure 3B, panels a',b'). Cells expressingRacV12 displayed a very polymorphic shape and edgesrich in lamella and ru�es, and were characterized bythe absence of actin organized in stress ®bers (Figure3B, panels c',d'). Thus the expression of the activeforms of Cdc42 or Rac in NIH3T3 cells does notmimic the morphology of Dbl transformed cells.
To evaluate the role of RhoA in Dbl-induced cellorganization upon adhesion on ®bronectin we
transiently transfected NIH3T3 cells with RhoV14.Cells expressing activated RhoA presented a con-tracted shape and a dense array of short, denselypacked and brightly stained actin stress ®bersthroughout their cytoplasm (Figure 3B, panels e',f').The same polygonal pro®le and massive organizationof actin in short stress ®bers was also detected inNIH3T3 ®broblasts treated with Cytotoxic Necrotiz-ing Factor 1 (CNF-1), which speci®cally activatesRhoA by deamination (Flatau et al., 1997) (Figure3B, compare panels e' and g'). Thus constitutiveactivation of RhoA prevents cells to acquire a®broblast-like shape, and enhances stress ®bersorganization leading to a contracted phenotype. Thismorphology reminds the characteristic features ob-served in Dbl-transformed cells plated on ®bronectinand indicates that the morphological rearrangementspromoted by Dbl expression may be mediated byRhoA activation rather than activation of Cdc42 orRac.
Figure 2 Dominant negative Cdc42 expression interferes with NIH3T3 spreading on ®bronectin. (A) NIH3T3 cells transientlytransfected with pEXV-mycCdc42N17 (panels a,b), pEXV-mycRacN17 (panels c,d) or treated by osmotic shock with puri®ed GST-C3transferase (panel e), puri®ed GST (panel f) or left untreated (panel g) were detached with EDTA, plated for 3 h on ®bronectin-coated coverslips in serum free medium and ®xed with PFA. (B) Stable pZIPNeoSV(X)1-Dbl transfectants were transientlytransfected with pEXV-mycCdc42N17 (panels a',b'), pEXV-mycRacN17 (panels c',d'), treated with puri®ed GST-C3 transferase (panele'), puri®ed GST (panel g') or left untreated (panel f'). Thirty-six hours later, cells were detached, plated for 3 h on ®bronectin-coated coverslips in serum free medium and ®xed with PFA. Transfected cells were visualized by anti-myc Mab 9E10 (panelsb,b',d,d'), while actin organization was detected with FITC-phalloidin (panels a,a',c,c',e,e',f,f',g,g'). Bar=5 mm. Panel h, h': C3 toxine�cacy was con®rmed by ADP-ribosylation assay. Control (7) and C3-treated cells (+) were harvested, protein extracts werenormalized and used for ADP ribosylation assay in presence of 2 mCi of NAD-32P. After SDS±PAGE, gels were exposed by the GS250 Molecular Imager
Oncogene
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1431
Expression of Dbl oncogene inhibits cell migration onfibronectin
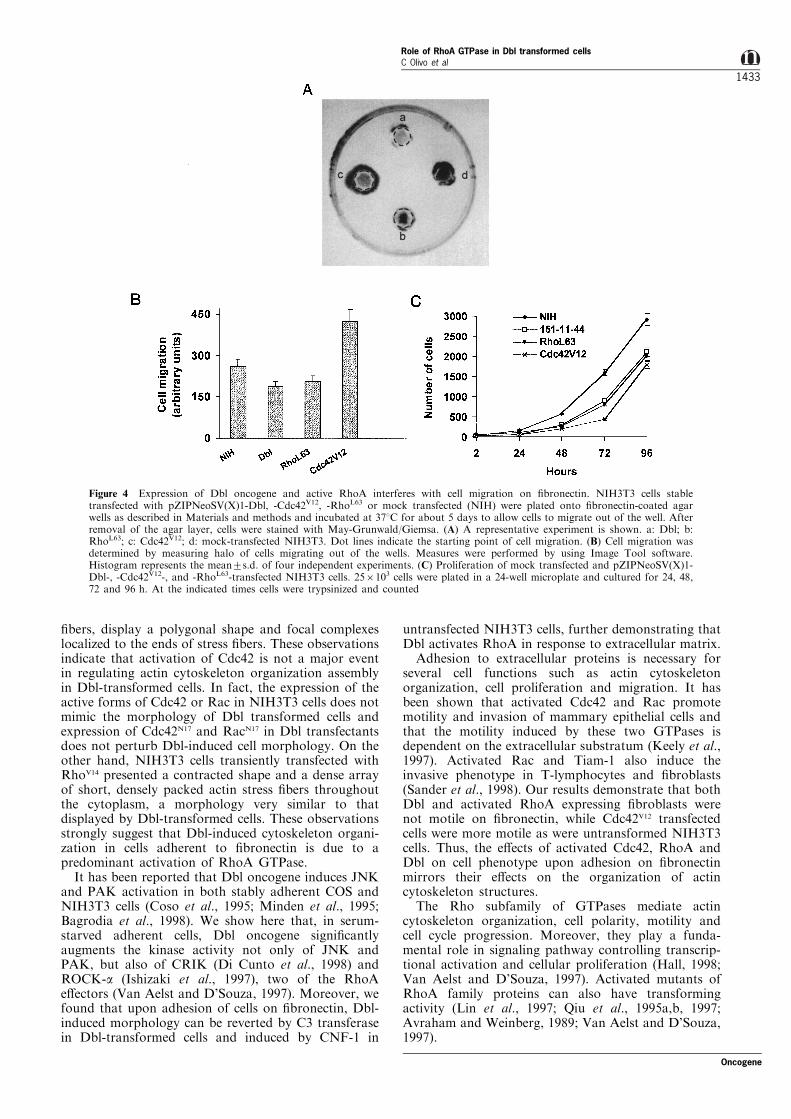
Cells injected with Cdc42 rapidly extend ®lopodia andthis is accompanied by concerted lamellipodia spread-ing. These associated structures are highly motile,frequently detaching from the substrate and eitherreattaching or folding back upon themselves toproduce membrane ru�es (Nobes and Hall, 1995).On the other hand, Dbl transformed cells plated on®bronectin show a well-organized actin cytoskeletonand numerous actin stress ®bers, a structure similar tothat observed in RhoA transformed cells. Thus todetermine whether activation of RhoA in Dbl-transformed cells a�ects their migratory properties,NIH3T3 cells transformed with oncogenic Dbl,
Cdc42V12 and RhoL63 were plated on ®bronectin-coatedPetri dishes and analysed for migration properties asdescribed in Materials and methods. As shown inFigure 4A,B, cells transformed with Dbl or activeRhoA migrate less and seem more compact thanCdc42V12-transfected NIH3T3 cells. Cdc42V12 transfec-tants exhibited the most motile phenotype, in agree-ment with the cellular structure of each cell line. Figure4C shows the growth curve of the di�erent cell linesanalysed. The growth pattern obtained indicates thatwhat we observed in the migration assay was due todi�erences in motility and not in proliferative potentialof the transformed cell lines. These data indicate thatexpression of Dbl oncogene, as well as of active RhoAleads to a reduced cell motility, while active Cdc42increases cell motility. Thus expression of Dblreproduces the functional e�ect of activation of RhoAand not of Cdc42, further supporting that RhoApathway is predominant in cytoskeletal organization ofDbl-transformed cells.
Discussion
Rho GTPases control actin cytoskeleton organizationand mediate signal transduction pathways leading totranscriptional control. Cdc42 and Rac induce respec-tively ®lopodia and lamellipodia formation, activateJNK and PAK kinases and cause anchorage indepen-dent growth, while RhoA is involved in the formationof actin stress ®bers and focal adhesion and activatesROCK-a and CRIK kinases. The Dbl oncogene acts`in vitro' as a GEF for Cdc42 and RhoA and theintegrity of the catalytic DH domain is indispensablefor both its exchange and transforming activity (Ron etal., 1991; Hart et al., 1994). This indicates that Dbl cantrigger constitutive activation of Cdc42 and RhoA,thus perturbing the organization of the actin cytoske-leton and causing alterations in cellular growth andmorphology. We have previously shown that Dbloncogene a�ects the structure of actin cytoskeleton asa consequence of adhesion to extracellular matricesand that, di�erently from other oncogenes, can induceactin stress ®bers upon adhesion to ®bronectin(De®lippi et al., 1997).
In this work we have investigated GTPases andsignaling pathway activation as a consequence of thee�ect of Dbl oncogene on cytoskeleton organization in®broblasts. Our data indicate that NIH3T3 cells platedon ®bronectin are elongated, organize actin into longand sparsely distributed stress ®bers along which focalcomplexes appear to be localized. Transfection ofNIH3T3 with activated Cdc42 causes depolymerizationof actin and enlargement of cells on the substrate.Moreover, transfection of the dominant negative formof Cdc42 strongly a�ects the ability of NIH3T3 cells tospread on ®bronectin. Therefore our results indicatethat Cdc42 activation is induced in the early phases ofcell adhesion in NIH3T3 cells in agreement withprevious reports showing that integrin-mediated adhe-sion of cells to ®bronectin leads to Cdc42-mediated cellspreading (Price et al., 1998; Clark et al., 1998).
The expression of Dbl oncogene in NIH3T3®broblasts a�ects cell shape, actin stress ®bersorganization and focal adhesions distribution. Cellsappear contracted, contain a thick array of short stress
Figure 3 Activation of RhoA determines Dbl cell morphology.(A) Mock (panel a) and pZIPNeoSV(X)1-Dbl transfectedNIH3T3 cells (panel b) were detached from culture plates withEDTA, washed twice, plated for 3 h on ®bronectin-coatedcoverslips, in serum free medium and ®xed with PFA. (B)NIH3T3 cells were transiently transfected with pEXV-mycCdc42V12 (panels a',b'), pEXV-mycRacV12 (panels c',d')pEXV-myc RhoV14 (panels e',f') or treated with CNF-1 (panelg') and processed as in (A). Actin cytoskeleton was stained withFITC phalloidin (a,a',b,c',e',g'). Positive transfected cell werevisualized by anti-myc 9E10 (panels b',d',f'). Bar=5 mm
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1432
Oncogene
®bers, display a polygonal shape and focal complexeslocalized to the ends of stress ®bers. These observationsindicate that activation of Cdc42 is not a major eventin regulating actin cytoskeleton organization assemblyin Dbl-transformed cells. In fact, the expression of theactive forms of Cdc42 or Rac in NIH3T3 cells does notmimic the morphology of Dbl transformed cells andexpression of Cdc42N17 and RacN17 in Dbl transfectantsdoes not perturb Dbl-induced cell morphology. On theother hand, NIH3T3 cells transiently transfected withRhoV14 presented a contracted shape and a dense arrayof short, densely packed actin stress ®bers throughoutthe cytoplasm, a morphology very similar to thatdisplayed by Dbl-transformed cells. These observationsstrongly suggest that Dbl-induced cytoskeleton organi-zation in cells adherent to ®bronectin is due to apredominant activation of RhoA GTPase.
It has been reported that Dbl oncogene induces JNKand PAK activation in both stably adherent COS andNIH3T3 cells (Coso et al., 1995; Minden et al., 1995;Bagrodia et al., 1998). We show here that, in serum-starved adherent cells, Dbl oncogene signi®cantlyaugments the kinase activity not only of JNK andPAK, but also of CRIK (Di Cunto et al., 1998) andROCK-a (Ishizaki et al., 1997), two of the RhoAe�ectors (Van Aelst and D'Souza, 1997). Moreover, wefound that upon adhesion of cells on ®bronectin, Dbl-induced morphology can be reverted by C3 transferasein Dbl-transformed cells and induced by CNF-1 in
untransfected NIH3T3 cells, further demonstrating thatDbl activates RhoA in response to extracellular matrix.
Adhesion to extracellular proteins is necessary forseveral cell functions such as actin cytoskeletonorganization, cell proliferation and migration. It hasbeen shown that activated Cdc42 and Rac promotemotility and invasion of mammary epithelial cells andthat the motility induced by these two GTPases isdependent on the extracellular substratum (Keely et al.,1997). Activated Rac and Tiam-1 also induce theinvasive phenotype in T-lymphocytes and ®broblasts(Sander et al., 1998). Our results demonstrate that bothDbl and activated RhoA expressing ®broblasts werenot motile on ®bronectin, while Cdc42V12 transfectedcells were more motile as were untransformed NIH3T3cells. Thus, the e�ects of activated Cdc42, RhoA andDbl on cell phenotype upon adhesion on ®bronectinmirrors their e�ects on the organization of actincytoskeleton structures.
The Rho subfamily of GTPases mediate actincytoskeleton organization, cell polarity, motility andcell cycle progression. Moreover, they play a funda-mental role in signaling pathway controlling transcrip-tional activation and cellular proliferation (Hall, 1998;Van Aelst and D'Souza, 1997). Activated mutants ofRhoA family proteins can also have transformingactivity (Lin et al., 1997; Qiu et al., 1995a,b, 1997;Avraham and Weinberg, 1989; Van Aelst and D'Souza,1997).
Figure 4 Expression of Dbl oncogene and active RhoA interferes with cell migration on ®bronectin. NIH3T3 cells stabletransfected with pZIPNeoSV(X)1-Dbl, -Cdc42V12, -RhoL63 or mock transfected (NIH) were plated onto ®bronectin-coated agarwells as described in Materials and methods and incubated at 378C for about 5 days to allow cells to migrate out of the well. Afterremoval of the agar layer, cells were stained with May-Grunwald/Giemsa. (A) A representative experiment is shown. a: Dbl; b:RhoL63; c: Cdc42V12; d: mock-transfected NIH3T3. Dot lines indicate the starting point of cell migration. (B) Cell migration wasdetermined by measuring halo of cells migrating out of the wells. Measures were performed by using Image Tool software.Histogram represents the mean+s.d. of four independent experiments. (C) Proliferation of mock transfected and pZIPNeoSV(X)1-Dbl-, -Cdc42V12-, and -RhoL63-transfected NIH3T3 cells. 256103 cells were plated in a 24-well microplate and cultured for 24, 48,72 and 96 h. At the indicated times cells were trypsinized and counted
Oncogene
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1433

In the past few years several investigators havesearched for e�ectors for Rho GTPases in order tounderstand the mechanisms lying behind the complexpattern of RhoA-regulated pathways. Of the manye�ector molecules so far identi®ed for the family ofGTPases, some are speci®c for a single member of thefamily, while others interact with more than oneGTPase. Moreover, many of these e�ector moleculesare ubiquitously expressed, and thus a cell typeselectivity does not necessarily explain speci®c re-sponses of GTPases to distinct upstream stimuli. Ithas been postulated that at least part of the speci®cityin RhoA GTPase signaling is the result of regulationthrough the action of RhoA GEFs which appear to becapable of distinct downstream signaling to speci®ce�ector pathways (Zhou et al., 1998). In our presentwork we have observed that Dbl oncogene, a GEF forboth Cdc42 and RhoA, does not induce activation ofCdc42 in response to adhesion to ®bronectin. We havealso found that Dbl expressing cells on ®bronectin arenot motile and present a very contracted morphologywith a thick array of ®bers. Lin et al. (1997) and Wu etal. (1998) have shown that a Cdc42-point mutant,which undergoes spontaneous exchange of GTP forGDP, while maintaining full GTPase activity, is able totransform NIH3T3 and to stimulate JNK activity. Theexpression of this CDC42 mutant in NIH3T3 cellscauses some of the e�ects induced by Dbl including thegeneration of giant multinucleated cells and growth insoft agar and in low serum medium suggesting thatCDC42 is a likely mediator of Dbl oncogene inducedtransformation. A possible explanation for theseapparent contradictory observation could be that thisoncogene activates distinct GTPases and thus varioussignaling pathways in response to di�erent upstreamstimuli. The relevance of RhoA activation in Dbl-induced transformation, possibly acting sinergisticallywith other activated GTPases, or the activation by Dblof other GTPases in response to adhesion to variousextracellular matrices, remains to be determined.
Materials and methods
Antibodies and reagents
Anti-Dbl polyclonal antibodies were previously generated asdescribed (Zangrilli and Eva, 1995). 9E10 anti-myc mono-clonal antibody (mAb) and ¯uorescein-(FITC)-conjugatedphalloidin were from Sigma; anti-Flag mAb was from Kodakand anti HA mAb (12CA5) was from Babco; rhodamine-labeled (RITC) antibody to mouse IgG was from KPL.Fibronectin was prepared as previously described (Tarone etal., 1982). Cytotoxic Necrotising Factor 1 (CNF-1), was akind gift of Dr P Boquet (U452, Inserm, Nice, France).
Plasmids
Construction of the pZIPNeo-DNDbl was previously de-scribed (Ron et al., 1989). RhoAL63 mutant was obtained withthe Transformer Site-Directed Mutagenesis Kit (Clontech)and cloned into the BamHI site of pZIPNeoSV(X)1 vector(Cepko et al., 1984). pZIPNeo-Cdc42V12 and pZIPNeo-Cdc42N17 constructs for stable expression in mammalian cellswere kindly provided by Dr R Cerione (Cornell University,Ithaca, NY, USA). Rho GTPases mutant vectors, pEXV-mycRhoV14, pEXV-mycCdc42V12, pEXV-mycCdc42N17,pEXV-mycRacV12, pEXV-mycRacN17 and the expression
plasmid pGEX-2T-C3, were kindly provided by Dr A Hall(MCR laboratory for Molecular Cell Biology, UK). pcDNA-Dbl, HA epitope-tagged JNK (HA-JNK) and the bacteriallyexpressed ATF fusion protein (GST-ATF2) were kindlysupplied by Dr S Gutkind (NIDCR, NIH, Bethesda, MA,USA). HA-tagged PAK-1 plasmid was a kind gift of Dr LLim (National University of Singapore, Singapore). Theexpression vector pcDNA3-HA-ROCK-a was a gift from DrNarumiya (University of Kyoto, Japan). pcDNA3-Flag-tagged CRIK was described in Di Cunto et al. (1998).
Cell cultures and transfections
COS cells were obtained from ATTC and were cultured inDMEM supplemented with 10% fetal calf serum. NIH3T3®broblasts were cultured in Dulbecco's modi®ed Eaglemedium (DMEM) supplemented with 10% calf serum. Masscultures of stable transfected cell lines were generated bytransfecting NIH3T3 cells by the calcium phosphatecoprecipitation method and cultured in DMEM supplemen-ted with 10% calf serum and 375 mg/ml of G418.For immuno¯uorescence experiments cells were plated at a
density of 126104 into a six-wells tissue culture plate andtransfected with 10 mg of plasmid DNA using the calciumphosphate procedure. Cells were used for experiments 36 hfrom DNA addition.For kinase assays cells grown to 80% con¯uence in
100 mm tissue culture dishes were transiently transfectedwith the indicated plasmids with the Lipofectamine Plusreagent as described by the manufacturer (Life Technologies).Twenty hours after transfection the medium was changed toDMEM containing 0.5% calf serum and cells were incubatedfor 24 h before harvest and lysis.
Toxins treatments and ADP-ribosylation assay
Treatment with CNF-1 was performed by pre-incubating cellswith 1076 M CNF-1 in culture medium overnight. C3transferase toxin was prepared as a GST-C3 fusion proteinaccording to Dillon and Feig (1995) and delivered into cellsby osmotic shock, according to Okada and Rechsteiner(1982). After osmotic shock, cells were rinsed in serum-freeDMEM, and incubated in serum-free DMEM containing20 mM HEPES for 4 h to allow cell recovery. Cells treatedwith GST-C3 transferase were washed with PBS, harvestedwith ice cold TBS (150 mM NaCl, 50 mM Tris-HCl pH 7.4),containing 250 mM sucrose, 3 mM imidazole, 1 mM PMSFand protease inhibitors and incubated for 30 min at 48C. Cellextracts were subjected to a three freezing-unfreezing cyclesand cleared by centrifugation at 14 000 r.p.m. for 10 min at48C. Samples were adjusted to the same protein concentra-tion and 20 mg per sample were incubated with 2 mCi[32P]NAD and C3 transferase in the ADP-ribosylation bu�er(50 mM Tris pH 7.5, 100 mM KCl, 1 mM DTT). ADP-ribosylation reaction was stopped by adding Laemmli Bu�erand boiling. Samples were separated onto a 12% SDS±PAGE gel and exposed using the GS 250 Molecular Imager(Bio-Rad).
Immunofluorescence analysis
Glass coverslips were coated with 10 mg/ml ®bronectin byover night incubation at 48C and washed twice with PBS.Cells were gently detached with 5 mM EDTA in PBS, washedtwice with 1 mM CaCl2, 1 mM MgCl2 and plated in serum-free DMEM supplemented with 20 mM HEPES for 3 h ontocoverslips. Cells were ®xed in 3% paraformaldehyde (PFA)for 10 min and permeabilized with 0.5% Triton-TBS(150 mM NaCl, 20 mM Tris-Cl pH 7.4) for 1.5 min. F-actinwas revealed by staining with FITC-phalloidin. Whentransient transfected cells were used, positive cells wererevealed with anti-myc mAb 9E10. Cells were observed by
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1434
Oncogene

a Zeiss 6361.4 oil immersion objective. Photographs weretaken on 400 Asa Kodak ®lm.
Kinase assays
JNK kinase activity in cells transfected with an epitope-tagged JNK (HA-JNK) was determined as described (Coso etal., 1995) using puri®ed bacterially expressed glutathione S-transferase ATF fusion protein as substrate. PAK andROCK-a kinase activity in cells transfected with epitopetagged plasmids (HA-PAK and HA-ROCK) was determinedin the same way using myelin basic protein (MBP) (Sigma) asa substrate. CRIK autokinase activity in cells transfectedwith an epitope tagged CRICK (Flag- CRIK) was performedas described (Di Cunto et al., 1998). Proteins were separatedby SDS±PAGE and transferred to Immobilon-P membranes(Millipore) which were used for both autoradiography andWestern blot analysis to evaluate protein expression level.Immunocomplexes were visualized by enhanced chemiolumi-nescence detection (Pierce) using Protein A coupled tohorseradish peroxidase.
Migration assay and cell growth curve
Sixty mm Petri dishes precoated with 20 mg/ml of ®bronectinwere covered with 10 ml of 0.4% noble agar (DIFCO) inDMEM supplemented with 10% calf serum and cloningcylinders were used to create wells in the agar. Cells were
plated at 56104 cells/well and incubated at 378C for about 5days to allow cells to migrate out of the well on the matrixunderneath the agar layer. After removal of the agar layercells were stained with May-Grunwald/Giemsa. Cell migra-tion was determined by measuring the halo of cells migratingout of the wells. Measures were performed by using ImageTool software (University of Texas, Health Science Center).Con¯uent cells were detached from culture dishes, counted
and plated in a 24 well microplate at 256103 cells/well. Cellswere grown for 24, 48, 72 and 96 h, detached and counted.The growth curve was repeated twice and each point wasperformed in triplicate. The values represent average of thetriplicates of one representative experiment.
AcknowledgmentsWe are indebted to Drs P Boquet, R Cerione, S Gutkind,A Hall, L Lim, and S Narumiya for providing reagents. Wealso thank Dr F Di Cunto for helpful discussion. C Olivowas supported by a short term fellowship by NationalResearch Council to visit Dr P Boquet laboratory. Thiswork was supported by grants of the Italian Associationfor Cancer Research (AIRC), MURST `Co®nanziamentoProgrammi di Ricerca', Istituto Superiore di Sanita',Progetto `Sostituzioni funzionali, organi arti®ciali e tra-pianti di organi' and from Ministero della SanitaÁ , Progetto®nalizzato N. ICSO70.2/RF95.221.
References
Aspenstrom P. (1999). Curr. Opin. Cell Biol., 11, 95 ± 102.Avraham H and Weinberg RA. (1989). Mol. Cell Biol., 9,
2058 ± 2066.Bagrodia S, Taylor SJ, Jordon KA, Van Aelst L and Cerione
RA. (1998). J. Biol. Chem., 273, 23633 ± 23636.Barry ST, Flinn HM, Humphries MJ, Critchley DR and
Ridley AJ. (1997). Cell Adhes. Commun., 4, 387 ± 398.Cepko CL, Roberts BE and Mulligan RC. (1984). Cell, 37,
1053 ± 1062.Chardin P, Boquet P, Madaule P, Popo� MR, Rubin EJ and
Gill DM. (1989). EMBO J., 8, 1087 ± 1092.Clark EA, King WG, Brugge JS, Symons M and Hynes RO.
(1998). J. Cell Biol., 142, 573 ± 586.Coso OA, Chiariello M, Yu JC, Teramoto H, Crespo P, Xu
N, Miki T and Gutkind JS. (1995). Cell, 81, 1137 ± 1146.Dartsch PC, Ritter M, Haussinger D and Lang F. (1994).
Eur. J. Cell Biol., 63, 316 ± 325.De®lippi P, Olivo C, Tarone G, Mancini P, Torrisi MR and
Eva A. (1997). Oncogene, 14, 1933 ± 1943.De®lippi P, Olivo C, Venturino M, Dolce L, Silengo L and
Tarone G. (1999). Micr. Res. Tech., 47, 67 ± 78.Di Cunto F, Calautti E, Hsiao J, Ong L, Topley G, Turco E
and Dotto GP. (1998). J. Biol. Chem., 273, 29706 ± 29711.Dillon ST and Feig LA. (1995). Methods Enzymol., 256,
174 ± 184.Felice GR, Eason P, Nermut MV and Kellie S. (1990). Eur. J.
Cell Biol., 52, 47 ± 59.Flatau G, Lemichez E, Gauthier M, Chardin P, Paris S,
Fiorentini C and Boquet P. (1997). Nature, 387, 729 ± 733.Hall A. (1998). Science, 279, 509 ± 514.Hart MJ, Eva A, Evans T, Aaronson SA and Cerione RA.
(1991). Nature, 354, 311 ± 314.Hart MJ, Eva A, Zangrilli D, Aaronson SA, Evans T,
Cerione RA and Zheng Y. (1994). J. Biol. Chem., 269, 62 ±65.
Hill CS, Winne J and Treisman R. (1995). Cell, 81, 1159 ±1170.
Ishizaki T, Naito M, Fujisawa K, MaekawaM, Watanabe N,Saito Y and Narumiya S. (1997). FEBS Lett, 404, 118 ±124.
Keely PJ, Westwick JK, Whitehead IP, Der CJ and PariseLV. (1997). Nature, 390, 632 ± 636.
Lin R, Bagrodia S, Cerione R and Manor D. (1997). Curr.Biol., 7, 794 ± 797.
Michiels F, Habets GG, Stam JC, van der Kammen RA andCollard JG. (1995). Nature, 375, 338 ± 340.
Minden A, Lin A, Claret FX, Abo A and Karin M. (1995).Cell, 81, 1147 ± 1157.
Miranti CK, Leng L, Maschberger P, Brugge JS and ShattilSJ. (1998). Curr. Biol., 8, 1289 ± 1299.
Montaner S, Perona R, Saniger L and Lacal JC. (1998). J.Biol. Chem., 273, 12779 ± 12785.
Nobes CD and Hall A. (1995). Cell, 81, 53 ± 62.Okada CY and Reichsteiner M. (1982). Cell, 29, 33 ± 41.Price LS, Leng J, Schwartz MA and Bokoch GM. (1998).
Mol. Biol. Cell, 9, 1863 ± 1871.Qiu RG, Abo A, McCormick F and Symons M. (1997). Mol.
Cell Biol., 17, 3449 ± 3458.Qiu RG, Chen J, Kirn D, McCormick F and Symons M.
(1995a). Nature, 374, 457 ± 459.Qiu RG, Chen J, McCormick F and Symons M. (1995b).
Proc. Natl. Acad. Sci., 92, 11781 ± 11785.Ren XD, Kiosses WB and Schwartz MA. (1999). EMBO J.,
18, 578 ± 585.Ron D, Graziani G, Aaronson SA and Eva A. (1989).
Oncogene, 4, 1067 ± 1072.Ron D, Zannini M, Lewis M, Wickner RB, Hunt LT,
Graziani G, Tronick SR, Aaronson SA and Eva A. (1991).New Biol., 3, 372 ± 379.
Sander EE, van Delft S, ten Klooster JP, Reid T, van derKammen RA, Michiels F and Collard JG. (1998). J. CellBiol., 143, 1385 ± 1398.
Tarone G, Cirillo D, Giancotti FG, Comoglio PM andMarchisio PC. (1985). Exp. Cell Res., 159, 141 ± 157.
Tarone G, Galetto G, Prat M and Comoglio PM. (1982). J.Cell Biol., 94, 179 ± 186.
Van Aelst L and D'Souza-Schorey C. (1997). Genes Dev., 11,2295 ± 2322.
Oncogene
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1435

Westwick JK, Lambert QT, Clark GJ, Symons M, Van AelstL, Pestell RG and Der CJ. (1997). Mol. Cell Biol., 17,1324 ± 1335.
Whitehead IP, Campbell S, Rossman KL and Der CJ. (1997).Biochim. Biophys. Acta, 1332, F1 ± F23.
Wu WJ, Lin R, Cerione RA and Manor D. (1998). J. Biol.Chem., 273, 16655 ± 16658.
Yron I, Deckert M, Re� ME, Munshi A, Schwartz MA andAltmen A. (1999). Cell Adhes. Commun., 7, 1 ± 11.
Zangrilli D and Eva A. (1995).Methods Enzymol., 256, 347 ±358.
Zhang S, Han J, Sells MA, Cherno� J, Knaus UG, UlevitchRJ and Bokoch GM. (1995). J. Biol. Chem., 270, 23934 ±23936.
Zhou K, Wang Y, Gorski JL, Nomura N, Collard J andBokoch GM. (1998). J. Biol. Chem., 273, 16782 ± 16786.
Role of RhoA GTPase in Dbl transformed cellsC Olivo et al
1436
Oncogene




![Arf nucleotide binding site opener [ARNO] promotes sequential activation of Arf6, Cdc42 and Rac1 and insulin secretion in INS 832/13 β-cells and rat islets](https://static.fdokumen.com/doc/165x107/6316194efc260b7102104d00/arf-nucleotide-binding-site-opener-arno-promotes-sequential-activation-of-arf6.jpg)















