
Diseases of Quinoa ( Chenopodium quinoa )
18
This article was downloaded by:[Danmarks Veternaer Og] On: 25 March 2008 Access Details: [subscription number 731846679] Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Food Reviews International Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713597252 Diseases of Quinoa (Chenopodium quinoa) Solveig Danielsen a ; Alejandro Bonifacio b ; Teresa Ames c a The Royal Veterinary and Agricultural University, Frederiksberg C, Denmark b PROINPA Foundation, Choquenaira, La Paz, Bolivia c International Potato Center (CIP), Lima, Peru Online Publication Date: 05 January 2003 To cite this Article: Danielsen, Solveig, Bonifacio, Alejandro and Ames, Teresa (2003) 'Diseases of Quinoa (Chenopodium quinoa)', Food Reviews International, 19:1, 43 - 59 To link to this article: DOI: 10.1081/FRI-120018867 URL: http://dx.doi.org/10.1081/FRI-120018867 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diseases of Quinoa ( Chenopodium quinoa )

This article was downloaded by:[Danmarks Veternaer Og]On: 25 March 2008Access Details: [subscription number 731846679]Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Food Reviews InternationalPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713597252
Diseases of Quinoa (Chenopodium quinoa)Solveig Danielsen a; Alejandro Bonifacio b; Teresa Ames ca The Royal Veterinary and Agricultural University, Frederiksberg C, Denmarkb PROINPA Foundation, Choquenaira, La Paz, Boliviac International Potato Center (CIP), Lima, Peru
Online Publication Date: 05 January 2003To cite this Article: Danielsen, Solveig, Bonifacio, Alejandro and Ames, Teresa(2003) 'Diseases of Quinoa (Chenopodium quinoa)', Food Reviews International,19:1, 43 - 59To link to this article: DOI: 10.1081/FRI-120018867URL: http://dx.doi.org/10.1081/FRI-120018867
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
Diseases of Quinoa (Chenopodium quinoa)
Solveig Danielsen,1,2,* Alejandro Bonifacio,3 and Teresa Ames1
1International Potato Center (CIP), Lima, Peru2The Royal Veterinary and Agricultural University, Thorvaldsensvej 40,
Frederiksberg C, Denmark3PROINPA Foundation, Choquenaira, La Paz, Bolivia
ABSTRACT
Downy mildew caused by Peronospora farinosa is the most damaging disease of
quinoa (Chenopodium quinoa), an ancient Andean grain crop. The disease has been
reported from all areas of quinoa cultivation. In the Andean highlands, it is considered
endemic. Despite the disease’s wide dissemination and significant effect on quinoa
crop production, little is known about its epidemiology, host specialization, population
structure, and host plant resistance. There is a similar knowledge gap regarding other
quinoa diseases, such as Rhizoctonia damping off, Fusarium wilt, leaf spot (Ascochyta
hyalospora), seed rot and damping off (Sclerotium rolfsii, Pythium zingiberum), and
brown stalk rot (Phoma exigua var. foveata). These diseases are less widespread than
downy mildew but are still considered potential production constraints, particularly
when the crop is introduced in areas outside its traditional growing regions. This article
provides an overview of current knowledge on downy mildew and other diseases
affecting quinoa production.
INTRODUCTION
Until recently, the diseases of quinoa (Chenopodium quinoa Willd.) received little
attention; few studies have been carried out on their epidemiology or their potential effects
43
DOI: 10.1081/FRI-120018867 8755-9129 (Print); 1525-6103 (Online)
Copyright q 2003 by Marcel Dekker, Inc. www.dekker.com
*Correspondence: Solveig Danielsen, Danish Government Institute of Seed Pathology for
Developing Countries (DGISP), Denmark; E-mail: [email protected].
FOOD REVIEWS INTERNATIONALVol. 19, Nos. 1 & 2, pp. 43–59, 2003
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
on crop production. The few available reports of quinoa disease occurrence around the
world have highlighted this knowledge gap. These reports include: Rhizoctonia damping
off and Fusarium wilt in Peru (Barboza et al., 2000); seed rot and damping off (caused by
Sclerotium rolfsii) in the United States (Beckman and Finch, 1980); damping off (Pythium
zingiberum) in Japan (Ikeda and Ichitani, 1985); and leaf spot (Ascochyta hyalospora)
(Boerema et al., 1977), and brown stalk rot (Phoma exigua var. foveata) in the Andean
region of South America (Otazu and Salas, 1977). These diseases have only been reported
sporadically, however, and are probably of minor importance as quinoa production
constraints.
In contrast, downy mildew caused by Peronospora farinosa is an important and
widespread disease in quinoa growing areas in Peru, Bolivia, Ecuador, and Colombia
(Alandia et al., 1979; Aragon and Gutierrez, 1992; Mujica et al., 1998). The disease is
consistently present in these areas and is thus regarded as endemic. Downy mildew has
also been detected in Canada (Tewari and Boyetchko, 1990) and Europe (John Hockenhull
and Sven-Erik Jacobsen, pers. comm., and Danielsen et al., 2002).
Despite the importance and wide dissemination of this disease, little is known about
its epidemiology, host specialization, and impact on grain yield, or the structure of its
pathogen populations and sources of resistance in quinoa. Estimates of yield losses due to
downy mildew vary between 20 and 25%, indicating that this disease is an important
constraint to quinoa production (Alandia et al., 1979). Because these data are based on
only one experiment with one cultivar, quantifying the effect of downy mildew on quinoa
yield requires further study.
Most research reports on downy mildew in quinoa focus on the detection of the
pathogen and the determination of differences in susceptibility among cultivars, under
natural growing conditions in the Andean region. This data is documented mainly in
technical reports from research center field stations, or in thesis reports, and is therefore
inaccessible to the public. Thus, the scientific literature on this disease is sparse compared
to that on downy mildew of spinach and beet, two economically important crops that are
also attacked by P. farinosa. A quick search of the CAB International abstracts database
(1973–2000) illustrates this disparity, generating 106, 109, and four hits for downy
mildew of beet, spinach, and quinoa, respectively.
To help fill this gap, we surveyed, compiled, and summarized existing data to provide
an overview of the current knowledge base on quinoa crop disease, with special emphasis
on downy mildew. The data presented below are based on a literature review and on recent
research carried out at the International Potato Center (CIP) in Lima, Peru.
LEAF AND STALK DISEASES
Downy Mildew
The Pathogen
Taxonomy
Peronospora farinosa (Fr.) Fr., the causal agent of downy mildew of quinoa, is an
oomycete belonging to the Peronosporaceae family (Peronosporales order), a group of
Danielsen, Bonifacio, and Ames44
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
highly specialized, biotrophic organisms that are parasitic on vascular plants and cause
downy mildew across a limited host range (Ingram, 1981). Oomycetes have recently been
excluded from the Fungi kingdom, but their exact classification remains controversial.
Some scientists place them in the Stramenopila kingdom, whereas others place them in the
Chromista kingdom. Peronospora farinosa attacks members of the Chenopodiaceae
family, including beet, spinach, and quinoa, and has been found on all continents
(Anonymous, 1988). A subdivision of P. farinosa was proposed by Byford (1967), who
found that isolates of P. farinosa from sugar beet (Beta vulgaris), fat hen (Chenopodium
album), and spinach (Spinacia oleracea) would only infect the host species from which
they were isolated. He therefore suggested the following classification: P. farinosa f.sp.
betae on Beta spp., P. farinosa f.sp. spinaciae on Spinacia spp., and P. farinosa f.sp.
chenopodii on Chenopodium spp. However, the failure of spores from C. album to infect
C. amaranticolor and C. capitatum (Byford, 1967) and the lack of cross-infection between
isolates from C. murale, C. album, and C. ambrosioides (Aragon and Gutierrez, 1992),
suggests that further specialization occurs. In other experiments, conducted in Peru, the
quinoa cv. Blanca de Junın was not infected by downy mildew isolates from the cv. La
Molina 89 (Aragon and Gutierrez, 1992), and another Andean grain crop, C. pallidicaule,
was not infected by P. farinosa isolates from quinoa (Alandia et al., 1979). This indicates
the existence of different pathotypes within this forma specialis, as well as different
sources of resistance among quinoa genotypes. In general, a high degree of physiological
specialization has been found within the Peronosporaceae family (Crute, 1981).
Infection Biology
The presence of free water and RH . 85% is necessary for conidia of the genus
Peronospora to germinate (Alandia et al., 1979; Develash and Sugha, 1996; Hartmann
et al., 1983). The germ tube penetrates the cuticle of the leaf and the mycelium ramifies
intercellularly, through the leaf and stalk tissues. Haustoria formed from hyphae penetrate
the cells, and sporangiophores emerge either individually or in groups, through the
stomata, to form ramifications with lemon-shaped sporangia. In general, Peronospora
species require moderate temperatures (10–208C) for optimal sporulation (Byford, 1981;
Develash and Sugha, 1996; Doshi and Thakore, 1993; Frinking and van der Stoel, 1987;
Hartmann et al., 1983; Shao et al., 1990). Species of the genus Peronospora do not form
zoospores. The asexual spores (sporangia; also called conidia) are dispersed by wind and
rain and are assumed to be of utmost importance for spread of the disease during the
growing season (Michelmore et al., 1988).
Sexual Reproduction
While heterothallism has been reported in P. farinosa f.sp. spinaciae (Asch and
Frinking, 1988; Inaba and Morinaka, 1984), the mating system of P. farinosa f.sp.
chenopodii has not yet been studied. Crosses of eight single-lesion isolates from different
regions in Peru and Bolivia in all possible combinations, using a detached leaf assay,
revealed the presence of two mating types, P1 and P2 (Danielsen et al., 2000), indicating
that P. farinosa f.sp. chenopodii is heterothallic. Oospores have been found in large
numbers in old, downy-mildew infected leaves from several quinoa growing areas in Peru
Diseases of Quinoa 45
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
(Danielsen and Ames, unpublished), indicating that both mating types are present in these
areas. In contrast, oospores were not found in naturally infected quinoa leaves from the
coastal areas of Lima (Peru), indicating that only one mating type exists in that area or that
environmental conditions are unfavorable for oospore formation.
The specific role of soil- and seed-borne oospores in the spread and initiation of
epidemics of downy mildew of quinoa is not known. However, considering the harsh
climatic conditions of the Andean highlands, which include long periods of drought and
extreme temperatures, it seems clear that oospores play a crucial role in the survival of the
disease. Past research also provides strong indications that seed- and soil-borne oospores
act as a primary source of downy mildew infection. Alandia et al. (1979) reported the
presence of oospores in the integument of quinoa seeds, and International Potato Center
(CIP) scientists observed heavy downy-mildew sporulation on 3–6% of the cotyledons of
six quinoa cultivars from Puno and Cusco (Peru) in 2- to 3-week-old quinoa seedlings
raised from seed and grown in pots of sterilized soil in greenhouses under humid and
temperate conditions, followed by rapid spread to all seedlings (data not shown). While
microscopy of the seed washing water did not reveal any oospores, numerous viable
oospores were found inside the flower residues and seed integument. Based on these
observations, seed-borne inoculum should be an important consideration in the future
commercialization and certification of quinoa seed.
Peronospora Farinosa Populations
Despite the scarcity of data, the following indicators imply that downy mildew
populations in Andean countries are highly diverse: 1) the downy mildew host has a high
degree of genetic diversity and plasticity, making it adaptable to a wide range of
agroecological zones (Risi and Galwey, 1989); 2) downy mildew of quinoa is detected in
climatically and geographically diverse areas, indicating a high degree of adaptability; and
3) sexual reproduction of P. farinosa in quinoa occurs frequently, allowing the pathogen to
constantly expand its genotypic diversity and develop new and more aggressive
pathotypes. Deciphering the pathogen’s genetic composition is an important prerequisite
for developing an effective control strategy. Therefore, determining the presence of
pathotypes and mating types, and their relative frequency and geographic distribution,
would be a first step in developing an effective control strategy for this pathogen.
The only research conducted on virulence factors in downy mildew of quinoa is that
reported by Ochoa et al. (1999). Twenty isolates collected from different parts of Ecuador
were tested on 39 quinoa lines, using a seedling assay to identify virulence factors. Four
virulence groups and three resistance factors were identified, and a preliminary differential
set of four quinoa lines was proposed for identification of virulence groups.
For some purposes, virulence factors are excellent markers for genotypic variation
within a population. However, these types of markers are vulnerable to the process of host
selection. As the use of molecular markers provides a means of testing genotypic variation
with neutral markers, DNA fingerprinting (Random Amplified Polymorphic DNA
[RAPD]) has been used to identify P. parasitica pathotypes (Jensen, 1996; Tham et al.,
1994), and Restriction Fragment Length Polymorphism (RFLP) and AFLP markers are
being used now to characterize strains of Phytophthora infestans from potato in Peru
(Gamboa et al., 1999). AFLPs offer a potentially powerful tool for detecting polymorphic
Danielsen, Bonifacio, and Ames46
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
DNA sequences in fungi (Lin et al., 1996; McDonald, 1997). The advantage of this
technique is that no previous knowledge on DNA sequences is required, and only small
amounts of DNA are needed to perform the test.
At present, research is being carried out at the CIP to characterize populations of the
downy mildew pathogen in Peru and Bolivia using virulence and Amplified Fragment
Length Polymorphism (AFLP) markers.
The Disease
Symptoms
Quinoa plants infected by P. farinosa present a variety of symptoms, depending on the
cultivar, the growth stage, and the environmental conditions. Typical symptoms include
pale or yellow chlorotic lesions on the surface of the leaf (which cause a reduction in the
photosynthetic area of the plant and eventually turn necrotic) and gray-violaceous
sporulating areas on the underside of the leaf. In some cultivars, the lesions are small and
numerous, whereas in others, the lesions are large, diffuse, and irregular. In some cultivars,
the lesions turn reddish to purple; a hypersensitive type of reaction (small necrotic flecks)
has also been observed. The degree of sporulation varies considerably within cultivars,
most likely due to genetic variation within the pathogen population and differences in the
cultivar response. It has also been noted that severe infection leads to heavy premature leaf
loss (Alandia et al., 1979). However, it is difficult to estimate how much defoliation is
caused specifically by downy mildew, as there are a number of other known factors, such
as physiological stress (e.g., drought, salinity, and frost) that cause defoliation. Systemic
infection has been observed, which may lead to dwarfing and yellowing (Alandia et al.,
1979; Frinking and Linders, 1986; Tewari and Boyetchko, 1990). Systemic infection is
probably caused by seed-transmitted oospores, as seen in pea and buckwheat (Vulsteke
et al., 1997; Zimmer et al., 1992).
Downy Mildew in the Andes
The endemic nature of downy mildew in the Andean region makes it an omnipresent
threat to quinoa production whenever conditions are favorable for disease development. It
has a high capacity for spread and development throughout the region, but it does not
occur with the same intensity in the various quinoa production zones defined for Andean
countries. In Bolivia, quinoa production regions have been divided into different types of
ecoregions, including Northern high plateau, Central high plateau, Southern high plateau,
and inter-Andean valley. In the high plateau, average annual rainfall and relative humidity
declines moving from north to south and from east to west. The temperature does not vary
much in this area, although the presence of Lake Titicaca causes a drop in temperatures
during the cropping period (Bonifacio, unpublished).
The conditions most favorable for disease development occur in the northern high
plateau of Bolivia, particularly in the areas surrounding Lake Titicaca. The valleys, which
tend to have a greater quantity of annual rainfall than the high plateau, are also endemic
zones for downy mildew. In the Southern high plateau, where rainfall is scarce, and
humidity is very low, the occurrence of downy mildew is extremely rare and, thus, is not
Diseases of Quinoa 47
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
considered an important production constraint. The conditions of the Central high plateau
(considered a transition zone between the Northern and Southern high plateaus) are only
favorable for the growth of downy mildew when average annual rainfall exceeds normal
levels or when precipitation is concentrated in short periods during the growing season.
Disease Assessment and Yield Loss
A reliable disease assessment method is crucial for disease epidemiology research,
crop-loss assessment, pathogenicity and virulence testing, and screening for resistance/
susceptibility. A large number of disease assessment scales have been proposed for
evaluation of downy mildew in several crops. Downy mildew in pea is measured on
a 0–10 severity scale by Falloon et al. (1995) and on a four-point scale by Mogling and
Broschewitz (1990), while Stegmark (1991) scores the percentage of leaf area infected.
Downy mildew in cabbage crops is measured on 1–9 and 0–5 scales by Jensen et al.
(1999b) and Sousa et al. (1997), respectively. Conclusions from comparison studies of the
various assessment methods indicate that care should be taken when choosing a scale for a
specific purpose. For example, assessment of the use of different methods in detecting the
resistance reaction in potato against early blight (Alternaria solani) showed that cultivar
ranking varied according to the assessment method used (Christ, 1991). Studies measuring
the severity of corky root (Rhizomonas suberifaciens) in lettuce indicated that no single
scale was optimal in all situations (O’Brien and van Bruggen, 1992). Another aspect that
should not be ignored is the effect of human error, which may influence results
significantly (O’Brien and van Bruggen, 1992; Stonehouse, 1994).
Different methods are now being used in Peru and Bolivia to assess the level of downy
mildew in quinoa. A 0–10-point scale has been defined by Otazu et al. (1976), where
0 ¼ no infection, 1 ¼ 1–10% leaf area infected, 2 ¼ 11–20%, etc. Based on this scale,
quinoa cultivars are divided into four groups: resistant ¼ 1–4% infection; moderately
resistant ¼ 5–19%; susceptible ¼ 20–49%, and very susceptible ^ 50%: However, this
scale does not take into account the defoliation caused by downy mildew. Another
evaluation method, developed by Bonifacio and Saravia (1999), considers the effect of
defoliation by only including the middle, foliated third of the plant in the evaluation. The
percentage of infected leaf area in this fraction of the plant is scored on a 0 to 5 scale,
where 0 ¼ no infection; 1 ¼ 1–20% leaf area infected, 2 ¼ 21–40%, etc.
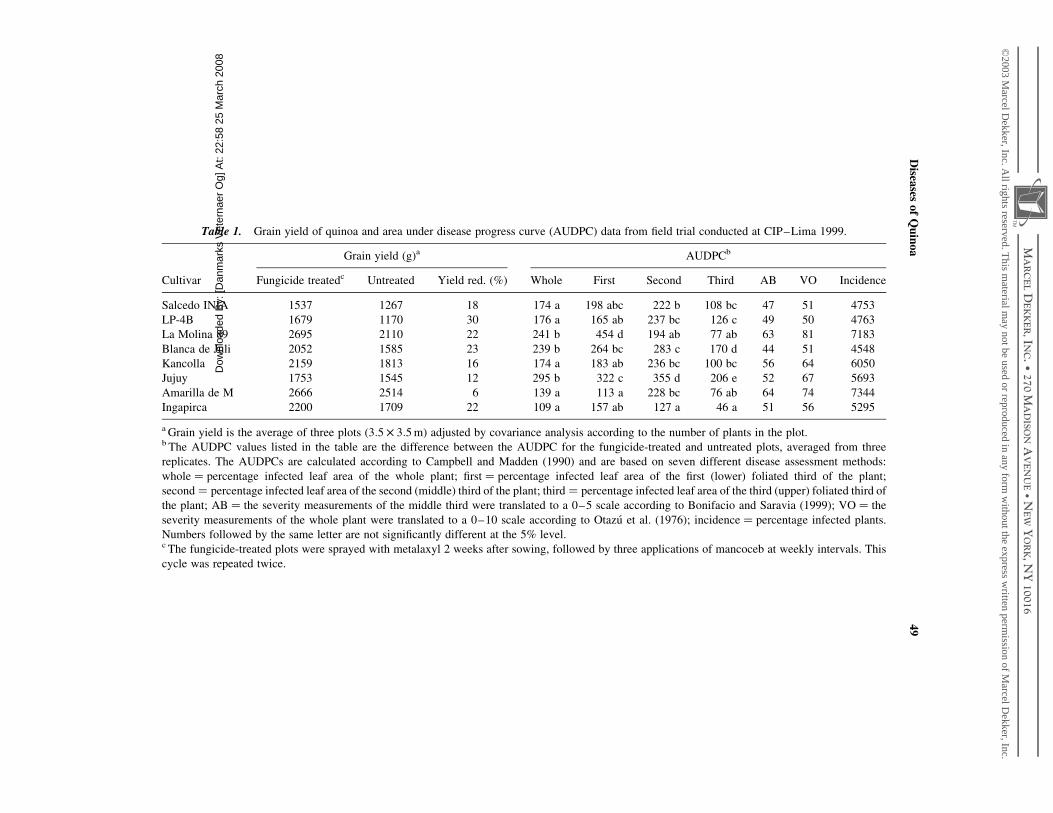
To evaluate existing disease assessment methods and to assess yield loss due to
downy mildew, a field trial was carried out at the International Potato Center (Lima, Peru)
during the winter months of 1999. Quinoa is not usually grown along the Peruvian coast,
where mean annual temperatures are higher than in the traditional quinoa growing areas in
the highlands. The trial included eight quinoa cultivars and two levels of downy mildew
infection (with and without fungicide treatment) in a split plot design with three replicates,
where fungicide treatment was the main plot and cultivar the subplot. Yield data were
adjusted according to the number of plants in each plot, using covariance analysis, and
then correlated to the area under the disease progress curves (AUDPCs), based on disease
severity measurements of the whole plant, the first (lower) section, the second (middle)
section, and the third (upper) section, and on the VO-scale (Otazu et al., 1976), the
AB-scale (Bonifacio and Saravia, 1999), and disease incidence (percent of plants infected)
(Table 1). The disease was assessed three times during the growth period. The downy
Danielsen, Bonifacio, and Ames48
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
Table 1. Grain yield of quinoa and area under disease progress curve (AUDPC) data from field trial conducted at CIP–Lima 1999.
Grain yield (g)a AUDPCb
Cultivar Fungicide treatedc Untreated Yield red. (%) Whole First Second Third AB VO Incidence
Salcedo INIA 1537 1267 18 174 a 198 abc 222 b 108 bc 47 51 4753
LP-4B 1679 1170 30 176 a 165 ab 237 bc 126 c 49 50 4763
La Molina 89 2695 2110 22 241 b 454 d 194 ab 77 ab 63 81 7183
Blanca de Juli 2052 1585 23 239 b 264 bc 283 c 170 d 44 51 4548
Kancolla 2159 1813 16 174 a 183 ab 236 bc 100 bc 56 64 6050
Jujuy 1753 1545 12 295 b 322 c 355 d 206 e 52 67 5693
Amarilla de M 2666 2514 6 139 a 113 a 228 bc 76 ab 64 74 7344
Ingapirca 2200 1709 22 109 a 157 ab 127 a 46 a 51 56 5295
a Grain yield is the average of three plots ð3:5 £ 3:5 mÞ adjusted by covariance analysis according to the number of plants in the plot.b The AUDPC values listed in the table are the difference between the AUDPC for the fungicide-treated and untreated plots, averaged from three
replicates. The AUDPCs are calculated according to Campbell and Madden (1990) and are based on seven different disease assessment methods:
whole ¼ percentage infected leaf area of the whole plant; first ¼ percentage infected leaf area of the first (lower) foliated third of the plant;
second ¼ percentage infected leaf area of the second (middle) third of the plant; third ¼ percentage infected leaf area of the third (upper) foliated third of
the plant; AB ¼ the severity measurements of the middle third were translated to a 0–5 scale according to Bonifacio and Saravia (1999); VO ¼ the
severity measurements of the whole plant were translated to a 0–10 scale according to Otazu et al. (1976); incidence ¼ percentage infected plants.
Numbers followed by the same letter are not significantly different at the 5% level.c The fungicide-treated plots were sprayed with metalaxyl 2 weeks after sowing, followed by three applications of mancoceb at weekly intervals. This
cycle was repeated twice.
Disea
seso
fQ
uin
oa
49
MA
RC
EL D
EK
KE
R, IN
C. •
270 MA
DISO
N A
VE
NU
E •
NE
W Y
OR
K, N
Y 10016
©2003 M
arcel Dekker, Inc. A
ll rights reserved. This m
aterial may not be used or reproduced in any form
without the express w
ritten permission of M
arcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
mildew level was kept low in the fungicide-treated plots. The general disease level in the
untreated plots was low, the AUDPC not exceeding 454 (as per measurement of the first
third part of the plant) in any cultivar.
A reduction in grain yield due to fungicide treatment was found in all cultivars
(6–29%); however, the yield loss was not significant in any cultivar, due to large
differences in variation coefficients ðR ¼ 1%–59%Þ: By contrast, a significant negative
correlation was found between yield and AUDPCwhole, AUDPCfirst, AUDPCsecond, and
AUDPCthird ðR ¼ 240–42%; p ¼ 0:0033 2 0:0054Þ: The AUDPCAB, AUDPCVO, and
AUDPCincidence values were not significantly correlated to yield. This shows that only
about 40% of the yield loss can be explained by downy mildew assessed by the first four
disease assessment methods, and that the five-point and 10-point scales and disease
incidence are not useful methods for downy mildew assessment when disease pressure is
low. The differences in yield and AUDPC (whole, first, second, third) between treated and
untreated plots were ranked using the Mann–Whitney test and subsequently compared
using a P2 test, where yield was the theoretical value and the AUDPCs the observed
values. None of the AUDPC rankings showed significant similarity to the yield ranking,
with AUDPCfirst showing the best fit and AUDPCsecond the worst. This underscores the fact
that, in this case, AUDPCs based on the measurement of percent leaf area infected may be
inadequate to predict yield loss. However, further study is required to test the value of the
different disease assessment methods under varying disease levels.
The tendency of the quinoa plant to defoliate under various stress conditions
(drought, salt, frost, downy mildew, etc.) complicates disease assessment in this crop, as
the measurement of the percent of diseased leaf area does not take into account the total
amount of foliage. One consequence of this leaf shed is that the level of downy mildew
infection of the lower part of the plant will not be registered properly, which may result in
skewed disease scores. This fact is clearly illustrated by the disease progress curve for cv.
La Molina 89 based on the lower third part of the foliage (Fig. 1). For the untreated plot,
there is a drastic drop in disease severity from the first to the second evaluation, due to the
fact that the infected leaves on the lower part of the plant were shed during this period, and
because the new and upper leaves were not reinfected at the same degree. Scoring shed
leaves at 100% disease severity does not correct the problem, due to the risk of including
leaves in this category that are shed for reasons other than downy mildew.
Figure 1. Disease progress curve for cv. La Molina 89 based on severity measurements of the
lower-third part of the foliage.
Danielsen, Bonifacio, and Ames50
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
Potential ways to overcome this problem include measurements of the green leaf area
(Bryson et al., 1997), percent reflectance at 800 nm wavelength (Nutter, 1989), or
reflectance at two wavelengths (Jacobsen et al., 1998). In the latter case, it is possible to
calculate a Relative Vegetation Index (RVI), which is highly correlated to biomass and
final seed yield and should, therefore, reveal any difference in leaf shed due to different
levels of downy mildew within a cultivar. Reflectance measurements may be a valuable
additional parameter for measuring disease severity and predicting yield loss, when a
negative control is available.
Control Measures
The traditional way of controlling downy mildew in various crops is through
application of fungicides such as metalaxyl. However, the use of such fungicides exposes
the environment and humans to health hazards. There is also a risk that the pathogen will
develop resistance to metalaxyl, as shown for P. parasitica (Crute and Gordon, 1986) and
Phytophthora infestans (Davidse et al., 1981). Moreover, the majority of quinoa growers
are resource-poor farmers who tend to practice low-input farming. Within a scenario of
rapidly spreading, endemic disease and sexually reproducing pathogen populations, these
farmers have only two options for disease control: effective cultural practices and the use
of durable host plant resistance.
Cultural Practices
Experiences from Bolivia show that cultural practices that minimize the relative
humidity in the microclimate may have an effect on the severity of downy mildew. Among
these are: adjusting the spacing between rows and individual plants, orienting the field
against the predominant wind direction, and avoiding excess water in the field (Bonifacio,
unpublished). However, cultural practices alone are insufficient for controlling a disease
such as downy mildew in quinoa, which has the ability to adapt, spread, and reproduce.
Host Resistance
Resistance to downy mildew has been the subject for genetic improvement for several
years. To date, however, improved varieties with reliable resistance are still not available,
although differences among quinoa cultivars in susceptibility to downy mildew have been
observed. A 1976 study from Peru describes 11 C. quinoa genotypes as resistant or
moderately resistant to P. effusa (former name of P. farinosa) at flowering and
fructification (Otazu et al., 1976). In Bolivia, plant breeders are working to increase the
level of durable resistance against downy mildew in quinoa, and a number of promising
accessions have been selected for future crossings (Bonifacio and Saravia, 1999). These
studies show that resistance sources are available, but the genetic mechanisms behind the
resistance (horizontal versus vertical) have not yet been investigated.
Quinoa farmers in the Andean region, however, have accumulated a large amount of
knowledge about the growing characteristics of quinoa cultivars, including grain
characteristics, adaptability to different geographic zones, and reaction to adverse factors
Diseases of Quinoa 51
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
such as drought, cold, salinity, and downy mildew. Susceptibility to downy mildew is
believed to be related to the length of the growth season, as the early cultivars appear to be
the most susceptible (Bonifacio and Saravia, 1999).
Within the C. quinoa species, the highest degree of resistance to downy mildew is
found in accessions from the valleys or lower ecoregions of Bolivia, Peru, and Ecuador. In
contrast, the ecotypes of the Southern high plateau are the most susceptible. In
evolutionary terms, this can be explained by the domestication process of the species,
where only genotypes with high resistance have survived in environments with favorable
conditions for downy mildew occurrence. Inversely, genotypes domesticated in very dry
areas have no resistance to downy mildew, either because the resistance genes have been
“lost” during evolution, or because genotypes without resistance have an evolutionary
advantage over resistant genotypes in these areas.
Genetic resistance to downy mildew is found not only in cultivated quinoa but also in
wild species that grow more or less associated with the cultivated crop. There are
indications that wild species such as C. hircinum, C. nuttalliae, C. petiolare, C. album, and
C. ambrosioides harbor downy mildew resistance genes. These sources may be useful for
incorporating resistance into commercial varieties, by traditional hybridization techniques
or by genetic transformation. At present, a large part of the genetic diversity of the genus
Chenopodium, mainly C. quinoa, is stored in germplasm collections in Bolivia, Peru, and
Ecuador (,5000 accessions). Many of these accessions have been characterized for
agronomic traits, including susceptibility to downy mildew, but a complete evaluation of
plant material for resistance to downy mildew has not been conducted with enough
continuity to allow for actual genetic improvement.
Screening for Resistance
Current knowledge is limited by the lack of experiments to identify the pathogen–
host interactions under controlled conditions. Therefore, it is not known whether the
variation in downy mildew incidence under different geographic conditions is due to
variation in environmental conditions or to variation within the pathogen populations.
Existing data on susceptibility and resistance of quinoa cultivars originates almost
exclusively from experiments based on natural infection. This creates uncertainty about
the inoculum level, the possible presence of focal infection points, and the composition of
the pathogen population. Screening for resistance in the field based on natural infection is
time-consuming and laborious. It should only be done in highly conducive areas, with
known pathogen populations and consistently high disease pressure, in order to ensure
reliable and reproducible results. Such “hot spot” areas have yet to be identified for the
quinoa/downy mildew system.
Screening methods based on artificial inoculation of seedlings or detached leaves
under controlled conditions may be a useful tool for quinoa breeders. A large amount of
literature is available on screening for resistance against downy mildew in several crops.
In pea and cabbage, cotyledons or seedlings with a few true leaves can be used in rapid
screening assays for breeding purposes; leaves are inoculated with a known concentration
of sporangia and incubated for 7 days under controlled conditions (Coelho et al., 1998;
Grontoft, 1993; Jensen et al., 1999a; Klodt-Bussmann and Paul, 1995; Nashaat and
Awasthi, 1995; Natti, 1958; Stegmark, 1991; Taylor et al., 1989). A detached leaf assay is
Danielsen, Bonifacio, and Ames52
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
currently being used at the International Potato Center (CIP) to evaluate resistance against
Phytophthora infestans in potato (Goth and Keane, 1997; Mizubuti and Fry, 1998). Such
screening systems offer many advantages, as they are quick and do not require much
space. A good correlation between seedling and adult plant resistance has been found
(Grontoft, 1993; Jensen et al., 1999b; Natti, 1958; Stegmark, 1991), although some
contradictions have been noted (Dickson and Petzoldt, 1993).
Ochoa et al. (1999) have established a seedling assay to identify virulence and
resistance factors in the quinoa/downy mildew system. Eighteen-day-old seedlings are
inoculated with a spore suspension (105 spores/mL) and incubated for 7 days under fixed
temperature and light regimes. The disease reaction is scored according to a 0–5 scale
(Table 2).
A similar assay is being used at the International Potato Center to characterize disease
reaction. Detached quinoa leaves from 4- to 6-week-old seedlings are placed upside down
on water agar plates moistened with 1 mL of spore suspension (105 spores/mL). After 1
week of incubation under controlled conditions, the degree of sporulation is scored
according to the scale shown in Fig. 2 (Danielsen and Ames, 2000).
These assays may be appropriate for large-scale screening of quinoa germplasm and
for studies of downy mildew populations. The method of choice would depend on the
availability of equipment and space.
Leaf Spot
Leaf spot of quinoa is a seed-borne disease caused by Ascochyta hyalospora (Cooke
and Ellis). Typical leaf spot symptoms include light-colored, circular spots (0.5–1.0 cm)
with a light-brown edge. The spots eventually turn necrotic, and black pycnidia can be
seen with the naked eye (Alandia et al., 1979; Boerema et al., 1977). Heavy infection leads
to leaf loss. In studies of Bolivian quinoa seed samples, observed levels of infection by the
Table 2. Scale for scoring disease reactions of downy mildew (Peronospora farinosa f.sp.
chenopodii) in primary leaves of quinoa (Chenopodium quinoa).
Reaction type Classification Symptoms
0 Resistant No evidence of symptoms, no perceptible necrosis
1 Resistant Small chlorotic-necrotic lesions (2–5 mm) with truncated
mycelium in the mesophyl of the leaf
2 Resistant Small chlorotic lesions (4–8 mm) with a little sporulation
3 Susceptible Medium-sized, confined chlorotic lesions with sporulation;
sporulation mainly on the lower-leaf surface
4 Susceptible Large, not clearly confined chlorotic lesions with sporulation;
sporulation mainly on the lower-leaf surface
5 Susceptible Mild chlorosis with abundant sporulation on upper and lower
leaf surfaces
Source: Ochoa et al., 1999.
Diseases of Quinoa 53
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
leaf spot pathogen ranged from 7.8–26.3% (Boerema et al., 1977). Germination testing in
the same study showed that the fungus produces mild to severe browning in the root and
hypocotyl; some of the most severely infected seedlings died.
During the growth season 1999–2000, leaf spot was observed in several quinoa
fields in Puno, Peru, mainly on cv. Kanccolla (data not shown). The geographic
distribution and economic importance of leaf spot are not known. The disease is
frequently reported from fields in the highlands of Peru, but it is not considered a major
constraint to production.
Brown Stalk Rot
Brown stalk rot caused by Phoma exigua var. foveata (Foister) Boerema was observed
for the first time in Puno in 1974–1975 (Otazu and Salas, 1977). Since then, the disease
has been reported frequently from the Andean highlands (Alandia et al., 1979). Initial
symptoms are small, humid spots on the upper-third part of the stalk. The lesions grow to
cover most of the stalk, and inflorescence and pycnidia become visible. The foliage
becomes chlorotic, and the stalk bends downward and tends to break easily.
Phoma exigua is a soil-borne pathogen that is seen in areas with low temperatures and
high humidity, common conditions in the Andean highlands during the growing season.
Pathogenicity tests have shown that mechanical injury of the plant is a prerequisite for
infection. In the Andean highlands, hailstorms frequently cause such mechanical damage
to plants (Otazu and Salas, 1977).
Figure 2. Sporulation index for determination of downy mildew (Peronospora farinosa f.sp.
chenopodii) susceptibility/resistance of quinoa genotypes in a detached leaf assay. The black dots
represent downy mildew sporangia. 0 ¼ no sporulation; 1 ¼ one to few single sporangiophores,
sporulation barely visible; 2 ¼ a few sporangiophores, scattered or in a small, dense group,
sporulation visible; 3 ¼ diffuse sporulation on the whole leaf, or dense sporulation on less than 50%
of the leaf; 4 ¼ moderate sporulation on the whole leaf, or dense sporulation on 50–90% of the leaf;
and 5 ¼ dense sporulation on more than 90% of the leaf. 0–2 ¼ resistant; 3–5 ¼ susceptible.
Danielsen, Bonifacio, and Ames54
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
Cross-inoculations showed that P. exigua var. foveata, which causes brown stalk
rot in quinoa, is the same pathogen that causes gangrene in potato (Logan, 1974; Otazu
and Salas, 1977). The frequent use of quinoa–potato crop rotations in the Andean
highlands may be a possible factor in the buildup of inoculum in the soil, which
increases disease pressure. However, neither of these diseases occurs at a level beyond
sporadic incidence.
ROOT AND SEEDLING DISEASES
Rhizoctonia Damping Off and Fusarium Wilt
Root and seedling diseases in quinoa caused by Rhizoctonia and Fusarium were
not mentioned in the literature. However, during the 1998 growing season, Rhizoctonia
solani and Fusarium spp. were isolated from a field of quinoa at the International
Potato Center (Barboza et al., 2000). In pathogenicity tests performed under
greenhouse conditions, R. solani produced symptoms similar to those observed in the
field caused by damping off: failure in seed germination and sunken lesions in the main
stems of old plants at ground level. None of the R. solani isolates anastomosed with
AG 1–7 tester strains, but they did anastomose with each other. Fusarium spp.
reproduced wilting and root rot in old plants.
Because both fungi are soil-borne pathogens that also affect several other hosts,
causing similar symptoms, the inoculum potential in the soil could be an important factor
in enabling the disease to cause damage.
Seed Rot and Damping Off
Seed rot and damping off of quinoa caused by Sclerotium rolfsii Sacc. was first
reported in California in 1980 (Beckman and Finch, 1980), when pre- and post-emergent
damping off of seedlings was observed in a quinoa field. Diseased seedlings showed stem
girdling, and collapse appeared in patchy areas in the field. Sclerotium rolfsii was
consistently isolated from diseased seedlings, and subsequent pathogenicity tests
confirmed that the isolated fungus was the causal agent of the disease.
Another potential constraint to quinoa is damping off caused by Pythium zingiberum,
a pathogen that causes rhizome rot in ginger (Ikeda and Ichitani, 1985). Inoculation of the
soil with P. zingiberum oospores caused damping off of quinoa seedlings within 10 days of
incubation at 308C; C. quinoa proved more susceptible than C. amaranticolor. In a
greenhouse experiment, an unidentified Pythium species killed emerging Amaranthus
seedlings, particularly at high moisture levels, whereas quinoa seedlings remained healthy
(Aufhammer et al., 1994). In both studies, however, differences in cultivar susceptibility
were not considered.
Seed rot and damping off caused by Sclerotium and Pythium have not yet been
reported in South America but should be considered potential production constraints when
quinoa is introduced into areas outside its traditional growing regions.
Diseases of Quinoa 55
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
ACKNOWLEDGMENTS
We thank John Hockenhull and Abigail Hollister for their valuable comments on the
manuscript. The research project on downy mildew of quinoa was supported by the Danish
government, Ministry of Foreign Affairs (RUF grant no. 90929).
REFERENCES
Alandia, S., Otazu, V., Salas, B. (1979). Enfermedades. In: Tapia, M., Gandarillas, H.,
Alandia, S., Cardozo, A., Mujica, A., Ortiz, R., Otazu, V., Rea, J., Salas, B., Sanabria,
E., eds. Quinua y Kaniwa. Bogota Colombia: IICA, pp. 137–148.
Anonymous (1988). Distributions Maps of Plant Diseases. UK No. 28. UK:
Commonwealth Mycological Institute (CMI), CAB International.
Aragon, L., Gutierrez, W. (1992). El mildiu en cuatro especies de Chenopodium.
Fitopatologıa 27:104–109.
Asch, M. V., Frinking, H. D. (1988). Heterothallism in Peronospora farinosa f.sp.
spinaciae. Trans. Br. Mycol. Soc. 91:692–693.
Aufhammer, W., Kaul, H. P., Kruse, M., Lee, H. J., Schwesig, D. (1994). Effects of sowing
depth and soil conditions on seedling emergence of amaranth and quinoa. Eur.
J. Agron. 3:205–210.
Barboza, J., Ames, T., Jacobsen, S. E. (2000). Reduccion de la germinacion y muerte de
plantas de Chenopodium quinoa causadas por Rhizoctonia y Fusarium. Memorias del
Primer Taller Internacional sobre Quinua., May 10–14, 1999, Lima, Peru, in
preparation La Molina: Centro Internacional de la Papa y Universidad Nacional
Agraria.
Beckman, P. M., Finch, H. C. (1980). Seed rot and damping-off of Chenopodium quinoa
caused by Sclerotium rolfsii. Plant Dis. 64:497–498.
Boerema, G. H., Mathur, S. B., Neergaard, P. (1977). Ascochyta hyalospora (Cooke &
Ell.) comb. nov. in seeds of Chenopodium quinoa. Neth. J. Plant Pathol. 83:153–159.
Bonifacio, A., Saravia, R. (1999). Evaluacion de la resistencia al mildiu en quinua. In
Tercer Taller de Preduza en Resistencia Duradera en Cultivos Altos en al Zona
Andina Cochabamba, Bolivia, de Septiembre de 27–29 1999, pp. 49–59.
Bryson, R. J., Paveley, N. D., Clark, W. S., Sylvester-Bradley, R., Scott, R. K. (1997). Use
of in-field measurements of green leaf area and incident radiation to estimate the
effects of yellow rust epidemics on the yield of winter wheat. Eur. J. Agron. 7:53–62.
Byford, W. J. (1967). Host specialization of Peronospora farinosa on Beta, Spinacia and
Chenopodium. Trans. Br. Mycol. Soc. 50:603–607.
Byford, W. J. (1981). Downy mildews of beet and spinach. In: Spencer, D. M., ed. The
Downy Mildews. London: Academic Press, pp. 531–543.
Campbell, C. L., Madden, L. V. (1990). Introduction to Plant Disease Epidemiology. New
York: John Wiley & Sons, p. 532.
Christ, B. J. (1991). Effect of disease assessment method on ranking potato cultivars for
resistance to early blight. Plant Dis. 75:353–356.
Coelho, P., Bahcevandziev, K., Valerio, L., Monteiro, A., Leckie, D., Astley, D., Crute,
I. R., Boukema, I. (1998). The relationship between cotyledon and adult plant
Danielsen, Bonifacio, and Ames56
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
resistance to downy mildew (Peronospora parasitica) in Brassica oleracea. Acta
Hortic.335–342.
Crute, I. R. (1981). The host specificity of peronosporaceous fungi and the genetics of the
relationship between host and parasite. In: Spencer, D. M., ed. The Downy Mildews.
London: Academic Press, pp. 237–253.
Crute, I. R., Gordon, P. L. (1986). Downy mildew. In 36th Annual Report National
Vegetable Research Station for 1985. Wellesbourne, UK, pp. 59–60.
Danielsen, S., Ames, T. (2000). El mildiu (Peronospora farinosa) de la quinua
(Chenopodium quinoa) en la zona andina. Manual practico para el estudio de la
enfermedad y del patogeno., in press Lima, Peru: Centro Internacional de la
Papa.
Danielsen, S., Echegaray, J., Ames, T. (2000). Heterotalismo en Peronospora farinosa
f.sp. chenopodii. XVI Congreso Peruano de Fitopatologıa, October 1–6. Peru: Univ.
Nacional de Piura
Danielsen, S., Jacobsen, S. E., Hockenhull, J. (2002). First report of downy mildew of
quinoa caused by Peronospora farinosa f.sp. chenopodii in Denmark. Plant Disease
86, 1175.
Davidse, L. C., Looijen, D., Turkensteen, L. J., Wal, D. V. d. (1981). Occurrence of
metalaxyl-resistant strains of Phytophthora infestans in Dutch potato fields. Neth.
J. Plant Pathol. 87:65–68.
Develash, R. K., Sugha, S. K. (1996). Sporangial viability and germination in
Peronospora destructor. Ind. Phytopathol. 49:157–166.
Dickson, M. H., Petzoldt, R. (1993). Plant age and isolate source affect expression of
downy mildew resistance in broccoli. HortScience 28:730–731.
Doshi, A., Thakore, B. B. L. (1993). Effect of certain environmental factors on
germination of conidia of Peronospora arborescens (Berk.) de Bary. J. Phytol. Res.
6:59–62.
Falloon, R. E., Viljanen-Rollinson, S. L. H., Coles, G. D., Poff, J. D. (1995). Disease
severity keys for powdery mildew and downy mildews of pea, and powdery scab of
potato. N. Z. J. Crop Hortic. Sci. 23:31–37.
Frinking, H. D., van den Stoel, M. C. (1987). Production of conidia by Peronospora
farinosa f.sp. spinaciae. Neth. J. Plant Pathol. 93:189–194.
Frinking, H. D., Linders, E. G. A. (1986). A comparison of two pathosystems: downy
mildew on Spinacia oleracea and on Chenopodium album. Neth. J. Plant Pathol.
92:97–106.
Gamboa, S., Perez, W., Nelson, R. (1999). Uso de marcadores moleculares en la
caracterizacion de poblaciones de Phytophthora infestans en Peru. 10th Latin-
american Phytopathological Congress, Guadalajara, Mexico, 27 September–1
October 1999.
Goth, R. W., Keane, J. (1997). A detached-leaf method to evaluate late blight resistance in
potato and tomato. Am. Potato J. 74:347–352.
Grontoft, M. (1993). A rapid screening method for testing the resistance of cotyledons to
downy mildew in Brassica napus and B. campestris. Plant Breed. 110:207–211.
Hartmann, H., Sutton, J. C., Procter, R. (1983). Effects of atmospheric water potential, free
water, and temperature on production and germination of sporangia in Peronospora
parasitica. Can. J. Plant Pathol. 5:70–74.
Diseases of Quinoa 57
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
Ikeda, A., Ichitani, T. (1985). Density of Pythium zingiberum oospores in relation to
infection of Chenopodium quinoa seedlings. Bull. Univ. Osaka Pref. Ser. B 37:13–18.
Inaba, T., Morinaka, T. (1984). Heterothallism in Peronospora effusa. Phytopathology
74:214–216.
Ingram, D. S. (1981). The biochemistry of host–parasite interaction. In: Spencer, D. M.,
ed. Downy Mildews. London: Academic Press, pp. 143–163.
Jacobsen, S.E., Pedersen, H., Jensen, C.R. (1998). Reflectance measurements, a quick and
non-destructive technique for use in agricultural research. International Conference
on Sustainable Agriculture in Tropical and Subtropical Highlands with Special
Reference to Latin America (SATHLA), Rio de Janeiro, Brazil, March 9–13, 1998.
Jensen, B. D. (1996). Resistance to downy mildew (Peronospora parasitica) in cole crops
(Brassica oleracea) Ph.D. Thesis, Copenhagen, Denmark: The Royal Veterinary and
Agricultural University,155.
Jensen, B. D., Værbak, S., Munk, L., Andersen, S. B. (1999a). Characterization and
inheritance of partial resistance to downy mildew, Peronospora parasitica, in
breeding material of broccoli, Brassica oleracea convar. botrytis var. italica. Plant
Breed. 118:549–554.
Jensen, B. D., Hockenhull, J., Munk, L. (1999b). Seedling and adult plant resistance to
downy mildew (Peronospora parasitica) in cauliflower (Brassica oleracea convar.
botrytis var. botrytis). Plant Pathol. 48:604–612.
Klodt-Bussmann, E., Paul, V. H. (1995). Studies on the preservation and aggressiveness of
Peronospora parasitica and results with regard to the disease resistance of winter
oilseed rape to the pathogen. J. Phytopathol. 143:613–617.
Lin, J. J., Kuo, J., Ma, J., Saunders, J. A., Beard, H. S., McDonald, M. H., Kenworthy, W.,
Ude, G. N., Matthews, F. (1996). Identification of molecular markers in soybean
comparing RFLP, RAPD and AFLP DNA mapping techniques. Plant Mol. Biol. Rep.
14:156–169.
Logan, C. (1974). The effect of soil- and tuber-borne inoculum on the incidence of potato
gangrene. Ann. Appl. Biol. 78:251–259.
McDonald, B. A. (1997). The population genetics of fungi: tools and techniques.
Phytopathology 87:448–453.
Michelmore, R. W., Ilott, T., Hulbert, S. H., Farrara, B. (1988). The downy mildews. Adv.
Plant Pathol. 6:53–79.
Mizubuti, E. S. G., Fry, W. E. (1998). Temperature effects on developmental stages of
isolates from three clonal lineages of Phytophthora infestans. Phytopathology
88:837–843.
Mogling, R., Broschewitz, B. (1990). Proposal for the scoring of severity of selected
diseases in fodder peas. Nachrbl. Pflanzenschutzd. DDR 44:115–116.
Mujica, A., Jacobsen, S. E., Izquierdo, J., Marathee, J. P. (1998). Libro de campo. Prueba
americana y europea de quinua. Proyecto Quinua—CIP DANIDA—UNAP. Puno,
Peru: Universidad Nacional del Altiplano, p. 41.
Nashaat, N. I., Awasthi, R. P. (1995). Evidence for differential resistance to Peronospora
parasitica (downy mildew) in accessions of Brassica juncea (mustard) at the
cotyledon stage. J. Phytopathol. 143:157–159.
Natti, J. J. (1958). Resistance of broccoli and other crucifers to downy mildew. Plant Dis.
Rep. 42:656–662.
Danielsen, Bonifacio, and Ames58
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.

Dow
nloa
ded
By:
[Dan
mar
ks V
eter
naer
Og]
At:
22:5
8 25
Mar
ch 2
008
Nutter, F. W. (1989). Detection and measurement of plant disease gradients in peanut with
a multispectral radiometer. Phytopathology 79:958–963.
O’Brien, R. D., van Bruggen, A. H. C. (1992). Accuracy, precision, and correlation to
yield loss of disease severity scales for corky root of lettuce. Phytopathology
82:91–96.
Ochoa, J., Frinking, H. D., Jacobs, Th. (1999). Postulation of virulence groups and
resistance factors in the quinoa/downy mildew pathosystem using material from
Ecuador. Plant Pathol. 48:425–430.
Otazu, V., Salas, B. (1977). Brown stalk of Chenopodium quinoa caused by Phoma exigua
var. foveata. Fitopatologıa 12:54–58.
Otazu, V., Agular, P. C., Canahua, A. (1976). Resistencia en quinua (Chenopodium
quinoa) al mildiu (Peronospora effusa). Fitopatologıa 11:47–49.
Risi, C. J., Galwey, N. W. (1989). The pattern of genetic diversity in the Andean grain crop
quinoa (Chenopodium quinoa Willd). II. Multivariate methods. Euphytica
41:135–145.
Shao, X. H., Chen, Q. Q., Zhang, F. Q. (1990). Research on relation between some
ecological factors and Peronospora parasitica (Pers.) Fr. on Chinese cabbage plants.
Acta Agric. Shanghai 6:78–81.
Sousa, M. E., Dias, J. S., Monteiro, A. A. (1997). Screening Portuguese cole landraces for
resistance to seven indigenous downy mildew isolates. Sci. Hortic. 68:49–58.
Stegmark, R. (1991). Selection for partial resistance to downy mildew in peas by means of
greenhouse tests. Euphytica 53:87–95.
Stonehouse, J. (1994). Assessment of Andean bean diseases using visual keys. Plant
Pathol. 43:519–527.
Taylor, P. N., Lewis, B. G., Matthews, P. (1989). Pathotypes of Peronospora viciae in
Britain. J. Phytopathol. 127:100–106.
Tewari, J. P., Boyetchko, S. M. (1990). Occurrence of Peronospora farinosa f.sp.
chenopodii on quinoa in Canada. Can. Plant Dis. Surv. 70:127–128.
Tham, F. Y., Lucas, J. A., Wilson, Z. A. (1994). DNA fingerprinting of Peronospora
parasitica, a biotrophic fungal pathogen of crucifers. Theor. Appl. Genet.
88:490–496.
Vulsteke, G., Callewaert, D., Meeus, P., Bosman, P. (1997). Control by seedcoating of
primary infection of downy mildew, Peronospora viciae Berk. (de Bary) on peas.
Parasitica 53:15–23.
Zimmer, R. C., McKeen, W. E., Campbell, C. G. (1992). Location of oospores in
buckwheat seed and probable roles of oospores and conidia of Peronospora ducometi
in the disease cycle on buckwheat. J. Phytopathol. 135:217–223.
Diseases of Quinoa 59
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
©2003 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.




















