
Discriminative behavioral assessment unveils remarkable reactive astrocytosis and early molecular...
9
Discriminative behavioral assessment unveils remarkable reactive astrocytosis and early molecular correlates in basal ganglia of 3-nitropropionic acid subchronic treated rats Giovanni Cirillo a,1 , Nicola Maggio a,c,1 , Maria Rosaria Bianco a , Cecilia Vollono a , Stefania Sellitti a , Michele Papa a,b, * a Laboratorio di Morfologia delle Reti Neuronali Dipartimento di Medicina Pubblica Clinica e Preventiva - Seconda Universita ` di Napoli, 80138 Napoli, Italy b Facolta ` di Medicina, Universita ` di Salerno, Israel c Weizmann Institute of Science, Rehovot 76100, Israel 1. Introduction The role of non-neuronal cells to the pathogenesis of neurodegenerative diseases has been widely investigated yet it remains poorly understood. Several studies reported that mod- ifications of neuro-glial network strongly contribute to several mental disorders (Musholt et al., 2009) and neurodegenerative diseases (Lobsiger and Cleveland, 2007) such as Alzheimer’s disease (Rodriguez et al., 2009), amyotrophic lateral sclerosis (Sheldon and Robinson, 2007), Parkinson’s disease (Saijo et al., 2009), and spinocerebellar ataxia type 1 (SCA1) (Giovannoni et al., 2007). Particularly, in an experimental model of SCA1, glial activation, characterized by astrocytic hypertrophy and increased production of glial fibrillary acidic protein (GFAP), correlates with changes of glial glutamate transporter expression (Giovannoni et al., 2007). Glial and microglial activation, through changes of cell phenotype and gene expression, might trigger glial-induced neuronal death (Bal-Price and Brown, 2001), having a key role to the pathophysiology of neurodegenerative disorders (Mrak and Griffin, 2005). Excitatory amino acids transporter 1 (EAAT1, also known as GLAST) is the major glutamate transporter expressed in the brain (Lehre and Danbolt, 1998). It is not detected in neurons (Ginsberg et al., 1995) but strongly expressed on astrocytes (Chaudhry et al., 1995). Its distribution correlates with the morphological changes reported in the astrocytes during inflammatory (Cavaliere et al., 2007; Xin et al., 2009; Tilleux et al., 2009) or degenerative processes (Dervan et al., 2004) and subsequent changes in neurotransmitter Neurochemistry International 56 (2010) 152–160 ARTICLE INFO Article history: Received 17 June 2009 Received in revised form 21 August 2009 Accepted 18 September 2009 Available online 30 September 2009 Keywords: 3-Nitropropionic acid Reactive astrocytosis Glial glutamate transporter GLAST Striatum ABSTRACT Reactive astrocytosis seems to be strongly implicated in the development and maintenance of inflammatory and neurodegenerative disorders. We design a new toxic model treatment with 3- nitropropionic acid (3-NP), a mitochondrial complex II irreversible inhibitor, to induce in rats Huntington’s disease (HD) like syndrome, characterized by hindlimb dystonia, involuntary choreiform movements and reduced global activity. In an attempt to find out whether molecular and morphological changes in the neuro-glial network could be involved in the pathogenesis of this disease, we developed a protocol of subchronic intra-peritoneal 3-NP intoxication. Moreover we set up specific, highly discriminative, behavioral tests to detect very early mild motor disabilities in 3-NP treated rats. This treatment did not cause severe cell death. However, in the Caudate-Putamen (CPu) of all 3-NP treated animals we found a massive astrogliosis, revealed by increased GFAP levels, paralleled by changes of the glial glutamate transporter GLAST distribution. To these glial changes we detected a transcriptional upregulation of c-fos and Sub-P in the striatal medium spiny neurons (MSN). We propose that this model of 3-NP intoxication along with the designed set of behavioral analyses allow to unmask in a very early phase the motor deficits and the underlying morpho-molecular changes associated to the onset of motor disabilities in the HD-like syndrome. Therefore this model unveil the key role played by the different components of the tripartite synapse in the pathogenesis of the HD, a putative non-cell-autonomous disease. ß 2009 Elsevier Ltd. All rights reserved. Abbreviations: 3-NP, 3-nitropropionic acid; i.p., intra-peritoneal; GFAP, glial fibrillary acidic protein; MSN, medium spiny neurons. * Corresponding author at: Department of Medicina Pubblica Clinica e Preventiva, Institute of Human Anatomy, Second University of Naples, via l. Armanni 5, 80100 Napoli, Italy. Tel.: +39 081 296636; fax: +39 081 296636. E-mail address: [email protected] (M. Papa). 1 These authors equally contributed to this work. Contents lists available at ScienceDirect Neurochemistry International journal homepage: www.elsevier.com/locate/neuint 0197-0186/$ – see front matter ß 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.neuint.2009.09.013
Transcript of Discriminative behavioral assessment unveils remarkable reactive astrocytosis and early molecular...

Discriminative behavioral assessment unveils remarkable reactive astrocytosisand early molecular correlates in basal ganglia of 3-nitropropionic acidsubchronic treated rats
Giovanni Cirillo a,1, Nicola Maggio a,c,1, Maria Rosaria Bianco a, Cecilia Vollono a,Stefania Sellitti a, Michele Papa a,b,*a Laboratorio di Morfologia delle Reti Neuronali Dipartimento di Medicina Pubblica Clinica e Preventiva - Seconda Universita di Napoli, 80138 Napoli, Italyb Facolta di Medicina, Universita di Salerno, Israelc Weizmann Institute of Science, Rehovot 76100, Israel
Neurochemistry International 56 (2010) 152–160
A R T I C L E I N F O
Article history:
Received 17 June 2009
Received in revised form 21 August 2009
Accepted 18 September 2009
Available online 30 September 2009
Keywords:
3-Nitropropionic acid
Reactive astrocytosis
Glial glutamate transporter GLAST
Striatum
A B S T R A C T
Reactive astrocytosis seems to be strongly implicated in the development and maintenance of
inflammatory and neurodegenerative disorders. We design a new toxic model treatment with 3-
nitropropionic acid (3-NP), a mitochondrial complex II irreversible inhibitor, to induce in rats
Huntington’s disease (HD) like syndrome, characterized by hindlimb dystonia, involuntary choreiform
movements and reduced global activity. In an attempt to find out whether molecular and morphological
changes in the neuro-glial network could be involved in the pathogenesis of this disease, we developed a
protocol of subchronic intra-peritoneal 3-NP intoxication. Moreover we set up specific, highly
discriminative, behavioral tests to detect very early mild motor disabilities in 3-NP treated rats. This
treatment did not cause severe cell death. However, in the Caudate-Putamen (CPu) of all 3-NP treated
animals we found a massive astrogliosis, revealed by increased GFAP levels, paralleled by changes of the
glial glutamate transporter GLAST distribution. To these glial changes we detected a transcriptional
upregulation of c-fos and Sub-P in the striatal medium spiny neurons (MSN). We propose that this model
of 3-NP intoxication along with the designed set of behavioral analyses allow to unmask in a very early
phase the motor deficits and the underlying morpho-molecular changes associated to the onset of motor
disabilities in the HD-like syndrome. Therefore this model unveil the key role played by the different
components of the tripartite synapse in the pathogenesis of the HD, a putative non-cell-autonomous
disease.
Contents lists available at ScienceDirect
Neurochemistry International
journa l homepage: www.e lsev ier .com/ locate /neuint
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
The role of non-neuronal cells to the pathogenesis ofneurodegenerative diseases has been widely investigated yet itremains poorly understood. Several studies reported that mod-ifications of neuro-glial network strongly contribute to severalmental disorders (Musholt et al., 2009) and neurodegenerativediseases (Lobsiger and Cleveland, 2007) such as Alzheimer’sdisease (Rodriguez et al., 2009), amyotrophic lateral sclerosis(Sheldon and Robinson, 2007), Parkinson’s disease (Saijo et al.,
Abbreviations: 3-NP, 3-nitropropionic acid; i.p., intra-peritoneal; GFAP, glial
fibrillary acidic protein; MSN, medium spiny neurons.
* Corresponding author at: Department of Medicina Pubblica Clinica e
Preventiva, Institute of Human Anatomy, Second University of Naples, via l.
Armanni 5, 80100 Napoli, Italy. Tel.: +39 081 296636; fax: +39 081 296636.
E-mail address: [email protected] (M. Papa).1 These authors equally contributed to this work.
0197-0186/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.
doi:10.1016/j.neuint.2009.09.013
2009), and spinocerebellar ataxia type 1 (SCA1) (Giovannoni et al.,2007). Particularly, in an experimental model of SCA1, glialactivation, characterized by astrocytic hypertrophy and increasedproduction of glial fibrillary acidic protein (GFAP), correlates withchanges of glial glutamate transporter expression (Giovannoniet al., 2007). Glial and microglial activation, through changes of cellphenotype and gene expression, might trigger glial-inducedneuronal death (Bal-Price and Brown, 2001), having a key roleto the pathophysiology of neurodegenerative disorders (Mrak andGriffin, 2005).
Excitatory amino acids transporter 1 (EAAT1, also known asGLAST) is the major glutamate transporter expressed in the brain(Lehre and Danbolt, 1998). It is not detected in neurons (Ginsberget al., 1995) but strongly expressed on astrocytes (Chaudhry et al.,1995). Its distribution correlates with the morphological changesreported in the astrocytes during inflammatory (Cavaliere et al.,2007; Xin et al., 2009; Tilleux et al., 2009) or degenerative processes(Dervan et al., 2004) and subsequent changes in neurotransmitter

G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160 153
uptake could represent the basis of morphological and functionalchanges underlying Huntington’s disease (HD). Clinical trials inpatients with HD (Seppi et al., 2001) and animal models of striatalneurodegeneration (Doble, 1999) designed to slow progression bydecreasing glutamatergic neurotransmission have used riluzole asattenuating agent of glutamate release.
Astrocytic activation in striatum is a common reaction aftertoxic, ischemic or degenerative process and seems to play a role inthe development of HD (Hickey et al., 2008). Huntington’s disease,a neurodegenerative autosomic dominant inherited disorder,characterized, in humans, by involuntary choreiform movementsand cognitive impairment (Landles and Bates, 2004) caused by aCAG expansion in the coding region of the gene huntingtin (1993),is characterized by marked striatal atrophy with neuronal loss,astrocytic activation and intra-neuronal protein aggregates(Aparicio and Lucas, 2002; Paulson and Fischbeck, 1996). Themechanism underlying the selective neuronal death of theGABAergic medium spiny neurons (MSN) in the striatum stillremains unknown. One hypothesis suggests that the genetic defectmay cause an impairment of energy metabolism that subsequentlymakes these neurons more vulnerable to slow excitotoxicdegeneration (Brouillet et al., 2005).
3-Nitropropionic acid (3-NP) is a suicide inhibitor of succinatedehydrogenase and has been shown to cause a HD-like syndromein primates (Brouillet et al., 2005) but also in rats (Borlongan et al.,1997a). I.p. 3-NP acute (60–100 mg/kg/die) or chronic treatment(2–5 mg/kg for 7 or 14 or more days) induced striatal lesions thatresulted in dystonia and other abnormal motor behaviorsmimicking the neurochemical and behavioral features of HD inhumans (Bogdanov et al., 1998; Fernagut et al., 2002). Bothintoxication protocols have established a link between mitochon-drial chain impairment and HD pathogenesis (Browne, 2008). 3-NPacute toxicity induces severe neurological deficits and produces aneuron depleted striatal lesion (Borlongan et al., 1997b) whilechronic intoxication has been reported to induce behavioral andneurochemical features (Borlongan et al., 1997b; Palfi et al., 1996)similar to those found in grade 1–2 HD patients (Chyi and Chang,1999). Unfortunately, both treatments failed to reveal the earlymolecular and morphological events that lead to the death of theMSN as, in both models, 3-NP treatment produces massiveneuronal death, a phenomenon that occurs only in late stage ofthe human disease (Paulsen et al., 2006).
To identify and better clarify the early modifications of theneurodegenerative process, we developed a new 3-NP treatmentprotocol which could mimic the phenotype of the humanHuntington’s disease in rats. This i.p. subchronic administrationinduced mild behavioral abnormalities depending on mitochon-drial chain impairment. We used a new designed behavioral test(progressive elevated beam test) and other well-accepted motorskill test to unmask the early motor deficits induced by 3-NPtreatment. In treated animals, we found an intense reactiveastrocytosis with no clear sign of neuronal death along with analtered expression of the glial glutamate transporter GLAST.Neurons, however, showed an increased expression of Sub-stance-P (Sub-P) and c-fos. We hypothesize that these molecularchanges could determine the functional as structural rearrange-ment of the tripartite synapse following mitochondrial insult, thatis the very early events leading to synaptic dysfunction and,possibly, in later stages, neuronal death.
2. Experimental procedures
2.1. Animals
Adult (250–300 g; Charles River, Calco, Italy) male Sprague Dawley rats (n = 20),
3 months old at the beginning of the intoxication, were used. Rats were maintained
on a 12/12 h light/dark cycle and allowed free access to food and water. Each animal
was housed under specific pathogen-free conditions in iron-sheet cages with solid
floors covered with 4–6 cm of sawdust during the experiments. All experimental
procedures were performed during the light cycle and were approved by the Animal
Ethics Committee of The Second University of Naples. Animal care was in
compliance with Italian (D.L. 116/92) and EC (O.J. of E.C. L358/1 18/12/86)
regulations on the care of laboratory animals. All efforts were made to reduce
animal numbers.
2.2. Treatment
3-Nitropropionic acid (3-NP, Sigma, Italy) was prepared as previously described
(Brouillet et al., 1995; Fernagut et al., 2002). In brief, 3-NP was dissolved in a few
drops of distilled water and the pH was adjusted to 7.4 with 1 M NaOH and the final
volume adjusted with 0.1 M phosphate-buffered saline (PBS), pH 7.4. Rats (n = 20)
were grouped as follows: animals undergoing subchronic i.p. 3-NP treatment
(n = 12) and vehicle (saline solution) treated rats (n = 8) assumed as control (CTR)
animals. We used an injection protocol characterized by three different decreasing
3-NP i.p. doses, in order to maintain a low 3-NP level for all the period of treatment.
In particular, the subchronic treatment consisted of twice a day i.p. injections of 3-
NP for 18 days. Twice a day injections were divided as follows: 7.5 mg/kg for the
first 2 days of treatment, followed by 3.75 mg/kg for 7 days, finally a dosage of 2 mg/
kg for the last 8 days of treatment. Animals were sacrificed 4 days after treatment.
2.3. Behavioral analysis
Progressive elevated beam test: This newly designed behavioral analysis was used
to evaluate the appearance of motor performance abnormalities, in the treated
animals vs. the controls, by progressively increasing the difficulty in the execution
of the task. The aim was to train the animals in crossing three beams of different
sizes to reach a platform on which their home cage was placed. Specifically, the
beams had a similar length (100 cm) but different width (8, 5 and 3 cm,
respectively). Before treatment, animals received 3 days of training (twice per
day), after which all rats were able to cross the board without rearing or stopping.
Animals were videotaped and analysis was tracked off-line. Each rat was scored for
time of crossing and numbers of foot slips for each beam. Occurrence of
bradykinesia was quantified by calculating the average velocity of walking along
each different bar for treated and CTR animals.
Footprint test: This test was used to assess gait abnormalities in 3-NP treated vs.
control animals. After coating the hind feet with a non-toxic black dye and forepaws
with a non-toxic blue dye, rats were allowed to walk on a beam (100 cm long, 8 cm
width) covered with paper. Step length, paws overlap and gait walking angle were
analyzed. Specifically, step length was quantified as the distance between two
subsequent feet on the y-axis. Paws overlap analysis was carried out by measuring
the centre distance between the anterior paw and the correlated rear paw
footprints. Gait angle walking was calculated by measuring the angle of the foot in
respect to the walking direction.
2.4. Tissue preparation
Rats were deeply anesthetized with an i.p. injection (300 mg/kg body weight) of
chloral hydrate and perfused transcardially with saline solution (TRIS HCl 0.1 M,
EDTA 10 mM) followed by 4% paraformaldehyde added to 0.1% glutaraldehyde in
0.01 M phosphate-buffer (PB), pH 7.4 at 4 8C. For light microscopy brains were
removed and post-fixed 2 h in the same fixative, then soaked in 30% sucrose PBS and
frozen in chilled isopentane on dry ice. Serial sections were cut at the cryo-sliding
vibratome at a thickness of 30 mm and collected in cold PBS for immunohis-
tochemistry.
2.5. Immunohistochemistry
Brain sections were blocked in 10% normal serum in 0.01 M phosphate-buffer
saline (PBS), 0.25% Triton for 1 h at room temperature. Each primary antibody was
diluted in 0.01 M PBS containing 10% normal serum 0.25% Triton. We used rabbit
anti Sub-P antiserum (1:5000) and guinea pig anti-GLAST antiserum (1:4000)
(Chemicon International, Temecula, CA, USA), rabbit anti c-fos antiserum (1:1000)
(Santa Cruz Biotechnology Inc., Santa Cruz, CA), mouse monoclonal anti glial
fibrillary acidic protein (1:400) (SIGMA, Saint Louis, MO, USA) and rabbit anti
DARPP-32 antiserum (1:100) (Cell Signaling Technology, USA). Sections were
incubated for 48 h at 4 8C. Sections were washed in PBS and incubated with the
appropriate biotinylated secondary antibody (Vector Labs, Inc., Burlingame, CA,
USA; 1:200) for 90 min at room temperature (RT). Next, they were processed using
the Vectastain avidin–biotin peroxidase kit (Vector Labs, Inc., Burlingame, CA, USA)
for 90 min, at RT. Sections were washed in 0.05 M Tris–HCl, reacted with 3,3-
diaminobenzidine tetrahydrochloride (DAB; Sigma, 0.5 mg/ml Tris–HCl) and 0.01%
hydrogen peroxide, then mounted on chrome-alume gelatin coated slides,
dehydrated, and coverslipped. Adjacent sections were Nissl-stained.
To analyze the spatial distribution of glial glutamate transporter GLAST, double
staining was performed as previously reported (Papa et al., 2003). In brief, sections
were incubated with either anti-GLAST and anti-GFAP primary antibodies following
2 h incubation with a solution containing either Alexa fluor 488 antigoat IgG (1:200)
and Alexa fluor 555 anti-mouse IgG (1:200; Molecular Probes). Sections were

G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160154
mounted and coverslipped with Vectashield (Vector Labs). Images were captured at
a resolution of 512 � 512 pixels. The appropriate argon laser fluorescence for
visualization of Alexa Fluor 488 was used with an excitation wavelength of 488 nm
and emission filter BP 505–530. HeNe laser fluorescence for the visualization of
Alexa Fluor 555 used an excitation wavelength of 543 nm and emission filter LP 560.
A sequence (n = 68) of single sections was acquired (0.23 mm apart).
2.6. Fluoro-Jade staining procedure
Brain sections were mounted with 0.1 M neutral phosphate-buffer onto 2%
gelatin coated slides and dried on a slide warmer at 50 8C for at least half an hour.
Sections were first immersed in a solution containing 1% NaOH in 80% alcohol for
5 min. This was followed by 2 min in 70% alcohol and 2 min in distilled water.
Afterwards, slices were transferred to a solution of 0.06% potassium permanganate
for 10 min, gently shaken on a rotating platform, then rinsed for 2 min in distilled
water, transferred to a 0.0004% Fluoro-Jade (Histo-Chem Inc., Jefferson, AR, USA)
staining solution, gently agitated for 20 min. Fluoro-Jade staining solution was
prepared as described elsewhere (Schmued and Hopkins, 2000a,b). After staining,
sections were rinsed with three 1 min changes of distilled water. Then, slides were
placed on a slide warmer, set at approximately 50 8C, until they were fully dry (5–
10 min). The dry slides were cleared by immersion in xylene for 1 min, then
coverslipped with a non-aqueous non-fluorescent plastic mounting media.
The tissue was examined using an epifluorescent microscope with blue (450–
490 nm) excitation light.
2.7. Measurements and statistical analysis
Behavior data were plotted as means � SEM and statistical significance was
calculated by t-Student’s test for unpaired data.
Slides were imaged with a Zeiss Axioskope 2 light microscope equipped with
high-resolution digital camera (C4742-95; Hamamatsu Photonics, Milan, Italy).
Analysis of the markers in striatum was accomplished by using computer assisted
image analysis system (MCID 7.0; Imaging Research, St. Catharines, Ontario,
Canada). All markers were analyzed by a morphometric approach because of the
perfect visualization of single positive elements. Therefore, the values were
expressed as a proportional area: number of positive elements relative to the
scanned area. The averages were obtained from five randomly selected brain
sections for each animal, and comparisons were made between treatment (3-NP)
and control group (CTR). Data were exported and converted to frequency
distribution histograms by using the Sigma-Plot 10.0 program with SigmaStat
3.5 integration (SPSS Erkrath, Germany). Data from all the quantitative analyses
were analyzed by one-way and two-way ANOVA, using all pairwise Holm-Sidak
method for multiple comparisons (p � 0.001). All data shown are presented as the
mean � SEM.
3. Results
3.1. General observations and behavioral tests
All 3-NP treated animals survived the treatment. No significantweight loss was reported: at the end of the subchronic 3-NPtreatment, CTR and treated rats weighted 355 � 8 and 324 � 19 g,respectively and did not significantly differ from the weight beforetreatment (295 � 5 and 312 � 12 g, respectively) (p � 0.05).
According to the studies of Fernagut (Fernagut et al., 2002), weevaluated in 3-NP treated rats the occurrence of these motorsymptoms: hindlimb clasping, reduced global activity (velocity,exploratory behavior, rearing and grooming) in a freely movingenvironment (open field test), hindlimb dystonia (crouchingposture, gait impairment), truncal dystonia (kyphotic posture)and balance adjustments to a postural challenge (rotating move-ments grasping rat tails). Apparently, the subchronic treated ratsdid not show any evident sign of these neurological deficits.
Two possibilities could explain this observation. On onehand, the absence of neurological impairments could have beendue to the weakness of the treatment which was unable toproduce any disease. On the other hand, it could have been thatneurological abnormalities were indeed developed, but thestandard scale used to quantify motor disabilities in rats was notsensitive enough to detect them. If this would have been thecase, one had to develop some more sensitive tests whichcould be able to detect small changes in the motor behavior ofthe animals.
To this aim, we designed a new test called the ‘‘Progressive
elevated beam test’’. The idea is based on the ability of rodents toeasily walk onto surfaces of very small width. Taken this intoaccount, we developed a new ‘‘crossing test’’: rats had to cross,once at time, three different beams whose width was progressivelydecreased. If any neurological deficit has been produced by 3-NPtreatment, treated rats had to show a progressive difficulty inaccomplishing a motor task whose complexity is increasing. Theresults of this experiment are reported in Fig. 1. The analysis of thetime to cross the first beam (Beam 1; 8 cm width) did not show anysignificant difference between control and treated rats (20 � 2.53and 20.87 � 1.78 s, respectively). Significant difference was obtainedwhen the time to cross the second beam (Beam 2; 5 cm width) wasmeasured (24.5 � 3.01 s vs. 28.87 � 1.78 s, for control vs. treated).Interestingly, the analysis of the time to cross the narrowest beam(Beam 3; 3 cm width) showed a remarkable difference between thetwo groups. In this condition, the treated animals needed 50 � 1.77 sto cross the bar compared to the 31.25 � 2.5 s of the control rats(Fig. 1A). This results indicates that the subchronic 3-NP protocol waseffective in causing motor impairments in treated animals.
As the animals were trained to execute the motor task withoutstopping before the beginning of the treatment, only a difference inthe velocity to cross beams and in the probability of foot slips couldexplain the difference between the two groups.
To check this possibility, we measured the average velocity ofcrossing (Fig. 1B) and the numbers of paw slips (Fig. 1C) in everybar crossed by CTR and treated rats. The average velocity to crossBeam 1 and Beam 2 was not statistically significant between thetwo groups (for Beam 1: 5.41 � 0.27 cm/s vs. 4.85 � 0.48 cm/s forcontrol vs. treated; for Beam 2: 4.58 � 0.58 cm/s vs. 4.34 � 0.56 cm/s, respectively). When crossing Beam 3, treated animals showed asignificant difference in average velocity compared to control rats(2.19 � 0.26 cm/s vs. 3.92 � 0.36 cm/s, respectively) (Fig. 1B).
Fig. 1C shows the analysis of the number of paw slips for eachbar. As it can be noted, the number of slips progressively increasesrelating to the increased difficulty of the motor task execution.When crossing Beam 1, the average of paw slips for treated animals(5.6 � 0.78) did not significantly differs from the CTR value(4.9 � 0.85). Significant differences were measured crossing Beam2 (for treated 8.2 � 0.98, for control 5.5 � 0.65) but more significantlycrossing Beam 3: in 3-NP treated rats, the number of foot slips was12.5 � 0.82 compared to CTR value, 6.5 � 0.65 (**p � 0.001,*p � 0.01).
Occurrence of paw slips could be interpreted as a sign ofabnormal gait. To check whether walking disabilities occurred intreated rats, a footprint test was performed.
The measurements of the step length provide evidence ofsignificant differences between the two groups (treated:3.1 � 0.32, control: 5.3 � 0.37) (Fig. 2A). An additional measurementof walking pattern is the paws overlap. The rational of this analysislies on the fact that rodents while walking usually tend to step theirhind paw in the same place previously occupied by their front paw.Quantifying this aspect of the motor behavior could then beconsidered a measure of locomotion ability. This analysis is showedin Fig. 2B: treated animals had a minor ability in overlapping theirpaws (64 � 6.7%) compared to control animals. A potential reason forthis anomaly could be that the gait’s angle in respect of the walkingdirection was altered in the treated animals. Therefore, the differenceof the gait’s angle with respect to the walking direction wasquantified in treated and control animals (Fig. 2C). This analysisproved that in treated animals, during walking, the angle formed bythe direction of the hind paw in respect to the walking track was2.1 � 0.188, while in control rats was 0.92 � 0.268 (**p � 0.001,*p � 0.01).
Altogether, this evidence demonstrates that 3-NP subchronictreatment is able to cause motor behavioral abnormalities that as it

Fig. 2. Footprint test. The foot print test revealed abnormal gait in the treated animals. Intoxicated animals had a deficit in step length (A), a minor degree of paws overlap (B)
and a different walking angle (C) compared to CTR rats (**p � 0.001; *p � 0.01).
Fig. 1. Progressive beam test analysis. A task of progressive difficulty unmasks behavioral disabilities in treated animals. Walking on beams of different width (8, 5, 3 cm,
respectively) detected an impairment of the performance in the intoxicated animals compared to control in respect to: time to cross the beam (A), average velocity to cross the
beam (B) and number of slips (C). It is important to note that the performance of the treated animals became worse as the difficulty of the task increased (**p � 0.001;
*p � 0.01).
G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160 155
has been already reported (Guyot et al., 1997; Teunissen et al.,2001), could most likely be due to extrapiramidal dysfunctions.
3.2. 3-NP subchronic treatment does not cause neuronal loss
It has been previously reported that 3-NP treatment causesneuronal cell death (Galas et al., 2004; Garcia et al., 2002; Wullneret al., 1994). To assess if the behavioral findings reported abovewere associated with neuronal death, we stain brain sections oftreated and control animals with Fluoro-Jade B (FJ-B). FJ-B is afluorescent dye that has been described to specifically stain alldegenerating neurons, regardless of specific insult or mechanismof cell death (Schmued et al., 2005; Schmued and Hopkins,
2000a,b). After staining, our results showed that 3-NP subchronictreatment did not produce any significant neuronal cell deathwhen compared to CTR (19.69 � 0.33 vs. 19.49 � 0.5) (Fig. 3A).
To further investigate this phenomenon, a DARPP-32 positivecell count was performed in both groups. DARPP-32 (dopamineand cyclic AMP regulated phosphoprotein) is a 32 kDa cytosolicprotein specifically expressed in striatal MSN (Reiner et al., 1998).Therefore, counting cells expressing this protein could give areliable index of the number of neurons in the striatum.Interestingly, no significant number differences of DARPP-32positive elements were detected between the two groups (forcontrol: 1988.54 � 48.01, for treated 1910.69 � 37.93) (p � 0.05)(Fig. 3B). These findings show that the subchronic protocol was
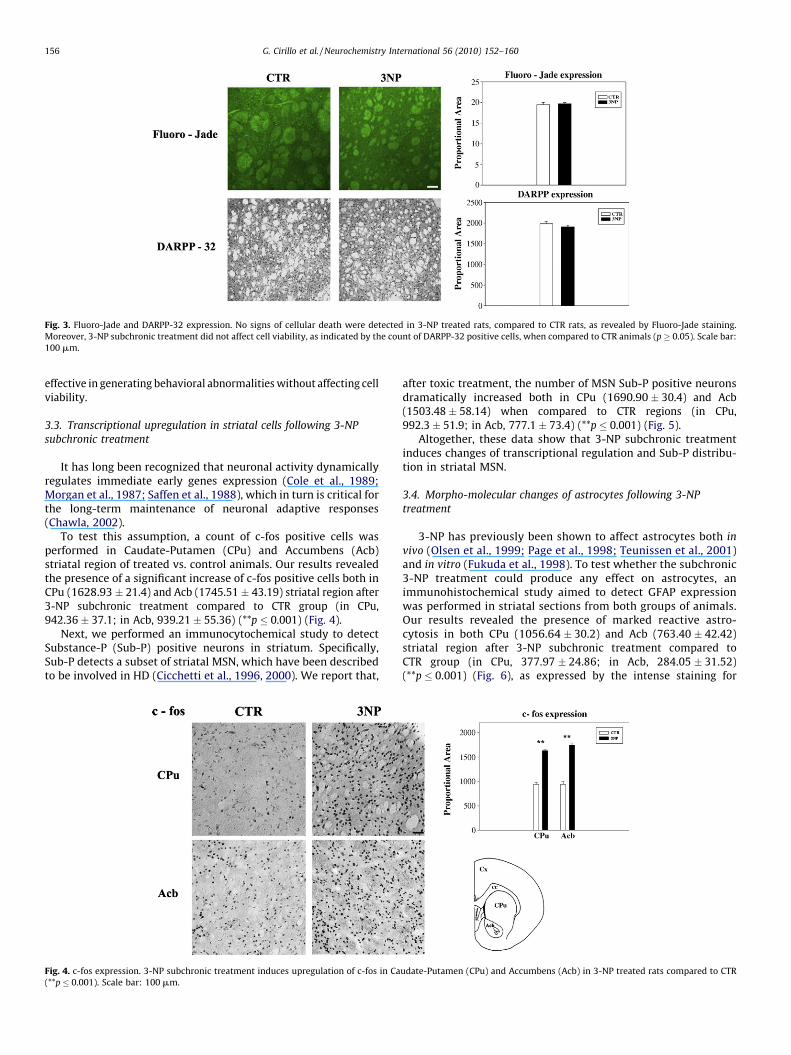
Fig. 3. Fluoro-Jade and DARPP-32 expression. No signs of cellular death were detected in 3-NP treated rats, compared to CTR rats, as revealed by Fluoro-Jade staining.
Moreover, 3-NP subchronic treatment did not affect cell viability, as indicated by the count of DARPP-32 positive cells, when compared to CTR animals (p � 0.05). Scale bar:
100 mm.
G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160156
effective in generating behavioral abnormalities without affecting cellviability.
3.3. Transcriptional upregulation in striatal cells following 3-NP
subchronic treatment
It has long been recognized that neuronal activity dynamicallyregulates immediate early genes expression (Cole et al., 1989;Morgan et al., 1987; Saffen et al., 1988), which in turn is critical forthe long-term maintenance of neuronal adaptive responses(Chawla, 2002).
To test this assumption, a count of c-fos positive cells wasperformed in Caudate-Putamen (CPu) and Accumbens (Acb)striatal region of treated vs. control animals. Our results revealedthe presence of a significant increase of c-fos positive cells both inCPu (1628.93 � 21.4) and Acb (1745.51 � 43.19) striatal region after3-NP subchronic treatment compared to CTR group (in CPu,942.36 � 37.1; in Acb, 939.21 � 55.36) (**p � 0.001) (Fig. 4).
Next, we performed an immunocytochemical study to detectSubstance-P (Sub-P) positive neurons in striatum. Specifically,Sub-P detects a subset of striatal MSN, which have been describedto be involved in HD (Cicchetti et al., 1996, 2000). We report that,
Fig. 4. c-fos expression. 3-NP subchronic treatment induces upregulation of c-fos in Ca
(**p � 0.001). Scale bar: 100 mm.
after toxic treatment, the number of MSN Sub-P positive neuronsdramatically increased both in CPu (1690.90 � 30.4) and Acb(1503.48 � 58.14) when compared to CTR regions (in CPu,992.3 � 51.9; in Acb, 777.1 � 73.4) (**p � 0.001) (Fig. 5).
Altogether, these data show that 3-NP subchronic treatmentinduces changes of transcriptional regulation and Sub-P distribu-tion in striatal MSN.
3.4. Morpho-molecular changes of astrocytes following 3-NP
treatment
3-NP has previously been shown to affect astrocytes both in
vivo (Olsen et al., 1999; Page et al., 1998; Teunissen et al., 2001)and in vitro (Fukuda et al., 1998). To test whether the subchronic3-NP treatment could produce any effect on astrocytes, animmunohistochemical study aimed to detect GFAP expressionwas performed in striatal sections from both groups of animals.Our results revealed the presence of marked reactive astro-cytosis in both CPu (1056.64 � 30.2) and Acb (763.40 � 42.42)striatal region after 3-NP subchronic treatment compared toCTR group (in CPu, 377.97 � 24.86; in Acb, 284.05 � 31.52)(**p � 0.001) (Fig. 6), as expressed by the intense staining for
udate-Putamen (CPu) and Accumbens (Acb) in 3-NP treated rats compared to CTR
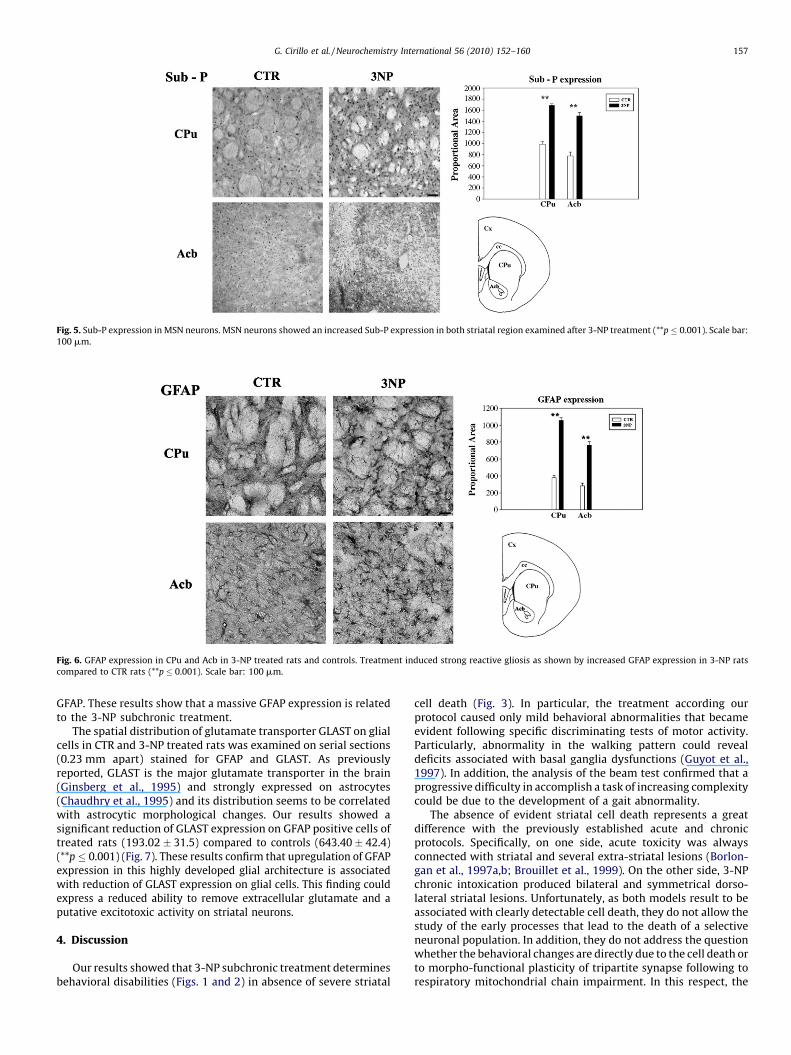
Fig. 5. Sub-P expression in MSN neurons. MSN neurons showed an increased Sub-P expression in both striatal region examined after 3-NP treatment (**p � 0.001). Scale bar:
100 mm.
Fig. 6. GFAP expression in CPu and Acb in 3-NP treated rats and controls. Treatment induced strong reactive gliosis as shown by increased GFAP expression in 3-NP rats
compared to CTR rats (**p � 0.001). Scale bar: 100 mm.
G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160 157
GFAP. These results show that a massive GFAP expression is relatedto the 3-NP subchronic treatment.
The spatial distribution of glutamate transporter GLAST on glialcells in CTR and 3-NP treated rats was examined on serial sections(0.23 mm apart) stained for GFAP and GLAST. As previouslyreported, GLAST is the major glutamate transporter in the brain(Ginsberg et al., 1995) and strongly expressed on astrocytes(Chaudhry et al., 1995) and its distribution seems to be correlatedwith astrocytic morphological changes. Our results showed asignificant reduction of GLAST expression on GFAP positive cells oftreated rats (193.02 � 31.5) compared to controls (643.40 � 42.4)(**p � 0.001) (Fig. 7). These results confirm that upregulation of GFAPexpression in this highly developed glial architecture is associatedwith reduction of GLAST expression on glial cells. This finding couldexpress a reduced ability to remove extracellular glutamate and aputative excitotoxic activity on striatal neurons.
4. Discussion
Our results showed that 3-NP subchronic treatment determinesbehavioral disabilities (Figs. 1 and 2) in absence of severe striatal
cell death (Fig. 3). In particular, the treatment according ourprotocol caused only mild behavioral abnormalities that becameevident following specific discriminating tests of motor activity.Particularly, abnormality in the walking pattern could revealdeficits associated with basal ganglia dysfunctions (Guyot et al.,1997). In addition, the analysis of the beam test confirmed that aprogressive difficulty in accomplish a task of increasing complexitycould be due to the development of a gait abnormality.
The absence of evident striatal cell death represents a greatdifference with the previously established acute and chronicprotocols. Specifically, on one side, acute toxicity was alwaysconnected with striatal and several extra-striatal lesions (Borlon-gan et al., 1997a,b; Brouillet et al., 1999). On the other side, 3-NPchronic intoxication produced bilateral and symmetrical dorso-lateral striatal lesions. Unfortunately, as both models result to beassociated with clearly detectable cell death, they do not allow thestudy of the early processes that lead to the death of a selectiveneuronal population. In addition, they do not address the questionwhether the behavioral changes are directly due to the cell death orto morpho-functional plasticity of tripartite synapse following torespiratory mitochondrial chain impairment. In this respect, the
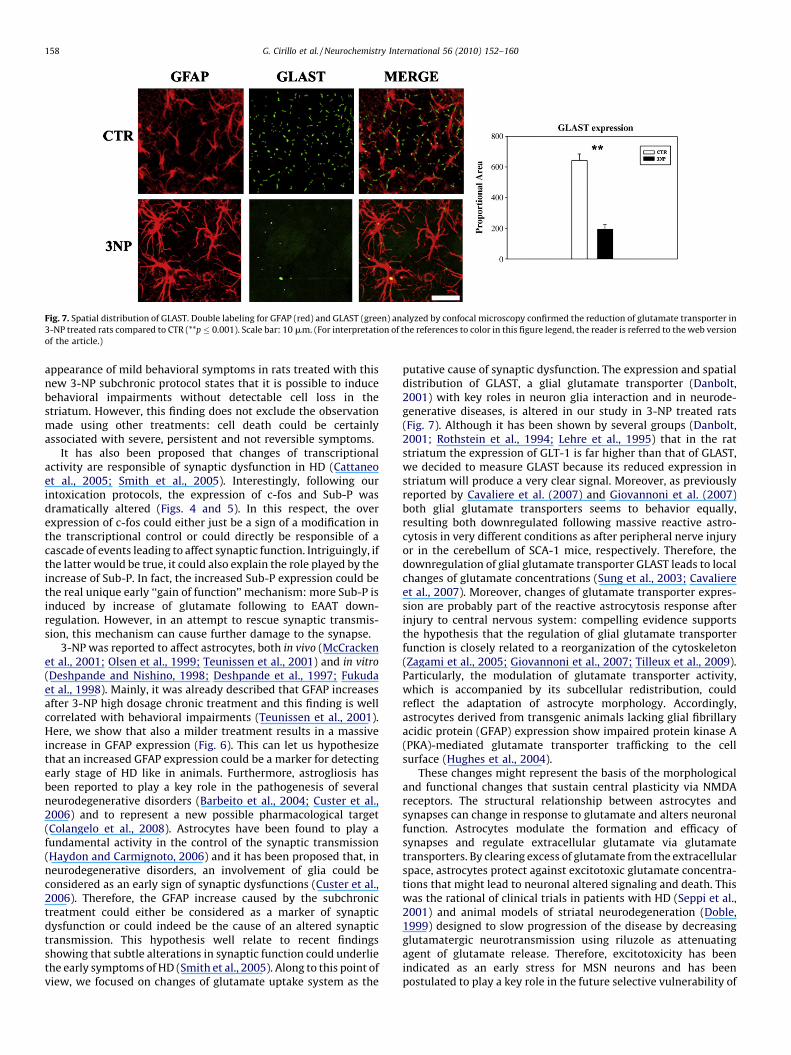
Fig. 7. Spatial distribution of GLAST. Double labeling for GFAP (red) and GLAST (green) analyzed by confocal microscopy confirmed the reduction of glutamate transporter in
3-NP treated rats compared to CTR (**p � 0.001). Scale bar: 10 mm. (For interpretation of the references to color in this figure legend, the reader is referred to the web version
of the article.)
G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160158
appearance of mild behavioral symptoms in rats treated with thisnew 3-NP subchronic protocol states that it is possible to inducebehavioral impairments without detectable cell loss in thestriatum. However, this finding does not exclude the observationmade using other treatments: cell death could be certainlyassociated with severe, persistent and not reversible symptoms.
It has also been proposed that changes of transcriptionalactivity are responsible of synaptic dysfunction in HD (Cattaneoet al., 2005; Smith et al., 2005). Interestingly, following ourintoxication protocols, the expression of c-fos and Sub-P wasdramatically altered (Figs. 4 and 5). In this respect, the overexpression of c-fos could either just be a sign of a modification inthe transcriptional control or could directly be responsible of acascade of events leading to affect synaptic function. Intriguingly, ifthe latter would be true, it could also explain the role played by theincrease of Sub-P. In fact, the increased Sub-P expression could bethe real unique early ‘‘gain of function’’ mechanism: more Sub-P isinduced by increase of glutamate following to EAAT down-regulation. However, in an attempt to rescue synaptic transmis-sion, this mechanism can cause further damage to the synapse.
3-NP was reported to affect astrocytes, both in vivo (McCrackenet al., 2001; Olsen et al., 1999; Teunissen et al., 2001) and in vitro
(Deshpande and Nishino, 1998; Deshpande et al., 1997; Fukudaet al., 1998). Mainly, it was already described that GFAP increasesafter 3-NP high dosage chronic treatment and this finding is wellcorrelated with behavioral impairments (Teunissen et al., 2001).Here, we show that also a milder treatment results in a massiveincrease in GFAP expression (Fig. 6). This can let us hypothesizethat an increased GFAP expression could be a marker for detectingearly stage of HD like in animals. Furthermore, astrogliosis hasbeen reported to play a key role in the pathogenesis of severalneurodegenerative disorders (Barbeito et al., 2004; Custer et al.,2006) and to represent a new possible pharmacological target(Colangelo et al., 2008). Astrocytes have been found to play afundamental activity in the control of the synaptic transmission(Haydon and Carmignoto, 2006) and it has been proposed that, inneurodegenerative disorders, an involvement of glia could beconsidered as an early sign of synaptic dysfunctions (Custer et al.,2006). Therefore, the GFAP increase caused by the subchronictreatment could either be considered as a marker of synapticdysfunction or could indeed be the cause of an altered synaptictransmission. This hypothesis well relate to recent findingsshowing that subtle alterations in synaptic function could underliethe early symptoms of HD (Smith et al., 2005). Along to this point ofview, we focused on changes of glutamate uptake system as the
putative cause of synaptic dysfunction. The expression and spatialdistribution of GLAST, a glial glutamate transporter (Danbolt,2001) with key roles in neuron glia interaction and in neurode-generative diseases, is altered in our study in 3-NP treated rats(Fig. 7). Although it has been shown by several groups (Danbolt,2001; Rothstein et al., 1994; Lehre et al., 1995) that in the ratstriatum the expression of GLT-1 is far higher than that of GLAST,we decided to measure GLAST because its reduced expression instriatum will produce a very clear signal. Moreover, as previouslyreported by Cavaliere et al. (2007) and Giovannoni et al. (2007)both glial glutamate transporters seems to behavior equally,resulting both downregulated following massive reactive astro-cytosis in very different conditions as after peripheral nerve injuryor in the cerebellum of SCA-1 mice, respectively. Therefore, thedownregulation of glial glutamate transporter GLAST leads to localchanges of glutamate concentrations (Sung et al., 2003; Cavaliereet al., 2007). Moreover, changes of glutamate transporter expres-sion are probably part of the reactive astrocytosis response afterinjury to central nervous system: compelling evidence supportsthe hypothesis that the regulation of glial glutamate transporterfunction is closely related to a reorganization of the cytoskeleton(Zagami et al., 2005; Giovannoni et al., 2007; Tilleux et al., 2009).Particularly, the modulation of glutamate transporter activity,which is accompanied by its subcellular redistribution, couldreflect the adaptation of astrocyte morphology. Accordingly,astrocytes derived from transgenic animals lacking glial fibrillaryacidic protein (GFAP) expression show impaired protein kinase A(PKA)-mediated glutamate transporter trafficking to the cellsurface (Hughes et al., 2004).
These changes might represent the basis of the morphologicaland functional changes that sustain central plasticity via NMDAreceptors. The structural relationship between astrocytes andsynapses can change in response to glutamate and alters neuronalfunction. Astrocytes modulate the formation and efficacy ofsynapses and regulate extracellular glutamate via glutamatetransporters. By clearing excess of glutamate from the extracellularspace, astrocytes protect against excitotoxic glutamate concentra-tions that might lead to neuronal altered signaling and death. Thiswas the rational of clinical trials in patients with HD (Seppi et al.,2001) and animal models of striatal neurodegeneration (Doble,1999) designed to slow progression of the disease by decreasingglutamatergic neurotransmission using riluzole as attenuatingagent of glutamate release. Therefore, excitotoxicity has beenindicated as an early stress for MSN neurons and has beenpostulated to play a key role in the future selective vulnerability of

G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160 159
striatal neurons in HD. Recently, in a mouse transgenic model ofHD (Graham et al., 2009), an enhanced sensitivity to NMDA hasbeen demonstrated before obvious phenotypic changes withparalleled increase of NMDA receptor expression and function.Moreover in this model, it has been possible for the first time toparallel the downregulation of glutamate transporter to theincrease of MSN neurotransmitter Sub-P, that could be theputative ‘‘gain of function’’ in striatal circuitry, at the very earlyphase in HD.
Taken together, our results support the hypothesis that theonset and progression of motor deficits is related to the reactiveastrocytosis and paralleled by changes of glial glutamatetransporter GLAST expression, altering the glutamate uptakesystem and exposing neurons to glutamate excitotoxicity. More-over, we detected increased transcriptional activity, as shown byincreased c-fos expression, in absence of detectable cellular death.In this regard, neuronal degeneration might be the ultimate step ofthe neuropathology, when rescue from the disease is no longerpossible. These could represent the early events leading toneuronal death, an event that occurs only in the late phase ofneurodegenerative diseases. The subchronic 3-NP treatment alongwith the progressive elevated beam test will allow to studyexhaustively changes of protein and gene expression, and whichmechanisms underlie these changes. To understand the causalrelationship of 3-NP-induced neuronal, glial and glutamatetransporter changes, it will be of crucial importance to focus onthe role played by gliosis performing experiments aimed to reducethe aberrant glial activity.
We believe this model to be helpful to address further questionsabout early mechanisms involved in the pathogenesis of HD and tofind new targeted therapeutical approaches to treat the disease inits early stages. Recently, Colangelo et al. (2008), have reported anovel, crucial role of intrathecal administration of nerve growthfactor as a strong modulator of plasticity in the neuronal–glialnetwork by reducing reactive astrocytosis and restoring glialglutamate transporters expression. Therefore, our model mightalso provide a new approach for testing glial-specific drugs that aredesigned to reduce CNS inflammatory or degenerative process inthe hope of curing this devastating disease.
Statement of interest
The authors declare that the research was conducted in theabsence of any commercial or financial relationships that shouldbe construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from Regione Campania(L.R. N.5 Bando 2003 to M.P.), the Italian Minister of Research andUniversity (PRIN 2007 to M.P.), Regione Campania (Prog. Spec art12 E.F. 2000 to M.P.), and the CNR (Neurobiotecnologie 2003 toM.P.). G.C. and M.R.B, are fellow of the Fondazione Rita Levi-Montalcini.
References
Bal-Price, A., Brown, G.C., 2001. Inflammatory neurodegeneration mediated bynitric oxide from activated glia-inhibiting neuronal respiration, causing gluta-mate release and excitotoxicity. J. Neurosci. 21, 6480–6491.
Barbeito, L.H., Pehar, M., Cassina, P., Vargas, M.R., Peluffo, H., Viera, L., Estevez, A.G.,Beckman, J.S., 2004. A role for astrocytes in motor neuron loss in amyotrophiclateral sclerosis. Brain Res. Rev. 47, 263–274.
Bogdanov, M.B., Ferrante, R.J., Kuemmerle, S., Klivenyi, P., Beal, M.F., 1998. Increasedvulnerability to 3-nitropropionic acid in an animal model of Huntington’sdisease. J. Neurochem. 71, 2642–2644.
Borlongan, C.V., Koutouzis, T.K., Sanberg, P.R., 1997a. 3-Nitropropionic acid animalmodel and Huntington’s disease. Neurosci. Biobehav. Rev. 21, 289–293.
Borlongan, C.V., Nishino, H., Sanberg, P.R., 1997b. Systemic, but not intraparench-ymal, administration of 3-nitropropionic acid mimics the neuropathology ofHuntington’s disease: a speculative explanation. Neurosci. Res. 28, 185–189.
Brouillet, E., Conde, F., Beal, M.F., Hantraye, P., 1999. Replicating Huntington’sdisease phenotype in experimental animals. Prog. Neurobiol. 59, 427–468.
Brouillet, E., Hantraye, P., Ferrante, R.J., Dolan, R., Leroy-Willig, A., Kowall, N.W., Beal,M.F., 1995. Chronic mitochondrial energy impairment produces selective stria-tal degeneration and abnormal choreiform movements in primates. Proc. Natl.Acad. Sci. U.S.A. 92, 7105–7109.
Brouillet, E., Jacquard, C., Bizat, N., Blum, D., 2005. 3-Nitropropionic acid: a mito-chondrial toxin to uncover physiopathological mechanisms underlying striataldegeneration in Huntington’s disease. J. Neurochem. 95, 1521–1540.
Browne, S.E., 2008. Mitochondria and Huntington’s disease pathogenesis: insightfrom genetic and chemical models. Ann. N. Y. Acad. Sci. 1147, 358–382.
Cattaneo, E., Zuccato, C., Tartari, M., 2005. Normal huntingtin function: an alter-native approach to Huntington’s disease. Nat. Rev. Neurosci. 6, 919–930.
Cavaliere, C., Cirillo, G., Bianco, M.R., Rossi, F., De Novellis, V., Maione, S., Papa, M.,2007. Gliosis alters expression and uptake of spinal glial amino acid transpor-ters in a mouse neuropathic pain model. Neuron Glia Biol. 3, 141–153.
Chaudhry, F.A., Lehre, K.P., van Lookeren Campagne, M., Ottersen, O.P., Danbolt, N.C.,Storm-Mathisen, J., 1995. Glutamate transporters in glial plasma membranes:highly differentiated localizations revealed by quantitative ultrastructuralimmunocytochemistry. Neuron 15, 711–720.
Chawla, S., 2002. Regulation of gene expression by Ca2+ signals in neuronal cells.Eur. J. Pharmacol. 447, 131–140.
Chyi, T., Chang, C., 1999. Temporal evolution of 3-nitropropionic acid-inducedneurodegeneration in the rat brain by T2-weighted, diffusion-weighted, andperfusion magnetic resonance imaging. Neuroscience 92, 1035–1041.
Cicchetti, F., Gould, P.V., Parent, A., 1996. Sparing of striatal neurons coexpressingcalretinin and substance P (NK1) receptor in Huntington’s disease. Brain Res.730, 232–237.
Cicchetti, F., Prensa, L., Wu, Y., Parent, A., 2000. Chemical anatomy of striatalinterneurons in normal individuals and in patients with Huntington’s disease.Brain Res. Rev. 34, 80–101.
Colangelo, A.M., Bianco, M.R., Vitagliano, L., Cavaliere, C., Cirillo, G., De Gioia, L.,Diana, D., Colombo, D., Redaelli, C., Zaccaro, L., Morelli, G., Papa, M., Sarmientos,P., Alberghina, L., Martegani, E., 2008. A new nerve growth factor-mimeticpeptide active on neuropathic pain in rats. J. Neurosci. 28, 2698–2709.
Cole, A.J., Saffen, D.W., Baraban, J.M., Worley, P.F., 1989. Rapid increase of animmediate early gene messenger RNA in hippocampal neurons by synapticNMDA receptor activation. Nature 340, 474–476.
Custer, S.K., Garden, G.A., Gill, N., Rueb, U., Libby, R.T., Schultz, C., Guyenet, S.J.,Deller, T., Westrum, L.E., Sopher, B.L., La Spada, A.R., 2006. Bergmann gliaexpression of polyglutamine-expanded ataxin-7 produces neurodegenerationby impairing glutamate transport. Nat. Neurosci. 9, 1302–1311.
Danbolt, N.C., 2001. Glutamate uptake. Prog. Neurobiol. 65, 1–105.Dervan, A.G., Meshul, C.K., Beales, M., McBean, G.J., Moore, C., Totterdell, S., Snyder,
A.K., Meredith, G.E., 2004. Astroglial plasticity and glutamate function in achronic mouse model of Parkinson’s disease. Exp. Neurol. 190, 145–156.
Deshpande, S.B., Fukuda, A., Nishino, H., 1997. 3-Nitropropionic acid increases theintracellular Ca2+ in cultured astrocytes by reverse operation of the Na+–Ca2+
exchanger. Exp. Neurol. 145, 38–45.Deshpande, S.B., Nishino, H., 1998. In vitro protection of 3-nitropropionic acid-
induced toxicity of astrocytes by basic fibroblast growth factor and thrombin.Brain Res. 783, 28–36.
Doble, A., 1999. The role of excitotoxicity in neurodegenerative disease: implica-tions for therapy. Pharmacol. Ther. 81, 163–221.
Fernagut, P.O., Diguet, E., Stefanova, N., Biran, M., Wenning, G.K., Canioni, P., Bioulac,B., Tison, F., 2002. Subacute systemic 3-nitropropionic acid intoxication inducesa distinct motor disorder in adult C57Bl/6 mice: behavioural and histopatho-logical characterisation. Neuroscience 114, 1005–1017.
Fukuda, A., Deshpande, S.B., Shimano, Y., Nishino, H., 1998. Astrocytes are morevulnerable than neurons to cellular Ca2+ overload induced by a mitochondrialtoxin, 3-nitropropionic acid. Neuroscience 87, 497–507.
Galas, M.C., Bizat, N., Cuvelier, L., Bantubungi, K., Brouillet, E., Schiffmann, S.N., Blum,D., 2004. Death of cortical and striatal neurons induced by mitochondrial defectinvolves differential molecular mechanisms. Neurobiol. Dis. 15, 152–159.
Garcia, M., Vanhoutte, P., Pages, C., Besson, M.J., Brouillet, E., Caboche, J., 2002. Themitochondrial toxin 3-nitropropionic acid induces striatal neurodegenerationvia a c-Jun N-terminal kinase/c-Jun module. J. Neurosci. 22, 2174–2184.
Ginsberg, S.D., Martin, L.J., Rothstein, J.D., 1995. Regional deafferentation down-regulates subtypes of glutamate transporter proteins. J. Neurochem. 65, 2800–2803.
Giovannoni, R., Maggio, N., Bianco, M.R., Cavaliere, C., Cirillo, G., Lavitrano, M., Papa,M., 2007. Reactive astrocytosis and glial glutamate transporter clustering areearly changes in a spinocerebellar ataxia type 1 transgenic mouse model.Neuron Glia Biol. 3, 335–351.
Graham, R.K., Pouladi, M.A., Joshi, P., Lu, G., Deng, Y., Wu, N.P., Figueroa, B.E., Metzler,M., Andre, V.M., Slow, E.J., Raymond, L., Friedlander, R., Levine, M.S., Leavitt, B.R.,Hayden, M.R., 2009. Differential susceptibility to excitotoxic stress in YAC128mouse models of Huntington disease between initiation and progression ofdisease. J. Neurosci. 29, 2193–2204.
Guyot, M.C., Hantraye, P., Dolan, R., Palfi, S., Maziere, M., Brouillet, E., 1997.Quantifiable bradykinesia, gait abnormalities and Huntington’s disease-likestriatal lesions in rats chronically treated with 3-nitropropionic acid. Neu-roscience 79, 45–56.

G. Cirillo et al. / Neurochemistry International 56 (2010) 152–160160
Haydon, P.G., Carmignoto, G., 2006. Astrocyte control of synaptic transmission andneurovascular coupling. Physiol. Rev. 86, 1009–1031.
Hickey, M.A., Kosmalska, A., Enayati, J., Cohen, R., Zeitlin, S., Levine, M.S., Chesselet,M.F., 2008. Extensive early motor and non-motor behavioral deficits are fol-lowed by striatal neuronal loss in knock-in Huntington’s disease mice. Neu-roscience 157, 280–295.
Hughes, E.G., Maguire, J.L., McMinn, M.T., Scholz, R.E., Sutherland, M.L., 2004. Loss ofglial fibrillary acidic protein results in decreased glutamate transport andinhibition of PKA-induced EAAT2 cell surface trafficking. Brain Res. Mol. BrainRes. 124, 114–123.
Landles, C., Bates, G.P., 2004. Huntingtin and the molecular pathogenesis of Hun-tington’s disease. Fourth in molecular medicine review series. EMBO Rep. 5,958–963.
Lehre, K.P., Levy, L.M., Ottersen, O.P., Storm-Mathisen, J., Danbolt, N.C., 1995. Differ-ential expression of two glial glutamate transporters in the rat brain: quantitativeand immunocytochemical observations. J. Neurosci. 15, 1835–1853.
Lehre, K.P., Danbolt, N.C., 1998. The number of glutamate transporter subtypemolecules at glutamatergic synapses: chemical and stereological quantificationin young adult rat brain. J. Neurosci. 18, 8751–8757.
Lobsiger, C.S., Cleveland, D.W., 2007. Glial cells as intrinsic components of non-cell-autonomous neurodegenerative disease. Nat. Neurosci. 10, 1355–1360.
Martin Aparicio, E., Lucas, J.J., 2002. Molecular basis of Huntington’s disease andpossible pathogenic mechanisms. Rev. Neurol. 35, 212–220.
McCracken, E., Dewar, D., Hunter, A.J., 2001. White matter damage followingsystemic injection of the mitochondrial inhibitor 3-nitropropionic acid in rat.Brain Res. 892, 329–335.
Morgan, J.I., Cohen, D.R., Hempstead, J.L., Curran, T., 1987. Mapping patterns of c-fosexpression in the central nervous system after seizure. Science 237, 192–197.
Mrak, R.E., Griffin, W.S., 2005. Glia and their cytokines in progression of neurode-generation. Neurobiol. Aging 26, 349–354.
Musholt, K., Cirillo, G., Cavaliere, C., Bianco, M.R., Bock, J., Helmeke, C., Braun, K.,Papa, M., 2009. Neonatal separation stress reduces glial fibrillary acidic protein-and S100beta-immunoreactive astrocytes in the rat medial precentral cortex.Dev. Neurobiol. 69, 203–211.
Olsen, C., Rustad, A., Fonnum, F., Paulsen, R.E., Hassel, B., 1999. 3-Nitropropionicacid: an astrocyte-sparing neurotoxin in vitro. Brain Res. 850, 144–149.
Page, K.J., Dunnett, S.B., Everitt, B.J., 1998. 3-Nitropropionic acid-induced changes inthe expression of metabolic and astrocyte mRNAs. Neuroreport 9, 2881–2886.
Palfi, S., Ferrante, R.J., Brouillet, E., Beal, M.F., Dolan, R., Guyot, M.C., Peschanski, M.,Hantraye, P., 1996. Chronic 3-nitropropionic acid treatment in baboons repli-cates the cognitive and motor deficits of Huntington’s disease. J. Neurosci. 16,3019–3025.
Papa, M., Boscia, F., Canitano, A., Castaldo, P., Sellitti, S., Annunziato, L., Taglialatela,M., 2003. Expression pattern of the ether-a-gogo-related (ERG) K+ channel-encoding genes ERG1, ERG2, and ERG3 in the adult rat central nervous system. J.Comp. Neurol. 466, 119–135.
Paulsen, J.S., Magnotta, V.A., Mikos, A.E., Paulson, H.L., Penziner, E., Andreasen, N.C.,Nopoulos, P.C., 2006. Brain structure in preclinical Huntington’s disease. Biol.Psychiatry 59, 57–63.
Paulson, H.L., Fischbeck, K.H., 1996. Trinucleotide repeats in neurogenetic disorders.Annu. Rev. Neurosci. 19, 79–107.
Reiner, A., Perera, M., Paullus, R., Medina, L., 1998. Immunohistochemical localiza-tion of DARPP32 in striatal projection neurons and striatal interneurons inpigeons. J. Chem. Neuroanat. 16, 17–33.
Rodriguez, J.J., Olabarria, M., Chvatal, A., Verkhratsky, A., 2009. Astroglia in dementiaand Alzheimer’s disease. Cell Death Differ. 16, 378–385.
Rothstein, J.D., Martin, L., Levey, A.I., Dykes-Hoberg, M., Jin, L., Wu, D., Nash, N.,Kuncl, R.W., 1994. Localization of neuronal and glial glutamate transporters.Neuron 13, 713–725.
Saffen, D.W., Cole, A.J., Worley, P.F., Christy, B.A., Ryder, K., Baraban, J.M., 1988.Convulsant-induced increase in transcription factor messenger RNAs in ratbrain. Proc. Natl. Acad. Sci. U.S.A. 85, 7795–7799.
Saijo, K., Winner, B., Carson, T.C., Collier, J.G., Boyer, L., Rosenfeld, M.G., Gage,F.H., Glass, C.K., 2009. A Nurr1/CoREST pathway in microglia and astrocytesprotects dopaminergic neurons from inflammation-induced death. Cell 137,47–59.
Schmued, L.C., Hopkins, K.J., 2000a. Fluoro-Jade B: a high affinity fluorescent markerfor the localization of neuronal degeneration. Brain Res. 874, 123–130.
Schmued, L.C., Hopkins, K.J., 2000b. Fluoro-Jade: novel fluorochromes for detectingtoxicant-induced neuronal degeneration. Toxicol. Pathol. 28, 91–99.
Schmued, L.C., Stowers, C.C., Scallet, A.C., Xu, L., 2005. Fluoro-Jade C results in ultrahigh resolution and contrast labeling of degenerating neurons. Brain Res. 1035,24–31.
Seppi, K., Mueller, J., Bodner, T., Brandauer, E., Benke, T., Weirich-Schwaiger, H.,Poewe, W., Wenning, G.K., 2001. J. Neurol. 248, 866–869.
Sheldon, A.L., Robinson, M.B., 2007. The role of glutamate transporters in neuro-degenerative diseases and potential opportunities for intervention. Neurochem.Int. 51, 333–355.
Smith, R., Brundin, P., Li, J.Y., 2005. Synaptic dysfunction in Huntington’s disease: anew perspective. Cell Mol. Life Sci. 62, 1901–1912.
Sung, B., Lim, G., Mao, J., 2003. Altered expression and uptake activity of spinalglutamate transporters after nerve injury contribute to the pathogenesis ofneuropathic pain in rats. J. Neurosci. 23, 2899–2910.
Teunissen, C.E., Steinbusch, H.W., Angevaren, M., Appels, M., de Bruijn, C., Prickaerts,J., de Vente, J., 2001. Behavioural correlates of striatal glial fibrillary acidicprotein in the 3-nitropropionic acid rat model: disturbed walking pattern andspatial orientation. Neuroscience 105, 153–167.
Tilleux, S., Goursaud, S., Hermans, E., 2009. Selective up-regulation of GLT-1 incultured astrocytes exposed to soluble mediators released by activated micro-glia. Neurochem. Int. 55, 35–40.
Wullner, U., Young, A.B., Penney, J.B., Beal, M.F., 1994. 3-Nitropropionic acid toxicityin the striatum. J. Neurochem. 63, 1772–1781.
Xin, W.J., Weng, H.R., Dougherty, P.M., 2009. Plasticity in expression of the gluta-mate transporters GLT-1 and GLAST in spinal dorsal horn glial cells followingpartial sciatic nerve ligation. Mol. Pain 26, 5–15.
Zagami, C.J., O’Shea, R.D., Lau, C.L., Cheema, S.S., Beart, P.M., 2005. Regulation ofglutamate transporters in astrocytes: evidence for a relationship betweentransporter expression and astrocytic phenotype. Neurotox. Res. 7, 143–149.




















