
Prospective identification of myogenic endothelial cells in human skeletal muscle

Acta Biomaterialia 8 (2012) 531–539
Contents lists available at SciVerse ScienceDirect
Acta Biomaterialia
journal homepage: www.elsevier .com/locate /actabiomat
Direct laser machining-induced topographic pattern promotes up-regulationof myogenic markers in human mesenchymal stem cells
Huaqiong Li a, Feng Wen a, Yee Shan Wong a, Freddy Yin Chiang Boey a, Venkatraman S. Subbu a,David Tai Leong b,c, Kee Woei Ng a, Gary Ka Lai Ng d, Lay Poh Tan a,⇑a Division of Materials Technology, School of Materials Science and Engineering, Nanyang Technological University, 50 Nanyang Avenue, 639798 Singapore, Singaporeb Department of Chemical and Biomolecular Engineering, National University of Singapore, 4 Engineering Drive, 117576 Singapore, Singaporec Cancer Science Institute of Singapore, National University of Singapore, 28 Medical Drive, 117456 Singapore, Singapored Singapore Institute of Manufacturing Technology, 71 Nanyang Drive, 638075 Singapore, Singapore
a r t i c l e i n f o
Article history:Received 24 May 2011Received in revised form 22 September 2011Accepted 22 September 2011Available online 28 September 2011
Keywords:hMSCsFemtosecond laserTopographyMyogenesis
1742-7061/$ - see front matter � 2011 Acta Materialdoi:10.1016/j.actbio.2011.09.029
⇑ Corresponding author. Tel.: +65 67906186; fax: +E-mail address: [email protected] (L.P. Tan).
a b s t r a c t
The engineering of tissue is preferably done with stem cells, which can be differentiated into the tissue ofinterest using biochemical or physical cues. While much effort has been focused on using biological factorsto regulate stem cell differentiation, recently interest in the contribution of physical factors has increased.In this work, three-dimensional (3-D) microchannels with topographic micropatterns were fabricated byfemtosecond laser machining on a biodegradable polymer (poly(L-lactide-co-e-caprolactone)) substrate.Two substrates with narrow and wide channels respectively were created. Human mesenchymal stemcells (hMSCs) were cultured on the scaffolds for cell proliferation and cellular organization. Gene expres-sion and the immunostaining of myogenic and neurogenic markers were studied. Both scaffolds improvedthe cell alignment along the channels as compared to the control group. Microfilaments within hMSCswere more significantly aligned and elongated on the narrower microchannels. The gene expression studyrevealed significant up-regulation of several hallmark markers associated with myogenesis for hMSCs cul-tured on the scaffold with narrow microchannels, while osteogenic and neurogenic markers were down-regulated or remained similar to the control at day 14. Immunostaining of myogen- and neurogen-specificdifferentiation markers were used to further confirm the specific differentiation towards a myogenic line-age. This study demonstrates that femtosecond laser machining is a versatile tool for generating control-lable 3-D microchannels with topographic features that can be used to induce specific myogenicdifferentiation of hMSCs in vitro, even in the absence of biological factors.
� 2011 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Tissue engineering aims to develop biocompatible cell–scaffoldconstructs that integrate with native tissue in situ to repair or re-place damaged tissue [1]. To achieve this aim, it is imperative touse cells which can augment the cellular functions in these cell–scaffold constructs. Stem cells, which are unspecialized precursorcells with the ability to self-renew and differentiate into differentcell types, are such cells [2]. Adult human mesenchymal stem cells(hMSCs) isolated from bone marrow have been proven capable ofself-renewal and differentiation into a wide variety of lineages inresponse to proper cues [3–5]. The cues include soluble/immobi-lized factors, chemical and physical signals from the extracellularmatrix [6]. Molecular mediators (e.g. growth factors, transcriptionfactors) capable of regulating stem cell fates are well characterized.However, increasing evidence has shown that non-molecular stim-
ia Inc. Published by Elsevier Ltd. A
65 67909081.
uli like physical environmental factors (e.g. matrix compliance [7]and surface topography [8]) can also control stem cell activities [9],whilst avoiding side effects such as uncontrollable cell growth,tumorigenesis and even cell death.
Topography, particularly controllable nanoscale topography, isone of the most important physical inductions, as increasing evi-dence has demonstrated that topography has great potential inregulating cell behavior, especially in the presence of biologicalor chemical induction media [10,11]. Interestingly, some recentstudies have revealed that, in the absence of any inductive biolog-ical agents, topographical cues alone may direct stem cell differen-tiation [8,12,13]. Electrospinning and lithography are twocommonly used techniques to construct scaffold with controllabletopographic features. Dang and Leong [12] have demonstrated thataligned electrospun hydroxybutyl chitosan fibers can up-regulatemyogenic genes of Pax-43, Pax-7 and myogenin of hMSCs to indi-cate myogenic lineage commitment. Similarly, Yim et al. [13]found that nanogratings of PDMS induced neuronal proteinsexpression of Tuj-1 and MAP2, as confirmed by both microarray
ll rights reserved.

532 H. Li et al. / Acta Biomaterialia 8 (2012) 531–539
and real-time polymerase chain reaction (RT-PCR) analyses. Osteo-progenitor cells and hMSCs, cultured on poly(methyl methacry-late)-based nanopits of ordered arrangements did not result inany expression of osteopontin or osteocalcin in hMSCs [8], whileirregular nanopits induced significantly higher amounts of thosetwo proteins and bone nodule formation. Though these methodswere broadly utilized to generate topography, there are distinctdrawbacks that limit their applications. One clear disadvantageof electrospinning is the limitation of scaling-up. For other conven-tional and popular lithography techniques, such as photolithogra-phy and electron beam lithography, the requirement oftemplates, facilities and multiple steps (e.g. spin coater, an appro-priate photoresist, plasma or chemical etching of the surface, re-moval of photoresist, each step needing special systems) makesthe technique expensive, less versatile (a separate template foreach pattern) and time consuming.
Recently, with advancements in laser technology, materialmodification using lasers has attracted much attention. Lasermachining is versatile, as it can be applied to a wide range of mate-rials. Compared to traditional techniques, laser machining offersmore versatility (no templates required) and scalability (to largerdimensions), and can be carried out rapidly and under ambientconditions. Femtosecond lasers can reduce the area of the heat-af-fected zone compared to lasers with a longer pulse width. Femto-second laser machining has been applied widely for the surfacemodification of polymers [14–22]. The application of femtosecondlaser machining in biomaterial is still very limited, even thoughthis method is highly efficient in creating unique surfaces and pat-terns. Femtosecond laser-generated perforation in acellular matrixgrafts is used for small-bore vascular implant. When transplanted,the perforated implants exhibited faster recellularization withendothelial cells than contralateral controls [20]. Another studyof micropatterning on collagen showed viable growth of rat mes-enchymal stem cells and fibroblasts in the ablated patterns, butfailed to illustrate the differentiation potential of hMSCs due tothe micropatterns [21]. We too have used femtosecond lasermachining to create scaffolds and investigated the lane effects oncell alignment [23]. In that study, we used the femtosecond laserto create highly uniform microchannels directly on the selectedbiodegradable polymer, poly(L-lactide-co-e-caprolactone) (PLLA–PCL) substrate, for enhanced cell alignment.
In this paper, we studied whether there were any lineage differ-entiations of hMSCs due to three-dimensional (3-D) microchannelsbeing formed by laser machining. We found that the optimizedchannel dimensions were able to direct hMSCs to align and elon-gate. This consequently induced myogenic differentiation withoutany exogeneously introduced induction medium. Other cellularbehaviors, such as proliferation and cellular organization, werealso investigated.
2. Materials and methods
2.1. Preparation of PLLA–PCL film
PLLA–PCL film was fabricated by the solvent casting method. Ablock copolymer PLLA–PCL (70:30) comprising 70% L-lactide and30% e-caprolactone, with a weight-averaged molecular weight of202,000 g mol�1, was used in this study (Purac Biomaterials, Lin-colnshire, IL). The copolymer PLLA–PCL granules were dissolvedin dichloromethane (Tedia, Fairfield, OH) in a 1:5 mass/volumeratio to achieve the optimal viscosity for the solvent casting pro-cess. The films were cast using an industrial film applicator(Sheen automatic film applicator, Paul N. Gardner Company,Inc., FL, USA). The polymer films were dried at 37 �C in a vacuumoven for 7 days. The thickness of the dry-cast films was approxi-mately 100 lm.
2.2. Direct laser machining of polymer
The micropatterning of the biopolymer film was carried out usinga Ti-sapphire (Clark-MXR CPA 2001, Dexter, MI) infrared solid-statelaser with a chirped pulse amplifier to produce high-intensitypulses, with the following technical specifications: pulse dura-tion = 150 fs; central wavelength = 800 nm; maximum power =800 mW; fixed repetition rate = 1 kHz. The laser pulse energy is ad-justed by an attenuator. The femtosecond laser beam is focused byan optical focus lens. With appropriate setting, the focused laserbeam can directly induce ablation in the polymer substrate. Twotypes of microchannels were produced: one of 30 lm width,30 lm interval and 20 lm depth (30–30–20), the other of 80 lmwidth, 30 lm interval and 20 lm depth (80–30–20). The scanningspeed of the femtosecond laser was 10 mm s�1. Narrower channelswere created by scanning two passes on the toolpath at a laser powerof 100 mW, while the wider channels only need one pass at a laserpower of 450 mW to get the same depth as that of the narrowerchannel.
2.3. Characterization of polymer films
The surface morphology of the microchannel scaffold was ob-served using scanning electron microscopy (SEM) performed ongold-sputtered samples. After coating, the samples were viewedunder a Jeol JSM-6360LV scanning electron microscope. AliconaInfiniteFocus was employed to generate a 3-D structure for sur-face analysis. The sample was placed on a motorized stage andilluminated with light. An objective lens of 10� magnificationwas used. The distance between the sample and the objectivewas adjusted to focus the image of the structure. Once the imagewas focused, the upper and lower limits of the structure surface(in the software) were set to generate the 3-D structure surface.The profile of the surface and its particular roughness could beobtained via the 3-D structure. Attenuated total reflectance Fou-rier transform infrared (ATR-FTIR) spectroscopic analysis of thePLLA–PCL substrates were performed on a PerkinElmer SpectrumGX spectrometer at a range of 600–4000 cm�1, using diamondcrystal as the internal reflection element. All spectra were calcu-lated means from eight scans at a resolution of 2 cm�1, with cor-rection for atmospheric water and carbon dioxide. The surfacechemical composition of the substrates was measured by X-rayphotoelectron spectroscopy (XPS) using a VG ESCALAB 250 spec-trometer (Thermo Electron, UK). All the binding energies werecorrected by referencing the binding energy of C1s in the C–Hgroup to 285.0 eV.
2.4. Cell culture
hMSCs and cell culture medium were both obtained fromLonza (Cambrex, UK). The hMSCs were expanded and culturedin mesenchymal stem cell basal medium according to the ven-dor’s instructions. For the experiments, the cells at passage 5were cultured with low-glucose Dulbecco’s modified Eagle’smedium (DMEM) containing L-glutamine (Sigma–Aldrich, USA),supplemented with 10% fetal bovine serum (FBS; PAA, Pasching,Austria) and 1% antibiotic/antimycotic solution (PAA) at 37 �C ina humidified atmosphere of 5% CO2. The culture medium waschanged every 2–3 days. Phosphate-buffered saline (PBS) and0.05% trypsin–ethylenediaminetetraacetic acid (EDTA) obtainedfrom Invitrogen were used for washing and cell detachment pur-poses, respectively. Scaffolds were sterilized using 70% ethanolfor 30 min. The scaffolds were then washed with deionizedwater and PBS three times, for 15 min each time. Before cellseeding, the scaffolds were immersed into the culture mediumfor 30 min.

H. Li et al. / Acta Biomaterialia 8 (2012) 531–539 533
2.5. Immunocytochemistry and microscopy
Briefly, cells were fixed with 4% paraformaldehyde for 15–20 min and then permeabilized with 0.1% Triton X-100 for5 min, followed by blocking with 5% bovine serum albumin(BSA) (Invitrogen) or 5% goat serum (Sigma). The primary anti-bodies used to target interested proteins were: rabbit polyclonalantifibronectin (1:400) (Sigma), chicken polyclonal antiMAP2(1:10000, Abcam) and mouse monoclonal antiheavy chain cardiacmyosin (1:200) (Abcam). Secondary antibodies were Cy3 goatanti-rabbit IgG (1:400, Millipore), Alexa Fluor� 568 goat anti-chicken (1:400, Molecular probes), and Alexa Fluor� 488 goatanti-mouse (1:400, Molecular probes). In order to verify the spec-ificity of the primary antibodies, C2C12 myoblasts and neuralstem cells were used as positive controls for major histocompat-ibility complex (MHC) and microtubule-associated protein 2(MAP2), respectively. The C2C12 myoblasts were induced to dif-ferentiate in 2% horse serum and stained for cardiac MHC. Neuralstem cells were induced to differentiate in DMEM F-12 andstained for MAP2. Negative controls in the absence of primaryantibodies were also performed. F-actin was stained with tetra-methyl rhodamine isothiocyanate-conjugated phalloidin (1:500,Chemicon). Nuclei were stained with 40,6-diamidino-2-phenylin-dole (DAPI, 1:1000; Chemicon). Fluorescence images were visual-ized with a Nikon 80i eclipse (Nikon, Japan) upright microscopeand captured with a Nikon DS-Fi1 digital camera (Nikon, Japan)using two 20� and 40� objective lens. A Leica TCS SP5 confocallaser scanning microscope system was also used to capture 3-Dimages using a 63� objective lens (oil).
2.6. PicoGreen assay
The scaffolds with cells were washed three times in PBS, priorto being lysed with 0.1 vol.% Triton X-100 solution with gentlepipetting for a duration of 30 min (0.5 ml per well of a 24-well
Fig. 1. Topography of microchannels fabricated directly using femtosecond laser on biod(SEM and 3-D profiler).
plate). The PicoGreen working solution was then prepared bydiluting the concentrated PicoGreen reagent in TE buffer(10 mM Tris, 1 mM EDTA, pH 7.4) at a dilution ratio of 1:200according to the manufacturer’s instructions. Subsequently, thecell lysate was mixed with the PicoGreen working solution at a1:1 ratio (100 ll within each well of a 96-well plate) and incu-bated for at least 5 min at room temperature, prior to readingthe fluorescence emission at 520 nm under an excitation wave-length of 480 nm with an Infinite200� microplate reader (TecanInc., Maennedorf, Switzerland).
2.7. Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA (tRNA) was extracted with RNeasy mini kit (Qiagen,Valencia, CA). The concentration and quality of the extractedtRNA was determined spectrophotometrically (Nanodrop-N100,Thermo Scientific). Subsequently, cDNA was obtained by reversetranscription of total RNA with random hexamers and SuperscriptIII reverse transcriptase (Invitrogen). qRT-PCR was performed on aCFX96 RT-PCR detection system (Bio-Rad Laboratories, Inc., USA)for 40 cycles. Primers specific to the targeted genes (listed inTable S1) were obtained from primerbank [24]. The relative quan-tification of gene expression was determined by comparing thetarget-amplified product against glyceraldehyde-3-phosphatedehydrogenase and b-actin (both were used as internal stan-dards). Three experiments and triplicates were done for theqRT-PCR. The results were analyzed using Relative ExpressionSoftware Tool software [25].
2.8. Statistical analysis
The mean and standard deviation were calculated for eachgroup of experimental data. Group means testing was performedusing analysis of variance for multiple comparisons. The signifi-cance level was set at 0.05.
egradable PLLA–PCL film, with sinusoidal surface topology inside the microchannels

0
5
10
15
20
25
30
35
% T
otal
Alignment (Degree)
Day 1A
B
C
D
control
30-30-20
80-30-20
0
10
20
30
40
50
% T
otal
Alignment (Degree)
Day 4control
30-30-20
80-30-20
0
10
20
30
40
50
60
70
% T
otal
Alignment (Degree)
Day 7 control
30-30-20
80-30-20
control 30-30-20 80-30-20
2D F
FT
(D
ay 7
)
Fig. 2. Alignment of hMSCs on unpatterned (control) and laser-micropatternedsubstrates. Quantitative study of cell orientation was done at days 1, 4 and 7;qualitative analysis using 2-D FFT analysis was done at day 7. Panel A: day 1; panelB: day 4; panel C: day 7; panel D: 2-D FFT analysis of day 7.
534 H. Li et al. / Acta Biomaterialia 8 (2012) 531–539
3. Results
3.1. Characterization of polymer surface
A femtosecond laser is able to apply highly focused energy forthe removal of material. By changing the laser power, scan speedand scan pass, microchannels with two desirable dimensions weremachined onto a 10 mm � 10 mm PLLA–PCL film. The featurepitches (the combination of groove and interval widths) were uni-form in each sample. As shown in Fig. 1, a 3-D microchannel topog-raphy with a lined structure on the top view (SEM images) and asinusoidal surface on the cross-section (3-D profiler images) wasachieved on the flat 2-D surface of the raw material throughfemtosecond laser machining. The channels were fabricated at apredetermined width, depth and interval distance. A 3-D profilerwas used to measure the roughness inside and outside themicrochannels. Two parameters, average roughness (Ra) androot-mean-square (Rq), were utilized to characterize the rough-ness of surfaces. The roughness in the channels for narrowchannels (30–30–20) and wider channels (80–30–20) wereRa = 0.80 lm, Rq = 0.98 lm and Ra = 0.83 lm, Rq = 1.05 lm,respectively. For the intervals (outside the channels), the rough-ness of both narrow and wider channels was approximatelyRa = 0.35 lm, Rq = 0.42 lm, which was clearly smaller than theroughness in the microchannels. Therefore, both types of channelwere considered to have similar roughness.
The substrates with and without laser machining were exam-ined by ATR-FTIR and XPS. As shown in the ATR-FTIR spectra(Fig. S1), there were almost no differences between laser-machinedsubstrates and the controls. All spectra revealed the characteristicpeaks of pure PLLA–PCL copolymer. The bands between 2870 and2995 cm�1 were present due to C–H stretching vibrations of alkylCH3, CH2 and CH groups. The bands from 1350 to 1460 cm�1 wereassigned to alkyl C–H bending vibrations [26]. The characteristicester absorption peak of PLLA–PCL was observed at 1744 cm�1,and C–O–C stretching vibrations were observed at 1084, 1128and 1182 cm�1 [27]. The results of the XPS spectra (Fig. S2) showedthe elemental and chemical states of specific atoms on the materialsurface. Identical carbon and oxygen signals were detected by anXPS survey scan of the substrates surface before and after lasermachining. The C1s detailed core-level spectra revealed three ma-jor peaks with similar binding energies in each high-resolution XPSspectrum (C–H, C–C: 285.0 ± 0.1 eV; C–O: 287.0 ± 0.1 eV; C@O:289.1 ± 0.1 eV) [28], again indicating that there was no significantchemical change after laser machining.
3.2. Cellular organization
hMSCs were cultured on the respective substrates (laser-ma-chined substrates with dimensions 30–30–20 and 80–30–20 mand control without laser machining) for 1, 4, 7, 11 and 14 days.To examine the cell morphology and microfilament arrangement,scaffolds with hMSCs were stained with phalloidin for F-actinand DAPI for nucleus. When hMSCs were cultured on the PLLA–PCL scaffolds with laser-machined microchannels, both narrowand wide channels helped the cells to elongate and align alongthe grating axis. In contrast, hMSCs cultured on controls showedno elongation or alignment in any specific direction.
The quantitative and qualitative analysis of cell alignment ondifferent substrates is shown in Fig. 2. The cells cultured on thecontrol were randomly oriented, with degrees of alignment from0� to 90� even when the cells were becoming confluent. The cellscultured on the scaffolds with narrow microchannels, 30–30–20,had better alignment than those on the wider channels, 80–30–20. This was observed from the beginning of cell growth on day1 (Fig. 2A). As seen in Fig. 2, for the narrower channel system
(30–30–20), the angular distribution of the cells changed from amaximum of around 10–19� on day 4 (Fig. 2B) to a maximum ofaround 0–5� on day 7 (Fig. 2C), clearly indicating progressivelygreater alignment of the cells along the grooved direction. This isnot as evident in the wider channel substrate (80–30–20), where
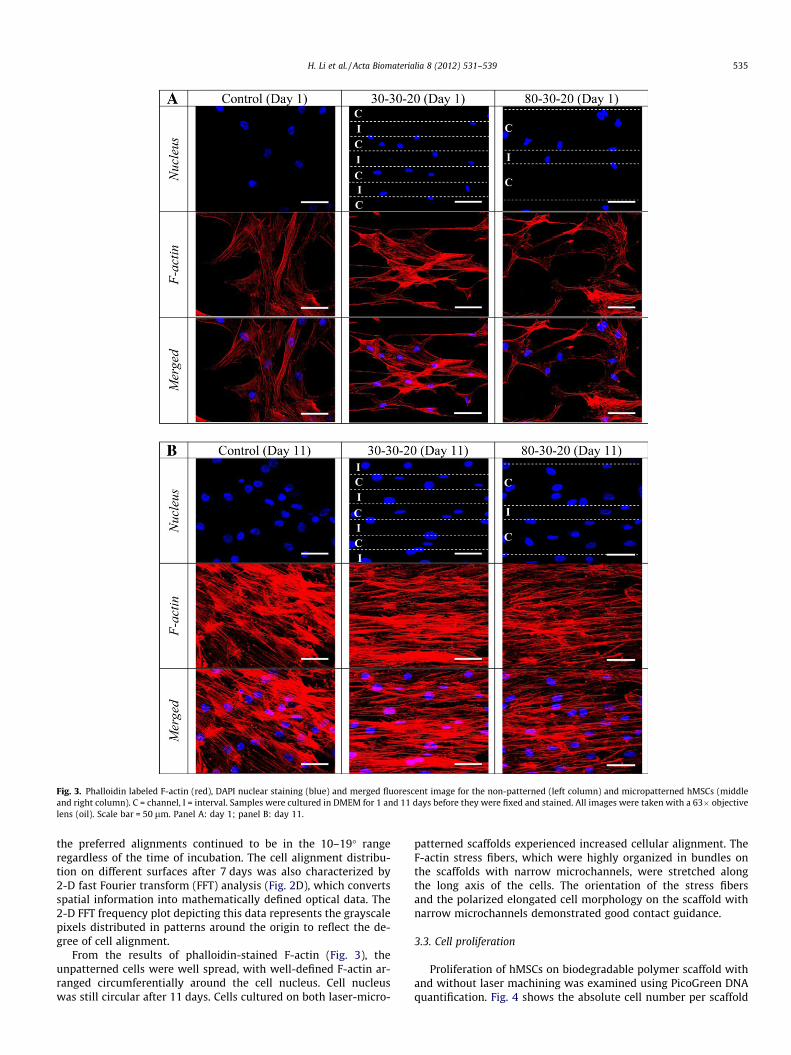
Fig. 3. Phalloidin labeled F-actin (red), DAPI nuclear staining (blue) and merged fluorescent image for the non-patterned (left column) and micropatterned hMSCs (middleand right column). C = channel, I = interval. Samples were cultured in DMEM for 1 and 11 days before they were fixed and stained. All images were taken with a 63� objectivelens (oil). Scale bar = 50 lm. Panel A: day 1; panel B: day 11.
H. Li et al. / Acta Biomaterialia 8 (2012) 531–539 535
the preferred alignments continued to be in the 10–19� rangeregardless of the time of incubation. The cell alignment distribu-tion on different surfaces after 7 days was also characterized by2-D fast Fourier transform (FFT) analysis (Fig. 2D), which convertsspatial information into mathematically defined optical data. The2-D FFT frequency plot depicting this data represents the grayscalepixels distributed in patterns around the origin to reflect the de-gree of cell alignment.
From the results of phalloidin-stained F-actin (Fig. 3), theunpatterned cells were well spread, with well-defined F-actin ar-ranged circumferentially around the cell nucleus. Cell nucleuswas still circular after 11 days. Cells cultured on both laser-micro-
patterned scaffolds experienced increased cellular alignment. TheF-actin stress fibers, which were highly organized in bundles onthe scaffolds with narrow microchannels, were stretched alongthe long axis of the cells. The orientation of the stress fibersand the polarized elongated cell morphology on the scaffold withnarrow microchannels demonstrated good contact guidance.
3.3. Cell proliferation
Proliferation of hMSCs on biodegradable polymer scaffold withand without laser machining was examined using PicoGreen DNAquantification. Fig. 4 shows the absolute cell number per scaffold

0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
day 1 day 4 day 7 day 11 day 14
Cel
l num
ber (
/cm
2 )
control
30-30-20
80-30-20
Fig. 4. Cell number of hMSCs 1, 4, 7, 11 and 14 days after seeding at an initialseeding density of 5000 cells per cm2 on scaffolds without laser machining,scaffolds with narrow channel width and scaffolds with wide channel width. Therewas no significant difference in the cell number between the two groups with lasermachining (p > 0.05).
536 H. Li et al. / Acta Biomaterialia 8 (2012) 531–539
at days 1, 4, 7, 11 and 14 after seeding at an initial cell seeding den-sity of 5000 cells per cm2. The control group without laser machin-ing had higher cell number compared with the laser-machinedscaffolds. Cells in the control group reached 100% confluence after11 days. Differences in cell numbers of both laser-machinedscaffolds were not statistically significant different (p > 0.05).
3.4. Gene expression analysis
Fig. 5 shows the profile of gene expression by hMSCs culturedon the laser micropatterned scaffolds after 7, 11 and 14 days, withcells cultured on unpatterned scaffolds as the control. Alkalinephosphatase (ALPL), osteonectin (ON) and runt-related transcrip-tion factor 2 (RUNX2) were chosen as markers of osteogenesis.GATA binding protein 4 (GATA4), myosin heavy chain 7 (MHC7),myogenic differentiation 1 (MyoD1) and NK2 transcription factorrelated, locus 5 (Nkx2.5) were used to confirm myogenesis, whilemicrotubule-associated protein 2 (MAP2), nestin and neurogenicdifferentiation 1 (NeuroD1) were taken as the markers ofneurogenesis.
Osteogenic genes did not show any statistically significant dif-ference (p > 0.05; Fig. 5A and B). ALPL is essential for the processof mineralization [29,30]. Osteonectin is a glycoprotein in the bonethat binds calcium, which was detected during bone formation andinitiating mineralization [31]. RUNX2 is a key transcription factorinvolved in bone mineralization maintenance [32,33]. At days 7and 11 of cell culture, RUNX2 levels were high, but had decreasedby day 14. For the other osteogenic markers, ALPL and osteonectin,there was no statistically significant down-regulation compared tothe control at the earlier time points, though there was a dip at day14.
MyoD1 is one of the basic helix–loop–helix (bHLH) transcrip-tion factors capable of regulating myogenesis [34]. The other threeselected myogenic genes, GATA4, MHC7 and Nkx2.5, are widelyknown to be important regulators of cardiomyogenesis. The tis-sue-restricted GATA-4 transcription factor and the Nkx2.5 homeo-domain protein are two early markers of precardiac cells [35]. TheMYH7 gene encodes a protein known as the cardiac beta (b)-myo-sin heavy chain, which plays a critical role in the skeletal and car-diac muscle functionality of the contractile machinery [36]. Thegene expression of hMSCs cultured on the scaffolds with narrowmicrochannels showed a definitive up-regulation of myogenesisand was found to gradually increase from day 7 to day 14 whencompared to control samples (Fig. 5C). The result can be taken to
imply that hMSCs cultured on the laser-patterned scaffolds withnarrow microchannels had differentiated towards the myogeniclineage as a result of elongated cell shape [37]. After 11 days of cellculture, cell–cell contact caused by confluence may lead to elon-gated cell shape formation. However, this effect may not be suffi-cient, unlike the elongated cell shape caused by narrow channelsresulting in the unstable myogenic genes up-regulation. This issupported by the fact that hMSCs cultured on the laser-patternedscaffolds with wider microchannels only showed up-regulation ofmyogenesis at the later time point of day 11 (Fig. 5D).
Nestin is a transient neurogenic marker during the early stageof development of neural stem/progenitor cells [38]. NeuroD1 isessential for the survival and maturation of adult-born neurons[39], while the MAP2 gene is involved in the process of microtu-bule assembly, which is critical for the formation and maintenanceof neuritis [40]. For cells cultured on the narrow microchannels,nestin, the chosen early stage marker for neurogenesis, was statis-tically significantly up-regulated on days 7 and 11, but then itgradually decreased, to reach nearly the same level as the controlon day 14. The bHLH transcription factor NeuroD1 and the lateneurogenic marker MAP2 both showed higher up-regulation ofgene expression than nestin at days 7 and 11; however, they thendecreased to a similar level as nestin on day 14 of cell culture(Fig. 5E). Meanwhile, for the laser-patterned scaffolds with widemicrochannels, only the NeuroD1 gene showed significant up-reg-ulation on day 11 of the hMSC culture (Fig. 5F).
3.5. Immunostaining of myogenic and neurogenic markers
In order to confirm the phenotypes and differentiation capaci-ties of hMSCs cultured on the laser-micropatterned scaffold,immunostainings for mesenchymal lineage-specific markers (ma-ture markers of myogenesis, cardiac MHC and neurogenesisMAP2) were performed. Dual labeling with antibodies againstMHC and MAP2 detected MHC staining on the laser-patterned scaf-folds with microchannels on days 11 and 14 of cell culture, butthere was no observable MAP2 staining on both groups of laser-patterned scaffolds at all time points (Fig. 6). The qRT-PCR andimmunostaining data suggest that our special microchannels helpdrive plated MSCs towards the myogenic lineage.
4. Discussion
The aim of our study was to investigate whether laser-etched 3-D microchannels could regulate hMSCs lineage differentiation. Ourhypothesis was that the femtosecond laser-formed channels, to-gether with their macro- and micropatterns, would influence cellalignment, which would in turn affect cell proliferation and differ-entiation. We observed that hMSCs were able to align themselvesalong the direction of the microchannels. The morphological alter-ations and cytoskeleton reorganization caused by microtopogra-phy were consistent with previous observations in other celltypes [41–43]. Much more interestingly, we observed that morecells were found inside the microchannels (on the bottom or at-tached to the wall of microchannels) than on the intervals, espe-cially when it was not passivated. This might be due to thesurface modifications created by the laser machining. Immunoflu-orescent staining of fibronectin on the laser-patterned scaffoldswithout hMSCs after 12 h of incubation in culture medium with10% FBS is shown in Fig. S3. This biased fibronectin patterningmay explain why more hMSCs were found inside the laser-pat-terned microchannels compared to the intervals. Many aggregateddots of fibronectin (only some aggregates are shown by arrows)were clearly observed inside the laser-scanned microchannels(Fig. S3). On the other hand, fewer fibronectin dots were found

30-30-20 80-30-20
Ost
eo-M
arke
rsM
yo-M
arke
rsN
euro
-Mar
kers
Day 7 Day 11 Day 14
0123456
A B
D
FE
CALPL ON RUNX2
Nor
mal
ized
fold
exp
ress
ion
*
01234567
ALPL ON RUNX2
Nor
mal
ized
fold
exp
ress
ion
0
1
2
3
4
5
6
7
7
GATA4 MHC7 MyoD1 Nkx2.5
Nor
mal
ized
fold
exp
ress
ion
* * * * * *
* * *
*
01234567
GATA4 MHC7 MyoD1 Nkx2.5N
orm
aliz
ed fo
ld e
xpre
ssio
n
* **
01234567
MAP2 Nestin NeuroD1
Nor
mal
ized
fold
exp
ress
ion
* *
* *
* *
01234567
MAP2 Nestin NeuroD1
Nor
mal
ized
fold
exp
ress
ion
*
Fig. 5. Relative gene level expression analyses of osteogenic, myogenic and neurogenic markers on days 7, 11 and 14 of cell culture. Bars represent mean ± standard deviation.Relative gene expression levels (i.e. DDCT) were normalized to the control samples (t = 7 days time point) without laser machining. ⁄Statistical difference between patternedand unpatterned samples (p < 0.05). Left panels A, C and E: osteo-, myo- and neuromarkers, respectively, for 30–30–20; right panels B, D and F: osteo-, myo- andneuromarkers for 80–30–20.
Fig. 6. Representative images showing immunostaining of myogenic and neurogenic markers (C = channel, I = interval). Cells were co-stained with distinctive and maturemarkers of myogenesis and neurogenesis using mouse monoclonal anti-heavy chain cardiac myosin (green) and chicken polyclonal anti MAP2 (red) (scale bar, 50 lm).
H. Li et al. / Acta Biomaterialia 8 (2012) 531–539 537

538 H. Li et al. / Acta Biomaterialia 8 (2012) 531–539
on the intervals; this may be due to the fact that the roughness ofthe channel is much greater than that of the interval. Following thesame logic, proteins from the culture medium will adsorb onto thesurface and act as cell adhesion nodes [44]. Consequently, thischanges the cell distribution between the intervals and channels,as well as that in the controls. To exclude the possibility that thechemical changes of the polymer surface after laser machininghad caused the differential attachment pattern, we used ATR-FTIRand XPS spectra to show that there were no notable differences inthe functional groups, and the chemical states of carbon and oxy-gen, between the laser-machined substrates and the control group.It can be concluded that the surface topography created by the la-ser machining is the dominant factor affecting protein adsorptionand the subsequent cell adhesion.
Many studies have been performed to investigate the effects ofsurface topography features on cell attachment, spreading and pro-liferation [45–47]. It was reported that nanoroughness plays a rolein regulating cell physiology, which is perhaps not surprising, sincecells are surrounded by nanoscale structures in their native envi-ronment [48]. One study demonstrated that the levels of short-term cell attachment were higher on rough surfaces compared tosmooth surfaces, and cell morphology was directly related to thenature of the underlying substrate [49]. Another in vitro studydemonstrated that osteoblasts exhibited greater initial attachmentto rough titanium surfaces [50]. On the other hand, it is known thatmicrotopographical features may impede cell migration and prolif-eration due to physical constrains [51]. When compared to thescaffolds with wide microchannels, the scaffolds with narrowmicrochannels had a similar surface roughness inside the channelsbut showed greater impedance to cell proliferation in the perpen-dicular direction of the channels, due to the narrower channelwidth.
It is well known that cell morphology alteration, cytoskeletonorganization and nuclear shape variation all play a critical role insignal transduction [11]. Macroscopic changing of the cell shapereveals that cells are able to sense micro- and nanoscale cues fromthe environment, such as surface topography or roughness. Thetopographical features used in cell culture are mainly basic shapes(e.g. lines, pillars or wells), among which line-shaped structureswere commonly used to align cells and influence cellular behavior.It is suggested that the mechanical tension caused deformation ofthe nucleus, which is physically linked to the cytoskeleton by inter-mediate filaments, thus altering the gene expression and subse-quently the cellular behavior [37,52]. In this reported work, afemtosecond laser was used to fabricate a system of micropatternsconsisting of a continuous sinusoidal surface topography andmicroroughness, both of which proved to enhance cell adhesionand defined cell orientation. Compared to conventional 2-D cellculture systems, our laser-machined scaffolds could partly providea pseudo-3-D environment that is essential for the maintenance ofcellular function and tissue formation in vivo.
The difference between the two laser-machined scaffolds is thesurface curvature produced inside the microchannels, which hasbeen shown to affect cell morphology and then tissue growth[53,54]. Cells cultured on these scaffolds have consistent morphol-ogies, with cells in the 30 lm width microchannels taking on amore elongated shape and appearing relatively smaller than thosein the 80 lm microchannels. This could be caused by the physicalconstraint induced by the narrower channels in the perpendiculardirection, which have relatively shorter radii of curvature. Thus,cells in the narrow channels would elongate more in the directionof the microchannels, leading to cell alignment. In contrast, thehMSCs grown on the control substrates grew in random directions,and were 30–50 lm wide and more than 100 lm long. Therefore,microchannels induce cell elongation by generating cytoskeletaltension, and this may direct myogenic differentiation [37,5].
The narrower 30 lm microchannels with smaller radii of curva-ture would give rise to a higher intracellular tension force than thewider 80 lm microchannels. This high intracellular tension recruitsa-smooth muscle actin to stress fibers [56,57]. Micropatterning ofeven narrower 20 lm wide fibronectin strips on poly(lactic-co-gly-colic acid) film could induce hMSCs towards the myocardial lineage[37,55]. The micropatterned matrix imposed cell shape restrictionsand led to cytoskeletal rearrangement and nucleus shape alter-ation; these changes translated into a biochemical response and ef-fected cell differentiation [37]. Compared to the 2-D surfaces, thelaser-machined 3-D scaffolds were able to increase the extent ofmyogenic gene up-regulation in the cells (MyoD1 gene up-regula-tion �5.1- vs. �3.4-fold; GATA4 gene up-regulation �4.1- vs.�3.0-fold). Since cell proliferation and differentiation are knownto have an inverse relationship [58], the directed myogenic differ-entiation of hMSCs might be a possible reason for the observationof lower cell numbers in laser-micropatterned scaffolds comparedto the control. However, it is believed that the higher surface areacreated by the microchannels could compensate for this slowerproliferation if sufficient time is given.
5. Conclusions
The results presented here show that laser-cut microchannels,together with the 3-D topographic micropatterns on the polymersubstrate, affects cell adhesion, proliferation, cytoskeleton reorga-nization and differentiation. The novelty of this work lies in theapplication of direct laser machining to create channels with 3-Dtopographic micropatterns that can coax differentiation of hMSCstowards a defined lineage without any exogeneous biochemicalfactors. Due to the biocompatibility and biodegradable propertiesof the PLLA–PCL polymer [59], combined with the availability ofhMSCs and their differentiation, this new strategy is promisingfor future tissue engineering applications. Further studies on themolecular level to check the signaling pathways behind the obser-vation in this study are in progress.
Acknowledgements
The authors would like to thank the National Research Founda-tion (Grant No. NRF-CRP2-2007-1) and Singapore Stem Cell Con-sortium (SSCC) (Grant No. SSCC/09/017) for their financial support.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.actbio.2011.09.029.
Appendix B. Figures with essential colour discrimination
Certain figures in this article, particularly Figures 1, 3, and 6, aredifficult to interpret in black and white. The full colour images canbe found in the on-line version, at doi:10.1016/j.actbio.2011.09.029.
References
[1] Vacanti JP, Langer R. Tissue engineering: the design and fabrication of livingreplacement devices for surgical reconstruction and transplantation. Lancet1999;354(Suppl. 1):S132–4.
[2] Hwang NS, Varghese S, Elisseeff J. Controlled differentiation of stem cells. AdvDrug Delivery Rev 2008;60:199–214.
[3] Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al.Multilineage potential of adult human mesenchymal stem cells. Science1999;284:143–7.
[4] Song H, Song B-W, Cha M-J, Choi I-G, Hwang K-C. Modification ofmesenchymal stem cells for cardiac regeneration. Expert Opin Biol Ther2010;10:309–19.

H. Li et al. / Acta Biomaterialia 8 (2012) 531–539 539
[5] Lund AW, Yener B, Stegemann JP, Plopper GE. The natural and engineered 3-Dmicroenvironment as a regulatory cue during stem cell fate determination.Tissue Eng Part B: Rev 2009;15:371–80.
[6] Ross AM, Jason AB. Controlling stem cell fate with material design. Adv Mater(Weinheim, Germany) 2010;22:175–89.
[7] Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem celllineage specification. Cell 2006;126:677–89.
[8] Dalby MJ, Gadegaard N, Tare R, Andar A, Riehle MO, Herzyk P, et al. The controlof human mesenchymal cell differentiation using nanoscale symmetry anddisorder. Nat Mater 2007;6:997–1003.
[9] Guilak F, Cohen DM, Estes BT, Gimble JM, Liedtke W, Chen CS. Control of stemcell fate by physical interactions with the extracellular matrix. Cell Stem Cell2009;5:17–26.
[10] Kulangara K, Leong KW. Substrate topography shapes cell function. Soft Matter2009;5:4072–6.
[11] Martínez E, Lagunas A, Mills CA, Rodríguez-Seguí S, Estévez M, Oberhansl S,et al. Stem cell differentiation by functionalized micro- and nanostructuredsurfaces. Nanomed 2009;4:65–82.
[12] Dang J, Leong K. Myogenic induction of aligned mesenchymal stem cell sheetsby culture on thermally responsive electrospun nanofibers. Adv Mater(Weinheim, Germany) 2007;19:2775–9.
[13] Yim EKF, Pang SW, Leong KW. Synthetic nanostructures inducingdifferentiation of human mesenchymal stem cells into neuronal lineage. ExpCell Res 2007;313:1820–9.
[14] Katayama S, Horiike M, Hirao K, Tsutsumi N. Structure induced by irradiationof femtosecond laser pulse in dyed polymeric materials. J Polym Sci Part B:Polym Phys 2002;40:2800–6.
[15] Baudach S, Bonse J, Kautek W. Ablation experiments on polyimide withfemtosecond laser pulses. Appl Phys A: Mater Sci Process 1999;69:S395–8.
[16] Tiaw KS, Goh SW, Hong M, Wang Z, Lan B, Teoh SH. Laser surface modificationof poly(e-caprolactone) (PCL) membrane for tissue engineering applications.Biomaterials 2005;26:763–9.
[17] Katayama S, Horiike M, Hirao K, Tsutsumi N. Structures induced by irradiationof femto-second laser pulse in polymeric materials. J Polym Sci Part B: PolymPhys 2002;40:537–44.
[18] Wochnowski C, Cheng Y, Meteva K, Sugioka K, Midorikawa K, Metev S.Femtosecond-laser induced formation of grating structures in planar polymersubstrates. J Opt A: Pure Appl Opt 2005;7:493–501.
[19] Tiaw KS, Goh SW, Hong M, Wang Z, Lan B, Teoh SH. Laser surface modificationof poly(epsilon-caprolactone) (PCL) membrane for tissue engineeringapplications. Biomaterials 2005;26:763–9.
[20] Bergmeister H, Boeck P, Kasimir M-T, Fleck T, Fitzal F, Husinsky W, et al. Effectof laser perforation on the remodeling of acellular matrix grafts. J BiomedMater Res B Appl Biomater 2005;74B:495–503.
[21] Liu Y, Sun S, Singha S, Cho MR, Gordon RJ. 3D femtosecond laser patterning ofcollagen for directed cell attachment. Biomaterials 2005;26:4597–605.
[22] Aguilar CA, Lu Y, Mao S, Chen SC. Direct micro-patterning of biodegradablepolymers using ultraviolet and femtosecond lasers. Biomaterials2005;26:7642–9.
[23] Yeong WY, Yu H, Lim KP, Ng KLG, Boey YCF, Subbu VS, et al. Multiscaletopological guidance for cell alignment via direct laser writing onbiodegradable polymer. Tissue Eng Part C: Methods 2010;16:1011–21.
[24] Spandidos A, Wang X, Wang H, Seed B. PrimerBank: a resource of human andmouse PCR primer pairs for gene expression detection and quantification.Nucleic Acids Res 2010;38:D792–9.
[25] Pfaffl MW, Horgan GW, Dempfle L. Relative expression software tool (REST�)for group-wise comparison and statistical analysis of relative expressionresults in real-time PCR. Nucleic Acids Res 2002;30:E36.
[26] Kweon H, Yoo MK, Park IK, Kim TH, Lee HC, Lee H-S, et al. A novel degradablepolycaprolactone networks for tissue engineering. Biomaterials2003;24:801–8.
[27] Prabhakaran MP, Venugopal JR, Ramakrishna S. Mesenchymal stem celldifferentiation to neuronal cells on electrospun nanofibrous substrates fornerve tissue engineering. Biomaterials 2009;30:4996–5003.
[28] Zhu Y, Chian KS, Chan-Park MB, Mhaisalkar PS, Ratner BD. Protein bonding onbiodegradable poly(L-lactide-co-caprolactone) membrane for esophagealtissue engineering. Biomaterials 2006;27:68–78.
[29] Weiss MJ, Henthorn PS, Lafferty MA, Slaughter C, Raducha M, Harris H.Isolation and characterization of a cDNA encoding a human liver/bone/kidney-type alkaline phosphatase. Proc Natl Acad Sci USA 1986;83:7182–6.
[30] Leong DT, Khor WM, Chew FT, Lim TC, Hutmacher DW. Characterization ofosteogenically induced adipose tissue-derived precursor cells in 2-dimensionaland 3-dimensional environments. Cells Tissues Organs 2006;182:1–11.
[31] Termine JD, Kleinman HK, Whitson SW, Conn KM, McGarvey ML, Martin GR.Osteonectin, a bone-specific protein linking mineral to collagen. Cell1981;26:99–105.
[32] Schroeder TM, Jensen ED, Westendorf JJ. Runx2: a master organizer of genetranscription in developing and maturing osteoblasts. Birth Defects Res Part C2005;75:213–25.
[33] Pereira BP, Zhou Y, Gupta A, Leong DT, Aung KZ, Ling L, et al. Runx2, p53, andpRB status as diagnostic parameters for deregulation of osteoblast growth anddifferentiation in a new pre-chemotherapeutic osteosarcoma cell line (OS1). JCell Physiol 2009;221:778–88.
[34] Kong Y, Flick MJ, Kudla AJ, Konieczny SF. Muscle LIM protein promotesmyogenesis by enhancing the activity of MyoD. Mol Cell Biol1997;17:4750–60.
[35] Durocher D, Charron F, Warren R, Schwartz RJ, Nemer M. The cardiactranscription factors Nkx2-5 and GATA-4 are mutual cofactors. EMBO J1997;16:5687–96.
[36] Thierfelder L, Watkins H, MacRae C, Lamas R, McKenna W, Vosberg H-P, et al.[alpha]-Tropomyosin and cardiac troponin T mutations cause familialhypertrophic cardiomyopathy: a disease of the sarcomere. Cell1994;77:701–12.
[37] Tay CY, Yu H, Pal M, Leong WS, Tan NS, Ng KW, et al. Micropatterned matrixdirects differentiation of human mesenchymal stem cells towards myocardiallineage. Exp Cell Res 2010;316:1159–68.
[38] Dahlstrand J, Zimmerman LB, McKay RD, Lendahl U. Characterization of thehuman nestin gene reveals a close evolutionary relationship toneurofilaments. J Cell Sci 1992;103:589–97.
[39] Gao Z, Ure K, Ables JL, Lagace DC, Nave K-A, Goebbels S, et al. Neurod1 isessential for the survival and maturation of adult-born neurons. Nat Neurosci2009;12:1090–2.
[40] Dehmelt L, Halpain S. Actin and microtubules in neurite initiation: are MAPsthe missing link? J Neurobiol 2004;58:18–33.
[41] Teixeira AI, Abrams GA, Bertics PJ, Murphy CJ, Nealey PF. Epithelial contactguidance on well-defined micro- and nanostructured substrates. J Cell Sci2003;116:1881–92.
[42] Webb A, Clark P, Skepper J, Compston A, Wood A. Guidance ofoligodendrocytes and their progenitors by substratum topography. J Cell Sci1995;108(Pt 8):2747–60.
[43] Flemming RG, Murphy CJ, Abrams GA, Goodman SL, Nealey PF. Effects ofsynthetic micro- and nano-structured surfaces on cell behavior. Biomaterials1999;20:573–88.
[44] Oh S, Brammer KS, Li YSJ, Teng D, Engler AJ, Chien S, et al. Stem cell fatedictated solely by altered nanotube dimension. Proc Natl Acad Sci USA2009;106:2130–5.
[45] Wan Y, Wang Y, Liu Z, Qu X, Han B, Bei J, et al. Adhesion and proliferation ofOCT-1 osteoblast-like cells on micro- and nano-scale topography structuredpoly(L-lactide). Biomaterials 2005;26:4453–9.
[46] Teixeira AI, Nealey PF, Murphy CJ. Responses of human keratocytes to micro-and nanostructured substrates. J Biomed Mater Res Part A 2004;71:369–76.
[47] Teixeira AI, McKie GA, Foley JD, Bertics PJ, Nealey PF, Murphy CJ. The effect ofenvironmental factors on the response of human corneal epithelial cells tonanoscale substrate topography. Biomaterials 2006;27:3945–54.
[48] Rebollar E, Frischauf I, Olbrich M, Peterbauer T, Hering S, Preiner J, et al.Proliferation of aligned mammalian cells on laser-nanostructured polystyrene.Biomaterials 2008;29:1796–806.
[49] Bowers KT, Keller JC, Randolph BA, Wick DG, Michaels CM. Optimization ofsurface micromorphology for enhanced osteoblast responses in vitro. Int J OralMaxillofac Implants 1992;7:302–10.
[50] Boyan BD, Batzer R, Kieswetter K, Liu Y, Cochran DL, Szmuckler-Moncler S,et al. Titanium surface roughness alters responsiveness of MG63 osteoblast-like cells to 1a,25-(OH)2D3. J Biomed Mater Res 1998;39:77–85.
[51] Lutolf MP, Gilbert PM, Blau HM. Designing materials to direct stem-cell fate.Nature 2009;462:433–41.
[52] Maniotis AJ, Chen CS, Ingber DE. Demonstration of mechanical connectionsbetween integrins, cytoskeletal filaments, and nucleoplasm that stabilizenuclear structure. Proc Natl Acad Sci USA 1997;94:849–54.
[53] Rumpler M, Woesz A, Dunlop JWC, van Dongen JT, Fratzl P. The effect ofgeometry on three-dimensional tissue growth. J Royal Soc Interface2008;5:1173–80.
[54] Nelson CM. Geometric control of tissue morphogenesis. Biochim Biophys ActaMol Cell Res 2009;1793:903–10.
[55] Tay CY, Pal M, Yu H, Leong WS, Tan NS, Ng KW, et al. Bio-inspiredmicropatterned platform to steer stem cell differentiation. Small2011;7:1416–21.
[56] Jakkaraju S, Zhe X, Schuger L. Role of stretch in activation of smooth musclecell lineage. Trends Cardiovasc Med 2003;13:330–5.
[57] Dugina V, Fontao L, Chaponnier C, Vasiliev J, Gabbiani G. Focal adhesionfeatures during myofibroblastic differentiation are controlled by intracellularand extracellular factors. J Cell Sci 2001;114:3285–96.
[58] Maione R, Amati P. Interdependence between muscle differentiation and cell-cycle control. Biochim Biophys Acta Rev Cancer 1997;1332:M19–30.
[59] Jeong SI, Kim B-S, Kang SW, Kwon JH, Lee YM, Kim SH, et al. In vivobiocompatibility and degradation behavior of elastic poly(L-lactide-co-e-caprolactone) scaffolds. Biomaterials 2004;25:5939–46.
Copyright © 2022 FDOKUMEN




















