
Dimer-based model for heptaspanning membrane receptors
69
Update | Research Focus 355 The membrane – water interface region of membrane proteins: structural bias and the anti-snorkeling effect Jie Liang, Larisa Adamian and Ronald Jackups Jr 357 Testis-specific histone H3 expression in somatic cells Je ´ro ˆ me Govin, Ce ´ cile Caron, Sophie Rousseaux and Saadi Khochbin Opinion 360 Dimer-based model for heptaspanning membrane receptors Rafael Franco, Vicent Casado ´ , Josefa Mallol, Sergi Ferre ´ , Kjell Fuxe, Antonio Corte ´ s, Francisco Ciruela, Carmen Lluis and Enric I. Canela 367 SNARE complexes and neuroexocytosis: how many, how close? Cesare Montecucco, Giampietro Schiavo and Sergio Pantano 373 Ultradian metronome: timekeeper for orchestration of cellular coherence David Lloyd and Douglas B. Murray 378 CIF and other mysteries of the store-operated Ca 21 -entry pathway Victoria M. Bolotina and Peter Csutora Review 388 Major cutbacks at chromosome ends Peter M. Lansdorp 396 Bridging gaps in phospholipid transport Dennis R. Voelker 405 Eukaryotic transcription factors as direct nutrient sensors Christopher A. Sellick and Richard J. Reece 413 Shedding light on ADAM metalloproteinases Ari-Pekka J. Huovila, Anthony J. Turner, Markku Pelto-Huikko, Iivari Ka ¨ rkka ¨ inen and Rebekka M. Ortiz TRENDS in July 2005 Vol. 30, No. 7 pp. 355–422 Biochemical Sciences Editor Fiona G. Hutton Assistant Editor Vicky Ashton Editorial Coordinator Joanna Pinto Illustrations The Studio Cover Design Geraldine Woods Publishing Manager O. Claire Moulton Editorial Enquiries Trends in Biochemical Sciences Elsevier, 84 Theobald’s Road, London, UK WC1X 8RR tel: +44 (0)20 7611 4400 fax: +44 (0)20 7611 4470 e-mail: [email protected] Subscription Enquiries E-mail: [email protected] Advisory Editorial Board J. Witkowski (Editor in Chief), Cold Spring Harbor Laboratory, USA M. Bienz, MRC, Cambridge, UK S. Cockcroft, University College London, UK J. Dixon, University of California, USA T. Gibson, EMBL, Germany C-H. Heldin, Ludwig Institute for Cancer Research, Sweden M. Hentze, EMBL, Germany R. Kornberg, Stanford University, USA T. de Lange, Rockefeller University, USA A. Lamond, University of Dundee, UK G. Petsko, Brandeis University, USA T. Pollard, Yale University, USA D.W. Russell, UT Southwestern Medical Center, USA P. Schimmel, The Scripps Research Institute, USA P.H. von Hippel, University of Oregon, USA M. Yanagida, Kyoto University, Japan Forthcoming articles Pumps, paradoxes and ploughshares: mechanism of the MCM2–7 DNA helicase T.S. Takahashi, D.B. Wigley and J.C. Walter Small non-coding RNAs as magic bullets F. Eckstein Ironing out the problem: new mechanisms of iron homeostasis E. Masse ´ and M. Arguin The spliceosome: a novel multi-faceted target for therapy J. Tazi, S. Durand and P. Jeanteur Oxidative activation of antioxidant defence P.G. Winyard, C.J. Moody and C. Jacob Detergent-resistant membranes should not be identified with membrane rafts D. Lichtenberg, F.M. Gon ˜ i and H. Heerklotz HIF-1a and p53: the ODD couple? D.R. Fels and C. Koumenis Scaffold proteins dictate Rho GTPase-signaling specificity M.J. Marinissen and J.S. Gutkind Cover: In a recent study, the membrane–water interface region was examined in detail for the first time using computational analysis, and the results revealed that this interface region has an important role in constraining protein secondary structure. On pages 355– 357, Jie Liang, Larisa Adamian and Ronald Jackups Jr highlight recent work demonstrating that, in the interface region, the side chains of tryptophan and tyrosine reverse orientation and tend to point towards the membrane core, thus changing from snorkeling to anti-snorkeling. www.iubmb.org
Transcript of Dimer-based model for heptaspanning membrane receptors

Update
|Research Focus
355 The membrane–water interface region ofmembrane proteins: structural bias and theanti-snorkeling effectJie Liang, Larisa Adamian and Ronald Jackups Jr
357 Testis-specific histone H3 expression insomatic cellsJerome Govin, Cecile Caron, Sophie Rousseaux and
Saadi Khochbin
Opinion
360 Dimer-based model for heptaspanning membrane receptorsRafael Franco, Vicent Casado, Josefa Mallol, Sergi Ferre, Kjell Fuxe, Antonio Cortes,
Francisco Ciruela, Carmen Lluis and Enric I. Canela
367 SNARE complexes and neuroexocytosis: how many, how close?Cesare Montecucco, Giampietro Schiavo and Sergio Pantano
373 Ultradian metronome: timekeeper for orchestration of cellular coherenceDavid Lloyd and Douglas B. Murray
378 CIF and other mysteries of the store-operated Ca21-entry pathwayVictoria M. Bolotina and Peter Csutora
Review
388 Major cutbacks at chromosome endsPeter M. Lansdorp
396 Bridging gaps in phospholipid transportDennis R. Voelker
405 Eukaryotic transcription factors as direct nutrient sensorsChristopher A. Sellick and Richard J. Reece
413 Shedding light on ADAM metalloproteinasesAri-Pekka J. Huovila, Anthony J. Turner, Markku Pelto-Huikko, Iivari Karkkainen and
Rebekka M. Ortiz
TRENDSin
July 2005
Vol. 30, No. 7
pp. 355–422
BiochemicalSciences
Editor Fiona G. Hutton
Assistant Editor Vicky Ashton
Editorial Coordinator Joanna Pinto
Illustrations The Studio
Cover Design Geraldine Woods
Publishing Manager O. Claire Moulton
Editorial Enquiries
Trends in Biochemical Sciences
Elsevier,
84 Theobald’s Road,
London, UK WC1X 8RR
tel: +44 (0)20 7611 4400
fax: +44 (0)20 7611 4470
e-mail: [email protected]
Subscription Enquiries
E-mail: [email protected]
Advisory Editorial Board
J. Witkowski (Editor in Chief), Cold
Spring Harbor Laboratory, USA
M. Bienz, MRC, Cambridge, UK
S. Cockcroft, University College London,
UK
J. Dixon, University of California, USA
T. Gibson, EMBL, Germany
C-H. Heldin, Ludwig Institute for Cancer
Research, Sweden
M. Hentze, EMBL, Germany
R. Kornberg, Stanford University, USA
T. de Lange, Rockefeller University, USA
A. Lamond, University of Dundee, UK
G. Petsko, Brandeis University, USA
T. Pollard, Yale University, USA
D.W. Russell, UT Southwestern Medical
Center, USA
P. Schimmel, The Scripps Research
Institute, USA
P.H. von Hippel, University of Oregon,
USA
M. Yanagida, Kyoto University, Japan
Forthcoming articles
Pumps, paradoxes and ploughshares: mechanism of the MCM2–7 DNA helicaseT.S. Takahashi, D.B. Wigley and J.C. Walter
Small non-coding RNAs as magic bulletsF. Eckstein
Ironing out the problem: new mechanisms of iron homeostasisE. Masse and M. Arguin
The spliceosome: a novel multi-faceted target for therapyJ. Tazi, S. Durand and P. Jeanteur
Oxidative activation of antioxidant defenceP.G. Winyard, C.J. Moody and C. Jacob
Detergent-resistant membranes should not be identified with membrane raftsD. Lichtenberg, F.M. Goni and H. Heerklotz
HIF-1a and p53: the ODD couple?D.R. Fels and C. Koumenis
Scaffold proteins dictate Rho GTPase-signaling specificityM.J. Marinissen and J.S. Gutkind
Cover: In a recent study, the membrane–water interface region was examined in detail for the first time using computational analysis,
and the results revealed that this interface region has an important role in constraining protein secondary structure. On pages 355–
357, Jie Liang, Larisa Adamian and Ronald Jackups Jr highlight recent work demonstrating that, in the interface region, the side
chains of tryptophan and tyrosine reverse orientation and tend to point towards the membrane core, thus changing from snorkeling
to anti-snorkeling.
www.iubmb.org

Research Focus
The membrane–water interface region of membraneproteins: structural bias and the anti-snorkeling effect
Jie Liang, Larisa Adamian and Ronald Jackups Jr
Department of Bioengineering, University of Illinois at Chicago, M/C563, 835 S. Wolcott Avenue, Chicago, IL 60612-7340, USA
Membrane proteins have important roles in many
cellular processes. Computational analysis of their
sequences and structures has provided much insight
into the organizing principles of transmembrane helices.
In a recent study, the membrane–water interface region
was examined in detail for the first time. The results
have revealed that this interface region has an important
role in constraining protein secondary structure. This
study raises new questions and opens up new directions
for studying membrane proteins.
Insights from computational analysis
Membrane proteins are abundant in most species andhave important roles, including signal transduction,proton pumping, cell trafficking and photosynthesis.Understanding their structural organization and theprinciples governing their folding and assembly is animportant task of biochemistry.
Computational analysis of membrane protein sequenceshas revealed fundamental insights. For instance, thesuccess of the prediction of transmembrane (TM) helicesfrom sequence hydropathy plots contributed to theformulation of the classic two-stage model of membrane-protein folding [1]. In addition, the observation of anasymmetric distribution of ionizable residues led to thediscovery of the ‘positive-inside’ rule [2], by which arginineand lysine are four times more abundant in thecytoplasmic segments of membrane proteins than in theextracellular segments. Further insights were gainedfrom analysis of the distribution [3] and sequence motifs[4] of amino acid types in the TM region.
Analysis of rapidly accumulating membrane-proteinstructures has been similarly fruitful. One example is thediscovery of aromatic girdles, namely, the two belt regionsof TM domains in which tryptophan and tyrosine arelocated in high proportions [5]. Studies of interactinghelices have revealed the important roles of regularhydrogen-bond, weak hydrogen-bond and packing inter-actions in helical assembly [6–8], confirming earlierpioneering experimental studies [9,10]. Recent analysisof membrane-protein structures continues to revealinsights about TM helices, such as side-chain preferencesand snorkeling effects [11,12].
Remarkably, results from computational analyses arelargely in good agreement with experimental data. Forexample, the stabilities of amino acids in lipids inferred by
Corresponding author: Liang, J. ([email protected]).Available online 2 June 2005
www.sciencedirect.com
computational analysis are consistent with experimentalstudies [13,14]. Recently, the sequence code for inserting apeptide into target membranes via the translocationmachinery was deciphered [15]. In this study, themeasured biological, physico-chemical and hydrophobicityscales all agree with each other, providing vital evidencethat direct protein–lipid interactions are crucial fortranslocon-mediated membrane insertion.
The membrane–water interface region
Much has been learned about the TM region, but little isknown about other regions of membrane proteins. Theinterface between the membrane and the aqueous solventis a special boundary region that has different physico-chemical properties compared with either the lipid (TMhelical) region or the bulk solvent. The crucial steps ofmembrane insertion occur here. What constraints doesthis region impose on the structure of membrane proteins?A recent study by Granseth et al. [16] is the first thatbrings this important region to the forefront ofinvestigation.
Secondary structures in the interface region
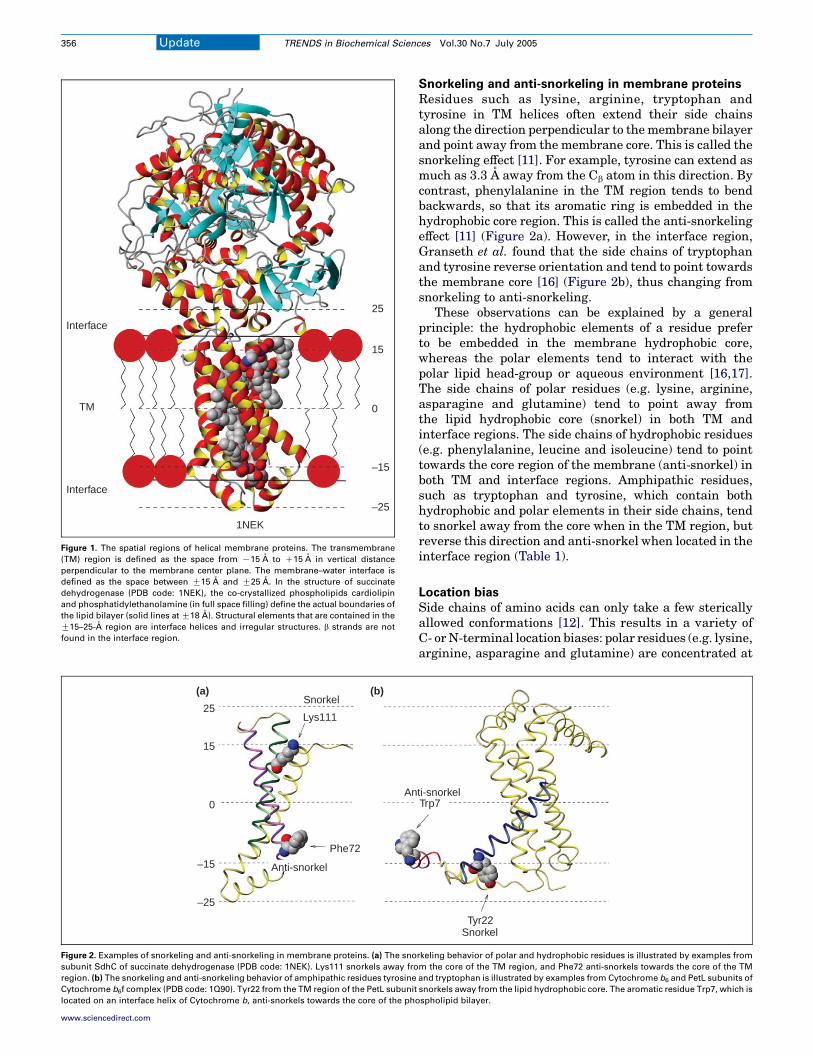
The membrane–water interface region can be defined bythe distance from the center of the membrane. Taking theregion that is G15–25 A from the center, Granseth et al.[16] analyzed 27 non-homologous protein structurescontaining 221 TM helices in total (Figure 1).
One of the main findings of Granseth et al. [16] is thatthe membrane–water interface region is dominated byirregular structures (w70%) and helices (w30%), butlacks b strands. The irregular structures are enrichedwith glycine and proline residues, which are well-knownturn promoters and helix breakers.
In most cases, interface helices are connected to TMhelices. Both types of helices are enriched with hydro-phobic residues, but interface helices have a much highercontent of polar aromatic residues (tryptophan andtyrosine). Frequently, a long peptide loop (O15 residues)connecting two TM helices contains an interface helix.There is little correlation between the end-to-end physicaldistance of two TM helices and the length of theconnecting loop. This suggests that interface helices helpto maintain the relative positions of TM helix ends whileaccommodating a large number of residues betweenhelices.
Update TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
Interface
TM
Interface
25
15
–25
0
–15
1NEK
Figure 1. The spatial regions of helical membrane proteins. The transmembrane
(TM) region is defined as the space from K15 A to C15 A in vertical distance
perpendicular to the membrane center plane. The membrane–water interface is
defined as the space between G15 A and G25 A. In the structure of succinate
dehydrogenase (PDB code: 1NEK), the co-crystallized phospholipids cardiolipin
and phosphatidylethanolamine (in full space filling) define the actual boundaries of
the lipid bilayer (solid lines at G18 A). Structural elements that are contained in the
G15–25-A region are interface helices and irregular structures. b strands are not
found in the interface region.
25
15
0
–15
–25
Snorkel
Lys111
Phe72
Anti-snorkel
An
(a) (b)
Figure 2. Examples of snorkeling and anti-snorkeling in membrane proteins. (a) The sno
subunit SdhC of succinate dehydrogenase (PDB code: 1NEK). Lys111 snorkels away fro
region. (b) The snorkeling and anti-snorkeling behavior of amphipathic residues tyrosine
Cytochrome b6f complex (PDB code: 1Q90). Tyr22 from the TM region of the PetL subunit
located on an interface helix of Cytochrome b, anti-snorkels towards the core of the ph
Update TRENDS in Biochemical Sciences Vol.30 No.7 July 2005356
www.sciencedirect.com
Snorkeling and anti-snorkeling in membrane proteins
Residues such as lysine, arginine, tryptophan andtyrosine in TM helices often extend their side chainsalong the direction perpendicular to themembrane bilayerand point away from the membrane core. This is called thesnorkeling effect [11]. For example, tyrosine can extend asmuch as 3.3 A away from the Cb atom in this direction. Bycontrast, phenylalanine in the TM region tends to bendbackwards, so that its aromatic ring is embedded in thehydrophobic core region. This is called the anti-snorkelingeffect [11] (Figure 2a). However, in the interface region,Granseth et al. found that the side chains of tryptophanand tyrosine reverse orientation and tend to point towardsthe membrane core [16] (Figure 2b), thus changing fromsnorkeling to anti-snorkeling.
These observations can be explained by a generalprinciple: the hydrophobic elements of a residue preferto be embedded in the membrane hydrophobic core,whereas the polar elements tend to interact with thepolar lipid head-group or aqueous environment [16,17].The side chains of polar residues (e.g. lysine, arginine,asparagine and glutamine) tend to point away fromthe lipid hydrophobic core (snorkel) in both TM andinterface regions. The side chains of hydrophobic residues(e.g. phenylalanine, leucine and isoleucine) tend to pointtowards the core region of the membrane (anti-snorkel) inboth TM and interface regions. Amphipathic residues,such as tryptophan and tyrosine, which contain bothhydrophobic and polar elements in their side chains, tendto snorkel away from the core when in the TM region, butreverse this direction and anti-snorkel when located in theinterface region (Table 1).
Location bias
Side chains of amino acids can only take a few stericallyallowed conformations [12]. This results in a variety ofC- orN-terminal location biases: polar residues (e.g. lysine,arginine, asparagine and glutamine) are concentrated at
SnorkelTyr22
Trp7ti-snorkel
rkeling behavior of polar and hydrophobic residues is illustrated by examples from
m the core of the TM region, and Phe72 anti-snorkels towards the core of the TM
and tryptophan is illustrated by examples from Cytochrome b6 and PetL subunits of
snorkels away from the lipid hydrophobic core. The aromatic residue Trp7, which is
ospholipid bilayer.

Table 1. Summary of the snorkeling and anti-snorkeling
behavior of different residues in the TM and interface regions
Region Polar Hydrophobic Amphopathic
Transmembrane
region
Snorkel Anti-snorkel Snorkel
Interface region Snorkel Anti-snorkel Anti-snorkel
Update TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 357
the N termini of helices, whereas hydrophobic residues(e.g. alanine, valine and isoleucine) and tyrosine areconcentrated at the C termini. Because residue side chainstend to point towards the N termini in a helices, thismakes N-terminal locations more favorable for polarresidues to snorkel. Tyrosine has a special rotamer toenable its hydroxyl group to extend further from the coreregion, hence, tyrosine is more favored at the C termini.The biased locations of residues can be largely explainedby the available side-chain rotamers and the propensity tosnorkel.
Concluding remarks
The work of Granseth and colleagues opens up a new areafor the study of membrane-protein biochemistry. Now,with the well-defined interface region and a clear pictureof its constraints on structures of membrane proteins, wecan start to ask new questions. For example, do interfacehelices form only in the constrained environment of theregions bordering the membrane? If the sequences ofinterface helices are introduced into soluble proteins, willthey still form stable helical structures? Does the entropiceffect of end-to-end distances for loops connecting two TMhelices differ from that of soluble proteins? How can suchthermodynamic considerations help to suggest mutantsfor enhanced stability or dynamics?
Undoubtedly, future studies of the interface region willfacilitate understanding of the folding mechanisms ofmembrane proteins, and might lead to the development ofengineering principles for designing novel and fullyfunctional membrane proteins.
Corresponding author: Khochbin, S. ([email protected]).Available online 26 May 2005
www.sciencedirect.com
References
1 Popot, J.L. and Engelman, D.M. (2000) Helical membrane proteinfolding, stability, and evolution. Annu. Rev. Biochem. 69, 881–922
2 von Heijne, G. (1986) The distribution of positively charged residues inbacterial inner membrane proteins correlates with the trans-membrane topology. EMBO J. 5, 3021–3027
3 Arkin, I.T. and Brunger, A.T. (1998) Statistical analysis of predictedtransmembrane a-helices. Biochim. Biophys. Acta 1429, 113–128
4 Senes, A. et al. (2000) Statistical analysis of amino acid patterns intransmembrane helices: the GxxxG motif occurs frequently and inassociation with b-branched residues at neighboring positions. J. Mol.Biol. 296, 921–936
5 Weiss, M.S. et al. (1991) Molecular architecture and electrostaticproperties of a bacterial porin. Science 254, 1627–1630
6 Adamian, L. and Liang, J. (2002) Interhelical hydrogen bonds andspatial motifs in membrane proteins: polar clamps and serine zippers.Proteins 47, 209–218
7 Eilers, M. et al. (2002) Comparison of helix interaction in membraneand soluble a-bundle proteins. Biophys. J. 82, 2720–2736
8 Senes, A. et al. (2001) The Ca–H.O hydrogen bond: a determinant ofstability and specificity in transmembrane helix interactions. Proc.Natl. Acad. Sci. U. S. A. 98, 9056–9061
9 Choma, C. et al. (2000) Asparagine-mediated self-association of amodel transmembrane helix. Nat. Struct. Biol. 7, 161–166
10 Zhou, F.X. et al. (2000) Interhelical hydrogen bonding drives stronginteractions in membrane proteins. Nat. Struct. Biol. 7, 154–160
11 Chamberlain, A.K. et al. (2004) Snorkeling preferences foster anamino acid composition bias in transmembrane helices. J. Mol. Biol.339, 471–479
12 Chamberlain, A.K. and Bowie, J.U. (2004) Analysis of side-chainrotamers in transmembrane proteins. Biophys. J. 87, 3460–3469
13 Beuming, T. and Weinstein, H. (2004) A knowledge-based scale for theanalysis and prediction of buried and exposed faces of transmembranedomain proteins. Bioinformatics 20, 1822–1835
14 Adamian, L. et al. (2005) Empirical lipid propensities of aminoacid residues in multispan a helical membrane proteins. Proteins59, 496–509
15 Hessa, T. et al. (2005) Recognition of transmembrane helices by theendoplasmic reticulum translocon. Nature 433, 377–381
16 Granseth, E. et al. (2005) A study of the membrane-water interfaceregion of membrane proteins. J. Mol. Biol. 346, 377–385
17 White, S.H. and Wimley, W.C. (1999) Membrane protein folding andstability: physical principles. Annu. Rev. Biophys. Biomol. Struct. 28,319–365
0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tibs.2005.05.003
Testis-specific histone H3 expression in somatic cells
Jerome Govin, Cecile Caron, Sophie Rousseaux and Saadi Khochbin
INSERM U309 – Laboratoire de Biologie Moleculaire et Cellulaire de la Differenciation Equipe Chromatine et expression des genes
Institut Albert Bonniot Faculte de Medecine, Domaine de la Merci 38706 La Tronche Cedex, France
Histone variants functionally differentiate individual
nucleosomes and, hence, act as key regulators of
chromatin structure and function. Large-scale proteomic
projects are now valuable sources of histone-variant
discovery, showing, in particular, that somatic mammal-
ian cells express a larger panel of histone H3 variants
than previously thought, including testis-specific vari-
ants and as yet uncharacterized species. These data
also suggest a tight relationship between the complex-
ity of histone-variant expression and physiopathological
states of the cells.

Update TRENDS in Biochemical Sciences Vol.30 No.7 July 2005358
Actors of nucleosome differentiation
The basic repeating unit of the genome packaging struc-ture in eukaryotes is the nucleosome, itself composed ofan octamer of the four core histones H3, H4, H2A andH2B. A fifth histone, the linker histone or H1, directs theformation of a higher-order structure in the nucleosomalarray. In addition to their structural role, histones mightalso serve as indicators, signalling the nature of thepackaged DNA via their post-translational modifications[1]. Indeed, these modifications provide the complexitynecessary to regionally and functionally specify thenucleosomes. The functional diversity of nucleosomes isincreased by the incorporation of histone variants [2,3].The genes encoding most of the histones are organized inclusters and have evolved to couple histone synthesis toDNA replication [4]. However, nucleosome assembly canalso occur independently of DNA replication, involvinghistones with divergent sequences. These histone variantsare mostly encoded by solitary genes that escape theregulatory constraints of replication-dependent genes buthave acquired sensitivity to other signalling pathwayssuch as those controlling embryonic development and celldifferentiation [5]. Therefore, the regulatory circuits ofhistone gene expression, in addition to the primarysequence of the encoding genes, are both of highestimportance to achieve a specific chromatin organizationat the right time and place. Accordingly, some of thehistone variants are essentially expressed in a tissue-specific manner. The testis best exemplifies this situationin which a massive synthesis of histone variants accom-panies a dramatic genome reorganization, which takesplace in the maturing germ cells and is characterized bythe replacement of the majority of the histones bytransition proteins and protamines [6]. Almost all knownhistone variants, including a panel of testis-specifichistone members, are synthesized before and during theassembly of transition proteins and protamines (Figure 1).Although almost nothing is known about the structuraland functional properties of the testis-specific histonevariants, recent investigations indicate that at least some
H1
H3
H2A
H2B
His
tone
s
Variants
H1t, H1t2, HILS1
H3t
TH2A
TH2B, TSH2B, H2BFWT
Somatic cells Spermatogenic cells
H1.1-H1.5, H1°
H3.1, H3.2, H3.3,CENP-A
H2A.X, H2A.Z,macroH2A,H2Abbd
Figure 1. Spermatogenic cells express all the known histone variants. In addition to
histone variants expressed in somatic cells (blue), spermatogenic cells express a
panel of specific members (yellow) [6]. Known histone variants expressed in human
somatic and spermatogenic cells are listed. H1.1–H1.5 signifies histone H1 variants
number one to number five. Histone H2BFWT has been recently identified [15].
www.sciencedirect.com
members have a crucial role in nuclear condensation andgenome reorganization [7].
Expression of new histone H3 variants in somatic cells
Somatic mammalian cells are known to express only fourhistone H3 variants: H3.1 and H3.2 are synthesized in areplication-dependent manner, whereas H3.3 is constitu-tively expressed and is enriched in transcriptionally activechromatin regions, and CENP-A (centromere protein A) isspecifically associated with centromeres [2,3]. In additionto these variants, one humanH3 variant, H3t – consideredas testis-specific – has been reported [8] (Figure 2). Therecent revolution in protein identification techniques nowenables a comprehensive analysis of the proteome ofdistinct subcellular structures. Surprisingly, examinationof the list of histones detected by these approaches showsthat, at least in the case of H3, the number of variantsexpressed in somatic cells is larger than previouslythought. Indeed, the identification of 692 proteins presentin HeLa-cell nucleoli has shown that H3t is among thehistones [9]. Histone H3t is normally expressed in humanprimary spermatocytes, and the analysis of multiplehuman cell lines, including HeLa cells and normal tissues,by RNase-protection assays shows no evidence for itsexpression outside the testis [8]. There is no informationavailable on the expression of the H3t protein inspermatogenic cells or on its intra-nuclear localizations.Therefore, it would be interesting to know whether H3tspecifically locates in the nucleoli chromatin of HeLacells and, moreover, where it locates in the chromatinof meiotic and post-meiotic spermatogenic cells. It ispossible that, like H3.3, H3t marks functionally definedchromatin regions.
Large-scale quantitative proteome studies undertakento evaluate quantitative and qualitative changes in theproteome occurring during particular physiopathologicalprocesses also revealed the expression of as yet unknownhistone H3 variants. One such study, aiming to iden-tify proteins with concentrations that vary during theearly stages of apoptosis after induction of PUMA(p53 up-regulated modulator of apoptosis), has shownthat an uncharacterized human histone variant is down-regulated in this process [10]. Although no expressedsequence tag (EST) corresponding to the reported proteincould be found in the databanks, a single cDNA fromhuman testis encoding a related and, as yet, uncharacter-ized histone H3 could be identified (GenBank accessionnumber: BI460089), suggesting that it could also corre-spond to another testis-specific H3 variant. A second studythat compared the proteome of human hepatocellularcarcinoma with that of normal hepatocytes also revealedthe up-regulation of another unknown histone H3 variantin cancerous cells [11].
With completion of the various genome-sequencingprojects, one might expect an easy identification of allthe genes encoding histone variants. In fact, the analysisof available mammalian genomic sequences databanksreveals the existence of several putative histoneH3-encoding genes, dispersed in the genome of human,mouse and rat, harbouring sometimes very divergentsequences. However, in most cases, a search in the EST

- A R T K Q T A R K S T G G K A P R K Q L A T K A A R K S A P A T G G V K K P H - R Y R P G T V A L R E- A R T K Q T A R K S T G G K A P R K Q L A T K A A R K S A P A T G G V K K P H - R Y R P G T V A L R E- A R T K Q T A R K S T G G K A P R K Q L A T K A A R K S A P S T G G V K K P H - R Y R P G T V A L R E- A R T K Q T A R K S T G G K A P R K Q L A T K V A R K S A P A T G G V K K P H - R Y R P G T V A L R E
G P R R R S R K P E A P R R R S P S P T P T P G P S R R - G P S L G A S S H Q H S R R R Q G - - W L K E
I R R Y Q K S T E L L I R K L P F Q R L V R E I A Q D F K T - - D L R F Q S S A V M A L Q E A C E A Y LI R R Y Q K S T E L L I R K L P F Q R L V R E I A Q D F K T - - D L R F Q S S A V M A L Q E A S E A Y LI R R Y Q K S T E L L I R K L P F Q R L V R E I A Q D F K T - - D L R F Q S A A I G A L Q E A S E A Y LI R R Y Q K S T E L L I R K L P F Q R L M R E I A Q D F K T - - D L R F Q S S A V M A L Q E A C E S Y L
I R K L Q K S T H L L I R K L P F S R L A R E I C V K F T R G V D F N W Q A Q A L L A L Q E A A E A F L
V G L F E D T N L C A I H A K R V T I M P K D I Q L A R R I R G E R A - - -V G L F E D T N L C A I H A K R V T I M P K D I Q L A R R I R G E R A - - -V G L F E D T N L C A I H A K R V T I M P K D I Q L A R R I R G E R A - - -V G L F E D T N L C V I H A K R V T I M P K D I Q L A R R I R G E R A - - -
V H L F E D A Y L L T L H A G R V T L F P K D V Q L A R R I R G L E E G L G
H3.1H3.2H3.3H3t
CENP-A
H3.1H3.2H3.3H3t
CENP-A
H3.1H3.2H3.3H3t
CENP-A
Figure 2. Sequence analysis of mammalian histone H3 variants. The sequence of the four known H3 variants expressed in human somatic cells is compared to that of testis-
specific H3t variant. H3 peptides identified in HeLa cells nucleoli are outlined by black rectangles [13]. Amino acids are numbered based on the H3.1 sequence. GenBank
accession numbers: H3.1, AAN10051; H3.2, AAN39283; H3.3, P84243; H3t, NP_003484; CENP-A, P49450.
Update TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 359
databanks does not show any evidence for theirexpression. It is, therefore, difficult to determine if theseare functional genes. Proteomic data now suggest that atleast some of them are functional genes. The geneencoding H3.4 illustrates this situation well; indeed, agene encoding a H3 variant was identified in 1981 andlater named H3.4 with no evidence for its functionality[12]. Now a comprehensive proteomic analysis of thecomponents of the mid-body isolated from synchronizedCHO cells shows the presence of a H3.4-related histone H3in this structure [13]. These data show for the first timethat proteomic approaches might be much more powerfulthan the transcriptomic methods in the discovery offunctional genes that encode new histone variants.Indeed, some of the genes, such as those encoding testis-specific histone variants, might be expressed at very lowlevels in somatic cells, or might encode unstable mRNAs,or be expressed only in response to specific physiopatho-logical stimuli, resulting in an under-representation oftheir corresponding cDNAs in the EST databanks. Bycontrast, the putative encoded proteins, althoughexpressed at low levels, could accumulate in specificcellular compartments and therefore be identified byproteomic analyses.
Concluding remarks
These new findings show that a complex set of histoneH3 variants could permanently, or occasionally, beexpressed in various cell types and emphasize their rolein the functional differentiation of nucleosomes. It is alsoimportant to keep inmind that the increased complexity ofthe expression of histone H3 variants, mainly that oftestis-specific members, might primarily concern onco-genically transformed cells. Indeed, although the expres-sion of testis-specific histone variants in tumour cells hasnot been previously reported, it has been known for manyyears that cancer cells express a variety of testis-specificproteins [14]. Therefore, it would be worth considering thepossibility that, at an early stage of malignant transform-ation of a cell, an initial alteration of DNA methylationand chromatin structure might induce an illegitimateexpression of a variety of histone variants, mainly
testis-specific members, thereby triggering a more exten-sive modification of chromatin structure and function,which could, in turn, actively enhance cell transformation.
Acknowledgements
This work is supported by the ‘Action Concertee Incitative’ and ‘RegionRhone Alpes’ emergence programs.
References
1 Strahl, B.D. and Allis, C.D. (2000) The language of covalent histonemodifications. Nature 403, 41–45
2 Sarma, K. and Reinberg, D. (2005) Histone variants meet their match.Nat. Rev. Mol. Cell Biol. 6, 139–149
3 Henikoff, S. et al. (2004) Histone variants, nucleosome assembly andepigenetic inheritance. Trends Genet. 20, 320–326
4 Marzluff, W.F. et al. (2002) The human and mouse replication-dependent histone genes. Genomics 80, 487–498
5 Khochbin, S. (2001) Histone H1 diversity: bridging regulatory signalsto linker histone function. Gene 271, 1–12
6 Govin, J. et al. (2004) The role of histones in chromatin remodellingduring mammalian spermiogenesis. Eur. J. Biochem. 271, 3459–3469
7 Martianov, I. et al. (2005) Polar nuclear localization of H1T2, a histoneH1 variant, required for spermatid elongation and DNA condensationduring spermiogenesis. Proc. Natl. Acad. Sci. U. S. A. 102, 2808–2813
8 Witt, O. et al. (1996) Testis-specific expression of a novel human H3histone gene. Exp. Cell Res. 229, 301–306
9 Andersen, J.S. et al. (2005) Nucleolar proteome dynamics.Nature 433,77–83
10 Gu, S. et al. (2004) Large-scale quantitative proteomic study ofPUMA-induced apoptosis using two-dimensional liquid chromato-graphy-mass spectrometry coupled with amino acid-coded masstagging. J. Proteome Res. 3, 1191–1200
11 Li, C. et al. (2004) Accurate qualitative and quantitative proteomicanalysis of clinical hepatocellular carcinoma using laser capturemicrodissection coupled with isotope-coded affinity tag and two-dimensional liquid chromatography mass spectrometry. Mol. Cell.Proteomics 3, 399–409
12 Sittman, D.B. et al. (1981) Isolation of two clusters of mouse histonegenes. Proc. Natl. Acad. Sci. U. S. A. 78, 4078–4082
13 Skop, A.R. et al. (2004) Dissection of the mammalian midbodyproteome reveals conserved cytokinesis mechanisms. Science 305,61–66
14 Scanlan, M.J. et al. (2004) The cancer/testis genes: review, standard-ization, and commentary. Cancer Immun. 4, 1
15 Churikov, D. et al. (2004) Novel human testis-specific histone H2Bencoded by the interrupted gene on the X chromosome. Genomics 84,745–756
0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tibs.2005.05.001
www.sciencedirect.com

Dimer-based model for heptaspanningmembrane receptorsRafael Franco1, Vicent Casado1, Josefa Mallol1, Sergi Ferre2, Kjell Fuxe3,
Antonio Cortes1, Francisco Ciruela1, Carmen Lluis1 and Enric I. Canela1
1Department Bioquımica i Biologia Molecular, Universitat de Barcelona, A. Diagonal, 645. 08028 Barcelona, Spain2National Institute on Drug Abuse, NIH, DHHS, Baltimore, MD 21224, USA3Department of Neuroscience; Division of Cellular and Molecular Neurochemistry, Karolinska Institutet (KF), S-171 77 Stockholm,
Sweden
Glossary
Non-linear Scatchard plots: Scatchard plots are used to determine the number
of binding sites and the equilibrium constant (Kd) for agonist, antagonist or any
other ligand. A Scatchard plot is linear when one molecule of ligand binds to
onemolecule of the receptor or when the binding of one molecule of the ligand
has no effect on the affinity of the other molecules for the vacant sites of the
receptor (all the intrinsic affinity constants are identical). Non-linear Scatchard
plots for ligand binding are the general consequence of the binding of more
than one molecule of ligand to one molecule of receptor or when one ligand
binds to two separated or non-interconvertible forms of a receptor having
different affinity constant values.
Hill coefficient: An indicator of cooperativity. When the Hill coefficient is!1 the
cooperativity is negative; if the value is O1, the cooperativity is positive.
Occupational theory of drug action: This theory assumes that themagnitude of
the receptor-mediated effect is proportional to the concentration of the drug-
receptor complex.
F-test: The F-test is used to compare the variances (or standard deviations) of
The existence of intramembrane receptor–receptor
interactions for heptaspanning membrane receptors is
now fully accepted, but a model considering dimers as
the basic unit that binds to two ligand molecules is
lacking. Here, we propose a two-state-dimer model in
which the ligand-induced conformational changes from
one component of the dimer are communicated to the
other. Our model predicts cooperativity in binding,
which is relevant because the other current models fail
to address this phenomenon satisfactorily. Our two-
state-dimer model also predicts the variety of responses
elicited by full or partial agonists, neutral antagonists
and inverse agonists. This model can aid our under-
standing of the operation of heptaspanning receptors
and receptor channels, and, potentially, be important for
improving the treatment of cardiovascular, neurological
and neuropsychyatric diseases.
Background
From the application of the pioneering ‘two-state’ theoryfor receptors [1–3] a variety of models have been devised toexplain the behaviour of heptaspanning, G protein-coupled receptors (GPCRs) [4–14]. Almost all currentmodels are based on the non-cooperative mechanismproposed in 1957 by del Castillo and Katz [15] for nicotinicreceptors, which do not belong to the GPCR superfamily.
Although the binding of agonists to heptaspanning recep-tors leads to non-linear Scatchard plots (see Glossary),current models fail to predict non-linear Scatchard plotsfor agonist binding. Non-linear Scatchard plots displayingcurves with a Hill coefficient of !1 could be explainedby assuming the existence of two separated or non-interconvertible forms of the receptor: a high-affinityform (R* or G protein-coupled) and a low-affinity form(R or G protein-uncoupled). However, existing modelsassume that R and R* are in equilibrium and, therefore,they would account for non-linear Scatchard plots only if(i) the concentration of G protein is lower or similar to thatof the receptor and (ii) the interconversion betweencoupled and uncoupled forms is almost irreversible. Thisis not an accurate assumption for many physiologicalconditions, mainly because G proteins are in excess [16].
Corresponding author: Canela, E.I. ([email protected]).Available online 13 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
Moreover, an agonist might induce changes in theproportions of the so-called ‘high-affinity’ and ‘low-affinity’states, which strongly suggests that the two states cannotexist separately but that they are interconnected [17]. Wehave provided compelling evidence that the apparent inter-conversion between states is independent of G proteins [18].If the agonist varies the proportion of high- and low-affinity sites, the existing models cannot accuratelyrepresent the behaviour of the receptors. Moreover,these models cannot explain positive cooperativity,which has been reported for 5-hydroxytryptamine1A
receptor [19,20]. Non-linear Scatchard plots require amodel of cooperativity that can explain both positive andnegative cooperativity.
A general model that is able to predict positive, neutraland negative cooperativity should consider inter- and/orintra-molecular interactions resulting from a multivalentreceptor molecule in terms of the number of ligand-binding sites per molecule of receptor; that is, a moleculecapable of binding more than one agonist molecule or,alternatively, the receptor molecule being oligomeric[21,22]. Cooperativity in the binding of agonists toreceptor oligomers would come as naturally as in thecase of oligomeric enzymes or oligomeric functionalproteins such as haemoglobin.
The hypothesis on the existence of intramembranereceptor–receptor interactions was introduced in the early
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
two sets of data to determine whether they differ significantly. The classical
t-test is a particular case of the F-test.
. doi:10.1016/j.tibs.2005.05.010

α K
µθ K
L
α L
θα L
K
µK
A + A + (RR) A + A + (RR)*
A + A(RR)*
A(RR)*AA(RR)A
A + A(RR)
Ti BS
Figure 1. Scheme of our proposed two-state-dimer-receptor model. K is the
equilibrium constant for the binding of the first ligand (A) molecule to the receptor
dimer (RR). L is the equilibrium constant between the resting (RR) and the active
(RR)* states of the dimer. A symmetric dimer is assumed and, therefore, the species
A(RR) and (RR)A are equivalent. A value of aZ1 suppresses any difference in the
affinity of (RR) and (RR)* for ligand A. A value of mZ1 would indicate that the affinity
of (RR) for the first and second A molecule is the same. Finally, qZ1 would indicate
that the equilibrium binding constant for the binding of A to A(RR) is identical to
that of A to A(RR)*. (See Supplementary Material for the equations derived from
this model).
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 361
1980s based on radio-ligand studies in membrane prepar-ations from brain regions [23–26]. The first demonstrationof GPCR homodimers was achieved with b-adrenergic andmuscarinic receptors [27,28]. It was suggested that themuscarinic receptor exists in oligomeric forms and that adimer and tetramer might exist as interconvertible species[28]. This was taken seriously ten years later with thedemonstration of further receptor homodimers in cellsexpressing recombinant receptors and in membranes frommammalian brain [29,30]. Among others, dopamine,adenosine, muscarinic, peptide P, GABA, metabotropicglutamate, opioid, adrenergic, histamine, serotonin andchemokine receptors can be found as homodimers in livingcells [27,31–33]. Recently, it has been shown that, whereasmonomers and dimers of A2A adenosine receptors arefound in living cells, the dimeric species is the pre-dominant one on the cell surface [34]. This means that, forA2A receptors and probably other receptors, dimers are thephysiological species that are activated by the physio-logical agonist. The available experimental evidenceindicates the impossibility of explaining the operation ofmany heptaspanning receptors without consideringdimers as the minimum structure. Thus, the existingmodels need to be revisited to consider dimers or a novelmodel should be devised that includes dimers as basicunits. Here, we consider a model – the ‘two-state-dimer’model – in which the minimal operating unit forheptaspanning membrane receptors is the dimer. Thismodel can explain not only cooperativity in agonistbinding (positive, neutral and negative) but also themolecular mechanism of full agonist, inverse agonist andantagonist operation.
The ligand-binding process
In the two-state-dimer model (Figure 1), the basic unit ofthe receptor is the homodimer. As in the case of the twoindependent-affinity-state model, each monomer of thereceptor molecule binds reversibly to an orthosteric ligand(A), but the receptor molecule – the homodimer – bindstwo molecules of ligand. The first molecule of ligand bindswith equilibrium constant K, forming a ternary complex[A(RR)]. A symmetrical dimer is assumed and, therefore,A(RR) and (RR)A species are equivalent. A secondmolecule of ligand binds to the ternary complex withequilibrium constant mK, where m is the binding coopera-tivity between the first and the second A molecule.
In terms of signalling or ‘receptor activity’, homodimerscan be in an inactive (RR) or active (RR)* form. Thefunctional response predicted by this model is pro-portional to the amount of active homodimers, that is,the sum of all the species containing the active homo-dimer: (RR)*, A(RR)* and A(RR)*A. In the absence ofligand, the proportion of (RR) and (RR)* is governed by L,the equilibrium constant for receptor isomerization. Asindicated, the ligand binds to the inactive state of thereceptor (RR) with equilibrium constant, K, and to theactive state of the receptor [(RR)*] with a modifiedequilibrium constant, aK. Binding of the ligand shiftsthe isomerization equilibrium constant from L to aL. Theconstant a reflects the intrinsic efficacy of A, that is, theratio of affinities of A for (RR)* and (RR); high values of a
www.sciencedirect.com
favour the formation of A(RR)*, and low values of a favourstabilization of the A(RR) states. A second molecule ofligand binds to the ternary complexes [A(RR)] and[A(RR)*] with equilibrium constants mK and mqK, respec-tively. The equilibrium between A(RR)A and A(RR)*A isthen governed by aqL. The meaning of the constant q issimilar to that of a; q is the intrinsic efficacy of the secondA molecule entering the dimer, that is, the ratio of affinitiesof A for A(RR)* and A(RR). Binding of the second ligandmolecule shifts the isomerization equilibrium constant fromaL to aqL (a, m and q can take any non-negative value). Avalue of one for any of these parameters indicates that thereis no effect of ligand binding on the particular equilibriuminvolved (see Supplementary Material).
Approximately 20 years ago there was already specu-lation about a possible dependence of cooperativity uponthe existence of receptor oligomeric structures [28]. As forenzymes, the rationale behind the ‘two-state-dimer’ modelis the communication of ligand-induced conformationalchanges [35] from one component of the dimer to the other.It should be noted that, although both monomers anddimers coexist in the cell, the functional species on the cellsurface seems to be the dimeric one [31,34].
The theoretical analysis of cooperativity in ligand bindingusing the two-state-dimer model (Figure 1) indicatesthat positive cooperativity, negative cooperativity or

Table 1. Effect of parameters a, q and m on cooperativity in ligand bindinga
mZ10 mZ1 mZ0.25 mZ0.1
q aZ1!10K3 aZ1 aZ1!103 aZ1!10K3 aZ1 aZ1!103 aZ1!10K3 aZ1 aZ1!103 aZ1!10K3 aZ1 aZ1!103
1!10K3 C C K C C K C K K K K K
1!10K2 C C K C C K C K K K K K
1!10K1 C C K C C K C K K K K K
1 C C K C C K C (1) K K K K
(8.5) (5.5)
1!101 C C C C C K C C K K C K
(84)
1!102 C C C C C C C C K K C K
(334) (836)
1!103 C C C C C C C C C K C CaThe performance of the proposed two-state-dimermodel (Figure 1) was assessed by calculating ligand (A) binding for an arbitrary equilibrium constant, K, value of 1 and the
ligand concentration in the range of 0.001–1000.The equilibrium constant between (RR) and (RR)*, L, was arbitrarily chosen as 0.5, which represents that, in the absence of A,
the constitutive activity is one third of the total activity, that is, the proportion of (RR)*with respect to the total receptor is 0.33. The values of a and qwere varied between 0.001
and 1,000, and m between 10 and 0.1. From a theoretical point of view, these values adequately cover all the possibilities of curves of binding versus ligand concentration. The
cooperativity was determined by considering the difference between the deduced saturation function (see Supplementary Material) and the reference saturation function,
which would correspond to the non-cooperative binding of molecule A to a dimer. The difference between these two functions is indicated: positive,C; negative,K; or non-
cooperativity, Hill coefficientZ1. Approximate q values for which non-cooperativity exists are in brackets.
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005362
non-cooperativity (or neutral cooperativity) can occur(Table 1). For given L and K values, there are a significantnumber of combinations of a, q and m values that give non-cooperative behaviour. Therefore, the existence of dimersdoes not necessarily account for positive or negativecooperativity (Table 1).
Bound (
1.5
0.6 1
1.0
0.5
0.40.2
0.5
0.3
0.1
0.40.2
0.6 10.40.2
8
4
0.40.2
700
0.6 1
350
0.40.2
300
150
0.40.2
25
15
5
(a) α = 0.001, µ = 10 (b)
(d) (e)
(g) (h)
α = 0.001,
α = 1, µ = 10 α = 1, µ =
Bou
nd (
dim
er)/
[A] fr
ee
α = 1000, µ = 10 α = 1000,
0.8
0.8
0.8
Figure 2. Scatchard plots corresponding to binding data deduced from the two-state dim
(c) aZ0.001, mZ0.1; (d) aZ1, mZ10; (e) aZ1, mZ1; (f) aZ1, mZ0.1; (g) aZ1000, mZ10; (h) a
1, 0.1 and 0.001. a reflects the intrinsic efficacy of the first ligand (A); q is the intrinsic e
between the first and the second A molecule. Ligand (A) concentrations in the range 0.
between (RR) and (RR)*, L, was 0.5.
www.sciencedirect.com
It has been deduced from the cooperativity analysisthat the shape of the Scatchard plot (Figure 2) isextremely dependent on a, q and m values. In generalterms, for low values of the intrinsic efficacy for the firstbinding, a, and high values of the intrinsic associationconstant for the second binding, m, convex upward curves,
dimer)
0.6 1
0.30
0.6 1
0.15
0.40.2
0.6 1
2.5
0.6 1
1.5
0.5
0.40.2
0.6 1
140
0.6 1
70
0.40.2
(c)
(f)
(i)
µ = 1 α = 0.001, µ = 0.1
1 α =1, µ = 0.1
µ = 1 α=1000, µ = 0.1
0.8 0.8
0.8 0.8
0.8 0.8
Ti BS
er-receptor model for different values of a. (a) aZ0.001, mZ10; (b) aZ0.001, mZ1;
Z1000, mZ1; and (i) aZ1000, mZ0.1. q (top to bottom in every plot)Z 1000, 100, 10,
fficacy of the second A molecule entering the dimer; m is the binding cooperativity
001–1000 and an arbitrary K value of 1 were considered. The equilibrium constant

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 363
which are indicative of positive cooperativity, are pre-dominantly found (Figure 2a,b,d,e). By contrast, for lowvalues of both m and a, negative cooperativity is found(Figure 2c). For higher values of a, positive or negativecooperativity would depend on the value of the intrinsicefficacy of the second binding (q) (Figure 2g,h,i). When q ishigh, positive cooperativity is predominantly found,whereas negative cooperativity is more probable when qis low (Figure 2d,g).
Receptor activation
The performance of our two-state-dimer model has beenassessed by a study of the dependence of receptor activityon ligand concentration. To enable direct comparison ofthe responses elicited by agonists and antagonists, theoccupational theory of drug action [36] was selected. Inthe symmetric two-state-dimer model, (RR)*, A(RR)* andA(RR)*A contribute to the overall response, whereas theinactive (RR), A(RR) and A(RR)A do not. The proportional-ity factor is set to one and the response, therefore, isequivalent to the ratio of active species versus totalamount of receptor [Equation 1, where A(RR)* isequivalent to (RR)*A; see Supplementary Material]:
0.30
-3 -2 -1 12 3
0.15
-3 -2 -1
-3 -2 -1 1 3 -3 -2 -1
-3 -2 -1 1 3 -3 -2 -1
0.30
0.15
1.0
0.5
1.0
0.5
1.0
0.5
1.0
0.5
0
0
0
α = 0.001, µ = 10 α = 0.00
α = 1, µ = 10 α = 1, µ
α = 1000, µ = 10 α = 1000
2
2
Bo
Bou
nd (
dim
er)/
[A] fr
ee
(a) (b)
(d) (e)
(g) (h)
Figure 3. Receptor-activation curves for the two-state-dimermodel. Curves dose–respons
0.001, mZ0.1; (d) aZ1, mZ10; (e) aZ 1, mZ1; (f) aZ1, mZ0.1; (g) aZ1000, mZ10; (h) aZ100
and 0.001. a reflects the intrinsic efficacy of the first ligand (A); q is the intrinsic efficacy of t
first and the second Amolecule. Ligand (A) concentrations in the range 0.001–1000 and a
(RR)*, L, was 0.5.
www.sciencedirect.com
Response Z½ðRRÞ��C ½AðRRÞ��C ½AðRRÞ � A�
½RTotal�Eqn 1
The variation of the response with respect to ligandconcentration has been analysed and the results aresummarized in Figure 3.
In the simplest case, when the intrinsic efficacy for thefirst ligand binding (a) is one (Figure 3d–f), the curves canbe flat (no variation in receptor activity upon ligandbinding) when the intrinsic efficacy for the second binding(q) is one, or can slope upwards (when qO1) or downwards(when q!1), until a plateau is reached. When a is high(e.g. aZ1000; Figure 3g–i), the proportion of activereceptor dimers increases with ligand concentrationuntil a plateau is reached, after which it eventuallydeclines. In this case, it is possible to find potentiation ofsignalling at low ligand concentrations and inhibition athigher ligand concentrations. Also, it is evident from thecurves depicted in Figure 3 that a decrease in the bindingcooperativity between the first and the second A molecule(m) makes a higher concentration of ligand necessary toproduce a given level of signalling. Also, the two-state-dimer model predicts that a compound can eventuallydecrease receptor activity further than the constitutive
12 3 -3 -2 -1 12 3
1 3 -3 -2 -1 1 3
1 3 -3 -2 -1 1 3
0.30
0.15
1.0
0.5
1.0
0.5
0 0
00
0 0
1, µ = 1 α = 0.001, µ = 0.1
= 1 α = 1, µ = 0.1
, µ = 1 α = 1000, µ = 0.1
2 2
2 2
und (dimer)
(c)
(f)
(i)
Ti BS
ewere devised for different values of a. (a) aZ0.001, mZ10; (b) aZ0.001, mZ1; (c) aZ0, mZ1; (i) aZ1000, mZ0.1. q (top to bottom in every plot)Z1000, 100, 10, 1, 0.1, 0.01
he second Amolecule entering the dimer; m is the binding cooperativity between the
n arbitrary K value of 1 were considered. The equilibrium constant between (RR) and

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005364
activity until a plateau or a minimum is reached(Figure 3a–c).
Full and partial agonists
Full agonists are defined by their ability to increasereceptor activity in a dose-dependent manner until themaximum signalling response is reached. Full agonistbinding to a dimeric receptor would require values of aZ1000 for any m or q, or aZ1 for qR100 and any m(Figure 3d–i). The two-state-dimer model can explainpositive and negative cooperativity in full agonist bindingto the receptor dimer (Table 1). A partial agonist is definedas a compound that activates the receptor but unable toinduce full receptor activation. Partial agonist binding to adimeric receptor would require aZ1 and qZ1–100 for anym value (Figure 3d,e,f).
Antagonists and inverse agonists
On defining a neutral antagonist as a molecule that doesnot change the proportion of inactive [(RR)] versus active[(RR)*] species, the combination of parameters in the two-state-dimer model (which allows such a possibility) is aZ1and qZ1 for any m value (Figure 3d–f). Often, it is reportedthat binding of a neutral antagonist is non-cooperative(linear Scatchard plot). In the existing models, it isassumed that the antagonist does not distinguish betweenthe high-affinity- and low-affinity-binding sites. By con-trast, in the two-state-dimer model, non-cooperativeantagonist binding would be a unique case because theonly combination of parameters that leads to flat con-stitutive activity and give linear Scatchard plots are: mZ0.25, aZ1 and qZ1. A m value of 0.25 is required for theoverall affinity of the second binding of the ‘non-cooperative’ antagonist to be identical to the overallaffinity for the first binding. Because there are twobinding sites for the antagonist, when aZ1 and qZ1, thevalue of mK is K/4. However, values that differ slightlyfrom those indicated can lead to apparently linearScatchard plots and, thus, detection of cooperativity inbinding can become a difficult task [37].
Inverse agonists are defined by their ability toantagonize the effect of full agonists, but also by theirability to decrease the constitutive activity of hepta-spanning membrane receptors. By inspecting Figure 3, acompound would be an inverse agonist when the values ofthe parameters are: a!1 for any q and m value, or aZ1 andq!1 irrespective of m (Figure 3a–f). The scarce availabledata indicate that cooperativity can exist in the binding ofinverse agonists [37] and that values representing non-cooperative binding to the dimer would be restrictive. Thetwo-state-dimer model can predict any type of cooperativ-ity for compounds that reduce the constitutive activity ofheptaspanning receptors (Table 1).
Predictions and performance of the two-state-dimer
model
For A1 adenosine receptors, which display non-linearScatchard plots, experimental data fit better to the two-state-dimer model than to the two independent-affinity-state model [38]. Although the difference between theactual values and the theoretical ones were similar in the
www.sciencedirect.com
two cases, the two-state-dimer model has less parametersand, therefore, the discriminative F-test selects it as thesimplest model able to explain the data. The parametersobtained when fitting data to the two-state-dimer modelgive valuable information about the equilibrium con-stants but also about the degree of cooperativity (seeSupplementary Material).
One of the main features of the two-state-dimer modelis its ability to predict quantitative changes in functionalactivity. Ligands with high values of a (e.g. aZ1000) andlow values of q (e.g. qZ0.1) show dual behaviour in termsof response at equilibrium; they increase the number ofactive species at low agonist concentration, but theydecrease them at high concentrations (Figure 3). Thiseffect is even more evident for mO0.1. Thus, the modelpredicts a homologous desensitization process at highconcentrations of agonist. The strength of this desensi-tization process would vary from agonist to agonist(Figure 3d,h,i). The limit of the curves at high concen-trations of A is the constitutive activity (when qaZ1) orsurpasses it (when qa!1). Interestingly, desensitizationdepends on the product aq, which reflects properties ofboth the binding of the first ligand molecule and thebinding of the second molecule to the dimer. If aqZ1, thelimit of the curve at high ligand concentrations is theconstitutive activity. This correlates with the well-knownobservation that, in terms of desensitization, two types ofreceptor exist: those that are desensitized easily and thosethat are not. Although desensitization is usually inter-preted to be a consequence of uncoupling of signallingmachinery, this is not known for certain. The two-state-dimer model predicts that homologous desensitization isdependent on the agonist in such a way that it is weakerfor agonists displaying positive cooperativity (Figures 2and 3). As measured experimentally, desensitizationincluding the well-known refractoriness to agonists afterreceptor activation would involve this homologousdesensitization plus receptor internalization, which isdependent on G protein-receptor kinase activation andreceptor phosphorylation.
The two-state-dimer model predicts that neutralantagonists can display any type of cooperativity onbinding to receptor dimers. However, as mentioned,detection of cooperativity in binding can become a difficulttask. Recently, Cheng [37] compared different methods ofanalysing cooperativity data. The author has reportedthat the Cheng–Prusoff equation, the Scatchard analysisand the Schild analysis give different results, and hasproposed a more robust method to calculate accuratevalues for equilibrium constants and Hill coefficients.Periyasami and Somani [39] have shown that theapparent Hill coefficient for yohimbine – an antagonistof a2 adrenoceptors both in equilibrium binding and incompetitive displacement assays – is !1. It should benoted that some data on dissociation of antagonists fromGABA and glycine receptors [40] are attributed to bindingof the antagonist to an orthosteric centre and to anallosteric centre. In addition, allosteric effectors – whichhave been reported for a variety of heptaspanningmembrane receptors – would affect the cooperativitybetween the two molecules in the dimer. The consideration

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 365
of allosteric centres will be a further development of themodel we present here.
In a recent survey of 105 published papers on theactivity of 380 antagonists on 73 biological GPCR targets,322 of the antagonists studied are reported to be inverseagonists and only 58 to be neutral antagonists [41]. Thepredominance of inverse agonism agrees with theoreticalpredictions of the two-state-dimer model, but also agreeswith those of other reported models (see SupplementaryMaterial). The parameters defining neutral antagonism inthe two-state-dimer model are restrictive and this could bethe reason that neutral antagonists are the minoritywithin the pharmacological space.
There are few reports on the cooperativity of inverse-agonist binding. The available data indicate, however,that cooperativity can exist in the binding of suchcompounds for a variety of receptors. The two-state-dimer model can predict any type of cooperativity forcompounds that reduce the constitutive activity; there-fore, the binding of these compounds to the receptordimer is represented by a!1 for any value of q and m, oraZ1 and q!1 for any value of m. Results of competitionassays for an inverse agonist of melacortin-4 receptors,carefully analysed using the Cheng protocol [37], showthat cooperativity in the competition occurs and that itvaries depending on the nature of the radioactivecompound bound. Higher (and positive) cooperativity isfound when the inverse agonist competes with the fullagonist, whereas the Hill coefficient decreases when thecompound competes with either a partial agonist or aneutral antagonist [37]. Moore and Scanlon [42] provideanother example of change in cooperativity dependingon the nature of compounds competing at the bindingsite of receptor. They demonstrate that antagonists shiftpositive cooperativity to negative cooperativity in thebinding of agonists to angiotensin receptors. Interestingly,Milligan et al. [43] have shown that two different inverseagonists that are specific for 5-hydroxytryptamine1A
receptors display different Hill coefficients. Whereasone of the inverse agonists has a Hill coefficient of O1(positive cooperativity), agonists and partial agonistsdisplay low Hill coefficients indicative of strong negativecooperativity. These varied behaviours can be easilyexplained by the two-state-dimer model: each of thesecompounds would have different a, q and m valuesand, therefore, cooperativity in binding-saturationcurves would be different for different compounds; andcooperativity for a given compound in competitionassays would vary depending on the nature of theradio-ligand bound and the nature of the competitor.Furthermore, the two-state-dimer model predicts thatcooperativity in the binding of all type of molecules tothe orthosteric centre would depend on the value of L(i.e. constitutive activity).
Another predictive valuable feature of the model isrelated to the possibility that an inverse agonist mightreduce constitutive activity at low concentrations butrevert this trend at higher concentrations. This should beconsidered when analysing data for novel putative inverseagonists before discarding them as ineffective.
www.sciencedirect.com
Concluding remarks
To explain ligand-binding and activation mechanisms ofGPCRs, their dimeric structure must be taken intoaccount. The model we propose here assumes the existenceof cooperative conformational changes in the GPCR dimer,which has been demonstrated for the leukotriene B4
receptor BLT1 [44]. In our opinion, we present a generalmodel for heptaspanning-membrane-receptor operationthat is based precisely in their (now) well-known dimericstructure. This two-state-dimer model can explain all thefeatures of the heptaspanning-membrane receptors andcan be useful for investigation of other types of receptorssuch as nicotinic acetylcholine receptors, which have twoidentical binding sites [45]. Furthermore, the modelpredicts other features (e.g. dual effects for a givencompound) of heptaspanning receptors – a tool thatcould be useful for improving current therapeutic strat-egies that target this type of receptor.
AcknowledgementsThis work was supported by grant SAF2001–3474 from Ministerio deCiencia y Tecnologia, Grant 02/056–00 from Fundacio la Caixa, Grants01/012710 and 02/021010 from Fundacio Marato TV3.
Supplementary data
Supplementary data associated with this article can befound at doi:10.1016/j.tibs.2005.05.010
References
1 Karlin, A. (1967) On the application of “a plausible model” of allostericproteins to the receptor for acetylcholine. J. Theor. Biol. 16, 306–320
2 Colquhoun, D. (1973) The relationship between classical andcooperative models for drug action. In A Symposium on DrugReceptors (Rang, H.P., ed.), pp. 149–182, University Park Press
3 Thron, C.D. (1973) On the analysis of pharmacological experiments interms of an allosteric receptor model. Mol. Pharmacol. 9, 1–9
4 De Lean, A. et al. (1980) A ternary complex model explains the agonist-specific binding properties of the adenylate cyclase-coupledb-adrenergic receptor. J. Biol. Chem. 255, 7108–7117
5 Costa, T. and Herz, A. (1989) Antagonists with negative intrinsicactivity at delta opioid receptors coupled to GTP-binding proteins.Proc. Natl. Acad. Sci. U. S. A. 86, 7321–7325
6 Samama, P. et al. (1993) A mutation-induced activated state of theb2-adrenergic receptor. Extending the ternary complex model. J. Biol.Chem. 268, 4625–4636
7 Onaran, H.O. et al. (1993) b g subunits of guanine nucleotide-bindingproteins and regulation of spontaneous receptor activity: thermo-dynamic model for the interaction between receptors and guaninenucleotide-binding protein subunits. Mol. Pharmacol. 43, 245–256
8 Samama, P. et al. (1994) Negative antagonists promote an inactiveconformation of the b2-adrenergic receptor. Mol. Pharmacol. 45,390–394
9 Leff, P. (1995) The two-state model of receptor activation. TrendsPharmacol. Sci. 16, 89–97
10 Weiss, J.M. et al. (1996) The cubic ternary complex receptor –occupancy model I. Model description. J. Theor. Biol. 178, 151–167
11 Weiss, J.M. et al. (1996) The cubic ternary complex receptor –occupancy model II. Understanding apparent affinity. J. Theor. Biol.178, 169–182
12 Weiss, J.M. et al. (1996) The cubic ternary complex receptor – occupancymodel III. Resurrecting efficacy. J. Theor. Biol. 181, 381–397
13 Hall, D.A. (2000) Modeling the functional effects of allostericmodulators at pharmacological receptors: an extension of the two-state model of receptor activation. Mol. Pharmacol. 58, 1412–1423
14 Lorenzen, A. et al. (2002) Modulation of agonist responses at the A1

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005366
adenosine receptor by an irreversible antagonist, receptor-G proteinuncoupling and by the G protein activation state. Biochem.Pharmacol. 64, 1251–1265
15 del Castillo, J. and Katz, B. (1957) A comparison of acetylcholine andstable depolarizing agents.Proc. R. Soc. Lond. B. Biol. Sci. 146, 362–368
16 Neubig, R.R. (1994) Membrane organization in G-protein mechan-isms. FASEB J. 8, 939–946
17 Wong, H.M. et al. (1986) Assessment of mechanistic proposals for thebinding of agonists to cardiac muscarinic receptors. Biochemistry 25,6995–7008
18 Casado, V. et al. (1991) The binding of [3H]R-PIA to A1 adenosinereceptors produces a conversion of the high- to the low-affinity state.FEBS Lett. 286, 221–224
19 Mattera, R. et al. (1985) Guanine nucleotide regulation of amammalian myocardial muscarinic receptor system. Evidence forhomo- and heterotropic cooperativity in ligand binding analyzed bycomputer-assisted curve fitting. J. Biol. Chem. 260, 7410–7421
20 Assie, M-B. et al. (1999) Correlation between low/high affinity ratiosfor 5-HT(1A) receptors and intrinsic activity. Eur. J. Pharmacol. 386,97–103
21 Franco, R. et al. (1996) The cluster-arranged cooperative model: amodel that accounts for the kinetics of binding to A1 adenosinereceptors. Biochemistry 35, 3007–3015
22 Franco, R. et al. (2003) Regulation of heptaspanning-membrane-receptor function by dimerization and clustering. Trends Biochem.Sci. 28, 238–243
23 Agnati, L.F. et al. (1980) Aspects on receptor regulation andisoreceptor identification. Med. Biol. 58, 182–187
24 Agnati, L.F. et al. (1982) New vistas on synaptic plasticity: the receptormosaic hypothesis of the engram. Med. Biol. 60, 183–190
25 Fuxe, K. et al. (1981) Modulation by cholecystokinins of 3H-spiroperidolbinding in rat striatum: evidence for increased affinity and reductionin the number of binding sites. Acta Physiol. Scand. 113, 567–569
26 Agnati, L.F. et al. (2003) Molecular mechanisms and therapeuticalimplications of intramembrane receptor/receptor interactions amongheptahelical receptors with examples from the striatopallidal GABAneurons. Pharmacol. Rev. 55, 509–550
27 Fraser, C.M. and Venter, J.C. (1982) The size of the mammalian lungb2-adrenergic receptor as determined by target size analysis andimmunoaffinity chromatography. Biochem. Biophys. Res. Commun.109, 21–29
28 Avissar, S. et al. (1983) Oligomeric structure of muscarinic receptors isshown by photoaffinity labeling: subunit assembly may explain high-and low-affinity agonist states. Proc. Natl. Acad. Sci. U. S. A. 80,156–159
Reproduction of material
Interested in reproducing part or all of an article published by ElGlobal Rights Department with details of how and where the req
on-line, plea
http://www.elsevier.com/wps/find/obtainpermi
Alternatively, ple
ElseviGlobal Rights D
PO BoxOxford OX5
Phone: (+44) 18Fax: (+44) 186
permissions@e
www.sciencedirect.com
29 Ciruela, F. et al. (1995) Immunological identification of A1 adenosinereceptors in brain cortex. J. Neurosci. Res. 42, 818–828
30 Ng, G.Y. et al. (1996) Dopamine D2 receptor dimers and receptor-blocking peptides. Biochem. Biophys. Res. Commun. 227, 200–204
31 Bouvier, M. (2001) Oligomerization of G-protein-coupled transmitterreceptors. Nat. Rev. Neurosci. 2, 274–286
32 Milligan, G. and White, J.H. (2001) Protein-protein interactions atG-protein-coupled receptors. Trends Pharmacol. Sci. 22, 513–518
33 Bai, M. (2004) Dimerization of G-protein-coupled receptors: roles insignal transduction. Cell. Signal. 16, 175–186
34 Canals, M. et al. (2004) Homodimerization of adenosine A2A receptors:qualitative and quantitative assessment by fluorescence and bio-luminescence energy transfer. J. Neurochem. 88, 726–734
35 Vilardaga, J.P. et al. (2003) Measurement of the millisecond activationswitch of G protein-coupled receptors in living cells. Nat. Biotechnol.21, 807–812
36 Kenakin, T. (2004) Principles: receptor theory in pharmacology.Trends Pharmacol. Sci. 25, 186–192
37 Cheng, H.C. (2004) The influence of cooperativity on the determin-ation of dissociation constants: examination of the Cheng–Prusoffequation, the Scatchard analysis, the Schild analysis and relatedpower equations. Pharmacol. Res. 50, 21–40
38 Saura, C. et al. (1996) Adenosine deaminase interacts with A1
adenosine receptors in pig brain cortical membranes. J. Neurochem.66, 1675–1682
39 Periyasamy, S. and Somani, P. (1986) Effect of proteases andphospholipases on [3H]yohimbine binding to human platelet mem-branes. Biochem. Pharmacol. 35, 3131–3136
40 Maksay, G. (1990) Dissociation of muscimol, SR 95531, and strychninefrom GABAA and glycine receptors, respectively, suggests similarcooperative interactions. J. Neurochem. 54, 1961–1966
41 Kenakin, T. (2004) Efficacy as a vector: the relative prevalence andpaucity of inverse agonism. Mol. Pharmacol. 65, 2–11
42 Moore, G.J. and Scanlon, M.N. (1989) Methods for analyzing andinterpreting cooperativity in dose-response curves – I. Antagonisteffects on angiotensin receptors in smooth muscle. Gen. Pharmacol.20, 193–198
43 Milligan, G. et al. (2001) S 14506: novel receptor coupling at 5-HT1A
receptors. Neuropharmacology 40, 334–34444 Mesnier, D. and Banerest, J-L. (2004) Cooperative conformational
changes in a G-protein-coupled receptor dimer, the leukotriene B(4)receptor BLT1. J. Biol. Chem. 279, 49664–49670
45 Edelstein, S.J. et al. (1996) A kinetic mechanism for nicotinicacetylcholine receptors based on multiple allosteric transitions. Biol.Cybern. 75, 361–379
from Elsevier articles
sevier, or one of our article figures? If so, please contact ouruested material will be used. To submit a permission requestse visit:
ssionform.cws_home/obtainpermissionform
ase contact:
erepartment800,1DX, UK.65-8438305-853333lsevier.com

SNAREcomplexesandneuroexocytosis:howmany, how close?Cesare Montecucco1,3, Giampietro Schiavo2 and Sergio Pantano3
1Dipartimento di Scienze Biomediche and Istituto CNR di Neuroscienze, Universita di Padova, Viale G. Colombo n. 3,
35121 Padova, Italy2Molecular NeuroPathoBiology Laboratory, Cancer Research UK, London Research Institute, Lincoln’s Inn Fields Laboratories,
44 Lincoln’s Inn Fields, Room 614-615, London WC2A 3PX, UK3Istituto Veneto di Medicina Molecolare, Via Orus 2, 35129 Padova, Italy
Regulated secretion is an essential process in all
eukaryotic cells. The release of molecules contained
inside exocytic granules and synaptic vesicles is
mediated by the assembly of a SNARE complex formed
by the coil-coiling of three proteins: SNAP-25, syntaxin
and VAMP/synaptobrevin. It seems that SNARE
complexes assemble together in rosette-shaped super-
complexes but there is controversy on the actual
number (N) of copies of SNARE complexes that are
necessary to mediate exocytosis. We discuss attempts
to determine the value of N and suggest that N varies
with the type of exocytic vesicles. In addition, we
propose that the N value in neuroexocytosis can be
estimated by the comparative use of different types of
botulinum neurotoxins.
Introduction
Neuroexocytosis is the fundamental physiological processthat leads a naıve cytosolic synaptic vesicle (SV) to bind toand fuse with the presynaptic membrane, therebydischarging its neurotransmitter contents into the syn-aptic cleft. Our interest in neuroexocytosis arose some20 years ago when we began to study the mechanism ofaction of the clostridial neurotoxins causing tetanus andbotulism. Meanwhile, an unprecedented wealth of infor-mation brought the field from the static, low-resolutionpicture provided by electron microscopy to the tantalizingmolecular and biophysical puzzle that is currentlyavailable. At present, many scientists are trying topinpoint the right place and time of action for the manymolecules that are suggested to be linked, in one way oranother, to neuroexocytosis.
The three synaptic SNARE proteins SNAP-25, VAMP/synaptobrevin and syntaxin occupy a central position inthis process by forming a heterotrimeric complex, whichbinds N-ethylmaleimide sensitive fusion ATPase (NSF)and soluble NSF attachment protein [a-SNAP; hence theacronym SNARE (SNAP receptor)] [1]. The synapticSNARE proteins are the specific substrates of the eightclostridial neurotoxins (one tetanus neurotoxin, TeNT,and seven botulinum neurotoxins: BoNT/A–G) [2,3].These neurotoxins specifically bind to nerve terminals
Corresponding author: Montecucco, C. ([email protected]).Available online 2 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
and deliver their zinc-endopeptidase N-terminal domaininside the cytosol, where it specifically cleaves a SNAREprotein at a single site within its cytosolic portion. Suchspecific cleavage leads to a prolonged, but eventuallyreversible, inhibition of neuroexocytosis, which in vivoresults in the paralytic syndromes of botulism andtetanus. Despite the availability of several pieces ofexperimental evidence demonstrating the involvement ofSNARE proteins in neuroexocytosis, and in the majority ofmembrane traffic events within eukaryotic cells [4,5], thisremains the most impressive proof of their central role inneuroexocytosis; and a badly unwanted one, if oneconsiders the hundreds of thousands of newborn babiesthat the WHO reports to die each year by tetanusneonatorum in the non tetanus-vaccinated areas of theworld [6].
There is evidence that SNARE complexes assembletogether in rosette super-complexes around the site ofmembrane fusion. Additional proteins, including thecalcium sensor synaptotagmin I and the syntaxin-inter-acting protein Munc-18 [5,7,8], cooperate with SNAREs toaccomplish neuroexocytosis, although their completenumber and precise mode- and time-of-action has not yetbeen established [5,7,8]. In addition, the number of copiesof SNARE complexes, termed N, that are necessary tomediate exocytosis is not known. Different methodsprovide different estimates and we discuss here someattempts at determining the value of N and the possibilitythat N varies with the type of exocytosis event. We willconclude by proposing the use of different types of BoNT toestimate the N value in neuroexocytosis.
The neuronal SNARE complex
Despite the large repertoire of SNAREs that are present ina typical mammalian cell (up to 26 have been identified inbudding yeast) [9], only a few of them are involved in therapid, highly controlled release of the neurotransmitter-containing vesicles, which follows nerve membranedepolarization. This event enables the influx of calciumthrough calcium channels located near the ‘active zones’,which are the preferred sites of neuroexocytosis [5,7].
VAMP/synaptobrevin 1 and 2 (here abbreviated asVAMP) are 13 kDa integral membrane proteins of SV andof large dense-core granules. They consist of four parts
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
. doi:10.1016/j.tibs.2005.05.002

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005368
(numbering refers to the rat isoform 2): (i) a N-terminal(27 residues) proline-rich segment whose sequence isisoform specific; (ii) segment 28–93, which is wellconserved among isoforms and contains heptad repeatstypical for coiled-coils and a short a-helix that inserts,together with nearby residues (77–92), into the interfacialregion of the lipid bilayer [10]; (iii) a single trans-membrane (TM) segment; and (iv) a poorly-conservedintra-lumenal tail of variable length due to alternativesplicing.
Syntaxin 1 and 2 are bound to the presynapticmembrane of neurons by a TM segment linked to a shortC-terminal domain exposed to the cell surface and a largecytosolic portion that includes two domains with distinctstructural features.TheN-terminaldomain (residues1–120)consists of three long a-helices that are likely to beinvolved in protein–protein interactions [10], whereas thecentral portion (residues 180–262) contains coiled-coil-forming heptad repeats. Several syntaxin isoforms co-exist within the nervous tissue.
Synaptosomal-associated membrane protein of 25 kDa(SNAP-25) is localized on the cytosolic face of the nervemembrane via palmitoylated cysteines located in themiddle of the polypeptide chain (Figure 1, right-handpanels). Both the N- and C-terminal halves of SNAP-25contain heptad repeats. SNAP-25 is highly conservedamong species with little variation in length. Uponinteraction with syntaxin, SNAP-25 forms a three-helix
0
80
60
40
20
[3H
] gly
cine
rel
ease
and
SN
AP
-25
clea
vage
(%
of c
ontr
ol)
[3H
] gly
cine
rel
ease
and
SN
AP
-25
clea
vage
(%
of c
ontr
ol)
Toxin concentration (Mo
Toxin concentration (Mo
100
0
80
60
40
20
100
10-12 10-11 10-10 1
10-12 10-11 10-10 1
(b) BoNT/E
(a) BoNT/A
Figure 1. Dose response curves of the effects of (a) botulinum neurotoxin A (BoNT/A) an
release (green circles, broken line). The vertical bars indicate the values (C/K SD) for SNA
scheme of SNAP-25 and the position of BoNT/A and BoNT/E cleavages. Adapted, with p
www.sciencedirect.com
bundle complex, which might function as a VAMP/synaptobrevin receptor on the plasma membrane [11,12].Moreover, SNAP-25 stoichiometrically binds to the puta-tive calcium sensor synaptotagmin and this interaction isbelieved to be important for the calcium-dependent phaseof neurotransmitter release [13]. Two isoforms (A and B)are generated by alternative splicing, with the B formpredominating in the adult nervous tissue. Both shorter(SNAP-23) and longer (SNAP-29) isoforms are also known,which can partially vicariate for SNAP-25 [14].
The three SNARE proteins associate before membranefusion into a 1:1:1 SNARE complex by bundling their coildomains starting from their N termini. Coiling proceedsuntil the formation of a w12 nm-long four-helices bundlewith a parallel orientation, which brings the TM segmentsof VAMP and syntaxin close to each other and close tothe C terminus and to the palmitoylated central loop ofSNAP-25 [15] (Figure 2a). This SNARE complex has atrans configuration with respect to the membrane locationof the two TM domains, one of which resides in the SVmembrane and the other is embedded in the synapticplasma membrane (Figure 2a). The surface of the SNAREcomplex has major grooves and charges enabling thepossibility of additional specific protein–protein inter-actions [5]. As discussed in detail elsewhere [5,7], thetrans-SNARE complexes bring the vesicle and plasmamembrane into close contact, thus permitting membranefusion, a process that has been mimicked in liposomes
CC CC
CCCC
lar)
lar)
0-9 10-8
0-9 10-8
BoNT/A
BoNT/E
d (b) BoNT/E on SNAP-25 proteolysis (purple squares, continuous line) and glycine
P-25 proteolysis (purple) and glycine release (green). The right-hand panels show a
ermission, from Ref. [43].

Ti BS
(a)(b)Vesicle membrane
Plasma membrane
BoNT/A
Figure 2. A model of the multimeric ring of SNARE complexes. The model is based on the X-ray structure of a complexin–SNARE complex (PDB code: 1KIL; [48] from which
complexin has been removed). (a) Side view of a cross section of SNARE super-complex. This model has been obtained by generating the transmembrane (TM) segments of
VAMP and syntaxin according to TMHMM predictions (at http://www.cbs.dtu.dk/services/TMHMM/) and imposing the constrains that the TM domains of VAMP (in blue) and
syntaxin (in red) are in contact with each other and also with the C-terminal and cysteine-rich palmytoylated regions of SNAP25, to account for the available experimental data
on their role in exocytosis. SNAP-25 is represented as green ribbons in the SNARE coiled-coil region. The middle, unstructured domain of SNAP-25 is generated randomly
and depicted in cyan. Palmitoylated cysteines are represented as yellow spheres and black coils inserted into the plasma membrane. (b) Top view of the membrane fusion
super-complex from the vesicle side. The solvent accessible surface of the peptide removed by BoNT/A in SNAP-25 is in ochre. The inset evidences the putative contacts
between contiguous SNAREs mediated by the C-terminal segment of SNAP-25 cleaved by BoNT/A. A model was first made with a trimeric SNARE super-complexes but this
enables limited phospholipid mixing to take place within the three TM helices. Here, a dodecamer (suggested by experiments with botulinum neurotoxins described in the
text) is shown. This leaves a large space for phospholipid mixing and for the (putative) presence of proteins catalyzing phospholipid intermixing.
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 369
[16,17], albeit with a time course that is much slower thanthat of neuroexocytosis. This has led to the proposal thatthe SNARE complex is the ‘minimal fusion machinery’[16], where ‘minimal’ presumably indicates that theSNARE complex is the core of the neuroexocytosisapparatus, and that an additional set of proteins isrequired to account for specific properties of neuroexo-cytosis, such as fast-tight regulation, calcium-dependenttriggering and inhibition of spontaneous fusion events.After fusion, the SNARE complex assumes a cis configur-ation (i.e. with the two TM regions within the samemembrane). Owing to the stability of the SNARE complex(see subsequent sections), additional proteins (e.g. NSF,SNAPs and others) enter into play to unwind the coiledfour-helix bundle and to retrieve the SV by membranefission [4,5].
Multimeric SNARE complexes
The SNARE complex is characterized by high stability, aproperty that is reflected in its resistance to denaturingconditions and makes it separable by SDS–PAGE [18]. Inaddition to heterotrimeric bundles, higher order multi-meric SNARE complexes form in vitro [18–21]. In thesquid giant synapse, high order SNARE complexesinduced by the cytosolic protein synaphin (also known ascomplexin) are required for a reaction that takes placeafter SV docking and before fusion, and which isindependent of calcium channel gating [22]. This processdoes not occur in vitro in the absence of the VAMP andsyntaxin TM domains and biological membranes [23].As expected from the ability of the TM domains of VAMPand syntaxin 1A to form both homo- and hetero-oligomers
www.sciencedirect.com
[24,25], the formation of multimers of SNARE complexes(SNARE super-complex) strongly depends on these TMregions, and on intact palmitoylated SNAP-25. Super-complex formation is abolished by the isolated N- andC-terminal halves of SNAP-25 or by SNAP-25 mutated inthe central cysteine-rich loop [22].
The precise stoichiometry of the multimeric SNAREsuper-complex has not been yet established, mainlybecause the interactions among SNARE complexes areweaker than those taking place within the complex itself.However, it should be noted that even weak interactionsmight be relevant in vivo where they occur betweenpartners embedded in a biological membrane, which canbe compared to a two-dimensional solvent of limitedvolume. These findings led to the proposal that severalSNARE complexes bridged by SNAP-25 molecules form arosette around the fusion pore and that such a ring ofSNARE complexes is necessary for rapid regulatedexocytosis [16,22]. Furthermore, this SNARE rosette wassuggested to function as a scaffold for the recruitment ofadditional proteins with regulatory and/or fusogenicactivities [22]. Such a proposal is similar to that proposedfor the fusion between the viral and the plasma mem-branes mediated by the influenza virus hemagglutinin[26] and by the E1 fusion protein of Semliki Forest virus[27]. In these cases, a minimum of three hemagglutinintrimers and five E1 trimers were found to cooperate in thefusion process.
The first attempt to determine the number of SNAREcomplexes that are necessary and sufficient for a vesiclefusion event has been performed by Hua and Scheller[28] using cracked pheochromocytoma cells (PC12, a

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005370
neuroendocrine tumor cell line). This ‘open cell’ experi-mental model enables the manipulation of the conditionsleading to the fusion of large dense-core granules at will.In particular, the inhibitory effect of increasing amounts ofthe cytosolic SNARE domain of VAMP on the kinetics ofcalcium-induced granule release was monitored. Thetruncated VAMP used in this study lacks the TM domainand therefore forms a non productive SNARE complexupon interaction with SNAP-25 and syntaxin on theplasma membrane [28]. Following some assumptionsand fittings of the experimental data, it was suggestedthat three SNARE complexes cooperate in the fusion of asingle granule [28]. More recently, the effect of mutationsin the syntaxin TM domain was assessed by amperometricrecordings of norepinephrine release from large dense-core granules and by conductance analysis of cell-attachedmembrane patches in PC12 cells [29]. This study providescompelling evidence that the fusion pore is formed bymultiple copies of the syntaxin TM segment, alone ortogether with other proteins. Considering steric hindrancefactors, it was suggested that, if syntaxin alone forms thefusion pore, between five and eight molecules are involved[29], therefore doubling the minimal number of SNAREcomplexes initially postulated for the fusion of a singlegranule [28]. However, an even larger figure can bederived from recent experiments performed with BoNTs,by measuring the release of neurotransmitters containedin SVs in intact spinal cord neurons.
Inhibition of synaptic vesicle exocytosis by botulinum
neurotoxins
A striking feature of the action of different BoNTs withinnerve terminals is the large difference in the duration oftheir inhibitory action both in vitro and in vivo. BoNT/Aand BoNT/C have a much longer inhibitory effect on nerveterminals than the other BoNTs, with BoNT/E causing theshortest paralysis [30–32]. Although part of this effectcan be accounted for by a longer lifetime of BoNT/A andBoNT/C [32], it is clear that other factors come into play.Co-injection of BoNT/A and BoNT/E in the human exten-sor digitoris brevis muscle leads to a short, BoNT/E-typeparalysis, rather than a long, BoNT/A-type effect, sug-gesting that the long duration of BoNT/A is not only dueto its persistence within the synapse, but also to theinhibitory effect mediated by the truncated SNAP-25 [31].Indeed, the BoNT/A-cleaved SNAP-25 (SNAP-25–197;Figure 1) does form a stable SNARE complex withVAMP and syntaxin, which can be disassembled by NSF/a-SNAP as the wild-type complex. By contrast, this is notthe case for the shorter BoNT/E truncated SNAP-25(SNAP-25–180) [33–35] (see Figure 1, right-hand panels).Moreover, SNAP-25–197 has been detected for O80 days[36] in cultured mouse spinal cord neurons and it remainswithin the poisoned neuromuscular junction much longerthan SNAP-25–180 [37]. In addition, SNAP-25–197 acts asa dominant-negative factor because it blocks exocytosis ofcatecholamines from PC12 cells and of insulin frompancreatic b cells [38,39].
The C-terminal region of SNAP-25 is crucial forconferring calcium sensitivity to exocytosis because theparalysis caused by BoNT/A, but not that caused by
www.sciencedirect.com
BoNT/E, can be partially reversed by raising intracellularcalcium [40]. Accordingly, the intracellular delivery of apeptide corresponding to the 26 C-terminal residues ofSNAP-25, which is released by BoNT/E, inhibits exo-cytosis [41]. However, it does not contribute to BoNT/Esynaptic inhibition because it diffuses away rapidly fromthe site of production. A comparison of the dose–responseeffects of BoNT/A and BoNT/E on SNAP-25 cleavage andon neurotransmitter release in mice spinal cord neuronshas highlighted a major difference among the two toxins[42]. In the case of BoNT/E, a direct relationship betweenSNAP-25 cleavage and inhibition of neuroexocytosis wasfound, whereas BoNT/A affected neurotransmitter releasemuch more than SNAP-25 cleavage, indicating thatSNAP-25–197, but not SNAP-25–180, interferes withneuroexocytosis.
A careful quantitative analysis on the same system wasperformed recently by synchronizing toxin uptake bybinding in the cold followed by its internalization at 378Cunder depolarizing conditions [43]. As shown in Figure 1b,the dose–response curves of BoNT/E cleavage of SNAP-25and glycine release almost overlap, with IC50 values of493G61 pM and 455G22 pM, respectively. By contrast,the neurotransmitter inhibition curve by BoNT/A isshifted toward lower concentrations (Figure 1a) with anIC50 value of 40G16 pM with respect to the IC50 value of589G62 pM for SNAP-25 cleavage [43]. Crucially, theseresults could have a direct bearing on the number ofSNARE complexes that are necessary for one event ofneurotransmitter release at a central synapse. In fact, ifwe consider the following: (i) BoNT/A-truncated SNAP-25can form a stable and inhibitory SNARE complex;(ii) several SNARE complexes assemble together andform a multimeric SNARE ring, which is ultimatelyresponsible of the exocytic event, as shown in Figure 2;and (iii) the truncated SNAP-25–197 might prevent theassembly of the multimer SNARE ring or might give riseto a non-functional SNARE ring, then the ratio betweenthe IC50 value for the BoNT/A-dependent SNAP-25cleavage and the IC50 value for the BoNT/A inhibition ofneurotransmitter release provides the number of SNAREcomplexes that are present in a functional multimer. Evenconsidering the uncertainty resulting from the variabilityof the assays, a figure ofw15 can be derived from the dataof Ref. [43]. This estimate correlates well with the value ofw10, which can be deduced from the data of a previousstudy [42], in which it was shown that these IC50 valuesare influenced by the external calcium concentration. Thisphenomenon is not limited to synapses of the centralnervous system because BoNT/A blocks neurotransmitterrelease at the neuromuscular junction by cleaving a smallpercentage of the SNAP-25 present, which unfortunatelycould not be quantified by immunofluorescence [44].
Another possible explanation of the different resultsobtained with BoNT/A and BoNT/E proposes the presenceof sub-pools of SNAP-25 with different roles in neuro-exocytosis and different rates of BoNT/A cleavage. BoNT/Acould preferentially act on a limited (1/15–1/10 of thetotal), yet ‘functional’ SNAP-25 sub-pool, whereas BoNT/Ecleaves the entire SNAP-25 population. However, therecent finding that BoNT/A tends to localize on the

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 371
cytosolic face of the plasma membrane, whereas BoNT/Eremains mainly cytosolic [45], argues against thispossibility.
The studies discussed here [28,29,42,43] stronglysupport the view that the assembly of a multimericSNARE super-complex is crucial for exocytosis, yet failto provide a consensus on the number of complexesinvolved, which varies from 3 to 5–8 to 10–15. However,it is worthy to consider that the experimental approachesused differ in many aspects, such as the cell system (PC12versus spinal neurons), their permeabilization (intactPC12 cells and neurons versus cracked PC12 cells) andthe manipulation of the internal medium. Crucially, theauthors have examined neurotransmitter release fromdifferent secretory organelles (SVs and large dense-corevesicles), which might have different requirements interms of size and opening speed of the membrane fusioncomplex. High-order multimers of SNARE complex couldbe required in tightly regulated fast exocytosis from SV,whereas lower-order multimers might be sufficient for theslower exocytosis of dense-core granules of chromaffin andPC12 cells.
Figure 2 shows lateral and top views of a putativedodecameric SNARE super-complex obtained by modelingmethods. This analysis does not lead to a defined numberof SNARE complexes in the assembled multimer butclearly shows the crucial position of the nine-residueSNAP-25 segment removed by BoNT/A (see inset ofFigure 2). This short segment lies within an area of inter-action among SNARE complexes and close to the TMregions of VAMP and syntaxin and to the palmitoylatedcysteine quartet of SNAP-25. Figure 2 provides cluesabout how BoNTs, or an anti-SNAP-25 antibody, can acton and inhibit a loose trans-SNARE complex undergoingoligomerization to generate a multimeric SNARE super-complex but cannot when oligomerization is accomplished[46,47]. We would like to suggest that a careful compari-son on the effects of BoNT/A and BoNT/E on SNAP-25cleavage, and on the release of different neurotrans-mitters from different neurons, will provide a novel andunique tool to study the role of high-order SNAREmultimers in neuroexocytosis in intact cells.
Concluding remarks
A crucial event in exocytosis is the assembly of a rosette-shaped oligomer of SNARE complexes. The trans-mem-brane domains of VAMP/synaptobrevin and syntaxin, thepalmitoylated cysteine quartet and the C terminus ofSNAP-25 seem to be directly involved in oligomerization.There is controversy on the number, N, of SNAREcomplexes involved. We propose that N is likely to varywith the type of exocytic vesicle and that the value of N inneurons can be estimated by the use of botulinumneurotoxins type A and E by comparing their effects onSNAP-25 cleavage and on the blockade of neurotrans-mitter release.
Acknowledgements
Our work is supported by grants from: the MIURCofin ProjectMM05192773–001, FIRB RBNE01RHZM_007 and Telethon GP0272Y01
www.sciencedirect.com
(C.M.), and Cancer Research UK (G.S.) and by Fondazione Cariparo(S.P.).
References
1 Sollner, T. et al. (1993) SNAP receptors implicated in vesicle targetingand fusion. Nature 362, 318–324
2 Schiavo, G. et al. (2000) Neurotoxins affecting neuroexocytosis.Physiol. Rev. 80, 717–766
3 Simpson, L.L. (2004) Identification of the major steps in botulinumtoxin action. Annu. Rev. Pharmacol. Toxicol. 44, 167–193
4 Bonifacino, J.S. and Glick, B.S. (2004) The mechanisms of vesiclebudding and fusion. Cell 116, 153–166
5 Jahn, R. et al. (2003) Membrane fusion. Cell 112, 519–5336 Galazka, A. and Gasse, F. (1995) The present status of tetanus and
tetanus vaccination. Curr. Top. Microbiol. Immunol. 195, 31–537 Duman, J.G. and Forte, J.G. (2003) What is the role of SNARE proteins
in membrane fusion? Am. J. Physiol. Cell Physiol. 285, C237–C2498 Toonen, R.F.G. and Verhage, M. (2003) Vesicle trafficking: pleasure
and pain from SM genes. Trends Cell Biol. 13, 177–1869 Burri, L. and Lithgow, T. (2004) A complete set of SNAREs in yeast.
Traffic 5, 45–5210 de Haro, L. et al. (2004) Ca2C/calmodulin transfers the membrane-
proximal lipid-binding domain of the v-SNARE synaptobrevin from cisto trans bilayers. Proc. Natl. Acad. Sci. U. S. A. 101, 1578–1583
11 An, S.J. and Almers, W. (2004) Tracking SNARE complex formation inlive endocrine cells. Science 306, 1042–1046
12 Rickman, C. et al. (2004) High affinity interaction of syntaxin andSNAP-25 on the plasma membrane is abolished by botulinum toxin E.J. Biol. Chem. 279, 644–651
13 Bai, J. and Chapman, E.R. (2004) The C2 domains of synaptotagmin –partners in exocytosis. Trends Biochem. Sci. 29, 143–151
14 Sorensen, J.B. et al. (2003) Differential control of the releasable vesiclepools by SNAP-25 splice variants and SNAP-23. Cell 114, 75–86
15 Lin, R.C. and Scheller, R.H. (1997) Structural organization of thesynaptic exocytosis core complex. Neuron 19, 1087–1094
16 Weber, T. et al. (1998) SNAREpins: minimal machinery for membranefusion. Cell 92, 759–772
17 Bowen, M.E. et al. (2004) Single molecule observation of liposome-bilayer fusion thermally induced by soluble N-ethyl maleimidesensitive-factor attachment protein receptors (SNAREs). Biophys. J.87, 3569–3584
18 Hayashi, T. et al. (1994) Synaptic vesicle membrane fusion complex:action of clostridial neurotoxins on assembly. EMBO J. 13, 5051–5061
19 Poirier, M.A. et al. (1998) Protease resistance of syntaxin–SNAP-25–VAMP complexes. Implications for assembly and structure. J. Biol.Chem. 273, 11370–11377
20 Fasshauer, D. et al. (1998) Identification of a minimal core of thesynaptic SNARE complex sufficient for reversible assembly anddisassembly. Biochemistry 37, 10354–10362
21 Antonin, W. et al. (2000) A SNARE complex mediating fusion of lateendosomes defines conserved properties of SNARE structure andfunction. EMBO J. 19, 6453–6464
22 Tokumaru, H. et al. (2001) SNARE complex oligomerization bysynaphin/complexin is essential for synaptic vesicle exocytosis. Cell104, 421–432
23 Pabst, S. et al. (2000) Selective interaction of complexin with theneuronal SNARE complex. J. Biol. Chem. 275, 19808–19818
24 Margittai, M. et al. (1999) A stable interaction between syntaxin 1aand synaptobrevin 2 mediated by their transmembrane domains.FEBS Lett. 446, 40–44
25 Laage, R. et al. (2000) A conserved membrane-spanning amino acidmotif drives homomeric and supports heteromeric assembly ofpresynaptic SNARE proteins. J. Biol. Chem. 275, 17481–17487
26 Danieli, T. et al. (1996) Membrane fusion mediated by the influenzavirus hemagglutinin requires the concerted action of at least threehemagglutinin trimers. J. Cell Biol. 133, 559–569
27 Gibbons, D.L. et al. (2004) Conformational change and protein–protein interactions of the fusion protein of Semliki Forest virus.Nature 427, 320–325
28 Hua, Y. and Scheller, R.H. (2001) Three SNARE complexescooperate to mediate membrane fusion. Proc. Natl. Acad. Sci.U. S. A. 98, 8065–8070

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005372
29 Han, X. et al. (2004) Transmembrane segments of syntaxin line thefusion pore of Ca-triggered exocytosis. Science 304, 289–292
30 Eleopra, R. et al. (1997) Botulinum neurotoxin serotype C: a noveleffective botulinum toxin therapy in human.Neurosci. Lett. 224, 91–94
31 Eleopra, R. et al. (1998) Different time courses of recovery afterpoisoning with botulinum neurotoxin serotypes A and E in humans.Neurosci. Lett. 256, 135–138
32 Foran, P.G. et al. (2003) Evaluation of the therapeutic usefulness ofbotulinum neurotoxin B, C1, E, and F compared with the long lastingtype A. Basis for distinct durations of inhibition of exocytosis incentral neurons. J. Biol. Chem. 278, 1363–1371
33 Pellegrini, L.L. et al. (1995) Clostridial neurotoxins compromise thestability of a low energy SNARE complex mediating NSF activation ofsynaptic vesicle fusion. EMBO J. 14, 4705–4713
34 Bajohrs, M. et al. (2004) A molecular basis underlying differences inthe toxicity of botulinum serotypes A and E. EMBO Rep. 5, 1090–1095
35 Otto, H. et al. (1995) Poisoning by botulinum neurotoxin A does notinhibit formation or disassembly of the synaptosomal fusion complex.Biochem. Biophys. Res. Commun. 212, 945–952
36 Keller, J.E. et al. (1999) Persistence of botulinum neurotoxin action incultured spinal cord cells. FEBS Lett. 456, 137–142
37 Meunier, F.A. et al. (2003) Dynamics of motor nerve terminalremodeling unveiled using SNARE-cleaving botulinum toxins: theextent and duration are dictated by the sites of SNAP-25 truncation.Mol. Cell. Neurosci. 22, 454–466
38 Huang, X. et al. (1998) Truncated SNAP-25 (1-197), like botulinumneurotoxin A, can inhibit insulin secretion from HIT-T15 insulinomacells. Mol. Endocrinol. 12, 1060–1070
AGORA initiative provides free agricult
The Health Internetwork Access to Research Initiative (HINARI) of the
and Agriculture O
As part of this enterprise, Elsevier has given 185 journals to Access to
institutions are now registered for the scheme, which aims to provid
ultimately help increase crop yields and e
According to the Africa University in Zimbabwe, AGORA has been w
information to our fingertips’ says Vimbai Hungwe. ‘The information
and research activities within the University. Given the economic hard
For more inform
http://www.healthin
www.sciencedirect.com
39 Criado, M. et al. (1999) A single amino acid near the C terminusof the synaptosome-associated protein of 25 kDa (SNAP-25) isessential for exocytosis in chromaffin cells. Proc. Natl. Acad. Sci.U. S. A. 96, 7256–7261
40 Lawrence, G.W. and Dolly, J.O. (2002) Ca2C-induced changes inSNAREs and synaptotagmin I correlate with triggered exocytosisfrom chromaffin cells: insights gleaned into the signal transductionusing trypsin and botulinum toxins. J. Cell Sci. 115, 2791–2800
41 Gutierrez, L.M. et al. (1997) A peptide that mimics the C-terminalsequence of SNAP-25 inhibits secretory vesicle docking in chromaffincells. J. Biol. Chem. 272, 2634–2639
42 Keller, J.E. and Neale, E.A. (2001) The role of the synaptic proteinSNAP-25 in the potency of botulinum neurotoxin type A. J. Biol.Chem. 276, 13476–13482
43 Keller, J.E. et al. (2004) Uptake of botulinum neurotoxin into culturedneurons. Biochemistry 43, 526–532
44 Raciborska, D.A. et al. (1998) Presynaptic protein interactions in vivo:evidence from botulinum A, C, D and E action at frog neuromuscularjunction. Eur. J. Neurosci. 10, 2617–2628
45 Fernandez-Salas, E. et al. (2004) Plasma membrane localizationsignals in the light chain of botulinum neurotoxin. Proc. Natl. Acad.Sci. U. S. A. 101, 3208–3213
46 Xu, T. et al. (1998) Multiple kinetic components of exocytosisdistinguished by neurotoxin sensitivity. Nat. Neurosci. 1, 192–200
47 Xu, T. et al. (1999) Inhibition of SNARE complex assembly differen-tially affects kinetic components of exocytosis. Cell 99, 713–722
48 Chen, X. et al. (2002) Three-dimensional structure of the complexin/snare complex. Neuron 33, 397–409
ure journals to developing countries
WHO has launched a new community scheme with the UN Food
rganization.
Global Online Research in Agriculture (AGORA). More than 100
e developing countries with free access to vital research that will
ncourage agricultural self-sufficiency.
elcomed by both students and staff. ‘It has brought a wealth of
made available goes a long way in helping the learning, teaching
ships we are going through, it couldn’t have come at a better time.’
ation visit:
ternetwork.net

Ultradian metronome: timekeeper fororchestration of cellular coherenceDavid Lloyd1 and Douglas B. Murray2,3
1Microbiology (BIOSI 1), Cardiff University, P O Box 915, Cardiff, CF10 3TL, UK2Dynamics Group, Department of Biology, Beckman Research Institute of the City of Hope Medical Center, Duarte, CA 91010, USA3Present address: The Systems Biology Institute, 9S3, Shinanomachi Research Park, Keio University School of Medicine,
35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan
Dynamic intracellular spatial and temporal organization
emerges from spontaneous synchronization of a mas-
sive array of weakly coupled oscillators; the majority of
subcellular processes are implicated in this integrated
expression of cellular physiology. Evidence for this view
comes mainly from studies of Saccharomyces cerevisiae
growing in self-synchronized continuous cultures, in
which a temperature-compensated ultradian clock
(period of w40 min) couples fermentation with redox
state in addition to the transcriptome and cell-division-
cycle progression. Functions for ultradian clocks have
also been determined in other yeasts (e.g. Schizo-
saccharomyces pombe and Candida utilis), seven pro-
tists (e.g. Acanthamoeba castellanii and Paramecium
tetraurelia), as well as cultured mammalian cells. We
suggest that ultradian timekeeping is a basic universal
necessity for coordinated intracellular coherence.
Glossary
Waveform: The repetitive shape of the oscillations.
Clock: A rhythm with a temperature-compensated period.
Parallel processing: Processes carried out simultaneously, in parallel.
Period: The time between successive maxima or minima of the oscillation.
The orchestrated auto-dynamics of life
One of the most characteristic properties of livingorganisms is their inherent rhythmicity – their auto-dynamic nature [1]. Their constituent functions operateover a wide range of time scales (attoseconds to days), andon each of these time scales coordination, regulation andcontrol is paramount [2]. Their systems have to becoordinated with the external environment [3]. Organismsmust accommodate (and, even more importantly, antici-pate) changes necessitated by the 24 h cycle, the circadiancycle. Strong survival pressures (e.g. avoidance ofpredators) have ensured the ubiquity of an endogenousclock (see Glossary) that provides an anticipatory advan-tage, and is reset by dawn and by dusk.
An ultradian-timekeeping requirement is also essentialfor the maintenance of the ordered complexity of life [4];this is the internal time base upon which all the diversebiochemical events, sequences and processes must becoordinated [5,6]. On each, and between every, level of thetemporal hierarchy of intracellular performance, there is anecessity for coordinated timekeeping so that accurateconvergence of parallel processing is ensured; despite itsdynamic complexity, the whole system is maintained in astate of continuing coherence [7]. The analogy with a
Corresponding author: Lloyd, D. ([email protected]).Available online 2 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
computer is apt, but only partial, because the machine alsocreates itself anew as it performs [1]; creation requirescontinuing destruction. The continuous yeast culture wehave studied, because of its tightly synchronized popu-lation, makes this complexity accessible in vivo as neverbefore, so as to reveal the remarkably pervasive charac-teristics of an ultradian timekeeper with a period ofw40 min. We suggest that this phenomenon is anarchetype. The strict rhythmic self-organization evidentin Saccharomyces cerevisiae is required in every livingorganism. In other yeasts, the period is different, as it is inprotozoa, higher animals and in humans [6].
Continuous culture of yeast
The yeast system has many advantages; its mostimportant property is the availability of continuous read-out methods, so that the ultradian clock output can befollowed for up to several months. Even though theconditions are kept constant, the O2 consumption rate ofthe growing culture oscillates with a period of w40 min(Box 1, Figure I). When the temperature of the culture isadjusted through a daily series of discrete steps from 308Cdown to 248C, and then up to 358C, the period of therespiratory oscillation is not changed [8]. Temperaturecompensation of period length has been intensivelystudied in circadian biology and is diagnostic of physio-logical timekeeping [9]. Thus, by analogy with thecircadian clock, this 40-min cycle of O2 demand representsa conveniently observable output of a shorter periodbiological clock. As is also the case for circadian clockcontrol, maintenance of ultradian-clock output depends onsignalling mechanisms that are sensitive to the A-typemonoamine oxidase inhibitors, phenelzine and iproniazid.These compounds prevent oxidation of biologically activeamines, including neurotransmitters; yeast employs path-ways not dissimilar to those found in nervous tissue,although detailed mechanisms await elucidation. [10].Furthermore, both clocks are slowed by LiC, an inhibitor
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
Phase shifting: The wave is displaced forwards or backwards in time.
Ultradian: An oscillation with a frequency greater than a day.
. doi:10.1016/j.tibs.2005.05.005

Box 1. Continuous culture of Saccharomyces cerevisiae
In the continuous culture vessel, temperature, pH, stirring rate,
aeration rate and nutrient supply are strictly and automatically
controlled. Residual dissolved O2 in the culture broth is monitored
continuously and, because the supply rate does not vary, any
changes in O2 consumption are reflected in a reciprocal change in
dissolved O2 levels; that is, we are measuring the respiratory activity
of the organisms [47]. To obtain a continuous culture of yeast that
can bemonitored over weeks, or evenmonths, a fermentation vessel
fitted with steam-sterilizable pH and dissolved O2 electrodes, filtered
air-supply, controlled heat supply, facility for alkali (2.5M NaOH)
addition and sterilemedium reservoir is employed. After autoclaving
the assembled fermentation system, organisms are inoculated and
grown as a batch (no fresh medium is supplied). When the cultured
organisms near the onset of nutrient (glucose) and stored-energy
reserve (glycogen and trehalose) depletion, the concentration of O2
dissolved in the culture begins to oscillate [48]; a continuous supply
of nutrients is then initiated. The immersed O2 electrode provides a
stable device for continuous measurement. The output has a distinct
waveform (Figure I). The trough–peak amplitude of the dissolved O2
is approximately half of the highest level attained; this indicates that
the respiratory activity undergoes major changes during the 40-min
cycle.
0.4
0.8
1.2
1.6
80
(b)
80
(a)
80 5
10
15
20
25
(d)
0 105 1350.0
0.8
1.6
2.4
Time (min)
200
160
120
200
160
120
160
120
160
120
80
15 30 45 60 75 90 120
0.4
0.8
1.2
[NA
D(P
)H]
arbi
trar
y un
its[G
luta
thio
ne] m
M[A
ceta
ldeh
yde]
mM
[Sul
phid
e]µm
oles
/g(c
ell)/
h
[O2]
µM
(c)
Figure I. The oscillation of dissolved O2, redox state indicators and the
signalling compounds, acetaldehyde and H2S, in continuous cultures of
Saccharomyces cerevisiae. (a) NAD(P)H (heavy continuous line). (b) Intracellu-
lar glutathione concentration (GSH). (c) Extracellular acetaldehyde. (d)
Extracellular H2S. The continuous fine line indicates dissolved O2 on all panels
(a–d). Data from Refs [8,14,25].
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005374
www.sciencedirect.com
of the phosphatidylinositol signalling pathway, and boththeir periods increase in a dose-dependent manner [10].
One of the most intriguing questions posed by thisoscillating yeast culture is the basis of its coherentbehaviour. If each organism was performing all its legiontasks independently, then we would see a steady state inwhich time averaging across the population would resultin a time-independent output. In fact, the individualorganisms (total number in the culture O109) havebecome synchronized and, because the oscillations ofdissolved O2 are of such large amplitude, this synchronymust have involved a majority of the individuals [11]. Atransiently produced and rapidly eliminated volatilemessenger substance responsible for extracellular com-munication between cells of the culture is acetaldehyde, afermentation product of the yeast; the appearance of thismetabolite also shows a 40-min period (Box 1, Figure Ic).Essential to the respiratory synchrony exhibited is asudden periodic production of H2S [12,13] (a potentinhibitor of cytochrome c oxidase, the terminal componentof the mitochondrial respiratory chain). This highlyreactive gas is rapidly oxidized by the O2 present in theculture [14], and so its waveform shows a series of spikesat 40-min intervals (Box 1, Figure Id). The importance ofacetaldehyde and H2S, and possibly that of other relatedeasily diffusible chemical species, has been systematicallyshown by their capacities for phase shifting of therespiratory oscillation when they are added to a cultureat specific times during the 40-min cycles [14].
Genetic alteration of the clock
GTS1, a gene isolated from a yeast cDNA library as acandidate for a biological rhythm-related gene, affects theultradian clock [15]. It has been claimed that complete lossof rhythmicity is a consequence of GTS1 deletion [16], buta more recent study indicates a change of period of therespiratory oscillations from 40 to 18 min, occasional lossof stability and a partial loss of temperature compensation[17]. In this study, the waveform of the oscillation was alsoaffected; the onset of enhanced respiration was morerapid, and there was a slower increase to the respiratorymaximum [17].
Redox switching and mitochondrial respiratory control
Cellular O2 consumption is almost entirely carried out bymitochondria, the respiratory organelles of yeasts thatgrow under aerobic conditions. The respiratory oscilla-tions that we have observed in yeast are accompanied bychanges in the intracellular redox state, as indicated bycontinuously monitored levels of reduced nicotinamidenucleotides [18] and reduced glutathione (GSH) [19](Box 1, Figure I). These observations provide conclusiveevidence that the fluctuating levels of O2 dissolved in theculture, which indicate changing activities of mitochon-drial oxygen consumption, are characterized by redoxswitching [20]. Further evidence for this comes fromthe perturbative effects of 5-nitro-2-furaldehyde (aninhibitor of glutathione synthesis) and DL-butathionine(S,R)-sulfoximine (a g-glutamyl-cysteine inhibitor).Furthermore, additions of GSH or its oxidized form,GSSG, perturb the respiratory oscillation. Nitrosonium

11.5
1
(a)
(b)
160
120
[O2]
µM
Sca
led
expr
essi
on
0.1
[H2S
] µM
[Ace
tald
ehyd
e] m
M
12.0 12.5 13.0
Time (h)
0.0
0.5
1.0
1.5
2.0
Figure 1. The transcriptome of Saccharomyces cerevisiae in continuous culture:
activity with respect to the respiratory oscillations. (a) Dissolved O2 in the culture,
acetaldehyde and H2S. (b) Transcripts in phase with the oxidative phase of the
culture (red), and those in phase with the reductive phase (green). The scale for the
transcripts was obtained by dividing the intensity of expression of that gene at that
point by the median intensity for the 32 points obtained over the duration of the
experiment. Data derived from Ref. [28].
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 375
cations derived from added sodium nitroprusside, NaNO2,or S-nitrosoglutathione also produce characteristicchanges in the phase and amplitude of the oscillation,thiols and metalloproteins are the preferred targets [21].All these experiments confirm that the intracellularoxidation–reduction balance changes as the cycles ofchanging respiratory activity proceed. When isolatedmitochondria are incubated with excess respiratorysubstrates, they become fully activated only when furthersupplemented with ADP [22]. This activated energy-conserving state, in which ATP generation proceeds, isreferred to as the ‘energized state’. That massive changesin ionic balances across the inner mitochondrial mem-brane accompany this transition (and its reversal torestore the initial state on completion of ADP phosphoryl-ation) is well documented [23]. These characteristicalterations in the structure and functions of mitochondriain vitro provide an understanding of the changingactivities of these organelles inside growing yeasts.Thus, the accelerated respiratory rates observed inorganisms in the continuous culture when the oscillationsof dissolved O2 are at their minimal phases correspondto phases of ADP stimulation, that is, mitochondrialenergization [24,25]. Mitochondrial events that accom-pany respiratory oscillations include changes in cyto-chrome redox states, changes in the inner-mitochondrial-membrane potential and changes in conformationalstates [24].
Perturbation of mitochondrial energy conservation
Confirmation that energy conservation within mitochon-dria is closely linked to the changing rates of respirationresponsible for the respiratory oscillation came fromstudies on the effects of addition of uncouplers ofmitochondrial energy-yielding reactions (protonophores)[26]. Addition of an uncoupler S13 (5-chloro-3-t-butyl-2-chloro-4 0-nitrosalicylanilide) or CCCP (m-chlorocarbonyl-cyanide phenylhydrazone) [27] produces a succession ofchanges that suggest that the ultradian clock controlsmitochondrial redox switching, phosphorylation capacityvia energization state and cell-division-cycle progression.The dynamics of responses clearly indicate that control isnot dominated by mitochondrial events because thekinetic competence of the organelle (e.g. as manifestedby the millisecond response times of redox components inisolated organelles) is far greater than that of the observedchanges seen in mitochondria in situ in the yeasts in theculture. The slow dynamics of redox switching andchanging respiratory activity monitored in organisms incontinuous culture is imposed by biosynthetic require-ments in vivo.
Transcriptional and cell-cycle-clock outputs
Analysis of the genome-wide transcriptional activity [28]during respiratory oscillations has shown that 5329transcripts were expressed and, of these, two blocks ofredox superclusters could be assigned: (i) 650 that givemaximal expression during the phase of active respiration(the oxidative phase of the cycle), and (ii) 4679 that aremaximally observed during the ‘reductive’ phase of thecycle (Figure 1). Furthermore, of the total population of
www.sciencedirect.com
organisms in the fermenter, w5–10% became synchronouswith respect to the cell-division cycle. The onset of DNAreplication occurs in this sub-population when respirationis at is lowest during the 40-min cycle. Therefore, bothDNA and RNA synthesis occurs preferentially when thecells are in a reductive state; this might be an evolutionarystrategy to avoid oxidative damage.
The pervasive nature of the 40-min oscillation – involvingas it does, not only the metabolic network but also themembrane-associated functions of mitochondria and thetranscriptome – imply a more complex mechanism than canbe explained by a simple limit cycle of the type proposed andmodelled for a small-scale regulatory network operating in alimited portion of metabolism. Sensitivity to perturbation isa function of network size [29]; thus, the Ca2C oscillator isvery sensitive, the glycolytic oscillator less so, and thecircadian oscillator is highly robust [30]. The robustness ofthe 40-min oscillator to the external environment (pH ortemperature changes) supports this proposed complexity;a simple feedback oscillator would be intrinsically fragileto perturbation. Instead, it seems likely that the oscillationmechanism involves a massive array of weakly coupledoscillators [29,30].
The phenomenon of collective synchronization in whicha large number of oscillators spontaneously entrain to acommon frequency (although their independent naturalindividual frequencies are quite different) has beenstudied extensively [31–34]. Although explanations forsynchronicity of fire flies and populations of pacemakerneurons have been analysed mathematically [35], appli-cation to the decentralized dynamic characteristics of thegenetically controlled networks and biochemical web ofthe cell, as revealed here in a spontaneously synchronizedyeast population, awaits a new chapter in systems biology[36]. One possible model for the multi-oscillatory nature of

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005376
the cell involves operation of a controlled chaoticattractor that underlies synchronicity of the proposedarray; this would be composed of the majority of cellularprocesses [37,38].
A correlation between ultradian and circadian time-keeping is evident in several studies. The period-lengthening effects of LiC in yeast have been describedboth for ultradian [10] and circadian clocks [39]. In cray-fish, supplementation of the environment with D2Oprolonged the normal ultradian rhythmicity (as indicatedby the 169-min cycles) seen in electrical outputs recordedfrom the retina. This treatment also commensuratelyincreased the circadian periods in a dose-dependentmanner [40]. In mammalian cells cultured in vitro, thetemperature-compensated (24-min) cycles of NADHoxidase cell-surface activity (ECTO-NOX) have beenengineered to show shorter or longer periods [41]. Akidney-cell line stably transformed with cDNAs encodingcysteine to alanine replacements gives transfectants withaltered ultradian and circadian properties [41]. Theseexperiments suggest that an ultradian clock might be usedto construct a circadian clock by a process that counts theshort (40-min) intervals; this is what happens in mostman-made clocks. If this is the case, then ultradiantimekeeping becomes even more important for biology.
Concluding remarks and future work
We propose that the 40-min oscillation percolates not onlythroughout the cellular network, including organelles,transcriptome, metabolome and proteome, but alsothroughout the entire population of organisms. Thisoscillatory state is not an exceptional curiosity foundonly in a peculiar system but, rather, a universal trait thatis necessary for the maintenance of the robust metabolicauto-dynamic state characteristic of normally healthycells. We have previously obtained ultradian-clock cycletimes for ten different strains of S. cerevisiae, in additionto ten other yeast and protist species [42]. As first shownby pioneering studies on sleep, an important humanultradian cycle has a period of 90 min [43]. Aging of cells iswidely held to be due, at least in part, to an accumulationof reactive O2 species-produced damage to cellularcomponents, especially lipids and proteins [44]. In yeast,rising and falling levels of lipid peroxides accompanyrespiratory oscillations; these changes correspond to therates of respiration [45]. Because oxidative stress variesduring the mitochondrial energization cycles, an intrigu-ing hypothesis exists that an important determinant ofcell aging derives from each successive ultradian-clockcycle [46].
Future work includes further analysis of transcriptomeexpression, of proteomic and metabolome changes in yeastduring the 40-min cycle to further characterize the detailsof the oscillatory response and of time-keeping functionsthat underlie cellular integrity. Limited by the responsetimes of our monitoring devices, we have as yet elucidatedonly those that occur on time scales of minutes. Reactionsoccurring with faster dynamics require new methods. Asincreases in resolution and speed of single-cell analyses,such as fast folding green fluorescence proteins (GFP) andfluorescence resonance energy transfer (FRET) improve,
www.sciencedirect.com
the high-frequency coordination of cellular process forcoherent operation on millisecond and even more rapidtime-scales will become further uncovered. This infor-mation, together with the temporal structure of cellpopulations (e.g. in tissues), will be crucial to producevalid system-wide theories regarding the organizationalstructure of organisms.
Time structure is necessary in biological systems forthe coordination and coherence of molecular transform-ations to provide integrated functions that emerge athigher levels (membranes, organelles, cells, tissues andorganisms). The ultradian clock described here in yeastprovides the time base for this synthesis.
AcknowledgementsWe thank The Royal Society for travel grants. D.B.M. received a JST-SORST Fellowship (Japan) for part of this work.
References
1 Lloyd, D. and Gilbert, D.A. (1998) Temporal organization of the celldivision cycle of eukaryotic microbes. Symp. Soc. Gen. Microbiol. 56,251–278
2 Aon, M.K. and Cortassa, S. (1997) Dynamic Biological Organization,Chapman and Hall
3 Dunlap, J.C. et al. (2003) Chronobiology, Sinaur Associates4 Gilbert, D. and Lloyd, D. (2000) The living cell: a complex autodynamic
system. Cell Biol. Int. 24, 569–5805 Lloyd, D. et al. (2001) Why homeodynamics, not homeostasis? The
Scientific World 1, 133–1456 Lloyd, D. and Stupfel, M. (1991) The occurrence and functions of
ultradian rhythms. Biol. Rev. Camb. Philos. Soc. 66, 275–2997 Reich, J.G. and Sel’kov, E.E. (1981) Energy Metabolism of the Cell,
Academic Press8 Murray, D.B. et al. (2001) Clock control of ultradian respiratory
oscillation found during yeast continuous culture. J. Bacteriol. 183,7253–7259
9 Hastings, J.W. and Sweeney, B.M. (1957) On the mechanism oftemperature independence in a biological clock. Proc. Natl. Acad. Sci.U. S. A. 43, 804–811
10 Salgado, L.E.J. et al. (2002) Some antidepressant agents (LiC,monoamine oxidase type A inhibitors) perturb the ultradian clock inSaccharomyces cerevisiae. Biol. Rhythm Res. 33, 351–361
11 Pikovsky, A. et al. (2002) Synchronization: a Universal Concept inNonlinear Science, Cambridge University Press
12 Sohn, H. and Kuriyama, H. (2001) Ultradian metabolic oscillation ofSaccharomyces cerevisiae during aerobic continuous culture: hydro-gen sulphide, a population synchronizer is produced by sulphitereductase. Yeast 18, 125–135
13 Sohn, H.Y. et al. (2000) Ultradian oscillation of Saccharomycescerevisiae: H2S mediates population synchrony. Yeast 16, 1185–1190
14 Murray, D.B. et al. (2003) Generation and maintenance of synchronyin Saccharomyces cerevisiae continuous culture. Exp. Cell Res. 287,10–15
15 Mitsui, K. et al. (1994) The GTS1 gene which contains a Gly-Thrrepeat, affects the timing of budding and cell size of the yeastSaccharomyces cerevisiae. Mol. Cell. Biol. 14, 5569–5578
16 Wang, J. et al. (2001) Evidence for the involvement of the GTS1 geneproduct in the regulation of biological rhythms in the continuousculture of the yeast Saccharomyces cerevisiae. FEBS Lett. 489, 81–86
17 Murray, D.B. et al. (2003) The GTS1 protein stabilizes the autonomousoscillator in yeast. Yeast 20, 463–470
18 Lloyd, D. and Murray, D.B. (2000) Redox cycling of intracellular thiols:state variables for ultradian, cell division cycle and circadian cycles?In The Redox State and Circadian Rhythms (Vanden Driessche, T.et al., eds), pp. 85–94, Kluwer Academic
19 Murray, D.B. et al. (1999) Involvement of glutathione in the regulationof respiratory oscillation during a continuous culture of Saccharo-myces cerevisiae. Microbiol. 145, 2739–2745
20 Holmgren, A. (2003) The role of thioredoxin and glutaredoxin systems

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 377
in disulfide reduction and thiol redox control. In Cellular Implicationsof Redox Signalling (Gitler, C. and Dannon, A., eds), pp. 1–26,Imperial College Press
21 Murray, D.B. et al. (1998) NOC, but not NO, inhibits respiratoryoscillations in ethanol-grown chemostat cultures of Saccharomycescerevisiae. FEBS Lett. 431, 297–299
22 Chance, B. and Williams, G.R. (1956) The respiratory chain andoxidative phosphorylation. Adv. Enzymol 17, 65–134
23 Hackenbrock, C.R. (1968) Ultrastructural bases for metabolically-linked mechanical activities in mitochondria. II Electron transport-linked ultrastructure transformations in mitochondria. J. Cell Biol.37, 345–369
24 Lloyd, D. et al. (2002) Respiratory oscillations in yeast: clock-drivenmitochondrial cycles of energization. FEBS Lett. 519, 41–44
25 Lloyd, D. et al. (2002) Cycles of mitochondrial energization driven bythe ultradian clock in a continuous culture of Saccharomycescerevisiae. Microbiology 148, 3715–3724
26 Hanstein, W.G. (1976) Uncoupling of oxidative phosphorylation.Biochim. Biophys. Acta 456, 129–148
27 Lloyd, D. (2003) Effects of uncoupling of mitochondrial energyconservation on the ultradian clock-driven oscillations in Sacchar-omyces cerevisiae continuous culture. Mitochondrion 3, 139–146
28 Klevecz, R.R. et al. (2004) A genomewide oscillation in transcriptiongates DNA replication and cell cycle. Proc. Natl. Acad. Sci. U. S. A.101, 1200–1205
29 Kori, H. and Kuramoto, Y. (2001) Slow switching in globally coupledoscillators: robustness and occurrence through delayed coupling.Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 63, 046214
30 Wolf, J. et al. Analysing the robustness of cellular rhythms, SystemsBiology (in press)
31 Strogatz, S. (2003) Synch: The Emerging Science of SpontaneousOrder, Hyperion
32 Wiener N. (1961) Cybernetics (2nd edn), MIT Press33 Winfree, A.T. (1967) Biological rhythms and the behaviour of
populations of coupled oscillations. J. Theor. Biol. 16, 15–4234 Kuramoto, Y. (1984) Chemical Oscillations, Waves and Turbulence,
Springer-Verlag
The Inside Story:Readings from Tren
Edited by(Cold Spring
From its first issue in 1976, TiBS has published articles on the hist
‘historical reflections’ are fascinating stories written by those who
2005 sees the 50th anniversary of the founding of the Internationa
celebrate, a selection of these delightful articles from TiBS over the
from the earliest days of biochemistry to protein and DNA sequen
find charming – and truthful – accounts of research that are, indee
Order your copy of The Inside Story: DNA to RNA to Protein b
(US$29.00/£
www.sciencedirect.com
35 Strogatz, S.H. (2000) From Kuramoto to Crawford: exploring the onsetof synchronization in populations of coupled oscillators. Physical D143, 1–20
36 Cortassa, S. et al. (2002) An Introduction to Metabolic and CellularEngineering, World Scientific
37 Lloyd, A.L. and Lloyd, D. (1993) Hypothesis: the central oscillatorof the circadian clock is a controlled chaotic oscillator. Biosystems 29,77–85
38 Klevecz, R.R. (1992) A precise circadian clock from chaotic cell cycleoscillations. In Ultradian Rhythms in Life Processes (Lloyd, D. andRossi, E.R., eds), pp. 41–70, Springer-Verlag
39 Engelmann, W. (1973) A slowing down of circadian rhythms bylithium ions. Z. Naturforsch 28c, 733–736
40 Fuentes-Pardo, B. and Saenz, E.M. (1988) Action of deuterium oxideupon the ERG circadian rhythm in crayfish, Procambarus bouvieri.Comp. Biochem. Physiol. 90A, 435–440
41 Morre, D.J. et al. (2002) Biochemical basis for the biological clock.Biochemistry 41, 11941–11945
42 Lloyd, D. (1992) Intracellular time keeping: epigenetic oscillationsreveal the functions of an ultradian clock. In Ultradian Rhythms inLife Processes (Lloyd, D. and Rossi, E.R., eds), pp. 5–22, Springer-Verlag
43 Kleitman, N. (1982) The basic rest-activity cycle 22 years later. Sleep5, 311–315
44 Laun, P. et al. (2001) Aged mother cells of Saccharomyces cerevisiaeshow markers of oxidative stress. Mol. Microbiol. 39, 1166–1173
45 Kwak, W-J. et al. (2003) Involvement of oxidative stress in theregulation of H2S production during ultradian metabolic oscillation ofSaccharomyces cerevisiae. FEMS Microbiol. Lett. 219, 99–104
46 Lloyd, D. et al. (2003) Respiratory oscillations in yeast: mitochondrialreactive oxygen species, apoptosis and time; an hypothesis. FEMSYeast Res 3, 333–339
47 Pirt, S.J. (1975) Principles of Microbe and Cell Cultivation, BlackwellScientific
48 Satroutdinov, A.D. et al. (1992) Oscillatory metabolism of Saccharo-myces cerevisiae in continuous culture. FEMS Microbiol. Lett. 77,261–267
DNA to RNA to Proteinds in Biochemical Sciences
Jan A. WitkowskiHarbor Laboratory Press)
ory of biochemistry, and cell and molecular biology. These
did the research or by the best historians of science.
l Union of Biochemistry and Molecular Biology (IUBMB). To
years have been put together to form a collection that ranges
cing. Within The Inside Story: DNA to RNA to Protein, you will
d, the inside story.
y Jan A. Witkowski from June 2005 at www.cshlpress.com
16.95).

CIF and other mysteries of thestore-operated Ca2C-entry pathwayVictoria M. Bolotina and Peter Csutora
Ion Channel and Calcium Unit, Boston University School of Medicine, Boston, MA 02118, USA
The molecular mechanism of the store-operated
Ca2C-entry (SOCE) pathway remains one of the most
intriguing and long lasting mysteries of Ca2C signaling.
The elusive calcium influx factor (CIF) that is produced
upon depletion of Ca2C stores has attracted growing
attention, triggered by new discoveries that filled the gap
in the chain of reactions leading to activation of store-
operated channels and Ca2C entry. Ca2C-independent
phospholipase A2 emerged as a target of CIF, and a
major determinant of the SOCE mechanism. Here, we
present our viewpoint on CIF and conformational-
coupling models of SOCE from a historical perspective,
trying to resolve some of the problem areas, and sum-
marizing our present knowledge on how depletion of
intracellular Ca2C stores signals to plasma membrane
channels to open and provide Ca2C influx that is required
for many important physiological functions.
Introduction
Every cell uses Ca2C as a key messenger to regulate abroad range of its vital functions. Inside the cell, Ca2C islocalized either within specific organelles that can serve asCa2C stores [1] or in different parts of the cytoplasm,where Ca2C can freely diffuse from one location to anotheror be restricted to specific domains [2,3]. Changes in theconcentration of Ca2C ions are tightly controlled by com-plex interactions among ion channels, transporters, pumpsand binding proteins, and is finely (spatially andtemporally) tuned to modulate a variety of intracellularfunctions. To increase Ca2C concentration in the cyto-plasm, Ca2C is either released from intracellular stores(providing a transient Ca2C elevation) or is taken into hecell from extracellular space (providing a sustainedincrease in intracellular Ca2C). The endoplasmic reticu-lum (ER) is well known for its role in storage and release ofCa2C upon agonist stimulation [4], and it is also a factoryfor Ca2C-dependent protein folding and processing, andinitiation of cell-death pathways [5], making refilling ofthe ER with Ca2C after each release event vital for itsfunctional integrity and cell survival.
A store-operated Ca2C-entry (SOCE) mechanism existsthat refills ER stores, and also provides a sustained Ca2C
influx for a variety of important functions in eukaryoticcells. Among those are regulated exocytosis, regulation ofenzymatic activity (e.g. glucose metabolism, nitric oxide
Corresponding author: Bolotina, V.M. ([email protected]).Available online 13 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
and cAMP synthesis), vascular contraction and relaxa-tion, Ca2C oscillations, gene transcription, the cell cycleand apoptosis [6]. The concept of SOCE seems to besimple: upon depletion of ER stores, a signal is producedthat activates specific Ca2C-conducting channels inplasma membrane that allow Ca2C entry into the cell.Because of its fundamental importance, this mechanismhas attracted much attention. However, there is still noconsensus on how SOCE operates, what the signal is thatis produced by the stores or which plasma membranechannels are activated upon depletion of the stores(see Science STKE open forum discussion and ScienceSTKE short reviews published 27 July, 2004; http://stke.sciencemag.org). Here, we present our outlook on some ofthe possible mechanisms of SOCE.
Store-operated Ca2C-entry
The concept of SOCE was formulated by James Putneyin 1986 [7]. Originally it was called a ‘capacitative modelfor receptor regulation of Ca2C release and Ca2C entry’(Box 1), or ‘capacitative Ca2C entry’, a term which, alongwith the more up-to-date term SOCE, is still widely usedfor this important phenomenon.
To account for new findings [8,9], the original capaci-tative model was revised [10] (Box 1) and, in its final andstill-valid form, proposed that some intermediate signal-ing mechanism is required to communicate informationfrom the empty stores to plasma membrane channels toenable Ca2C to enter the cell and refill depleted stores.This model opened discussion on the nature of this elusivemechanism that has continued for the past 15 years.However, before proceeding to the mechanism linkingdepleted stores with Ca2C entry, we will first brieflyoutline and clarify some important, but rather confusingpoints, related to the store-operated channels.
Store-operated channels
By definition, a store-operated channel (SOC) is a specificchannel that is activated when Ca2C stores become empty.To be qualified as a SOC, it needs to be demonstrated thatdepletion of the stores is required and sufficient for itsactivation. Not every channel activated by agonist-induced Ca2C release from the stores will pass thesecriteria. In many cases, 1,4,5-inositol trisphosphate[Ins(1,4,5)P3], diacylglycerol, Ca
2C rise or other agonist-triggered pathways might possibly lead to activation ofCa2C-conducting channels that are different from SOCs,
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
. doi:10.1016/j.tibs.2005.05.009

Box 1. Store-operated Ca2C entry (SOCE)
In 1986, James Putney formulated the concept of the store-operated
(capacitative) Ca2C entry [7]. Originally, it was presented as a
‘capacitative model’ for receptor regulation of Ca2C release and
Ca2C entry. Citing the author, ‘[Figure Ia] represents the attempt to
summarize aspects of the model which can be empirically supported
by experimental observations: that [1,4,5-inositol trisphosphate]
Ins(1,4,5)P3 can empty a component of the endoplasmic reticulum,
the receptor-regulated Ca2C pool, and that an empty pool seems to
serve as a signal for Ca2C entry’. In this original model, incoming Ca2C
was thought to pass directly into the stores from the extracellular
space, and only then released into the cytosol through an Ins(1,4,5)P3
receptor (IP3R) opened due to the agonist stimulation. New experi-
mental evidence that accumulated over the next few years that
followed this original model showed that this is not entirely true. It was
demonstrated that Ca2C release precedes Ca2C influx, and extra-
cellular Ca2C cannot gain access directly to the intracellular Ca2C
stores, but must first enter the cytosol and be taken up from there into
the stores [8,9]. To account for these and some other findings, Putney
revised his model in 1990 [10]. In its final and still-valid form
(Figure Ib), the model postulated that ‘activation of surface membrane
receptors (R) by agonists, through the action of a guanine-nucleotide-
dependent regulatory protein leads to the activation of a polyphos-
phoinositide-specific phospholipase C (PLC) which catalyzes the
breakdown of phosphatidylinositol 4,5-bisphosphate [PtdIns(4,5)P2]
and release of Ins(1,4,5)P3. Ins(1,4,5)P3 activates a Ca2C channel (IRC)
on the organelle, leading to the initial, transient phase of Ca2C
mobilization. Secondary to the depletion of the Ca2C content of this
intracellular pool, and by an unknown mechanism, a plasma
membrane Ca2C channel is opened, allowing Ca2C entry to the
cytoplasm during the second, sustained phase of the Ca2C signal’.
This revised model [10] sparked discussion on the possible
mechanisms that might be responsible for store-dependent activation
of plasma membrane channels, which has been ongoing for more
than 15 years. Citing Putney again, ‘should we seek novel soluble
mediators, or physical connections?’ At the same time, Putney also
acknowledged that ‘it seems doubtful that capacitative Ca2C entry is
the only mechanism by which agonists increase Ca2C entry in cells. It
seems more likely that it is but one of several mechanisms by which
this might be accomplished, depending on the cell and stimulus
involved’. Indeed, other Ca2C-entry mechanisms can be activated
upon agonist-induced cell stimulation, which (because of the lack of
specific inhibitors of SOCE) were often attributed to store-operated
pathway, creating a lot of confusion in the Ca2C field. The more recent
discoveries brought more clarity, and enable us to separate the store-
operated pathway from other Ca2C influx pathways that might be
regulated in different ways, but that are all important for cell function.
R1
–
R A
Ca2+
Ca2+
Ca2+
Agonist
DG
Ins(1,4,5)P3
Ins(1,4,5)P3
Ca2+
Ca2+
Ca2+
Ca2+
PLCGp
R
Agonist
IRC
Leak
Pump
(a) (b)
Entry
channelP
tdIn
s(4,
5)P 2
Figure I. Model for store-operated Ca2C entry as proposed in 1986 (a) and in a revised form in 1990 (b). Abbreviations: DG, diacylglycerol; RA, receptor for agonist; R1,
receptor for 1,4,5-inositol trisphosphate. Part (a) reproduced, with permission, from Ref. [7]. Part (b) reproduced, with permission, from Ref. [10].
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 379
which, over the years, created a lot of uncertainty andconfusion in the SOCE field.
CRAC and SOC, or how to solve a terminological puzzle
Howmany SOCs exist and howmany are needed to satisfyphysiological needs of different types of cells? For manyyears, the assumption was that there should be only onespecific channel that is activated by Ca2C-store depletion.The first SOC was identified in mast cells [11], and wasnamed Ca2C release-activated Ca2C channel (CRAC).Because of its extremely small conductance, CRAC couldnot be resolved on the single channel level, but the whole-cell CRAC current (ICRAC) was recorded in non-excitable
www.sciencedirect.com
cells [12–14], and is rightfully considered to be the best-defined SOC current that is activated by depletion of Ca2C
stores. Does this mean that SOCs in other cell typesshould have the same CRAC-like properties? Growingevidence tells us that CRAC (which is simply a specificSOC with high Ca2C selectivity, or Ca2C-SOC; PCa2C/PNaCO1000) is not the only member of the SOC family.Indeed, when other investigators used standard pro-cedures to deplete the stores in various cell types, insteadof ICRAC they recorded inward cation currents thatdeveloped with a similar time-course and had a similarpharmacological profile, but were much less selective forCa2C[6,15]. To distinguish them from Ca2C-SOC (CRAC),

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005380
we call these cation channels ‘cat-SOCs’. Because of theirpoor cation selectivity (PCa2C/PNaCw1–50), cat-SOCscould be resolved at the single channel level givingimportant advantages in studying the mechanism oftheir operation.
Why might cells need SOCs with different relativeselectivity to Ca2C and NaC? Ca2C-SOC might be perfectfor non-excitable cells, whereas cat-SOCs might be moreuseful for excitable cells. Indeed, activation of cat-SOCmight provide not only the path for Ca2C entry, but also adepolarizing trigger to produce a secondary activation ofvoltage-gated Ca2C channels [16], which might amplifyCa2C influx required for different kinds of physiologicalresponses in excitable cells. Another possible scenario isthat NaC influx through cat-SOCs, accompanied bymembrane depolarization, would trigger secondary Ca2C
influx through the NaC–Ca2C exchanger working in areverse mode [17,18]. These are only few of the advantagesof poorly selective cat-SOCs responding to store depletion.
Thus, accumulating evidence suggests that the SOCfamily has a growing number of members that may bedivided based on their selectivity into two major cate-gories: Ca2C-SOC (or CRAC) and numerous cat-SOCs.Better classification might become possible when themolecular identity and subunit composition of each SOCis finally identified.
Molecular identity of SOCs: TRPs or not?
The molecular architecture of SOCs is largely controver-sial. The potential role of transient receptor potentialchannels (TRPs) in SOCE has attracted a lot of attention,but so far there is no consensus on which TRPs (if any) orin which specific way they contribute to the molecularstructure and/or function of SOCs. For a detailed updateon the role of TRPs in the store-operated pathway seeRefs [6,19–21]. Here, we only emphasize that accumulat-ing evidence suggests that combinations of different TRPsubunits might form ion channels with highly variablecation permeability and regulatorymechanisms, but so farnone of themmatch the exact properties of native SOCs. Itremains possible that some TRPs (or a combination ofdifferent TRPs) could be a part of the pore-forming com-plex of a SOC, which should also contain some commonstructural domain (i.e. a channel subunit or specialaccessory protein) that would be responsible for theirstore-dependent activation.
Mechanisms of SOC activation
One of the most intriguing mysteries of the store-operatedpathway is the mechanism of its activation. How do thestores communicate with the plasmamembrane channels?What is the signal produced by the stores upon their deple-tion? How is this signal created, and how is it delivered toplasma membrane? How does it open Ca2C-conductingchannels? How is the whole process of Ca2C entry ter-minated when the stores are refilled? These and manyother related questions have been asked over and overagain, motivating investigators to study this elusivemechanism.
Several attractive models have been proposed toaccount for the link between store depletion and Ca2C
www.sciencedirect.com
entry [6]. As a result of many years of intensive studies bymany different groups, some original models survived andrecently gained a new momentum, whereas others facedmajor setbacks. It is not our goal to cover all the models;instead, we concentrate on the two major ones that havebeen a point of continuous debate and confrontation formore than 15 years.
To convey information from one location (intracellularstores) to another (plasma membrane channels) (Box 1),the two easiest ways would be via a diffusible messengeror via conformational coupling. The idea of a messengerwas first formulated by Putney and colleagues [8], whoproposed that, upon store depletion, a messenger isreleased from the stores into the cytosol, where it thendiffuses to the plasma membrane and opens Ca2C
channels. Around the same time, Irvine [22] and Berridge[23] proposed that Ins(1,4,5)P3 receptors (IP3R) form aphysical bridge between the intracellular Ca2C stores andthe plasma membrane channels. These two models pro-vided elegant, but principally different, mechanisms ofstore-dependent activation of Ca2C entry, and for manyyears these models co-existed. Several recent discoveriesgave a strong preference for a diffusible messengermechanism.
Calcium influx factor
Randriamampita and Tsien [24] were the first to reportthe existence of a specific factor produced by the cellsfollowing depletion of their Ca2C stores. This factor wasnamed calcium influx factor (CIF), which is the diffusiblemessenger that is produced by Ca2C stores upon theirdepletion, and which travels to the plasma membrane andactivates SOCs and Ca2C entry. Among many benefits ofthis pioneering work was the development of a method toobtain a crude extract from donor cells, which, whenapplied to the unstimulated recipient cells, could activateCa2C entry (Table 1). Ironically, this work not onlyspurred excitement and interest in CIF model, but alsoraised many concerns and questions that could not beanswered at that time, leading to frustration and long-lasting disbelief in the whole idea of CIF. To name only few,the original extract activated Ca2C influx in recipient cellswhen applied extracellular, suggesting that CIF mighteither penetrate membranes, have an extracellulartarget or have a special mechanism way for deliveryinside the cell. In addition, in some types of recipient cells[24,25], this extract also induced Ins(1,4,5)P3-dependentCa2C release (presumably through muscarinic receptors)and, thus, might work simply by emptying the stores(far upstream from the desired CIF location in the store-operated cascade). These issues questioned the specificityand even the existence of CIF as a mechanism.
Hanley’s group [26,27] made an important subsequentstep in CIF purification; they provided evidence that thecrude extracts used in previous studies [24,25] wereindeed a complex mixture of different factors. Theyintroduced a sequence of additional purification steps[size-exclusion chromatography and high-performancethin-layer chromatography (HPTLC)], and successfullypurified a specific component of the original extract thatacted exclusively when microinjected into the cells, and

Table 1. Characterization of CIF extracts
Source of extract Bioassay system Bioassay
method
Properties Refs
Jurkat cells Macrophages Ca2C imaging Activates Ca2C rise in recipient cells when applied extracellular [24]
Fibroblasts
Astrocytoma cells
Chemical characterization done only on crude extract with
major extracellular activity
Jurkat cells,
lacrimal cells
Astrocytoma cells
Hepatocytes
Lacrimal cells
Xenopus oocytes
Ca2C imaging
ClKCa current
Contains both extracellular and intracellular activities
Activates Ca2C influx and Ca2C release
Extracellular activity is also present in extracts from
unstimulated cells
[25]
Jurkat cells Xenopus oocytes ClKCa current Intracellular activity could be isolated from crude extracts by
HPTLC and had the following characteristics: estimated size
w600 Da, resistant to heat, sensitive to phosphatase, sensitive
to phosphatase, activity potentiated by okadaic acid, does not
bind to albumin or to C18 column
[26,27]
Jurkat cells,
Saccharomyces
cerevisiae
Xenopus oocytes
Jurkat cells
Ca2C imaging
ICRAC
Contains only intracellular activity
Triggers Ca2C influx spreading through the oocyte from the
point of injection
Activates and/or accelerates ICRACEstimated size w700 Da
Binds to anion-exchange HPLC column
Activity does not require proximity of ER
[28]
Platelets,
Saccharomyces
cerevisiae
Xenopus oocytes
Smooth muscle cells
Contains only
intracellular activity
Ca2C imaging
Cat-SOC
Specific HPLC fraction found that contains only Ca2C influx,
and no Ca2C release activity
Contains only intracellular activity
Crude extract and HPLC-purified fraction activate single
cat-SOC in inside-out membrane patches
Diffuses from permeabilized platelets only after Ca2C-store
depletion
[29,47]
Saccharomyces
cerevisiae
Xenopus oocytes
RBL cells
Jurkat cells
ICRACCat-SOC
Activates ICRAC in RBL cells, and both, ICRAC and cat-SOC in
lymphocytes
Activates single cat-SOC in inside-out membrane patches
[30]
Platelets In vitro enzymatic assay
Smooth muscle cells
iPLA2 activity;
Cat-SOC
Activates iPLA2
Displaces inhibitory CaM from iPLA2
Activates single cat-SOC by displacing CaM bound to iPLA2 in
the inside-out patch
[47]
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 381
caused activation of Ca2C-dependent ClK currents (as adetector of Ca2C influx). CIF activity was absent inunstimulated cells, and appeared in a time-dependentmanner in donor cells after depletion of their stores [26],which comprised the anticipated features of CIF.Hanley et al. not only resolved some problems posed byoriginal studies but, most importantly, confirmed theauthenticity of CIF activity, provided its basic chemicalcharacterization and refined the methods for its purifi-cation (Table 1).
Further evidence for the validity of CIF model camefrom the groups of Marchase and Bolotina [28,29]. Usingadvanced methods described by Hanley’s group [26,27],and a new donor system for CIF purification (a pmr1strain of genetically modified yeast, in which Ca2C storeswere irreversibly depleted), Csutora and others [28]carefully analyzed the properties of CIF extracts obtainedfrom yeast and found them to be identical to that isolatedfrom Jurkat cells (Table 1). Using Ca2C imaging inXenopus oocytes, they confirmed that refined CIF extractscontain only intracellular activity, and monitored thespreading of the Ca2C influx originating from the point ofCIF injections that was consistent with the diffusion of aw700-Da messenger. Trepakova et al. [29] demonstratedthat CIF extract with the same properties could also beobtained from human platelets. Importantly, a further
www.sciencedirect.com
purification step was added (anion-exchange HPLC) thatenabled isolation of a specific fraction of the original crudeextract containing only Ca2C influx and no Ca2C-releaseactivity. Thus, the combined efforts and persistence of theresearch groups of Tsien, Hanley, Marchase and Bolotinaresulted in finding several good sources from which CIFcould be extracted, and developing the sequence ofpurification steps that finally enabled authentic CIFactivity to be separated from most contaminants.
In a continuous search for a better bioassay approach,CIF extracts were tested and shown to produce Ca2C
influx in Jurkat cells and Xenopus oocytes (Table 1). CIFinjection also accelerated the development of ICRAC [28,30]in Jurkat and rat basophilic leukemia (RBL) cells, andactivated Ca2C-dependent ClK currents in oocytes [26,27].Although these approaches were good for testing CIFactivity, they were not good enough to convince the toughcritics in the validity of CIF model, who requireddemonstration that CIF could indeed activate SOCs inplasma membrane.
Bolotina and coworkers [29] first validated the CIFeffect on plasma membrane channels by demonstratingthat CIF (both crude extract and HPLC-purified CIFfraction from platelets with depleted stores) is able toproduce robust activation of single cat-SOCs in inside-outmembrane patches. Moreover, they demonstrated that,

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005382
upon depletion of the stores in donor cells (permeabilizedplatelets), CIF is produced, diffuses out (only frompermeabilized, but not from intact platelets) and activatessingle cat-SOCs in inside-out membrane patches, whichserve as biological recipients of the signal. These data notonly confirm the authentic activity of CIF, but alsodemonstrate that CIF is indeed a diffusible messengerthat is produced by depleted stores and activates plasmamembrane SOC.
Table 1 summarizes all currently available informationabout CIF extracts and bioassay techniques used bydifferent investigators, including how the data wereaccumulated. Based on the present knowledge, andconfirmed by different independent studies, CIF is adiffusible non-protein messenger with molecular size ofw600 Da that is resistant to heat, alkaline pH andproteases, but can be destroyed by phosphatase andchemical treatments that target sugars and nucleotides,making it most likely to be a phosphorylated sugarnucleotide or related molecule. It is produced by ERCa2C stores upon their depletion, and activates singlecat-SOC in inside-out plasma membrane patches, whole-cell Ca2C-SOC currents (ICRAC) and Ca2C influx in avariety of bioassay systems, leaving little doubt about itsexistence and validity as a diffusible messenger.
iPLA2 – the odd enzyme that turned out to be a central
player in the SOCE pathway
So, how does CIF activate SOCs in the plasmamembrane?After the original demonstration that CIF can activatesingle cat-SOCs in inside-out membrane patches [29], itwas intuitive to think that CIF directly interacts withSOCs to open them. However, if that was the case, oneobservation could not be easily explained: after storedepletion and activation of cat-SOC in intact cells, thechannels remained active even after the membranepatches were excised and CIF was washed away [31].This led our group to suspect that an additional cascade ofplasma membrane-delimited reactions might be involvedin CIF-induced activation of SOC channels. In reviewingCa2C-independent processes that can be activated bydepletion of intracellular Ca2C stores in the presence ofstrong Ca2C chelators, the Ca2C-independent phospho-lipase A2 (iPLA2) attracted our attention because it wasreported to be activated under the same conditions asthose known to trigger activation of SOC channels [32].
iPLA2 turned out to be one of the most complex, poorlyunderstood and under-represented members of the phos-pholipase A2 family of enzymes. Its main role was thoughtto be in remodeling of cellular phospholipids [33]. Only afew devoted enthusiasts believed that iPLA2 could beinvolved in a variety of agonist-triggered signalingcascades and, over the years, they accumulated strongevidence that iPLA2 has multiple physiologically import-ant functions in numerous cell types and tissues [34–43].One of the peculiar features of a specific b isoform of iPLA2
(iPLA2b) is that it is active independent of the presenceor absence of Ca2C, but its activation is regulated bycalmodulin (CaM), which binds to iPLA2b and inhibits itsfunction [44,45]. Displacement of the inhibitory CaM leads
www.sciencedirect.com
to iPLA2 activation and production of arachidonic acid andlysophospholipids (LysoPLs).
Novel players in the store-operated game, and how they
all clicked together
The major break-through in our understanding of themolecular mechanism of the store-operated CIF-mediatedCa2C-entry pathway came from recent studies [46,47] thatdemonstrate that functional iPLA2 is required for acti-vation of SOCs and SOCE in a variety of cell types [46].Bolotina and co-investigators [47] further demonstratedthat single cat-SOC channels can be activated in amembrane-delimited manner by displacement of inhibi-tory CaM from iPLA2. Exogenously applied LysoPLs (butnot arachidonic acid) are capable of activating SOCE inintact cells and single cat-SOC in inside-out membranepatches. Importantly, experimental evidence was providedthat displacement of inhibitory CaM from iPLA2 can beproduced by CIF [47].
Based on these important findings, a novel model wasproposed [47], in which CIF-induced displacement ofinhibitory CaM from iPLA2 is the key event, leading toiPLA2 activation and generation of LysoPLs that, in turn,activate SOC channels in membrane-delimited manner.Figure 1 illustrates this model, and highlights SOC andCIF as the main determinants that are waiting for theirmolecular identification. It also follows individual steps inthe SOCE pathway (Figure 1, steps 1–10) that need to befurther investigated to provide a complete understandingof how this simple, yet complex, mechanism works. Thismodel provides a mechanism not only for activation, butalso for termination of SOC activity. Upon refilling of thestores and termination of CIF production, CaM re-binds toiPLA2 to inhibit it, thus, production of LysoPLs stops andthe activity of SOC and Ca2C influx is terminated. Thismechanism is consistent with the previously describedphysiological regulation of SOCE by nitric oxide, whichwas shown to inhibit SOCE by activation of sarco/endoplasmic reticulum calcium ATPase (SERCA)-inducedrefilling of Ca2C stores [48–50]. The crucial role of iPLA2
in SOC activation has been confirmed in platelets [46],smooth muscle cells [46,47], Jurkat T-lymphocytes [46],RBL cells [46,47] and prostate cancer epithelial cells [51].
The new CIF–iPLA2–SOC mechanism can finallyexplain some findings that have been used for years tochallenge the CIF model [52–54]. Among those, it is nowclear: (i) how CIF injected into the cells producesactivation of SOCE independently of localization of thestores and even in the absence of their depletion [28];(ii) that dialysis of cells with the Ca2C chelator BAPTAmight not only produce depletion of Ca2C stores but canalso directly activate iPLA2 by displacing inhibitory CaM,both leading to Ca2C-SOC (CRAC) activation; (iii) whySOC channels that have been pre-activated in intact cellsremain active after membrane patches have been excised[31] – when iPLA2 is already activated, CIF is not needed,and inhibitory CaM could be easily lost during membranepatch extraction; (iv) how direct application of CIF toinside-out membrane patches causes a robust activation ofSOC independently of the presence or absence of Ca2C andother cytosolic components [29]; and (v) how addition of

Figure 1. Calcium influx factor (CIF) has a key role in activation of store-operated Ca2C entry. Two separate Ca2C-entry pathways can be initiated by agonist-induced
phospholipase C (PLC)-mediated production of inositol (1,4,5)-trisphosphate [Ins(1,4,5)P3]: store-operated and Ins(1,4,5)P3-receptor (IP3R) operated. The store-operated
channel (SOC) (1) is activated when Ca2C stores in endoplasmic reticulum (ER) become empty [e.g. as a result of Ins(1,4,5)P3-induced Ca2C release]. Depleted Ca2C stores
generate a key messenger molecule, Ca2C influx factor (CIF) (2) by cleavage from a larger precursor or synthesis from a smaller precursor (3) via an unknown mechanism
(4) [see inset (b)]. CIF is released into the cytoplasm (5), where it diffuses (6) to the plasma membrane. A cascade of plasma-membrane-delimited reactions in which CIF
displaces (7) inhibitory calmodulin (CaM) from the membrane-bound Ca2C-independent phospholipase A2 (iPLA2) (8), leading to iPLA2 activation and the generation of
lysophospholipids (LysoPL) (9) that, in turn, activate SOC channels (1) either directly or via an, as yet, unknown intermediate [see inset (a)]. SOCs mediate store-operated
Ca2C entry (SOCE) (10), which might refill the stores via the Ca2C ATPase (SERCA), and also activate many important physiological processes. Despite its simplicity, this
pathway still has a lot of mysteries. Not only the molecular identities of SOCs and CIF remain unknown, but also the details of many of the steps in this pathway still need to be
established. The IP3R-operated channel (IP3ROC) (11) is activated by conformational coupling (12) with an IP3R, which is triggered by (IP3R) release of Ca2C from the stores
and ER-store depletion is neither required nor sufficient for activation of IP3ROC and the corresponding Ca2C influx (13). Ca2C entering via SOC (10) or IP3ROC (13) can be used
for a variety of cellular functions that might be different depending on which Ca2C-entry pathway is involved.
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 383
CaM inhibits active SOCs [46,47]. Moreover, it explainswhy it might be important to keep iPLA2 activity andLysoPL levels in the plasma membrane under strictcontrol. Indeed, in case of excessive activation of iPLA2
(e.g. by higher than physiological levels of CIF) over-production of LysoPLs might occur, which could disruptthe integrity of the lipid bilayer and cause additional non-specific Ca2C influx [28]. Also, the ability of CIF to displaceCaM might explain why it could be physiologicallyimportant for CIF-producing stores to be located in closeproximity to the plasma membrane where their target islocated, which was always considered important only forthe conformational-coupling model.
Discovery of a previously missing iPLA2-dependentlink in the chain of store-operated reactions has given anew, exciting twist to the CIF mechanism of SOCEactivation. Although old questions were resolved, newquestions have emerged about the details of eachindividual step in the ER–CIF–iPLA2–CaM–LysoPL–SOC pathway (see Box 2).
CIF versus other diffusible messengers
Is CIF the only diffusible messenger that can activateSOCE? Other messengers were suggested to triggerSOCE, and some were even called CIF, which resulted in
www.sciencedirect.com
a lot of confusion. Indeed, not every messenger that iscapable of activating Ca2C entry can be named CIF.This term is already reserved for a specific messengerthat, by definition, is produced by the Ca2C stores onlyupon their depletion and activates SOCE via specificSOCs. Thus, messengers that can trigger Ca2C release[e.g. Ins(1,4,5)P3, cADP-ribose, sphingosine-1-phosphate,lysophosphatidic acid], or are produced in compartmentsother than Ca2C stores, or regulate SOCs at the later steps(e.g. LysoPLs) could not be called CIF. So, the term ‘CIF’should be used with caution when referring to diffusiblemessengers that might activate Ca2C entry via a variety ofother mechanisms or serve as side regulators of the SOCEpathway.
For example, 5,6-epoxyeicosatrienoic acid (5,6-EET) – aproduct of the cytochrome P450 system (localized at theCa2C stores) – was proposed to serve as a short-lived mes-senger that activates SOCE [55–57]. However, 5,6-EETwas shown to function from outside the cells, it mightparticipate in paracrine signaling mechanisms and hasa very short half-life. Overall, it does not share thecharacteristics reported for CIF extracts [26,28] and,based on its molecular nature, cannot be present in theHPLC fractions containing authentic CIF activity [29,30];thus, 5,6-EET is an unlikely candidate for being CIF.

Box 2. Outstanding questions
† What is the molecular identity of SOCs? Although we know more
and more about the functional properties and mechanism of SOC
regulation, the molecular composition and functional arrangement
of these channels remain unknown.
† What is the molecular identity of CIF? The mechanism of the action
of CIFs has been uncovered, and its purification from numerous
cellular extracts and powerful bioassay approaches are established.
Now, the chemical structure of this elusive and most mysterious
messenger needs to be identified.
† Where and how is CIF made? It is believed that specific small
compartments of ER beneath the plasma membrane are the major
factories of CIF. However, it is unknown what is the Ca2C sensor in
ER, what is the precursor for CIF production, by what mechanism is it
converted into active CIF, and how is it released from the ER into the
cytoplasm.
† What is the molecular mechanism of CIF-induced displacement of
CaM from iPLA2? Although CIF-induced removal of inhibitory CaM
has been demonstrated, further characterization is necessary to
understand how this occurs at the molecular level, and how this
mechanism is physiologically regulated.
† Is there any functional co-localization of the main determinants of
the store-operated cascade? Close co-localization of iPLA2 and SOC
in plasma membrane is suggested based on the fact that both are
present in excised membrane patches. Lipid rafts might play an
important part in SOCE. It is attractive to speculate that iPLA2 and
SOC are located and held together in the same rafts, enabling
LysoPLs produced by iPLA2 to reach and activate the channels
quickly without disturbing the overall membrane structure.
† What is the mechanism of LysoPL action? Do LysoPLs affect the
channels directly or is yet another intermediate required for SOC
activation? These questions await answers.
† What is the inactivation mechanism of store-operated Ca2C entry?
When the stores are refilled, CIF production is terminated, and the
Ca2C entry process is turned off? It is unknown how CIF is degraded
in the cytoplasm, how LysoPLs are metabolized and if there are any
additional rearrangements in the pathway during termination of
SOCE.
† Which pathologies might be associated with impairment of CIF
production or iPLA2 function? A growing number of pathophysio-
logical conditions have been associated with, or demonstrated to be
caused by, impaired SOCE. But which parts of the CIF–iPLA2–SOC
cascade might be responsible for the development of any diseases is
not known.
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005384
Sphingosine-1-phosphate (Sp1P) was recently proposedas another CIF candidate [58]. Depletion of Ca2C storeswas found to induce Sp1P production and Ca2C influx,which was blocked by inhibition of Sp1P synthesis inHL60 cells [58]. However, Sp1P acts from outside the cells,and its injection into oocytes did not mimic the propertiesof CIF extracts [26]. The absence of store depletion inHL60 cells was not clearly verified [58], and Sp1P wasfound to release Ca2C from Ins(1,4,5)P3-sensitive stores insome other cells [59]. In addition, Sp1P does not triggerCa2C influx in some cell types that are known to haveSOCE (e.g. smooth muscle and RBL cells; V. Bolotina,unpublished observations). Overall, Sp1P does not fit themajor properties of CIF extracts [26] (Table 1), and is wellknown to participate in many signaling mechanisms thatare able to cause Ca2C entry unrelated to SOCE [60]; thus,this messenger is an unlikely candidate for CIF.
Conformational coupling
The conformational-coupling model was proposed byIrvine [22] and Berridge [23], and since had manyrevisions and variations [61,62]. The original model
www.sciencedirect.com
postulated that IP3R on the ER could be physicallyconnected to plasma membrane Ca2C-influx channelsand that information about Ca2C-store depletion couldbe translated via direct protein–protein coupling toplasma-membrane-channel opening and Ca2C entry.This elegant hypothesis was an attractive alternative tothe diffusible messenger model; over the years, manyimportant data accumulated – some of which werebelieved to support it and some to strongly contradictedit [6,61]. Here, we summarize the most recent evidencethat leads us believe that IP3R is neither required norsufficient for activation of SOCE, and, although it ispossible that conformational coupling exists, it mostprobably targets another type of channel that is operatedby IP3R and not by depletion of the stores.
Some pros and cons of the conformational-coupling
model
Although it was clear from the beginning that confor-mational coupling based on protein–protein interactions(which should provide a fast response, within milli-seconds) could not explain slow development of SOCE(which usually needs tens of seconds), the conformational-coupling model was considered, for a long time, analternative to the diffusible-messenger mechanism.
One of the most exciting findings that seemed tosupport the conformational-coupling model came fromstudies showing that IP3R can bind to TRP canonical(TRPC) channels [63–66]. But surprisingly, almost allTRPCs (not only the ones that are considered as molecularcandidates for SOC) were found to bind to IP3R, and somecan even bind to ryanodine receptors. This suggests thatTRPC–IP3R coupling might not be specific enough tooperate SOCs, or it could serve other needs that are yet tobe determined. Moreover, the specificity of such inter-action was also questioned because large signal complexeswere pulled down that include not only IP3R and TRPC1,but also SERCA, caveolin-1, phospholipase C (PLC)-b andother components [67]. At the same time, many experi-ments and conclusions favoring the conformational-coupling model were based on the assumption that2-aminoethoxydiphenyl borate (2-APB) is a specificinhibitor of IP3R. However, 2-APB was later found toinhibit SOCs directly [68–71], leaving IP3R out of theequation.
Further strong evidence supporting the conformational-coupling model was the finding that IP3R can activateTRPC3 channels and some endogenous Ca2C-conductingchannels in excised membrane patches [64,72,73]. How-ever, ryanodine receptors, Ca2C, diacylglycerol and fattyacids [6] are also capable of opening TRPC3 channels,making depletion of the stores neither required norsufficient for their activation [6]. Because it is still apoint of debate if TRPC3 [74] or any other TRP familymembers are related to SOC [6,75], the results of thesestudies leave many possibilities for alternativeinterpretations. One of which is that the channelsthat are activated by IP3R are different from SOC andmight belong to a new family of IP3R-operated cationchannels. Evidence for this possibility has started toaccumulate.

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 385
There is a growing body of evidence indicating thatIP3R might be not at all that is required for the activationof SOCs and SOCE. Indeed, cell dialysis with heparin – acompetitive antagonist of IP3R – fails to affect activation ofCa2C–SOC current (ICRAC) [6,11,13]. But the majorchallenge for the conformational-coupling model camefrom the studies in triple IP3R knockout DT40 cells, inwhich all three known types of IP3R were deleted [76].These cells did not respond to Ins(1,4,5)P3, but seemed tohave completely normal SOCE mechanism [76]. Thesefindings were confirmed and further extended by severalother groups [69–71], all of which demonstrated that IP3Rmight have an important role in agonist-induced Ca2C
influx, but not in SOCE. Furthermore, expression oftruncated IP3R, which did not produce a functionalCa2C-release channel [but did cause Ins(1,4,5)P3-inducedCa2C release and store depletion], restored Ins(1,4,5)P3-induced Ca2C influx. This confirmed that IP3R, but notdepletion of the stores, is important for IP3R-dependentCa2C influx [77]. Thus, it now seems clear that IP3Rs arenot required for activation of SOCs, and SOCE isregulated by a mechanism other than conformationalcoupling involving IP3R.
In an attempt to rescue the main idea of theconformational-coupling model (namely, the existenceand importance of plasma membrane SOC coupling toER structures that might translate Ca2C-store depletioninto SOC opening), several variations of the original modelhave been proposed. First, to account for normal ICRACand SOCE in IP3R-deficient DT40 cells, it was suggestedthat ryanodine-sensitive Ca2C-release channels mightsubstitute for IP3R in coupling to SOC and activatingCa2C influx [78]. Second, to account for the slow activationof SOCE, an interestingmodification of the conformational-coupling model was proposed [79]: the ‘secretion-likecoupling’ or ‘trafficking’ model. In this version, a part ofperipheral ER moves to the plasma membrane upon storedepletion to enable IP3R to couple to SOC (this traffickingrequires additional time). This model was based onobservations that stabilizing cortical actin cytoskeletonmight prevent ER from making a contact with the plasmamembrane, thus inhibiting activation of SOCE in some(but not all) cell types [70,79,80]. Later it was shown thatthis maneuver induces loss of channel activity (and Ca2C
entry) due to internalization of the TRPC3-signalingcomplex, not disruption of IP3R–TRPC3 interaction [81].Moreover, this model is also based on the presence offunctional IP3R, which (as discussed) is not required fornormal activation of SOCE.
SOC and IP3ROC: close relatives or strangers?
Strong evidence suggests that the conformational coup-ling is unlikely to be a mechanism of activation of SOCsand SOCE. However, it could be important for activationof some other non-store-operated channels that might playan important part in agonist-induced Ca2C entry, andtherefore requires further study. The idea that otherchannels might be conformationally coupled to IP3R issupported by recent studies [51,82,83] that have demon-strated that different types of Ca2C-influx channels, withdifferent activation modes and molecular origin, could co-
www.sciencedirect.com
exist in human prostate cancer epithelial cells [51] and incarcinoma A431 and HEK293 cells [82,83]. In epithelialcells [51], one cation current was activated by storedepletion and was dependent on iPLA2; another wasactivated by Ins(1,4,5)P3 and was insensitive to iPLA2, butwas sensitive to manipulations that might disruptconformational coupling of IP3R with the plasma mem-brane channels. The first, clearly SOC, was dependent onexpression of TRPC4 but not TRPC1, whereas the oneactivated by Ins(1,4,5)P3 was dependent on expression ofTRPC1, but not TRPC4. It is still believed that thesedifferent channels might be SOCs that are duallyregulated by CIF and IP3R-dependent conformationalcoupling.
To account for these and other experimental evidence,we recently proposed an alternative interpretation [84],and speculated that two distinct classes of Ca2C-conductingchannels might co-exist in the plasma membrane thatrespond to different signals (Figure 1): (i) SOCs, which areactivated by depletion of Ca2C stores via the CIF–iPLA2
pathway [no Ins(1,4,5)P3 needed], and (ii) IP3R-operatedchannels (IP3ROCs), which are activated by IP3R via adirect-coupling mechanism (no store-depletion required).The existence of two functionally different pathways thatactivate distinct channels could explain why the SOCEpathway works in the absence of IP3Rs in DT40 cells[69–71] and how expression of an IP3R mutant that doesnot release Ca2C from the stores can restore Ins(1,4,5)P3-induced Ca2C influx in DT40 cells [85]. Thus, twoapparently different Ca2C-influx pathways might co-exist in the same cells: one that is store-operated andanother that is operated by conformational coupling.Further studies are needed to clearly discriminate SOCand IP3ROC not only by the mode of their activation, butalso based on their biophysical properties.
Concluding remarks and future perspectives
Substantial progress has been made in recent years inunderstanding the mechanism of SOCE activation follow-ing store depletion. Presently, the diffusible-messengermodel is gaining strong support: CIF existence has beenverified by different groups, iPLA2 was found to be aplasma membrane target for CIF, a missing link in acascade of plasma membrane-delimited reactions linkingCIF with SOC activation was unveiled, and novelmolecular determinants and regulators of the store-operated pathway have been identified. Despite theexciting progress in our understanding of the overallmechanism of the store-operated pathway, the molecularidentity of the key players (CIF and SOCs) remains amystery. Further studies are needed to establish allthe details in each individual step in this importantCa2C-entry mechanism (Figure 1), and to answer manyoutstanding questions (Box 2).
References
1 Pozzan, T. et al. (1994) Molecular and cellular physiology ofintracellular calcium stores. Physiol. Rev. 74, 595–636
2 Van Breemen, C. and Saida, K. (1989) Cellular mechanisms regulating[Ca]i smooth muscle. Annu. Rev. Physiol. 51, 315–329
3 Rizzuto, R. et al. (2004) Flirting in little space: the ER/mitochondriaCa2C liaison. Sci. STKE 2004, re1

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005386
4 Berridge, M.J. (2002) The endoplasmic reticulum: a multifunctionalsignaling organelle. Cell Calcium 32, 235–249
5 Ferri, K.F. and Kroemer, G. (2001) Organelle-specific initiation of celldeath pathways. Nat. Cell Biol. 3, E255–E263
6 Parekh, A.B. and Putney, J.W., Jr. (2005) Store-operated calciumchannels. Physiol. Rev. 85, 757–810
7 Putney, J.W., Jr. (1986) A model for receptor-regulated calcium entry.
Cell Calcium 7, 1–128 Takemura, H. et al. (1989) Activation of calcium entry by the
tumor promoter thapsigargin in parotid acinar cells. Evidencethat an intracellular calcium pool and not an inositol phosphateregulates calcium fluxes at the plasma membrane. J. Biol. Chem.
264, 12266–122719 Montero, M. et al. (1991) Agonist-induced Ca2C influx in human
neutrophils is secondary to the emptying of intracellular calciumstores. Biochem. J. 277, 73–79
10 Putney, J.W., Jr. (1990) Capacitative calcium entry revisited. Cell
Calcium 11, 611–624
11 Hoth, M. and Penner, R. (1992) Depletion of intracellular calciumstores activates a calcium current in mast cells. Nature 355, 353–356
12 Hoth, M. et al. (1993) Ion channels and calcium signaling inmast cells.Ann. N. Y. Acad. Sci. 707, 198–209
13 Parekh, A.B. and Penner, R. (1997) Store depletion and calcium influx.Physiol. Rev. 77, 901–930
14 Lewis, R.S. (2001) Calcium signaling mechanisms in T lymphocytes.Annu. Rev. Immunol. 19, 497–521
15 McFadzean, I. and Gibson, A. (2002) The developing relationshipbetween receptor-operated and store-operated calcium channels insmooth muscle. Br. J. Pharmacol. 135, 1–13
16 Poteser, M. et al. (2003) Crosstalk between voltage-independent Ca2C
channels and L-yype Ca2C channels in A7r5 vascular smooth musclecells at elevated intracellular pH: evidence for functional couplingbetween L-type Ca2C channels and a 2-APB-sensitive cation channel.Circ. Res. 92, 888–896
17 Arnon, A. et al. (2000) NaC entry via store-operated channelsmodulates Ca2C signaling in arterial myocytes. Am. J. Physiol. Cell
Physiol. 278, C163–C17318 Nazer, M.A. and Van Breemen, C. (1998) Functional linkage of
Na(C)–Ca2C exchange and sarcoplasmic reticulum Ca2C release
mediates Ca2C cycling in vascular smooth muscle. Cell Calcium 24,275–283
19 Vazquez, G. et al. (2004) The mammalian TRPC cation channels.Biochim. Biophys. Acta 1742, 21–36
20 Clapham, D.E. (2003) TRP channels as cellular sensors. Nature 426,517–524
21 Nilius, B. (2003) From TRPs to SOCs, CCEs, and CRACs: consensusand controversies. Cell Calcium 33, 293–298
22 Irvine, R.F. (1990) ‘Quantal’ Ca2C release and the control of Ca2C
entry by inositol phosphates – a possible mechanism. FEBS Lett. 263,5–9
23 Berridge, M.J. (1990) Calcium oscillations. J. Biol. Chem. 265,9583–9586
24 Randriamampita, C. and Tsien, R.Y. (1993) Emptying of intracellularCa2C stores releases a novel small messenger that stimulates Ca2C
influx. Nature 364, 809–81425 Gilon, P. et al. (1995) The Ca(2C)-mobilizing actions of a Jurkat cell
extract onmammalian cells andXenopus laevis oocytes. J. Biol. Chem.
270, 8050–805526 Thomas, D. and Hanley, M.R. (1995) Evaluation of calcium influx
factors from stimulated Jurkat T– lymphocytes by microinjection into
Xenopus oocytes. J. Biol. Chem. 270, 6429–643227 Kim, H.Y. et al. (1995) Chromatographic resolution of an intracellular
calcium influx factor from thapsigargin-activated Jurkat cells.Evidence for multiple activities influencing calcium elevation inXenopus oocytes. J. Biol. Chem. 270, 9706–9708
28 Csutora, P. et al. (1999) Calcium influx factor is synthesized by yeastand mammalian cells depleted of organellar calcium stores. Proc.
Natl. Acad. Sci. U. S. A. 96, 121–12629 Trepakova, E.S. et al. (2000) Calcium influx factor (CIF) directly
activates store-operated cation channels in vascular smooth musclecells. J. Biol. Chem. 275, 26158–26163
www.sciencedirect.com
30 Su, Z. et al. (2001) A store-operated nonselective cation channel inlymphocytes is activated directly by Ca2C influx factor and diacyl-glycerol. Am. J. Physiol. Cell Physiol. 280, C1284–C1292
31 Trepakova, E.S. et al. (2001) Properties of a native cation channelactivated by Ca2C store depletion in vascular smooth muscle cells.J. Biol. Chem. 276, 7782–7790
32 Wolf, M.J. et al. (1997) Depletion of intracellular calcium stores
activates smooth muscle cell calcium-independent phospholipase A2.J. Biol. Chem. 272, 1522–1526
33 Winstead, M.V. et al. (2000) Calcium-independent phospholipase A2:structure and function. Biochim. Biophys. Acta 1488, 28–39
34 Ford, D.A. et al. (1991) The rapid and reversible activation of acalcium-independent plasmalogen-selective phospholipase A2 duringmyocardial ischemia. J. Clin. Invest. 88, 331–335
35 Lehman, J.J. et al. (1993) Arachidonic acid release from aorticsmooth muscle cells induced by [Arg8]vasopressin is largelymediated by calcium-independent phospholipase A2. J. Biol. Chem.
268, 20713–20716
36 Gross, R.W. et al. (1995) Nitric oxide activates the glucose-dependentmobilization of arachidonic acid in a macrophage-like cell line (RAW264.7) that is largely mediated by calcium-independent phospholipaseA2. J. Biol. Chem. 270, 14855–14858
37 Wolf, M.J. et al. (1995) Long-term potentiation requires activation ofcalcium-independent phospholipase A2. FEBS Lett. 377, 358–362
38 Seegers, H.C. et al. (2002) Calcium-independent phospholipaseA2-derived arachidonic acid is essential for endothelium-dependentrelaxation by acetylcholine. J. Pharmacol. Exp. Ther. 302, 918–923
39 Mancuso, D.J. et al. (2003) Cardiac ischemia activates calcium-independent phospholipase A2b, precipitating ventricular tachyar-
rhythmias in transgenic mice: rescue of the lethal electrophysiologicphenotype by mechanism-based inhibition. J. Biol. Chem. 278,22231–22236
40 Bao, S. et al. (2004) Male mice that do not express group VIAphospholipase A2 produce spermatozoa with impaired motility andhave greatly reduced fertility. J. Biol. Chem. 279, 38194–38200
41 Ramanadham, S. et al. (2004) Apoptosis of insulin-secreting cellsinduced by endoplasmic reticulum stress is amplified by overexpres-sion of group VIA calcium-independent phospholipase A2 (iPLA2b)and suppressed by inhibition of iPLA2b. Biochemistry 43, 918–930
42 Wang, Z. et al. (2004) Group VIA phospholipase A2 (iPLA2b) forms asignaling complex with the calcium/calmodulin-dependent proteinkinase IIb expressed in pancreatic islet b cells. J. Biol. Chem. 280,6840–6849
43 Ma, Z. et al. (2001) Studies of insulin secretory responses and ofarachidonic acid incorporation into phospholipids of stably transfectedinsulinoma cells that overexpress group VIA phospholipase A2(iPLA2b) indicate a signaling rather than a housekeeping role foriPLA2b. J. Biol. Chem. 276, 13198–13208
44 Wolf, M.J. and Gross, R.W. (1996) The calcium-dependent associationand functional coupling of calmodulin with myocardial phospholipase
A2. J. Biol. Chem. 271, 20989–2099245 Jenkins, C.M. et al. (2001) Identification of the calmodulin-binding
domain of recombinant calcium-independent phospholipase A2b.Implications for structure and function. J. Biol. Chem. 276, 7129–7135
46 Smani, T. et al. (2003) Ca2C-independent phospholipase A2 is anovel determinant of store-operated Ca2C entry. J. Biol. Chem. 278,11909–11915
47 Smani, T. et al. (2004) A novel mechanism for the store-operatedcalcium influx pathway. Nat. Cell Biol. 6, 113–120
48 Cohen, R.A. et al. (1999) Mechanism of nitric oxide-induced
vasodilation. Refilling of intracellular stores by sarcoplasmic reticu-lum Ca2C ATPase and inhibition of store-operated Ca2C influx. Circ.Res. 84, 210–219
49 Trepakova, E.S. et al. (1999) Nitric oxide inhibits capacitative cationinflux in human platelets by promoting sarcoplasmic/endoplasmicreticulum Ca2C-ATPase – dependent refilling of Ca2C stores. Circ.Res. 84, 201–209
50 Dedkova, E.N. and Blatter, L.A. (2002) Nitric oxide inhibitscapacitative Ca2C entry and enhances endoplasmic reticulum Ca2C
uptake in bovine vascular endothelial cells. J. Physiol. 539, 77–9151 Vanden Abeele, F. et al. (2004) Two types of store-operated Ca2C

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 387
channels with different activation modes and molecular origin inLNCaP human prostate cancer epithelial cells. J. Biol. Chem. 279,30326–30337
52 Clapham, D.E. (1995) Calcium signaling. Cell 80, 259–26853 Petersen, C.C.H. and Berridge, M.J. (1996) Capacitative calcium
entry is colocalised with calcium release in Xenopus oocytes: evidenceagainst a highly diffusable calcium influx factor. Pflugers Arch. 432,286–292
54 Jaconi, M. et al. (1997) Calcium release and influx colocalize to theendoplasmic reticulum. Curr. Biol. 7, 599–602
55 Alvarez, J. et al. (1991) Cytochrome P-450 may link intracellular Ca2C
stores with plasma membrane Ca2C influx. Biochem. J. 274, 193–19756 Rzigalinski, B.A. et al. (1999) Calcium influx factor, further evidence it
is 5,6-epoxyeicosatrienoic acid. J. Biol. Chem. 274, 175–18257 Watanabe, H. et al. (2003) Anandamide and arachidonic acid use
epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 424,434–438
58 Itagaki, K. and Hauser, C.J. (2003) Sphingosine 1-phosphate, adiffusible calcium influx factor mediating store-operated calciumentry. J. Biol. Chem. 278, 27540–27547
59 Ghosh, T.K. et al. (1994) Sphingosine 1-phosphate generated in theendoplasmic reticulum membrane activates release of stored calcium.J. Biol. Chem. 269, 22628–22635
60 Pyne, S. and Pyne, N.J. (2000) Sphingosine 1-phosphate signalling inmammalian cells. Biochem. J. 349, 385–402
61 Berridge, M.J. (1995) Capacitative calcium entry. Biochem. J. 312,1–11
62 Spassova,M.A. et al. (2004) Calcium entrymediated by SOCs and TRPchannels: variations and enigma. Biochim. Biophys. Acta 1742, 9–20
63 Boulay, G. et al. (1999) Modulation of Ca2C entry by polypeptides ofinositol 1,4,5-trisphosphate receptor (IP3R) that bind transientreceptor potential (TRP): evidence for roles of TRP and IP3R instore-depletion-activated Ca2C entry. Proc. Natl. Acad. Sci. U. S. A.96, 14955–14960
64 Kiselyov, K. et al. (1999) The N-terminal domain of the IP3 receptorgates store-operated hTrp3 channels. Mol. Cell 4, 423–429
65 Tang, J. et al. (2001) Identification of common binding sites forcalmodulin and inositol 1,4,5-trisphosphate receptors on the carboxyltermini of Trp channels. J. Biol. Chem. 276, 21303–21310
66 Rosado, J.A. and Sage, S.O. (2000) Coupling between inositol1,4,5-triphosphate receptors and human transient receptor potentialchannel 1 when intracellular Ca2C stores are depleted. Biochem. J.350, 631–635
67 Lockwich, T.P. et al. (2000) Assembly of Trp1 in a signaling complexassociated with caveolin-scaffolding lipid raft domains. J. Biol. Chem.275, 11934–11942
68 Braun, F.J. et al. (2001) Stable activation of single Ca2C release-activated Ca2C channels in divalent cation-free solutions. J. Biol.Chem. 276, 1063–1070
69 Prakriya, M. and Lewis, R.S. (2001) Potentiation and inhibition of
Free journals for dev
The WHO and six medical journal publishers have launched the Acc
poorest countries to gain free access to bio
Gro Harlem Brundtland, director-general for the WHO, said that this in
the health information gap betw
See http://www.healthinternetw
www.sciencedirect.com
Ca2C release-activated Ca2C channels by 2-aminoethyldiphenylborate (2-APB) occurs independently of IP3 receptors. J. Physiol.536, 3–19
70 Bakowski, D. et al. (2001) An examination of the secretion-likecoupling model for the activation of the Ca2C release-activated Ca2C
current ICRAC in RBL-1 cells. J. Physiol. 532, 55–7171 Ma, H.T. et al. (2001) Assessment of the role of the inositol
1,4,5-trisphosphate receptor in the activation of transient receptorpotential channels and tore-operated Ca2C entry channels. J. Biol.Chem. 276, 18888–18896
72 Kiselyov, K. et al. (1998) Functional interaction between InsP3
receptors and store-operated Htrp3 channels. Nature 396, 478–48273 Kaznacheyeva, E. et al. (2000) Plasma membrane calcium channels in
human carcinoma A431 cells are functionally coupled to inositol1,4,5-trisphosphate receptor-phosphatidylinositol 4,5-bisphosphatecomplexes. J. Biol. Chem. 275, 4561–4564
74 Hofmann, T. et al. (1999) Direct activation of human TRPC6 andTRPC3 channels by diacylglycerol. Nature 397, 259–263
75 Clapham, D.E. et al. (2001) The TRP ion channel family. Nat. Rev.Neurosci. 2, 387–396
76 Sugawara, H. et al. (1997) Genetic evidence for involvement of type 1,type 2 and type 3 inositol 1,4,5-trisphosphate receptors in signaltransduction through the B-cell antigen receptor. EMBO J. 16,3078–3088
77 Raghu, P. et al. (2000) Normal phototransduction in Drosophilaphotoreceptors lacking an InsP(3) receptor gene. Mol. Cell. Neurosci.15, 429–445
78 Kiselyov, K.I. et al. (2000) Gating of store-operated channels byconformational coupling to ryanodine receptors. Mol. Cell 6, 421–431
79 Patterson, R.L. et al. (1999) Store-operated Ca2C entry: evidence for asecretion-like coupling model. Cell 98, 487–499
80 Rosado, J.A. et al. (2000) A role for the actin cytoskeleton in theinitiation and maintenance of store-mediated calcium entry in humanplatelets. Evidence for conformational coupling. J. Biol. Chem. 275,7527–7533
81 Lockwich, T. et al. (2001) Stabilization of cortical actin inducesinternalization of transient receptor potential 3 (Trp3)-associatedcaveolar Ca2C signaling complex and loss of Ca2C influx withoutdisruption of Trp3–inositol trisphosphate receptor association. J. Biol.Chem. 276, 42401–42408
82 Bugaj, V. et al. (2005) Functional properties of endogenous receptor-and store-operated calcium influx channels in HEK293 cells. J. Biol.Chem. 280, 16790–16797
83 Gusev, K. et al. (2003) The store-operated calcium entry pathways inhuman carcinoma A431 cells: functional properties and activationmechanisms. J. Gen. Physiol. 122, 81–94
84 Bolotina, V.M. (2004) Store-operated channels: diversity and acti-vation mechanisms. Science STKE. doi: 10.1126/stke.2432004pe34
85 van Rossum, D.B. et al. (2004) Agonist-induced Ca2C entrydetermined by inositol 1,4,5-trisphosphate recognition. Proc. Natl.Acad. Sci. U. S. A. 101, 2323–2327
eloping countries
ess to Research Initiative, which enables nearly 70 of the world’s
medical literature through the Internet.
itiative was ‘perhaps the biggest step ever taken towards reducing
een rich and poor countries’.
ork.net for more information.

Major cutbacks at chromosome endsPeter M. Lansdorp
Terry Fox Laboratory, British Columbia Cancer Agency, Vancouver, BC, V5Z 1L3, Canada
Department of Medicine, University of British Columbia, Vancouver, BC, V5Z 4E3, Canada
To distinguish a telomere from a double-strand break, a
minimum number of telomere repeats must ‘cap’ each
chromosome end. The length of each repeat array will
reflect a unique history of addition and losses. Telomere
losses are predicted to occur slowly but surely with
every replication cycle (referred to as ‘typical’ telomere
loss) in addition to intermittently and, potentially,
rapidly (‘sporadic’). Recent studies have shown that
sporadic telomere losses can result from failure to
properly repair (oxidative) damage to telomeric DNA,
from failure to properly process higher-order structures
of G-rich DNA and from homologous recombination
reactions. Differences in telomere-erosion pathways
between normal and malignant cells provide novel
targets for the prevention and therapy of disease.
Introduction
Based on extensive studies by many different groups, ithas become clear that telomeres and telomerase havecrucial roles in proliferation of normal and malignantcells. The loss of telomere repeats with replication andwith age in human cells was first proposed as a molecularmechanism of replicative senescence in 1990 [1–3].Further data supporting this idea were provided instudies showing that transfer of the telomerase reversetranscriptase gene could extend the replicative lifespan ofapparently normal diploid human cells [4,5]. Since then,many papers have reported data compatible with thenotion that telomere shortening limits the number oftimes most normal human somatic cells can divide in vitroand in vivo. In humans, the telomere length in humanvascular endothelium is inversely correlated with age [6,7],and the rate of telomere loss with age in T lymphocytesexceeds that in other cell types [8]. Limitations in theproliferative potential of cells of the cardiovascular andimmune system are consistent with the recent findingthat the telomere length in human blood cells correlateswith the onset of age-dependent mortality [9]. In healthywomen, sustained psychological stress was found to beassociated with indicators of accelerated cellular andorganismal aging including reduced telomere length inblood leukocytes [10]. Interest in telomere biology isalso derived from studies of human tumors. Numerousstudies have shown that tumor cells and immortal celllines typically express high levels of telomerase [11]and are required to do so to sustain their proliferativeactivity [12].
Corresponding author: Lansdorp, P.M. ([email protected]).Available online 4 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
The main focus of telomere biology research over thepast two decades has been the mechanisms that elongatetelomeres (mainly the action of telomerase) and variousproteins such as telomere-repeat-binding factor (TRF)-1and TRF2 that regulate telomerase action [13]. Much lessattention has been paid to the mechanisms that lead toloss of telomere repeats. Indeed, telomere loss is typicallyexplained as resulting from incomplete DNA replication(the ‘end-replication problem’) [14,15], and errors inchromosome-end processing following replication [16,17]are sometimes mentioned. Studies reporting that (oxi-dative) damage to telomere repeats could be the majorcause of telomere shortening in human cells [18,19] areless frequently cited, perhaps because these findingscomplicate notions about telomere loss acting as a simple‘mitotic clock’.
Recently, two other mechanisms of telomere shorteningwere proposed: (i) the failure to unwind or correctlyprocess higher-order structures of G-rich telomeric DNA[20,21] and (ii) the deletion of T loops by homologousrecombination [22]. Ding et al. [20] reported that RTEL(regulator of telomere length), a DNA helicase-likemolecule predicted to recognize or unwind G quadruplexDNA, is required to maintain long tracts of telomererepeats in the mouse. Crabbe et al. [21] proposed that thehelicase protein deficient in patients with Werner’ssyndrome (WRN) is necessary for efficient replication ofG-rich telomeric DNA, preventing telomere dysfunctionand consequent genomic instability. Wang et al. [22],showed that a mutant allele of TRF2 can induce deletionsof telomeric DNA and it was proposed that such deletionscould also occur in normal cells. Ideally, the importance ofeach of the various telomere-attrition pathways that havebeen proposed should be established for different cell types(Table 1). This is a particularly difficult challenge not onlybecause telomerase could compensate for the loss oftelomeric DNA to different degrees in different celltypes, but also because the importance of varioustelomere-erosion pathways might vary between cells, forexample, as a function of levels of telomerase expression(see later), the length of telomere tracts, the amount ofdamage to telomeric DNA and the efficiency of variouspathways involved in the repair of such damage. Never-theless, studies in this general area should be encouragedbecause knowledge of the pathways that cause telomereattrition might result in effective strategies to minimizetelomere loss. It is possible that such strategies are moreeasily implemented than strategies aimed at elongationof telomeres that target telomerase.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
. doi:10.1016/j.tibs.2005.05.004

Table 1. Typical and sporadic loss of telomeric DNA
Mechanism of telomere loss Occurrence at individual chromosome
ends
Molecules involved Refs
The ‘end-replication problem’ Typical, occurring at ends replicated
by lagging-strand-DNA synthesis
Replication proteins [14,15]
Nucleolytic processing of 5 0 ends Typical, occurring at ends replicated
by leading-strand-DNA synthesis
Mre11–Rad50–Nbs1(Xrs2) complex;
the Ku70–Ku80 heterodimer; Exo1a
[16,17,34,36,50]
Oxidative damage, failed repair or
recombination
Sporadic, dependent on signaling
pathways and redox state in cells,
local oxygen radical production and
detoxification, mitochondrial function
[18,19,54]
T-loop deletion, rapid telomere
deletion
Sporadic XRCC3 [22,52]
Recombination following G-rich DNA
folding into stable higher-order
structures such as G-quadruplex
DNAb
Sporadic RTELa
WRNa
BACH1a
DDX11a
[20]
[21]
[79]
[80]aThe involvement of these molecules in the processing of telomeric DNA remains to be proven or clarified.bRepair of certain telomeric DNA lesions, T-loop deletion and resolution of telomeric G4 DNA could involve similar recombination reactions.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 389
Structure and function of telomeres in mammals
Telomeres, or the ends of linear chromosomes, consist inall vertebrates of tandem repeats of (TTAGGG/CCCTAA)nand associated proteins [13,23]. Telomeres typically end ina single-strand overhang at the 3 0 end, which folds backonto duplex telomeric DNA to form a ‘T-loop’ structure[24]. The length of the repeats varies between chromo-somes and between species. In humans and mice, thelength of telomere repeats at individual chromosome endsin individual cells is strikingly variable [25–27]. Humanchromosome ends are typically capped with 2–10 kilobasepairs of detectable telomere repeats depending on the typeof tissue, the age of the donor and the replicative history ofthe cells. Individual human chromosome ends showmarked variation in average telomere length, chromo-some 17p, for example, typically has one of the shortestarray of repeats [28]. Telomeres prevent the ends of linearchromosomes from appearing as DNA breaks and, thus,protect chromosome ends from degradation and fusion. Ithas been proposed that telomeres can switch between anopen state (in principle, enabling elongation by telomer-ase) and a closed state (inaccessible to telomerase) andthat the open state is inversely related to the length of therepeat tract [29]. This model is supported by recent data inyeast [30]. In most human cells, telomerase seems to bepresent at limiting levels, thereby enabling elongation ofonly a limited number of critically short telomeres. Recentdata showing a link between telomeres and DNA-damage-response proteins in senescent cells have provided furthersupport for the involvement of telomeres in the replicativesenescence of human cells [31,32].
In summary, a minimum number of repeats is requiredat every chromosome end to recruit sufficient telomere-specific proteins for the formation of a fully functionaltelomere that distinguishes a normal end from a double-strand break. Telomerase is present in limiting amountsin most human cells and cannot prevent the progressiveaccumulation of short telomeres. Apoptosis or replicativesenescence typically results when the number of criticallyshort telomeres exceeds the limited number that telomer-ase can maintain.
www.sciencedirect.com
Regulation of telomere-length
In immortal cells, the average telomere length reflects theequilibrium between telomere loss and telomere elonga-tion. The situation is more complicated at individualchromosome ends because elongation and loss eventscould differ in degree and frequency, and do not have tooccur during every cell cycle. Most of our knowledge aboutthe molecules involved in telomere-length regulation hasderived from studies of the average telomere length inunicellular organisms such as Tetrahymena, Saccharo-myces cerevisiae and S. pombe. Tetrahymena was used inthe discovery of telomerase [33] and S. cerevisiae has beeninstrumental in elucidating pathways of telomere replica-tion [34,35] and checkpoint activation [36]. Interestingly,recent studies of S. cerevisiae have shown that telomerasedoes not act on every telomere in each cell cycle and seemsto prefer short telomeres [30]. A large number (O200)of genes are known to affect telomere metabolism inS. cerevisiae [37]. Yet, telomere-length regulation in thisspecies is expected to be simple compared with that inhigher eukaryotes. The major complicating factor is thatcells in multicellular organisms belong to either repro-ductive or somatic tissues and that, within those tissues,cells are typically organized into stem cells, progenitorcells and end cells [38]. Maintenance of genome integrity(and telomeres) is most important for the cells of thegermline and the early embryo, but is probably lessefficient in various somatic cells, especially fully differ-entiated cells. This is most dramatically illustrated by thecomplete lack of nuclei in human red blood cells.Differences in the efficiency of DNA-repair pathwaysbetween (stem) cell types are increasingly being recog-nized [39–42] and could play an important part in theaging of cells and tissues. Such differences add complexityto generalizations about telomere-length regulation inmammals. To complicate things further, the situationseems to differ between mammalian species. This isillustrated by the different phenotypes of telomerasedeficiency in humans and mice. In laboratory mice,complete lack of telomerase activity is tolerated for up toseveral generations [43]. By contrast, a modest twofoldreduction in telomerase levels in humans (resulting, for

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005390
example, from haplo-insufficiency for the telomeraseRNA-template gene [23,44,45] or the human telomerasereverse transcriptase gene [46]) can result in prematuredeath, typically, from complications of aplastic anemia orimmune deficiency. Together with the age-related declinein telomere length observed in humans (but not in mice),these observations have provided support for the idea thattelomerase levels are extremely tightly regulated andlimiting in most human somatic cells. One possibility isthat telomere loss evolved as a mechanism to suppress thegrowth of tumors in long-lived species, such as humans,but not in small and short-lived animals, such as rodents[47,48]. Most studies of telomere-length regulation havefocused on telomerase. From such studies, it is knownthat regulation of telomerase occurs at the level ofsynthesis of the enzyme and via the state of its substrate– the telomere itself [13,23]. The regulation of telo-merase remains an active research area, but a detaileddiscussion of the studies in this area is beyond the scopeof this review.
Mechanisms of telomere loss
The loss of telomeric DNA can be broadly separated intotwo distinct classes: typical and sporadic (Table 1). Thetypical loss of telomeric DNA is expected to occur at everychromosome end at every replication cycle (unless theexpected loss is countered by elongation events) (Figure 1).
∗
{
∗
X
Chromosome end
+
+
3′
(a)
Figure 1. Loss of telomeric DNA with replication. Replication from internal origins results
3 0 end of chromosomes (half arrow) on the G-rich strand (red) is not fully replicated be
Following removal of the primer, a gap of a few nucleotides on the newly formed stran
primer is deposited at the very end of the chromosome terminus (box), a gap of X numbe
in a 3 0 single-strand overhang, loss of telomeric DNA with replication must also result from
(green) template strand (box). The actual amount of telomeric DNA (X plus Y) lost from
losses encountered from sporadic losses that result from DNA damage and errors in re
www.sciencedirect.com
A basic assumption in models of typical telomere loss isthat telomeric DNA is replicated from internal origins andthat DNA replication is not initiated from the very end ofchromosomes. This assumption is supported by data inyeast [49] but not (yet) in vertebrates. The typical ‘end-replication problem’ (Figure 1a) results from the failure ofconventional DNA polymerases to replicate the very 3 0 endof the chromosome [14,15]. The problem is caused bylimitations imposed by lagging-strand-DNA replication ofthe G-rich template sequences (Figure 1a) and has twocomponents. First, the removal of the RNA primer used forinitiation of DNA replication will result in a gap of a fewnucleotides in the newly formed strand. Second, assumedfailure by the replisome to deposit the last RNA primerat the very 3 0 end will result in a further extension ofthe gap (Figure 1a).
With the finding that chromosomes typically end with asingle 3 0 overhang came the realization that some form ofexonucleolytic processing must take place on the C-richtemplate strand to create a new 3 0 overhang [16,17].According to a current model, the blunt end arising fromleading-strand replication of the C template strand(Figure 1b) must be processed to yield a 3 0 overhang.The nature of the molecules involved in the recruitment,activation and arrest of the processing steps at the‘leading-strand’ telomere are topics of active research.Various molecules, including Mre11–Rad50–Nbs1(Xrs2)
Ti BS
∗
{X
{Y
+
+
5′
Chromosome end(b)
in two typical problems. (a) According to the classical ‘end-replication problem’, the
cause lagging-strand replication of the 30 strand requires a RNA primer (asterisks).
d (broken lines) will emerge. Furthermore, because it is not expected that the RNA
r of nucleotides is formed in the newly formed strand. (b) Because all telomeres end
exonucleolytic processing of a number of nucleotides (Y) of the 5 0 end of the C-rich
these typical replication problems is not known but could be small relative to the
plication, repair and recombination.

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 391
complex, the Ku70–Ku80 heterodimer, DNA-PK and Exo1have been implicated in these processes [34,36,50].
It is interesting to note that the loss of telomeric DNAin the absence of mechanisms to extend telomeres hasbeen estimated to be !10 base pairs (bp) per divisionper chromosome end in various organisms [31]. Themuch higher rate of telomere attrition observed withmost human cells (typically 50–200 bp per division)supports the idea that telomeres in human cells couldbe actively degraded [31]. At present, it is not knownif the ‘extra’ DNA is lost primarily from the typicalprocessing steps in the current model of telomerereplication (Figure 1b) or from sporadic loss oftelomeric DNA. If sporadic losses were to contributesignificantly to the overall loss of telomeric DNA,(tumor) cells that can avoid or counter such sporadicloss events are predicted to have a greatly increasedreplicative potential.
∗
∗
Resolution Holliday JunctionRestart replication fork
∗∗
X
∗∗
Helicase removal oRetry elongation bTruncation of telom
Processing Creation of
Strand invasFailure to cr
Processing 5′ endCreation of a free 3′ end
Strand invasionBreak-induced replicationHolliday Junction formation
(a) (b)
(i) (i)
(ii) (ii)
(iii) (iii)
(iv) (iv)
+
+
+
Figure 2. Speculative models of repair events following collapse of a replication fork en
internal genomic site. (b) Nick in telomeric DNA, no telomerase present. (c) Nick in telom
internal replication origins resulting in lagging-strand replication of the G-rich strand (r
between replicated and non-replicated DNA dictate that replication can only resume at D
could require extension of the 3 0 end by telomerase [exemplified by the box in (c)] before
might prevent extension of the invading 3 0 strand that could occur by break-induced repli
because a Holliday Junction cannot form. In this case, replication can only resume if t
template sequences. The likelihood of such a recombination event is predicted to depen
loss could result. DNA, solid lines; G-rich DNA, red; C-rich DNA, green; newly synthesize
lagging-strand-DNA synthesis, asterisks; replication direction (50/3 0 newly formed stra
www.sciencedirect.com
Sporadic loss of telomeric DNA
Telomere loss can also result from sporadic events such asfailed repair of damage to telomeric DNA or from problemsarising occasionally during replication, repair or recombi-nation. The study of sporadic-loss events represents achallenge because typical telomere-length measurementsare biased towards measuring the average telomerelength. Interestingly, using single telomere-length analy-sis (STELA; a PCR-based approach that accuratelymeasures the full spectrum of telomere lengths fromindividual chromosomes [27]), a significant proportion ofvariably shortened telomeres next to a major population ofaverage-sized telomeres have been observed in DNA fromhumans [27] and Caenorhabditis elegans [51]. Theseobservations support the idea that sporadic telomerelosses could contribute significantly to telomere-lengthhomeostasis in amanner that is, perhaps, similar to be therapid telomere deletion described in yeast [52].
Ti BS
∗
∗
Reolution Holliday JunctionRestart replication fork
∗∗
f invading strand?y telomerase?ere?
Processing 5′ endElongation 3′ end by telomerase
Strand invasionHolliday Junction formation
5′ enda free 3′ end
ioneate Holliday Junction
(c)
(i)
(ii)
(iii)
(iv)
+
countering a nick in the template for lagging-strand-DNA synthesis. (a) Nick at an
eric DNA, telomerase present. (i) It is assumed that telomeres are replicated from
ed) and leading-strand replication of the C-rich strand (green), and that differences
NA that has not yet been replicated. (ii) Effective repair of lesions in telomeric DNA
strand invasion (iii) because telomere-end-binding proteins such as Pot1 [X in b(iii)]
cation [a(iii)]. In the absence of telomerase, repair by recombination [b(iv)] could fail
he free end invades out of register beyond the original lesion into non-replicated
d on the length of the repeat array. With or without recombination, rapid telomere
d DNA, red and green broken lines; 30 end, half arrow; RNA primer for initiation of
nd), full arrows.

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005392
Oxidative damage to telomeric DNA
DNA is a relatively unstable molecule that can accumu-late a perplexing diversity of lesions arising fromenvironmental agents (ranging fromUV light and ionizingradiation to tobacco smoke), normal cellular metabolism(in particular, reactive oxygen species derived fromoxidative respiration and lipid peroxidation) and spon-taneous disintegration [53]. Free radicals play an import-ant part in the normal function of cells [54] and it has beenshown that telomeric G-rich DNA is 5–10-fold morevulnerable to oxidative damage than non-telomericgenomic DNA [55,56]. It is not clear, however, if oxidativelesions to telomeric DNA are actively repaired indepen-dently of DNA replication. Repair of oxidative damage tonucleotides is typically achieved using nucleotide-excisionrepair [53], but this pathway is known to be less efficientfor lesions in non-transcribed sequences [57]. Repair ofUV-light-induced pyrimidine dimers in telomeric DNAdeclined with age, and was less efficient than repair ofsimilar lesions at endogenous genes but more efficientthan that of inactive, non-coding regions [58].
Relative to genomic DNA, the repair of lesions intelomeric DNA could be complicated by several factors.First, the repeat nature of the DNA-target sequences isexpected to complicate the correct alignment of sequencesduring repair by homologous recombination [59]. Second,during typical recombinational repair, a 3 0 end invades thehomologous duplex and is extended by DNA synthesisusing the homologous template; telomere-binding proteinscould actively prevent such extension. Third, because it isnot known if replication is initiated from telomere ends,replication forks that have stalled or collapsed duringreplication of telomeric DNA, presumably, cannot berescued by convergence of replication forks [60]. Theconsequences of a replication-fork collapse at telomereshave not been studied in mammalian cells. A diversity ofreactions have been found in prokaryotes [60,61], indicat-ing a complex interplay between DNA replication,recombination and repair, and the situation at telomeresin eukaryotes is currently unclear.
Is telomerase involved in repair of (telomeric) DNA?
What are the molecular mechanisms involved in therepair of (oxidative) lesions in telomeric-DNA lesions orthe repair of replication forks stalled in telomeric DNA?Some examples of hypothetical (and speculative) DNA-repair reactions following collapse of a replication forkat a nick in the DNA template for lagging-strand-DNAsynthesis are shown in Figure 2. One potentially import-ant difference between repair events at telomeric andgenomic sequences could be the presence of proteins thatbind to telomere ends, such as Pot1 [62], that couldinterfere with a crucial step in recombinational repair –break-induced replication [63]. At telomeres (and perhapsselected sites in genomic DNA), repair by recombinationcould require the extension of a 3 0 end by telomerasebefore strand invasion (Figure 2). A role for telomerase inthe repair of telomeric DNA by recombination is consistentwith observations that support a telomere-length-inde-pendent role for telomerase [64]. Alternatively, telomerasecould be required to elongate telomeres that are critically
www.sciencedirect.com
short owing to errors in telomere repair, replication orrecombination. This mechanism was proposed to explainthe survival of murine embryonic stem cells deficient inRTEL before differentiation [20] and the survival ofnormal cells following T-loop deletion [22]. Becausesporadic losses from any cause do not have to result indetectable changes in the average telomere length, therepair of such lesions (and the survival of cells) couldrequire telomerase in a manner independent of telomerelength. Furthermore, telomerase could also be required forthe repair of lesions in genomic DNA by providing limitedextension of 3 0 G-rich sequences that cannot be extendedby break-induced replication. Such a role for telomerase inDNA repair is in agreement with the observations thatloss of the PIF1 DNA helicase in S. cerevisiae results inde novo telomere formation and gross chromosomalrearrangements [65,66]. PIF1 is postulated to dissociatetelomerase from DNA [67] and could aid in the repair ofDNA double-strand breaks by enabling telomerase toprovide limited extension of 3 0 ends while preventingde novo telomere formation. Subsequent processing bymismatch-repair pathways could ensure correct basepairing. Notably, limited extension of 3 0 ends, which isrequired for repair of telomeric and genomic DNA, couldpossibly be accomplished by telomerase variants that areincapable of net telomere elongation [64].
In summary, sporadic loss of telomeric DNA resultingfrom replication errors or from failure to properly repairdamage to telomeric DNA could, in theory, contributesignificantly to the overall loss of telomeres observed withage and with replication. Telomerase could have a rolein the repair of telomeric DNA by elongating telomeresthat have become critically short. In addition, telomerasecould play a similar part in the repair of genomic G-richsequences. Application of novel techniques, such aschromosome orientation fluorescence in situ hybridization(CO-FISH) [68,69], have revealed high frequencies of‘telomere’ sister-chromatid exchange in human andmurine cells, which is compatible with the idea thatrepair of damage to telomeric DNA could use differentpathways than those of the repair of lesions withingenomic DNA. Further studies on the replication,recombination and repair of telomeric DNA using tech-niques, such as STELA [27] and CO-FISH [68,69], areneeded to delineate the role of various DNA-repairpathways in countering sporadic telomere loss in differ-ent cell types.
Deletion of T loops
Telomeres are known hot-spots for recombination events[70] including exchanges between sister chromatids [69].Recombination between telomere sister chromatids couldpossibly enable cells to bypass a requirement for telomer-ase in repair of telomeric lesions (Figure 2b) and has beenproposed as a mechanism that could delay senescence intelomerase-negative cells [69,71]. High rates of recombi-nation have, indeed, been observed in telomerase-negativetumor cells [72] including high rates of sister-telomereexchange [73]. The T-loop structure of mammaliantelomeres is thought to provide a protective cap tochromosome ends and repress non-homologous end

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 393
joining (NHEJ) at natural chromosome ends. Recently, amutant allele of TRF2 was described that suppressedNHEJ but induced catastrophic deletions of telomericDNA [22]. The deletion events were stochastic andoccurred rapidly to generate dramatically shortenedtelomeres that were accompanied by a DNA-damageresponse and induction of senescence. T-loop-sized dele-tions were shown to require XRCC3, a protein implicatedin Holliday junction resolution, and created T-loop-sizedtelomeric circles. Telomeric circles were also detected inunperturbed cells and it was suggested that T-loop dele-tion by homologous recombination (HR) could contributeto telomere attrition in normal cells. T-loop deletions wereshown to have great preference for telomeric sequencesreplicated by leading-strand-DNA synthesis. The reasonfor this preference remains to be elucidated but it could berelated to the differential processing of leading- andlagging-strand telomeres following replication (Figure 1b).It was suggested that the T-loop deletions observed inTRF2 mutant cells are similar to the rapid telomeredeletions described in yeast [52].
Deletions of G-rich DNA
Recent observations in C. elegans [51] and mice [20]suggest that certain G-rich-DNA sequences can cause
G-rich DNA on template lagging strand
G4 forstochaon len
Resolution G4 DNANo deletions
∗
∗ ∗
∗X
DOG-1
(a)
ColRecTem
No DOG-1Polymerase switch?Lesion bypass?
Figure 3. Model of the replication problem in Caenorhabditis elegans lacking the dog-1
replicated by lagging-strand synthesis occasionally form a stable G4-DNA structure that
with the major variable being the length of the G tract. In the presence of DOG-1, the G4 D
could result in replication past the lesion. Alternatively, the replication fork could co
recombinational repair using the G-rich DNA from the sister chromatid as a template. Va
and the G4 lesion could be excised later, for example, by the nucleotide-excision-repa
structures of G-rich DNA are not expected to form, which explains why deletions occur
DNA, red; C-rich DNA, green; newly synthesized DNA, red and green broken lines; 30 en
replication direction (5 0/3 0 newly formed strand), full arrows.
www.sciencedirect.com
specific problems during replication, and it was proposedthat loss of telomeric DNA resulting from such problems isa dominant factor in the regulation of telomere length inmice [20].C. eleganswith amutation in the dog-1 (deletionof G-rich DNA-1) gene have a highly unusual mutatorphenotype characterized by short (typically 100–200nucleotides) deletions throughout their genomic DNA[51]. Strikingly, deletions invariably originate aroundthe 3 0 end of polyguanine tracts and are observed inw50% of genomic G-tracts above a threshold length ofO18 consecutive guanine residues. Because only half ofthe G-tracts show these uni-directional deletions, it wasproposed that deletions could result from failure to resolvehigher-order structures of G-rich DNA such as guaninequadruplex (G4) DNA [74], which arise stochasticallyduring lagging-strand replication of G-rich templatesequences [51] (Figure 3).
Following the observations in dog-1-deficient C. elegans,the murine Rtel gene (for regulator of telomere length)was cloned and characterized [20].Rtel encodes a helicase-like protein similar to the DOG-1 protein. It was foundthat RtelK/K mice died at day 10 or 11 of gestation withsevere proliferative defects of cells in the central nervousand cardiovascular system. Rtel-deficient embryonic stemcells were viable, but displayed telomere loss and genetic
Ti BS
mation:stic, dependinggth of G tract
No deletions
∗
∗
ssDNA
lapse replication fork?ombinational repair?plate switch?
G-rich DNA on template leading strand
(b)
gene (based on [51]). (a) Polyguanine tracks of O18 consecutive guanine residues
requires the DOG-1 protein for resolution. G4 formation is predicted to be stochastic
NA is resolved. Without DOG-1, several scenarios are possible. A polymerase switch
llapse and resume downstream. The exposed single-stranded DNA could trigger
rious molecules, including the WRN helicase, could be involved in repair reactions
ir pathway. (b) If the G tract is replicated by leading-strand synthesis higher-order
in only w50% of poly-G tracts in dog-1 mutants. Template DNA, solid lines; G-rich
d, half arrow; RNA primer for initiation of lagging-strand DNA synthesis, asterisks;

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005394
instability upon differentiation in vitro. Based on theseobservations, it was proposed that RTEL, similar toDOG-1, is required to recognize or resolve higher-orderstructures of G-rich DNA such as G4 DNA. Furtherstudies of the DOG-1 and RTEL proteins are required todetermine whether these proteins are indeed helicases aspredicted, and to study the type of structures that arerecognized and/or resolved by these proteins.
G4 DNA and telomere-length regulation
A variety of G4-DNA structures obtained from single-stranded G-rich telomeric DNA in vitro have beendescribed [75]. Several authors have proposed that suchstructures can form at the single-strand 3 0 overhangpresent at telomeres and that the formation of G4 DNA atthis site inhibits telomerase action [76,77]. Indeed, a largeamount of work has been done to identify compounds thatinhibit telomerase by favoring the formation or stabiliz-ation of G4 telomeric DNA [78]. In principle, G4 DNAcould arise at any place where single-strand G-rich DNAis exposed. At telomeres, this could be at the 3 0 end ofthe chromosome but also at the base of the T loop [24] or,indeed, anywhere along the track of telomere repeatsduring replication, repair and recombination. Specializedhelicases, such as DOG-1, RTEL, WRN and BLM(and, perhaps, also BACH1 [79] – a protein even moresimilar to DOG-1 than RTEL and DDX11, a DNA helicaserecently implicated in human telomere-length regulation[80]), could be required (with or without additionalnucleases) to resolve G4-DNA arising at telomeric andgenomic sites [81]. The BLM and WRN helicases areknown to resolve G4-DNA in vitro [82,83] and bothmolecules have been implicated in telomere-lengthregulation [21,84]. For telomere repeats, the success-ful outcome of recombination-based repair of G4-DNAlesions could be complicated by the repeat nature ofthe target sequence and by the possible involvementof the single-strand overhang in the recombinationreaction [20,52,85,86].
Concluding remarks
The notion that telomere loss in somatic cells primarilyresults from the ‘end-replication problem’ has becomeincreasingly untenable. Instead, telomere loss seems toreflect DNA losses at every chromosome end, which areinherent to replication and processing of telomeric DNA,in addition to sporadic losses of variable length, which arerelated to the repair of stalled replication forks, damage totelomeric DNA and T-loop deletion. Sporadic telomere-lossevents are expected to affect only a minority of telomeresat any given cell cycle, yet such events could be the mostimportant factor in the overall rate of telomere attrition incells. Because sporadic telomere-loss events are notreadily detectable using methods that measure theaverage telomere length, novel methods that can measureevents at single telomeres are needed to advance studiesin this area. Such studies will no doubt yield many moresurprises regarding the mechanisms of telomere loss ineukaryotic cells.
www.sciencedirect.com
AcknowledgementsI apologize to colleagues whose work could not be cited owing to spacelimitations. I thank anonymous reviewers, Gerry Krystal and themembers of my laboratory for helpful comments and discussions. Workin my laboratory is supported by grants from the National Institutes ofHealth (AI29524), the Canadian Institute of Health Research(MOP38075) and the Terry Fox Foundation.
References
1 Hastie, N.D. et al. (1990) Telomere reduction in human colorectalcarcinoma and with ageing. Nature 346, 866–868
2 Harley, C.B. et al. (1990) Telomeres shorten during ageing of humanfibroblasts. Nature 345, 458–460
3 Counter, C.M. et al. (1992) Telomere shortening associated withchromosome instability is arrested in immortal cells which expresstelomerase activity. EMBO J. 11, 1921–1929
4 Bodnar, A.G. et al. (1998) Extension of life-span by introduction oftelomerase into normal human cells. Science 279, 349–353
5 Vaziri, H. and Benchimol, S. (1998) Reconstitution of telomeraseactivity in normal human cells leads to elongation of telomeres andextended replicative life span. Curr. Biol. 8, 279–282
6 Chang, E. and Harley, C.B. (1995) Telomere length and replicativeaging in human vascular tissues. Proc. Natl. Acad. Sci. U. S. A. 92,11190–11194
7 Aviv, H. et al. (2001) Age dependent aneuploidy and telomere length ofthe human vascular endothelium. Atherosclerosis 159, 281–287
8 Rufer, N. et al. (1999) Telomere fluorescence measurements ingranulocytes and T lymphocyte subsets point to a high turnover ofhematopoietic stem cells and memory T cells in early childhood.J. Exp. Med. 190, 157–167
9 Cawthon, R.M. et al. (2003) Association between telomere length inblood and mortality in people aged 60 years or older. Lancet 361,393–395
10 Epel, E.S. et al. (2004) Accelerated telomere shortening in response tolife stress. Proc. Natl. Acad. Sci. U. S. A. 101, 17312–17315
11 Kim, N.W. et al. (1995) Specific association of human telomeraseactivity with immortal cells and cancer. Science 266, 2011–2015
12 Shay, J.W. and Roninson, I.B. (2004) Hallmarks of senescence incarcinogenesis and cancer therapy. Oncogene 23, 2919–2933
13 Smogorzewska, A. and de Lange, T. (2004) Regulation of telomerase bytelomeric proteins. Annu. Rev. Biochem. 73, 177–208
14 Watson, J.D. (1972) Origin of concatameric T4 DNA. Nat. New Biol.239, 197–201
15 Olovnikov, A.M. (1973) A theory of marginotomy. The incompletecopying of template margin in enzymic synthesis of polynucleotidesand biological significance of the phenomenon. J. Theor. Biol. 41,181–190
16 Wellinger, R.J. et al. (1996) Evidence for a new step in telomeremaintenance. Cell 85, 423–433
17 Makarov, V.L. et al. (1997) Long G tails at both ends of humanchromosomes suggest a C strand degradation mechanism for telomereshortening. Cell 88, 657–666
18 von Zglinicki, T. et al. (2000) Accumulation of single-strand breaks isthe major cause of telomere shortening in human fibroblasts. FreeRadic. Biol. Med. 28, 64–74
19 Kawanishi, S. and Oikawa, S. (2004) Mechanism of telomere short-ening by oxidative stress. Ann. N. Y. Acad. Sci. 1019, 278–284
20 Ding, H. et al. (2004) Regulation of murine telomere length by Rtel: anessential gene encoding a helicase-like protein. Cell 117, 873–886
21 Crabbe, L. et al. (2004) Defective telomere lagging strand synthesis incells lacking WRN helicase activity. Science 306, 1951–1953
22 Wang, R.C. et al. (2004) Homologous recombination generates T-loop-sized deletions at human telomeres. Cell 119, 355–368
23 Collins, K. and Mitchell, J.R. (2002) Telomerase in the humanorganism. Oncogene 21, 564–579
24 Griffith, J.D. et al. (1999) Mammalian telomeres end in a large duplexloop. Cell 97, 503–514
25 Lansdorp, P.M. et al. (1996) Heterogeneity in telomere length ofhuman chromosomes. Hum. Mol. Genet. 5, 685–691
26 Zijlmans, J.M.J.M. et al. (1997) Telomeres in the mouse have largeinter-chromosomal variations in the number of T2AG3 repeats. Proc.Natl. Acad. Sci. U. S. A. 94, 7423–7428

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 395
27 Baird, D.M. et al. (2003) Extensive allelic variation and ultrashorttelomeres in senescent human cells. Nat. Genet. 33, 203–207
28 Martens, U.M. et al. (1998) Short telomeres on human chromosome17p. Nat. Genet. 18, 76–80
29 Blackburn, E.H. (2001) Switching and signaling at the telomere. Cell106, 661–673
30 Teixeira, M.T. et al. (2004) Telomere length homeostasis is achievedvia a switch between telomerase-extendible and -nonextendiblestates. Cell 117, 323–335
31 Takai, H. et al. (2003) DNA damage foci at dysfunctional telomeres.Curr. Biol. 13, 1549–1556
32 d’Adda di Fagagna, F. et al. (2003) A DNA damage checkpointresponse in telomere-initiated senescence. Nature 426, 194–198
33 Greider, C.W. and Blackburn, E.H. (1985) Identification of a specifictelomere terminal transferase activity in Tetrhymena extracts. Cell43, 405–413
34 Lundblad, V. (2003) Telomere replication: an Est fest. Curr. Biol. 13,R439–R441
35 Makovets, S. et al. (2004) Anatomy and dynamics of DNA replicationfork movement in yeast telomeric regions. Mol. Cell. Biol. 24,4019–4031
36 Lydall, D. (2003) Hiding at the ends of yeast chromosomes: telomeres,nucleases and checkpoint pathways. J. Cell Sci. 116, 4057–4065
37 Askree, S.H. et al. (2004) A genome-wide screen for Saccharomycescerevisiae deletion mutants that affect telomere length. Proc. Natl.Acad. Sci. U. S. A. 101, 8658–8663
38 Lansdorp, P.M. (1997) Self-renewal of stem cells. Biol. Blood MarrowTransplant. 3, 171–178
39 Nouspikel, T. and Hanawalt, P.C. (2002) DNA repair in terminallydifferentiated cells. DNA Repair (Amst.) 1, 59–75
40 Mitchell, J.R. et al. (2003) Divide and conquer: nucleotide excisionrepair battles cancer and ageing. Curr. Opin. Cell Biol. 15, 232–240
41 Saretzki, G. et al. (2004) Stress defense in murine embryonic stemcells is superior to that of various differentiated murine cells. StemCells 22, 962–971
42 Miura, T. et al. (2004) Cellular lifespan and senescence signaling inembryonic stem cells. Aging Cell 3, 333–343
43 Blasco, M.A. et al. (1997) Telomere shortening and tumor formation bymouse cells lacking telomerase RNA. Cell 91, 25–34
44 Dokal, I. (2001) Dyskeratosis congenita. A disease of prematureageing. Lancet 358, S27
45 Fogarty, P.F. et al. (2003) Late presentation of dyskeratosis congenitaas apparently acquired aplastic anaemia due to mutations intelomerase RNA. Lancet 362, 1628–1630
46 Yamaguchi, H. et al. (2005) Mutations in TERT, the gene fortelomerase reverse transcriptase, in aplastic anemia. N. Engl.J. Med. 352, 1413–1424
47 Verfaillie, C.M. et al. (2002) Stem cells: hype and reality. Hematology(Am. Soc. Hematol. Educ. Program. pp. 369–391
48 Campisi, J. (2003) Cancer and ageing: rival demons?Nat. Rev. Cancer3, 339–349
49 Wellinger, R.J. and Zakian, V.A. (1989) Lack of positional require-ments for autonomously replicating sequence elements on artificialyeast chromosomes. Proc. Natl. Acad. Sci. U. S. A. 86, 973–977
50 Larrivee, M. et al. (2004) The generation of proper constitutive G-tailson yeast telomeres is dependent on the MRX complex. Genes Dev. 18,1391–1396
51 Cheung, I. et al. (2002) Disruption of dog-1 in Caenorhabditis eleganstriggers deletions upstream of guanine-rich DNA. Nat. Genet. 31,405–409
52 Lustig, A.J. (2003) Clues to catastrophic telomere loss in mammalsfrom yeast telomere rapid deletion. Nat. Rev. Genet. 4, 916–923
53 Hoeijmakers, J.H.J. (2001) Genome maintenance mechanisms forpreventing cancer. Nature 411, 366–374
54 Droge, W. (2002) Free radicals in the physiological control of cellfunction. Physiol. Rev. 82, 47–95
55 Henle, E.S. et al. (1999) Sequence-specific DNA cleavage by Fe2C-mediated fenton reactions has possible biological implications. J. Biol.Chem. 274, 962–971
56 Oikawa, S. and Kawanishi, S. (1999) Site-specific DNA damage atGGG sequence by oxidative stress may accelerate telomere short-ening. FEBS Lett. 453, 365–368
www.sciencedirect.com
57 Nouspikel, T. and Hanawalt, P.C. (2003) When parsimony backfires:neglecting DNA repair may doom neurons in Alzheimer’s disease.Bioessays 25, 168–173
58 Kruk, P.A. et al. (1995) DNA damage and repair in telomeres: relationto aging. Proc. Natl. Acad. Sci. U. S. A. 92, 258–262
59 Petes, T.D. and Hill, C.W. (1988) Recombination between repeatedgenes in microorganisms. Annu. Rev. Genet. 22, 147–168
60 McGlynn, P. and Lloyd, R.G. (2002) Recombinational repair andrestart of damaged replication forks. Nat. Rev. Mol. Cell Biol. 3,859–870
61 Marians, K.J. (2004) Mechanisms of replication fork restart inEscherichia coli. Philos. Trans. R. Soc. Lond. B Biol. Sci. 359, 71–77
62 Baumann, P. and Cech, T.R. (2001) Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 292, 1171–1175
63 Kraus, E. et al. (2001) Break-induced replication: a review and anexample in budding yeast. Proc. Natl. Acad. Sci. U. S. A. 98, 8255–8262
64 Masutomi, K. et al. (2003) Telomerase maintains telomere structure innormal human cells. Cell 114, 241–253
65 Zhou, J. et al. (2000) Pif1p helicase, a catalytic inhibitor of telomerasein yeast. Science 289, 771–774
66 Myung, K. et al. (2001) Multiple pathways cooperate in thesuppression of genome instability in Saccharomyces cerevisiae.Nature 411, 1073–1076
67 Schulz, V.P. and Zakian, V.A. (1994) The saccharomyces PIF1 DNAhelicase inhibits telomere elongation and de novo telomere formation.Cell 76, 145–155
68 Bailey, S.M. et al. (2004) Strand-specific fluorescence in situ hybrid-ization: the CO-FISH family. Cytogenet. Genome Res. 107, 14–17
69 Bailey, S.M. et al. (2004) Frequent recombination in telomeric DNAmay extend the proliferative life of telomerase-negative cells. NucleicAcids Res. 32, 3743–3751
70 Kass-Eisler, A. and Greider, C.W. (2000) Recombination in telomere-length maintenance. Trends Biochem. Sci. 25, 200–204
71 Lansdorp, P.M. (1997) Lessons from mice without telomerase. J. CellBiol. 139, 309–312
72 Neumann, A.A. and Reddel, R.R. (2002) Telomere maintenance andcancer–look, no telomerase. Nat. Rev. Cancer 2, 879–884
73 Londono-Vallejo, J.A. et al. (2004) Alternative lengthening oftelomeres is characterized by high rates of telomeric exchange.Cancer Res. 64, 2324–2327
74 Sen, D. and Gilbert, W. (1988) Formation of parallel four-strandedcomplexes by guanine-rich motifs in DNA and its implications formeiosis. Nature 334, 364–366
75 Neidle, S. and Parkinson, G.N. (2003) The structure of telomeric DNA.Curr. Opin. Struct. Biol. 13, 275–283
76 Gomez, D. et al. (2004) Interaction of telomestatin with the telomericsingle-strand overhang. J. Biol. Chem. 279, 41487–41494
77 Cuesta, J. et al. (2003) The design of G-quadruplex ligands astelomerase inhibitors. Mini Rev. Med. Chem. 3, 11–21
78 Rezler, E.M. et al. (2002) Telomeres and telomerases as drug targets.Curr. Opin. Pharmacol. 2, 415–423
79 Cantor, S.B. et al. (2001) BACH1, a novel helicase-like protein,interacts directly with BRCA1 and contributes to its DNA repairfunction. Cell 105, 149–160
80 Vasa-Nicotera, M. et al. (2005) Mapping of a major locus that deter-mines telomere length in humans. Am. J. Hum. Genet. 76, 147–151
81 Duquette, M.L. et al. (2004) Intracellular transcription of G-richDNAs induces formation of G-loops, novel structures containing G4DNA. Genes Dev. 18, 1618–1629
82 Mohaghegh, P. et al. (2001) The Bloom’s and Werner’s syndromeproteins are DNA structure-specific helicases. Nucleic Acids Res. 29,2843–2849
83 Opresko, P.L. et al. (2002) Telomere-binding protein TRF2 binds to andstimulates the Werner and Bloom syndrome helicases. J. Biol. Chem.277, 41110–41119
84 Du, X. et al. (2004) Telomere shortening exposes functions for themouse Werner and Bloom syndrome genes. Mol. Cell. Biol. 24,8437–8446
85 Zhu, X.D. et al. (2003) ERCC1/XPF removes the 3 0 overhang fromuncapped telomeres and represses formation of telomeric DNA-containing double minute chromosomes. Mol. Cell 12, 1489–1498
86 Liu, L. et al. (2002) Mitochondrial dysfunction leads to telomereattrition and genomic instability. Aging Cell 1, 40–46

Bridging gaps in phospholipidtransportDennis R. Voelker
Program in Cell Biology, Department of Medicine, National Jewish Medical and Research Center, 1400 Jackson St.,
Denver, CO 80206, USA
Phospholipid transport between membranes is a funda-
mental aspect of organelle biogenesis in eukaryotes;
however, little is know about this process. A significant
body of data demonstrates that newly synthesized
phospholipids can move betweenmembranes by routes
that are independent of the vesicular traffic that carries
membrane proteins. Evidence continues to accumulate
in support of a system for phospholipid transport that
occurs at zones of apposition and contact between
donor membranes – the source of specific phospholipids
– and acceptor membranes that are unable to synthesize
the necessary lipids. Recent findings identify some of
the lipids and proteins that must be present on
membranes for inter-organelle phospholipid transport
to occur between the endoplasmic reticulum and
mitochondria or Golgi. These data suggest that protein
and lipid assemblies on donors and acceptors promote
membrane docking and facilitate lipid movement.
Introduction
The maturation of organelles within eukaryotic cellsprimarily requires the selective transport of specificproteins and lipids to the limiting membrane and interiorof the developing structures. In the past two decades,large amounts of information and fine mechanistic detailabout protein sorting to many organelles has beenobtained [1,2]. By contrast, our understanding about theprocess of phospholipid transport for the purpose of neworganelle assembly remains small. However, recentadvances, especially the identification of mutant strainsof yeast and mammalian cells, are now providing the toolsto begin to unravel some of the complexities of phospho-lipid transport. This discipline is in the early stages ofidentifying some of the molecules involved in transportand experimenting to test their mechanisms-of-action.These endeavors are providing a framework for how a fewof these polar lipids can be transported.
One feature of intermembrane phospholipid transportthat is not widely appreciated is the resistance of theprocess to inhibitors that affect membrane protein andsecretory protein transport. This resistance is surprisingfrom several perspectives. It is well established that manymembrane and secreted proteins travel between organ-elles in vesicles whose structure is primarily defined by
Corresponding author: Voelker, D.R. ([email protected]).Available online 13 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
phospholipid [3]. Thus, some phospholipid movementbetween organelles must occur via vesicles. When theproduction of these vesicles is arrested by mutation [2] orintoxication with poisons such as brefeldin A [4,5], neitherthe lipid in the vesicle nor the membrane proteins orencapsulated cargo proteins are delivered to theirdestinations. However, in several reports the traffic ofbiosynthetically radiolabeled phospholipids betweenorganelles proceeds unabated under the same conditionsthat arrest membrane or secreted protein transport [6–9].In addition, reconstitution studies with permeabilizedcells and isolated organelles in many cases fail todemonstrate any dependence of newly synthesized phos-pholipid transport upon cytosolic factors, ATP or GTP[10–16]. These findings strongly support a mechanism forphospholipid transport between many organelles that canproceed via routes that are independent of vesicleformation, migration and fusion. It is not clear whythere should be a non-vesicular mechanism to movephospholipids between membranes or why it should beso predominant. One possibility is that non-vesicularphospholipid transport might be an evolutionarily primi-tive system for moving components between membranesthat preceded the development of vesicular mechanisms.The efficiency and efficacy of the non-vesicular transportcould be such that it has been retained as a defaultmechanism for the majority of phospholipid transport.
Progress in addressing the mechanisms of intracellularphospholipid transport in eukaryotes has been hamperedby the lack of strong genetic selections and screens, andalso convenient biochemical methods for measuring theprocesses. Despite these impediments, modest progresscontinues to be made and a growing number of mutantshave been isolated from mammalian cells and yeast thatnow implicate specific genes and their products in theprocesses [17–21]. Most importantly, the genetic advancesprovide crucially important raw materials for reconstitu-tion studies that can be used to probe the mechanisms ofphospholipid transport.
Organelle specific metabolism of
aminoglycerophospholipids
One combined biochemical and genetic approach forexamining phospholipid transport makes use of theorganelle specific metabolism of the aminoglycerophos-pholipids (Box 1), phosphatidylserine (PtdSer), phospha-tidylethanolamine (PtdEtn) and phosphatidylcholine
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
. doi:10.1016/j.tibs.2005.05.008

Box 1. Aminoglycerophospholipids
In many eukaryotic cells, including those from mammals and the yeast
Saccharomyces cerevisiae, the aminoglycerophospholipids comprise
w70–80% of the total phospholipids present in cell membranes. The
structures of these lipids are shown in Figure I. On average, intracellular
membranes in many eukaryotes have a phospholipid composition of
50% PtdCho, 10–25% PtdEtn and 1–10% PtdSer. Yeast can synthesize the
full complement of their aminoglycerophospholipids by decarboxylat-
ing PtdSer and methylating PtdEtn. The methyl groups are transferred to
PtdEtn from S-adenosylmethionine. Nucleated mammalian cells also
synthesizePtdSer and decarboxylate it to formPtdEtn.However,with the
exception of hepatocytes, mammalian cells do not synthesize significant
amounts of PtdCho from PtdEtn.
CH CH
H2C
HC
H 2C
O
O PO
O –
O
R1
O R 2
2
CO2
NH3 CH CH
H2C
HC
H2C
O
O PO
O –
O
R1
O R 2
2 N2
CH
CH
CH
3
3
3
CH CH
H2C
HC
H2C
O
O PO
O –
O
R1
O R 2
2 NH32
Phosphatidylserine Phosphatidylethanolamine Phosphatidylcholine
Figure I. Phosphatidylserine (PtdSer), phosphatidylethanolamine (PtdEtn) and phosphatidylcholine (PtdCho) comprise a family of lipids known as the aminoglycerophos-
pholipids. These lipids are closely related structurally and linked metabolically. PtdSer is decarboxylated to form PtdEtn, which is subsequently methylated to form
PtdCho. The CO2 moiety (red) of PtdSer that is removed to form PtdEtn and the methyl groups (green) that are added to PtdEtn to form PtdCho are shown.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 397
(PtdCho), as shown schematically in Figure 1. In the yeastSaccharomyces cerevisiae, PtdSer is synthesized in theendoplasmic reticulum (ER) and a subdomain of thisorganelle known as the mitochondria-associated mem-brane (MAM) [15]. After its synthesis, PtdSer is trans-ported from the ER and MAM to numerous organellesincluding the plasma membrane, mitochondria and Golgi.Upon arrival of PtdSer at the mitochondria, it is importedto the inner membrane where it becomes a substrate forPtdSer decarboxylase 1 (Psd1p; GenBank accessionnumber: NP_014230) [22], which catalyzes the conversionto PtdEtn. Transport of PtdSer to the Golgi also providesthe substrate for PtdSer decarboxylase 2 (Psd2p;NP_011686) and generates PtdEtn in this locale [23,24].In either case, the formation of PtdEtn constitutes achemical reporter for the transport of PtdSer to therespective organelles. The subsequent transport of PtdEtnout of the mitochondria or Golgi to the ER results in thefurther metabolism of a significant portion (but not all) of
PtdSer PtPsd2p
Ser PtdSerPss1p
PtdSer PtPsd1p
Mitochondri
Golgi
ER/MAM
Figure 1. The transport and metabolic itinerary of the aminoglycerophospholipids. PtdSe
associated membrane (MAM) by the action of PtdSer synthase (Pss1p). After synthesis, th
is imported to the inner membrane and decarboxylated by the Psd1p to form PtdEtn. Like
at this location. The PtdEtn is exported from either organelle to the ER, where it is sequ
PtdCho. The arrows between the organelles define major transport steps for these phosph
Psd1p and the PtdEtn is subsequently transported out of the mitochondria. In mammal
www.sciencedirect.com
this lipid to PtdCho by the combined actions of PtdEtnmethyltransferases 1 and 2 (Pem1p and Pem2p;NP_011673 and NP_012607, respectively) [25,26]. Thissynthesis of PtdCho (from what was originally PtdSer)constitutes another chemical reporter for the transport ofPtdEtn from the loci of Psd1p and Psd2p to the ER.Mammalian cells also synthesize PtdSer in a MAMfraction [27] and transport the lipid to the mitochondriawhere it is decarboxylated by Psd1p [28]. However,mammalian cells have not been reported to contain aPsd2p enzyme and, with the exception of hepatocytes, donot methylate the resultant PtdEtn to form PtdCho.Consequently, the majority of studies with mammaliancells use the action of Psd1p to examine PtdSer transportto the mitochondria [29].
Genetic screens for phospholipid transport mutants
The organelle specific metabolism of PtdSer and PtdEtnhas been used to develop genetic screens in yeast to
Ti BS
dEtn
dEtn
PtdEtn PtdCho
Pem1pPem2p
a
ER
r is synthesized in the endoplasmic reticulum (ER) and closely related mitochondria-
e PtdSer is transported to other organelles. Upon arrival at the mitochondria, PtdSer
wise, when PtdSer arrives at the Golgi it is decarboxylated by Psd2p to form PtdEtn
entially methylated in three reactions by PtdEtn methyltansferases 1 and 2 to form
olipids. All steps shown occur in yeast. In mammalian cells, PtdSer is transported to
s, only hepatocytes express significant levels of PtdEtn methyltransferase.

Ti BS
Ser PtdSer PtdSer PtdEtn PtdEtn
PtdCho
Etn
(a) PSTA and PEEA pathways
Ser PtdSer PtdSer PtdEtn PtdEtn
PtdCho
Etn
(b) PSTB and PEEB pathways
Mitochondria ER
MAM
pstAmutations
peeAmutations
Golgi ER
ER
pstBmutations
peeBmutations
Figure 2. Genetic screening in yeast for phospholipid transport mutants. (a) In psd2D mutant strains lacking Psd2p, aminoglycerophospholipid synthesis proceeds along the
PtdSer transport A and PtdEtn export A pathways (PSTA and PEEA), which involve lipid movement into and out of the mitochondria. The psd2D strains can be mutagenized
and then screened for Etn auxotrophs. Among the Etn auxotrophs will be new strains with defects in PtdSer transport (pstA mutants) and PtdEtn export (peeA mutants). The
PSTA portion of the pathway is highly active in mammalian cells and in yeast. (b). In psd1D mutant strains lacking Psd1p, aminoglycerophospholipid synthesis proceeds
along the PtdSer transport B and PtdEtn export B pathways (PSTB and PEEB), which involve lipid movement into and out of the Golgi in yeast. The psd1D strains can be
mutagenized and then screened for Etn auxotrophs. Among the Etn auxotrophs will be new strains with defects in PtdSer transport (pstB mutants) and PtdEtn export (peeB
mutants). Abbreviations: ER, endoplasmic reticulum; MAM, mitochondria-associated membrane.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005398
identify mutations, genes and proteins that participate inlipid transport. The general features of this screening areoutlined in Figure 2. In yeast that have been manipulatedto delete the PSD1gene encoding Psd1p (i.e. strains with apsd1D mutation), or to delete the PSD2 gene encodingPsd2p (i.e. strains with a psd2D mutation), growth isrelatively normal in minimal media [22–24]. By contrast,when doubly mutated psd1D psd2D strains are con-structed, they are unable to synthesize sufficient PtdEtnfor survival in minimal media [23]. However, like manyeukaryotes, yeasts possess multiple pathways for thesynthesis of PtdEtn [30]. One of these pathways, usuallycalled the Kennedy pathway in recognition of the seminalcontributions of Eugene P. Kennedy, enables yeast to useethanolamine (Etn) to synthesize PtdEtn. Empirically, onefinds that the psd1D psd2D double mutant strains of yeastcan be rescued by Etn supplementation [23]. From thesefindings, Trotter and Voelker postulated that if Etn canrescue PtdEtn deficiency due to inactive Psd1p and Psd2pactivity, then it might also rescue PtdEtn deficiency due toa defect in transport of PtdSer to the loci of either Psd1p orPsd2p [18]. From a theoretical standpoint and forsimplicity, the transport of PtdSer to Psd1p was namedthe PtdSer transport A (PSTA) pathway and the transportof PtdSer to Psd2p was named the PtdSer transport B(PSTB) pathway. Likewise, newly formed PtdEtn must beexported from the mitochondria and Golgi back to the ERfor the synthesis of PtdCho and these pathways are namedPEEA and PEEB.
www.sciencedirect.com
In execution, the screen for yeast mutants defective inPtdSer transport to Psd1p uses strains harboring a psd2Dmutation that are mutagenized and analyzed for Etnauxotrophy [20]. The psd2D genetic background forcesalmost all PtdEtn synthesis to proceed via transport ofPtdSer to Psd1p, as shown in Figure 2a. The mutation ofpsd2D strains and screening for Etn auxotrophs enablesthe identification of new strains defective in the transportof PtdSer to Psd1p, in addition to strains with defects inthe activity of Psd1p. In yeast, this screen has produced amutant named pstA1 (described in more detail later) andits corresponding complementing gene. Similar types ofanalyses can be performed with strains that have a psd1Dmutation, which are dependent on the transport of PtdSerto Psd2p for the synthesis of the majority of their PtdEtn.Mutation and screening for Etn auxotrophs in strains witha psd1Dmutation enables the identification of new strainsthat are defective in transport of PtdSer to Psd2p, or thatare defective in the activity of this enzyme, as illustratedin Figure 2b. Thus far, this second screen has yieldedtwo mutants (pstB1 and pstB2) and their correspondingcomplementing genes, in addition to a variant of Psd2p,that have provided important new insight into the processof PtdSer transport [16,18,19,21]. In principle, both ofthese screens should also yield new mutants (peeA andpeeB) in the export of PtdEtn from the mitochondria andGolgi back to the ER.
A screen related to that described previously for thePSTA pathway has also been applied to mammalian cells.

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 399
This screen was designed to identify strains that have aplasma membrane deficiency in PtdEtn and are resistantto the effects of a toxin that binds the lipid. The toxin isthe cyclic peptide Ro09–0198, which recognizes PtdEtnand causes cytolysis. This screen produced a variant ofCHO-K1 cells (R41) that is defective in PtdSer transport tothe locus of Psd1p and results in a deficiency in cellularPtdEtn [17].
Biochemical studies
In addition to the genetic approach, biochemical studieswith intact cells, permeabilized cells and isolated organ-elles have used the action of Psd1p and Psd2p to examineand characterize PtdSer transport. For lipid transport toPsd1p, yeast and mammalian cells exhibit some differ-ences, if the process is dissected spatially and temporally.The biosynthetic machinery for PtdSer is also differentbetween yeast and mammalian cells. Yeast uses a PtdSersynthase (NP_010943), which requires CDP-diacylglycerolas the source of the phosphatidic acid moiety thatcondenses with serine to make PtdSer [30]. This PtdSeris preferentially synthesized in the MAM compartmentof the ER and transferred to the mitochondria in anATP-independent reaction [14,15,31]. Morphological evi-dence indicates that PtdSer transfer occurs at the zones ofapposition between the MAM and the outer mitochondrialmembrane. The machinery that affects the transfer ofPtdSer between the organelles remains unknown butgenetic studies (described later) are providing some cluesabout the regulation of the process. Once the PtdSer isincorporated into the outer mitochondrial membrane,there are no demonstrable energetic requirements for itto gain access to Psd1p present in the inner membrane[11,15]. The mitochondrial outer membrane can also beloaded with a PtdSer analog (1-acyl, 2-(NBD-aminoca-proyl)-PtdSer, and this lipid is also imported to the locus ofPsd1p in an ATP-independent manner [20,32].
In mammalian systems, studies using metabolicinhibitors with intact cells reveal that there is an ATPrequirement for PtdSer transport to the locus of Psd1p[28]. Evidence for this ATP requirement is also producedin transport studies using permeabilized cells [13,33].However, experiments with liposomal or isolated micro-somal donors and purified mitochondria do not demon-strate an ATP requirement for PtdSer transport betweenthe membranes [12]. These results strongly suggest thatthe terminal event of lipid transfer from a donor mem-brane to the outer mitochondrial membrane does notrequire ATP in either mammalian cells or yeast (asdescribed earlier). However, unlike yeast, mammaliancells require ATP at multiple steps preceding the finallipid transfer between the membranes. Mammalian cellssynthesize PtdSer by an exchange reaction in which theamino alcohol substituents (Etn and Cho) present inPtdEtn and PtdCho are exchanged for Ser [34].
Thus, in contrast to yeast, the phosphatidic acid moietyof mammalian PtdSer is derived from PtdEtn and PtdCho.The mammalian reaction requires Ca2C and in intact andpermeabilized cells is coupled to the ability of the ER andMAM to sequester the ion via a Ca2C-ATPase [13,32].After PtdSer is formed, there is another ATP requirement
www.sciencedirect.com
that precedes the final lipid transfer to mitochondria. Thisstep is known to be separate from the ATP requirementnecessary for synthesis of PtdSer because it is possible topulse-radiolabel the PtdSer pool and subsequently arrestfurther synthesis of this lipid. Even under conditions ofarrested PtdSer synthesis, the previously synthesizedradioactive lipid is still transported to the mitochondria.The ATP requirement that follows the synthesis of PtdSeris not well understood because isolated mitochondriareadily import PtdSer when incubated with liposomescontaining this lipid in the absence of ATP [12]. Thecurrent assessment of these data is that the newly synthe-sized PtdSer requires ATP to enter a portion of the MAMthat enables it to interact directly with the mitochondria[35]. Although it is tempting to speculate that thenascent PtdSer is lumenally sequestered and requiresthe ATP-dependent action of a transbilayer translocase tomove it to the cytosolic surface of the bilayer, to date, thereis no experimental evidence to support this idea.
The reconstitution of PtdSer synthesis and transport inpermeabilized mammalian cells has also been used toscreen for macromolecular factors that augment thetransport reaction. A successful application of thisapproach led to the identification in brain cytosol of anEF-hand protein capable of increasing the rate oftransport of PtdSer from the ER and MAM to the mito-chondria [36]. The protein, named S100B (NP_006236),increases the rate of lipid transport approximatelythreefold. It is not known whether this protein actuallyparticipates in the transport reaction or promotes thestability and/or assembly of interactions between the ERor MAM and mitochondria.
Studies with transport mutants
Examination of the biochemical characteristics of PtdSertransport using the mutants identified in the screensdescribed previously has provided important insights intothe types of molecules involved in the process and theirmechanisms-of-action (Figure 3). The yeast mutant pstA1is an Etn auxotroph that shows a defect in the rate ofconversion of nascent PtdSer to PtdEtn in intact cells [20].The mitochondria purified from the mutant strain have amarkedly reduced level of PtdEtn and examination ofthese mitochondria on sucrose density gradients revealsthat they are much denser than the organelles from wild-type cells. This is expected if the phospholipid:proteinratio is reduced in mutant mitochondria as a consequenceof reduced phospholipid transport to the organelle. PtdSertransport between the outer and inner mitochondrialmembranes is normal in the pstA1 mutant whenmeasured using the analog 1-acyl, 2-(NBD-aminoca-proyl)-PtdSer. Reconstitution experiments using MAMand mitochondria reveal a defect in the transport ofPtdSer between the organelles. When MAM and mito-chondria are purified from wild-type and mutant cells andrecombined in different combinations as donors andacceptors, the lesion in pstA1 maps to both of themembranes. This finding means that the MAM fromthe pstA1 strain are defective as donor membranes, andthe mitochondria from the pstA1 strain are defective asacceptor membranes. The gene that rectifies all the

Ti BS
Ser
Ser
PtdSer
PtdSer
ATP
Met30p
PtdSer PtdEtnPtdSer
X Y
R41p
PtdIns
PtdSer
PtdIns4P
PtdOH
PtdEtn
Stt4p
Psd2p
PstB2p
S100B
Psd1p
Mitochondria
Golgi
PtdSerdomain
C2domain
ER
MAM
Adapterproteins?
Figure 3. Molecular components and properties of PtdSer transport to mitochondria
and the Golgi. PtdSer synthesized in the mitochondria-associated membrane
(MAM) domain of the endoplasmic reticulum (ER) is transported to the outer
mitochondrial membrane under the regulation of yeast Met30p, a ubiquitin ligase
subunit. Met30p affects the properties of both the MAM and the mitochondria and
this is proposed to occur via protein components (designated X and Y) on the
respective organelles. It is not yet known if Met30p ubiquitinates X and Y directly on
the MAM and mitochondria, or if its action regulates their degradation or
transcription. The rate of transfer of PtdSer to mammalian mitochondria can be
augmented several-fold by the action of mammalian S100B, an EF-hand-Ca2C
binding protein. Genetic screening identifies a CHO-K1 mutant (R41) that is
proposed to have a lesion in a putative protein, R41p, required for efficient transport
of PtdSer between the outer (om) and inner (im) mitochondrial membranes. In
mammalian cells, but not yeast, there is an ATP requirement for newly synthesized
PtdSer to become competent for transport to the mitochondria. In yeast, PtdSer
synthesized in the ER is transported to the Psd2p in the Golgi for the synthesis of
PtdEtn. The Psd2p has not been identified in mammalian cells. Reconstitution
studies with chemically defined donors demonstrate that PtdSer-rich domains are
the preferred source of the lipid destined for transport to Psd2p, and PtdIns4P and
PtdOH can augment the transport reaction. Genetic studies implicate Stt4p as a
source of Ptd-Ins-4-P required for PtdSer transport but there is uncertainty how this
occurs because the enzyme is primarily found in the plasma membrane. The
presence of Psd2p and PstB2p on the Golgi membrane is required for
intermembrane PtdSer transport to occur. The C2 domain of Psd2p is not required
for catalysis by the enzyme but is necessary for the transport reaction. The C2
domain might function by recognizing anionic lipids in the donor membrane and
promoting docking with the acceptor membranes. It is also likely that adaptor
proteins have a role in promoting interactions among proteins and lipids between
the membranes.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005400
phospholipid synthesis and transport abnormalities of thepstA1 mutation was cloned by complementation andidentified as MET30 (NP_012218). MET30 encodes aprotein subunit (Met30p) of a multi-component E3ubiquitin ligase [37]. The Met30p is an F box proteinthat dictates substrate specificity of the ubiquitin ligase[38]. This result is quite striking and provides unantici-pated linkage between protein ubiquitination and phos-pholipid transport. A growing body of data now implicatesprotein ubiquitination in many membrane traffickingevents, including viral budding at the cell surface,endosomal protein sorting and multivesicular bodyformation [39]. Ubiquitination is also known to regulategene expression by controlling the half life and/or activity
www.sciencedirect.com
of transcription factors. One known substrate for E3ubiquitin ligase containing Met30p is the transcriptionfactor Met4p [40–42] (NP_014296). Upon ubiquitination,Met4p is inactivated. Biochemical studies reveal that thepstA1 mutant is defective in ubiquitination of Met4p [20].Currently, it is not known how many proteins can serve assubstrates for recognition by Met30p.
At this stage, it is unclear howMet30p regulates PtdSertransport between the MAM and the mitochondria. Threepossible explanations seem most likely. The first hypoth-esis – and perhaps the simplest – is that protein substratespresent on both the MAM and mitochondria are recog-nized by Met30p. Ubiquitination of these putative targetproteins (designated as X and Y in Figure 3) could have arole in either direct activation of transport molecules, orMAM-mitochondria recognition and docking for thepurpose of lipid transport. A second hypothesis is thatan inhibitor of PtdSer transport resides on both the MAMand mitochondria. In this case, Met30p directed ubiqui-tination of a protein substrate could lead to its degradationand alleviation of an inhibitory block upon transport. Athird hypothesis is that the action of Met30p upon Met4pregulates transcription of a factor involved in lipidtransport. Because ubiquitination directed by Met30pinactivates Met4p, it is likely that the factor would be anegative regulator of PtdSer transport.
Biochemical studies in mammalian cells, with the R41mutant cell-line that is resistant to the cytolytic effects ofthe toxin Ro09–0198, also reveal important informationabout the process of PtdSer transport within the mito-chondria [17]. The R41 cells show a reduction in totalcellular PtdEtn content and the metabolism of nascentPtdSer to PtdEtn. Enzymatic studies reveal that Psd1pactivity is normal but the transport of 1-acyl, 2-(NBD-aminocaproyl)-PtdSer between the outer and innermitochondrial membranes is markedly reduced. Interest-ingly, this defect in lipid import into the mitochondria doesnot affect protein import into the organelle. These resultsprovide significant evidence for the existence of specificproteins involved in the transport of phospholipidsbetween the outer and inner mitochondrial membranesthat are independent of the protein transport machinery.Thus far, the cDNA and gene required to complement theR41 strain have not been cloned.
Examination of the mutants obtained in the PSTBpathway is also providing some new definition to themolecular requirements for the transport of PtdSer toPsd2p in yeast cells. The first mutant described in thispathway was pstB1 [18]. The pstB1mutant shows a defectin the conversion of nascent PtdSer to PtdEtn by Psd2p.The reduced PtdEtn synthesis is not due to an alterationin Psd2p activity but in the access of the substrate to theenzyme. The gene that complements the pstB1 growthdefect (Etn auxotrophy) and abnormality in lipid syn-thesis is STT4 (NP_013408), which encodes a phospha-tidylinositol 4-kinase (PtdIns4-kinase; Stt4p) [43]. TheStt4p is one of three PtdIns4-kinases that have beendescribed in yeast and shows both plasma membraneand microsomal distribution in subcellular fractions.The enzyme is tethered to the plasma membrane bya structural protein named Sfk1p [44] (NP_012873).

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 401
There is reasonable evidence that the major pool ofphosphatidylinositol 4-phosphate (PtdIns4P) producedby Stt4p resides in the plasma membrane [45]. However,it remains unclear whether the PtdIns4P generated bythis enzyme can also be found in other intracellularmembranes. Although the genetic experiments with pstB1strains implicate PtdIns4P at some stage of PtdSertransport to Psd2p, much remains uncertain. It isunknown whether PtdIns4P or its downstream product,phosphatidylinositol (4,5)-bisphosphate PtdIns(4,5)P2, isimportant in the transport process. Analysis of thesequence of Psd2p reveals the presence of a C2 domain.Typically, C2 domains are involved in Ca2C binding,protein–lipid and protein–protein interactions [46].Anionic lipids, including PtdSer and polyphosphoinosi-tides, are recognized by C2 domains and this raises thepossibility that Psd2p might participate directly inrecognition of these lipids on the donor membrane aspart of the transport reaction.
To test the role of the C2 domain of Psd2p in PtdSertransport, deletion mutants lacking this domain(psd2-C2D) were constructed [21]. In strains lackingwild-type copies of Psd1p and Psd2p, the mutant protein,Psd2-C2Dp, is catalytically active and can be expressed atlevels 10-fold higher than that normally required toproduce the PtdEtn required for cell growth. The catalyticactivity of Psd2-C2Dp can be measured using a 1-acyl,2-(NBD-aminocaproyl)-PtdSer substrate that spon-taneously inserts into membranes harboring the enzyme.Further analysis reveals that the Psd2-C2Dp undergoesthe same post-translational processing as its wild-typecounterpart and is localized correctly within the cell.Despite the high catalytic activity of the Psd2-C2Dpexpressing strains, the cells fail to grow unless they aresupplemented with Etn. Thus, the C2 domain of Psd2p hasan important non-catalytic role in the action of theprotein. Measurement of PtdSer transport in intact andpermeabilized cells reveals that this lipid cannot bedelivered to the enzyme lacking the C2 domain. Collec-tively, these results define a direct role for the C2 domainof Psd2p in the transport process. The mechanism ofhow the C2 domain is functioning remains to beelucidated. It could form a docking module that recognizesanionic phospholipids (e.g. PtdSer, PtdIns, PtdIns4P,PtdIns(4,5)P2) and perhaps proteins on the donor mem-brane. If the C2 domain only acts in docking, then otherproteins would need to contribute to the assembly of atransport apparatus for moving PtdSer to the acceptormembrane. Alternatively, the C2 domain might bind toand physically participate in the transport of the lipidbetween the membranes. With either mechanism it seemsprobable that Psd2p can function in concert with otherproteins present on the acceptor membrane to form aPtdSer transport complex.
The genetic screening of the PSTB pathway identifiedanother mutant strain (pstB2), and its corresponding gene(PSTB2) and encoded protein (PstB2p), as a component inthe process of PtdSer transport to the locus of Psd2p [19].The mutant strain displays a profound defect in thetransport dependent metabolism of PtdSer to PtdEtn inintact and permeabilized cells and reconstitution
www.sciencedirect.com
reactions with isolated organelles. The PstB2p is amember of the phospholipid exchange and transferprotein family and is structurally similar to Sec14p, aPtdIns and PtdCho transfer protein that participates inprotein trafficking events [47]. The gene complementingthe pstB2 mutant strain has been named PDR17(NP_014135) and SFH4 in other screens [48,49] but forconsistency it is referred to as PSTB2 in this review. ThePstB2p transfers PtdIns between liposomes and mito-chondria in cell-free assays but does not transfer PtdSer.The protein is amphitropic and can be either soluble ormembrane bound in yeast and in insect cell expressionsystems. The membrane bound form of the protein ishighly resistant to removal by alteration of ionic con-ditions. Reconstitution assays with permeabilized cellsand organelle preparations demonstrate that the proteinmust be present on the acceptor membrane for transfer ofnascent PtdSer to Psd2p to occur [16]. The mechanism-of-action of PstB2p is still unknown and many outstandingquestions need to be resolved. The protein is capable ofbinding phosphatidylinositol (PtdIns) but it is not clearhow this relates to PtdSer transport. This lipid bindingcould account for the amphitropic properties of PsdB2pand it could facilitate interactions between donor andacceptor membranes. The requirement for PstB2p on theacceptor membranes also suggests that it interacts withspecific proteins (perhaps Psd2p) at that location but thishas not yet been demonstrated.
Additional studies with the PSTB pathway haveprovided further insights into the role of lipids in theprocess of PtdSer transport. In an effort to define theseparate molecular requirements of the donor andacceptor membranes in lipid transport, a chemicallydefined donor membrane system was developed [50]. Thedefined donors exhibit some interesting and unexpectedcharacteristics. The first unusual property is thatmembranes with a relatively planar geometry madefrom large unilamellar vesicles (with diameters ofw400 nm) are superior donors compared with highlycurved membranes made from small unilamellar vesicles.A second unusual property is that PtdSer-rich donors arethe preferred substrate for transport. Pure PtdSer vesicles(400 nm diameter) transfer the lipid to acceptors at20-times the rate of vesicles containing 50% PtdSer and50% PtdCho. The decline in PtdSer transfer with changein liposome composition is exponential and, at 50%PtdSer,the transport reaction is negligible. Interestingly, phos-phatidic acid (PtdOH) and PtdIns4P can reverse theinhibition caused by surface dilution with lipids such asPtdCho. The PtdOH completely reverses the inhibition,whereas the PtdIns4P partially reverses the inhibition. Inreconstitution studies with chemically defined donors, thefidelity of the process to the properties of PtdSer transportobserved in vivo and in permeabilized cells, was cruciallytested using both pstB2D and psd2-C2D mutants. Theseexperiments function as important tests to rule outspurious fusion of chemically defined donors with theacceptor membranes. The results reveal that the recon-stitution of transport with chemically defined donorsfaithfully recapitulates the properties of PtdSer transportfound in both intact and permeabilized cells. These

Box 2. Emerging lipid and protein motifs implicated in phospholipid traffic
UbiquitinUbiquitin is a 76-amino acid protein that is attached to target proteins
via an isopeptide linkage at specific lysine residues. In many cases,
ubiquitin serves as a modification that destines the modified target
protein for degradation via proteasomes. Recently, ubiquitin has been
recognized in many systems to also serve as an important signaling
motif for either activating or inactivating proteins without degradation.
This activation and inactivation can serve to regulate catalysis,
transcription, subcellular trafficking and protein–protein interactions.
Ubiquitin ligasesUbiquitin ligases attach ubiquitin to target proteins. The ubiquitin
ligase subunit of interest in phospholipid traffic, Met30p, is part of a
large multiprotein complex. The role of Met30p is to bind to the
holoenzyme and dictate the choice of target protein to be modified by
ubiquitin.
S100BS100B is an EF-hand domain-Ca2C binding protein that seems to
function as a regulator of lipid transport machinery on mammalian
mitochondria-associated membrane (MAM) and/or mitochondria.
Phosphatidylinositol-4-kinasePhosphatidylinositol-4-kinase phosphorylates phosphatidylinositol to
yield the product PtdIns4P. This modification can serve to mark
membrane domains for interactions with specific binding proteins.
Genetic and biochemical studies implicate the PtdIns4 kinase, Stt4p, in
phospholipid transport. The PtdIns4P also functions as an important
precursor to PtdIns(4,5)P2, which also acts as a membrane recognition
domain for interaction with binding proteins. Cleavage of
PtdIns(4,5)P2 produces two important signaling molecules, diacylgly-
cerol and inositol (1,4,5)-trisphosphate (Ins(1,4,5)P3).
Phospholipid exchange and transfer proteins
Phospholipid exchange and transfer proteins can function to
exchange lipids between membranes in vitro but their function
in vivo remains uncertain. The exchange protein described in this
report, PstB2p, recognizes and transfers PtdIns in vitro. However,
in vivo it is an essential component in PtdSer transport to Psd2p. The
property of PtdIns recognition might promote the attachment to
membranes in vivo, where it functions as part of a larger molecular
complex that transports PtdSer between membranes.
C2 domains
C2 domains are Ca2C- and phospholipids-binding domains involved in
protein–lipid interactions. A C2 domain that is present on Psd2p is not
required for catalysis but is necessary for the transfer of PtdSer
between donor and acceptor membranes.
PtdSer domainsReconstitution studies with liposomes as donor membranes demon-
strate that PtdSer-rich domains are the preferred source of the lipid for
transfer to Psd2p. Dilution of pure PtdSer domains with small amounts
of PtdCho dramatically inhibits intermembrane transport of the lipid.
Acidic phospholipidsReconstitution studies show that dilution of PtdSer domains with
phosphatidic acid does not disrupt transport like PtdCho but has a
stimulatory effect on the process. PtdIns4P, the product of Stt4p,
shows similar albeit weaker activity than phosphatidic acid.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005402
findings raise the possibility that segregated domains ofPtdSer, or PtdSer and PtdOH, or PtdSer and PtdIns4Pwithin the donor membrane, could serve as specific localregions for intermembrane lipid transport. Such domainscould arise as a consequence of specific proteins present inthe donor membrane that corral PtdSer and other anioniclipids. Alternatively, the anionic lipid-rich domains mightalso be induced in the donor membrane by interactionswith proteins in the acceptor membrane. Soluble adaptorproteins might also function to recognize specific lipid andprotein elements in donor and acceptor membranes. Thesemodels are all highly speculative at this stage but theyprovide a provocative hypothetical framework for futureexperiments.
Summary
Phospholipid transport associated with new organelleformation is a fundamental process of biochemistry andcell biology whose mechanisms remain to be elucidated.Genetic and biochemical tools applied to the problem areimplicating specific proteins and lipids in the transportand its regulation, as summarized in Figure 3 and Box 2.The proteins or protein motifs implicated thus far includeubiquitin, ubiquitin ligase (Met30p), PtdIns4-kinase(Stt4p), a phospholipid binding protein (PstB2p), a solubleEF-hand protein (S100B) and a C2 domain (present onPsd2p). The lipids implicated in the process includePtdSer, PtdIns, PtdIns4P and PtdOH. PtdSer serves notonly as a substrate for transport and decarboxylation butalso as a potential ligand for the C2 domain of Psd2p.PtdIns, PtdIns4P and PtdOH are also potential
www.sciencedirect.com
recognition ligands for docking reaction between thedonor and acceptor membranes. In addition, the anioniclipids could also function as nucleation sites for inter-action with membrane proteins either in their resident orapposed membranes that can serve to assemble dockingand transport machinery at regions of membrane contact.These molecules are likely to constitute an early list ofcomponents involved in non-vesicular polar lipid trans-port between organelles. From this current list ofmolecular participants, the focus now needs to movetowards defining their precise roles in the transportreactions by addressing the questions: (i) what interactswith what? (ii) Do protein–protein and protein–lipidinteractions between membranes introduce physicalchanges in local membrane environments (e.g. segre-gation of PtdSer) that are essential for transport? (iii) Howare intermembrane docking and phospholipid transportinterrupted to stop the process and disengage the donorsfrom the acceptors? (iv) How applicable are the molecularmotifs used for aminophospholipid transport to othertypes of polar lipids? (v) Do mammalian cells use the samemolecular motifs for aminophospholipid transport asthose uncovered in yeast? These current, challengingquestions require new genetic, biochemical and physicalapproaches to the problem. The resolution of these issuesis likely to provide new insights into the mechanisms ofphospholipid transport during membrane biogenesis.
Acknowledgements
This work was supported by a grant from the National Institutes ofHealth 2R37 GM 32453.

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 403
References
1 Rothman, J.E. (2002) Lasker Basic Medical Research Award. Themachinery and principles of vesicle transport in the cell. Nat. Med. 8,1059–1062
2 Schekman, R. (2002) Lasker Basic Medical Research Award. SECmutants and the secretory apparatus. Nat. Med. 8, 1055–1058
3 Schekman, R. (2004) Merging cultures in the study of membranetraffic. Nat. Cell Biol. 6, 483–486
4 Orci, L. et al. (1991) Brefeldin A, a drug that blocks secretion, preventsthe assembly of non-clathrin-coated buds on Golgi cisternae. Cell 64,1183–1195
5 Bednarek, S.Y. et al. (1995) COPI- and COPII-coated vesiclesbud directly from the endoplasmic reticulum in yeast. Cell 83,1183–1196
6 Sleight, R.G. and Pagano, R.E. (1983) Rapid appearance of newlysynthesized phosphatidylethanolamine at the plasma membrane.J. Biol. Chem. 258, 9050–9058
7 Vance, J.E. et al. (1991) Brefeldin A does not inhibit the movement ofphosphatidylethanolamine from its site of synthesis to the cell surface.J. Biol. Chem. 266, 8241–8247
8 Kaplan, M.R. and Simoni, R.D. (1985) Intracellular transport ofphosphatidylcholine to the plasmamembrane.J.Cell Biol. 101, 441–445
9 Gnamusch, E. et al. (1992) Transport of phospholipids betweensubcellular membranes of wild-type yeast cells and of the phospha-tidylinositol transfer protein-deficient strain Saccharomyces cerevi-siae sec 14. Biochim. Biophys. Acta 1111, 120–126
10 De Kroon, A.I. et al. (2003) Continuous equilibration of phosphatidyl-choline and its precursors between endoplasmic reticulum andmitochondria in yeast. Mol. Biol. Cell 14, 2142–2150
11 Simbeni, R. et al. (1993) Import of phosphatidylserine into isolatedyeast mitochondria. Biochim. Biophys. Acta 1145, 1–7
12 Voelker, D.R. (1989) Reconstitution of phosphatidylserine import intorat liver mitochondria. J. Biol. Chem. 264, 8019–8025
13 Voelker, D.R. (1990) Characterization of phosphatidylserine synthesisand translocation in permeabilized animal cells. J. Biol. Chem. 265,14340–14346
14 Achleitner, G. et al. (1995) Synthesis and intracellular transport ofaminoglycerophospholipids in permeabilized cells of the yeast,Saccharomyces cerevisiae. J. Biol. Chem. 270, 29836–29842
15 Achleitner, G. et al. (1999) Association between the endoplasmicreticulum and mitochondria of yeast facilitates interorganelletransport of phospholipids through membrane contact. Eur.J. Biochem. 264, 545–553
16 Wu, W.I. and Voelker, D.R. (2001) Characterization of phosphatidyl-serine transport to the locus of phosphatidylserine decarboxylase 2 inpermeabilized yeast. J. Biol. Chem. 276, 7114–7121
17 Emoto, K. et al. (1999) Isolation of a Chinese hamster ovary cellmutant defective in intramitochondrial transport of phosphatidyl-serine. Proc. Natl. Acad. Sci. U. S. A. 96, 12400–12405
18 Trotter, P.J. et al. (1998) A genetic screen for aminophospholipidtransport mutants identifies the phosphatidylinositol 4-kinase,STT4p, as an essential component in phosphatidylserine metabolism.J. Biol. Chem. 273, 13189–13196
19 Wu, W.I. et al. (2000) A new gene involved in the transport-dependent metabolism of phosphatidylserine, PSTB2/PDR17,shares sequence similarity with the gene encoding thephosphatidylinositol/phosphatidylcholine transfer protein, SEC14.J. Biol. Chem. 275, 14446–14456
20 Schumacher, M.M. et al. (2002) Phosphatidylserine transport to themitochondria is regulated by ubiquitination. J. Biol. Chem. 277,51033–51042
21 Kitamura, H. et al. (2002) The C2 domain of phosphatidylserinedecarboxylase 2 is not required for catalysis but is essential for in vivofunction. J. Biol. Chem. 277, 33720–33726
22 Trotter, P.J. et al. (1993) Phosphatidylserine decarboxylase fromSaccharomyces cerevisiae: isolation of mutants, cloning of thegene and creation of the null phenotype. J. Biol. Chem. 268,21416–21424
23 Trotter, P.J. et al. (1995) Phosphatidylserine decarboxylase 2 ofSaccharomyces cerevisiae: cloning and mapping of the gene, hetero-logous expression and creation of the null allele. J. Biol. Chem. 270,6071–6080
24 Trotter, P.J. and Voelker, D.R. (1995) Identification of a non-
www.sciencedirect.com
mitochondrial phosphatidylserine decarboxylase activity (PSD2)in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 270,6062–6070
25 Summers, E.F. et al. (1988) Saccharomyces cerevisiae cho2 mutantsare deficient in phospholipid methylation and cross-pathway regu-lation of inositol synthesis. Genetics 120, 909–922
26 Kodaki, T. and Yamashita, S. (1987) Yeast phosphatidylethanolaminemethylation pathway. Cloning and characterization of two distinctmethyltransferase genes. J. Biol. Chem. 262, 15428–15435
27 Vance, J.E. (1990) Phospholipid synthesis in a membrane fractionassociated with mitochondria. J. Biol. Chem. 265, 7248–7256
28 Voelker, D.R. (1985) Disruption of phosphatidylserine translocation tothe mitochondria in baby hamster kidney cells. J. Biol. Chem. 260,14671–14676
29 Wu, W.I. and Voelker, D.R. (2002) Biochemistry and genetics ofinterorganelle aminoglycerophospholipid transport. Semin. Cell Dev.Biol. 13, 185–195
30 Carman, G.M. andHenry, S.A. (1999) Phospholipid biosynthesis in theyeast Saccharomyces cerevisiae and interrelationship with othermetabolic processes. Prog. Lipid Res. 38, 361–399
31 Gaigg, B. et al. (1995) Characterization of a microsomal subfractionassociated with mitochondria of the yeast, Saccharomyces cerevisiae.Involvement in synthesis and import of phospholipids into mitochon-dria. Biochim. Biophys. Acta 1234, 214–220
32 Voelker, D.R. (1991) Adriamycin disrupts phosphatidylserine importinto the mitochondria of permeabilized CHO-K1 cells. J. Biol. Chem.266, 12185–12188
33 Voelker, D.R. (1989) Phosphatidylserine translocation to the mito-chondrion is an ATP dependent process in permeabilized animal cells.Proc. Natl. Acad. Sci. U. S. A. 86, 9921–9925
34 Kuge, O. and Nishijima, M. (2003) Biosynthetic regulation andintracellular transport of phosphatidylserine in mammalian cells.J. Biochem. (Tokyo) 133, 397–403
35 Shiao, Y.J. et al. (1995) Evidence that phosphatidylserine isimported into mitochondria via a mitochondria-associated mem-brane and that the majority of phosphatidylethanolamine isderived from decarboxylation of phosphatidylserine. J. Biol.Chem. 270, 11190–11198
36 Kuge, O. et al. (2001) Enhancement of transport-dependentdecarboxylation of phosphatidylserine by S100B protein inpermeabilized Chinese hamster ovary cells. J. Biol. Chem. 276,23700–23706
37 Thomas, D. et al. (1995) Met30p, a yeast transcriptional inhibitor thatresponds to S-adenosylmethionine, is an essential protein with WD40repeats. Mol. Cell. Biol. 15, 6526–6534
38 Deshaies, R.J. (1999) SCF and Cullin/RingH2-based ubiquitin ligases.Annu. Rev. Cell Dev. Biol. 15, 435–467
39 Schnell, J.D. and Hicke, L. (2003) Non-traditional functions ofubiquitin and ubiquitin-binding proteins. J. Biol. Chem. 278,35857–35860
40 Kaiser, P. et al. (2000) Regulation of transcription by ubiquitinationwithout proteolysis: Cdc34/SCF(Met30)-mediated inactivation of thetranscription factor Met4. Cell 102, 303–314
41 Kuras, L. et al. (2002) Dual regulation of the met4 transcription factorby ubiquitin-dependent degradation and inhibition of promoterrecruitment. Mol. Cell 10, 69–80
42 Flick, K. et al. (2004) Proteolysis-independent regulation of thetranscription factor Met4 by a single Lys48-linked ubiquitin chain.Nat. Cell Biol. 6, 634–641
43 Yoshida, S. et al. (1994) A novel gene, STT4, encodes a phosphatidyl-inositol 4-kinase in the PKC1 protein kinase pathway of Saccharo-myces cerevisiae. J. Biol. Chem. 269, 1166–1171
44 Audhya, A. and Emr, S.D. (2002) Stt4 PI 4-kinase localizes to theplasma membrane and functions in the Pkc1-mediated MAP kinasecascade. Dev. Cell 2, 593–605
45 Roy, A. and Levine, T.P. (2004) Multiple pools of phosphatidylinositol4-phosphate detected using the pleckstrin homology domain of Osh2p.J. Biol. Chem. 279, 44683–44689
46 Cho, W. and Stahelin, R. (2005) Membrane protein interactions in cellsignaling and membrane trafficking. Annu. Rev. Biophys. Biomol.Struct. 34, 119–151
47 Bankaitis, V.A. et al. (1990) An essential role for a phospholipidtransfer protein in yeast Golgi function. Nature 347, 561–562

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005404
48 van den Hazel, H.B. et al. (1999) PDR16 and PDR17, two homologousgenes of Saccharomyces cerevisiae, affect lipid biosynthesis andresistance to multiple drugs. J. Biol. Chem. 274, 1934–1941
49 Phillips, S.E. et al. (1999) Yeast Sec14p deficient in phosphatidyl-inositol transfer activity is functional in vivo. Mol. Cell 4, 187–197
Have you contributed to a
Did you know that you are entitle
A 30% discount is available to ALL Elsevier book and journal contr
from us.
To take advantage of your discount:
1. Choose your book(s) from www.elsevier.com or www.books.elsev
2. Place your order
Americas:
TEL: +1 800 782 4927 for US customers
TEL: +1 800 460 3110 for Canada, South & Central America cu
FAX: +1 314 453 4898
E-MAIL: [email protected]
All other countries:
TEL: +44 1865 474 010
FAX: +44 1865 474 011
E-MAIL: [email protected]
You’ll need to provide the name of the Elsevier book or journa
orders within the US, Canada, and the UK.
If you are faxing your order, please enclose a copy of this pag
3. Make your payment
This discount is only available on prepaid orders. Please note
Elsevier Health Sciences products.
For more information, visit w
www.sciencedirect.com
50 Wu, W.I. and Voelker, D.R. (2004) Reconstitution of phosphatidylser-ine transport from chemically defined donor membranes to phospha-tidylserine decarboxylase 2 implicates specific lipid domains in theprocess. J. Biol. Chem. 279, 6635–6642
n Elsevier publication?
d to a 30% discount on books?
ibutors when ordering books or stand-alone CD-ROMs directly
ier.com
stomers
l to which you have contributed. Shipping is FREE on pre-paid
e.
that this offer does not apply to multi-volume reference works or
ww.books.elsevier.com

Eukaryotic transcription factors asdirect nutrient sensorsChristopher A. Sellick and Richard J. Reece
The University of Manchester, Faculty of Life Sciences, The Michael Smith Building, Oxford Road, Manchester M13 9PT, UK
The recognition of changes in environmental conditions,
and the ability to adapt to these changes, is essential for
the viability of cells. There are numerous well-charac-
terized systems by which the presence or absence of an
individual metabolite can be recognized by a cell. The
recognition of a metabolite is, however, just one step of
a process that often results in changes in the expression
of sets of genes required to respond to that metabolite.
The signalling pathway between metabolite recognition
and transcriptional control is often complex. However,
recent evidence from yeast suggests that complex sig-
nalling pathways might be circumvented via the direct
interaction between individual metabolites and regula-
tors of RNA polymerase II transcription.
Introduction
The genome of an organism is complex. Within its DNAsequence, an organism contains all the information notonly required to define cell type, but also to respond to avariety of external conditions and signals. For example,the genome of the simple eukaryotic yeast Saccharomycescerevisiae contains 12 million base pairs of DNA split into16 separate chromosomes and comprises w6000 differentgenes. The protein products of these genes rarely actindividually, and whole pathways are often regulated inresponse to a particular signal. Indeed, the complexprocesses of the cell depend on differential expression ofsets of genes either in particular cell types (e.g. when celldifferentiation takes place within an embryo), at a parti-cular time (e.g. when a microbe produces an antibiotic latein its growth cycle) or under certain environmentalconditions (e.g. changes in the metabolic flux within acell depending on metabolite availability). Thus, withincells, there are programmes whereby sets of genes can becoordinately switched on or off. Efficient switches must beable to operate in two directions: on/off and off/on.Genetic switches must also operate in this manner, forexample, enabling appropriate gene expression whencertain nutrients are available and turning geneexpression off when nutrients are limiting or unavailable.It is becoming increasingly apparent that what weretraditionally thought of as metabolic enzymes can havediverse roles in a wide range of cellular processes [1].Similarly, metabolites seem not to serve only as substratesfor enzymes. Indeed, the link between metabolic sub-strates and their ability to directly influence transcription
Corresponding author: Reece, R.J. ([email protected]).Available online 9 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
is long established in prokaryotes. For example, inEscherichia coli, the interaction between lactose and thelac repressor [2], or galactose and GalR [3], to modulatethe DNA-binding properties of each respective protein iswell understood. Here, we discuss two eukaryotic geneticswitches and the ways in which they are regulated by theaction of small-molecule metabolites. Insights into theprocesses of metabolically regulated gene expression ineukaryotes have been gained from the study of the simplegenetic switches of yeast – although the mechanisticdetails have turned out to be somewhat complex.
Direct modulation of transcription factor function –
Put3p
S. cerevisiae can use a variety of compounds as nitrogensources. When preferred sources of nitrogen such asammonia or glutamate are available, the transcription ofgenes involved in the use of less-preferred nitrogensources are repressed [4]. Proline, a comparatively poornitrogen source, is transported into the cell by the generalamino acid permease Gap1p and by the proline-specifictransporter Put4p (Figure 1). Both Gap1p and Put4p arelocated in the plasma membrane, and the activity of eachis regulated in response to the quality of the nitrogenavailable to the cell [5,6]. For its use as a nitrogen source,intracellular proline is converted to glutamate in themitochondria by the products of PUT1 and PUT2, thegenes encoding proline oxidase and D1-pyrroline-5-car-boxylate dehydrogenase, respectively [7]. The presence ofproline, and absence of preferred nitrogen sources, resultsin the high-level transcriptional induction of both PUT1and PUT2 [8]. In response to the overall quality ofnitrogen available to the cell, Nil1p and Ure2p – twoGATA factor proteins (for review, see Ref. [9]) – regulatethe expression of both PUT1 and PUT2 [10,11]. However,neither Nil1p nor Ure2p are essential for the growth ofyeast with proline as the sole source of nitrogen, whereasthe specific regulator of the proline-utilization pathway,Put3p, is absolutely required for growth on proline [10].
Put3p architecture
Put3p is a transcriptional activator that has an overallarchitecture typical of the Zn(II)2Cys6 binuclear clusterfamily of proteins [12]. It possesses an N-terminalDNA-binding and dimerization domain [13] and an acidicactivation domain at its C terminus [14]. The proteinis bound to its target DNA sites upstream of PUT1and PUT2 irrespective of whether those genes are
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
. doi:10.1016/j.tibs.2005.05.007

Put4p
Proline
PUT1/PUT2
Nucleus
Put3p
Cell wall
Mitochondrion
CH2H2C
H2CNH
CH
COOH
CH2H2C
H2CNH
CH
COOHPut1pPut2p
Gap1p
Ti BS
Figure 1. Use of proline by yeast cells. Proline enters the cell via one of two transporters – Gap1p (predominantly) or Put4p. Once inside the cell, proline proceeds to the
mitochondria where it is enzymatically converted to glutamate. In addition, proline probably enters the nucleus to convert Put3p to a transcriptionally active form.
The presence of proline, but only as the sole source of nitrogen, results in induction of Put3p, which activates transcription of the proline-utilization genes, PUT1 and PUT2.
The enzymes encoded by these genes, Put1p and Put2p, are transported into the mitochondria where they convert proline to glutamate.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005406
transcriptionally active [15]. Put3p has been identified asa phosphoprotein, and the extent of phosphorylation hasbeen correlated with transcriptional activity [16]. Phos-phorylation of Put3p, however, does not seem to be depen-dent on the presence of proline. Rather, it is differentiallyphosphorylated in response to the quality of the nitrogensource available to cells. This was further confirmed by theobservation that treatment of yeast cells with rapamycin,which mimics nitrogen-limiting conditions, resulted in thehyperphosphorylation of Put3p [11].
Conformational changes in Put3p
The activation of transcription by Put3p in responseto proline seems to be mediated by a conformationalchange within the protein. The addition of prolineto immuno-precipitated Put3p results in increased resist-ance to digestion with certain proteases [17]. In addi-tion, immuno-precipitated glutathione S-transferase(GST)-tagged Put3p from proline-grown yeast cells hasincreased resistance to thrombin cleavage of the GST tagcompared with the same protein prepared from ammonia-grown cells [17]. These data suggest that Put3p undergoesa conformational change in the presence of proline. It isconceivable that this conformational change results inunmasking of the activation domain, thereby enabling therecruitment of the transcriptional machinery.
Transcriptional activation induced by proline
In vivo foot-printing has demonstrated clearly that Put3pbinds to its DNA-recognition sites in a proline-indepen-dent manner [18]. Therefore, transcriptional activationby Put3p cannot be regulated by the control of itsDNA-binding properties but, instead, changes in thetranscriptional activity of DNA-tethered Put3p mustoccur in response to proline. In vitro transcriptionexperiments have demonstrated that the activity ofPut3p is induced in the presence of proline itself [19].
www.sciencedirect.com
Direct interaction between Put3p and proline
The ability of proline to modulate the activity of Put3pin vitro suggests that the metabolite is directly affectingthe activity of the transcriptional regulator. Prolineaffinity chromatography has demonstrated that therecognition of proline by Put3p is indeed via a directphysical interaction. When Put3p was applied to a columnbearing immobilized proline attached via its carboxylgroup, the protein was able to bind to the matrix and couldbe eluted with proline [19]. Put3p was eluted from thecolumn at a proline concentration of w25 mM, whichsuggests that the interaction between Put3p and proline isweak [19]. However, this is a level of the metabolitecomparable to that required for the activation of PUT1and PUT2 in vivo [16], and to elicit a response usingin vitro transcription assays [19]. When a similar columnwas produced in which the proline was attached via itsimino-group nitrogen, no Put3p binding was detected.This, and other data, suggests that that the pyrrolidinering is vital for recognition by Put3p. Pyrrolidine itselfwas found to activate Put3p in an in vitro transcriptionassay [19], whereas the addition of hydroxyl residues tothe pyrrolidine ring ablated its ability to modulate theactivity of Put3p. Therefore, Put3p seems to recognize andinteract with a simple pyrrolidine ring and the carboxylgroup found in proline (Table 1) might be removedor modified without unduly affecting the interactionwith the protein.
The interaction between Put3p and proline is sufficientto bring about a transcriptional response. However, theposition and structure of the proline-binding site withinPut3p are not known. The activation domain of Put3p isnot proline responsive and, when fused to a heterologousDNA-binding domain, it activates transcription in aconstitutive manner [14]. This suggests that regulationof the transcriptional activity of Put3p occurs elsewherein the protein.

Table 1. Some metabolically regulated transcriptional switches in Saccharomyces cerevisiae
Pathway Activator Ancillary proteins Metabolic inducer
Proline utilization Put3p –
NH
C
OH
O
Proline
Galactose metabolism Gal4p Gal80p, Gal3p
O
OH
OH
OH
HO
CH2OH Galactose
Pyrimidine biosynthesis Ppr1p –
NH
O
HN
O
O
OH
Orotic acid
Leucine biosynthesis Leu3p –OH
O
OHO
OH
CH3H3C
a-isopropylmalate
Lysine biosynthesis Lys14p –
HOH
O
O
NH2 a-aminoadipate semialdehyde
Arginine metabolism Arg81p Mcm1p NH2HN
NH
H2N
O
OH
Arginine
Weak acid response War1p –
O
OH Benzoic acid
Purine biosynthesis Bas1p Bas2p
NH
NH2N
N
O
OHOH
O
CH
P
O
COOH
CH2
COOHOH
HO
O
5 0-phosphoribosyl-5-aminoimid-
azole-4-N-succinocarboxamide
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 407
Model for transcriptional activation by Put3p
At present, the model for transcriptional activation byPut3p is that a dimeric, DNA-bound form of Put3p sensesthe presence of proline within a cell (Figure 2). The pre-sence of proline, but only as the sole source of nitrogen,results in the activation of transcription of PUT1 andPUT2. The conversion of Put3p from a transcriptionallyinert form to a transcriptionally active form is thought tooccur via a conformational change in the protein. Asmentioned, it is possible that this conformational changeresults in unmasking of the activation domain in thepresence of proline.
Phosphorylation might also play a part in regulatingthe overall transcriptional activity of Put3p. It hasbeen proposed that phosphorylation of Put3p is requiredfor transcriptional activation, and that under-phosphory-lated forms of the protein are unable to activate tran-scription [16]. However, analysis of the phosphorylationprofile of recombinant purified Put3p that activatestranscription in vitro has revealed that the protein isunphosphorylated [19]. This suggests that proline can act
www.sciencedirect.com
on unphosphorylated Put3p to convert it into a transcrip-tional activator in vitro, and raises the question as to therole of phosphorylation in regulating transcriptionalactivation by Put3p. It is possible that phosphorylatedforms of Put3p are able to respond to lower levels ofproline, or that the phosphorylation of Put3p is involved inthe global regulation of the proline-utilization pathway bythe GATA factors. The mechanism by which the overallquality of the nitrogen source available to a yeast cell isdetected is still largely unknown. Moreover, the molecularmechanism by which this information is relayed to thePUT genes is unclear, although a complex interplaybetween nitrogen transporters, GATA factors and Put3pis likely to be involved.
Metabolic signalling via protein complex formation –
GAL induction
The expression of the yeast GAL genes is often consideredas a paradigm for eukaryotic transcriptional regulation.The GAL genetic switch provides the transcriptionalcontrol of genes whose products are required for the

L-proline
Put3p dimer
Ti BS
Transcriptionalmachinery
Figure 2. Proposed mechanism for proline-specific transcriptional activation by
Put3p. A DNA-bound Put3p dimer functions as a proline sensor. When the proline
concentration is sufficiently high, proline interacts directly with Put3p. The proline-
binding domain of Put3p is likely to be within the central domain of the protein
(red). The direct interaction results in a conformation change in Put3p that unmasks
the activation domain (yellow) and enables the protein to activate transcription. The
DNA-binding and dimerization domain of Put3p is shown in green.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005408
metabolism of the sugar galactose (the enzymes of theLeloir pathway) [20]. When yeast cells are grown inthe absence of galactose, the GAL genes are transcrip-tionally inert. If galactose is the only available carbonsource, then the GAL genes will become transcriptionallyactive – rapidly and to a high level – and galactose isconverted to glucose 1-phosphate [21]. Galactose is acomparatively poor sugar source for the cell, and yeastwill metabolize other sources of carbon (e.g. glucose) inpreference to galactose, even if a mixture of glucose andgalactose is available [22]. Glucose will trigger cataboliterepression of the GAL genes, and the expression of theactivator upon which they depend, Gal4p, is severelyreduced [23]. In the presence of other carbon sources, suchas raffinose or glycerol, Gal4p is produced in the cell andcan be found tethered upstream of the GAL genes. Like inthe case of Put3p, the modulation of the DNA-bindingactivity of Gal4p does not play a major part in controllingthe activity of the genetic switch in response to induction.The activity of DNA-bound Gal4p is inhibited by itsinteraction with another protein, Gal80p [24]. Theactivation of gene expression by Gal4p is thought toinvolve the ordered recruitment of various protein com-plexes to the promoter of the GAL genes. The transcrip-tionally active form of DNA-tethered Gal4p interactsdirectly with the Tra1p subunit of the SAGA (Spt/Ada/GCN5/acetyltransferase) co-activator–histone-modifying
www.sciencedirect.com
complex [25–27]. Other subunits of SAGA (Spt3p andSpt8p) interact directly with specific subunits of the RNApolymerase II mediator complex (Med2p and Pgd1p,respectively) [28]. Gal4p might also interact with com-ponents of themediator directly [29]. Finally, the mediatorrecruits the RNA polymerase II holoenzyme to the pro-moter, and then initiation of transcription can occur.
Activation of the transcriptional properties of Gal4p
Although the presence of galactose within the cell triggersthe activation of Gal4p, neither Gal4p nor Gal80p functionas the galactose sensor. A transcriptional inducer Gal3pinteracts with the transcriptional repressor Gal80p in agalactose- and ATP-dependent manner [30]. Gal3p seemsto require galactose and ATP so that it can adopt a con-formation to enable it to interact with Gal80p [30]. The netresult of this interaction is that Gal4p becomes active andtranscription of the GAL genes proceeds.
The molecular mechanism by which the activation oftheGAL genes occurs has been the subject of much debate.Two, somewhat conflicting, models for its molecular mode-of-action have been proposed (Figure 3). It has been sug-gested that the induction of the GAL genes occurs via theassociation of a tripartite complex formed between Gal4p,Gal80p and Gal3p [24]. In favour of this model (Figure 3a)are the observations that: (i) Gal4p purified from yeastgrown either in the presence or absence of galactose isassociated with Gal80p [31], (ii) artificially constructedGal80p molecules that contain an activation domain canregulate transcription in the presence and absence ofgalactose [32], (iii) the in vitro assembly of the threeproteins in a gel-shift assay [24], and (iv) the recentobservation using fluorescence resonance energy transfer(FRET) that Gal4p and Gal80p do not dissociate from eachother in the presence or absence of galactose [27]. Otherevidence, however, suggests that this model is incorrectand that Gal80p dissociates fromGal4p and interacts withGal3p in the cytoplasm of yeast cells [33]. This wouldresult in the freeing of Gal4p from the inhibitory effects ofGal80p and enabling transcriptional activation to occur[34]. The dissociation model (Figure 3b) is supported bydata indicating that Gal3p is predominately, and perhapsexclusively, cytoplasmic [33] and that the expression of amyristoylated version of the protein (which is targetedto the plasma membrane of the cell) does not undulyimpair the induction of the GAL genes [34]. In addition,chromatin immunoprecipitation experiments [34] andpull-down assays [35] suggest that the Gal4p–Gal80pcomplex is somewhat weakened (although perhaps notcompletely dissociated) when cells are grown in thepresence of galactose. In either of these two models, theability of Gal3p to interact with Gal80p in a galactose-dependent manner is essential for the transcriptionalinduction of the GAL genes. Consequently, understandingthis interaction is key to understanding the transcrip-tional activation in response to galactose.
Gal3p is similar to galactokinases
Gal3p has proven to be a difficult protein to studybiochemically. When produced in E. coli, the protein islargely insoluble, and it is difficult to produce in large

Gal3p
Gal4p Gal4p
Gal80p Gal80p
Gal4p Gal4p
Gal80p
Gal80p
Gal3p Gal3p
Gal80p Gal80p
Gal3p
Cytoplasm
Nucleus
Gal4p Gal4p
Gal3p
Gal4p Gal4p
Gal80p Gal80p
Gal3p
Cytoplasm
Nucleus
Gal3p
+ Galactose+ ATP
+ Galactose+ ATP
(a)
(b)
Ti BS
Transcriptional
Transcriptional
machinery
machinery
Figure 3. Models for the activation of the yeast GAL genes. (a) A non-dissociation model in which activation occurs via the formation of a tripartite complex of Gal4p, Gal80p
and Gal3p in the presence of galactose and ATP. A conformational change in Gal80p enables Gal4p-mediated recruitment of the transcriptional machinery. (b) A dissociation
model in which the interaction between Gal3p and Gal80p results in a reduction in the nuclear concentration of Gal80p, thereby, enabling Gal4p to interact with the
transcriptional machinery.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 409
amounts in yeast. However, the analysis of Gal3p functionhas been greatly assisted by its similarity to Gal1p – yeastgalactokinase. Gal1p and Gal3p are extremely similar,beingw70% identical andw90% similar at the amino acidlevel [36,37]. Both proteins require galactose and ATP tofunction – Gal3p requires these to interact with Gal80p[38], whereas Gal1p produces galactose-1-phosphate atthe expense of ATP. Despite the high level of similaritybetween the two proteins, Gal3p does not itself possess agalactokinase activity [39]. The removal of Gal1p fromyeast cells results in the inability to grow on galactose,whereas Gal3p-deficient cells can grow on galactose butshow a severe long-term adaptation phenotype. Ratherthan inducing theGAL genes within 30 min after additionof the sugar to cultures, cells lacking Gal3p only fullyinduce the GAL genes approximately three or four dayslater [40]. The effect of loss of Gal3p can be overcome ifGal1p is produced at higher than wild-type levels [41].However, yeast cells bearing a gal3 mutation and a gal1mutation are unable to induce the GAL genes at all [40].Recent modelling data have suggested that theGAL genesbecome induced in the presence of Gal1p but the absenceof Gal3p due to stochastic changes in the cellular con-centration of Gal80p [42,43]. Other galactokinases, forexample, those from E. coli [40], can substitute for the lossof Gal1p in yeast to rescue the galactokinase deficiency,but cannot overcome the transcriptional defect in theGALgenes resulting from the loss of Gal3p. Combined, thesedata suggest that, although Gal1p is primarily a galacto-kinase enzyme, it can also function as a weak transcrip-tional inducer. This conclusion has also been supported by
www.sciencedirect.com
results from several in vitro experiments [24,39]. Otheryeasts, for example, the milk yeast Kluyveromyces lactis,contain a single galactokinase-like molecule that is bothan efficient transcriptional inducer (Gal3p-like function)and a galactokinase enzyme (Gal1p-like function) [44].
Strong evidence indicates that Gal1p and Gal3p haveevolved from each other. Gal3p seems to be a defectivegalactokinase because it contains a two-amino-acid dele-tion within its galactokinase signature motif [39]. There-introduction of these residues (a serine and an alaninestarting at position 164) imparts the resulting proteinwith, albeit weak, galactokinase activity [39]. Thestructures of the galactokinase enzymes from Lactococcuslactis [45], Pyrococcus furiosus [46] and human [47] haverecently been solved. The equivalent residue to Ser164 inboth the P. furiosus and human galactokinase structuresinteracts with the magnesium ion and the b-phosphate ofthe nucleotide [46,47]. Despite the relative lack of overallsequence similarity between the enzymes from diverseorganisms, the galactose- and nucleotide-binding of eachcrystallized protein show a high level of conservation(Figure 4). A model for the bifunctional Gal1p fromK. lactis has been proposed based on its homology withother sugar kinases, the structures of which have beensolved [48]. The overall shape of the molecule within theregion of the protein involved in the interaction with theligands might, indeed, be similar to the galactokinases,but the likely Gal80p-interaction site is not complete inthese models. Therefore, it seems likely that Gal3palso shares similar galactose- and nucleotide-bindingpockets, and is likely to interact with its small molecules

Tyr233
His43
Glu42
PO4
Arg36
Asp183
Gly180
Asp45
Lactococcus lactis
ADP
Tyr200
Arg11
Asp151
Gly148
Asp20
His18
Glu17
Pyrococcus furiosus
ADPNP
Tyr236
Arg37
Asp186
Gly183
Asp46
His44
Glu43
Human
Figure 4. The similarity in ligand-binding domains of different galactokinase enzymes. The galactose-binding site of the galactokinase enzymes from Lactococcus lactis (PDB
code: 1PIE), Pyrococcus furiosus (PDB code: 1S4E) and human (PDB code: 1WUU) are shown. The relative position of either a phosphate ion, ADP or ADPNP that were present
in the crystal structure are also shown. The P. furiosus and human enzymes also contain a magnesium ion (green sphere). The broken lines indicate distances of 2.3–3.3 A
between the protein and the ligands, and might indicate potential hydrogen-bond interactions. Amino acids with the capacity to form hydrogen bonds with galactose are
labelled.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005410
in a similar manner to the galactokinases. Crystals ofS. cerevisiae Gal1p that diffract to beyond 2.6 A haverecently been grown (J. Thoden and H. Holden, personalcommunication), the structure of which will resolve thedifferences between the purely enzymatic proteins andthose capable of transcriptional induction.
The reasons for the evolution of a galactokinase enzyme(Gal1p) into a transcriptional regulator (Gal3p) areunknown. Gal1p and Gal3p are larger than their bacterialor mammalian galactokinase counterparts [39]. Specifi-cally, these yeast transcriptional inducers contain anextra w70 amino acids located between the nucleotide-binding site (homology region III) and the second halfof the galactose-binding site (homology region V) [39].Although it is tempting to speculate that this region of theprotein is responsible for the interaction with Gal80p, andhence transcriptional induction, mutations in GAL3 thatgive rise to a constitutive phenotype (activation of tran-scription in the absence of galactose) are spread through-out its length [49,50]. In addition, Gal80p mutants thatshow impaired Gal3p interaction are spread throughoutthe length of the protein [51,52]. Gal80p has beensuggested to interact with Gal4p as a dimeric protein[53], but its interaction with Gal3p occurs as a monomeronly in the presence of galactose and ATP [54]. Theinfluence of Gal3p and Gal4p on the monomer–dimerinteraction of Gal80p is not fully characterized, and thestructure of Gal80p is not currently known. Modelling theprimary amino sequence of Gal80p onto the previouslysolved structure of the sequence-related, but functionallydistinct, glucose-fructose oxidoreductase from Zymomo-nas mobilis [55] suggests that the mutations in Gal80pthat affect Gal3p interaction might cluster on one surfaceof the protein, whereas mutations that affect Gal4p
www.sciencedirect.com
interaction reside on another surface [52]. Caution shouldbe exercised in interpreting a homology model based ontwo distantly related proteins (15% amino acid identity,30% similarity), but the similarity between Gal80p andan oxidoreductase might suggest that Gal80p is also adinucleotide-binding protein. This could provide yetanother layer of complexity to the control of expressionof the GAL genes.
An additional impediment to the biochemical analysisof Gal3p is that the interaction between galactose andGal3p, or Gal1p, seems to be somewhat weak in nature.The GAL genetic switch is activated maximally in vitrousing galactose at a 1.5 mM concentration [24], and theKm for galactose of Gal1p is 1.2 mM [37]. The weakness ofthis interaction might be a controlling influence on theoverall activation of the GAL genes in vivo. The weakinteraction between Gal3p and galactose might mean thata high concentration of the metabolite must be reachedinside the cell before the GAL genes are switched on fully.Thus, the GAL system offers an intriguing, and complex,interplay between a transcriptional activator, regulatorproteins and small molecules. The recent structural ana-lysis of related proteins is providing a mechanism todissect the molecular details of this elegant switch.
Other metabolically regulated transcriptional switches
The systems described represent only two well-character-ized transcriptional programmes that respond directly tochanges in the intracellular levels of metabolites. Thereare, however, others in which activation is controlled bymetabolites (Table 1). For example: the expression ofgenes required for pyrimidine biosynthesis are controlledby orotic acid (a biosynthetic intermediate of the pyrimi-dine pathway), which controls the activity of the

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 411
transcriptional regulator Ppr1p [56]; the formation of anactive transcriptional complex between Bas1p and Bas2pto control de novo AMP biosynthesis is dependent on5 0-phosphoribosyl-5-aminoimidazole-4-N-succinocarbox-amide (SAICAR) [57]; leucine biosynthesis is regulated bythe action of a-isopropylmalate on Leu3p [58]; lysinebiosynthesis is regulated by the presumed interactionbetween a-aminoadipate semialdehyde and Lys14p [59];arginine biosynthesis is regulated by the interactionbetween arginine and Arg81p which promotes theDNA-binding properties of the Arg81p–Mcm1p complex[60]; in response to weak acid stress, War1p activatescertain stress-response genes via the likely interactionbetween the protein and weak acids [61,62]. In each ofthese cases, the small molecule is presumed to modulatethe transcriptional properties of the activator protein bysome mechanism yet to be determined.
Concluding remarks
The direct modulation of transcription factor function bysmall molecule metabolites is well established in prokary-otes. As we have discussed here, this mechanism oftranscriptional regulation is also common in single-celledeukaryotes. Yeast cells contain many of the signallingpathways found in higher eukaryotes, for example,mitogen-activated protein kinase cascades [63–65], butlack the metabolic homeostasis systems present in higher,multicellular eukaryotes. In higher eukaryotes, the inter-actions between, say, hormones and transcription factorsare understood in detail [66], but it seems that the directmodulation of transcription factor function by metabolitesis largely missing. Single-celled organisms have generallyless control over their metabolic content and this seems tohave been exploited to provide a series of elegant andprecise genetic controls. Understanding the diverse waysin which metabolites can directly influence and regulatetranscription is vital if transcription and metabolism areto be understood in a fully integrated manner.
Acknowledgements
We are grateful to Laila Beynon, Hazel Holden, Amanda Hughes, KarineRebora and David Timson for discussions and their comments on, andsuggestions for, this article. Work in the authors’ laboratory was fundedby The Wellcome Trust and the BBSRC.
References
1 Kim, J.W. and Dang, C.V. (2005) Multifaceted roles of glycolyticenzymes. Trends Biochem. Sci. 30, 142–150
2 Lewis, M. et al. (1996) Crystal structure of the lactose operon repressorand its complexes with DNA and inducer. Science 271, 1247–1254
3 Chatterjee, S. et al. (2002) Ligand specificity and ligand-inducedconformational change in gal repressor. Proteins 49, 554–559
4 Brandriss, M.C. and Magasanik, B. (1979) Genetics and physiology ofproline utilization in Saccharomyces cerevisiae: enzyme induction byproline. J. Bacteriol. 140, 498–503
5 Jauniaux, J.C. and Grenson, M. (1990) GAP1, the general amino acidpermease gene of Saccharomyces cerevisiae. Nucleotide sequence,protein similarity with the other bakers yeast amino acid permeases,and nitrogen catabolite repression. Eur. J. Biochem. 190, 39–44
6 Jauniaux, J.C. et al. (1987) Nitrogen catabolite regulation of prolinepermease in Saccharomyces cerevisiae. Cloning of the PUT4 gene andstudy of PUT4 RNA levels in wild-type and mutant strains. Eur.J. Biochem. 164, 601–606
7 Brandriss, M.C. (1987) Evidence for positive regulation of prolineutilization in Saccharomyces cerevisiae. Genetics 117, 429–435
www.sciencedirect.com
8 Marczak, J.E. and Brandriss, M.C. (1991) Analysis of constitutive andnoninducible mutations of the PUT3 transcriptional activator. Mol.Cell. Biol. 11, 2609–2619
9 Cooper, T.G. (2002) Transmitting the signal of excess nitrogen inSaccharomyces cerevisiae from the Tor proteins to the GATA factors:connecting the dots. FEMS Microbiol. Rev. 26, 223–238
10 Xu, S. et al. (1995) Roles of URE2 and GLN3 in the proline utilizationpathway of Saccharomyces cerevisiae. Mol. Cell. Biol. 15, 2321–2330
11 Saxena, D. et al. (2003) Rapamycin treatment results in GATA factor-independent hyperphosphorylation of the proline utilization pathwayactivator in Saccharomyces cerevisiae. Eukaryot. Cell 2, 552–559
12 Todd, R.B. and Andrianopoulos, A. (1997) Evolution of a fungalregulatory gene family: the Zn(II)2Cys6 binuclear cluster DNA bindingmotif. Fungal Genet. Biol. 21, 388–405
13 Swaminathan, K. et al. (1997) Crystal structure of the PUT3–DNAcomplex reveals a novel mechanism for DNA recognition by a proteincontaining a Zn2Cys6 binuclear cluster. Nat. Struct. Biol. 4, 751–759
14 des Etages, S.A. et al. (1996) Functional analysis of the PUT3transcriptional activator of the proline utilization pathway inSaccharomyces cerevisiae. Genetics 142, 1069–1082
15 Selleck, S.B. and Majors, J.E. (1987) In vivo DNA-binding propertiesof a yeast transcription activator protein.Mol. Cell. Biol. 7, 3260–3267
16 Huang, H.L. and Brandriss, M.C. (2000) The regulator of the yeastproline utilization pathway is differentially phosphorylated in responseto the quality of the nitrogen source.Mol. Cell. Biol. 20, 892–899
17 des Etages, S.A. et al. (2001) Conformational changes play a role inregulating the activity of the proline utilization pathway-specificregulator in Saccharomyces cerevisiae. Mol. Microbiol. 40, 890–899
18 Axelrod, J.D. et al. (1991) Proline-independent binding of PUT3transcriptional activator protein detected by footprinting in vivo. Mol.Cell. Biol. 11, 564–567
19 Sellick, C.A. and Reece, R.J. (2003) Modulation of transcription factorfunction by an amino acid: activation of Put3p by proline. EMBO J. 22,5147–5153
20 Reece, R.J. (2000)Molecular basis of nutrient controlled geneexpressionin Saccharomyces cerevisiae. Cell. Mol. Life Sci. 57, 1161–1171
21 Yarger, J.G. et al. (1984) Regulation of galactokinase (GAL1) enzymeaccumulation in Saccharomyces cerevisiae. Mol. Cell. Biochem. 61,173–182
22 Broach, J.R. (1979) Galactose regulation in Saccharomyces cerevisiae.The enzymes encoded by the GAL7, 10, 1 cluster are co-ordinatelycontrolled and separately translated. J. Mol. Biol. 131, 41–53
23 Griggs, D. and Johnston, M. (1991) Regulated expression of the GAL4activator gene in yeast provides a sensitive genetic switch for glucoserepression. Proc. Natl. Acad. Sci. U. S. A. 88, 8597–8601
24 Platt, A. and Reece, R.J. (1998) The yeast galactose genetic switch ismediated by the formation of a Gal4p/Gal80p/Gal3p complex. EMBOJ. 17, 4086–4091
25 Larschan, E. and Winston, F. (2001) The S. cerevisiae SAGA complexfunctions in vivo as a coactivator for transcriptional activation byGal4. Genes Dev. 15, 1946–1956
26 Bhaumik, S.R. and Green, M.R. (2001) SAGA is an essential in vivo
target of the yeast acidic activator Gal4p. Genes Dev. 15, 1935–194527 Bhaumik, S.R. et al. (2004) In vivo target of a transcriptional activator
revealed by fluorescence resonance energy transfer. Genes Dev. 18,333–343
28 Roberts, S.M. and Winston, F. (1997) Essential functional interactionsof SAGA, a Saccharomyces cerevisiae complex of Spt, Ada, and Gcn5proteins, with the Snf/Swi and Srb/mediator complexes. Genetics 147,451–465
29 Larschan, E. and Winston, F. (2005) The Saccharomyces cerevisiaeSrb8-Srb11 complex functions with the SAGA complex duringGal4-activated transcription. Mol. Cell. Biol. 25, 114–123
30 Zenke, F.T. et al. (1996) Activation of Gal4p by galactose-dependentinteraction of galactokinase and Gal80p. Science 272, 1662–1665
31 Parthun,M.R. andJaehning, J.A. (1992) A transcriptionally active formof GAL4 is phosphorylated and associated with GAL80. Mol. Cell. Biol.12, 4981–4987
32 Leuther, K.K. and Johnston, S.A. (1992) Nondissociation of GAL4 andGAL80 in vivo after galactose induction. Science 256, 1333–1335
33 Peng, G. and Hopper, J.E. (2000) Evidence for Gal3p’s cytoplasmiclocation and Gal80p’s dual cytoplasmic-nuclear location implicates

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005412
new mechanisms for controlling Gal4p activity in Saccharomyces
cerevisiae. Mol. Cell. Biol. 20, 5140–514834 Peng, G. and Hopper, J.E. (2002) Gene activation by interaction of an
inhibitor with a cytoplasmic signaling protein. Proc. Natl. Acad. Sci.
U. S. A. 99, 8548–855335 Sil, A.K. et al. (1999) The Gal3p–Gal80p–Gal4p transcription switch
of yeast: Gal3p destabilizes the Gal80p–Gal4p complex in response to
galactose and ATP. Mol. Cell. Biol. 19, 7828–784036 Schell, M.A. and Wilson, D.B. (1977) Purification and properties
of galactokinase from Saccharomyces cerevisiase. J. Biol. Chem. 252,1162–1166
37 Timson, D.J. and Reece, R.J. (2002) Kinetic analysis of yeastgalactokinase: Implications for transcriptional activation of the GAL
genes. Biochimie 84, 265–27238 Yano, K.I. and Fukasawa, T. (1997) Galactose-dependent reversible
interaction of Gal3p with Gal80p in the induction pathway ofGal4p-activated genes of Saccharomyces cerevisiae. Proc. Natl.
Acad. Sci. U. S. A. 94, 1721–1726
39 Platt, A. et al. (2000) The insertion of two amino acids into atranscriptional inducer converts it into a galactokinase. Proc. Natl.
Acad. Sci. U. S. A. 97, 3154–315940 Bhat, P.J. et al. (1990) Analysis of the GAL3 signal transduction
pathway activating GAL4 protein-dependent transcription inSaccharomyces cerevisiae. Genetics 125, 281–291
41 Bhat, P.J. and Hopper, J.E. (1992) Overproduction of the GAL1 orGAL3 protein causes galactose-independent activation of the GAL4protein: evidence for a newmodel of induction for the yeastGAL/MEL
regulon. Mol. Cell. Biol. 12, 2701–270742 Verma, M. et al. (2004) Expression of GAL genes in a mutant strain
of Saccharomyces cerevisiae lacking GAL80: quantitative model andexperimental verification. Biotechnol. Appl. Biochem. 39, 89–97
43 Bhat, P.J. and Venkatesh, K.V. (2005) Stochastic variation in theconcentration of a repressor activates GAL genetic switch: impli-cations in evolution of regulatory network. FEBS Lett. 579, 597–603
44 Meyer, J. et al. (1991) Galactokinase encoded by GAL1 is abifunctional protein required for induction of the GAL genes inKluyveromyces lactis and is able to suppress the gal3 pheontype inSaccharomyces cerevisiae. Mol. Cell. Biol. 11, 5454–5461
45 Thoden, J.B. and Holden, H.M. (2003) Molecular structure of
galactokinase. J. Biol. Chem. 278, 33305–3331146 Hartley, A. et al. (2004) Substrate specificity and mechanism from
the structure of Pyrococcus furiosus galactokinase. J. Mol. Biol. 337,387–398
47 Thoden, J.B. et al. (2005)Molecular structure of human galactokinase:implications for type II galactosemia. J. Biol. Chem. 280, 9662–9670
48 Menezes, R.A. et al. (2003) Sites for interaction betweenGal80p andGal1pin Kluyveromyces lactis: structural model of galactokinase based onhomology to the GHMP protein family. J.Mol. Biol. 333, 479–492
Endea
Coming soon in the qua
the history and philo
The accidental conservat
Powering the porter bre
Female scientists in fi
Locate Endeavour on ScienceDirect
www.sciencedirect.com
49 Blank, T.E. et al. (1997) Novel Gal3 proteins showing altered Gal80pbinding cause constitutive transcription of Gal4p-activated genes inSaccharomyces cerevisiae. Mol. Cell. Biol. 17, 2566–2575
50 Suzuki-Fujimoto, T. et al. (1996) Analysis of the galactose signaltransduction pathway in Saccharomyces cerevisiae: interactionbetween Gal3p and Gal80p. Mol. Cell. Biol. 16, 2504–2508
51 Nogi, Y. et al. (1977) Interaction of super-repressible and dominantconstitutive mutations for the synthesis of galactose pathwayenzymes in Saccharomyces cerevisiae. Mol. Gen. Genet. 152, 137–144
52 Pilauri, V. et al. (2005) Gal80 dimerization in the yeast GAL geneswitch. Genetics 169, 1903–1914
53 Melcher, K. and Xu, H.E. (2001) Gal80–Gal80 interaction on adjacentGal4p binding sites is required for complete GAL gene repression.EMBO J. 20, 841–851
54 Timson, D.J. et al. (2002) Gal3p and Gal1p interact with the tran-scriptional repressor Gal80p to form a complex of 1:1 stoichiometry.Biochem. J. 363, 515–520
55 Kingston, R.L. et al. (1996) The structure of glucose-fructose oxido-reductase from Zymomonas mobilis: an osmoprotective periplasmicenzyme containing non-dissociable NADP. Structure 4, 1413–1428
56 Flynn, P.J. and Reece, R.J. (1999) Activation of transcription bymetabolic intermediates of the pyrimidine biosynthetic pathway. Mol.Cell. Biol. 19, 882–888
57 Rebora, K. et al. (2001) Yeast AMP pathway genes respond to adeninethrough regulated synthesis of a metabolic intermediate. Mol. Cell.Biol. 21, 7901–7912
58 Kohlhaw, G.B. (2003) Leucine biosynthesis in fungi: enteringmetabolism through the back door. Microbiol. Mol. Biol. Rev. 67, 1–15
59 El Alami, M. et al. (2002) The proper folding of a long C-terminalsegment of the yeast Lys14p regulator is required for activation ofLYS genes in response to the metabolic effector. Mol. Microbiol. 43,1629–1639
60 Amar, N. et al. (2000) ArgRII, a component of the ArgR–Mcm1complex involved in the control of arginine metabolism in Saccharo-myces cerevisiae, is the sensor of arginine. Mol. Cell. Biol. 20,2087–2097
61 Schuller, C. et al. (2004) Global phenotypic analysis and transcrip-tional profiling defines the weak acid stress response regulon inSaccharomyces cerevisiae. Mol. Biol. Cell 15, 706–720
62 Kren, A. et al. (2003) War1p, a novel transcription factor controllingweak acid stress response in yeast. Mol. Cell. Biol. 23, 1775–1785
63 Gustin, M.C. et al. (1998) MAP kinase pathways in the yeastSaccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 62, 1264–1300
64 Posas, F. et al. (1998) Signal transduction by MAP kinase cascades inbudding yeast. Curr. Opin. Microbiol. 1, 175–182
65 Banuett, F. (1998) Signalling in the yeasts: an informational cascadewith links to the filamentous fungi. Microbiol. Mol. Biol. Rev. 62,249–274
66 Bassett, J.H. et al. (2003) Mechanisms of thyroid hormone receptor-specificnuclearandextranuclearactions.Mol.Cell.Endocrinol.213,1–11
vour
rterly magazine for
sophy of science:
ionist by M.A. Andrei
wery by J. Sumner
lms by B.A. Jones
(http://www.sciencedirect.com)

Shedding light on ADAMmetalloproteinasesAri-Pekka J. Huovila1, Anthony J. Turner2, Markku Pelto-Huikko3, Iivari Karkkainen1
and Rebekka M. Ortiz1
1Institute of Medical Technology, University of Tampere and Tampere University Hospital, Biokatu 6, FIN-33520 Tampere, Finland2Proteolysis Research Group, School of Biochemistry and Microbiology, University of Leeds, Leeds LS2 9JT, UK3Departments of Developmental Biology and Pathology, University of Tampere Medical School and Tampere University Hospital,
33014 University of Tampere, Finland
ADAM metalloproteinase disintegrins have emerged as
the major proteinase family that mediates ectodomain
shedding, the proteolytic release of extracellular
domains from their membrane-bound precursors.
Recent gene-manipulation studies have established
the role of ADAM-mediated shedding in mammalian
physiology and, in addition, raised the issue of func-
tional redundancy among ADAM sheddases. ADAM
sheddases activate, for example, growth factors and
cytokines, thus regulating signalling pathways that are
important in development and pathological processes
such as cancer. The recent studies have also begun to
elucidate the substrate specificity and the mechanisms
that control ADAM-mediated shedding events that regu-
late, for example, growth-factor and Notch signalling, and
the processing of the amyloid precursor protein.
ADAM metalloproteinases comprise the major sheddase
family
Cellular membrane proteins can be proteolytically cleavedat their juxta-membrane region in a process called ecto-domain shedding, which results in detachment of theirextracellular region (the ectodomain). Shedding can releasecytokines, growth factors or other mediators from theirmembrane-bound precursors or, conversely, down-regulatereceptors and other proteins from the cell surface.Membersof the ADAM (a metalloprotease and disintegrin) familyhave been identified as proteinases that mediate diverseshedding events [1–4]. Although ADAMproteins seem to beresponsible of majority of known shedding, other proteases,including members of the matrix metalloproteinase (MMP)family, also shed several substrates [1,4,5].
ADAMs have been found in animal species fromprotozoans to mammals; O30 ADAM orthologues havebeen identified, 29 of which are found in mammals. Theybelong to the metzincin subfamily of metalloproteinasesand share a characteristic domain structure that isresponsible for their proteolytic, adhesion and putativesignalling activities [1,3,6] (Figure 1). Genome databasesindicate the existence of several additional human andmouse ADAMs (see Box 1). However, not all ADAMs are
Corresponding author: Huovila, A.-P.J. ([email protected]).Available online 9 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
proteinases:w50% do not contain all the amino acids thatare crucial for the catalytic activity [1,2].
Here, we review recent studies that have addressed thesubstrate specificity of ADAMs and regulation of theirsheddase activity, and discuss select key findings such asthe role of ADAMs in the regulation of epidermal growthfactor (EGF)-receptor (EGFR) and Notch signalling, andin the processing of the amyloid precursor protein (APP).We refer to ADAMs with an intact catalytic site asmetalloproteinase ADAMs and to those implicated inectodomain shedding as sheddase ADAMs.
Table 1 lists the ADAM substrates indicated in cell-based shedding experiments or in vitro cleavage assays,including type I and II, glycosylphosphatidyl inositol-anchored and indirectly membrane-associated proteins.Conspicuously, ADAM-17 and ADAM-10 seem to be theprincipal sheddases. Although the contribution of otherADAMs is increasingly evident, the current understand-ing of ADAM-mediated shedding is largely based onstudies of ADAM-17 and ADAM-10.
ADAM-17, the prototype sheddase
ADAM-17 was the first ADAM implicated in ectodomainshedding by its identification as the tumour necrosisfactor-a (TNFa) convertase [5,7]. Since then, ADAM-17has been established as an essential sheddase inmammalian development [8–11]. However, in addition tothis role in development, the number and variety of othersubstrates (e.g. TNF receptors and other cytokines andcytokine receptors; Table 1) indicate a wider role forADAM-17 in adults [5].
The major contribution of ADAM-17 to the regulation ofcytokine signalling, in addition to TNFa activation, is indi-cated by its participation in shedding of several cytokinereceptors. Such a role is supported by the increasedADAM17 expression during inflammation [3]. The mis-regulated sheddase activity of ADAM-17 in TIMP (tissueinhibitor of metalloproteinases)-3-deficient mice corrobor-ates its importance in the inflammatory response [12].
ADAM-17 also regulates growth-factor and hormonesignalling. As the major sheddase of several EGF-familygrowth factors [13,14], ADAM-17 has been implicatedin both ‘direct’ activation and G-protein-coupledreceptor (GPCR)-induced transactivation of EGFR [13–15].
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
. doi:10.1016/j.tibs.2005.05.006

Ti BS
100 aa
Cysteine switch
N- -CDI ACRMP E CTTMPRO
TIMP inhibition
TIMP-1 ADAM-10, (-17)TIMP-2 ADAM-17, (-33)TIMP-3 ADAM-10, -12, -17, -28, -33TIMP-4 ADAM-17, -28, -33
Pro-domainremoval
PPC,autocatalysis(ADAM-8, -28)
Integrin binding
α4β1 ADAM-28α5β1 ADAM-15, -17α6β1 ADAM-9α7β1 ADAM-12α9β1 ADAM-12, -15αVβ3 ADAM-15αVβ5 ADAM-9
Cytosolic proteininteraction motifs
SH3 (PxxP)SH2 (p-Tyr)PDZ (e.g. GDxV-cooh)WW (e.g. PPxY)EVH1 (PPP+PSKxxR)14-3-3 (e.g. RSx[pS/pT]xP)Tyr phosphorylationSer/Thr phosphorylation
Figure 1. Schematic representation of the domain structure and molecular interactions of ADAM metalloproteinases. In the ‘cysteine-switch’ mechanism, a cysteine residue
in the pro-domain forms an intramolecular bond with the zinc atom at the catalytic site (red arrow), thus blocking the active site [3,6]. Proteolytic removal of the pro-domain
activates the metalloproteinase domain by unblocking the active site [3,6]. ADAM metalloproteinases can be differentially inhibited by the TIMP. The TIMP-inhibition profiles
of the sheddase ADAMs are included (ADAM-8 and ADAM-9 are not inhibited by TIMPs) [31,35]. ADAM-17 and ADAM-33 are shown in brackets because inhibition by TIMP-1
and TIMP-2, respectively, is minor. The integrin-binding abilities of the sheddase ADAMs are listed, the arrow points to the putative integrin-binding motif (‘disintegrin loop’)
in the disintegrin domain [6]. The putative binding motifs for different interaction domains and kinase target sites found in the ADAM cytosolic tail sequences are also listed
(the sequence motif in parentheses). Abbreviations: aa, amino acids; ACR, ADAM cysteine-rich domain; CT, cytosolic tail; DI, disintegrin domain; E, Epidermal growth factor-
like domain; EVH1, enabled/vasodilator-stimulated phosphoprotein homology 1; MP, metalloproteinase domain; PDZ, post-synaptic density-95/discs large/zonula occudens-
1; PPC, pro-protein convertase; PRO, pro-domain; TIMP, tissue inhibitor of metalloproteinases; TM, transmembrane region; SH2/SH3, Src-homology domain; WW, domain
with two conserved tryptophan (W) residues; 14–3-3, 14–3-3 domain.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005414
ADAM-17 regulates growth-hormone signalling by releas-ing growth-hormone-receptor ectodomains, which can actas circulating decoy receptors [16].
From MADM to ADAM-10
A membrane proteinase, originally isolated as a myelinbasic protein-degrading enzyme from bovine brain, was
Box 1. Web resources related to ADAM metalloproteinases
† The White laboratory ADAMs page, the ‘official ADAM database’,
well linked to other resources and regularly updated: http://www.
people.virginia.edu/%7Ejw7 g/Table_of_the_ADAMs.html
† Other updated ADAMs pages with links to other resources: http://
www.uta.fi/%7Eloiika/ADAMs/HADAMs.htm; http://www.uta.fi/
%7Eloiika/ADAMs/MMADAMs.htm
† ADAM gene family page of Human Gene Nomenclature Commit-
tee. Links to related databases: http://www.gene.ucl.ac.uk/nomen-
clature/genefamily/metallo.html
† The Apte laboratory web page. Excellent resource on the related
ADAMTS family (ADAMs with thrombospondin motifs). Links to
other resources: http://www.lerner.ccf.org/bme/apte/adamts/
nomenclature.php
† An information resource for proteinases and proteins that inhibit
them. Information about ADAMs can be found by following the links
to the M12 (astacin/adamalysin) family: http://merops.sanger.ac.uk/
www.sciencedirect.com
subsequently identified as an ADAM and termed MADM(mammalian disintegrin metalloproteinase). It was latershown to be identical with ADAM-10 and, its orthologue,Kuzbanian, was implicated in early neural developmentin Drosophila by its proteolytic activation of Notchsignalling [17]. The severe phenotype of adam10K/K
embryos (see later; Table 2) suggests that ADAM-10 isalso involved in the shedding of Notch in mammals [18].Moreover, ADAM-10 has been implicated in other shed-ding events in the regulation of developmental processessuch as axonal pathfinding [19,20].
In addition to neural development, ADAM-10 seems tobe involved in physiological shedding events in the adultbrain. Strong evidence suggests that ADAM-10 mediatesthe non-amyloidogenic processing of APP [21]. In cell-based studies, ADAM-10 and ADAM-17 have been shownto carry out the constitutive and regulated ‘normal’processing of the cellular prion protein, respectively[22,23]. These results suggest that defective or mis-regulated ADAM-10 function might contribute to neuro-degenerative processes such as Alzheimer’s disease andspongiform encephalopathies [17,21,22].
Together with ADAM-17, ADAM-10 plays a central partin the regulation of EGFR signalling. In an extensive

Table 1. ADAM sheddase substrates and regulationa
Substrate ADAMsc Regulation Refs
Cytokines
CX3CL-1 10C, 17R PMA [57,58]
CXCL-16 10C,R IFNg, TNFa [59,60]
TNFa 17R, 10, 9, 8 PMA, LPS, glutamate [5,7]b
TRANCE 19C, 17 [6,30]b
Growth factors
Amphiregulin 17R, 15R PMA, carbachol, LPA, tobacco smoke, cannabinoids [13–15]b
Betacellulin 10C [13]
EGF 10C [13]
Epiregulin (9, 12, 15)R, 17R PMA [13,14]
HB-EGF 9R, 10R, 12R, 17R TPA, PMA, GPCR agonists [13–15,25,36,55]b
Neuregulins 17, 19C,R PMA [29,61]b
TGFa 17R, 15R PMA, FGF, PDGF, LPA [8,13–15]b
Receptors
CD23 8, 15, 28 [62]
CD30 17R PMA [39]
CD40 17C [63]
c-Kit 17R PMA [64]
ErbB4 17C,R PMA, PV [65]
GHR 17R PMA [16]
IL-1 receptor-II 17R, 8 PMA [5,35]b
IL-6 receptor 10C, 17R PMA, MCD, IL-8, C5a, PAF, LTB4, FMLP [40,66]b
IL-15 receptor-a 17C,R PMA [67]
M-CSFR 17R TPA, LPS [68]
P75NTR 17R PMA, PV [48]
TNF receptor-I 17R, 8 PMA [5,7]b
TNF receptor-II 17R, 9 PMA, TNFa [5,6,8]b
trkA (NGFR) 17R PMA, PV, UV, osmotic stress [46]
Adhesion proteins
CD44 10R, 17R Ca2C, PMA [69,70]
CHL1 8C [71]
L1 10R PMA, MCD [72]
L-selectin 17R PMA, DPA-NSAIDs [8,73]
VCAM-1 17R PMA [74]
Proteinases
BACE1 10R PMA [75]
Meprin-b 17R PMA [76]
Oxytocinase 9, 12 [77]
Other proteins
APP 9, 10C,R, 17R PMA [21,51]b
Collagen XVII 9, 10, 17 [78]
Notch 10, 17 [18]b
Delta 10 [79–81]b
Jagged 17R (or 10R) PMA [81]
Ephrin-A2 10R [19]
KL-1 8, 9, 17, 33C [31,35]
Mucin-1 17R PMA [82]
PrP 10C,R, 17R TPA [22,23]b
Slit 10 [20]aAbbreviations: APP, amyloid precursor protein; BACE, b-site APP-cleaving enzyme; CD, cluster of differentiation; CHL1, close homologue of L1; CXCL/CX3CL, CXC/CX3CL
chemokine ligand; DPA-NSAIDs, diphenylamine-based non-steroid anti-inflammatory drugs; EGF, epidermal growth factor; FGF, fibroblast growth factor; GHR, growth
hormone receptor; GPCR, G-protein-coupled receptor; HB-EGF, heparin-binding EGF-like growth factor; IFN, interferon; IL, interleukin; LPA, lysophosphatidic acid; LPS,
lipopolysaccharide; LTB4, leukotriene B4; MCD, methyl b-cyclodextrin; M-CSFR, macrophage-colony-stimulating factor receptor; NGFR, nerve growth factor receptor;
p75NTR, p75 neurotensin receptor; PAF, platelet-activating factor; PDGF, platelet-derived growth factor; PMA, phorbol myristate acetate; PrP, prion protein; PV, pervanadate;
TGF, transforming growth factor; TNF, tumour necrosis factor; TPA, tetradecanoylphorbol myristate acetate; TRANCE, TNF-related activation induced cytokine; trkA,
neurotrophic tyrosine kinase type 1; VCAM, vascular cell-adhesion molecule; UV, ultraviolet radiation.bThe reference contains additional relevant references.cThe ADAMs shown in parentheses indicate that the evidence is not conclusive or unambiguous, see the text for further explanation. Superscript ‘C’ and ‘R’ denote reported
constitutive or regulated sheddase activity, repectively.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 415
study, Sahin et al. [13] demonstrated that ADAM-10 isthe main sheddase of EGF and betacellulin, and otherstudies have indicated ADAM-10 in GPCR–EGFR trans-activation [2,15].
Lessons from gene knockout experiments
Mice lacking functional ADAM-8, ADAM-9, ADAM-12or ADAM-15 largely develop normally, although some
www.sciencedirect.com
adam12K/K mice show minor defects during development[24–27]. As indication of these ADAMs in diverse sheddingevents in cell-based assays (Table 1), corroborated by theirwide expression in mammalian tissues, suggests that theyfunction as physiological sheddases, Sahin et al. [13]generated adam9K/K, adam12K/K and adam15K/K mice,all of which also showed apparently normal phenotype.Although characterization of fibroblasts isolated from

Table 2. ADAM metalloproteinase gene manipulation in micea,b
Genotype Phenotype Refs
adam8K/K Viable, fertile, no obvious pathology [27]
adam9K/K Viable, fertile, no obvious pathology [24]
adam10K/K Embryonic lethal (E9.5), defective heart and CNS development, vasculogenesis and somitogenesis;
defective Notch signalling. APP processing normal in mEFs
[18]
adam10 overexpression;
APP[V717I]
Increased secretion of APPsa; reduced b-amyloid and amyloid plaque formation; impaired LTP and
cognitive deficits alleviated
[21]
Dominant negative adam10
overexpression; APP[V717I]
Increased number and size of amyloid plaques in the brains [21]
adam12K/K Viable, fertile, grossly normal; high post-natal death rate; minor brown fat and neck muscle
hypotrophy. Reduced PMA-stimulated shedding of HB-EGF in mEFs
[25]
adam12 overexpression Dystrophic pathology alleviated in mdx mice. Increased expression and redistribution of a7B
integrin and utrophin in mdx and normal mice. Induction of adipogenesis
[28]
adam15K/K Viable, healthy, no obvious pathology. Decreased neovascularization in experimental pathologies [26]
adam17K/K Perinatal lethal; general epithelial dysgenesis, defective heart and lung development, deficient
ectodomain shedding
[8–11]
adam19K/K Most died within a few days after birth; defective heart development, normal bone and lung
development
[30]
adam19K/K Most died within a week; defective heart development [29]
adam9/15K/K Viable, fertile, no obvious pathology [13]
adam9/12/15K/K Viable, fertile, no obvious pathology. mEFs showed reduced PMA-stimulated shedding of
epiregulin
[13]
adam9/12/15/17K/K Like adam17K/K (slightly higher embryonic lethality) [13]aInformation on mice lacking non-proteolytic ADAMs are not included.bAbbreviations: APP, amyloid precursor protein; APP[V717I], mice transgenic for human APP[V717I]; CNS, central nervous system; HB-EGF, heparin-binding epidermal growth
factor; LTP, long-term potentiation; mEF, mouse embryonic fibroblast; PMA, phorbol myristyl acetate; wt, wild type.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005416
these mice indicates possible contribution to the sheddingof EGFR ligands, the physiological functions of ADAM-9,ADAM-12, or ADAM-15 (alone or in combination) do notseem to be essential for normal development. See Table 2for a summary of the sheddase adam-manipulationexperiments in mice.
In a mouse model, overexpression of adam12 has beenshown to alleviate the pathology of muscular dystrophy,suggesting a role in muscle regeneration [28]. In addition,upregulated adipose tissue formation in adam12-over-expressing mice and (minor) defects in brown fat andadjacent muscle formation in some adam12K/K micemight indicate a regulatory function for ADAM-12 inmuscle and adipose development [25].
ADAM-10 is crucial to early embryogenesis [18]: Notch-signalling is defective in adam10K/K mice. Gene knockoutresults in decreased expression of a Notch target gene andup-regulation of a Notch ligand in the neural tube. Dueto early embryonic death, the putative function as APPa-secretase could not be tested in adam10K/K neuronalcultures. However, overexpression of wild-type anddominant-negative adam10 in mice that are geneticallydisposed to Alzheimer’s disease has provided convincingevidence for ADAM-10 as the principal a-secretase inmouse brain [21].
ADAM-17-deficient embryos show defects reminiscentof those in transforming growth factor-a (TGFa)-deficientmice, with additional defects similar to those caused bythe lack of EGFR [8]. Further characterization hasverified that ADAM-17 is needed for epithelial differen-tiation and growth in developing heart and lung, and forshedding of several substrates, including several EGFRligands [8–11,13,14].
The defects in adam19K/K mice indicate an importantrole of ADAM-19 in morphogenesis and remodelling ofthe endocardial cushion of the developing heart [29,30].The conclusions on the actual sheddase function are
www.sciencedirect.com
controversial: one study showed evidence for ADAM-19as a neuregulin sheddase in the developing endocardialcushion mesenchyme [29], whereas another indicated thatADAM-19 is not a neuregulin sheddase therein [30].However, because these cell-based shedding assays wereessentially identical, it remains to be investigatedwhether these results reflect actual differences betweenthe two lines of adam19K/K mice.
The lack of obvious pathological phenotype in micedeficient of functional ADAM-9, ADAM-12 or ADAM-15was suggested to reflect possible functional redundancywith other proteinases (probably ADAMs) [24–26].
Substrate selectivity and potential mechanisms for
regulation of ADAM sheddases
To understand how individual sheddase ADAMs mightcleave different substrates and several substrates can becleaved by different ADAMs, it is important to understandhow ADAMs recognize their substrates and how theirsheddase activity might be regulated.
Intrinsic substrate selectivity at the active site of someADAMs is indicated by the preference for residues flank-ing the scissile bond in a subset of substrates [3,5,31]. Bycontrast, the variability of the cleavage-site sequences andtheir tolerance of mutations indicates some pliability inthe substrate recognition at the active sites [1,5].
Despite the lack of a consensus cleavage motif, it isclear from in vitro studies that there can be substantialdifferences in the cleavage efficiency of a given substratecut by different ADAMs, and in different substrates cut bya given ADAM [3,5,6]. This, however, cannot be directlyextrapolated to shedding of full-length proteins on the cellsurface because the cleavage efficiency can be different formembrane-bound substrates and their peptide analogues[5]. Whereas both native membrane-bound TNFa andpeptide analogues of its cleavage site are readily cut byADAM-17, peptide mimetics of other substrates are cut far

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 417
less efficiently [5], indicating weak contact at the activesite and suggesting that interactions distal to the cleavagesite contribute to the binding energy [1,5]. Interestingly, amembrane-distal region of TNFa ectodomain enablesphorbol myristate acetate (PMA)-stimulated shedding byADAM-17 only, but allows constitutive shedding by otherADAMs [32].
Membrane attachment of both the sheddase and thesubstrate is required for at least some shedding events[1,5,6]. Moreover, the secondary structure of the juxta-membrane stalk might contribute to substrate recognition[3,31]. By contrast, some soluble ADAM isoforms canprocess certain substrates [3], further emphasizing thatprediction of shedding activities based on differences incleavage efficiency in vitro must be made with caution.
Shedding activity of ADAMs can be regulated withpharmacological agents or by subjecting the cells toconditions that induce shedding (Table 1). Althoughthe regulation of ADAM proteinases is, as yet, poorlyunderstood, potential regulatory mechanisms have beenrecognized.
Transcriptional control
The expression of genes encoding ADAMs can be tran-scriptionally regulated in response to pathological con-ditions and experimentally, for example, by extracellularfactors such as cytokines and growth factors [3,6]. Forinstance, ADAM17 expression is upregulated duringinflammatory response, and oxygen-glucose deprivationcan induce TNFa shedding in rat forebrain slices andneuronal cells by upregulation of ADAM17 expression [33].
Alternative splicing
Splice variants have been reported for several ADAMgenes [3]. Interestingly, alternative splicing often affectsthe cytosolic region of the deduced ADAM protein, givingrise to different cytosolic tails or resulting in soluble, andthus probably secreted, isoforms [3]. For example, severalhuman ADAM15 mRNA variants arise from differentialuse of alternative cytosolic tail-coding exons [34]. This useof alternative exons seems to be non-random and, hence,probably regulated by as yet unknown mechanisms [34].Because several cytosolic interactions have been reportedfor ADAM-15, alternative splicing could affect theregulation of ADAM-15 function or subcellular location.
Post-translational processing
The proper function of ADAM metalloproteinases must becontrolled by preventing potential harmful effects withinthe cells themselves or in their environment while pro-moting the action at an appropriate location and time. Inmost cases, newly synthesized proteolytic ADAMs arekept latent by the pro-domain by a ‘cysteine switch’ [3,6].Removal of the pro-domain by pro-protein convertases isan established key step inADAMactivation [3,6] (Figure 1).
ADAM ectodomains and external factors
Another function of the pro-domain is to facilitate foldingand passage through the quality-control checks along thesecretory pathway [2,3]. Interestingly, following the proteo-lytic separation of the pro-domains and metalloproteinase
www.sciencedirect.com
domains the pro-domains of ADAM-12, ADAM-17 andADAM-33 can, at least to some extent, remain non-covalently bound to the metalloproteinase domain intransfected cells (see Ref. [31] and references therein).
Regulated binding of another protein by the ADAMcatalytic domain might link intracellular signals to theactivation of the actual shedding. PMA stimulation canenhance peptide-substrate processing by ADAM-17 inde-pendently of its cytosolic domain, suggesting that theintrinsic activity could be enhanced by removal of aninhibitor or by binding of an activator [5].
The proteolytic activity of several ADAMs can beinhibited by the TIMPs (Figure 1), which are physio-logical inhibitors of MMPs [31,35]. Although in most casesthe physiological relevance of TIMPs remains unclear,inactivation of the gene encoding TIMP-3 causes anincrease of ADAM-17 activity and sustained TNFashedding in mice [12].
The structural features discussed here, and the some-what promiscuous substrate recognition of ADAMs, haveprimed proposals that their downstream extra-cytosolicdomains might also regulate shedding activity eitherdirectly by binding to the substrate or indirectly via athird protein [1,3,5]. The models presented by White [1]and Moss and Bartsch [7] suggest that an interactionbetween the sheddase ADAM, tetraspanins, integrins andthe substrate might regulate the sheddase function.Indeed, Yan et al. [36] have demonstrated that theGPCR-induced heparin-binding EGF-like growth factor(HB-EGF) shedding by ADAM-10 involves its associationwith a signalling complex containing a tetraspanin andthe substrate. In addition, the importance of the dis-integrin and/or cysteine-rich domains of ADAM-17 andADAM-13 for their sheddase activity [1,2,5] is consistentwith these models.
Cytosolic interactions
Cytosolic tails of most sheddase ADAMs contain putativerecognition motifs for signalling proteins and adaptors(Figure 1). Several such interactions have been demon-strated [3,6] and it is plausible that they could participatein regulation of metalloproteinase activity or subcellularlocalization of sheddase ADAMs. Recently, two mouseADAM-15 tail isoforms were shown to interact differen-tially with Src family protein tyrosine kinases in vitro [37],corroborating the potential regulation of cytosolic ADAMinteractions by alternative splicing. By contrast, as men-tioned, experimental stimulation does not always requirethe cytosolic tail of the sheddase ADAM [2,5].
Location – membrane environment
The majority of the shedding events are thought occur atthe cell surface, but ADAM sheddases might also actintracellularly, possibly depending on cell type and theshedding process [3].
The aforementioned tetraspanin-complex model [1,7]suggests regulation of ADAMs by association with selectmembrane micro-domains. Shedding of neuregulin byADAM-19 was recently demonstrated to depend on thelocation of both the sheddase and the substrate in lipidrafts (local cholesterol-enriched lateral assemblies in the

Ti BS
p38 ?
MEK
ERK
Shedding activation
UV PV
PKC
p38
SorbitolGPCR
PMAEGF
TGFα
Figure 2. MAPK pathways regulate ADAM-mediated ectodomain shedding. MAPKs
Erk and p38 are central to the activation of ADAM-mediated ectodomain shedding.
This scheme combines the observations from several recent studies (see Refs
[46–49] and references therein). Stimulation by phorbol esters (denoted here by
PMA) or growth factors (EGF, TGFa) is mediated by PKC and the Mek/Erk pathway
(MEK, ERK). PMA stimulation can also be mediated independently of Erk, via
unknown route(s) (denoted by a question mark). Stimulation of PV is also signalled
via the Mek/Erk pathway. Osmotic stress-induced stimuli (e.g. sorbitol) can be
mediated via both Mek/Erk and p38 pathways, whereas p38 mediates the UV-
induced stress stimulus. Stimulation of GPCR by, for example, muscarine or N-
formyl-methionyl-leucyl-phenylalanine, can activate shedding via both Mek/Erk
and p38 pathways. The basal constitutive shedding seems to depend on signals
mediated by p38 MAPK (depicted as p38). Abbreviations: EGF, epidermal growth
factor; ERK, extracellular signal-regulated kinase; GPCR, G-protein-coupled
receptors; MAPK, mitogen-activated protein kinase; MEK, MAPK/ERK kinase; PKC,
protein kinase C; PMA, phorbol myristate acetate; PV, pervanadate; TGFa,
transforming growth factor a; UV, ultraviolet radiation.
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005418
plasma membrane) [38]. By contrast, decreasing cellularcholesterol and disruption of the rafts increases sheddingof APP, interleukin-6 receptor, and CD30 by ADAM-10 andADAM-17 [39,40]. Because targeting of APP and b-siteAPP-cleavage enzyme 1 (BACE1) to rafts seems to benecessary for the amyloidogenic-b cleavage [41,42], APPlocation inside or outside rafts might regulate the balancebetween the amyloidogenic and non-amyloidogenicprocessing.
Substrate regulation
It is commonly assumed that the sheddase enzyme medi-ates the stimulation of ectodomain shedding by PMA [3,5].However, the ability of the membrane-distal domain ofL-selectin and the angiotensin-converting enzyme toconfer the inducibility to other, natively non-regulatedsubstrates, is consistent with a conformational changethat exposes a cryptic recognition site to the sheddase uponstimulation [43,44].Therefore, thesedomainsmightprovideexamples of substrate-mediated activation of shedding.
Sheddase stability
High-dose PMA-stimulation decreases the half-life ofmature ADAM-17 on the cell surface, probably viaendocytic degradation [6,45]. This might be specific toADAM-17 because ADAM-10 was not down-regulated[45]. Nevertheless, the cells might control the specificityand kinetics of PMA-induced shedding by down-regulat-ing ADAM-17 or a subset of sheddases.
It is obvious that at least some of these mechanismsmight be involved in the regulation of shedding eventsbut it is unclear how they might be linked to theregulatory stimuli.
MAP kinase pathways signal the activation of
ADAM-mediated shedding
Accumulating studies have informed the view thatmitogen-activated protein kinases (MAPKs) Erk and p38mediate most, albeit not all, of the shedding-activationsignals triggered by growth factors, stress and commonlyused non-physiological drugs [46–49]. An emerging themeis that regulated shedding is often activated via theMek/Erk pathway, whereas the constitutive basal shed-ding seems to be dependent on signals via p38 [3,5](Figure 2). For example, both the muscarine-stimulatedand the EGF-dependent shedding of APP by thea-secretase pathway are predominantly dependent onthe Erk/protein kinase Ca (PKCa) pathway [49]. Bycontrast, shedding activation by GPCR stimulation canbe induced through both Erk and p38 pathways (see Ref.[47] and references therein). As depicted in Figure 2,phorbol-ester stimulation can also activate sheddingindependently of Erk [48].
It is not understood, however, how these pathways arecoupled to the shedding event itself. A reasonablehypothesis is that some of the regulatory mechanismscould be engaged. Another question awaiting clarificationis to what extent the experimental activation correspondsto the physiological regulation of shedding. In thefollowing, we present examples illustrating how ADAM-mediated shedding contributes to mammalian physiology
www.sciencedirect.com
and how mis-regulated shedding might contribute tohuman disease.
ADAM sheddases in the processing of APP and Notch
ADAMs have been implicated in several processesduring central nervous system (CNS) development, andtheir wide expression also indicates diverse roles inthe adult CNS [50]. Recent studies provide strong evi-dence for ADAMs in the physiological processing of APPand Notch [3,17].
b-Amyloid plaques are a hallmark diagnostic ofAlzheimer’s disease (AD). APP is normally processed bythe a-secretase pathway, which does not lead to depositionof the b-amyloid fragments that are generated by thealternative b-secretase pathway. However, it has not beenclear whether b-amyloid deposition actually contributes tothe AD pathogenesis or whether the a-sectetase andb-secretase pathways are mutually exclusive. The recentstudy addressing the putative role of ADAM-10 in thea-processing provides novel clarification to this issue [21].
ADAM-9, ADAM-10 and ADAM-17 can mediate non-amyloidogenic a-processing of APP in cell lines and inpeptide-cleavage assays [17]. They all contribute to thea-processing in glioblastoma cells [51]. In a neuroblastomaline, ADAM-10 and, to a lesser extent, ADAM-17,contribute to this processing event [52]. ADAM-10 and

Ti BS
(4)
(3)(2)(1)
Notch
APP
(5)
(6)
(2) (1)
(3)
GPCREGFR
α α β
γ γ
(a)
(b)
ProteasomeIDE
Nucleus transcription
Figure 3. Regulated intramembrane proteolysis and GPCR–EGFR transactivation
mediated by ADAMs. (a) Notch and amyloid precursor protein (APP) are integral
plasma membrane proteins (1) that can undergo ectodomain shedding and regulated
intramembrane proteolysis (RIP; 2 and 3) – so named because the proteinase (the
presenilin complex) at the cytosolic side seems to cleave a peptide bond that is
predicted to reside within the phospholipid bilayer of the cell membrane [53]. Notch
processing (dark blue ectodomain) is an example of ligand-induced RIP, and APP
processing (yellow ectodomain) represents ligand-independent RIP. Processing of
Notch is initiated by binding of its ligand (e.g. Delta, which is, for simplification,
depicted in its soluble form; orange), which triggers the ectodomain shedding (2) by
a-secretase (magenta). Followinga-cleavage, the intracellulardomainofboth proteins
(green) is released to the cytosol by the presenilin complex (light blue; g, for
g-secretase) at a position within the predicted transmembrane part of the proteins (3).
The released intracellular domains (ICD; green) are translocated into the nucleus (4)
where their association with transcription factors can activate specific gene
expression. Cytosolic ICDs of Notch and APP are eventually degraded (5) by the
proteasome or the insulin-degrading enzyme (IDE). Alternatively to the non-
amyloidogenic a-processing of APP, the b-site APP-cleaving enzyme (BACE; black)
can cleave at a different position (2), which, followed by the g-secretase cleavage,
results in the release of the amyloidogenic b-peptide (red; 6). a-secretase cleaves APP
within the b-peptide sequence, thus preventing the b-peptide formation. This
schematic presentation is based on the studies reviewed recently by Allinson et al.
[17]. Interestingly, Notch–Delta signalling might be a two-way RIP affair. Although the
signalling activity of the soluble Delta ectodomain is controversial, its membrane-
bound form can activate Notch on an adjacent cell. During such juxtacrine interaction,
Delta seems to be processed in a similar manner to Notch. First, its ectodomain is shed
by an ADAM, followed by g-secretase-mediated release of the ICD, which can be
translocated into the nucleus [79–81]. (b) Transactivation of epidermal growth factor
receptor (EGFR;cyan)byG-protein-coupledreceptor (GPCR; orange)signallingoccurs
via EGFR-ligand ectodomain shedding. ADAMs seem to be the key mediators of the
cross-talk between the two signalling systems [2,15]. Stimulation of GPCR signalling
(orange arrows) by a GPCR agonist (1) leads to the induction of EGFR ligand (green)
release by an ADAM (2, magenta), which in turn leads to EGFR dimerization and
mitogenic signalling (3) upon ligand binding, and, similarly, to ‘direct’ paracrine EGFR
activation via ADAM-mediated EGFR-ligand shedding [13,14]. Autocrine transactiva-
tion is depicted here for simplicity, but paracrine transactivation of EGFR signalling on
nearby cells would occur by the same mechanism. In cancer cells and tumour
xenografts, the juxtacrine activation of EGFR on an adjacent cell by membrane-
anchored pro-transforming growth factor-a was recently shown to require ADAM
sheddase activity [54].
Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 419
www.sciencedirect.com
ADAM-17 cleave APP at the consensus physiologicala-cleavage site (Lys–Leu), whereas ADAM-9 cuts a His–Glnbond two residues upstream, corresponding to an alter-native cleavage site identified in primary hippocampalneurons [17]. However, ADAM-9 is unlikely to be anessential a-secretase because adam9K/K hippocampalneurons show no defect in APP processing [24].
Hartmann et al. [18] showed that ADAM-10-deficientfibroblasts process APP normally, indicating that otherproteinases function as the a-secretase in this cell type.This, however, cannot be extrapolated to the CNS, and theauthors suggested that a-processing-competent ADAMsmight work as teams in a tissue-specific manner [18], andhence another ADAM could deputize for the missingADAM-10. The recent report showing that adam10overexpression prevents amyloid plaque formation andalleviates the neurological deficits in a mouse model ofAlzheimer’s disease provides compelling evidence forADAM-10 as a major a-secretase in the living brain [21].Furthermore, dominant-negative overexpression of cata-lytically inactive ADAM-10 worsens the condition of thesemice, and leads to increased deposition of b-amyloidplaques [21]. These results strongly corroborate the causalrole of b-amyloid in Alzheimer’s disease and the reciprocalrelationship between the actions of a- and b-secretase inamyloid production. Altogether, although ADAM-10 isstrongly indicated as a major physiological a-secretase, itis likely that there is not a single a-secretase but that thephysiological processing of APP is a co-operative action ofseveral activities [17].
Ectodomain shedding is an essential part of Notchactivation, a conserved lateral inhibition system in neuraldevelopment. Again, both ADAM-10 and ADAM-17 arecapable of correct cleavage. Biochemical and geneticevidence has established Kuzbanian (ADAM-10) as theNotch sheddase in Drosophila and Caenorhabditis elegans,whereas the assignment of the relevant mammaliansheddase(s) has been more controversial [3]. The factsthat adam17K/K mice do not phenocopy the notch knock-out defects, whereas Notch signalling is severely dis-rupted in adam10K/K mice, argue strongly for ADAM-10as the major mammalian Notch sheddase [8,18].
Processing of APP and Notch provide excellentexamples of ADAM-mediated shedding in the regulationof gene expression, a process called regulated intramem-brane proteolysis [53]. Shedding can be ligand-induced(Notch) or ligand-independent (APP) (Figure 3a). Subse-quently, proteolytically released cytosolic domain of thetarget protein translocates into the nucleus where its inter-action with transcription factor(s) leads to gene activation.
Emerging roles of ADAM-mediated ectodomain shed-
ding in human disease
GPCR-induced transactivation of EGFR signalling(Figure 3b) has emerged as an important mechanismthat regulates normal cellular responses, but it has alsobeen implicated in pathologies such as cancers [2,7,15,54].As discussed, ADAMs are key regulators of EGFRsignalling via shedding of EGFR ligands. ADAM-10,ADAM-15 and ADAM-17 were recently shown to con-tribute to tumourigenesis, migration and invasion by

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005420
GPCR–EGFR transactivation in different tumours andcell lines [7,15,54].
ADAM-mediated EGFR transactivation is also linkedto cardiac pathophysiology. ADAM-12 was recently shownto play a part in the physiological response of cardiomyo-cytes to elevated blood pressure by EGFR-transactivation,leading to cardiac hypertrophy, which, in turn, can lead tocardiovascular disease [55].
Another recent discovery that ADAM33 is an asthma-susceptibility gene [56] has had a major impact on asthmaand ADAM research. The proteolytic activity of ADAM-33suggests a sheddase function [31] and, thus, might linkADAM-mediated ectodomain shedding to asthma.
Identification of novel ADAM metalloproteinase
substrates
Because all the ADAMs with an intact active-sitesequence that have been studied so far show metallopro-teinase activity [1,2], it is plausible that physiologicalsubstrates also exist for those ADAMs not yet investi-gated. Furthermore, there are several shed membraneproteins, such as angiotensin-converting enzyme, withoutidentified sheddase(s) [52].
Identification of the particular ADAM that cleaves aspecific protein substrate in vivo or in vitro has provedproblematic. Although the ability of a specific ADAM tohydrolyse a small peptide spanning the sequence aroundthe known cleavage site is indicative, it is by no meansconclusive. Other factors also determine the specificity(as discussed), and the kinetics of hydrolysis of suchpseudo-substrates are often several orders of magnitudelower than for the intact protein. All too often, the precisecleavage site is not known and other strategies foridentifying the relevant ADAM(s) are therefore required.
One approach is to use selective inhibitors to dis-tinguish between ADAMs, although, at present, fewcompounds are sufficiently discriminatory. A class ofhydroxamate-based compounds originally identified asMMP inhibitors, such as batimastat and marimastat,have conventionally been used to inhibit ADAM protein-ases. The compound TAPI (TNFa proteinase inhibitor)was originally shown to block shedding of the TNFa andits receptor, and has commonly been used as an ADAM-17inhibitor, although it is, in fact, a rather broad-spectruminhibitor. Also, more selective inhibitors are sometimesused at concentrations beyond their range of selectivity.The most exhaustive comparative study of hydroxamate-based compounds as shedding inhibitors was reported byParkin et al. [23]. The compound GI254023X is reported tobe a selective ADAM-10 inhibitor and has been used toidentify the constitutive sheddase for fractalkine asADAM-10 [57]. Unfortunately, most of the inhibitorstudies have considered only ADAM-10 and ADAM-17,and a thorough comparison of the potency of the variousinhibitors against each of the purified ADAMs is notavailable.
Given the similarity of their active sites, it is unlikelythat a panel of small molecule, potent and highly selectiveinhibitors of each of the ADAMs will emerge in the nearfuture. Alternative approaches to identify physiologicallyrelevant sheddases have relied on increased shedding
www.sciencedirect.com
upon overexpression of specific ADAM genes or, alter-natively, depleting the ADAM using siRNA or geneknockout. Given the evident redundancy of the ADAMs,even these approaches can rarely give an unequivocalanswer, and a one-to-one relationship between sheddaseand substrate is probably an unusual event. Finally,a definitive requirement for a shedding event is theencounter of the sheddase and the substrate, emphasizingthe importance of the localization studies in vivo.
Concluding remarks
Although gene knockout experiments have shown thatparticular ADAMs are indispensable during mammaliandevelopment, functional redundancy among at least asubset of ADAMs is increasingly evident. Besides, alter-native splicing is emerging as a mechanism by whichfunctional complexity can be increased. This undoubtedlycomplicates both identification of the shedding-machinerycomponents and the development of specific inhibitorsagainst particular shedding events. Nevertheless, recentadvances towards the elucidation of the control mechan-isms that regulate ADAM-mediated shedding and identi-fication of key players in important events, such as APPprocessing, have set the stage for more comprehensivephysiological understanding of this challenging field.
Note added in proof
The recent study by Bohm et al. [83] shows that thedevelopment of osteoarthritic lesions is accelerated inaging adam15K/K mice.
AcknowledgementsWe apologize to colleagues whose original work could not be cited owing tospace limitations. This work was supported by the INTAS grant 2001–245(A.J.T. and A-P.J.H.) and grants from the Medical Research Council ofGreat Britain (A.J.T.), the Wellcome Trust (A.J.T.), British HeartFoundation (A.J.T.), and the Medical Research Fund of TampereUniversity Hospital (A-P.J.H. and M.P.).
References
1 White, J.M. (2003) ADAMs: modulators of cell–cell and cell–matrixinteractions. Curr. Opin. Cell Biol. 15, 598–606
2 Blobel, C.P. (2005) ADAMs: key components in EGFR signalling anddevelopment. Nat. Rev. Mol. Cell Biol. 6, 32–43
3 Seals, D.F. and Courtneidge, S.A. (2003) The ADAMs family ofmetalloproteases: multidomain proteins with multiple functions.Genes Dev. 17, 7–30
4 Kheradmand, F. andWerb, Z. (2002) Shedding light on sheddases: rolein growth and development. Bioessays 24, 8–12
5 Black, R.A. et al. (2003) Substrate specificity and inducibility of TACE(tumour necrosis factor a-converting enzyme) revisited: the Ala-Valpreference, and induced intrinsic activity. Biochem. Soc. Symp. 70,39–52
6 Becherer, J.D. and Blobel, C.P. (2003) Biochemical properties andfunctions of membrane-anchored metalloprotease-disintegrin pro-teins (ADAMs). Curr. Top. Dev. Biol. 54, 101–123
7 Moss, M.L. and Bartsch, J.W. (2004) Therapeutic benefits fromtargeting of ADAM family members. Biochemistry 43, 7227–7235
8 Peschon, J.J. et al. (1998) An essential role for ectodomain shedding inmammalian development. Science 282, 1281–1284
9 Zhao, J. et al. (2001) Pulmonary hypoplasia in mice lacking tumornecrosis factor-a converting enzyme indicates an indispensable rolefor cell surface protein shedding during embryonic lung branchingmorphogenesis. Dev. Biol. 232, 204–218

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 421
10 Jackson, L.F. et al. (2003) Defective valvulogenesis in HB-EGF andTACE-null mice is associated with aberrant BMP signaling. EMBO J.22, 2704–2716
11 Shi, W. et al. (2003) TACE is required for fetal murine cardiacdevelopment and modeling. Dev. Biol. 261, 371–380
12 Mohammed, F.F. et al. (2004) Abnormal TNF activity in Timp3K/K
mice leads to chronic hepatic inflammation and failure of liverregeneration. Nat. Genet. 36, 969–977
13 Sahin, U. et al. (2004) Distinct roles for ADAM10 and ADAM17 inectodomain shedding of six EGFR ligands. J. Cell Biol. 164, 769–779
14 Hinkle, C.L. et al. (2004) Selective roles for tumor necrosis factora-converting enzyme/ADAM17 in the shedding of the epidermalgrowth factor receptor ligand family: the juxtamembrane stalkdetermines cleavage efficiency. J. Biol. Chem. 279, 24179–24188
15 Schafer, B. et al. (2004) Distinct ADAM metalloproteinases regulateG protein-coupled receptor-induced cell proliferation and survival.J. Biol. Chem. 279, 47929–47938
16 Zhang, Y. et al. (2000) Tumor necrosis factor-alpha converting enzyme(TACE) is a growth hormone binding protein (GHBP) sheddase: themetalloprotease TACE/ADAM-17 is critical for (PMA-induced) GHreceptor proteolysis and GHBP generation. Endocrinology 141,4342–4348
17 Allinson, T.M. et al. (2003) ADAMs family members as amyloidprecursor protein alpha-secretases. J. Neurosci. Res. 74, 342–352
18 Hartmann, D. et al. (2002) The disintegrin/metalloprotease ADAM 10is essential for Notch signalling but not for a-secretase activity infibroblasts. Hum. Mol. Genet. 11, 2615–2624
19 Hattori, M. et al. (2000) Regulated cleavage of a contact-mediatedaxon repellent. Science 289, 1360–1365
20 Schimmelpfeng, K. et al. (2001) The function of leak and kuzbanianduring growth cone and cell migration. Mech. Dev. 106, 25–36
21 Postina, R. et al. (2004) A disintegrin-metalloproteinase preventsamyloid plaque formation and hippocampal defects in an Alzheimerdisease mouse model. J. Clin. Invest. 113, 1456–1464
22 Checler, F. and Vincent, B. (2002) Alzheimer’s and prion diseases:distinct pathologies, common proteolytic denominators. TrendsNeurosci. 25, 616–620
23 Parkin, E.T. et al. (2004) Dual mechanisms for shedding of the cellularprion protein. J. Biol. Chem. 279, 11170–11178
24 Weskamp, G. et al. (2002) Mice lacking the metalloprotease-disintegrin MDC9 (ADAM9) have no evident major abnormalitiesduring development or adult life. Mol. Cell. Biol. 22, 1537–1544
25 Kurisaki, T. et al. (2003) Phenotypic analysis of Meltrin a (ADAM12)-deficient mice: involvement of Meltrin a in adipogenesis andmyogenesis. Mol. Cell. Biol. 23, 55–61
26 Horiuchi, K. et al. (2003) Potential role for ADAM15 in pathologicalneovascularization in mice. Mol. Cell. Biol. 23, 5614–5624
27 Kelly, K. et al. (2005) Metalloprotease-disintegrin ADAM8: expressionanalysis and targeted deletion in mice. Dev. Dyn. 232, 221–231
28 Moghadaszadeh, B. et al. (2003) Compensation for dystrophin-deficiency: ADAM12 overexpression in skeletal muscle results inincreased a7 integrin, utrophin and associated glycoproteins. Hum.
Mol. Genet. 12, 2467–247929 Kurohara, K. et al. (2004) Essential roles of Meltrin b (ADAM19) in
heart development. Dev. Biol. 267, 14–2830 Zhou, H.M. et al. (2004) Essential role for ADAM19 in cardiovascular
morphogenesis. Mol. Cell. Biol. 24, 96–10431 Zou, J. et al. (2004) Catalytic activity of human ADAM33. J. Biol.
Chem. 279, 9818–983032 Zheng, Y. et al. (2004) Evaluation of the contribution of different
ADAMs to tumor necrosis factor a (TNFa) shedding and of thefunction of the TNFa ectodomain in ensuring selective stimulatedshedding by the TNFa convertase (TACE/ADAM17). J. Biol. Chem.
279, 42898–4290633 Hurtado, O. et al. (2002) TACE/ADAM17-TNF-a pathway in rat
cortical cultures after exposure to oxygen-glucose deprivation orglutamate. J. Cereb. Blood Flow Metab. 22, 576–585
34 Ortiz, R.M. et al. (2004) Aberrant alternative exon use and increasedcopy number of human metalloprotease-disintegrin ADAM15 gene inbreast cancer cells. Genes Chromosomes Cancer 41, 366–378
35 Amour, A. et al. (2002) The enzymatic activity of ADAM8 and ADAM9is not regulated by TIMPs. FEBS Lett. 524, 154–158
www.sciencedirect.com
36 Yan, Y. et al. (2002) The metalloprotease Kuzbanian (ADAM10)mediates the transactivation of EGF receptor by G protein-coupledreceptors. J. Cell Biol. 158, 221–226
37 Shimizu, E. et al. (2003) Structure and expression of the murineADAM 15 gene and its splice variants, and difference of interactionbetween their cytoplasmic domains and Src family proteins. Biochem.Biophys. Res. Commun. 309, 779–785
38 Wakatsuki, S. et al. (2004) Lipid rafts identified as locations ofectodomain shedding mediated by Meltrin b/ADAM19. J. Neurochem.89, 119–123
39 von Tresckow, B. et al. (2004) Depletion of cellular cholesterol and lipidrafts increases shedding of CD30. J. Immunol. 172, 4324–4331
40 Matthews, V. et al. (2003) Cellular cholesterol depletion triggersshedding of the human interleukin-6 receptor by ADAM10 andADAM17 (TACE). J. Biol. Chem. 278, 38829–38839
41 Ehehalt, R. et al. (2003) Amyloidogenic processing of the Alzheimerb-amyloid precursor protein depends on lipid rafts. J. Cell Biol. 160,113–123
42 Cordy, J.M. et al. (2003) Exclusively targeting b-secretase to lipid raftsby GPI-anchor addition up-regulates b-site processing of the amyloidprecursor protein. Proc. Natl. Acad. Sci. U. S. A. 100, 11735–11740
43 Zhao, L. et al. (2001) Regulation of membrane metalloproteolyticcleavage of L-selectin (CD62l) by the epidermal growth factor domain.J. Biol. Chem. 276, 30631–30640
44 Pang, S. et al. (2001) Roles of the juxtamembrane and extracellulardomains of angiotensin-converting enzyme in ectodomain shedding.Biochem. J. 358, 185–192
45 Endres, K. et al. (2003) Tumor necrosis factor-alpha convertingenzyme is processed by proprotein-convertases to its mature formwhich is degraded upon phorbol ester stimulation. Eur. J. Biochem.270, 2386–2393
46 Diaz-Rodriguez, E. et al. (2002) Extracellular signal-regulated kinasephosphorylates tumor necrosis factor a-converting enzyme at threo-nine 735: a potential role in regulated shedding. Mol. Biol. Cell 13,2031–2044
47 Fan, H. et al. (2003) Characterization of growth factor-inducedserine phosphorylation of tumor necrosis factor-a convertingenzyme and of an alternatively translated polypeptide. J. Biol.Chem. 278, 18617–18627
48 Weskamp, G. et al. (2004) Evidence for a critical role of the tumornecrosis factor a convertase (TACE) in ectodomain shedding of the p75neurotrophin receptor (p75NTR). J. Biol. Chem. 279, 4241–4249
49 Canet-Aviles, R.M. et al. (2002) Muscarine enhances soluble amyloidprecursor protein secretion in human neuroblastoma SH-SY5Y by apathway dependent on protein kinase Ca, src-tyrosine kinase andextracellular signal-regulated kinase but not phospholipase C. BrainRes. Mol. Brain Res. 102, 62–72
50 Karkkainen, I. et al. (2000) Metalloprotease-disintegrin (ADAM)genes are widely and differentially expressed in the adult CNS. Mol.Cell. Neurosci. 15, 547–560
51 Asai, M. et al. (2003) Putative function of ADAM9, ADAM10, andADAM17 as APP a-secretase. Biochem. Biophys. Res. Commun. 301,231–235
52 Allinson, T.M. et al. (2004) The role of ADAM10 and ADAM17 in theectodomain shedding of angiotensin converting enzyme and theamyloid precursor protein. Eur. J. Biochem. 271, 2539–2547
53 Selkoe, D. and Kopan, R. (2003) Notch and Presenilin: regulatedintramembrane proteolysis links development and degeneration.Annu. Rev. Neurosci. 26, 565–597
54 Borrell-Pages, M. et al. (2003) TACE is required for the activation ofthe EGFR by TGF-a in tumors. EMBO J. 22, 1114–1124
55 Asakura, M. et al. (2002) Cardiac hypertrophy is inhibited byantagonism of ADAM12 processing of HB-EGF: metalloproteinaseinhibitors as a new therapy. Nat. Med. 8, 35–40
56 Van Eerdewegh, P. et al. (2002) Association of the ADAM33 gene withasthma and bronchial hyperresponsiveness. Nature 418, 426–430
57 Hundhausen, C. et al. (2003) The disintegrin-like metalloproteinaseADAM10 is involved in constitutive cleavage of CX3CL1 (frac-talkine) and regulates CX3CL1-mediated cell–cell adhesion. Blood102, 1186–1195
58 Garton, K.J. et al. (2001) Tumor necrosis factor-a-converting enzyme(ADAM17) mediates the cleavage and shedding of fractalkine(CX3CL1). J. Biol. Chem. 276, 37993–38001

Review TRENDS in Biochemical Sciences Vol.30 No.7 July 2005422
59 Abel, S. et al. (2004) The transmembrane CXC-chemokine ligand 16 isinduced by IFN-g and TNF-a and shed by the activity of thedisintegrin-like metalloproteinase ADAM10. J. Immunol. 172,6362–6372
60 Gough, P.J. et al. (2004) A disintegrin andmetalloproteinase 10-mediatedcleavage and shedding regulates the cell surface expression of CXCchemokine ligand 16. J. Immunol. 172, 3678–3685
61 Montero, J.C. et al. (2000) Differential shedding of transmembraneneuregulin isoforms by the tumor necrosis factor-a-convertingenzyme. Mol. Cell. Neurosci. 16, 631–648
62 Fourie, A.M. et al. (2003) Catalytic activity of ADAM8, ADAM15, andMDC-L (ADAM28) on synthetic peptide substrates and in ectodomaincleavage of CD23. J. Biol. Chem. 278, 30469–30477
63 Contin, C. et al. (2003) Membrane-anchored CD40 is processed by thetumor necrosis factor-a-converting enzyme. Implications for CD40signaling. J. Biol. Chem. 278, 32801–32809
64 Cruz, A.C. et al. (2004) Tumor necrosis factor-a-converting enzymecontrols surface expression of c-Kit and survival of embryonic stemcell-derived mast cells. J. Biol. Chem. 279, 5612–5620
65 Rio, C. et al. (2000) Tumor necrosis factor-a-converting enzyme isrequired for cleavage of erbB4/HER4. J. Biol. Chem. 275, 10379–10387
66 Marin, V. et al. (2002) Chemotactic agents induce IL-6Ra sheddingfrom polymorphonuclear cells: involvement of a metalloproteinase ofthe TNF-a-converting enzyme (TACE) type. Eur. J. Immunol. 32,2965–2970
67 Budagian, V. et al. (2004) Natural soluble IL-15Ra is generated bycleavage that involves the tumor necrosis factor-a-converting enzyme(TACE/ADAM17). J. Biol. Chem. 279, 40368–40375
68 Rovida, E. et al. (2001) TNF-a-converting enzyme cleaves themacrophage colony-stimulating factor receptor in macrophagesundergoing activation. J. Immunol. 166, 1583–1589
69 Murai, T. et al. (2004) Engagement of CD44 promotes Rac activationand CD44 cleavage during tumor cell migration. J. Biol. Chem. 279,4541–4550
70 Nagano, O. et al. (2004) Cell-matrix interaction via CD44 isindependently regulated by different metalloproteinases activated inresponse to extracellular Ca2C influx and PKC activation. J. Cell Biol.165, 893–902
71 Naus, S. et al. (2004) Ectodomain shedding of the neural recognition
Elsevier joins major health
Elsevier has joined with scientific publishers and leading voluntary h
initiative to help patients and caregivers close a crucial information g
disseminating medical research and is scheduled to launch in 2005.
Elsevier will provide the voluntary health organizations with increase
immediately upon publication, together with content from back issues
into materials for patients and link to the full text of selected researc
patientINFORM has been created to allow patients seeking the la
most up-to-date, reliable research available for specific diseases.
‘Not only will patientINFORM connect patients and their caregivers w
making it easier to understand research findings, patientINFORM will
physicians and make well-informed decisions about care’, said Harmo
Society.
For more information, visit
www.sciencedirect.com
molecule CHL1 by the metalloprotease-disintegrin ADAM8 promotesneurite outgrowth and suppresses neuronal cell death. J. Biol. Chem.279, 16083–16090
72 Gutwein, P. et al. (2003) ADAM10-mediated cleavage of L1 adhesionmolecule at the cell surface and in released membrane vesicles.FASEB J. 17, 292–294
73 Gomez-Gaviro, M.V. et al. (2002) Structure–function relationship androle of tumor necrosis factor-a-converting enzyme in the down-regulation of L-selectin by non-steroidal anti-inflammatory drugs.J. Biol. Chem. 277, 38212–38221
74 Garton, K.J. et al. (2003) Stimulated shedding of vascular celladhesion molecule 1 (VCAM-1) is mediated by tumor necrosis factor-a-converting enzyme (ADAM 17). J. Biol. Chem. 278, 37459–37464
75 Hussain, I. et al. (2003) Characterization of the ectodomain sheddingof the b-site amyloid precursor protein-cleaving enzyme 1 (BACE1).J. Biol. Chem. 278, 36264–36268
76 Hahn, D. et al. (2003) Phorbol 12-myristate 13-acetate-inducedectodomain shedding and phosphorylation of the human meprinbetametalloprotease. J. Biol. Chem. 278, 42829–42839
77 Ito, N. et al. (2004) ADAMs, a disintegrin and metalloproteinases,mediate shedding of oxytocinase. Biochem. Biophys. Res. Commun.314, 1008–1013
78 Franzke, C.W. et al. (2002) Transmembrane collagen XVII, anepithelial adhesion protein, is shed from the cell surface by ADAMs.EMBO J. 21, 5026–5035
79 Bland, C.E. et al. (2003) Notch-induced proteolysis and nuclearlocalization of the Delta ligand. J. Biol. Chem. 278, 13607–13610
80 Six, E. et al. (2003) The Notch ligand Delta1 is sequentially cleaved byan ADAM protease and g-secretase. Proc. Natl. Acad. Sci. U. S. A. 100,7638–7643
81 LaVoie, M.J. and Selkoe, D.J. (2003) The Notch ligands, Jagged andDelta, are sequentially processed by a-secretase and presenilin/g-secretase and release signaling fragments. J. Biol. Chem. 278,34427–34437
82 Thathiah, A. et al. (2003) Tumor necrosis factor-a convertingenzyme/ADAM 17 mediates MUC1 shedding. J. Biol. Chem. 278,3386–3394
83 Bohm, B.B. et al. (2005) Homeostatic effects of the metalloproteinasedisintegrin ADAM15 in degenerative cartilage remodeling. ArthritisRheum. 52, 1100–1109
information initiative
ealth organizations to create patientINFORM, a groundbreaking
ap. patientINFORM is a free online service dedicated to
d online access to our peer-reviewed biomedical journals
. The voluntary health organizations will integrate the information
h articles on their websites.
test information about treatment options online access to the
ith the latest research, it will help them to put it into context. By
empower patients to have a more productive dialogue with their
n Eyre, M.D., national chief medical officer of the American Cancer
www.patientinform.org




















