
Embryogenesis, Histology, and Organology of the Ovary of ...
Upload
independentCategory
view
2download
0
ARTICLE IN PRESS
Differentiation 79 (2010) 159–170
Contents lists available at ScienceDirect
Differentiation
0301-46
Join the
doi:10.1
Abbre$Joinn Corr
E-m
journal homepage: www.elsevier.com/locate/diff
Mini-Series on Germ Cell Development
Differentiation potential of germ line stem cells derived fromthe postnatal mouse ovary$
Jason Pacchiarotti a, Chad Maki a, Thomas Ramos a, Joel Marh a, Kyle Howerton a,Jadelind Wong a, Jane Pham a, Sandra Anorve a, Yung-Chiong Chow b, Fariborz Izadyar a,n
a PrimeGen Biotech, 213 Technology Drive, Irvine, CA 92618, USAb Mackay Memorial Hospital, Department of Urology, Taipei, Taiwan
a r t i c l e i n f o
Article history:
Received 16 July 2009
Received in revised form
10 December 2009
Accepted 7 January 2010
Keywords:
Stem cell
Oogenesis
Follicle
Engineering
Tissue
Regeneration
81/$ - see front matter & 2010 International
International Society for Differentiation (ww
016/j.diff.2010.01.001
viations: OGSCs, ovarian germ line stem cells
the International Society for Differentiation
esponding author: Tel: +1 949 428 0511; fax
ail address: [email protected] (F
a b s t r a c t
General belief in reproductive biology is that in most mammals female germ line stem cells are
differentiated to primary oocytes during fetal development and oogenesis starts from a pool of
primordial follicles after birth. This idea has been challenged previously by using follicle kinetics studies
and demonstration of mitotically active germ cells in the postnatal mouse ovary (Johnson et al., 2004;
Kerr et al., 2006; Zhang et al., 2008). However, the existence of a population of self-renewing ovarian
germ line stem cells in postnatal mammals is still controversial (Eggan et al., 2006; Telfer et al., 2005;
Gosden, 2004). Recently, production of offspring from a germ line stem cell line derived from the
neonatal mouse ovary was reported (Zou et al., 2009). This report strongly supports the existence of
germ line stem cells and their ability to expand in vitro. Recently, using a transgenic mouse model in
which GFP is expressed under a germ cell-specific Oct-4 promoter, we isolated and generated
multipotent cell lines from male germ line stem cells (Izadyar et al., 2008). Using the same strategy we
isolated and derived cell lines from postnatal mouse ovary. Interestingly, ovarian germ line stem cells
expanded in the same culture conditions as the male suggesting that they have similar requirements for
their self-renewal. After 1 year of culture and many passages, ovarian germ line stem cells maintained
their characteristics and telomerase activity, expressed germ cell and stem cell markers and revealed
normal karyotype. As standard protocol for differentiation induction, these cells were aggregated and
their ability to form embryoid bodies (EBs) was investigated. EBs generated in the presence of growth
factors showed classical morphology and expressed specific markers for three germ layers. However, in
the absence of growth promoting factors EBs were smaller and large cells with the morphological and
molecular characteristics of oocytes were formed. This study shows the existence of a population of
germ line stem cell in postnatal mouse ovary with multipotent characteristics.
& 2010 International Society of Differentiation. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Germ-line stem cells are responsible for gametogenesis andtransmission of genetic information to the next generation. Inmale mammals, distinct populations of stem cells known asspermatogonial stem cells maintain spermatogenesis via the self-renewal and generation of spermatogonia destined to differenti-ate into sperm. In the female however, there has been alongstanding dogma that mammalian females produce a finitenumber of oocytes in the prenatal phase of life, and thisproduction ceases after birth (Bristol-Gould et al., 2006; Huttand Albertini, 2006). Recently, emergent data have come to refute
Society of Differentiation. Publish
w.isdifferentiation.org)
(www.isdifferentiation.org).
: +1 949 271 2398
. Izadyar).
the dogma and support postnatal development of oocytes andfollicles in female mouse ovaries (Johnson et al., 2004; Kerr et al.,2006). These studies, by characterizing the kinetics of thedecreasing number of follicles through the life concluded thatwithout postnatal production of follicles from germ line stem cellsthe mouse would not reach its expected reproductive lifespan(Johnson et al., 2004). Another study using immunolabelling ofgerm cell nuclear antigen (GCNA) and proliferating cell nuclearantigen (PCNA) revealed that there is a postnatal renewal offollicles in adult mouse ovaries (Kerr et al., 2006). Although thesestudies strongly indicate de novo oogenesis in the mouse ovary,they were unsuccessful to conclusively prove the existence ofgerm line stem cells. The strongest evidence so far for theexistence of a germ line stem cell pool in the ovary is from Zouet al. (2009) in which using immunomagnetic cell sorting by germcell marker VASA they generated cell lines from neonatal mouseovary with the ability to regenerate oogenesis and produce
ed by Elsevier Ltd. All rights reserved.

ARTICLE IN PRESS
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170160
offspring. However, VASA is a general germ cell marker (Toyookaet al., 2000; Castrillon et al., 2000) and does not specifically enrichgerm line stem cells and could not be used for identification andcharacterization of germ line stem cells in the ovary.
In the present study to unambiguously identify and track germline stem cells, we utilized a transgenic mouse model expressinggreen fluorescent protein (GFP) under the control of a germline-specific Pou5f1 (Oct-4) promoter (Yeom et al., 1996). POU5F1(previously known as octamer-binding transcription factor 3
4,Oct-4) was originally identified as an ES cell and germline-specific marker (Okamota et al., 1990; Scholer et al., 1990).At gastrulation, Pou5f1 expression is down-regulated and there-after is maintained only in primordial germ cells (Yeom et al.,1996). PGCs, of both males and females, continue to expressPou5f1 as they proliferate and migrate to genital ridges. In themales, the expression in germ cells persists throughout fetaldevelopment and is maintained postnatally in proliferatinggonocytes, prospermatogonia and undifferentiated spermatogo-nia. In female germ cells however, Pou5f1 is expressed ingonocytes and undifferentiated oogonia and undergoes downregulation during oogenesis, coincident with entry into meiosisand is up-regulated in the final stage of meiotic prophase I (Pesceet al., 1998; Tadokora et al., 2002). Therefore, in this mouse modeldifferent sizes of GFP positive cells could be visualized duringdifferent stages of oogenesis. Immunohistochemical staining withspecific germ cell and stem cell markers was used to characterizea small population of GFP-Oct-4 cells as being ovarian germ linestem cells.
In addition, based on the dogma, there should not be anyoogonia present in the postnatal mouse ovary and oogenesisresumes from a pool of primary oocytes enclosed in primordialfollicles. As the chromosome content of oogonia (2n) is half of theprimary oocytes (4n), we used this as another parameter in thisstudy to track the presence of germ line stem cell in the ovary.Furthermore, using flow cytometry, we successfully isolated GFP-Oct-4 positive cells and have been able to expand them in culturefor more than 1 year. Cultured ovarian germ line stem cells(OGSCs) were further characterized for their marker expression,telomerase activity and karyotype normalcy. Finally, differentia-tion potential of OGSC were analyzed using traditional embryoidbody formation and their ability to form more advanced germcells similar to oocyte as well as their multipotentiality fordifferentiation to other lineages was investigated.
2. Materials and methods
2.1. Isolation and purification of germ-line stem cells
For cell isolation, ovaries from 40 to 60 OG2 pups between 2and 5 days old or one adult mouse were dissected under a micro-dissection microscope. Ovaries were first collected in a Petri dishcontaining cold DPBS (Cellgro, Herndon, VA, USA) supplementedwith 5 mM EDTA (Sigma, St. Louis, MO, USA). Using a 5 ml pipette,ovaries were then transferred to a 50 ml conical tube. Aftercentrifugation DPBS was removed and ovaries were re-suspendedin Collagenase (1 mg/mL) (Roche, Indianapolis, IN, USA) andDNase-I (10 U/mL) (Invitrogen, Carlsbad, CA, USA) and placed in a37 1C water bath for 30 min. Every 10 min ovaries were broken tosmaller pieces by pipette and at the end 10% FBS (Hyclone,Invitrogen, Logan, UT, USA) was added to inactivate the enzyme.Cells were then passed through a 40 mm strainer (Milipore,Billerica, MA, USA) and were centrifuged at 400g for 10 min,supernatant was removed and cells were re-suspended in culturemedium and kept on ice. Ovarian germ-line stem cells as detectedby GFP were sorted using fluorescent-assisted cell sorting (FACS).
2.2. Culture of ovarian germ line stem cells
Enriched GFP positive germ line cells were cultured on mouseembryonic fibroblast (MEF) feeder in a culture medium previouslyused for expansion of multipotent germ line stem cells (Okamotaet al., 1990) in a concentration of 5000–10,000 per well of a four-well plate (Nunc, Pittsburgh, PA, USA). After a week, smallcolonies formed and these colonies were manually split for 4–5passages before trypsinization. For trypsinization, contents of afour-well plate (10–12 big colonies) were manually picked in aneppendorf tube and 0.25% trypsin (Gibco, Gaithersburg, MD, USA)was added for 2 min. After repeated agitation trypsin wasinactivated by adding FBS (10%). Cells were centrifuged at 400g
for 5 min and re-suspended in above-mentioned culture medium.Culture was maintained at 37 1C in a Forma Series 2, 5% CO2
humidified incubator (Thermo Electron Corporation, Waltham,MA, USA). Half of the medium was changed every other day andevery 2 weeks cells were transferred to a new MEF plate.
2.3. Characterization of ovarian germ-line stem cells
Mouse ovaries were dissected and fixed in 4% paraformalde-hyde (PFA) (Electron Microscopy Science, Hatfield, PA, USA)overnight at 4 1C and localization of GFP positive cells weredetermined using fluorescent microscopy. In addition, someovaries were transferred into 20% cold sucrose, embedded andfrozen in OCT compound (VWR, West Chester, PA, USA) and keptat �80 1C. The 8–10 mm sections were prepared and localizationof germ line stem cells in ovarian sections was confirmed usingOct-4 and VASA immunolabelling. Freshly isolated GFP positivecells were collected for telomerase determination and geneexpression profiling. Cultured ovarian germ line stem cells werealso collected for telomerase activity and gene analysis. Inaddition some cultured cells were fixed in 2% PFA for 30 min atroom temperature washed in PBS and kept at 4 1C. To characterizecultured ovarian germ line stem cells, GCNA, c-Kit, Oct-4,Nanog and GFR-a1 staining was performed using fluorescentimmunohistochemistry.
2.4. Immunohistochemical staining
For fluorescent immunocytochemistry, cells were permea-blized with 1� Cytoperm (BD Biosciences, San Jose, CA, USA) or0.2% Triton X-100 (Sigma) for 15 min and subsequently incubatedin 2% (w/v) bovine serum albumin (BSA) (Sigma), 2% (v/v) normalgoat serum (Cedarlane Laboratories Limited, Burlington, Ontario,Canada)/1� Cytoperm–PBS for 30–60 min both at room tem-perature. Primary antibody was either diluted at the optimalconcentration in 2%BSA 2% goat serum/1� Cytoperm–PBS andincubated for 3 h at 4 1C, or diluted in blocking buffer overnight at4 1C. After two washes, fluorescent secondary antibody wasdiluted accordingly in 2%BSA 2% goat serum/1� Cytoperm–PBSand incubated for 1 h at 4 1C in the dark. Cells were washed twicein PBS, wrapped in foil and stored at 4 1C until microscopicanalysis. Images were recorded using an Olympus BX61 micro-scope with SlideBookTM imaging. For negative controls, theprimary antibody was omitted. The primary antibodies used inthis study were acquired from various companies. GCNA antibodywas kindly provided by Dr. Enders, University of Kansas MedicalCenter. Each antibody was validated and the concentrationsoptimized in our laboratory. The source and working dilutions ofthese antibodies are presented in Table 1.

ARTICLE IN PRESS
Table 1Antibodies and reagents used for immunohistochemical localization and flow cytometry analysis of ovarian germ line stem cells.
Name of antigen/reagent Source Working dilution Method
DDX4 (VASA) Abcam 1:200 ICC
SSEA-1 Chemicon 1:200 ICC
CD117 Santa Cruz Biotechnology 1:100 ICC
CD117 BD Pharmingen 1:100 Flow cytometry
SSEA-1 eBioscience 1:200 Flow cytometry
GFR-a1 R&D Systems 1:100 Flow cytometry ICC
Oct-4 Santa Cruz 1:50 ICC
Nanog Bethyl 1:100 ICC
PNA-FITC Vector 1:100 ICC
Phalloidin-Alexa 568 Invitrogen 1:40 ICC
GCNA Provided by Dr. Enders 1:20 ICC
CD133-PE eBiosciences 1:100 Flow cytometry
Nestin Chemicon 1:100 ICC
Troponin abcam 1:100 ICC
GATA-4 Chemicon 1:100 ICC
Alexa 488 Invitrogen 1:500 ICC
Alexa 568 Invitrogen 1:500 ICC
FITC Jackson 1:200 Flow cytometry
DAPI Invitrogen 1:10,000 ICC
TO-PRO-3 Invitrogen 1:100 Flow cytometry
Hoechst 33342 Invitrogen 1:1000 Flow cytometry
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170 161
2.5. Flow cytometry
For c-Kit sorting, freshly isolated testicular cells containing theOct-4-GFP construct were stained with CD117 APC (BD Pharmin-gen, San Diego, CA, USA). GFP excitation was attained with 488 nmsolid state Coherent laser (Coherent, Santa Clara, CA, USA) andemission light collected through a 530-40 dichroic mirror. APCexcitation was attained with 638 nm solid state Coherent laser(Coherent) and emission light collected through a 710-40 dichroicmirror. Cells stained at final dilution of 1:200 c-Kit APC incomplete culture medium for 45 min on ice in the dark. Afterwashing twice in fresh cold complete medium, cells wereconcentrated at 1�107 cells/ml and kept on ice for sorting. Tosort for GFP/c-Kit+/� cells, first we gated the whole GFPpopulation with the non-stained cells to establish a base line forAPC channel. A region was placed based on the base line todiscriminate between negative and positive c-Kit stained cells. Anycells/events falling into this region was considered to be positivelystained for c-Kit and was sorted away from the negative c-Kit cells.
2.6. Determination of ploidy of GFP-Oct-4 cells
GFP-Oct-4 cells were collected from 2 to 10 transgenic mouseovaries and cells were used for DNA content determination. ForDNA analysis, cells were washed and re-suspended in 200 mL coldPBS. In a 12�75 mm round-bottom tube 4 ml of ice cold 70%ethanol was added and cells were fixed overnight at �20 1C. Forstaining, cells were washed in cold PBS and re-suspended in PBScontaining 5 mg/ml PI at a final cell density of 0.5�106 cells/mland were incubated at 37 1C for 30 min prior to flow cytometryanalysis.
2.7. RT-PCR analysis
Total cellular RNA was isolated using RNeasy mini kit (QiagenInc., Valencia, CA, USA) according to the manufacturer’s recom-mendations. The isolated RNA was then transcribed to cDNA usingthe Quantitect RT kit (Qiagen Inc.) and later purified with theQIAquick PCR purification kit (Qiagen Inc.). All primers exceptSCP3 used in this study were designed and purchased fromQiagen. Primer sets for SCP3 were 50-GTGGAAGAAAG-CATTCTGGG-30, and 50-TTTGCCATCTCTTGCTGCTG-30. For each
RT-PCR reaction, 20 ng of cDNA template was used in a 25 mlreaction volume with HotStar Taq Plus (Qiagen Inc.) andrespective primers. All targets were amplified for 30 cycles.Amplification products were identified by size on a 2% agarose gel.
2.8. Telomerase assay and karayotyping
The methods used to evaluate telomerase activity havebeen previously described (Hou et al., 2001). Briefly, cells werere-suspended in CHAPS buffer (Chemicon, Temecula, CA, USA)supplemented with 0.4 units RnaseOut Inhibitor (Invitrogen)/mlCHAPS buffer. Cells were re-suspended at 1000 cells/ml CHAPSbuffer and then incubated on ice for 30 min. Cell lysates werecleared by centrifugation at 4 1C and at 16,000g for 10 min. Proteinconcentration was assayed with Bradford reagent using BSA as astandard. The total volume of each reaction mixture was 25 ml andcontained 250 mg protein, 12.5 ml Quantitect SYBR Green PCRMaster Mix, 1 mg TS primer, 0.5 mg ACX primer, and water to thefinal 25 ml volume. In the sample set, ESC lysate was used as thepositive control and two negative controls were used, a no templatecontrol, and a heat inactivated telomerase ESC sample. The rtq-trapreaction was initiated with a single 20 min incubation at 25 1C.Following this step, the samples were incubated for 15 min at 95 1Cand then subjected to a two step 40-cycle amplification (30 s at95 1C and 90 s at 60 1C). The PCR was performed on the Bio-Rad iQ5and fluorescent signals were captured and analyzed using theBio-Rad iQ5 Optical System Software. Karyotyping was performedat Coriell Cell Repositories, Cytogenetics Laboratory. For karyotyp-ing, proliferating cells were incubated in culture with 0.1 mg/mlKaryoMAX Colcemid (Invitrogen) for 3–4 h before they werere-suspended in hypotonic solution (0.075 M KCl) and incubatedat room temperature for 10 min. Cells were then re-suspended incold fixative (3:1 methanol:acetic acid) and stored at 4 1C for atleast 30 min. Following washing with fixative, cells were applied toclean glass slides and air dried. Metaphase chromosomes wereprepared and karyotypes created using an Applied Spectral ImagingBand View digital imaging system.
2.9. EB formation and in vitro differentiation
Ovarian germ line stem cell colonies were dissociated fromculture by trypsinization and re-suspended in PM-1 medium

ARTICLE IN PRESS
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170162
without growth factors and b-mercaptoethanol in ultra-lowadhesive culture dishes. Culture was performed at 37 1C for 11days and at day 0, 2, 5 and 11 following morphologicalexamination samples were taken for RNA extraction and RT-PCRanalysis. In addition, spontaneous differentiation of the EBs tothree germ layers was confirmed using immunohistochemicallocalization of Nestin (ectoderm), GATA-4 (endoderm) andTroponin (mesoderm) markers.
2.10. In vitro production of primordial follicle-like structures
To further support development of oocyte-like structures,these cells were aggregated with immature granulose cells fromneonatal pups. This method has been shown to be effective information of new primordial follicles from primary oocytes andtheir development to fertilizable metaphase-II oocytes (Pesceet al., 1998; Tadokora et al., 2002). For isolation of granulosa cells,ovaries from 4 to 8 days old female pups were dissected andgranulosa cells were separated from germ cells by a two-stepdifferential adhesion procedure.
2.11. Teratomas formation test
Mouse Oct-4-GFP ovarian germ line stem cells were examinedfor their ability to form teratomas in vivo by subcutaneous, intramuscular or injection into the seminiferous tubules of nude mice(Harlan, USA). As positive controls for teratoma formation, ES cellswere injected in some mice. For subcutaneous, intramuscular ortesticular injections, approximately 1�106 cells were injected.Mice were sacrificed 6 weeks later, and tissues were harvested formorphological and histological analysis. Some animals weresacrificed 7 months after transplantation and in addition tomorphological examination, the harvested tissue was used fordetection of the GFP-DNA using PCR amplification. All animalexperiments were conducted in accordance with the NationalResearch Council’s Guidelines for the Care and Use of LaboratoryAnimals.
3. Results
3.1. Characterization of ovarian germ line stem cells
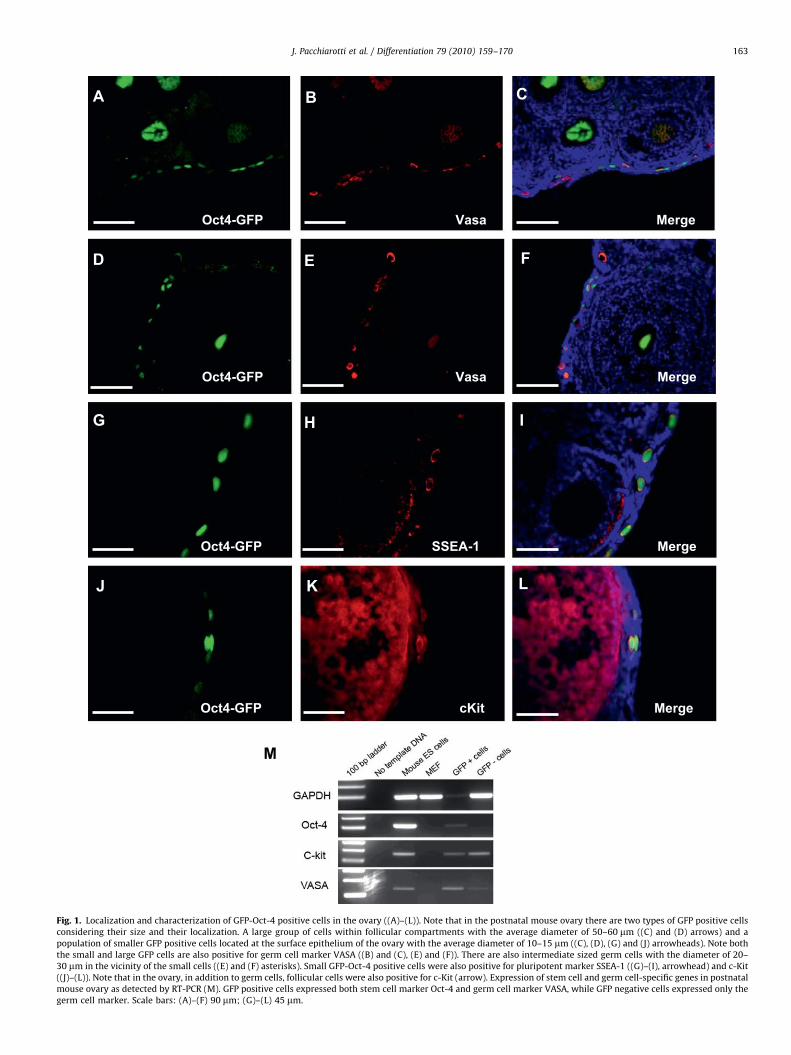
Localization and size of the GFP-Oct-4 positive cells in bothneonatal and adult mouse ovary indicates the presence of germline stem cells in postnatal ovary. Two distinct populations ofGFP-Oct-4 positive cells were found in the mouse ovary, based ontheir size and distribution. A small group of cells, with the averagediameter of 10–15 mm, located at the surface epithelium of theovary and much larger cells, with the average diameter of50–60 mm, located in the center of the follicular compartment,representing oocytes. There were also some medium size (averagediameter of 20–30 mm) GFP-Oct-4 positive cells located in mouseovary indicating the development of large GFP-Oct-4 positiveoocytes from the small GFP-Oct-4 positive cells (Fig. 1).Immunohistochemical staining confirmed that GFP-Oct-4positive cells at the surface epithelium of the mouse ovary arealso positive for germ cell marker VASA and c-Kit and pluripotentmarker SSEA-1. In addition, RT-PCR analysis showed that GFP-Oct-4 positive cells isolated from neonatal mouse ovary expressedmRNA for both pluripotent marker Oct-4 and germ cell markersVASA and c-Kit indicating the presence of germ stem cells in thispopulation, while the GFP negative cells showed only theexpression of germ cell markers. GFP-Oct-4 positive cells also
showed moderate levels of telomerase activity as compared tomouse embryonic stem cells (Supplementary Fig. 2).
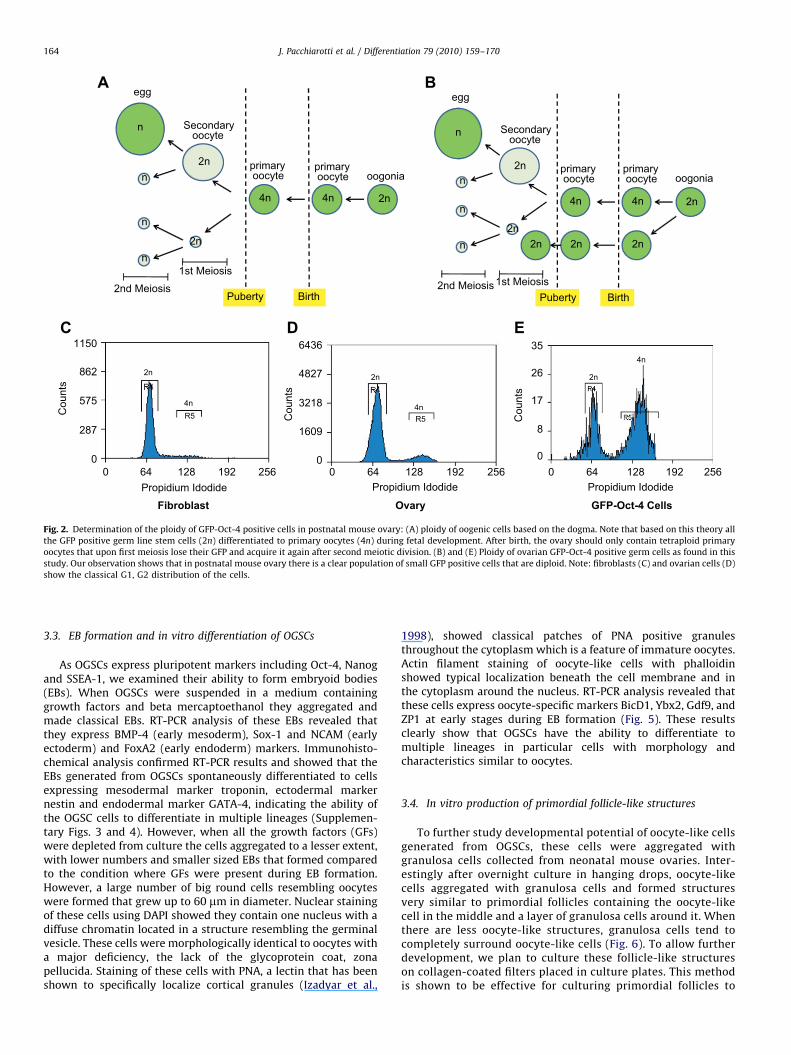
Flow cytometry analysis revealed that the percentage ofGFP-Oct-4 positive cells in the mouse ovary significantly dimin-ished with age. While 1–2% GFP positive cells were found in theovaries of the neonatal mice, only 0.05% was present in the adultovary. Among the GFP-Oct-4 positive cells, 60% were negative orexpressed low levels of c-Kit and 40% showed high levels of c-Kitexpression (Supplementary Fig. 1). Next, we analyzed the ploidyof GFP-Oct-4 positive cells using propidium iodide and flowcytometry. While about 70% of GFP-Oct-4 positive cells weretetraploid (4n), 30% of these cells were diploid (2n). Thesecharacterization studies clearly demonstrate the presence of adiploid cell population in postnatal mouse ovary with germ celland stem cell characteristics (Fig. 2). To verify that the stem cellpopulation found in the postnatal mouse ovary is different fromthe circulating stem cells with hematopoietic origin, we studiedCD133 expression in the OG2 mouse ovary. Interestingly therewas a clear population of CD133 cells in the ovary; however, noneof the CD133 positive cells co-localized GFP-Oct-4 indicating thatgerm line stem cells in the mouse ovary are distinct from thecirculating CD133 cells (Supplementary Fig. 5).
3.2. Generation of ovarian germ line stem cell lines (OGSC)
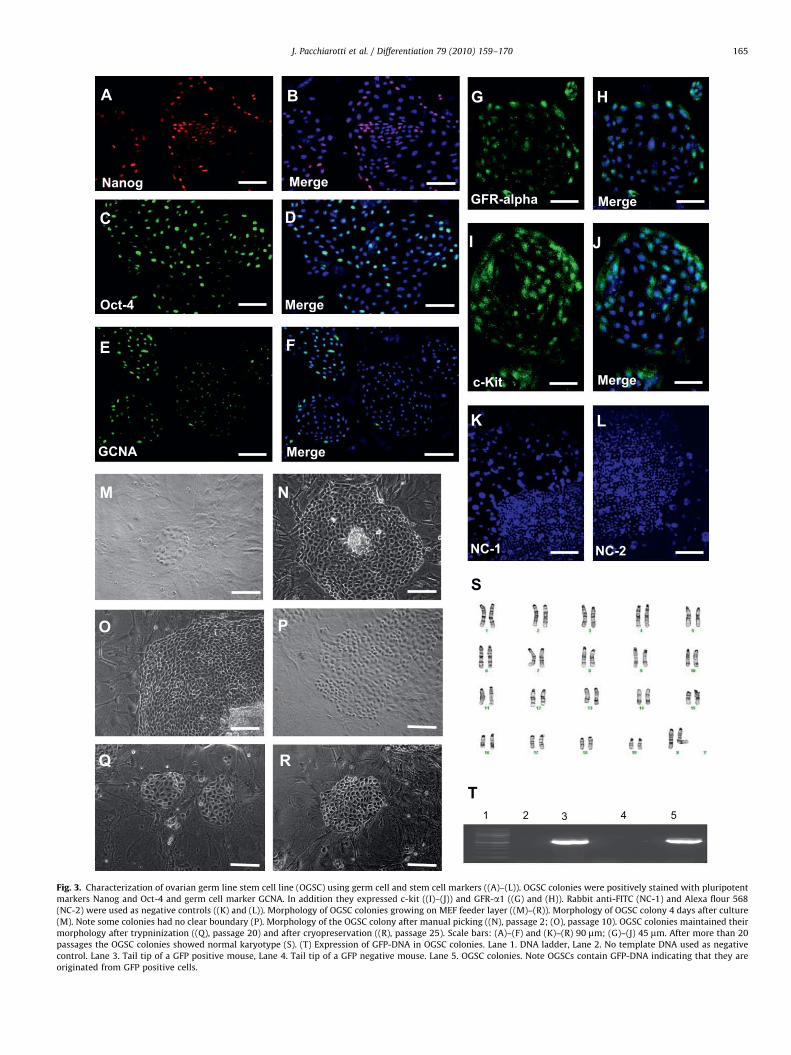
Using the same culture method we recently developed forexpansion of male germ line stem cells (Izadyar et al., 2008), wesucceeded to expand GFP-Oct-4 positive sorted cells from thepostnatal ovary in culture. GFP positive cells cultured on MEFsformed round flat colonies, some showed a clear boundary andsome appeared as a monolayer without a clear border. In total,seven attempts have been made for cell line derivation and onaverage 10–20 OG2 pups were used in each attempt. OGSC lineswith similar morphology and characteristics were generatedsuccessfully in all seven attempts indicating the reproducibilityof this protocol. These colonies maintained their telomeraseactivity and normal karyotype after more than 20 passages(Supplementary Fig. 2). In addition, OGSC colonies were stainedpositive for germ cell markers GCNA and c-Kit, pluripotentmarkers Oct-4, Nanog and GFR-a1, the receptor of GDNF(Fig. 3). Furthermore, subcutaneous, intramuscular, or ovariantransplantation of these cells did not result in malignancyindicating that they are not pluripotent (Supplementary Fig. 6).
OGSC colonies could be expanded by enzymatic digestionusing trypsin and generated new colonies. After several passagesthese colonies retain their morphology; however, they did notshow GFP expression in the majority of the cells, but GFP-DNAwas present (Fig. 3). Only a few cells in each colony, mainly at thecenter of the colony become larger and gradually show GFPexpression. These cells move apart from the colony and makeindividual round shaped cells with different sizes (up to 40 mm indiameter). We speculate these cells might be primary oocytes thathave been generated from the ovarian germ line stem cell colony.Flow cytometry using standard beads also confirmed the presenceof large cells with the diameter of 40 mm in culture. Interestinglythese large cells had a big nucleus containing diffuse chromatinsimilar to the germinal vesicle stage oocytes. These observationsindicate that ovarian germ line stem cells might have beenspontaneously differentiated to primary oocytes. RT-PCR analysisrevealed that cultures containing these cells express oocyte-specific markers. Moreover, few of these oocyte-like cells weresurrounded by a layer of other cells resembling primordial folliclestructures (Fig. 4). Under this culture condition, differentiationbeyond this stage was not observed.
ARTICLE IN PRESS
Oct4-GFP
Oct4-GFP
Vasa
SSEA-1
Merge
Merge
Oct4-GFP cKit Merge
A
M
B
F
C
ED
IHG
J K L
Oct4-GFP Vasa Merge
Fig. 1. Localization and characterization of GFP-Oct-4 positive cells in the ovary ((A)–(L)). Note that in the postnatal mouse ovary there are two types of GFP positive cells
considering their size and their localization. A large group of cells within follicular compartments with the average diameter of 50–60 mm ((C) and (D) arrows) and a
population of smaller GFP positive cells located at the surface epithelium of the ovary with the average diameter of 10–15 mm ((C), (D), (G) and (J) arrowheads). Note both
the small and large GFP cells are also positive for germ cell marker VASA ((B) and (C), (E) and (F)). There are also intermediate sized germ cells with the diameter of 20–
30 mm in the vicinity of the small cells ((E) and (F) asterisks). Small GFP-Oct-4 positive cells were also positive for pluripotent marker SSEA-1 ((G)–(I), arrowhead) and c-Kit
((J)–(L)). Note that in the ovary, in addition to germ cells, follicular cells were also positive for c-Kit (arrow). Expression of stem cell and germ cell-specific genes in postnatal
mouse ovary as detected by RT-PCR (M). GFP positive cells expressed both stem cell marker Oct-4 and germ cell marker VASA, while GFP negative cells expressed only the
germ cell marker. Scale bars: (A)–(F) 90 mm; (G)–(L) 45 mm.
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170 163
ARTICLE IN PRESS
128 192 256
2n
4n
0
8
17
26
35
Cou
nts
Propidium Idodide0 128 192 256
0
1609
3218
4827
6436
R5
2n
4n
Cou
nts
Propidium Idodide
BirthPubertyBirthPuberty
oogoniaprimary oocyte
primary oocyte
Secondary oocyte
egg
1st Meiosis2nd Meiosis
oogoniaprimary oocyte
primary oocyte
Secondary oocyte
egg
1st Meiosis2nd Meiosis
2n
n
n
n2n
n
n
n
2n4n4n
2n
n
2n4n4n
2n
n
2n2n2n
Fibroblast Ovary GFP-Oct-4 Cells
0 128 1920
287
575
862
1150
R5
2n
4n
Cou
nts
Propidium Idodide64 256 64 0 64
Fig. 2. Determination of the ploidy of GFP-Oct-4 positive cells in postnatal mouse ovary: (A) ploidy of oogenic cells based on the dogma. Note that based on this theory all
the GFP positive germ line stem cells (2n) differentiated to primary oocytes (4n) during fetal development. After birth, the ovary should only contain tetraploid primary
oocytes that upon first meiosis lose their GFP and acquire it again after second meiotic division. (B) and (E) Ploidy of ovarian GFP-Oct-4 positive germ cells as found in this
study. Our observation shows that in postnatal mouse ovary there is a clear population of small GFP positive cells that are diploid. Note: fibroblasts (C) and ovarian cells (D)
show the classical G1, G2 distribution of the cells.
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170164
3.3. EB formation and in vitro differentiation of OGSCs
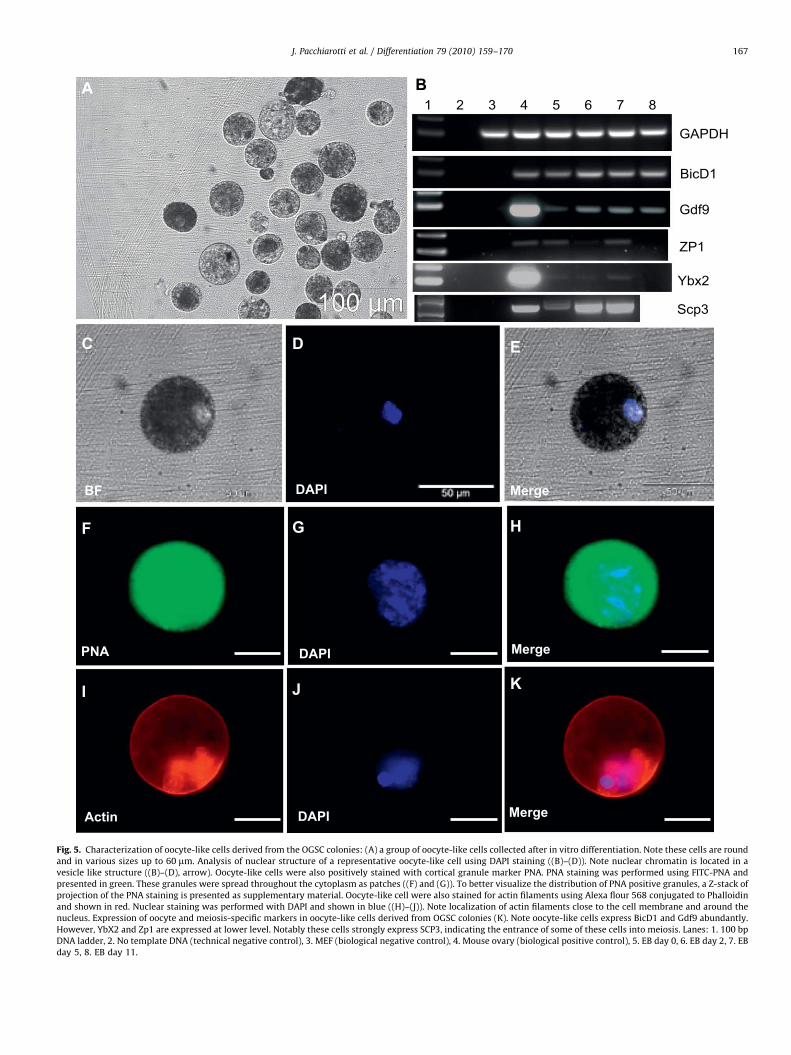
As OGSCs express pluripotent markers including Oct-4, Nanogand SSEA-1, we examined their ability to form embryoid bodies(EBs). When OGSCs were suspended in a medium containinggrowth factors and beta mercaptoethanol they aggregated andmade classical EBs. RT-PCR analysis of these EBs revealed thatthey express BMP-4 (early mesoderm), Sox-1 and NCAM (earlyectoderm) and FoxA2 (early endoderm) markers. Immunohisto-chemical analysis confirmed RT-PCR results and showed that theEBs generated from OGSCs spontaneously differentiated to cellsexpressing mesodermal marker troponin, ectodermal markernestin and endodermal marker GATA-4, indicating the ability ofthe OGSC cells to differentiate in multiple lineages (Supplemen-tary Figs. 3 and 4). However, when all the growth factors (GFs)were depleted from culture the cells aggregated to a lesser extent,with lower numbers and smaller sized EBs that formed comparedto the condition where GFs were present during EB formation.However, a large number of big round cells resembling oocyteswere formed that grew up to 60 mm in diameter. Nuclear stainingof these cells using DAPI showed they contain one nucleus with adiffuse chromatin located in a structure resembling the germinalvesicle. These cells were morphologically identical to oocytes witha major deficiency, the lack of the glycoprotein coat, zonapellucida. Staining of these cells with PNA, a lectin that has beenshown to specifically localize cortical granules (Izadyar et al.,
1998), showed classical patches of PNA positive granulesthroughout the cytoplasm which is a feature of immature oocytes.Actin filament staining of oocyte-like cells with phalloidinshowed typical localization beneath the cell membrane and inthe cytoplasm around the nucleus. RT-PCR analysis revealed thatthese cells express oocyte-specific markers BicD1, Ybx2, Gdf9, andZP1 at early stages during EB formation (Fig. 5). These resultsclearly show that OGSCs have the ability to differentiate tomultiple lineages in particular cells with morphology andcharacteristics similar to oocytes.
3.4. In vitro production of primordial follicle-like structures
To further study developmental potential of oocyte-like cellsgenerated from OGSCs, these cells were aggregated withgranulosa cells collected from neonatal mouse ovaries. Inter-estingly after overnight culture in hanging drops, oocyte-likecells aggregated with granulosa cells and formed structuresvery similar to primordial follicles containing the oocyte-likecell in the middle and a layer of granulosa cells around it. Whenthere are less oocyte-like structures, granulosa cells tend tocompletely surround oocyte-like cells (Fig. 6). To allow furtherdevelopment, we plan to culture these follicle-like structureson collagen-coated filters placed in culture plates. This methodis shown to be effective for culturing primordial follicles to
ARTICLE IN PRESS
Merge
Merge
Merge
Merge
Nanog
Oct-4
GFR-alpha
c-Kit
GCNA Merge
A B
C D
E F
G H
I J
T
S
NM
O P
Q R
1 2 3 4 5
LK
NC-1 NC-2
Fig. 3. Characterization of ovarian germ line stem cell line (OGSC) using germ cell and stem cell markers ((A)–(L)). OGSC colonies were positively stained with pluripotent
markers Nanog and Oct-4 and germ cell marker GCNA. In addition they expressed c-kit ((I)–(J)) and GFR-a1 ((G) and (H)). Rabbit anti-FITC (NC-1) and Alexa flour 568
(NC-2) were used as negative controls ((K) and (L)). Morphology of OGSC colonies growing on MEF feeder layer ((M)–(R)). Morphology of OGSC colony 4 days after culture
(M). Note some colonies had no clear boundary (P). Morphology of the OGSC colony after manual picking ((N), passage 2; (O), passage 10). OGSC colonies maintained their
morphology after trypninization ((Q), passage 20) and after cryopreservation ((R), passage 25). Scale bars: (A)–(F) and (K)–(R) 90 mm; (G)–(J) 45 mm. After more than 20
passages the OGSC colonies showed normal karyotype (S). (T) Expression of GFP-DNA in OGSC colonies. Lane 1. DNA ladder, Lane 2. No template DNA used as negative
control. Lane 3. Tail tip of a GFP positive mouse, Lane 4. Tail tip of a GFP negative mouse. Lane 5. OGSC colonies. Note OGSCs contain GFP-DNA indicating that they are
originated from GFP positive cells.
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170 165

ARTICLE IN PRESS
MEF only
0.16
MEF + OG2 cells
12.40
E
A B
C D
Fig. 4. Spontaneous differentiation of OGSC colonies to oocyte-like cells. Note two large round cells (arrows) that are GFP positive and are separated from the OGSC
colonies (A). A large oocyte-like cell with the diameter of 50 mm adjacent to a OGSC colony as shown by hematoxylin–eosin stain (B). An OGSC colony containing numerous
round cells (C). A very large cell embedded on the MEF feeder and surrounded by a layer of cell resembling primordial follicle (D). Scale bars: (A)–(C) 90 mm; (D) 5 mm.
Determination of the size of the cells by flow Cytometry ((E) and (F)). R indicates cells that are smaller than 15 mm and R7 indicates cells that are larger than 15 mm. Note
that MEF cells are mostly at R1 region. Plates contained OGSC spontaneously differentiated to oocyte-like cells contained 12% cells that are larger than 15 mm. Based on the
side scatter the largest cells are 40–50 mm in diameter.
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170166
mature follicles (Eppig and O’Brien, 1996; Epigg andWigglesworth, 2000). The functionality of OGSC-derivedoocytes using in vitro maturation, fertilization and embryodevelopment as well as ovarian transplantation is currentlyunder investigation.
4. Discussion
In this study using a transgenic mouse model in which GFP isconstructed in a germ line-specific region of Oct-4 promoter wehave been able to track germ line stem cells in postnatal mouse
ARTICLE IN PRESS
1 2 3 4 5 6 7 8
GAPDH
BicD1
Gdf9
ZP1
Ybx2
Scp3
B
J
C D E
A
DAPI
DAPI
PNA
F G H
Merge
MergeBF
DAPI
I
Actin
K
Merge
Fig. 5. Characterization of oocyte-like cells derived from the OGSC colonies: (A) a group of oocyte-like cells collected after in vitro differentiation. Note these cells are round
and in various sizes up to 60 mm. Analysis of nuclear structure of a representative oocyte-like cell using DAPI staining ((B)–(D)). Note nuclear chromatin is located in a
vesicle like structure ((B)–(D), arrow). Oocyte-like cells were also positively stained with cortical granule marker PNA. PNA staining was performed using FITC-PNA and
presented in green. These granules were spread throughout the cytoplasm as patches ((F) and (G)). To better visualize the distribution of PNA positive granules, a Z-stack of
projection of the PNA staining is presented as supplementary material. Oocyte-like cell were also stained for actin filaments using Alexa flour 568 conjugated to Phalloidin
and shown in red. Nuclear staining was performed with DAPI and shown in blue ((H)–(J)). Note localization of actin filaments close to the cell membrane and around the
nucleus. Expression of oocyte and meiosis-specific markers in oocyte-like cells derived from OGSC colonies (K). Note oocyte-like cells express BicD1 and Gdf9 abundantly.
However, YbX2 and Zp1 are expressed at lower level. Notably these cells strongly express SCP3, indicating the entrance of some of these cells into meiosis. Lanes: 1. 100 bp
DNA ladder, 2. No template DNA (technical negative control), 3. MEF (biological negative control), 4. Mouse ovary (biological positive control), 5. EB day 0, 6. EB day 2, 7. EB
day 5, 8. EB day 11.
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170 167
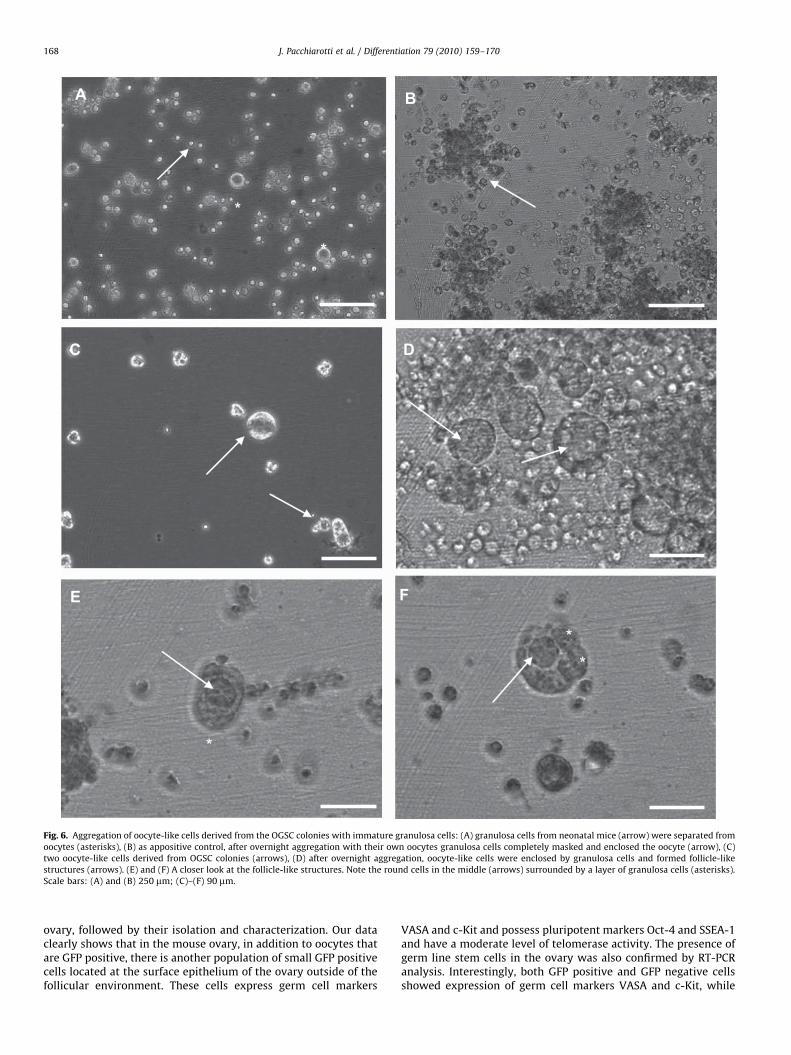
ARTICLE IN PRESS
A B
C D
E F
*
*
**
*
Fig. 6. Aggregation of oocyte-like cells derived from the OGSC colonies with immature granulosa cells: (A) granulosa cells from neonatal mice (arrow) were separated from
oocytes (asterisks), (B) as appositive control, after overnight aggregation with their own oocytes granulosa cells completely masked and enclosed the oocyte (arrow), (C)
two oocyte-like cells derived from OGSC colonies (arrows), (D) after overnight aggregation, oocyte-like cells were enclosed by granulosa cells and formed follicle-like
structures (arrows). (E) and (F) A closer look at the follicle-like structures. Note the round cells in the middle (arrows) surrounded by a layer of granulosa cells (asterisks).
Scale bars: (A) and (B) 250 mm; (C)–(F) 90 mm.
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170168
ovary, followed by their isolation and characterization. Our dataclearly shows that in the mouse ovary, in addition to oocytes thatare GFP positive, there is another population of small GFP positivecells located at the surface epithelium of the ovary outside of thefollicular environment. These cells express germ cell markers
VASA and c-Kit and possess pluripotent markers Oct-4 and SSEA-1and have a moderate level of telomerase activity. The presence ofgerm line stem cells in the ovary was also confirmed by RT-PCRanalysis. Interestingly, both GFP positive and GFP negative cellsshowed expression of germ cell markers VASA and c-Kit, while

ARTICLE IN PRESS
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170 169
only the GFP positive cells showed Oct-4 expression, indicatingthat the GFP positive cells are enriched in germ line stem cells.More importantly we found that there is a distinct population ofsmall GFP-Oct-4 positive cells which are diploid. As primaryoocytes tend to be larger in diameter and are known to havetetraploid DNA content (Kleckner, 1994), the presence of smalldiploid GFP-Oct-4 positive germ cells strongly indicates thepresence of oogonia residing in the postnatal mouse ovary.
Search for the germ line stem cells in the female reproductivesystem resulted in contradictory conclusions. Several indepen-dent studies point to the possibility of the presence of germ linestem cells in the mouse ovary. First, studies about the kinetics ofthe follicle atrasia in the mouse ovary revealed that underphysiological condition, without postnatal production of folliclesthe mouse would not reach its expected reproductive lifespan(Johnson et al., 2004). Similar studies using cytotoxic agents alsosuggested the novo production of follicles and the possibility ofthe existence of a germ line stem cell pool in the postnatal mouseovary (Generoso et al., 1971). Studies about the presence ofmitotically active germ cells in postnatal ovary also resulted incontroversial conclusions. While a previous study by Byskov et al.(2005), using BRDU labeling in the ovary, showed no active celldivision in the germ cells, a recent study using proliferation cellnuclear antigen (PCNA) and germ cell nuclear antigen (GCNA)antibodies shows the presence of actively dividing cells at thesurface epithelium of ovaries of prepubertal mouse (Kerr et al.,2006). Recent study by Zou et al., (2009) is the strongest evidenceso far showing the existence of a germ line stem cell pool in thepostnatal mouse ovary that could be multiplied in vitro andregenerate oogenesis following ovarian transplantation resultingin live offspring. However, the localization and characterization ofgerm line stem cells in the ovary as well as their possible role forde novo production of oocytes in vivo under physiologicalconditions remains to be understood.
To the best of our knowledge, our results clearly demonstratefor the first time the localization of a germ line stem cellpopulation outside follicular environment in the postnatal mouseovary. Our preliminary results using PCNA antibody, concerningthe proliferation activity of GFP-Oct-4 cells at the surfaceepithelium of ovary revealed that these cells are negative (Datanot shown). We speculate that ovarian germ line stem cells mostlikely do not play an active role in oogenesis after birth and are areserved pool in the quiescent state which can be activated andmultiplied under certain conditions, i.e. stimulation of certainfactors. Our culture experiments strongly support this idea as wehave been able to expand these cells under the influence of amixture of growth factors for many passages. Interestingly,growth factors used for expansion of male germ line stem cells(Izadyar et al., 2008) also supported female germ line stem cells.Among the four growth factors used in this mixture, GDNF isknown to be a major regulator of male germ line stem cells(Costoya et al., 2004; Oatley and Brinster, 2008). Interestingly,OGSC colonies were positively stained for GFR-a1, the receptor forGDNF, indicating that both male and female germ line stem cellsmight share similar pathways for their self-renewal. OGSCcolonies maintained their morphological characteristics andtelomerase activity after over 20 passages and expressed germcell marker VASA and pluripotent markers Oct-4 and Nanog,indicating their long-term self-renewal capacity. OGSC coloniesalso expressed c-Kit receptor molecule, indicating that thismolecule might have a role at early stages of oogenesis. C-Kitand its ligand stem cell factor (SCF) are one of the key regulatorsof differentiation of germ line stem cells (De Miguel et al., 2002)and are required during various stages of oogenesis and folliculardevelopment (Yoshida et al., 1997; Kang et al., 2003). In ourprevious study about germ line stem cells in neonatal OG2 mouse
testes, we found that the c-Kit positive population of germ linestem cells can generate multipotent germ line stem cells inculture. Recently, using specific culture conditions, pluripotentcell lines have been generated from unipotent germ line stemcells from the mouse testes (Ko et al., 2009). This indicates thattheir female counterpart also might have a multipotent ability.Indeed, female germ line stem cells were capable of aggregatingand forming EBs expressing markers of three germ layers. Moreadvanced studies using specific differentiation protocols areneeded to study differentiation of these cells to various pheno-types. Transplantation of OGSCs into immunodefficient mice didnot result in teratomas formation indicating that they are notpluripotent.
In search of a proper protocol for in vitro differentiation of OGSCsto oocytes, we found that aggregation and removal of growth factorsfrom mouse ES cell cultures induce an efficient oocyte differentiation(Hubner et al., 2003). Using the same strategy OGSCs producedabundant numbers of large, round cells of various sizes (30–60 mm)with the morphology and characteristics of oocytes. Immunohisto-chemical observations using Phalloidin, an actin cytoskeletal marker,and PNA, a marker for cortical granules (Izadyar et al., 1998), incombination with nuclear stain DAPI revealed that these cells arelarge round cells surrounded by actin filaments and contained onediffuse nuclear chromatin and numerous patches of cortical granulesthroughout the cytoplasm. Moreover, RT-PCR analysis using oocyte-specific markers showed that these cells express early and lateoocyte markers including Gdf9 and ZP1. Meiotic-specific markerSCP3 was also expressed in oocyte-like cells indicating some of thesecells have already entered meiosis.
In their natural environment, oocytes grow and develop inprimordial follicles surrounded and supported by granulosa cells.In an attempt to mimic this system in vitro, we have isolatedgranulosa cells from neonatal mice and aggregated them with theoocyte-like cells generated from OGSCs. Previous studies haveshown that granulosa cells obtained from primordial follicles canre-aggregate to primary oocytes, form follicle structures similar toprimordial follicles that can be fully matured and support growthand development of the enclosed oocyte to fertilizable eggs (Eppigand O’Brien, 1996; Epigg and Wigglesworth, 2000). Interestingly,OGSC-derived oocyte-like cells aggregated with granulosa cellsand formed structures similar to primordial follicles. In vitro andin vivo development of these follicles is under investigation in ourlaboratory. In conclusion, our results clearly demonstrate thepresence of a population of germ line stem cells in postnatalmouse ovary with the ability to self-renew and differentiate tooocyte-like cells that might be useful for follicle engineering andassisted reproductive technologies. In addition our resultsindicate that OGSCs are multipotent and might be used for othertissue regeneration purposes.
Acknowledgments
We would like to thank Carl Javier, Jessie Kinjo, MichaelPascual, Cyril Sirk, Michael Rosenberg, Min Chun and Stacy Phamfor technical and administrative support. We also acknowledgeDr. Enders from the University of Kansas Medical Center forgenerous providing the GCNA antibody.
Appendix A. Supplementary data
Supplementary data associated with this article can be foundin the online version at doi:10.1016/j.diff.2010.01.001.

ARTICLE IN PRESS
J. Pacchiarotti et al. / Differentiation 79 (2010) 159–170170
References
Bristol-Gould, S.K., Kreeger, P.K., SelKirk, C.G., Kilen, S.M., Mayo, K.E., Shea, L.D.,Woodruff, T.K., 2006. Fate of the initial follicle pool: empirical andmathematical evidence supporting its sufficiency for adult fertility. Dev. Biol.298, 149–154.
Byskov, A.G., Faddy, M.J., Lemmen, J.G., Andersen, C.Y., 2005. Eggs forever?Differentiation 73, 438–446.
Castrillon, D.H., Quade, B.J., Wang, T.Y., Quigley, C., Crum, C.P., 2000. The humanVASA gene is specifically expressed in the germ cell lineage. PNAS 97,9585–9590.
Costoya, J.A., Hobbs, R.M., Barna, M., Cattoretti, G., Manova, K., Sukhwani, M.,Orwig, K.E., Wolgemuth, D.J., Pandolfi, P.P., 2004. Essential role of PLZF inmaintenance of spermatogonial stem cells. Nat. Genet. 36, 551–553.
De Miguel, M.P., Cheng, L., Holland, E.C., Federspiel, M.J., Donovan, P.J., 2002.Dissection of the c-Kit signaling pathway in mouse primordial germ cells byretroviral mediated gene transfer. PNAS 99, 10458–10463.
Eggan, K., Jurga, S., Gosden, R., Min, I.M., Wagers, A.J., 2006. Ovulated oocytes inadult mice derived from non-circulating germ cells. Nature 441, 1109–1114.
Eppig, J.J., O’Brien, M.J., 1996. Development in vitro of mouse oocytes fromprimordial follicles. Biol Reprod 54, 197–207.
Epigg, J.J., Wigglesworth, K., 2000. Development of mouse and rat oocytes inchimeric reaggregated ovaries after interspecific exchange of somatic andgerm cell components. Biol. Reprod. 63, 1014–1023.
Gosden, R.G., 2004. Germline stem cells in the postnatal ovary: is the ovary morelike a testis? Hum. Reprod. Update 10, 193–195.
Generoso, W.M., Stout, S.K., Huff, S.W., 1971. Effects of alkylating chemicals onreproductive capacity of adult female mice. Mutat. Res. 13, 172–184.
Hutt, K., Albertini, D., F., 2006. Clinical applications and limitations of currentovarian stem cell research: a review. J. Exp. Clin. Assist. Reprod. 3, 6–10.
Hubner, K., Fuhrmann, G., Christenson, L.K., Kehler, J., Reinbold, R., De La Fuente, R.,Wood, J., Strauss, J.F., Boiani, M., Scholer, H.R., 2003. Derivation of oocytes frommouse embryonic stem cells. Science 300, 1251–1256.
Hou, M., Xu, D., Bjorkholm, M., Gruber, A., 2001. Real-time quantitative telomericrepeat amplification protocol assay for the detection of telomerase activity.Clin. Chem. 47, 519–524.
Izadyar, F., Pau, F., Marh, J., Slepko, N., Wang, T., Gonzalez, R., Ramos, T., Howerton,K., Sayre, C., Silva, F., 2008. Generation of multipotent cell lines from distinctpopulation of male germline stem cells. Reproduction 135, 771–784.
Izadyar, F., Hage, W.J., Colenbrander, B., Bevers, M.M., 1998. The promotory effectof growth hormone on the developmental competence of in vitro maturedbovine oocytes is due to improved cytoplasmic maturation. Mol. Reprod. Dev.49, 444–453.
Johnson, J., Canning, J., Kaneko, T., Pru, J.K., Tilly, J., L., 2004. Germline stem cells andfollicular renewal in the postnatal mammalian ovary. Nature 428, 145–150.
Kerr, JB., Duckett, R., Myers, M., Britt, KL., Mladenovska, T., Findlay, JK., 2006.Quantification of healthy follicles in the neonatal and adult mouse ovary:evidence for maintenance of primordial follicle supply. Reproduction 132,95–109.
Kleckner, N., 1994. Germ cells and fertilization, Molecular Biology of the Cell 3rded. Garland Publishing Inc., New York, pp. 1014–1026.
Kang, J.S., Lee, C.J., Lee, J.M., Rha, J.Y., Song, K.W., Park, M.H., 2003. Follicularexpression of c-kit/SCF and inhibin-a in mouse ovary during development.J. Histochem. Cytochem. 51, 1447–1458.
Ko, K., Tapia, N., Wu, G., Kim, J.B., Bravo, M.J., Sasse, P., Glaser, T., Ruau, D., Han,D.W., Greber, B., Hausdorfer, K., Sebastiano, V., Stehling, M., Fleischmann, B.K.,Brustle, O., Zenke, M., Scholer, H.R., 2009. Induction of pluripotency in adultunipotent germline stem cells. Cell Stem Cell 5, 87–96.
Okamota, K., Okazawa, H., Okuda, A., et al., 1990. A novel octamer bindingtranscription factor is differentially expressed in mouse embryonic stem cells.Cell 60, 461–472.
Oatley, J.M., Brinster, R.L., 2008. Regulation of spermatogonial stem cell self-renewal in mammals. Annu. Rev. Cell Dev. Biol. 24, 263–286.
Pesce, M., Wang, X., Wolgemuth, D.J., 1998. Differential expression of the oct-4transcription factor during mouse germ cell differentiation. Method Dev. 71,89–98.
Scholer, H.R., Ruppert, S., Suzuki, N., 1990. New type of POU domain in germ line-specific protein Oct-4. Nature 344, 435–439.
Telfer, E.E., Gosden, R.G., Byskov, A.G., Spears, N., Albertini, D., Andersen, C.Y.,Anderson, R., Braw-Tal, R., Clarke, H., Gougeon, A., McLaughlin, E, Mclaren, A.,McNatty, K., Schatten, G., Silber, S., Tsafriri, A., 2005. On regenerating the ovaryand generating controversy. Cell 122, 821–822.
Toyooka, Y., Tsunekawa, N., Takahashi, Y., Matsui, Y., Satoh, M., Noce, T., 2000.Expression and intracellular localization of mouse vasa-homologue proteinduring germ cell development. Mech. Dev. 93, 139–149.
Tadokora, Y., Yomogida, K., Ohta, H., 2002. Homeostatic regulation ofgerminal stem cell proliferation by GDNF/FSH pathway. Mech. Dev. 113,29–39.
Yeom, YI., Fuhramann, G., Ovitt, C.E., 1996. Germline regulatory element of Oct-4specific for the totipotent cycle of embryonal cells. Development 122,881–894.
Yoshida, H., Takakura, N., Kataoka, H., Kunisada, T., Okamura, H., Nishikawa, S-I.,1997. Stepwise requirement of c-kit tyrosine kinase in mouse ovarian follicledevelopment. Dev. Biol. 184, 122–137.
Zhang, D., Fouad, H., Zoma, D., Salama, S.A., Wentz, M.J, Al-Hendy, A., 2008.Expression of stem cell and germ cell markers within non follicle structures inadult mouse ovary. Reprod. Sci. 15, 139–146.
Zou, K., Yuan, Z., Yang, Z., Luo, H., Sun, K., Zhou, L., Xiang, J., Shi, L., Yu, Q., Zhang, Y.,Hou, R., Wu, J., 2009. Production of offspring from a germline stem cell linederived from neonatal ovaries. Nat. Cell Biol. 11, 631–636.
Copyright © 2022 FDOKUMEN




















