
Differential rRNA Genes Expression in Hexaploid Wheat Related to NOR Methylation
10
Differential rRNA Genes Expression in Hexaploid Wheat Related to NOR Methylation Ana Carvalho & Carlos Polanco & José Lima-Brito & Henrique Guedes-Pinto Published online: 12 January 2010 # Springer-Verlag 2010 Abstract Ribosomal RNA genes expression was analysed in 18 Portuguese bread wheat accessions (Triticum aestivum L. em Thell.), and the number of argyrophilic-nucleolar organiser regions (Ag-NORs) per cultivar was scored. Ten accessions presented six Ag-NORs per metaphase and six nucleoli per interphase, and eight accessions presented four Ag-NORs per metaphase and four nucleoli per interphase. Fluorescent in situ hybridisation with the 45S rDNA sequence pTa71 and genomic DNA from Aegilops tauschii (2n =2x=14, DD) confirmed the Ag-NOR location and identified the six satellited chromosomes as being the chromosome pairs 1B, 6B and 5D. The methylation pattern of the NOR region was studied by Southern blot using pTa71 as probe, which represented a complete rDNA unit of bread wheat. DNA digestions performed by MspI and HpaII resulted in different patterns revealing the high level of cytosine methylation at their recognition sequences. The total percentage values of NOR methylation indicated that wheat accessions with a maximum of four Ag-NORs were more heavily methylated at the NOR region than accessions with a maximum of six Ag-NORs. Keywords Cytosine methylation . Epigenetics . NOR polymorphism . Ribosomal DNA (rDNA) . Triticum aestivum L. Introduction Polyploidy is considered one of the most well-studied phenomenon, namely in cereals, and is considered one of the most important evolutionary processes in higher plants. The first generations of newly formed polyploids are subjected to rapid genetic and epigenetic changes on their way to functional diploidy (Leitch and Bennett 1997; Ozkan et al. 2001). These changes include the elimination of chromosome/genome-specific sequences and transcrip- tional or posttranscriptional modifications caused by DNA methylation or histone acetylation, promoting their silencing. Altered patterns of DNA methylation have been shown to be a frequent outcome in new allopolyploids (Liu et al. 1998; Shaked et al. 2001). Natural allopolyploids such as hexaploid wheat could have probably experimented similar changes through its evolutionary process. DNA methylation is a frequent epigenetic modification in both plants and mammals and does not occur in yeast, Drosophila or Caenorhabditis elegans (Paszkowski and Whitham 2001). Studies of epigenetic regulation in plants and mammals are yielding complementary results that reveal the evolutionary conservation of epigenetic mechanisms, including methylation (Habu et al. 2001). The Arabidopsis thaliana mutant, ddm1, has contributed for epigenetic studies in order to evaluate the DNA methylation mechanisms and consequences in plants (Scheid et al. 1998). Recently, Carvalho et al. (2008) reported that the hypomethylation agent 5-azacytidine induced secondary constrictions in wheat chromosomes and simultaneous nucleolar organiser region (NOR) rearrangements in one F 1 triticale × tritordeum interspecific hybrid. In wheat, ribosomal RNA (rRNA) genes are organised in multiple repetitive families arranged in tandem arrays. Each rDNA repeat unit is composed by the conserved coding region 18S-5.8S-25S rRNA genes, including A. Carvalho : J. Lima-Brito (*) : H. Guedes-Pinto Institute of Biotechnology and Bioengineering (IBB), Centre of Genetics and Biotechnology (CGB), University of Trás-os-Montes and Alto Douro, P. O. Box 1013, 5000-801 Vila Real, Portugal e-mail: [email protected] C. Polanco Department of Genetics and Molecular Biology, Faculty of Biologic and Environmental Sciences, University of Leon, 24007 Leon, Spain Plant Mol Biol Rep (2010) 28:403–412 DOI 10.1007/s11105-009-0165-5
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Differential rRNA Genes Expression in Hexaploid Wheat Related to NOR Methylation

Differential rRNA Genes Expression in Hexaploid WheatRelated to NOR Methylation
Ana Carvalho & Carlos Polanco & José Lima-Brito &
Henrique Guedes-Pinto
Published online: 12 January 2010# Springer-Verlag 2010
Abstract Ribosomal RNA genes expression was analysed in18 Portuguese bread wheat accessions (Triticum aestivum L.em Thell.), and the number of argyrophilic-nucleolarorganiser regions (Ag-NORs) per cultivar was scored. Tenaccessions presented six Ag-NORs per metaphase and sixnucleoli per interphase, and eight accessions presented fourAg-NORs per metaphase and four nucleoli per interphase.Fluorescent in situ hybridisation with the 45S rDNAsequence pTa71 and genomic DNA from Aegilops tauschii(2n=2x=14, DD) confirmed the Ag-NOR location andidentified the six satellited chromosomes as being thechromosome pairs 1B, 6B and 5D. The methylation patternof the NOR region was studied by Southern blot usingpTa71 as probe, which represented a complete rDNA unit ofbread wheat. DNA digestions performed by MspI and HpaIIresulted in different patterns revealing the high level ofcytosine methylation at their recognition sequences. The totalpercentage values of NOR methylation indicated that wheataccessions with a maximum of four Ag-NORs were moreheavily methylated at the NOR region than accessions with amaximum of six Ag-NORs.
Keywords Cytosine methylation . Epigenetics .
NOR polymorphism . Ribosomal DNA (rDNA) .
Triticum aestivum L.
Introduction
Polyploidy is considered one of the most well-studiedphenomenon, namely in cereals, and is considered one ofthe most important evolutionary processes in higher plants.The first generations of newly formed polyploids aresubjected to rapid genetic and epigenetic changes on theirway to functional diploidy (Leitch and Bennett 1997;Ozkan et al. 2001). These changes include the eliminationof chromosome/genome-specific sequences and transcrip-tional or posttranscriptional modifications caused by DNAmethylation or histone acetylation, promoting their silencing.Altered patterns of DNAmethylation have been shown to be afrequent outcome in new allopolyploids (Liu et al. 1998;Shaked et al. 2001). Natural allopolyploids such as hexaploidwheat could have probably experimented similar changesthrough its evolutionary process.
DNA methylation is a frequent epigenetic modificationin both plants and mammals and does not occur in yeast,Drosophila or Caenorhabditis elegans (Paszkowski andWhitham 2001). Studies of epigenetic regulation in plantsand mammals are yielding complementary results that revealthe evolutionary conservation of epigenetic mechanisms,including methylation (Habu et al. 2001). The Arabidopsisthaliana mutant, ddm1, has contributed for epigenetic studiesin order to evaluate the DNA methylation mechanisms andconsequences in plants (Scheid et al. 1998). Recently,Carvalho et al. (2008) reported that the hypomethylationagent 5-azacytidine induced secondary constrictions in wheatchromosomes and simultaneous nucleolar organiser region(NOR) rearrangements in one F1 triticale × tritordeuminterspecific hybrid. In wheat, ribosomal RNA (rRNA) genesare organised in multiple repetitive families arranged intandem arrays. Each rDNA repeat unit is composed by theconserved coding region 18S-5.8S-25S rRNA genes, including
A. Carvalho : J. Lima-Brito (*) :H. Guedes-PintoInstitute of Biotechnology and Bioengineering (IBB),Centre of Genetics and Biotechnology (CGB),University of Trás-os-Montes and Alto Douro,P. O. Box 1013, 5000-801 Vila Real, Portugale-mail: [email protected]
C. PolancoDepartment of Genetics and Molecular Biology, Facultyof Biologic and Environmental Sciences, University of Leon,24007 Leon, Spain
Plant Mol Biol Rep (2010) 28:403–412DOI 10.1007/s11105-009-0165-5

the variable non-coding intergenic spacer (IGS). IGS itselfconsists of a non-transcribed spacer region containing motifsthat are designated as subrepeats and is itself flanked byexternal transcribed spacers. The 5.8S rRNA gene is flanked bytwo internal transcribed spacers, designed as ITS1 and ITS2.The rRNA genes are located in the NORs, which constitute thechromosomal secondary constriction, being the centromerethe primary constriction (McClintock 1934; Navashin 1934).The NOR loci in hexaploid wheat are located on the shortarms of four chromosomes 1A (Nor-A1), 1B (Nor-B1), 6B(Nor-B2) and 5D (Nor-D3) (Crosby 1957; Bhowal 1972), asdetected after nucleolar activity, chromosome morphology,rRNA/DNA in situ hybridisation (Pardue and Gall 1969) andrDNA/DNA in situ hybridisation (Appels et al. 1980;Hutchinson and Miller 1982) analyses. Additional rDNA lociwere also found on the long arms of the chromosomes: 1B(Nor-B6; Jiang and Gill 1994), 7D (Nor-D4; Mukai et al.1991) and 5A (Nor-A7; Jiang and Gill 1994). Nonetheless,only the NORs located in the short arms of chromosomes 1A,1B, 6B and 5D have the capacity of organising nucleolifollowing the hierarchy of dominance 1B>6B>5D>1A(Darvey and Driscoll 1972). The differential NOR activity isnot a direct consequence of their number of rRNA genes(Flavell and O’Dell 1979; Martini and Flavell 1985) but itrelies on the excess of rRNA gene transcription in hexaploidwheat, obligating to the regulation of the total rRNAproduction per cell (Flavell et al. 1988; Sardana et al. 1993).Some authors reported that the number of rRNA genes onhomologous chromosomes could vary substantially amongvarieties (Flavell and Smith 1974; Mohan and Flavell 1974;Sardana et al. 1993) due to factors involving the proportion oftotal potentially active rRNA genes per cell (Martini andFlavell 1985) and the genetic control exerted by genesdispersed in the wheat genome. These genes are located onthe short arm of chromosome 1A (Viegas and Mello-Sampayo 1975), chromosome 5D (Viegas 1980), the longarm of chromosome 6B and at least 13 of the remaining non-nucleolar chromosomes (Flavell and O’Dell 1979; Martiniand Flavell 1985). Moreover, the gene activity could beregulated by trans-acting factors produced by other genes,which bind to regulatory sequences within the IGS (Polancoand Pérez De La Vega 1997).
In the Triticeae tribe, the rye NOR locus is suppressed inthe F1 wheat × rye hybrids (Vieira et al. 1990) and thesynthetic amphiploid triticale (X Triticosecale Wittmack)(Lacadena et al. 1988; Neves et al. 1997), a phenomenoncalled nucleolar dominance or amphyplasty, that was alsodetected in F1 triticale × tritordeum interspecific hybrids(Lima-Brito et al. 1998). In wheat, both active and inactiverRNA genes subjected to nucleolar dominance are heavilymethylated within their IGS regions, but studies withrestriction endonucleases suggested that inactive genes aremethylated somewhat more heavily (Flavell et al. 1988).
Lima-Brito et al. (1998) indicated that the variability at therRNA genes expression in Triticeae could be due to DNAmethylation or variability of the IGS rDNA subrepeatsequences. Other authors suggested that natural selection isthe major force that directs differentiation at the level ofrDNA repeat unit (Flavell et al. 1986; Saghai-Maroof et al.1984; Gupta et al. 2002; Sharma et al. 2004).
In the present work, we intend to analyse the rRNAgenes expression in 18 bread wheat accessions usingsequential silver nitrate staining and fluorescent in situhybridisation (FISH) technique performed with the rDNAsequence pTa71 and genomic DNA from Aegilops tauschii,as probes. We also aim to assess the cytosine methylationlevels at the NOR regions using the Southern blot techniquewith two methylation-sensitive restriction enzymes, HpaIIand MspI.
Materials and Methods
Plant Material
Eighteen bread wheat (2n=6×=42; AABBDD) accessionswere used for this study. Fifteen out of the 18 accessionsbelong to an Old Portuguese collection that was kindlygiven by the Instituto Nacional de Investigação Agrária edas Pescas (INIAP), Elvas, Portugal, and the remainingthree accessions belong to the Portuguese bread wheatcultivar ‘Barbela’ (Table 1).
Sequential Silver Nitrate Staining and FISH
Seeds were germinated in distilled water at 25°C, and threeroot-tips were collected per seed, treated in ice-cold waterfor 24 h and fixed in ethanol/acetic acid (3:1). Fixed root-tips were used for mitotic chromosome spreads preparationsby squash. Salt-nylon silver nitrate staining (see Stack et al.1991) was performed on the chromosome spreads for thevisualisation and scoring of Ag-NORs on metaphase cellsand nucleoli on the interphase nuclei. Then, the silvernitrate staining was removed by immerging the chromosomespreads on a solution of sodium tiossulfate (20 g/L) andpotassium ferrocianide (8 g/L) (9:1) for 1–3 min, followed bya thorough wash in distilled water, air dry and a dehydration in100% ethanol (−20°C) for 10 min. Both the 45S rDNAsequence pTa71 (Gerlach and Bedbrook 1979) and genomicDNA from A. tauschii (2n=2x=14; DD) were labelled bynick translation with digoxigenin-11-dUTP (Roche) andbiotin-16-dUTP (Roche), respectively. The FISH experimentswere performed according to Schwarzacher and Heslop-Harrison (2000). Hybridisation occurred for 16 h at 37°Con the Omnislide Hybaid modified thermocycler. Thehybridisation signals were detected with antibodies conju-
404 Plant Mol Biol Rep (2010) 28:403–412

gated with fluorochromes, namely avidin D-fluorescein(Vector Laboratories) for the biotin-labelled probe and anti-digoxigenin-rhodamine (Roche) for the digoxigenin-labelledprobe. Slides were mounted with Vecta Shield and DAPI(Vector Laboratories), and the hybridisation patterns wereobserved on an AxioPlan 2.0 epifluorescence microscope(Zeiss). The images were captured by a digital cameraAxioCAM (Zeiss) and prepared for printing with the AdobePhotoshop 6.0 software.
The identification of the satellited chromosomes wasbased on previous reported silver-staining banding patternsand ideograms (Amado et al. 1997; Mukai et al. 1993).
Genomic DNA Extraction and Digestion
Four to five young leaves were collected from each breadwheat seedling and conserved at −80°C. DNA extractionfollowed the CTAB protocol (Doyle and Doyle 1987) withminor modifications such as the grinding of the plantmaterial with liquid nitrogen, the cellular lyses at 65°C for 2 hand two consecutive extractions with chloroform-isoamylalcohol (24:1).
Genomic DNA samples (10 μg) were digested overnightwith two methylation-sensitive restriction endonucleases,HpaII and MspI, according to the manufacturer’s (NewEngland Biolabs, Beverly, MA, USA) instructions. Bothenzymes have a 5′-CCGG-3′ recognition sequence. However,MspI is not sensitive to dam, dcm or mammalian CpG
methylation, while HpaII fails to cleave the rDNA when theinner C is methylated. Both MspI and HpaII cannot cleavewhen the external cytosine of the CCGG sequence ismethylated.
Southern Blot Analysis
Southern blots (Southern 1975) were prepared by transfer-ring the digested genomic DNA to a nylon N+ membrane(Roche) using the saline transfer buffer 20× SSC (sodiumcitrate; sodium chloride). The pTa71 cloned sequence wasdigested with the restriction endonuclease EcoRI (NewEngland Biolabs, Beverly, MA, USA) in order to isolate therRNA genes 18S-5.8S-25S, including the IGS region. This8.9-kb fragment was labelled with digoxigenin using the“DIG-High Prime DNA Labeling and Detection Starter KitII” (Roche) following the manufacturer’s instructions. Theprobe labelling was evaluated by the dot blot technique asrecommended by the DIG-High Prime kit. Hybridisationsignals were detected by conventional autoradiography.
The NOR methylation levels were determined from theSouthern blots using the ImageJ software (Abramoff et al.2004). Considering the HpaII/MspI restriction map of onebread wheat rDNA repeat unit (GenBank X07841; Barkeret al. 1988), performed by the Vector NTI software(Invitrogen), the DNA fragments produced by a completedigestion range from 5 bp to 1 kb (data not shown). Thus,any restriction fragment above 1 kb will result from
Table 1 Accession number, botanical variety and common name of 18 bread wheat accessions used in this study
Accession number Botanical variety Common name
7 T. aestivum ssp. aestivum var. ferrugineum ‘Ardito’
24 T. aestivum ssp. aestivum var. aestivum ‘Egípcio’
30 T. aestivum ssp. aestivum var. aestivum ‘Fronteiriço’
36 T. aestivum ssp. aestivum var. graecum ‘Grécia’
46 T. aestivum ssp. aestivum var. ferrugineum ‘Mestiço’
48 T. aestivum ssp. aestivum var. lutescens ‘Mocho Cabeçudo’
50 T. aestivum ssp. aestivum var. lutescens ‘Mocho de Espiga Branca’
51 T. aestivum ssp. aestivum var. milturum ‘Mocho de Espiga Ruivo’
53 T. aestivum ssp. aestivum var. aestivum ‘Mole Algarvio’
75 T. aestivum ssp. aestivum var. aestivum ‘Rieti’
79 T. aestivum ssp. aestivum var. aestivum ‘Sacho’
80 T. aestivum ssp. aestivum var. ferrugineum ‘Saloio’
92 T. aestivum ssp. aestivum var. ferrugineum ‘Temporão de Coruche’
93 T. aestivum ssp. aestivum var. aestivum ‘Transmontano’
101 T. aestivum ssp. aestivum var. aestivum ‘Restauração’
Accessions not included in the Old Portuguese collection
T. aestivum ssp. aestivum var. erythrospermum ‘Barbela’ 6/92 (code: B6)
T. aestivum ssp. aestivum var. erythrospermum ‘Barbela’ 93/92 (code: B93)
T. aestivum ssp. aestivum var. erythrospermum ‘Barbela’ 94/92 (code: B94)
Plant Mol Biol Rep (2010) 28:403–412 405

cytosine methylation in the recognition sites of theserestriction enzymes. The percentage of NOR methylationwas calculated by measuring the percentage of hybrid-isation signal in each lane that corresponded to restrictionfragments above 1 kb. A similar analysis was performed inArabidopsis rRNA genes by Riddle and Richards (2005).
Statistical Analyses
The ANOVA factorial and the post hoc Fisher statisticaltests were performed in order to analyse the effects of eachrestriction enzyme, bread wheat accession and/or maximumnumber of Ag-NORs in the percentage values of NORmethylation. For this purpose, we used the StatView software.The F values significance, due to the different effects andtheir interaction, was established for probabilities higherthan 5% (P>0.05), between 1% and 5% (0.01<P<0.05),between 0.1% and 1% (0.001<P<0.01) and lower than0.1% (P<0.001).
Results and Discussion
rRNA Genes Expression and FISH Analyses
The metaphase Ag-NORs analysis provides the chromosomaldistribution of active versus silent rRNA genes due to thepresence of acidic/argyrophilic domains in the RNA poly-merase I (Pol I) transcription machinery, stainable by silvernitrate (Goodpasture and Bloom 1975). The number of Ag-NORs per metaphase and the number of nucleoli perinterphase nuclei scored on the 18 bread wheat accessionsare presented in Table 2. Ten of the 18 accessions presenteda maximum of six Ag-NORs per metaphase (Fig. 1c) and sixnucleoli per interphase nuclei (Fig. 1e). The remaining eightaccessions showed a maximum of four Ag-NORs permetaphase (Fig. 1a) and four nucleoli per interphase cell(Fig. 1f, g). Sequential FISH performed in the same cellswith the 45S rDNA probe (pTa71) confirmed the Ag-NORslocation (Fig. 1b, d). The pTa71 hybridisation signalsobserved here were compared with those reported by Mukaiet al. (1993). Four major Ag-NORs were identified asbelonging to the chromosome pairs 1B and 6B (Nor-B1 andNor-B2, respectively). The third satellited chromosome pair(5D) was identified by the simultaneous use of pTa71 andgenomic DNA from A. tauschii, as probes (Fig. 1b, d). Eightout of the 18 bread wheat accessions studied showed amaximum of four Ag-NORs (Table 2) due to the Nor-D3locus suppression on 5D (Fig. 1a).
Silent NORs do not appear as secondary constrictionsprobably due to the absence of Pol I transcription factors orother mechanism that operate in organisms with multipleNORs allowing the silence of an entire NOR (McStay
2006). Previous analysis of wheat rDNA revealed that theactivity of the rRNA genes at different chromosomal loci isdifferentially regulated, with some loci exhibiting full orpartial dominance over others (Martini and Flavell 1985;Flavell et al. 1988). The differential silver-staining patternsof the NORs reflected the hierarchy among the rDNAclusters in the wheat genome. Our results confirm that theNor-B1 and Nor-B2 loci have dominance over the Nor-D3locus. The four major Ag-NORs located on the chromosomepairs 1B and 6B were easily detected in contrast to the minorAg-NOR from the 5D chromosome pair (Fig. 1c). As reportedby Chen and Pikaard (1997), nucleolar dominance is anepigenetic phenomenon, which describes the nucleolusformation around the rRNA genes inherited from only oneprogenitor of an interspecific hybrid or allopolyploid. Theallopolyploid nature of hexaploid wheat could explain thedifferential silver-staining patterns between the NORs ofthe B genome (1B and 6B) and the D genome (5D) and theintra-specific nucleolar dominance in bread wheat. This trendhas been widely studied in wheat-rye hybrids and triticale(Vieira et al. 1990; Lacadena et al. 1988). Rye rRNA genessuppression could be reverted by demethylating agents suchas 5-azacytidine (Neves et al. 1995; Caperta et al. 2002). Byother hand, epigenetic changes could be maintained throughgenerations, by natural selection driving forces, constitutinga probable explanation for the Ag-NOR polymorphismdetected here.
RFLP Analysis
NOR silencing has been correlated with DNA methylationof IGS rDNA regions (Sharma et al. 2005). Consequentlythe rRNA genes expression is affected by the methylationof regulatory sequences or enhancer elements present in theIGS region. After the cytogenetic analyses, we consideredthe possibility that cultivars with a maximum of four Ag-NORs might have higher methylated NOR regions thanthose with six Ag-NORs. To test this hypothesis, weanalysed genomic Southern blots, obtained independentlywith the methylation-sensitive endonucleases MspI andHpaII, hybridised with the pTa71 probe (homologous to eachrDNA repeat unit).
Different digestion patterns were achieved by MspI(Fig. 2) and HpaII (Fig. 3), and there was variation in themethylation status at the rDNA regions among the breadwheat accessions analysed.
The results obtained by Sharma et al. (2005) in wildbarley (Hordeum spontaneum), using a similar approach,showed that the CCGG restriction site of HpaII/MspI in therDNA repeat units are heavily methylated because HpaIImethylation patterns of the rDNA were represented by onlyuncut DNA, in contrast to MspI that gave a pattern typicalof rDNA in the autoradiograms. In the present study, we
406 Plant Mol Biol Rep (2010) 28:403–412
have obtained similar results, with higher methylation levelin the HpaII digestion. This enzyme does not cleave theCCGG sequence if the inner C is methylated. Thus, we maysuggest that most CCGG sequences at the NOR region havethe inner cytosine methylated avoiding the HpaII digestion.The total percentage of NOR methylation was determinedfrom the Southern blots using the ImageJ software.Considering the HpaII/MspI restriction map of one breadwheat rDNA repeat unit (GenBank X07841; Barker et al.1988), achieved by the Vector NTI software (Invitrogen),we expected DNA fragments ranging from 5 bp to 1 kb(data not shown). Thus, any restriction fragment above 1 kbwill result from cytosine methylation in the recognitionsites of these restriction enzymes. The percentage of NORmethylation was calculated by measuring the percentage ofhybridisation signal in each lane that corresponded tofragments above 1 kb. Regarding the HpaII blots(Fig. 4a), all bread wheat accessions presented high levelof NOR methylation, independently of their maximumnumber of Ag-NORs. Bread wheat accessions with fourAg-NORs and digested with HpaII presented percentagesof total NOR methylation that ranged from 64.29% to74.56%, whereas the accessions with six Ag-NORs variedfrom 65.77% to 78.36% (Fig. 4a). However, no significantstatistical differences were found among accessions with a
different maximum number of Ag-NORs that were digestedwith HpaII (P>0.05).
Considering the MspI digestions (Fig. 4b), the percentagevalues of NOR methylation in bread wheat accessions withsix Ag-NORs ranged from 40.04% to 54.05%, whereas inaccessions with four Ag-NORs, it ranged from 43.69% to63.63%. No significant statistical differences were foundamong accessions with a different maximum number of Ag-NORs that were digested with MspI (P>0.05). Nonetheless,when we analysed the NOR methylation values achieved byHpaII and MspI among the 18 bread wheat accessions, wefound out significant differences between the two enzymes(P<0.0001). These results suggest that the cytosine residuesof the HpaII/MspI recognition sites in the rDNA of breadwheat are heavily methylated and explain the differentmethylation patterns achieved by the two enzymes.
The complementary approaches used in the presentstudy, cytogenetics and Southern blot, both performed withthe rDNA probe, pTa71, allowed us to determine that mostbread wheat accessions with a maximum of four Ag-NORs(major 1B and 6B NORs) have a higher level of NORmethylation. This is most probably due to the methylationof the 5D rDNA locus, because in these lines it was notactively expressed as shown by the absence of silver nitratestaining (Fig. 1a). Considering the NORs hierarchy in
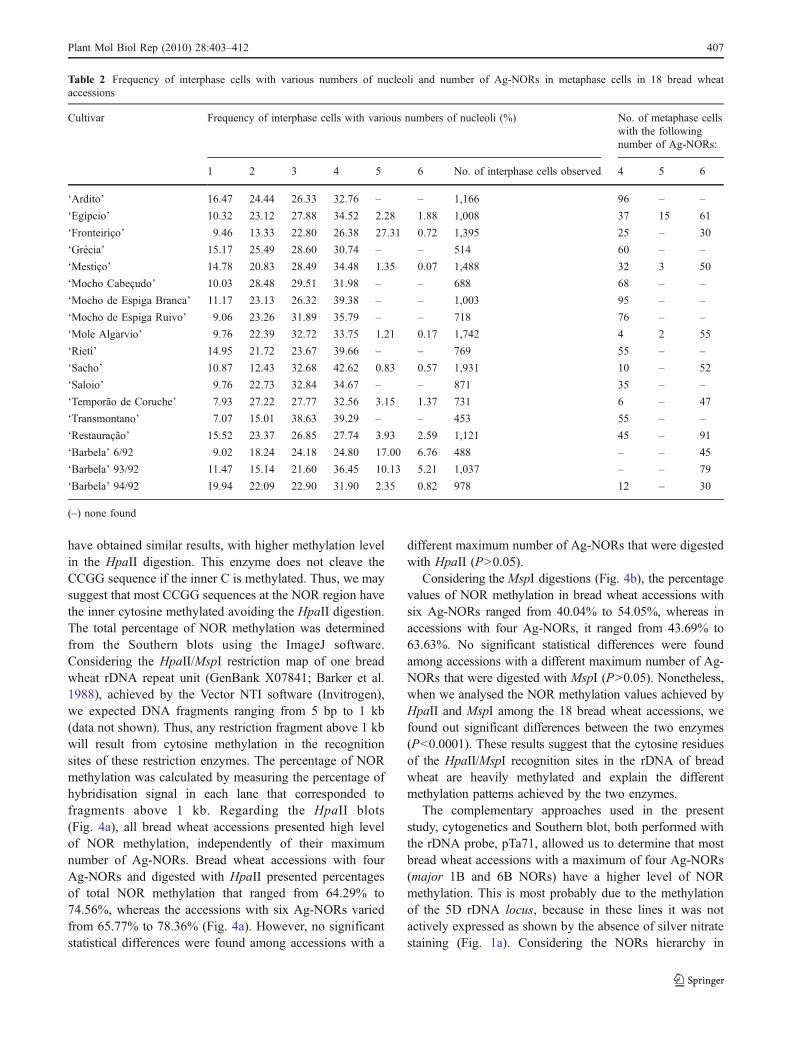
Table 2 Frequency of interphase cells with various numbers of nucleoli and number of Ag-NORs in metaphase cells in 18 bread wheataccessions
Cultivar Frequency of interphase cells with various numbers of nucleoli (%) No. of metaphase cellswith the followingnumber of Ag-NORs:
1 2 3 4 5 6 No. of interphase cells observed 4 5 6
‘Ardito’ 16.47 24.44 26.33 32.76 – – 1,166 96 – –
‘Egípcio’ 10.32 23.12 27.88 34.52 2.28 1.88 1,008 37 15 61
‘Fronteiriço’ 9.46 13.33 22.80 26.38 27.31 0.72 1,395 25 – 30
‘Grécia’ 15.17 25.49 28.60 30.74 – – 514 60 – –
‘Mestiço’ 14.78 20.83 28.49 34.48 1.35 0.07 1,488 32 3 50
‘Mocho Cabeçudo’ 10.03 28.48 29.51 31.98 – – 688 68 – –
‘Mocho de Espiga Branca’ 11.17 23.13 26.32 39.38 – – 1,003 95 – –
‘Mocho de Espiga Ruivo’ 9.06 23.26 31.89 35.79 – – 718 76 – –
‘Mole Algarvio’ 9.76 22.39 32.72 33.75 1.21 0.17 1,742 4 2 55
‘Rieti’ 14.95 21.72 23.67 39.66 – – 769 55 – –
‘Sacho’ 10.87 12.43 32.68 42.62 0.83 0.57 1,931 10 – 52
‘Saloio’ 9.76 22.73 32.84 34.67 – – 871 35 – –
‘Temporão de Coruche’ 7.93 27.22 27.77 32.56 3.15 1.37 731 6 – 47
‘Transmontano’ 7.07 15.01 38.63 39.29 – – 453 55 – –
‘Restauração’ 15.52 23.37 26.85 27.74 3.93 2.59 1,121 45 – 91
‘Barbela’ 6/92 9.02 18.24 24.18 24.80 17.00 6.76 488 – – 45
‘Barbela’ 93/92 11.47 15.14 21.60 36.45 10.13 5.21 1,037 – – 79
‘Barbela’ 94/92 19.94 22.09 22.90 31.90 2.35 0.82 978 12 – 30
(–) none found
Plant Mol Biol Rep (2010) 28:403–412 407
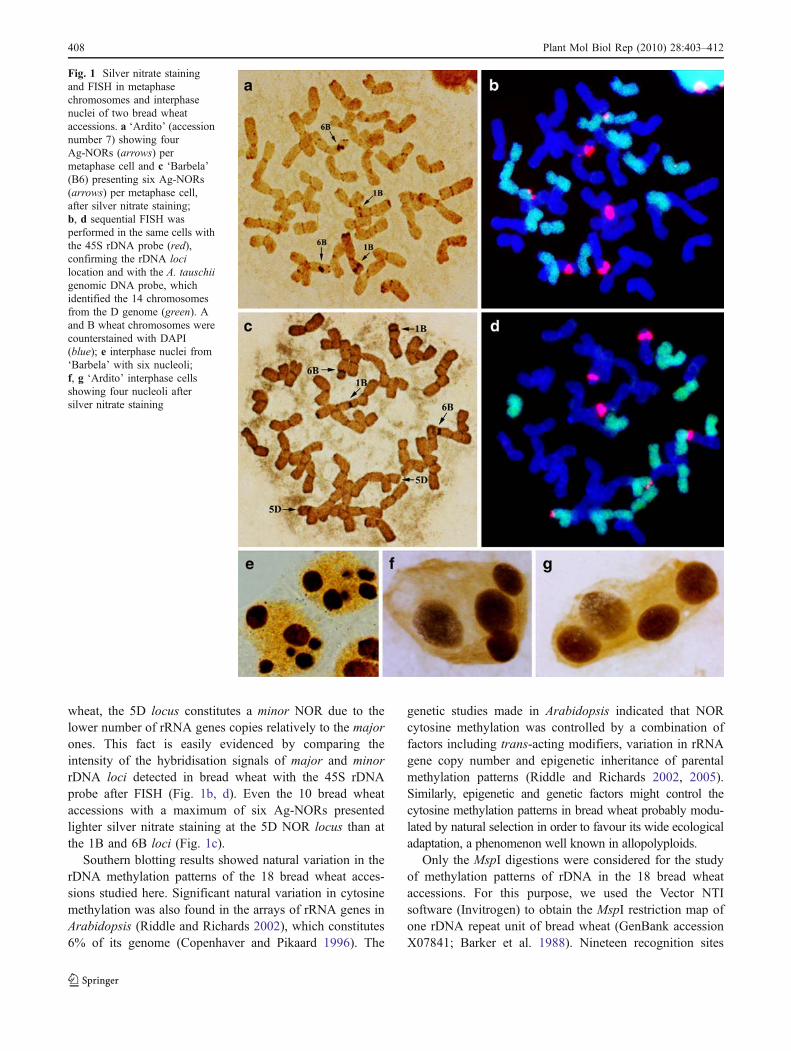
wheat, the 5D locus constitutes a minor NOR due to thelower number of rRNA genes copies relatively to the majorones. This fact is easily evidenced by comparing theintensity of the hybridisation signals of major and minorrDNA loci detected in bread wheat with the 45S rDNAprobe after FISH (Fig. 1b, d). Even the 10 bread wheataccessions with a maximum of six Ag-NORs presentedlighter silver nitrate staining at the 5D NOR locus than atthe 1B and 6B loci (Fig. 1c).
Southern blotting results showed natural variation in therDNA methylation patterns of the 18 bread wheat acces-sions studied here. Significant natural variation in cytosinemethylation was also found in the arrays of rRNA genes inArabidopsis (Riddle and Richards 2002), which constitutes6% of its genome (Copenhaver and Pikaard 1996). The
genetic studies made in Arabidopsis indicated that NORcytosine methylation was controlled by a combination offactors including trans-acting modifiers, variation in rRNAgene copy number and epigenetic inheritance of parentalmethylation patterns (Riddle and Richards 2002, 2005).Similarly, epigenetic and genetic factors might control thecytosine methylation patterns in bread wheat probably modu-lated by natural selection in order to favour its wide ecologicaladaptation, a phenomenon well known in allopolyploids.
Only the MspI digestions were considered for the studyof methylation patterns of rDNA in the 18 bread wheataccessions. For this purpose, we used the Vector NTIsoftware (Invitrogen) to obtain the MspI restriction map ofone rDNA repeat unit of bread wheat (GenBank accessionX07841; Barker et al. 1988). Nineteen recognition sites
Fig. 1 Silver nitrate stainingand FISH in metaphasechromosomes and interphasenuclei of two bread wheataccessions. a ‘Ardito’ (accessionnumber 7) showing fourAg-NORs (arrows) permetaphase cell and c ‘Barbela’(B6) presenting six Ag-NORs(arrows) per metaphase cell,after silver nitrate staining;b, d sequential FISH wasperformed in the same cells withthe 45S rDNA probe (red),confirming the rDNA locilocation and with the A. tauschiigenomic DNA probe, whichidentified the 14 chromosomesfrom the D genome (green). Aand B wheat chromosomes werecounterstained with DAPI(blue); e interphase nuclei from‘Barbela’ with six nucleoli;f, g ‘Ardito’ interphase cellsshowing four nucleoli aftersilver nitrate staining
408 Plant Mol Biol Rep (2010) 28:403–412

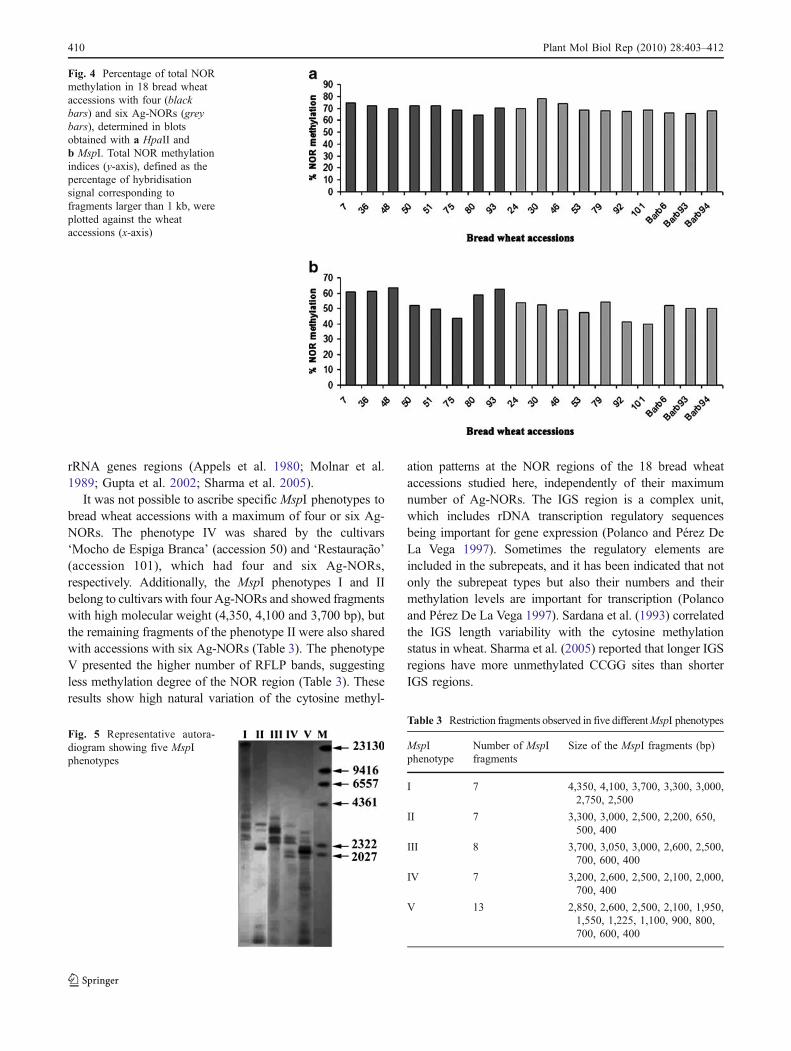
were found, with 12 of them located within the IGS region.Considering this restriction map, a maximum of 20fragments was expected. The MspI digestions hybridisedwith the pTa71 probe revealed five different bandingpatterns throughout the 18 bread wheat accessions, rangingfrom seven to 13 fragments of varying sizes (Fig. 5,Table 3). The five banding patterns were classified as I to VMspI phenotypes (Fig. 5, Table 3).
The results showed in Fig. 5 and Table 3 demonstratedthat some MspI recognition sites are heavily methylated at
their external cytosine residue, avoiding the enzymaticcleavage of the rDNA repeats. Since most MspI recognitionsequences are located within the IGS region, we mightsuggest that the cytosine methylation of the spacer is higherthan that present at the rDNA coding regions. Earlierstudies reported that the heterogeneity for methylation-sensitive enzymes could be either a consequence of themodification in sequences/methylation of recognition siteslocated in the IGS rDNA region or a result of the partial/complete methylation of the restriction sites at the coding
Fig. 3 Different patterns of NOR methylation in T. aestivumaccessions achieved after a genomic DNA digested with HpaII andsize fractionated by gel electrophoresis, b autoradiogram resulted fromhybridisation with pTa71 after 40 min of exposure. Lanes 1 ‘Ardito’,2 ‘Egípcio’, 3 ‘Fronteiriço’, 4 ‘Grécia’, 5 ‘Mestiço’, 6 ‘Mocho
Cabeçudo’, 7 ‘Mocho de Espiga Branca’, 8 ‘Mocho de Espiga Ruivo’,9 ‘Mole Algarvio’, 10 ‘Rieti’, 11 ‘Sacho’, 12 ‘Saloio’, 13 ‘Temporãode Coruche’, 14 ‘Transmontano’, 15 ‘Restauração’, 16 ‘Barbela’ 6/92,17 ‘Barbela’ 94/92, 18 ‘Barbela’ 93/92
Fig. 2 Different patterns of NOR methylation in T. aestivumaccessions (accession numbers are identified per lane) achieved aftera genomic DNA digested with MspI and size fractionated by gelelectrophoresis; b autoradiogram that resulted from hybridisation withpTa71 after 40 min of exposure and c the same autoradiogram after an
overnight exposure. Lanes1 ‘Ardito’, 2 ‘Egípcio’, 3 ‘Grécia’, 4‘Mocho Cabeçudo’, 5 ‘Mocho de Espiga Branca’, 6 ‘Mocho deEspiga Ruivo’, 7 ‘Mole Algarvio’, 8 ‘Rieti’, 9 ‘Restauração’, 10‘Barbela’ 6/92, 11 ‘Barbela’ 94/92, 12 ‘Barbela’ 93/92
Plant Mol Biol Rep (2010) 28:403–412 409
rRNA genes regions (Appels et al. 1980; Molnar et al.1989; Gupta et al. 2002; Sharma et al. 2005).
It was not possible to ascribe specific MspI phenotypes tobread wheat accessions with a maximum of four or six Ag-NORs. The phenotype IV was shared by the cultivars‘Mocho de Espiga Branca’ (accession 50) and ‘Restauração’(accession 101), which had four and six Ag-NORs,respectively. Additionally, the MspI phenotypes I and IIbelong to cultivars with four Ag-NORs and showed fragmentswith high molecular weight (4,350, 4,100 and 3,700 bp), butthe remaining fragments of the phenotype II were also sharedwith accessions with six Ag-NORs (Table 3). The phenotypeV presented the higher number of RFLP bands, suggestingless methylation degree of the NOR region (Table 3). Theseresults show high natural variation of the cytosine methyl-
ation patterns at the NOR regions of the 18 bread wheataccessions studied here, independently of their maximumnumber of Ag-NORs. The IGS region is a complex unit,which includes rDNA transcription regulatory sequencesbeing important for gene expression (Polanco and Pérez DeLa Vega 1997). Sometimes the regulatory elements areincluded in the subrepeats, and it has been indicated that notonly the subrepeat types but also their numbers and theirmethylation levels are important for transcription (Polancoand Pérez De La Vega 1997). Sardana et al. (1993) correlatedthe IGS length variability with the cytosine methylationstatus in wheat. Sharma et al. (2005) reported that longer IGSregions have more unmethylated CCGG sites than shorterIGS regions.
Fig. 5 Representative autora-diogram showing five MspIphenotypes
Fig. 4 Percentage of total NORmethylation in 18 bread wheataccessions with four (blackbars) and six Ag-NORs (greybars), determined in blotsobtained with a HpaII andb MspI. Total NOR methylationindices (y-axis), defined as thepercentage of hybridisationsignal corresponding tofragments larger than 1 kb, wereplotted against the wheataccessions (x-axis)
Table 3 Restriction fragments observed in five differentMspI phenotypes
MspIphenotype
Number of MspIfragments
Size of the MspI fragments (bp)
I 7 4,350, 4,100, 3,700, 3,300, 3,000,2,750, 2,500
II 7 3,300, 3,000, 2,500, 2,200, 650,500, 400
III 8 3,700, 3,050, 3,000, 2,600, 2,500,700, 600, 400
IV 7 3,200, 2,600, 2,500, 2,100, 2,000,700, 400
V 13 2,850, 2,600, 2,500, 2,100, 1,950,1,550, 1,225, 1,100, 900, 800,700, 600, 400
410 Plant Mol Biol Rep (2010) 28:403–412

We have analysed the intra-specific IGS length hetero-geneity of these Old Portuguese bread wheat accessions andfound out high levels of IGS polymorphism among them(unpublished data). This IGS length variability might be thereason of the high natural variation in methylation patternsof the NOR region determined here. The IGS region playsan important role in the regulation of the neighbouringrRNA genes expression (Doerfler 1983; Cedar 1988;Sardana et al. 1993), and its variability has been correlatedwith biological and economic important traits such asdevelopment rates in Drosophila (Cluster et al. 1987) andgrain yield in maize (Rocheford et al. 1990). Sharma et al.(2005) reported that the presence of specificMspI phenotypessuggests that natural selection plays an important role inthe methylation of the IGS nucleotides and, therefore, in theswitching on or off of the rRNA genes expression. TherDNA loci differ in their response towards methylation dueto selection forces, which regulate the IGS sequence activity(Sharma et al. 2005). Other studies also suggested a role ofecogeographical factors in the differentiation of rDNA inwild barley (Gupta et al. 2002; Sharma et al. 2004).
In conclusion, we considered that the variation in themaximum number of Ag-NORs of the bread wheat accessionsstudied here is related with the cytosine methylation levels ofthe NOR region. Although we could not attribute specificMspI phenotypes to accessions with a maximum of four orsix Ag-NORs, we noticed that in general, the formeraccessions presented high levels of NOR methylation.
Acknowledgements This work was supported by the PTDC/AGR-GPL/65876/2006 project and the Ph.D. grant SFRH/BD/17348/2004both financed by the Portuguese Foundation for Science and theTechnology (FCT).
References
Abramoff MD, Magelhaes PJ, Ram SJ (2004) Image processing withImageJ. Biophotonics International 11(7):36–42
Amado L, Abranches R, Neves N, Viegas W (1997) Development-dependent inheritance of 5-azacytidine-induced epimutations intriticale: analysis of rDNA expression patterns. Chromosome Res5:445–450
Appels R, Gerlach WL, Dennis ES, Swift H, Peacock WJ (1980)Molecular and chrosomal organization of DNA sequences codingfor the ribosomal RNAs in cereals. Chromosoma 78:293–311
Barker RF, Harberd NP, Jarvis MG, Flavell RB (1988) Structure andevolution of the intergenic region in a ribosomal DNA repeat unitof wheat. J Mol Biol 201:1–17
Bhowal JG (1972) Nucleolar chromosomes in wheat. Z Pflanzenzücht68:253–257
Caperta A, Neves N, Morais-Cecilio L, Malhó R, Viegas W (2002)Genome restructuring in rye affects the expression, organization anddisposition of homologous rDNA loci. J Cell Sci 115:2839–2846
Carvalho A, Guedes-Pinto H, Heslop-Harrison JS, Lima-Brito J(2008) Wheat neocentromeres found in F1 triticale × tritordeumhybrids (AABBRHch) after 5-azacytidine treatment. Plant MolBiol Rep 26:46–52
Cedar H (1988) DNA methylation and gene activity. Cell 53:3–4Chen ZJ, Pikaard CS (1997) Epigenetic silencing of RNA polymerase
I transcription: a role for DNA methylation and histonemodification in nucleolar dominance. Genes Dev 11:2124–2136
Cluster PD, Marinkovic D, Allard RW, Ayala FJ (1987) Correlationsbetween development rates, enzyme activities, ribosomal DNAspacer-length phenotypes, and adaptation in Drosophila mela-nogaster. Proc Natl Acad Sci USA 84:610–614
Copenhaver GP, Pikaard CS (1996) Two-dimensional RFLP analysesreveal megabase-sized clusters of rRNA gene variants inArabidopsis thaliana, suggesting local spreading of variants asthe mode for gene homogenization during concerted evolution.Plant 9:273–282
Crosby AR (1957) Nucleolar activity of lagging chromosomes inwheat. Am J Bot 44:813–822
Darvey NL, Driscoll CJ (1972) Nucleolar behaviour in Triticum.Chromosoma 36(2):131–139
Doerfler W (1983) DNA methylation and gene activity. Annu RevBiochem 52:93–124
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for smallquantities of fresh leaf tissue. Phytochem Bull 19:11–15
Flavell RB, O’Dell M (1979) The genetic control of nucleolusformation in wheat. Chromosoma 71:135–152
Flavell RB, Smith DB (1974) The role of homoeologous group 1chromosomes in the control of rRNA genes in wheat. BiochemGenet 12(4):271–279
Flavell RB, O’Dell M, Sharp P, Nevo E, Beiles A (1986) Variation inthe intergenic spacer of ribosomal DNA of wild wheat, Triticumdicoccoides, in Israel. Mol Biol Evol 3:547–558
Flavell RB, O’Dell M, Thompson WF (1988) Regulation of cytosinemethylation in ribosomal DNA and nucleolus organizer expressionin wheat. J Mol Biol 204:523–534
Gerlach WL, Bedbrook JR (1979) Cloning and characterization ofribosomal RNA genes from wheat and barley. Nucl Acids Res7:1869–1885
Goodpasture C, Bloom SE (1975) Visualization of nucleolar organizerin mammalian chromosomes using silver staining. Chromosoma53:37–50
Gupta PK, Sharma PK, Balyan HS, Roy JK, Sharma S, Beharav A,Nevo E (2002) Polymorphism at rDNA loci in barley and itsrelation with climatic variables. Theor Appl Genet 104:473–481
Habu Y, Kakutani T, Paszkowski J (2001) Epigenetic developmentmechanisms in plants: molecules and targets of plant epigeneticregulation. Curr Opin Genet Dev 11:215–220
Hutchinson J, Miller TE (1982) The nucleolar organizers of tetraploidand hexaploid wheats revealed by in situ hybridization. TheorAppl Genet 61:285–288
Jiang J, Gill BS (1994) New 18S.26S ribosomal RNA gene loci:chromosomal landmarks for the evolution of polyploid wheats.Chromosoma 103:179–185
Lacadena JR, Cermeño MC, Orellana J, Santos JL (1988) Nucleolarcompetition in Triticeae. In: Brandham PE (ed) Kew ChromosomeConference III. HMSO, London, pp 151–165
Leitch IJ, Bennett MD (1997) Polyploidy in angiosperms. TrendsPlant Sci 2(12):470–476
Lima-Brito J, Guedes-Pinto H, Heslop-Harrison JS (1998) Theactivity of nucleolar organizing chromosomes in multigenericF1 hybrids involving wheat, triticale and tritordeum. Genome41:763–768
Liu B, Vega JM, Segal G, Abbo S, Rodova M, Feldman M (1998)Rapid changes in newly synthesized amphiploids of Triticum andAegilops. I. Changes in low-copy noncoding DNA sequences.Genome 41:272–277
Martini G, Flavell R (1985) The control of nucleolus volume in wheat,a genetic study at three developmental stages. Genetical Societyof Great Britain 54:11–120
Plant Mol Biol Rep (2010) 28:403–412 411

McClintock B (1934) The relationship of a particular chromosomalelement to the development of the nucleoli in Zea mays. ZZellforsch Mikrosk Anat 21:294–328
McStay B (2006) Nucleolar dominance: a model for rRNA genesilencing. Genes Dev 20:1207–1214
Mohan J, Flavell RB (1974) Ribosomal RNA cistron multiplicity andnucleolar organizers in hexaploid wheat. Genetics 76:33–44
Molnar SJ, Gupta PK, Fedak G, Wheatcroft R (1989) RibosomalDNA repeat unit polymorphism in 25 Hordeum species. TheorAppl Genet 78:387–392
Mukai Y, Endo TR, Gill BS (1991) Physical mapping of the 18S.26SrRNA multigene family in common wheat: identification of anew locus. Chromosoma 100:71–78
Mukai Y, Nakahara Y, Yamamoto M (1993) Simultaneous discrimi-nation of the three genomes in hexaploid wheat by multicolourfluorescence in situ hybridization using total genomic and highlyrepeated DNA probes. Genome 36:489–494
Navashin M (1934) Chromosomal alterations caused by hybridizationand their bearing upon certain general genetic problems. Cytologia5:169–203
Neves N, Heslop-Harrison JS, Viegas W (1995) rRNA gene activityand control of expression mediated by methylation and imprintingduring embryo development in wheat × rye hybrids. Theor ApplGenet 91:529–533
Neves N, Silva M, Heslop-Harrison JS, Viegas W (1997) Nucleolardominance in triticales: control by unlinked genes. ChromosomeRes 5:125–131
Ozkan H, Levy AA, Feldman M (2001) Allopolyploidy-induced rapidgenome evolution in the wheat (Aegilops-Triticum) group. PlantCell 13:1735–1747
Pardue ML, Gall JG (1969) Molecular hybridization of radioactiveDNA to the DNA of cytological preparations. Proc Natl Acad Sci64:600–604
Paszkowski J, Whitham SA (2001) Gene silencing and DNAmethylation processes. Current Opinions in Plant Biology 4:123–129
Polanco C, Pérez De La Vega M (1997) Intergenic ribosomal spacervariability in hexaploid oat cultivars and landraces. Heredity 78:115–123
Riddle NC, Richards EJ (2002) The control of natural variation incytosine methylation in Arabidopsis. Genetics 162:355–363
Riddle NC, Richards EJ (2005) Genetic variation in epgeneticinheritance of ribosomal RNA gene methylation in Arabidopsis.Plant J 41:524–532
Rocheford TR, Osterman JC, Gardner CO (1990) Variation in theribosomal DNA intergenic spacer of a maize population mass-selected for high grain yield. Theor Appl Genet 79:793–800
Saghai-Maroof MA, Soliman KM, Jorgensen RA, Allard RW (1984)Ribosomal DNA spacer length polymorphisms in barley:Mendelian inheritance, chromosomal location, and populationdynamics. Proc Natl Acad Sci 81:8014–8018
Sardana R, O’Dell M, Flavell RB (1993) Correlation between the sizeof the intergenic regulatory region, the status of cytosinemethylation of rRNA genes and nucleolar expression in wheat.Mol Gen Genet 236:155–162
Scheid OM, Afsar K, Paszkowski J (1998) Release of epigeneticsilencing by trans-acting mutations in Arabidopsis. Proc NatlAcad Sci 95:632–637
Schwarzacher T, Heslop-Harrison JS (2000) Practical in situ hybrid-ization. BIOS Scientific, Oxford, p 204
Shaked H, Kashkush HK, Ozkan H et al (2001) Sequence eliminationand cytosine methylation are rapid and reproducible responses ofthe genome to wide hybridization and allopolyploidy in wheat.Plant Cell 13:1749–1759
Sharma S, Beharav A, Balyan HS, Nevo E, Gupta PK (2004)Ribosomal DNA polymorphism and its association with geo-graphical and climatic variables in 27 wild barley populationsfrom Jordan. Plant Sci 166:467–477
Sharma S, Balyan HS, Gupta PK (2005) Adaptive methylation patternof ribosomal DNA in wild barley from Israel. Barley GeneticsNewsletter 35:27–35
Southern EM (1975) Detection of specific sequences among DNAfragments separated by gel electrophoresis. J Mol Biol 98:503–517
Stack S, Herikhoff L, Sherman J, Anderson L (1991) Staining plant cellswith silver. I. The salt nylon technique. Biotech Histochem 1:69–78
Viegas WS (1980) Modulation of rRNA gene content by chromosome5D in wheat. Genetica 51(3):233–239
Viegas W, Mello-Sampayo T (1975) Nucleolar organization in thegenus Triticum. Broteria, Ser Trimest Cien Nat 44:121–133
Vieira R, Queiroz A, Morais L, Barão A, Mello-Sampayo T, Viegas W(1990) 1R chromosome nucleolus organizer region activation by5-azacytidine in wheat × rye hybrids. Genome 33:707–712
412 Plant Mol Biol Rep (2010) 28:403–412












![Sitti Ghaiyah dan Anisa Nor Fitria [Energi Terbarukan]](https://static.fdokumen.com/doc/165x107/63169096c32ab5e46f0dd650/sitti-ghaiyah-dan-anisa-nor-fitria-energi-terbarukan.jpg)







