
Differential regulation of the yeast isozymes of pyruvate carboxylase and the locus of action of...
13
The International Journal of Biochemistry & Cell Biology 39 (2007) 1211–1223 Differential regulation of the yeast isozymes of pyruvate carboxylase and the locus of action of acetyl CoA Sarawut Jitrapakdee a , Abdussalam Adina-Zada b , Paul G. Besant b , Kathy H. Surinya c , W. Wallace Cleland d , John C. Wallace c , Paul V. Attwood b,∗ a Department of Biochemistry, Faculty of Science, Mahidol University, Bangkok 10400, Thailand b Biochemistry and Molecular Biology (M310), School of Biomedical, Biomolecular and Chemical Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia c School of Molecular Biosciences, University of Adelaide, Adelaide, SA 5005, Australia d Institute for Enzyme Research and Department of Biochemistry, University of Wisconsin-Madison, Madison, WI 53726, USA Received 15 January 2007; received in revised form 14 March 2007; accepted 22 March 2007 Available online 30 March 2007 Abstract Unlike other eukaryotes studied to date, yeast has two genes for pyruvate carboxylase coding for very similar, but not identical, isozymes (Pyc1 and Pyc2), both of which are located in the cytoplasm. We have found that there are marked differences in the kinetic properties of the isozymes potentially leading to differential regulation of Pyc1 and Pyc2 activity by both activators and substrates. For example, Pyc2 is only activated 3.7-fold by acetyl CoA, and 9.6-fold by NH 4 + , whilst the figures for Pyc1 are 16 and 14.6-fold, respectively. Pyc1 and Pyc2 display different allosteric properties with respect to acetyl CoA activation and aspartate inhibition, with Pyc1 showing a higher degree of cooperativity than Pyc2, even in the absence of aspartate. We have investigated the locus of action in the amino acid sequence of the isozymes of this activator by measuring its regulation of various chimeric constructs of the two isozymes. In this way, we conclude that the main locus of action of acetyl CoA lies in the N-terminal half of the enzyme, within the biotin-carboxylation domain, between amino acids 99 and 478 of Pyc1. © 2007 Elsevier Ltd. All rights reserved. Keywords: Pyruvate carboxylase; Isozyme; Enzyme kinetics; Regulation of enzyme activity; Biotin; Acetyl CoA activation; Chimera ∗ Corresponding author. Tel.: +61 8 6488 3329; fax: +61 8 6488 1148. E-mail addresses: [email protected] (S. Jitrapakdee), [email protected] (A. Adina-Zada), [email protected] (P.G. Besant), [email protected] (K.H. Surinya), [email protected] (W.W. Cleland), [email protected] (J.C. Wallace), [email protected] (P.V. Attwood). 1. Introduction Pyruvate carboxylase (EC 6.4.1.1) is a biotin- dependent enzyme that catalyses the carboxylation of pyruvate using bicarbonate as a substrate, with ATP cleavage used to drive the carboxylation of biotin by bicarbonate to form an enzyme–carboxybiotin inter- mediate which then carboxylates the pyruvate to form oxaloacetate (for reviews see Attwood, 1995; Attwood & Wallace, 2002; Jitrapakdee & Wallace, 1999). The enzyme has an anaplerotic role in that it supplies oxaloacetate for gluconeogenesis and to replenish tricar- boxylic acid cycle intermediates that have been removed 1357-2725/$ – see front matter © 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocel.2007.03.016
Transcript of Differential regulation of the yeast isozymes of pyruvate carboxylase and the locus of action of...

A
iksaitct©
K
pkcjp
1
The International Journal of Biochemistry & Cell Biology 39 (2007) 1211–1223
Differential regulation of the yeast isozymes of pyruvatecarboxylase and the locus of action of acetyl CoA
Sarawut Jitrapakdee a, Abdussalam Adina-Zada b, Paul G. Besant b,Kathy H. Surinya c, W. Wallace Cleland d, John C. Wallace c, Paul V. Attwood b,∗
a Department of Biochemistry, Faculty of Science, Mahidol University, Bangkok 10400, Thailandb Biochemistry and Molecular Biology (M310), School of Biomedical, Biomolecular and Chemical Sciences,
The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australiac School of Molecular Biosciences, University of Adelaide, Adelaide, SA 5005, Australia
d Institute for Enzyme Research and Department of Biochemistry, University of Wisconsin-Madison, Madison, WI 53726, USA
Received 15 January 2007; received in revised form 14 March 2007; accepted 22 March 2007Available online 30 March 2007
bstract
Unlike other eukaryotes studied to date, yeast has two genes for pyruvate carboxylase coding for very similar, but not identical,sozymes (Pyc1 and Pyc2), both of which are located in the cytoplasm. We have found that there are marked differences in theinetic properties of the isozymes potentially leading to differential regulation of Pyc1 and Pyc2 activity by both activators andubstrates. For example, Pyc2 is only activated 3.7-fold by acetyl CoA, and 9.6-fold by NH4
+, whilst the figures for Pyc1 are 16nd 14.6-fold, respectively. Pyc1 and Pyc2 display different allosteric properties with respect to acetyl CoA activation and aspartate
nhibition, with Pyc1 showing a higher degree of cooperativity than Pyc2, even in the absence of aspartate. We have investigatedhe locus of action in the amino acid sequence of the isozymes of this activator by measuring its regulation of various chimericonstructs of the two isozymes. In this way, we conclude that the main locus of action of acetyl CoA lies in the N-terminal half ofhe enzyme, within the biotin-carboxylation domain, between amino acids 99 and 478 of Pyc1.2007 Elsevier Ltd. All rights reserved.
eywords: Pyruvate carboxylase; Isozyme; Enzyme kinetics; Regulation of e
∗ Corresponding author. Tel.: +61 8 6488 3329;fax: +61 8 6488 1148.
E-mail addresses: [email protected] (S. Jitrapakdee),[email protected] (A. Adina-Zada),
[email protected] (P.G. Besant),[email protected] (K.H. Surinya),[email protected] (W.W. Cleland),[email protected] (J.C. Wallace),[email protected] (P.V. Attwood).
357-2725/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.doi:10.1016/j.biocel.2007.03.016
nzyme activity; Biotin; Acetyl CoA activation; Chimera
1. Introduction
Pyruvate carboxylase (EC 6.4.1.1) is a biotin-dependent enzyme that catalyses the carboxylation ofpyruvate using bicarbonate as a substrate, with ATPcleavage used to drive the carboxylation of biotin bybicarbonate to form an enzyme–carboxybiotin inter-mediate which then carboxylates the pyruvate to formoxaloacetate (for reviews see Attwood, 1995; Attwood
& Wallace, 2002; Jitrapakdee & Wallace, 1999). Theenzyme has an anaplerotic role in that it suppliesoxaloacetate for gluconeogenesis and to replenish tricar-boxylic acid cycle intermediates that have been removed
of Bioc
1212 S. Jitrapakdee et al. / The International Journalfor synthetic purposes. In vertebrates, the enzyme isprimarily regulated by the allosteric activator, acetylCoA, whereas in some microorganisms, acetyl CoA isnot regulatory (e.g. Aspergillus niger) whilst in othersthere is a complete dependence on its presence (e.g.Bacillus thermodenitrificans) (Wallace, 1985). In manymicroorganisms, aspartate is a negative regulator ofpyruvate carboxylase activity (Wallace, 1985). Monova-lent cations such as K+ and NH4
+ have also been shownto activate pyruvate carboxylase (Barden & Scrutton,1974; McClure, Lardy, & Kneifel, 1971), although thephysiological relevance of this regulation is not clear.
In S. cerevisiae there are two isozymes of pyruvatecarboxylase (Pyc1 and Pyc2), each encoded by a sepa-rate gene (Stucka, Dequin, Salmon, & Gancedo, 1991;Walker, Val, Rohde, Devenish, & Wallace, 1991). Bothisozymes are expressed in the cytoplasm, unlike thepyruvate carboxylase of higher eukaryotes, where theenzyme is found in the mitochondrial matrix (Rohde,Lim, & Wallace, 1991). The amino acid sequences of thetwo isozymes are 91% identical (Stucka et al., 1991; Val,Chapman-Smith, Walker, Cronan, & Wallace, 1995).Fig. 1 shows the major domain structure of Pyc1 inrelation to the amino acid sequence and shows thatthe most N-terminal of the domains is the ATP/HCO3-binding domain, where biotin carboxylation occurs.This is followed by the pyruvate-binding domain whereoxaloacetate formation occurs and then the biotinyldomain that contains Lys1112 to which biotin is cova-lently bonded.
The expression of the two isozymes is differentiallyregulated, depending on factors such as growth phase,carbon source and nitrogen source, especially ammo-nium ions (Brewster, Val, Walker, & Wallace, 1994;Huet, Menendez, Gancedo, & Francois, 2000). Thisdifferential regulation of gene expression and the co-localisation of Pyc1 and Pyc2 in the cytoplasm wouldsuggest that Pyc1 and Pyc2 have different kinetic or reg-
ulatory properties. All of the early characterisation ofpyruvate carboxylase from S. cerevisiae was performedon enzyme preparations from baker’s yeast, which poten-tially contained a mixture of both Pyc1 and Pyc2. TheFig. 1. Schematic representation of the primary structure of yeast pyru-vate carboxylase, showing regions of homology with other, relatedenzymes, based on the work of Lim, Morris, Occhiodoro, and Wallace(1988).
hemistry & Cell Biology 39 (2007) 1211–1223
kinetic parameters of Pyc1 in terms of substrate Kms havebeen recently determined (Branson, Nezic, Jitrapakdee,Wallace, & Attwood, 2004; Branson, Nezic, Wallace, &Attwood, 2002). However, the kinetics and regulation ofpurified Pyc2 have not been investigated.
In the current work, we have measured the Kms foreach of the substrates of Pyc2 and the dependence of itsactivity on acetyl CoA concentration, including the Ka.We compare the kinetic parameters of Pyc1 and Pyc2and their regulation by acetyl CoA, NH4
+ and aspar-tate. We discuss the differences in kinetic parametersand regulation of the two isozymes with respect to thedifferences in the regulation of their expression. In addi-tion, we have investigated the locus of action of acetylCoA in the amino acid sequences of the isozymes byexamining the regulation of various chimeric constructsof Pyc1 and Pyc2.
2. Materials and methods
2.1. Chemicals and reagents
The sodium salt of acetyl CoA was purchased fromSigma and all other reagents were of analytical quality.
2.2. Yeast strains and constructs
S. cerevisiae DM18 (MAT�, ura3, trp1, �pyc1:LEU2, �pyc2: HIS3) (Brewster et al., 1994) harbor-ing pVT100-PYC1 (Branson et al., 2002) was used asa source to prepare PYC1 isozyme.
The PYC2 construct was generated by PCR-amplification of PYC2 gene from S. cerevisiae W303chromosomal DNA. The gene was cloned from twooverlapping PCR fragments using two pairs of primerdesigned from the PYC2 gene sequence depositedin the Saccharomyces genome database (http://www.yeastgenome.org/). PYC2-F1 (5′-cccaagcttaagtaaaat-atgagcagtagcaag-3′, underline indicates HindIII restric-tion site, bold indicates an ATG initiation codon) andPYC2-R1 (5′-accaccggccttcttgacagcattc-3′) were syn-thesized to generate a 1.9 kb 5′-end of PYC2 gene whilstPYC2-F2 (5′-agtgctactggaaaagggaccatg-3′) and PYC2-R2 (5′-ccctctagattaacgagtaaaaattactttttttggggatgg-3′;underline indicates XbaI restriction site) were syn-thesized to generate a 1.8 kb 3′-end of PYC2 gene.These two fragments were sequenced and assembled bydigestion with HindIII/BamHI and with BamHI/XbaI,
respectively, and cloned into HindIII/XbaI-digestedpVT100u (Vernet, Dignard, & Thomas, 1987), pro-ducing pVT100-PYC2 construct. Pyc2 construct withN-terminal 9His tag was constructed by PCR-amplifying
of Bioc
agcaP(pomfN0gc9uwFwcpp
cPcPgbctcggtrrPrP
Fcso
S. Jitrapakdee et al. / The International Journal
0.8 kb 5′-end of Pyc1 gene using 9His F primer (5′-cccctcgagatg-caccaccaccatcatcaccaccatcacagcagtag-aagaaattggccggtctt-3′, bold indicates codons for 9His)nd Pyc2-R primer (5′-atagcgtcacgaacttcacgg-3′). ThisCR product was cloned into pDrive cloning systemQiagen) and sequenced. The insert was excised fromDrive vector by digesting with XhoI, located upstreamf an initiation codon and an AccI, which located in theiddle of PCR product, and replaced the corresponding
ragment in pVT100-PYC2. Pyc1 construct with-terminal 9His was generated by PCR-amplifying a.9 kb 5′-end of Pyc2 gene using 5′-His Pyc1 primer (5′-ccctcgagtaaaaataatgcatcatcatcaccaccatcatcatggctcg-aaagaaaattcgccggcttg-3′; bold indicates codons forHis) and PYC1-R (5′-tctgtgatggtatgttccacttgg-3′)sing pV100-PYC1 as template. This PCR productas cloned into pDrive cloning system and sequenced.inally, the insert was excised from pDrive by digestingith XhoI, locating at the most 5′-end of initiation
odon, and EcoRI, locating in the middle of the PCRroduct, and replaced the corresponding fragment inVT100-PYC1.
Six Pyc1/Pyc2 chimeras were generated as follows:himera 1 containing the N-terminal 616 residues ofyc1 and the C-terminal 562 residues of Pyc2 andhimera 2 containing the N-terminal 617 residues ofyc2 and the C-terminal 562 residues of Pyc1 wereenerated by utilizing a unique BamHI site presenceetween codons 616 and 617 of Pyc1, and betweenodons 617 and 618 of Pyc2 genes, allowing thesewo fragments to be swapped in frame. These twohimeras were constructed by digesting Pyc1 and Pyc2enes with XhoI, locating at the most 5′-end of theene and BamHI. These XhoI-BamHI fragments werehen replaced each other to form chimeras 1 and 2,espectively. Chimera 3 containing the N-terminal 437
esidues of Pyc1 and the C-terminal 741 residues ofyc2 and chimera 4 containing the N-terminal 438esidues of Pyc2 and the C-terminal 741 residues ofyc1 were generated by introducing a SacII site betweenig. 2. Reaction schemes for the binding of NH4+ to the isozymes or
himeras. In both schemes initial binding of a single NH4+ to each
ubunit results in enzyme activation, however the subsequent bindingf (i) one or (ii) two NH4
+ results in inhibition.
hemistry & Cell Biology 39 (2007) 1211–1223 1213
codons 437 and 438 of Pyc1 and between codons 438and 439 of Pyc2 genes, without altering their encodedamino acids and reading frames by site-directedmutagenesis. The mutagenic primers used to introduceSacII site in Pyc1 gene were PYC1/SacII-Forward(5′-aatcgagttcagaatccgcggtgtcaagaccaac-3′; underlineindicates SacII restriction site) and PYC1/SacII-Reverse (5′-gttggtcttgacaccgcggattctgaactcgatt-3′) andin Pyc2 gene were PYC2/SacII-Forward (5′-atcgaattca-gaatccgcggtgtgaagaccaac-3′) and PYC2-SacII-Reverse(5′-gttggtcttcacaccgcggattctgaattcgat-3′). These SacII-modified Pyc1 and Pyc2 constructs cloned in pBluescriptSK II plasmid (Stratagene) were then excised with XhoIlocating at the most 5′-end of both Pyc1 and Pyc2 genes,and SacII, and replaced each other to produce chimeras 3and 4, respectively. Chimera 5 containing the N-terminal95 residues of Pyc1 and the C-terminal 1083 residuesof Pyc2 and chimera 6 containing the N-terminal 96residues of Pyc2 and the C-terminal 1083 residues ofPyc1 were generated by introducing SalI site betweencodons 95 and 96 of Pyc1 and between codons 96 and97 of Pyc2 genes, without altering their encoded aminoacids and reading frames. The mutagenic primers usedto introduce a SalI site in Pyc1 gene were Pyc1-MluI-Forward (5′-ccaaaaacaccaggtcgacttcatccatccaggt-3′)and Pyc1-MluI-Reverse (5′-acctggatggatgaagtcgacc-tggtgtttttgg-3′) and for Pyc2 gene are Pyc2-MluI-Forward (5′-aaagaagcataaggtcgacttcatccatccaggt-3′)and Pyc2-MluI-Reverse (5′-acctggatggatgaagtcgacct-tatgcttcttt-3′). The SalI-modified Pyc1 and Pyc2 con-structs cloned in pBluescript SK II plasmid were thenexcised with XhoI and SalI and replaced each other toproduce chimeras 5 and 6, respectively. Finally, all ofthe above chimeric Pyc1-Pyc2 fragments were excisedfrom pBluescript and cloned into the equivalent sitesin pVT100u. The positions of various regions of Pyc1and Pyc2 being swapped to form these six chimerasare shown in Tables 2 and 3. All of these constructswere transformed into S. cerevisiae W303 where thepyc1 gene had been disrupted with a HIS3 cassette byhomologous recombination (Brachmann et al., 1998).
2.3. Preparation and purification of isozymes andchimeras
Preparation and purification of the untagged Pyc1 andPyc2 isozymes and the untagged chimeras was essen-tially as described by Branson et al. (2004) using anion
exchange chromatography on DEAE Sepharose CL-6Band affinity chromatography on avidin-Sepharose as alast purification step. The 9xHis-tagged chimeras werepurified by affinity chromatography on Ni-NTA resin,
of Bioc
1214 S. Jitrapakdee et al. / The International Journalusing a linear imidazole gradient (0–250 mM). Purifiedenzymes were stored at −80 ◦C in storage buffer com-prising 20% (v/v) glycerol in 0.1 M Tris–Cl, pH 7.8. Priorto assay, the enzyme was transferred to 0.1 M Tris–Cl, pH7.8 by centrifuging through Sephadex G-25 as describedby Helmerhorst and Stokes (1980).
2.4. Assays of pyruvate carboxylase activity
The conditions for this spectrophotometric assaywhere oxaloacetate formation is measured in a coupledreaction with malate dehydrogenase, were essentiallyas described by Attwood and Cleland (1986). One unitof enzyme activity is defined as the amount of enzymerequired to catalyze the formation of 1 �mol of oxaloac-etate min−1 at 30 ◦C. All assays were performed at30 ◦C. Assays were performed in 0.1 mM Tris–Cl, pH 7.8with the following concentrations of substrates unlessstated otherwise: 2.5 mM ATP; 20 mM NaHCO3; 7 mMMgCl2; 10 mM pyruvate; 0.21 mM NADH. Note thatin the experiment where [ATP] was varied, the con-centration of free Mg2+ was maintained at 4.5 mM andin all experiments where substrate concentrations werevaried the reactions were performed in the presence of0.25 mM acetyl CoA. Assays were initiated by the addi-tion of either isozymes or chimeras and activities werecalculated as units ml−1 of enzyme solution.
2.5. Data analysis
The kinetic parameters given in Table 1 were obtainedfrom non-linear regression fits of the Michaelis-Mentenequation to the data. The ammonium chloride acti-vation/inhibition data were analysed using non-linearleast-squares regression analysis to fit of the followingEqs. ((i) and (ii)) to the data:
v = (v0 + V [NH4Cl]/Ka)/
(1 + [NH4Cl](1 + [NH4Cl]/KI)/Ka) (i)
v = (v0 + V [NH4Cl]/Ka)/
(1 + [NH4Cl](1 + [NH4Cl]2/KI)/Ka) (ii)
Table 1Comparison of kinetic parameters of Pyc1 and Pyc2
Kinetic parameter Value of paramete(mM)b for Km or
Km for HCO3− 1.36 ± 0.12
Km for pyruvate 0.50 ± 0.06Km for MgATP 0.07 ± 0.01kcat (with saturating substrates and acetyl CoA) 60 ± 2
a Standard errors were obtained from the non-linear least-squares regressiob Values of kinetic parameters for Pyc1 are taken from Branson et al. (2004
hemistry & Cell Biology 39 (2007) 1211–1223
where v is the measured velocity of the reaction; v0 isthe measured velocity of the reaction in the absence ofNH4Cl; V is the velocity of the reaction if all of theenzyme subunits had a single NH4
+ bound (E·NH4, seeFig. 2); Ka is the activation constant for the binding ofNH4
+ to the enzyme (see Fig. 2); KI is the inhibitionconstant for the subsequent binding of NH4
+ (see Fig. 2)and has units of mM in (i) and mM2 in (ii).
Eqs. (i) and (ii) were derived based on the respectivereaction schemes shown in Fig. 2 with the assumptionsthat E·NH4 represents the maximally active form ofthe enzyme and that E·(NH4)2 and E·(NH4)3 have noenzymic activity.
The data for activation of the isozymes and chimerasby acetyl CoA, in the presence or absence of aspartatewere initially analysed by linear regression according tothe Hill Eq. (iii):
log(v/(Vmax − v)) = n log([acetyl CoA]) − log Ka
(iii)
where v is the velocity of the reaction at any acetylCoA concentration; Vmax is the velocity of the reac-tion at saturating acetyl CoA; n is the Hill coefficientof cooperativity; Ka is the activation constant.
In order to obtain an estimate of Ka with a standarderror, the data were re-analysed according to Eq. (iv):
(v/(Vmax − v)) = ([acetyl CoA]/Ka)n (iv)
where the value of n used was taken from the Hill plotanalysis.
3. Results and discussion
3.1. Kinetic parameters for the reaction catalysedby Pyc1 and Pyc2
The values of the K s for the substrates and k val-
m catues are shown in Table 1. The value of kcat in the presenceof saturating concentrations of acetyl CoA and the sub-strates of the reaction for Pyc2 is 78% of that for Pyc1indicating that Pyc2 is somewhat less catalytically activer for Pyc1 (±S.E.)a
(s−1)b for kcat
Value of parameter for Pyc2 (S.E.)a
(mM) for Km or (s−1) for kkcat
21.5 ± 1.80.79 ± 0.040.17 ± 0.02
47 ± 4
n analysis of the kinetic data.).

S. Jitrapakdee et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 1211–1223 1215
Fig. 3. The effects of NH4+ concentration on the activity of (a) Pyc1, (b) Pyc2, (c) chimera 1, and (d) chimera 2 in the absence of acetyl CoA.
R ng the ni s repreE
tseK
prrmmtc
PttA(wPteer
eactions were performed in 0.125 M Tris–Cl at pH 7.8 and 30 ◦C, usin the presence of different concentrations of NH4Cl. The dashed lineq. (ii).
han Pyc1. In addition, the Km for pyruvate of Pyc2 isomewhat higher than that of Pyc1. The largest differ-nces between Pyc1 and Pyc2 however, occur in theirm values for MgATP and HCO3
−.The Km for MgATP is 2.4-fold higher for Pyc2 com-
ared to that of Pyc1 and is closer to the value of 0.2 mMeported by Myers, Tolbert, and Utter (1983) for a prepa-ation of pyruvate carboxylase from S. cerevisiae, whichay have contained a mixture of Pyc1 and Pyc2. Thisay indicate that there is a greater degree of control of
he activity of Pyc2 than that of Pyc1 activity by theoncentration of ATP.
The Km for HCO3−, of Pyc2 is nearly 16 times that of
yc1. The value of the Km for HCO3− of Pyc2 lies closer
o the value reported by Myers et al. (1983) (19.1 mM) inhe presence of acetyl CoA than that measured by Ruiz-mil, de Torrontegui, Palacian, Catalina, and Losada
1965) (2.4 mM) for enzyme isolated from S. cerevisiae,hich in both cases may have contained a mixture ofyc1 and Pyc2. However, in the presence of 8.4 mM K+
he enzyme preparation studied by Myers et al. (1983)xhibited a Km for HCO3
− of 3.1 mM and in the pres-nce of a combination of acetyl CoA and K+, this Km waseduced further. Thus, in the yeast cell, where the con-
ormal components of the spectrophotometric assay (see Section 2.4),sent fits of Eq. (i) to the activity data, the solid lines represent fits of
centration of K+ is estimated to be about 17 mM (Myerset al., 1983) and in the presence of acetyl CoA, the Kmfor HCO3
− may be much closer to that measured forPyc1. When we repeated the kinetic experiments vary-ing [NaHCO3] in the presence of 17 mM KCl, we foundthat at concentrations of NaHCO3 above about 50 mM,inhibition was evident (data not shown). Estimates ofthe Kms obtained from a fit of an equation of the formsimilar to that of Eq (i) to the data were 4.1 ± 1.8 mMfor Pyc2 and 34 ± 10 mM for Pyc1. The inhibition wassomewhat similar to that seen with increasing concen-trations of NH4
+ (see below) suggests that there may beboth activatory and inhibitory effects of the monovalentcations (K+ and Na+) (McClure et al., 1971) that differbetween Pyc1 and Pyc2 and that make determination ofthe true Km for HCO3
− difficult.
3.2. Effect of NH4+on the activity of Pyc1, Pyc2
and chimeras 1 and 2
In view of the amino sequence similarity betweenPyc1 and Pyc2 (92.8% sequence identity), it seemedfeasible to prepare chimeras of Pyc1 and Pyc2 with-out incurring any great disruption of the structure or

of Bioc
1216 S. Jitrapakdee et al. / The International Journalfunction of the enzyme whilst enabling the investigationas to which domain(s) was associated with the regula-tory properties with respect to acetyl CoA and NH4
+.To perform an initial investigation, two chimeras wereconstructed, each containing roughly half the amino acidsequence of each isozyme (see Table 2). Both chimeraswere expressed as 9His-tagged recombinant proteins.
Fig. 3 shows the effects of increasing concentrationof NH4
+ on the activity of Pyc1, Pyc2 and chimeras1 and 2. Enzyme activity increases with [NH4
+] up toabout 75 mM. However, above this concentration, theNH4Cl becomes inhibitory. The estimated Kas and KIsfor NH4
+ and velocities in the absence of NH4+ are given
in Table 2. Both Pyc1 and chimera 1 are somewhat lesssensitive to activation by NH4
+, with their Kas beingabout 49% higher than those for Pyc2 and chimera 2,respectively. On the other hand, both Pyc2 and chimera2 are less sensitive to inhibition by NH4
+, with theirKIs being 2.5 and 4-fold higher than those of Pyc1 andchimera 1, respectively. Whilst the v0 values for Pyc1and chimera 1 are lower than those for Pyc2 and chimera2, respectively, the maximum velocity for Pyc1 is lowerthan that for Pyc2 whilst the reverse is the case for thechimeras.
Huet et al. (2000) found a four to five-fold up-regulation of the expression of Pyc1, but little change inPyc2 expression when the yeast was grown with ammo-nium as the nitrogen source compared to aspartate. Huetet al. (2000) offered the explanation that with NH4
+ asthe nitrogen source there would be a high demand for�-ketoglutarate as a substrate for glutamate dehydroge-nase which catalyses the formation of glutamine, usingNH4
+ as the other substrate. Pyruvate carboxylase activ-ity would therefore need to be increased to replenish�-ketoglutarate in the tricarboxylic acid cycle removedfor this purpose. One hypothesis that might explain the
difference in regulation of expression Pyc1 compared toPyc2 is that Pyc1 activity is more highly regulated bythe intracellular concentration of NH4+. However, ourdata do not really support this hypothesis, since the dif-
Table 2Analysis of effects of NH4Cl on Pycl, Pyc 1 and Chimeras 1 and 2
Isozyme/chimera Ka (mM)
Pycl 11.1 ± 0.6Pyc2 7.5 ± 0.8
Chimera 1 13.7 ± 17
Chimera 2 9.2 ± 1.0
Values of Ka, Kt and Vmax arc derived from the fits of Eq. (ii) to the data as shin the absence of NH4C1 as a percentage of Vmax.
hemistry & Cell Biology 39 (2007) 1211–1223
ferences in activation of the two isozymes by NH4+ are
relatively small and indeed the Ka of activation for Pyc1is higher than that for Pyc2.
We did find a larger difference between the isozymesin terms of their inhibition at high NH4Cl concentrations,with Pyc2 being less sensitive to inhibition. However, theconcentrations of NH4Cl at which this inhibition has apronounced effect on enzymic activity are unlikely tobe of physiological relevance and may be a salt con-centration effect rather than a specific effect of NH4
+
binding.In general, chimera 2 is more like Pyc2 in being some-
what more sensitive to activation and less sensitive toinhibition by NH4
+ than chimera 1, which is more likePyc1. In terms of the velocities in the absence of NH4
+
(v0) chimera 1 is more similar to Pyc1 in having a some-what lower v0 than chimera 2, however the maximumvelocity of chimera 1 is lower than that of chimera 2whilst the reverse is true for Pyc1 and Pyc2. Thus, in themajority of ways, the effects of NH4
+ on yeast pyruvatecarboxylase appear to be determined by the N-terminalhalf of the amino acid sequence, although the maxi-mum velocity may be influenced by the carboxy-terminalsequence.
3.3. Activation of Pyc1 and Pyc2 by acetyl CoA andthe effects of aspartate
Fig. 4a and b shows Hill plots of the activation ofPyc1 and Pyc2, respectively, by acetyl CoA in the pres-ence and absence of aspartate. Table 3 shows the Hillcoefficients of cooperativity calculated from these Hillplots, the Kas of activation by acetyl CoA in the absenceof aspartate and the velocities of the reaction catalysedby Pyc1 and Pyc2 in the absence of acetyl CoA as apercentage of those in the presence of saturating acetyl
CoA. In the absence of aspartate, the Hill coefficient forPyc2 is not significantly different from 1 indicating thatthere is that there is no cooperativity in the binding ofacetyl CoA. However, that for Pyc1 in the absence ofKI (mM2 × 10−4) Vmax (u mg−1) v0 (% Vmax)
7.8 ± 0.3 5.8 6.819.8 ± 1.7 7.9 10.4
4.5 ± 0.5 8.0 9.7
17.8 ± 1.5 3.8 11.6
own in Fig. 3. Values of v0 were obtained by expressing the velocity
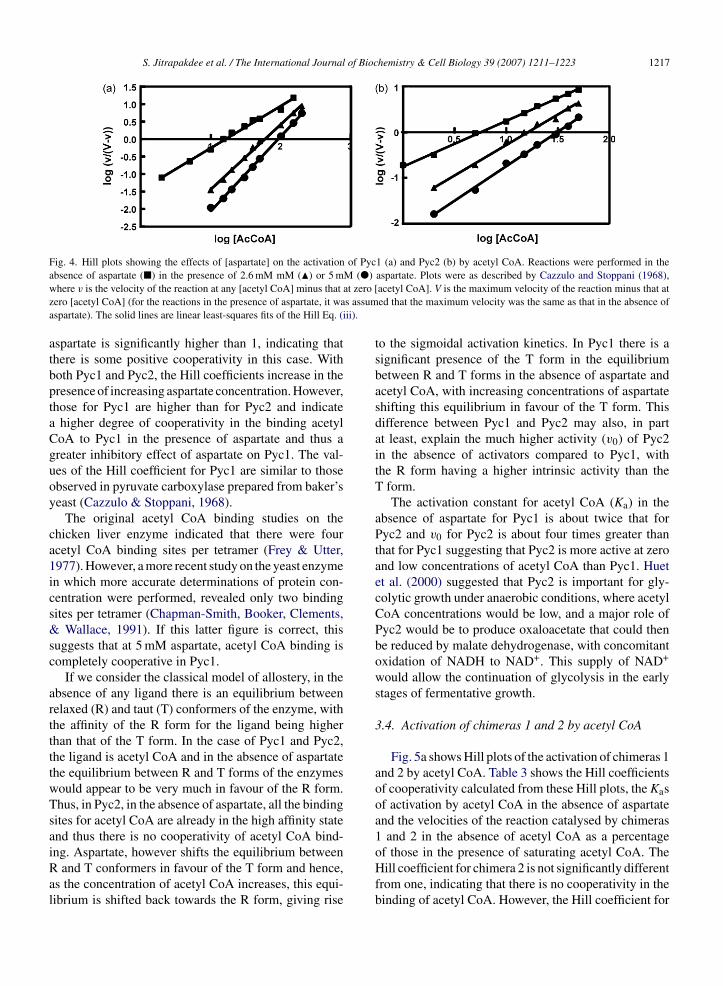
S. Jitrapakdee et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 1211–1223 1217
Fig. 4. Hill plots showing the effects of [aspartate] on the activation of Pyc1 (a) and Pyc2 (b) by acetyl CoA. Reactions were performed in thea M (�)w at zero [z s assuma ii).
atbptaCguoy
ca1ics&sc
arttttwTsaiRal
bsence of aspartate (�) in the presence of 2.6 mM mM (�) or 5 mhere v is the velocity of the reaction at any [acetyl CoA] minus that
ero [acetyl CoA] (for the reactions in the presence of aspartate, it waspartate). The solid lines are linear least-squares fits of the Hill Eq. (i
spartate is significantly higher than 1, indicating thathere is some positive cooperativity in this case. Withoth Pyc1 and Pyc2, the Hill coefficients increase in theresence of increasing aspartate concentration. However,hose for Pyc1 are higher than for Pyc2 and indicate
higher degree of cooperativity in the binding acetyloA to Pyc1 in the presence of aspartate and thus areater inhibitory effect of aspartate on Pyc1. The val-es of the Hill coefficient for Pyc1 are similar to thosebserved in pyruvate carboxylase prepared from baker’seast (Cazzulo & Stoppani, 1968).
The original acetyl CoA binding studies on thehicken liver enzyme indicated that there were fourcetyl CoA binding sites per tetramer (Frey & Utter,977). However, a more recent study on the yeast enzymen which more accurate determinations of protein con-entration were performed, revealed only two bindingites per tetramer (Chapman-Smith, Booker, Clements,
Wallace, 1991). If this latter figure is correct, thisuggests that at 5 mM aspartate, acetyl CoA binding isompletely cooperative in Pyc1.
If we consider the classical model of allostery, in thebsence of any ligand there is an equilibrium betweenelaxed (R) and taut (T) conformers of the enzyme, withhe affinity of the R form for the ligand being higherhan that of the T form. In the case of Pyc1 and Pyc2,he ligand is acetyl CoA and in the absence of aspartatehe equilibrium between R and T forms of the enzymesould appear to be very much in favour of the R form.hus, in Pyc2, in the absence of aspartate, all the bindingites for acetyl CoA are already in the high affinity statend thus there is no cooperativity of acetyl CoA bind-
ng. Aspartate, however shifts the equilibrium betweenand T conformers in favour of the T form and hence,s the concentration of acetyl CoA increases, this equi-ibrium is shifted back towards the R form, giving rise
aspartate. Plots were as described by Cazzulo and Stoppani (1968),acetyl CoA]. V is the maximum velocity of the reaction minus that ated that the maximum velocity was the same as that in the absence of
to the sigmoidal activation kinetics. In Pyc1 there is asignificant presence of the T form in the equilibriumbetween R and T forms in the absence of aspartate andacetyl CoA, with increasing concentrations of aspartateshifting this equilibrium in favour of the T form. Thisdifference between Pyc1 and Pyc2 may also, in partat least, explain the much higher activity (v0) of Pyc2in the absence of activators compared to Pyc1, withthe R form having a higher intrinsic activity than theT form.
The activation constant for acetyl CoA (Ka) in theabsence of aspartate for Pyc1 is about twice that forPyc2 and v0 for Pyc2 is about four times greater thanthat for Pyc1 suggesting that Pyc2 is more active at zeroand low concentrations of acetyl CoA than Pyc1. Huetet al. (2000) suggested that Pyc2 is important for gly-colytic growth under anaerobic conditions, where acetylCoA concentrations would be low, and a major role ofPyc2 would be to produce oxaloacetate that could thenbe reduced by malate dehydrogenase, with concomitantoxidation of NADH to NAD+. This supply of NAD+
would allow the continuation of glycolysis in the earlystages of fermentative growth.
3.4. Activation of chimeras 1 and 2 by acetyl CoA
Fig. 5a shows Hill plots of the activation of chimeras 1and 2 by acetyl CoA. Table 3 shows the Hill coefficientsof cooperativity calculated from these Hill plots, the Kasof activation by acetyl CoA in the absence of aspartateand the velocities of the reaction catalysed by chimeras1 and 2 in the absence of acetyl CoA as a percentage
of those in the presence of saturating acetyl CoA. TheHill coefficient for chimera 2 is not significantly differentfrom one, indicating that there is no cooperativity in thebinding of acetyl CoA. However, the Hill coefficient for
1218S.Jitrapakdee
etal./The
InternationalJournalofBiochem
istry&
CellB
iology39
(2007)1211–1223
Table 3Analysis of acetyl CoA activation of Pycl, Pyc2 and chimeras 1–6 and the effects of aspartate on this activation
Isozyme/chimeraa Ka (�M)b Vmax (u mg−1) v0 (% max vel)c n (0 mM Asp)d n (2.6 mM Asp)d n (5.0 mM Asp)d
Pycl 16.5 ± 0.6 8.3 6.1 1.20 ± 0.04 1.86 ± 0.04 2.13 ± 0.04
Pyc2 8.0 ± 1.7 4.0 26.7 0.99 ± 0.02 1.32 ± 0.04 1.51 ± 0.039HisPycl 6.9 ± 0.3 5.1 11.3 1.18 ± 0.04 ND ND
9His Pyc2 4.0 ± 0.2 2.1 20.0 0.98 ± 0.15 ND ND
Chimera 1 10.9 ± 0.7 4.5 10.5 1.12 ± 0.06 ND ND
Chimera 2 1.9 ± 0.1 2.3 22.2 0.94 ± 0.07 ND ND
Chimera 3 7.6 ± 0.4 1.4 15.2 1.18 ± 0.07 ND ND
Chimera 4 1.8 ± 0.1 0.3 22.3 1.07 ± 0.04 ND ND
Chimera 5 5.7 ± 0.4 0.06 17.1 1.03 ± 0.06 ND ND
Chimera 6 3.4 ± 0.1 1.3 11.9 1.37 ± 0.03 ND ND
ND is not determined.a Diagrams representing the polypeptide chains of the various constructs of Pycl, Pyc2 and the chimeras where the numbers indicate the amino acid position in Pycl at the boundary between the
Pycl and Pyc2 sequences.b Ka is the activation constant for acetyl CoA in the absence of aspartate ± standard error derived from the fit of Eq. (iv) to the data.c v0 is the velocity of the reaction in the absence of acetyl CoA (and aspartate) given as a percentage of the maximum velocity of the reaction at saturating [acetyl CoA].d n is the Hill coefficient of cooperativity ± standard error estimated by linear regression analysis of the Hill plot.

S. Jitrapakdee et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 1211–1223 1219
Fig. 5. Hill plots showing the activation of: (a) chimeras 1 (�) and 2 (�); (b) chimeras 3 (�) and 4 (�); (c) chimeras 5 (©) and 6 (�) by acetylC v is theC cetyl Cot e). The
ctrs
P2atarcCPaeilamwotta
oA. Plots were as described by Cazzulo and Stoppani (1968), whereoA]. V is the maximum velocity of the reaction minus that at zero [a
he maximum velocity was the same as that in the absence of aspartat
himera 1 is significantly higher than one, indicating thathere is some positive cooperativity in this case. In thisespect, chimera 1 is similar to Pyc1 whilst chimera 2 isimilar to Pyc2.
In terms of the values of v0, chimera 1 is similar toyc1 and chimera 2 is more like Pyc2. Similarly, chimerahas a lower activation constant (Ka) than chimera 1,
gain making it more like Pyc2 than Pyc1. However,he absolute values of Ka for both chimeras 1 and 2re 1.5 and 4-fold lower than those for Pyc1 and Pyc2,espectively. To investigate whether the His tag on thehimeras was affecting the activation constant for acetyloA, N-terminally 9His-tagged constructs of Pyc1 andyc2 were expressed and purified and their activation bycetyl CoA measured as for Pyc1 and Pyc2. The param-ters for the activation of these constructs are also shownn Table 3. Thus, the Ka for 9HisPyc1 was found to beess than that of Pyc1, suggesting that the 9His tag isffecting the activation of the enzyme by acetyl CoA, pri-arily by reducing the Ka. Similarly, the Ka for 9HisPyc2as reduced compared to Pyc2. In addition, the values
f Vmax of the 9His-tagged constructs were lower thanhose of the untagged isozymes and similar to those ofhe corresponding chimeras whilst the v0 and n valuesppear not to be markedly affected.velocity of the reaction at any [acetyl CoA] minus that at zero [acetylA] (for the reactions in the presence of aspartate, it was assumed that
solid lines are linear least-squares fits of the Hill Eq. (iii).
From the studies of chimeras 1 and 2 and the com-parisons with the Pyc1 and Pyc2 and their 9His-taggedforms, we can conclude that the major determinants ofthe response of the enzyme to acetyl CoA activationand the effects of NH4Cl lie in the N-terminal half ofthe amino acid sequence. This is in agreement with thestudy by Islam, Sueda, and Kondo (2005) who made achimera of the biotin-carboxylation subunit (472 aminoacids) of the Aquifex aeolicus enzyme and the C-terminalsequence of the B. thermodenitrificans enzyme. The A.aeolicus enzyme activity is independent of acetyl CoAand Islam et al. (2005) found that the activity of thechimera was also acetyl CoA-independent. From ourstudy, however, it is important to note the marked effectof the 9His tag on the Ka for acetyl CoA in both Pyc1and Pyc2 and illustrates the potential dangers of usingsuch tags which, although extremely useful for the pur-poses of enzyme purification, can have unforseen effectson enzyme action.
3.5. Further investigation of the locus of activation
of the Pyc isozymes by acetyl CoAIn order to further investigate the locus of action ofacetyl CoA, two more sets of chimeras were created

of Bioc
1220 S. Jitrapakdee et al. / The International Journal(see Table 3). As the 9His tag on both Pyc1 and Pyc2appeared to affect Ka for acetyl-CoA, the next sets ofchimeras were constructed without this N-terminal tag.In chimeras 3 and 4, the N-terminal section comprisesthe first 437 amino acids from Pyc1 and Pyc2, respec-tively. This N-terminal sequence encompasses the wholebiotin-carboxylation domain (see Fig. 1) and a sequenceof about 100 amino acids, N-terminal to this. In chimeras5 and 6, the N-terminal section comprises most of thisN-terminal sequence, i.e. the first 95 amino acids fromPyc1 and Pyc2, respectively.
Fig. 5b and c shows the Hill plots for activation ofthese chimeras by acetyl CoA and Table 3 shows param-eters of activation of these chimeras by acetyl CoA. Theparameters for chimeras 3 and 4 most resemble thoseof Pyc1 and Pyc2, respectively, with chimera 4 show-ing little cooperativity, having a higher v0 than chimera3 and a lower Ka than chimera 3. The absolute valuesof Ka for both chimeras are however, lower than thosefor Pyc1 (about 50%) and Pyc2 (about 23%) and the v0value for chimera 3 is 2.5 times that for Pyc1, whilst thatfor chimera 4 is similar to that of Pyc2.
The parameter values for chimeras 5 and 6 in Table 3,show that chimera 5 exhibits little cooperativity, whilstchimera 6 shows significant cooperativity. In addition,the v0 value for chimera 5 is larger than that for chimera6. In these respects chimera 6 behaves like Pyc1 andchimera 5 more like Pyc2. However, the Ka value forchimera 5 is still larger than that for chimera 6, althoughthere is less than a two-fold difference between them. TheKa value for chimera 6 is almost double that of chimera4 and 43% of that of Pyc2, whilst that of chimera 5is 75% of that of chimera 3 and 35% of that of Pyc1.The low Vmax values observed with chimeras 3–6 sug-gest that fusion of parts of the isozyme sequences hasintroduced some structural instability. This is especiallysevere in chimera 5, suggesting that there may be somestructurally important interactions in Pyc1 that have beenseverely disrupted in this chimera.
Analysis of the activation of chimeras 3 and 4 byacetyl CoA indicate that, whilst overall they behave ina similar manner to Pyc1 and Pyc2, respectively, thereare some marked differences. The Hill coefficients forchimeras 3 and 4 are essentially the same as for Pyc1and Pyc2, respectively, and the v0 value for chimera 4is similar to that of Pyc2, the v0 value for chimera 3 isconsiderably higher than for Pyc1. In addition, the val-ues of K for chimeras 3 and 4 are lower than for Pyc1
aand Pyc2, respectively. This suggests that there is part ofthe amino acid sequence between amino acids 437 and616 that contributes to the action of acetyl CoA on theenzyme. However, analysis of Pyc1 and Pyc2 sequenceshemistry & Cell Biology 39 (2007) 1211–1223
between these regions reveal only three amino acid dif-ferences, i.e. V491 in Pyc1 (I492 in Pyc2), A545 (S546)and R549 (K550) (see Fig. 6).
Chimera 5 shows essentially no cooperativity whilstchimera 6 exhibits positive cooperativity and the acetylCoA-independent activity of chimera 5 is greater thanthat of chimera 6. In these respects chimera 5 is behav-ing more like Pyc2 and chimera 6 is more like Pyc1.On the other hand, the Ka for chimeras 5 is larger thanthat for chimera 6, although that for chimera 6 is almosttwice that for chimera 4. Thus, the main determinantsfor cooperativity and acetyl-CoA-independent activityappear to lie somewhere between amino acids 95 and437. Analysis of Pyc1 and Pyc2 sequences shows 13amino acid differences across this region including N141
(H142 in Pyc2), K145 (R146), E162 (Q163), E285 (V286),A330 (S331), S340 (T341), P342 (T343), F347 (L348), A357
(S358), A368 (S369), I378 (L379), T400 (A401) and I401
(T402). These sequence differences between Pyc1 andPyc2 may form distinct structural motifs that mediatedifferential activation by acetyl-CoA.
Sueda, Islam, and Kondo (2004) found that there is aninteraction between the biotin-carboxylase domains inthe enzyme from B. thermodenitrificans and postulatedthat this was important for the formation of the tetramericquaternary structure. Since cooperativity is dependenton inter-subunit interactions, this is in agreement withour data that the basis for the cooperativity lies in thebiotin-carboxylation domain.
The situation is more complex with respect to theKa for acetyl CoA. The differences in the Ka valuesbetween all of the chimeras and the isozymes suggestthat determinants of Ka are more dispersed and thereis an interdependence between these determinants thataffects Ka. The fact that the Ka values of chimeras 5 and6 are still quite different from those of Pyc1 and Pyc2indicates that one or more of these determinants lie inthe N-terminal 95 amino acid sequence.
Fig. 6 shows a multiple sequence alignment betweenthe N-terminal halves of the amino acid sequences ofPyc1, Pyc2 and pyruvate carboxylase from A. niger,whose activity is completely independent of acetyl CoA(Feir & Suzuki, 1969). As can be seen, there are rel-atively few differences between Pyc1 and Pyc2 aloneand with the exception of the arginine in position 595 ofPyc1 where the corresponding residue in Pyc2 is alanine,the differences are fairly conservative in terms of aminoacid properties. Where there are differences between the
sequences of Pyc1 and Pyc2 that are not so conservative,these also coincide with different residues in the A. nigersequence. For example at the position corresponding to71 in Pyc1 there is a valine in Pyc1, a glutamate in Pyc2
S. Jitrapakdee et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 1211–1223 1221
Fig. 6. Multiple sequence alignment of the N-terminal halves of amino acid sequences of Pyc1 (PYC1 YEAST) (Lim et al., 1988), Pyc2(PYC2 YEAST) (Stucka et al., 1991) and pyruvate carboxylase from Aspergillus niger (PYC ASPNG) (Panneman, Ruijter, Van den Broeck,& Visser et al., 1998). The alignment was performed using the ClustalW program. The consensus line represents the consensus amino acid sequencewhere, at those positions, the amino acids in two or more of the enzymes are identical. Amino acids are given in each individual enzyme sequencewhen the amino acid at that position is different from the consensus or where there is no consensus, i.e. the amino acids at such a position aredifferent in all three sequences. The position numbers given above the consensus sequence refer to the Pyc1 sequence and indicate points in thealignment where there is either no consensus or there is a non-conservative difference in amino acids between Pyc1 and Pyc2.

of Bioc
1222 S. Jitrapakdee et al. / The International Journaland lysine in the A. niger enzyme. Thus, at this posi-tion, the range of amino acids varies from non-polar, toacidic to basic. There is a similar arrangement at posi-tion 88. At other positions there is a combination of polar,non-polar and basic or acidic residues (positions 1, 91,94, 285, 342 and 545) or aromatic, aliphatic and polar(positions 5 and 347). Thus, in N-terminal region of thesequence these amino acid differences are scattered. Thedifferences in the alignment at positions 285 and 342 ofPyc1 do lie within the span between positions 95 and437 and will be of interest for future site-directed muta-genesis studies aimed at investigating the basis of acetylCoA-independent activity and the cooperativity of acetylCoA activation.
3.6. Conclusions
In conclusion, we have shown a number of differencesbetween the kinetic properties of the isozymes Pyc1 andPyc2 from S. cerevisiae, which may have physiologicalrelevance in the differential control of their activities.However, in the current work all activation experimentswere performed in the presence of high substrate concen-trations and in the presence of a single activator and thuswe are essentially studying activator effects on Vmax. Inthe yeast cell there is likely to be a combination of theeffects of different activator concentrations and chang-ing substrate and inhibitor concentrations that contributeto the fine differential regulation of Pyc1 and Pyc2 activ-ities.
The main determinants of acetyl CoA activation ofthe enzyme lie in the N-terminal half of the aminoacid sequence, with those that affect the degree ofacetyl-CoA-independent activity and the cooperativityof acetyl CoA action being more localised to the biotin-carboxylation domain.
Acknowledgements
This work was supported in part by the AustralianResearch Council grant DP0346807 to J.C.W., theNational Institutes of Health grant 1 R01 GM070455 toW.W. Cleland., J.C.W. and P.V.A. and the Career Devel-opment Grant RSA460002 from the Thailand ResearchFund to S.J.
References
Attwood, P. V. (1995). The structure and the mechanism of action ofpyruvate carboxylase. International Journal of Biochemistry andCell Biology, 27, 231–249.
hemistry & Cell Biology 39 (2007) 1211–1223
Attwood, P. V., & Cleland, W. W. (1986). Decarboxylation ofoxalacetate by pyruvate carboxylase. Biochemistry, 25, 8191–8196.
Attwood, P. V., & Wallace, J. C. (2002). Chemical and catalytic mecha-nisms of carboxyl transfer reactions in biotin-dependent enzymes.Accounts of Chemical Research, 35, 113–120.
Barden, R. E., & Scrutton, M. C. (1974). Pyruvate carboxylase fromchicken liver. Effects of univalent and divalent cations on catalyticactivity. Journal of Biological Chemistry, 249, 4829–4838.
Brachmann, C. B., Davies, A., Cost, G. J., Caputo, E., Li, J., Hieter,P., et al. (1998). Designer Deletion Strains derived from Saccha-romyces cerevisiae S288C: a useful set of strains and plasmids forPCR-mediated gene disruption and other applications. Yeast, 14,115–132.
Branson, J. P., Nezic, M., Jitrapakdee, S., Wallace, J. C., & Attwood,P. V. (2004). Kinetic characterization of yeast pyruvate carboxy-lase isozyme Pyc1 and the Pyc1 mutant, C249A. Biochemistry, 43,1075–1081.
Branson, J. P., Nezic, M., Wallace, J. C., & Attwood, P. V. (2002).Kinetic characterization of yeast pyruvate carboxylase isozymepyc1. Biochemistry, 41, 4459–4466.
Brewster, N. K., Val, D. L., Walker, M. E., & Wallace, J. C. (1994).Regulation of pyruvate carboxylase isozyme (PYC1, PYC2) geneexpression in Saccharomyces cerevisiae during fermentative andnonfermentative growth. Archives of Biochemistry and Biophysics,311, 62–71.
Cazzulo, J. J., & Stoppani, A. O. M. (1968). The regulation ofyeast pyruvate carboxylase by acetyl-coenzyme A and l-aspartate.Biochimica et Biophysica Acta, 127, 563–567.
Chapman-Smith, A., Booker, G. W., Clements, P. R., & Wallace,J. C. (1991). Further studies on the localization of the reactivelysyl residue of pyruvate carboxylase. Biochemical Journal, 276,759–764.
Feir, H. A., & Suzuki, I. (1969). Pyruvate carboxylase of Aspergillusniger: kinetic study of a biotin-containing carboxylase. CanadianJournal of Biochemistry, 47, 697–710.
Frey, W. H., & Utter, M. F. (1977). Binding of acetyl CoA to chickenliver pyruvate carboxylase. Journal of Biological Chemistry, 252,51–56.
Helmerhorst, E., & Stokes, G. B. (1980). Microcentrifuge desalting: arapid, quantitative method for desalting small amounts of protein.Analytical Biochemistry, 104, 130–135.
Huet, C., Menendez, J., Gancedo, C., & Francois, J. M. (2000). Regula-tion of pyc1 encoding pyruvate carboxylase isozyme I by nitrogensources in Saccharomyces cerevisiae. European Journal of Bio-chemistry, 267, 6817–6823.
Islam, M. N., Sueda, S., & Kondo, H. (2005). Construction of newforms of pyruvate carboxylase to assess the allosteric regulationby acetyl CoA. Protein Engineering Design and Selection, 18, 71–78.
Jitrapakdee, S., & Wallace, J. C. (1999). Structure, function and regu-lation of pyruvate carboxylase. Biochemical Journal, 340, 1–16.
Lim, F., Morris, C. P., Occhiodoro, F., & Wallace, J. C. (1988).Sequence and domain structure of yeast pyruvate carboxylase.Journal of Biological Chemistry, 263, 11493–11497.
McClure, W. R., Lardy, H. A., & Kneifel, H. P. (1971). Rat liver pyru-vate carboxylase. I. Preparation, properties, and cation specificity.
Journal of Biological Chemistry, 246, 3569–3578.Myers, D. E., Tolbert, B., & Utter, M. F. (1983). Activation of yeastpyruvate carboxylase: interactions between acyl coenzyme A com-pounds, aspartate, and substrates of the reaction. Biochemistry, 22,5090–5096.

of Bioc
P
R
R
S
S. Jitrapakdee et al. / The International Journal
anneman, H., Ruijter, G. J. G., Van den Broeck, H. C., & Visser, J.(1998). Aspergillus niger pyruvate carboxylase. European Journalof Biochemistry, 258, 223–232.
ohde, M., Lim, F., & Wallace, J. C. (1991). Electron microscopiclocalization of pyruvate carboxylase in rat liver and Saccharomycescerevisiae by immunogold procedures. Archives of Biochemistryand Biophysics, 290, 197–201.
uiz-Amil, M., de Torrontegui, G., Palacian, E., Catalina, L., &Losada, M. (1965). Properties and function of yeast pyru-vate carboxylase. Journal of Biological Chemistry, 240, 3492–3845.
tucka, R., Dequin, S., Salmon, J. M., & Gancedo, C. (1991). DNAsequences in chromosomes II and VII code for pyruvate carboxy-lase isoenzymes in Saccharomyces cerevisiae: analysis of pyruvatecarboxylase-deficient strains. Molecular Genes and Genetics, 229,307–315.
hemistry & Cell Biology 39 (2007) 1211–1223 1223
Sueda, S., Islam, M. N., & Kondo, H. (2004). Protein engineeringof pyruvate carboxylase. European Journal of Biochemistry, 271,1391–1400.
Val, D. L., Chapman-Smith, A., Walker, M. E., Cronan, J. E., Jr., & Wal-lace, J. C. (1995). Polymorphism of the yeast pyruvate carboxylase2 gene and protein—effects on protein biotinylation. BiochemicalJournal, 312, 817–825.
Vernet, T., Dignard, D., & Thomas, D. Y. (1987). A family of yeastexpression vectors containing the phage f1 intergenic region. Gene,52, 225–233.
Walker, M. E., Val, D. L., Rohde, M., Devenish, R. J., & Wallace, J. C.
(1991). Biochemical and Biophysical Research Communications,176, 1210–1217.Wallace, J. C. (1985). Distribution and biological functions of pyruvatecarboxylase in nature. In D. B. Keech & J. C. Wallace (Eds.),Pyruvate carboxylase (pp. 5–64). Boca Raton: CRC Press.




















