
Structural and biochemical studies of alcohol dehydrogenase isozymes from Kluyveromyces lactis

Expl Agric. (2013), volume 49 (1), pp. 53–73 C© Cambridge University Press 2012
doi:10.1017/S0014479712001056
CHANGES IN ANTIOXIDANT ISOZYMES AS A BIOMARKERFOR CHARACTERIZING HIGH TEMPERATURE STRESS
TOLERANCE IN RICE (ORYZA SATIVA L.) SPIKELETS
By SMRUTI DAS†,‡, P. KRISHNAN†,‡,¶, MONALISA NAYAK‡ and
B. RAMAKRISHNAN§
†Division of Agricultural Physics, Indian Agricultural Research Institute, New Delhi 110012,
India, ‡Division of Plant Physiology, Central Rice Research Institute, Cuttack 753 006, Orissa,
India and §Division of Microbiology, Indian Agricultural Research Institute, New Delhi
110012, India
(Accepted 27 October 2012)
SUMMARY
High temperature stress at flowering can adversely affect rice yield, largely due to failure of fertilization.Oxidative damage can be a major reason inducing spikelet sterility in rice. In the present study, the effectof high temperatures on antioxidant metabolism in rice spikelets was characterised using nine differentgenotypes. Exposure to different temperatures at flowering stage revealed significant differences amongvarious antioxidant enzymes in spikelets, both quantitatively and qualitatively. Spikelets of susceptiblegenotypes withstood temperature stress of up to 35 ◦C, those of moderately tolerant between 35 ◦C and38 ◦C and those of tolerant genotypes up to 40 ◦C. Presence or absence, and changes in the isozymeintensities were consistent with alterations in their activities. Superoxide dismutase (SOD) isozymes II andIII were present after exposure at 30 ◦C and 35 ◦C, while SOD I appeared above 40 oC. Intensities ofcatalase isozymes I and III and the only isozyme of ascorbate peroxidase altered, while the only isozymeof guaical peroxidase and two (III and IV) of the four isozymes of catechol peroxidase disappeared afterhigh temperature exposure of 45 ◦C. Thus, this work provides an evidence of the role of antioxidantmetabolism in spikelets under high temperature stress conditions. Hence, changes in antioxidant isozymesin rice spikelets can be used as a biomarker for characterizing high temperature stress tolerance in ricespikelets.
I N T RO D U C T I O N
Climate change predictions indicate an increase in the surface air temperature from1.7 ◦C to 4.4 ◦C in the next few decades (Intergovernmental Panel on ClimateChange (IPCC), 2007). Higher temperatures may diminish the likely benefits ofincreasing atmospheric concentrations of carbon dioxide (CO2) on productivity insome of the rice-growing environments (Krishnan et al., 2007). Temperatures higherthan 35 ◦C cause injuries with varying intensity in rice, depending on developmentalstages. White leaf tips, chlorotic bands and blotches, white bands and specks on leaves,reduced tillering and reduced height may occur at vegetative stages (Krishnan et al.,2011, Yoshida et al., 1981). At reproductive stage, white spikelets, white panicles andreduced spikelet number can occur (Yoshida et al., 1981). Temperature coinciding
¶Corresponding author. Email: [email protected]

54 S M RU T I DA S et al.
with flowering determines fertility (Krishnan and Surya Rao, 2005) and spikeletswith floral organs, such as lodicules (two), stamens (six) and pistil (one), are mostsensitive to heat stress (Morita et al., 2004). The magnitude of thermal burden onspikelets is largely a function of their size and shape and their immediate environment,particularly the relative humidity (Wassmann et al., 2009), wind speed and radiation.The rice spikelets can poise the magnitude of thermal burden to some extent bytranspirational cooling (Sheehy et al., 1998). Spikelet tissue temperature can be abetter descriptor for high temperature stress damage than ambient air temperature(Jagadish et al., 2007). Floral tissue damage due to high temperature stress could bedue to certain combinations of temperature, relative humidity, radiation, relativehumidity and wind speed resulting in inadequate transpirational cooling. Undercontrolled environments, micro thermocouples can be used to estimate the spikelettissue temperature. Nevertheless, there exist many difficulties under field conditionsin making accurate measurements of spikelet tissue temperature using short focallength infrared thermometer or a thermal infrared camera due to canopy/paniclearchitecture, colour and hairiness of spikelets, and time of the day.
Reactive oxygen species (ROS) are produced during plant photosynthesis,photorespiration, respiration, flowering and other reactions of cellular metabolism(Zhao et al., 2011). Hydroxyl radicals (OH−), superoxide anion (O2
−), singlet oxygen(1O2) or less reactive hydrogen peroxide (H2O2
−) can be harmful to the overall plantmetabolic activity and damage membrane lipids, proteins, chlorophyll and nucleicacids. Under unfavourable environments, ROS production is increased and thisincrease is much higher due to high temperature stress (Djanaguiraman et al., 2011).Activation of antioxidant enzymes, such as superoxide dismutase (SOD), catalase(CAT), ascorbate peroxidase (APX) and peroxidase (POD), can help to scavengeROS. While SOD converts superoxide radicals (O2−) into hydrogen peroxide (H2O2),CAT dismutates H2O2 into water and oxygen, and APX by utilizing ascorbate as anelectron donor to reduce H2O2 to water, POD reduces H2O2 to water using varioussubstrates (e.g. phenolic compounds and/or antioxidants) as electron donors. Non-enzymatic constituents, such as lipid-soluble and membrane-associated tocophenols,water-soluble reductants, ascorbic acid and glutathione, are also involved in theantioxidant defence system of plants (Wahid et al., 2007). Generally, healthy plantsmaintain a balance between generating free radical oxygen species (ROS) and theirscavenging mechanisms to maintain growth and metabolism. Multiple molecularforms of antioxidant enzymes can occur within tissues, cells or organelles and theirchanges can imply their potential roles in detoxification of ROS. There exists arelationship between enhanced or constitutive antioxidant enzyme activities andincreased resistance to plant stress (Türkan et al., 2005).
Investigations on enzymatic scavenging mechanisms for ROS will help to definetheir involvement and provide new insights into high temperature tolerance inspikelets. We propose a hypothesis that higher activities of antioxidant enzymes,coupled with changes in their isozyme profiles in rice spikelets provide an importantmechanism of tolerance to high temperature stress. In the present work, the effectof high temperature stress on the activities of antioxidant enzymes like SOD (EC

High temperature stress effects on spikelets of rice 55
1.15.1.1), CAT (EC 1.11.1.6), APX (EC 1.11.1.11), POD (EC 1.11.1.7), guaicolperoxidase (GP) (EC 1.11.1.7) and catechol oxidase (CO) (EC 1.10.3.1), togetherwith their isozyme profiles, were studied in spikelets of nine different rice genotypes,grouped as tolerant, moderate and susceptible from their responses to temperatureafter an agronomic evaluation from field experiments (Supplementary Table 1).
M AT E R I A L S A N D M E T H O D S
Plant material and growth conditions
Rice genotypes. Seeds of rice genotypes (cv. N22, Annapurna and Khitish, consideredto be tolerant to high temperature stress, and those of highly susceptible such as,cv. Ramakrishna, ADT 43 and Basmati 370, along with moderately tolerant cv.Sasyashree, Prasad and Divya), were obtained from the Division of Crop Improvement,Central Rice Research Institute (CRRI), Cuttack, India. The seeds were sown ontotrays containing 2-mm sieved clay loam soil (2 kg per tray for each genotype), mixedthoroughly with (NH4)2SO4 (2.5 g), KCl (0.5 g) and single superphosphate (SSP) (0.5 g).Sterile water was used regularly to facilitate germination and growth of seedlings.After 15 d, three seedlings each for a particular genotype were transplanted into potscontaining clay loam soil (7.4 kg), mixed thoroughly with (NH4)2SO4 (7.5 g), KCl(1.5 g) and SSP (1.5 g). An additional dose of (NH4)2SO4 (2.5 g) was provided on25–30 d after transplanting. For each of the nine genotypes, 35 pots were maintainedseparately under flooded conditions throughout the crop cycle. Pots were maintainedunder natural sunlight conditions with the day/night temperature of 29 ± 0.9/21 ±0.6 ◦C and RH of 75% (SD = 2.95) in the greenhouse of the experimental farm ofCRRI, Cuttack.
Growth condition and high temperature treatment
On the first day of flowering (anthesis, i.e. the appearance of anthers), pots ofeach genotype were transferred at 08.00 h into controlled environment of the PlantGrowth Chambers (Cassia Siamia Ltd., Hyderabad, India). The temperatures (SDin parentheses) of the growth chambers were maintained at 25 (SD = 1.6), 30(1.4), 35 (1.2), 40 (1.5), 45 (1.7) and 50 (1.6) ◦C with an RH of 75% for six daysseparately in accordance with specific treatments for all the genotypes. Investigationswere performed after exposure of plants at the flowering stage to high temperaturesonly up to 50 ◦C. Temperature of the ambient air cycled from outside the plantgrowth chambers was increased with the help of heaters inside the chambers.Photosynthetic photon flux density was maintained at 640 μmol m−2 s−1 inside,while the concentration of CO2 was not measured. After temperature treatments, allthe plants were restored to the natural sunlit conditions (∼25–32 ◦C). There were nopest or disease problems within the greenhouse.
Sampling
Spikelet fertility of the control and high temperature-treated plants was testedon those which opened between 09.00 h and 15.00 h on the first day of anthesis,

56 S M RU T I DA S et al.
and labelled using acrylic marker (Jagadish et al., 2010). After observing for 12 d,fertility in percentage was scored by pressing the marked spikelets individually. Inan effort to estimate the activities of antioxidant enzymes and their isozyme profiles,fertile spikelets of all the plants after different levels of high temperature exposurewere collected between 08.00 h and 08.30 h on the following day. A minimum ofthree samples of spikelets (∼1500 spikelets) were collected with each sample froma minimum of three plants of a particular genotype and pooled. All the sampledspikelets were frozen in liquid nitrogen, extracted immediately and the supernatantwas collected for analysis.
Extraction and estimation of antioxidant enzyme activities
The antioxidant enzymes were extracted from the frozen spikelets by grindingthe tissue using a pestle and mortar with liquid nitrogen and 25% (w/w) polyvinylpolypyrrolidone (PVPP). After grinding, the powdered tissue (0.5 g ∼300 spikelets)was then extracted with 10 mL of ice cold potassium phosphate buffer (50 mM, pH7.8) containing EDTA (1 mM), ascorbate (1 mM), sorbitol (10% w/v) and Triton X100 (0.1). The extract was centrifuged twice for 10 min at 15,000 rpm at 4 ◦C. Thesupernatant as enzyme extract was then used for all the assays.
Superoxide dismutase (SOD) assay
The SOD assay was performed according to the photochemical method(Chowdhury and Chowdhury, 1985). The reaction mixture (3 mL) contained 2.4 mLof 50 mM sodium phosphate (Na2PO4) buffer (pH 7.8) containing 0.1 mM EDTA,63 μM nitroblue tetrazolium chloride (NBT), 13 μM L-methionine and 0.2 mL ofenzyme extract and 0.5 mL riboflavin (1.3 μM), which was added last. The reactionwas monitored in the presence of 40 W fluorescent lamp for 20 min. One unit of SODactivity is defined as the amount of enzymes required to cause 50% inhibition of therate of NBT reduction at 560 nm and was expressed as unit per gram of fresh weight.
Catalase (CAT) assay
The assay mixture contained 2.3 mL of 50 mM Na2PO4 buffer (pH 7.0), 0.5 mLof 10 mM H2O2 and 0.2 mL of enzyme extract. The decomposition of H2O2 wasmonitored at 240 nm for 3 min (Cakmak and Marschner, 1992). The activity isexpressed as change in Optical Density (OD) per minute per gram fresh weight.
Ascorbate peroxidase (APX) assay
The reaction mixture contained 1.8 mL of 50 mM Na2PO4 buffer (pH 7.0), 0.5 mLof 100 mM H2O2, 0.5 mL of 0.5 mM ascorbic acid and 0.2 mL of enzyme aliquot.The assay was allowed to equilibrate at 25 ◦C for 1 min before the addition of H2O2,which initiated the reaction. The reaction was monitored as decrease in absorbanceat 290 nm for 1.0 min at 25 ◦C after the determination of initial rate. A controlreaction was prepared by replacing ascorbate with HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) buffer. A unit of APX is defined as the amount necessary

High temperature stress effects on spikelets of rice 57
to oxidize 1.0 μmol of ascorbate per min at 25 ◦C (290 nm extinction coefficients of2.8 mm−1 cm−1) (Nakano and Asada, 1981).
Guaiacol peroxidase (GP) assay
The activities were measured as oxidation of guaiacol (8.26 mM, ε = 26.6 mM− 1
cm−1) in the presence of H2O2 (8.8 mM) within 2 min (Rao et al., 1996). Thesupernatant (0.2 mL) was reacted with 1.8 mL of 50 mM Na2PO4 buffer (pH 7.0)and 0.5 mL of 0.1 mM H2O2. The reaction was started with addition of 0.5 mL of0.1 mM guaicol and followed in a spectrophotometer (Specord 200, Analytik Jena,Germany). The enzyme activity is measured by increase in absorbance at 470 nmcaused by guaicol oxidation (E = 26.6 mM per cm), and the activity is expressed inmillimolar guaicol oxidized per minute per gram fresh weight.
Catechol oxidase (CO) assay
Catechol (or polyphenol) oxidase (CO) was measured by monitoring the change incolour intensity by oxidation of catechol at 420 nm. The reaction mixture contained1.8 mL of 50 mM Na2PO4 buffer (pH 7.0), 0.5 mL of 0.5% (w/v) catechol, 0.5 mLof 0.1 mM H2O2 and 0.2 mL of enzyme aliquot. The enzyme activity is measuredby increase in absorbance at 420 nm caused by catechol oxidation and expressed aschange in optical density per minute per gram fresh weight.
Isozyme profiling
Native polyacrylamide gel electrophoresis (PAGE) was performed on the crudeextract using the stacking gel (with 4.5% acrylamide) and the separating gel (with10% acrylamide) with running buffer composed of 4 mM Tris-HCl (pH 8.8) andglycine (38 mM). The crude extract with a protein content of 100 μg was loadedper well. The electrophoresis was performed at 4 ◦C with the application of 30 mAcurrent.
Superoxide dismutase (SOD)
Isozyme profiling of SOD was done by incubating the gel in the solution containing100 mL of 50 mM sodium phosphate buffer (pH 7.5) containing 200 mg of NBT indark at 30 ◦C for 20 min. The solution was then drained off and incubated in anothersolution containing 50 mM Na-Phosphate buffer (pH 7.5), 0.4 mL TEMED and 1 mgriboflavin for about 15 min under 40 W fluorescent tubes (two in number) at roomtemperature for development of achromatic bands.
Catalase (CAT)
For the isozymes of CAT, the polyacrylamide gel was incubated in the solutioncontaining H2O2 (0.1% v/v) for 5 min. Then the solution was drained off and the gelwas placed in a solution containing potassium ferricyanide (0.5%) and ferric chloride(0.5%) solutions for 10 min. The zymogram was recorded as soon as the achromaticbands became evident.

58 S M RU T I DA S et al.
Ascorbic acid peroxidase (APX)
Isozyme profiles of APX were obtained by incubating the gel in a solution of 50 mMpotassium phosphate buffer containing 4 mM ascorbic acid and H2O2 (0.1 mM) for15 min, then washing it with distilled water and incubating in a solution of 0.125 MHCl containing potassium ferricyanide (0.1%) and ferric chloride (0.1%) for 10 min.APX was located as achromatic bands on coloured background, as a result of thereaction between ferric chloride and potassium ferrocyanide, the latter having beenproduced by the reduction of potassium ferricyanide with un-reacted ascorbic acid.
Guaicol peroxidase (GP)
For determining the isozymes of GP, the polyacrylamide gel was prepared with10% separating- and 4.5% stacking-gel. After electrophoresis the gel was stained witha solution containing 0.5 mM guaicol and 2 mL of 30% (w/v) H2O2 for about 5–10 min. The brownish colour bands appeared in the gel after washing with distilledwater.
Catechol oxidase (CO)
Isozymes of CO were stained with a solution containing 0.1% catechol and 2 mLof 30% H2O2 for 5–10 min. The brownish colour bands appeared in the gel afterwashing with distilled water.
All the gels were scanned using Densitometer FlorchemTM 5500 (Alpha Innotech,California, USA). The gels were analysed for densitometric values using UVI Techsoftware (St. John’s Innovation Center, Cambridge, UK).
Yield and its components
For analyses of yield components and grain quality parameters, five replicate plantsfor each genotype during harvest stage were selected. After measuring the height,plants were harvested carefully along with the soil and washed to remove adheringsoil. Plants were then divided into culms and leaf sheaths, leaf blades and panicles; thenumbers of tillers and panicles were determined. To minimize wilting and respirationduring separation of tillers and other plant parts, the samples were kept in ice-coldwater. Immediately after processing, all samples were placed in an oven at 60 ◦C. Thedry weight of each plant part was determined after a constant weight was attained.Panicles were hand-threshed, and the yield of filled grains was weighed after drying at60 ◦C for 12 h. The 1000-grain weight for each rice genotype was estimated as a meanof three replicate samples, one from each replicate of five plants. Spikelet sterility wasmeasured as the number of filled spikelets and expressed as a proportion of the totalnumber of spikelets (Krishnan and Surya Rao, 2005). In the present experiment, thetemperature treatments above 45 ◦C invariably led to drastic reduction in yield in allthe genotypes. Although observations on different yield components were made for allthe temperature treatments, data from two temperature treatments (25 ◦C and 45 ◦Cfor six days) were only presented.

High temperature stress effects on spikelets of rice 59
Grain quality parameters
Another set of five plants in each replicate of a particular genotype was sampledand their panicles were sun-dried and hand-threshed. The grains from each replicatesamples were dried in an oven at 30 ◦C to constant moisture content of 10%, and asub-sample (50 g) of seeds from these was immediately stored in sealed and laminatedaluminium foil packets at 1–2 ◦C until they were tested for grain quality parameters.
Membrane stability index (MSI)
The MSI was measured by an electrolytic leakage technique. Briefly, three weighedreplicates of rice spikelets (2 g ∼100 spikelets) from each treatment were soaked in50 mL of distilled water at 25 ◦C for 16 h. The cell membrane thermal stability(CMTS) was estimated using the following equation:
CMTS = 1 – (1 – (T1/T2))/(1 – (C1/C2)),
where T and C refer to conductivity in the control and high temperature treatedsamples and subscripts 1 and 2 refer to conductance before and after autoclavingrespectively. A control with distilled water (but without seeds) was also maintained.Membrane Thermal Stability (MSI) was expressed using the following equation:
MSI (%) = 100 – CMTS.
Protein concentration
For the estimation of protein concentration, grain sample (0.5 g) was maceratedwith 5 mL of 0.05 M Tris HCl buffer (pH 7.2) and transferred to a centrifuge tube.The extract was then centrifuged at 1200× g at 4 ◦C for 15 min. The supernatantwas immediately used for the estimation of protein (Das et al., 2010).
Concentrations of sugar and starch
Dried grain sample (50 mg) was extracted in 10 mL of 80 percent ethanol forextraction of sugar and starch estimation. A glass ball was placed on top of the tubeand kept in a water bath at 80 to 85 ◦C for 30 min. Cooled solution was centrifugedand decanted into a 50 mL beaker. This extraction was repeated three more times.The alcohol extract was evaporated on a water bath at 80–85 ◦C until most of thealcohol was removed (e.g. volume was reduced to about 3 mL) and made up to 25 mLwith distilled water. While the concentration of sugar in the extract was analyzed, theresidue left after sugar estimation was used for starch estimation (Das et al., 2010).
Statistical analyses
All the experiments were repeated three times (three technical replicate experimentswith three biological replicates for each sample) for ensuring reproducibility of results.Significant differences between mean values of the observed parameters of controland temperature treatments of spikelets of individual genotype were evaluated bytwo-factor factorial analysis of variance. Analysis of variance was performed with theIRRISTAT Random Effect Model procedure (International Rice Research Institute,Los Banos, Philippines). Data on sterility (as percentage) and membrane stability

60 S M RU T I DA S et al.
index were transformed to arcsin values prior to analysis. Means differing significantlywere compared using Duncan’s Multiple Range Test (DMRT) at p ≤ 0.05. Pearsoncorrelation matrices were calculated across all environments for each genotype pooled,and their homogeneity was determined. The significance of individual correlationcoefficients was evaluated (Gomez and Gomez, 1984).
R E S U LT S
Antioxidant enzyme activities
Antioxidant enzyme activities (SOD, CAT, APX, GP and CO) in spikelets amongdifferent genotypes did not significantly differ at the exposure to the lowest (25 ◦C) aswell as the highest temperature (50 ◦C) (Figure 1). Increasing temperature from 25 ◦Cto 40 ◦C at anthesis led to significant increase in the activities of antioxidant enzymesin all the genotypes. While maximum activities of different enzymes were observedbetween 35 ◦C and 40 ◦C, there was concomitant decrease with further increasein temperature. Among the genotypes examined, cv. N22, Annapurna and Khitishshowed maximum activities at 40 ◦C, while cv. ADT 43, Ramakrishna and Basmati370 had their maximum activities at 35 ◦C. Rapid decrease in enzyme activities attemperatures above 40 ◦C was also a characteristic of genotypes tested.
Expression of antioxidant isozymes in response to different temperatures
Antioxidant isozyme profiling revealed temperature-dependent changes in all thegenotypes examined. In addition to the highly active isozymes, which were detectedin the spikelets of the temperature-treated plants, there appeared to be high specificitypatterns. The SOD isozymes II and III initially appeared under low temperatures(25 ◦C), while the isozyme-I was observed after the 40 ◦C treatment (Figure 2). Afterthe treatment of 50 ◦C, the isozymes I and II disappeared in all the genotypes.Three CAT isozymes, with CAT I and III being major isozymes, were detected uponseparation by the non-denaturing PAGE, followed by staining (Figure 3). There wasonly one isozyme of APX in all the nine rice genotypes after the exposure of 25, 30,35, 40, 45, 50 and 55 ◦C for six days (Figure 4). The most significant enhancementof GP was observed at 40 ◦C, where two isoenzymes of GP preserved their activities(Figure 5). However, there was only one isozyme of GP at all other temperaturetreatments. There were about four isozymes of CO in these rice genotypes (Figure 6).The isozyme I of CO was present at all temperature treatments, the isozyme II waspresent after the 30 ◦C treatment, and isozymes III and IV were present only at 35 ◦Ctreatment. However, the CO isozymes III and IV disappeared after 35 ◦C treatment;isozyme II disappeared at 45 ◦C and only isozyme I was present after 50 ◦C treatment.
Relative quantification of isozymes and their changes
The relative quantification of isozymes was performed using densitometricvalues, which represented band intensities. Antioxidant enzymes, such as SOD(Supplementary Table 2) and CAT (Supplementary Table 3), showed three, APX
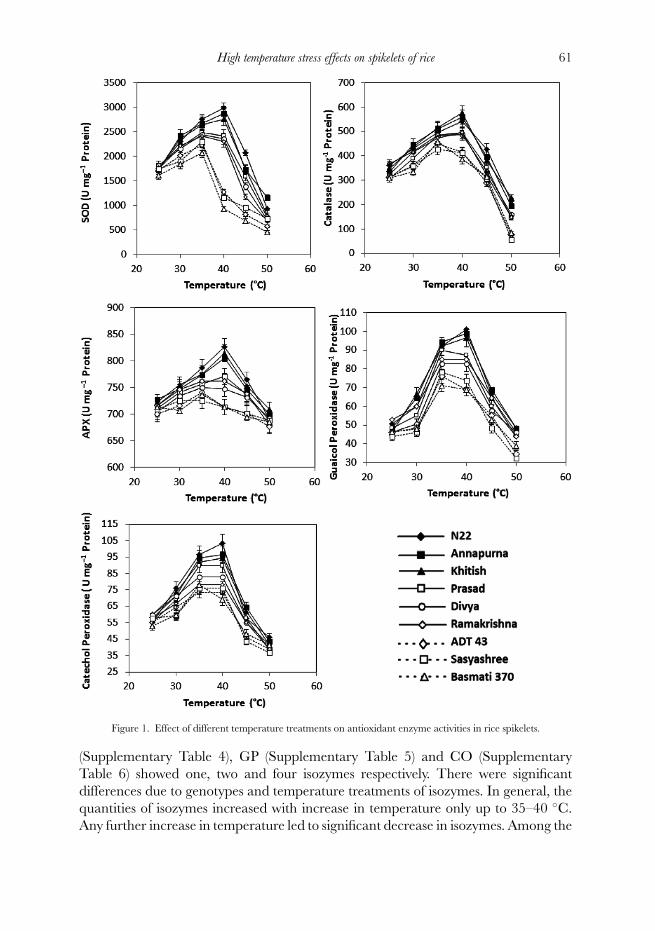
High temperature stress effects on spikelets of rice 61
Figure 1. Effect of different temperature treatments on antioxidant enzyme activities in rice spikelets.
(Supplementary Table 4), GP (Supplementary Table 5) and CO (SupplementaryTable 6) showed one, two and four isozymes respectively. There were significantdifferences due to genotypes and temperature treatments of isozymes. In general, thequantities of isozymes increased with increase in temperature only up to 35–40 ◦C.Any further increase in temperature led to significant decrease in isozymes. Among the

62 S M RU T I DA S et al.
(a)
25 oC 30 oC 35 oC C C 40 o 45 o 50 oC (b)
Densitometric scans
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
Genotype Isozyme II III
Isozyme I II III
Isozyme I II III
Isozyme I II III
Isozyme III
Isozyme III
1. N22
2. Annapurna
3. Khitish
4. Sasyashree
5. Prasad
6. Divya
7. Ramakrishna
8. ADT 43
9. Basmati 370
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
Figure 2. (Colour online) (a) Native PAGE analysis, and (b) densitometric scans of native PAGE electrophoretic patternof SOD enzyme.
nine rice genotypes tested, cv. N22, Annapurna and Khitish showed higher amountsof isozymes at 35–40 ◦C, while the genotypes, such as Sasyashree, Prasad and Divya,had only moderate increase in their isozymes. In contrast, there were drastic decreasein the isozymes of cv. Ramakrishna, ADT 43 and Basmati 370 after the temperaturetreatment beyond 35 ◦C. There were no significant differences in the amounts ofisozymes of all the genotypes both at 25 ◦C and 55 ◦C. Genotypes of cv. Ramakrishna,ADT 43 and Basmati 370 invariably had lower amounts of isozymes at all temperaturetreatments.

High temperature stress effects on spikelets of rice 63
(a)
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC (b)
Densitometric scan
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
Genotype Isozyme I II III
Isozyme I II III
Isozyme I II III
Isozyme I III
Isozyme I III
Isozyme I III
1. N22
2. Annapurna
3. Khitish
4. Sasyashree
5. Prasad
6. Divya
7. Ramakrishna
8. ADT 43
9. Basmati 370
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
Figure 3. (Colour online) (a) Native PAGE electrophoretic pattern of catalase enzyme, and (b) densitometric scans ofnative PAGE electrophoretic pattern of catalase enzyme.
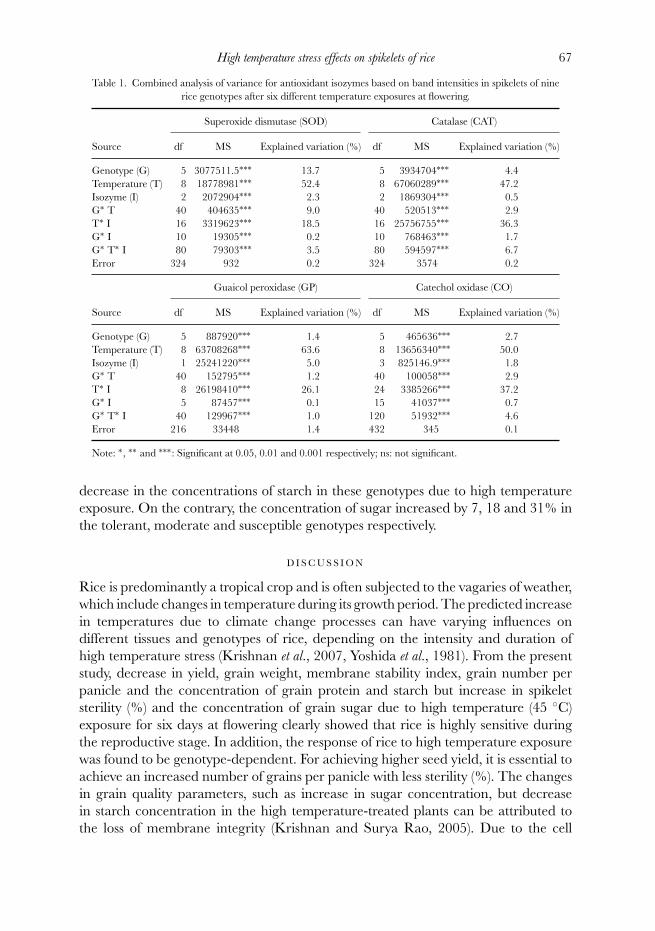
Analysis of variance
The interaction mean square (MS) value was less compared with that of main effects(Table 1). The value of the main effect MS for the proportion of antioxidant enzymesof SOD, CAT, APX, GP and CO was higher due to the temperature treatment or thegenotypes tested. The changes in the relative quantities of isozymes of SOD, CAT,GP and CO were found to be influenced more by the temperature treatment than thegenotype tested. Only in the case of APX, the influence due to genotype was found tobe more than that of high temperature treatment. In general, the explained variation(%) due to the temperature treatment was found to be maximum, being 52.5, 47.2,

64 S M RU T I DA S et al.
(a)
30 oC 35 oC 40 oC 45 oC C 50 o 55oC
(b)
Densitometric scans
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
Genotype Isozyme I
Isozyme I
Isozyme I
Isozyme I
Isozyme I
Isozyme I
1. N22
2. Annapurna
3. Khitish
4. Sasyashree
5. Prasad
6. Divya
7. Ramakrishna
8. ADT 43
9. Basmati 370
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 12 3 4 5 6 7 8 9
Figure 4. (Colour online) (a) Native PAGE electrophoretic pattern of APX enzyme, and (b) densitometric scans ofnative PAGE electrophoretic pattern of APX enzyme.
63.6 and 50.0% for the SOD, CAT, GP and CO respectively. Only the APX showedthe maximum explained variation of 87.8% due to the genotypes.
Yield attributes and grain quality
Exposure of rice plants to a high temperature of 45 ◦C at flowering for six days ledto significant reduction in yield per plant, while there were only marginal reductionsin the 1000 grain weight of all the genotypes tested (Table 2). In contrast, there were

High temperature stress effects on spikelets of rice 65
(a)
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
(b)
Genotype
Densitometric scans
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
Isozyme I
Isozyme I
Isozyme I II
Isozyme I
Isozyme I
Isozyme I
1. N22
2. Annapurna
3. Khitish
4. Sasyashree
5. Prasad
6. Divya
7. Ramakrishna
8. ADT 43
9. Basmati 370
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 12 3 4 5 6 7 8 9
Figure 5. (Colour online) (a) Native PAGE electrophoretic pattern of guaicol peroxidase enzyme, and (b) densitometricscans of native PAGE electrophoretic pattern of guaicol peroxidase enzyme.
two- to three-fold increase in spikelet sterility (%), especially in the genotypes such asSasyashree, Prasad, Divya, Ramakrishna, ADT 43 and Basmati 370. Only marginalincrease was observed in the tolerant genotypes (N22, Annapurna and Khitish). Inadditional, the number of grains per panicle of high temperature (45 ◦C)-treatedplants of tolerant genotypes was comparable to 25 ◦C. Interestingly, decrease in themembrane stability indices (MSI) was also indicative of agronomic grouping based on

66 S M RU T I DA S et al.
(a)
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
(b)
Densitometric scans
25 oC 30 oC 35 oC 40 oC 45 oC 50 oC
Genotype Isozyme I
Isozyme I II
Isozyme I II III IV
Isozyme I II
Isozyme I
Isozyme I
1. N22
2. Annapurna
3. Khitish
4. Sasyashree
5. Prasad
6. Divya
7. Ramakrishna
8. ADT 43
9. Basmati 370
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
Figure 6. (Colour online) (a) Native PAGE electrophoretic pattern of catechol oxidase enzyme, and (b) densitometricscans of native PAGE electrophoretic pattern of catechol oxidase enzyme.
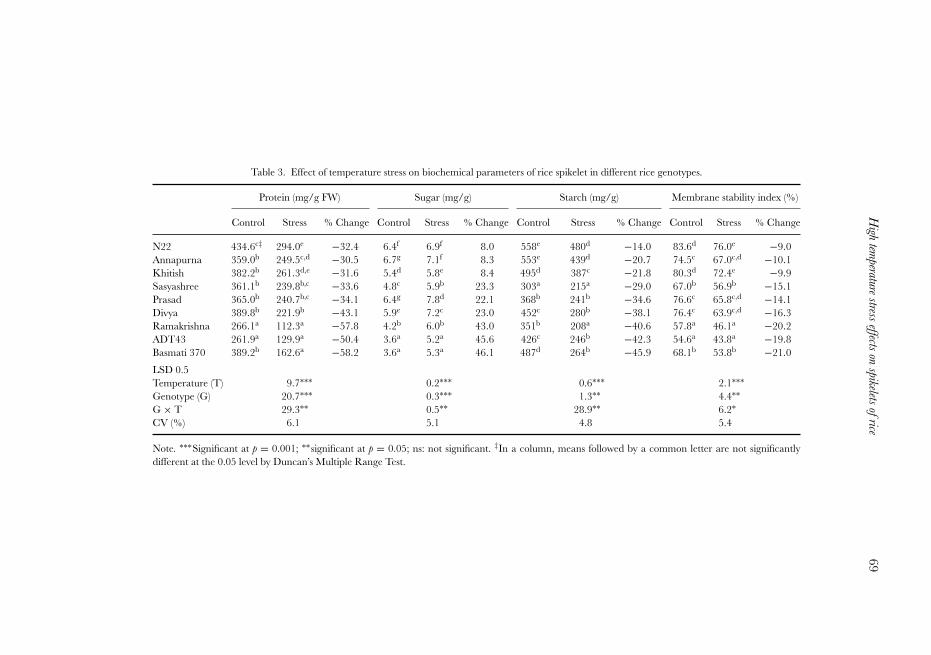
the levels of tolerance to high temperature stress (Table 3); MSI declined by 9–10%in the tolerant group (cv. N22, Annapurna and Khitish), about 15% in the moderategroup (cv. Sasyashree, Prasad and Divya) and around 20% in the susceptible group(cv. Ramakrishna, ADT 43 and Basmati 370). The concentration of grain protein,an important quality parameter, decreased by 31, 37 and 55% respectively in thetolerant, moderate and susceptible genotypes (Table 3). There were similar trend of
High temperature stress effects on spikelets of rice 67
Table 1. Combined analysis of variance for antioxidant isozymes based on band intensities in spikelets of ninerice genotypes after six different temperature exposures at flowering.
Superoxide dismutase (SOD) Catalase (CAT)
Source df MS Explained variation (%) df MS Explained variation (%)
Genotype (G) 5 3077511.5∗∗∗ 13.7 5 3934704∗∗∗ 4.4Temperature (T) 8 18778981∗∗∗ 52.4 8 67060289∗∗∗ 47.2Isozyme (I) 2 2072904∗∗∗ 2.3 2 1869304∗∗∗ 0.5G∗ T 40 404635∗∗∗ 9.0 40 520513∗∗∗ 2.9T∗ I 16 3319623∗∗∗ 18.5 16 25756755∗∗∗ 36.3G∗ I 10 19305∗∗∗ 0.2 10 768463∗∗∗ 1.7G∗ T∗ I 80 79303∗∗∗ 3.5 80 594597∗∗∗ 6.7Error 324 932 0.2 324 3574 0.2
Guaicol peroxidase (GP) Catechol oxidase (CO)
Source df MS Explained variation (%) df MS Explained variation (%)
Genotype (G) 5 887920∗∗∗ 1.4 5 465636∗∗∗ 2.7Temperature (T) 8 63708268∗∗∗ 63.6 8 13656340∗∗∗ 50.0Isozyme (I) 1 25241220∗∗∗ 5.0 3 825146.9∗∗∗ 1.8G∗ T 40 152795∗∗∗ 1.2 40 100058∗∗∗ 2.9T∗ I 8 26198410∗∗∗ 26.1 24 3385266∗∗∗ 37.2G∗ I 5 87457∗∗∗ 0.1 15 41037∗∗∗ 0.7G∗ T∗ I 40 129967∗∗∗ 1.0 120 51932∗∗∗ 4.6Error 216 33448 1.4 432 345 0.1
Note: ∗, ∗∗ and ∗∗∗: Significant at 0.05, 0.01 and 0.001 respectively; ns: not significant.
decrease in the concentrations of starch in these genotypes due to high temperatureexposure. On the contrary, the concentration of sugar increased by 7, 18 and 31% inthe tolerant, moderate and susceptible genotypes respectively.
D I S C U S S I O N
Rice is predominantly a tropical crop and is often subjected to the vagaries of weather,which include changes in temperature during its growth period. The predicted increasein temperatures due to climate change processes can have varying influences ondifferent tissues and genotypes of rice, depending on the intensity and duration ofhigh temperature stress (Krishnan et al., 2007, Yoshida et al., 1981). From the presentstudy, decrease in yield, grain weight, membrane stability index, grain number perpanicle and the concentration of grain protein and starch but increase in spikeletsterility (%) and the concentration of grain sugar due to high temperature (45 ◦C)exposure for six days at flowering clearly showed that rice is highly sensitive duringthe reproductive stage. In addition, the response of rice to high temperature exposurewas found to be genotype-dependent. For achieving higher seed yield, it is essential toachieve an increased number of grains per panicle with less sterility (%). The changesin grain quality parameters, such as increase in sugar concentration, but decreasein starch concentration in the high temperature-treated plants can be attributed tothe loss of membrane integrity (Krishnan and Surya Rao, 2005). Due to the cell

68S
MR
UT
ID
AS
etal.
Table 2. Effect of temperature stress on yield attributes of different rice genotypes.
Yield/plant (g) Sterility (%)1000 grain weight
(g)No. of
grains/panicle
Control Stress % Change Control Stress % Change Control Stress % Change Control Stress % Change
N22 9.9c‡ 9.7c,d −5.6 13.1a 16.9a,b 28.6 22.3b,c 22.3c,d −0.2 73g 62e −15.1Annapurna 9.9c 9.3c,d −5.7 21.3d 27.4b 28.4 20.9a,b 20.7a,b,c −1.1 62e 50d −19.3Khitish 10.6d,e 10.1d,e −4.9 18.0c 25.5b 56.8 22.3c 22.0c,d −1.1 67f 56d −16.4Sasyashree 10.7e 9.9d,e −7.7 16.3b 32.3c 97.9 21.7b,c 21.0b,c −3.2 64e 49d −23.4Prasad 10.2d,e 9.4c,d −7.7 17.6c 33.7c 91.8 22.1c 21.3c −3.6 48c 36c −25.0Divya 11.0e 10.2d,e −8.0 15.5b 31.9c 105.4 21.7b,c 21.0b,c −3.0 59d 37c −37.3Ramakrishna 8.4a 6.6a,b −21.8 14.8b 64.6e 337.3 20.7a 19.3a −6.6 37b 18b −51.4ADT43 9.4b 7.2b,c −12.4 18.1c 67.7e 273.3 21.5a,b,c 20.4a,b −5.3 38b 19b −50.0Basmati 370 8.6a 6.5a −14.1 21.3d 60.2d 186.3 23.4c 22.3c,d −4.0 34a 15a −55.9
LSD 0.5Temperature (T) 0.4∗∗∗ 0.7∗∗∗ 0.6∗∗∗ 1.4∗∗∗Genotype (G) 0.9∗∗∗ 1.5∗∗∗ 1.2∗∗ 3.0∗∗∗G × T 1.2 ns 2.1∗∗ 1.8 ns 4.3∗∗CV (%) 7.7 4.8 5.1 5.8
Note. ∗∗∗Significant at p = 0.001; ∗∗ significant at p = 0.05; ns: not significant. ‡In a column, means followed by a common letter are not significantlydifferent at the 0.05 level by Duncan’s Multiple Range Test.
High
temperature
stresseffects
onspikelets
ofrice
69
Table 3. Effect of temperature stress on biochemical parameters of rice spikelet in different rice genotypes.
Protein (mg/g FW) Sugar (mg/g) Starch (mg/g) Membrane stability index (%)
Control Stress % Change Control Stress % Change Control Stress % Change Control Stress % Change
N22 434.6c‡ 294.0e −32.4 6.4f 6.9f 8.0 558e 480d −14.0 83.6d 76.0e −9.0Annapurna 359.0b 249.5c,d −30.5 6.7g 7.1f 8.3 553e 439d −20.7 74.5c 67.0c,d −10.1Khitish 382.2b 261.3d,e −31.6 5.4d 5.8e 8.4 495d 387c −21.8 80.3d 72.4e −9.9Sasyashree 361.1b 239.8b,c −33.6 4.8c 5.9b 23.3 303a 215a −29.0 67.0b 56.9b −15.1Prasad 365.0b 240.7b,c −34.1 6.4g 7.8d 22.1 368b 241b −34.6 76.6c 65.8c,d −14.1Divya 389.8b 221.9b −43.1 5.9e 7.2c 23.0 452c 280b −38.1 76.4c 63.9c,d −16.3Ramakrishna 266.1a 112.3a −57.8 4.2b 6.0b 43.0 351b 208a −40.6 57.8a 46.1a −20.2ADT43 261.9a 129.9a −50.4 3.6a 5.2a 45.6 426c 246b −42.3 54.6a 43.8a −19.8Basmati 370 389.2b 162.6a −58.2 3.6a 5.3a 46.1 487d 264b −45.9 68.1b 53.8b −21.0
LSD 0.5Temperature (T) 9.7∗∗∗ 0.2∗∗∗ 0.6∗∗∗ 2.1∗∗∗Genotype (G) 20.7∗∗∗ 0.3∗∗∗ 1.3∗∗ 4.4∗∗G × T 29.3∗∗ 0.5∗∗ 28.9∗∗ 6.2∗CV (%) 6.1 5.1 4.8 5.4
Note. ∗∗∗Significant at p = 0.001; ∗∗significant at p = 0.05; ns: not significant. ‡In a column, means followed by a common letter are not significantlydifferent at the 0.05 level by Duncan’s Multiple Range Test.

70 S M RU T I DA S et al.
membrane injury, the enzymes lose their catalytic activities, the cell reserves getdepleted and the by-products of catabolic reactions become toxic (Wahid et al., 2007).High temperature exposure leading to decrease in the concentrations of protein andstarch, and increase in soluble sugar concentration as observed in this present study,may lead to irreversible degradation of important cellular machinery during graindevelopment. From an experiment using the phytotron facilities at the InternationalRice Research Institute, Philippines, it was observed that flowering was the mostsensitive stage to high temperature in three indica (tropical) rice selections (N22,IR747B2-6 and BKN6624-46-2) (Satake and Yoshida, 1978). The spikelet tissuetemperature of 33.7 ◦C even for an hour at anthesis induced sterility (Jagadish et al.,2007). Even the temperatures of 38 ◦C and 41 ◦C for an hour before or after anthesisaffected spikelet fertility (Matsui and Omasa, 2002). Similar observations are welldocumented but the underlying biochemical mechanisms and gene expression are tobe investigated in detail (Hu et al., 2009).
Many antioxidant enzymes are activated to scavenge the ROS generated duringmetabolic reactions. Isozyme profiling of antioxidant enzymes can suggest whetherinduction of new isoforms or only the quantitative changes in the constitutive isoformsoccur due to the high temperature stress. Agronomic characteristics, such as survival,plant height, yield and its components and grain quality parameters, offer theadvantage of ease in their measurements. Because of the complex nature and rangeof plant responses, it is difficult to define tolerance to high temperature exposure bythe agronomic traits alone. Our results suggested that the antioxidant defense systemwould be a more appropriate criteria for evaluating and selecting genotypes tolerantto high temperature. There was induction of new isozymes of SOD, GP and CO inspikelets after the high temperature treatment. No new isozymes were inducted forCAT and APX but there were alterations in the relative quantitation of constitutiveisoforms. The high temperature treatments of up to 40 ◦C activated the isoenzymescompared with the control treatment at 30 ◦C. Similar results were reported for PODin rice (Srivalli et al., 2003).
In the present study of high temperature-stressed rice spikelets, both CO and GPincreased significantly whereas APX did not show any variation due to different levelsof high temperature exposure over 25 ◦C. In an earlier study on antioxidant defencemechanisms in rice seedlings exposed to low temperature stress, POD was found toshow transient increase in its activity (Oidaira et al., 2000). Peroxidases are known to beinvolved in the biosynthesis of cell wall and lignification. Higher activities of peroxidasemay contribute not only to the ROS scavenging but also to the maintenance of cellularmembranes and cell wall function. In the present study, there were about 1–3 SODisozymes in rice spikelets and one of them responded directly to the high temperaturestress treatment.
Catalase activity of rice seed embryo and germination rate were closely correlatedunder low temperature conditions (Tanida, 1996). The tolerance in rice to chillinginjury was attributed to the cold stability of CAT and APX. On the contrary, Oidairaet al. (2000) did not observe significant changes in CAT activities in rice seedlingsunder low temperature. In the present study, three major bands representing isozymes

High temperature stress effects on spikelets of rice 71
of CAT were detected in rice spikelets. We observed that increase in sterility anddecrease in CAT activities in spikelets were related under high temperature stressconditions. The activities of all three isozymes of CAT increased with high temperaturetreatment of 45 ◦C for six days (Figure 3). Likewise, severe water stress was found toinduce a new CAT isoform in rice seedlings (Srivalli et al., 2003). In the present studyof high temperature stress, higher intensities of all CAT isoenzymes were detected,suggesting the role of ROS scavenging mechanisms directly and the generation ofROS indirectly. No new isoform of APX was found to be induced in the spikelets afterthe high temperature treatment in the present study. Probably, the isozyme of APX,which was constitutive in nature, was either activated or inhibited. The activation ofthe existing isozyme suggests that the same isozyme may be produced under hightemperature stress. In agreement with many previous observations (Scandalios et al.,1984), our results indicate that only those isoforms which are needed to protect thehigh temperature stress condition are expressed. It is also well known that antioxidantisozymes respond differentially to biotic and abiotic stresses. Any limitation of one ofthe antioxidant systems may be compensated through the up-regulation of another.Although no such dependence was clearly observed in the present study, there existeda close similarity between the enzyme profiles of GP and CO in rice spikelets, whensubjected to high temperature stress. Likewise, the profiles of CAT, GP and SODsuggest their interrelationships.
High or low temperature stress can have adverse effect on cellular homeostasis, anduncoupling of many physiological processes in plants (Wahid et al., 2007). Usingmicroarray hybridization for 51,279 probe sets for japonica and indica transcripts,the genome-wide response patterns under heat stress for O. sativa subsp. japonica (cv.Zhonghua 11) was found to be different from those of cold, drought and salt, eventhough there were 240 genes, which responded to both heat and drought (Hu et al.,2009). The Os04g01740 (OsHsp90-1) gene of the Hsp70 family was found to bestrongly induced under heat stress, but not induced by cold, drought or salt. Hightemperature can increase the accumulation of sucrose and pyruvate/oxaloacetate-derived amino acids and decrease levels of sugar phosphates and organic acids involvedin glycolysis/gluconeogenesis and the tricarboxylic acid cycle respectively (Yamakawaand Hakata 2010). The DNA microarray analysis revealed that genes for alternativeoxidases (AOXs) were up-regulated to a Ratio of Cumulative Expression Level (RECL,that is the ratio of the sum of expression levels for all genes encoding aminoacyl-tRNAsynthetase for a given amino acid in the high temperature condition to that in thecontrol) of 1.40 under high temperature conditions. Alternative oxidases can preventthe over-production of respiratory components leading to the formation of ROS.
The antioxidant defense mechanism is a part of high temperature stress adaptation,and its strength is correlated with acquisition of thermotolerance. The ROS areprimary damage causing agents, and at the same time they play protective role byacting as signalling molecules. In the cellular defence mechanisms against the effectof ROS on different biomolecules, antioxidant enzymes are expressed. Enhancedactivities of antioxidant enzymes and their levels help to protect from the harmfuleffects of ROS. Our results clearly suggest that high temperature stress at the flowering

72 S M RU T I DA S et al.
stage of spikelets affects oxidative reactions. The activities of antioxidant enzymes(CAT, SOD, APX, GP and CO) responded to the high temperature stress, whichis also genotype-dependent. The levels of tolerance to high temperature stress areprimarily due to the differences in genotypic resistance by multiple molecular forms ofantioxidant enzymes and their relative compositions. Increase in the activity of theseantioxidant enzymes appears almost simultaneously, which can lead to the avoidanceof ROS generation or can scavenge ROS or repair after the injury. The synthesis ofnew isozymes of antioxidant enzymes with altered kinetic properties is more beneficialthan the existing antioxidant enzymes for scavenging of ROS. In conclusion, the resultsof this study confirmed our hypothesis that activities of antioxidant enzymes increaseddue to high temperature exposure but declined suddenly after 35 ◦C and beyond,depending on the varying levels of capabilities of genotypes to high temperaturestress. Differential modulation of SOD, CAT, APX, GP and CO isozymes and theiractivities in rice spikelets suggest their potential use as biomarkers for identificationand selection of genotypes that are tolerant to high temperature stress.
Acknowledgements. P. Krishnan and B. Ramakrishnan thank the Science andEngineering Research Council (SERC), Department of Science and Technology(DST), Government of India for the support with a research project on ‘Hightemperature stress response and associated changes in growth and yield of rice (Oryza
sativa L)’. Smruti Das is grateful to DST for the Junior Research Fellowship. Weacknowledge the facilities and support provided by the Director, CRRI, Cuttack andthe Director, IARI, New Delhi.
R E F E R E N C E S
Cakmak, I. and Marschner, H. (1992). Magnesium deficiency and high light intensity enhance activities of superoxidedismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiology 98:1222–1227.
Chowdhury, S. R. and Choudhury, M. A. (1985). Hydrogen peroxide metabolism as an index of water stress tolerancein Jute. Physiologia Plantarum 65:503–507.
Das, S., Nayak, M., Patra, B. C., Ramakrishnan, B. and Krishnan, P. (2010). Characterization of seeds of selected wildspecies of rice stored under high temperature and humidity conditions. Indian Journal of Biochemistry and Biophysics
47:178–184.Djanaguiraman, M., Prasad, P. V. V. and Al-Khatib, K. (2011). Ethylene perception inhibitor 1-MCP decreases
oxidative damage of leaves through enhanced antioxidant defense mechanisms in soybean plants grown underhigh temperature stress. Environmental and Experimental Botany 71:215–223.
Gomez, K. A. and Gomez, A. A. (1984). Statistical Procedures for Agricultural Research, 2nd edn. New York: John Wiley.Hu, W., Hu, G. and Han, B. (2009). Genome-wide survey and expression profiling of heat shock proteins and heat shock
factors revealed overlapped and stress specific response under abiotic stresses in rice. Plant Science 176:583–590.Intergovernmental Panel on Climate Change (IPCC) (2007). Fourth Assessment Report of the Intergovernmental Panel on
Climate Change: The Impacts, Adaptation and Vulnerability (Working Group III). New York, NY: Cambridge UniversityPress.
Jagadish, S. V. K., Craufurd, P. Q. and Wheeler, T. R. (2007). High-temperature stress and spikelet fertility in rice(Oryza sativa L.). Journal of Experimental Botany 58:1627–1635.
Jagadish, S. V. K., Muthurajan, R., Oane, R., Wheelerm, T. R., Heuer, S., Bennett, J. and Craufurd, P. Q. (2010).Physiological and Proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). Journal
of Experimental Botany 61:143–156.Krishnan, P., Ramakrishnan, B., Raja Reddy, K. and Reddy, V. R. (2011). High temperature stress effects on rice
plant growth and yield. Advances in Agronomy (USA) 111:87–206.

High temperature stress effects on spikelets of rice 73
Krishnan, P. and Surya Rao, A. V. (2005). Effects of genotype and environment on seed yield and quality of rice. The
Journal of Agricultural Science (Cambridge) 143:283–292.Krishnan, P., Swain, D. K., Baskar, C., Nayak, S. K. and Dash, R. N. (2007). Simulation studies on the effects of
elevated CO2 and temperature on rice yield in Eastern India. Agriculture, Ecosystem and Environment 122:233–242.Matsui, T. and Omasa, K. (2002). Rice (Oryza sativa L.) cultivars tolerant to high temperature at flowering: anther
characteristics. Annals of Botany 89:683–687.Morita, S., Shiratsuchi, H., Takahashi, J. and Fujita, K. (2004). Effect of high temperature on grain ripening in rice
plants. Analysis of the effects of high night and high day temperatures applied to the panicle and other parts ofthe plant. Japanese Journal of Crop Science 73:77–83.
Nakano, Y. and Asada, K. (1981). Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinachchloroplasts. Plant and Cell Physiology 22:867–880.
Oidaira, H., Sano, S., Koshiba, T. and Ushimaru, T. (2000). Enhancement of antioxidative enzyme activities in chilledrice seedlings. Journal of Plant Physiology 156:811–813.
Rao, M.V., Paliyath, G. and Ormrod, D. P. (1996). Ultraviolet-B and ozone-induced biochemical changes in antioxidantenzymes of Avabidopsis thaliana. Plant Physiology 110:125–136.
Satake, T. and Yoshida, S. (1978). High temperature induced sterility in indica rices at flowering. Japanese Journal of Crop
Science 47:6–17.Scandalios, J. G., Tsaftaris, A. S., Chandlee, J. M. and Skadsen, R. M. (1984). Expression of the developmentally
regulated catalase (Cat) genes in maize. Developmental Genetics 4:281–293.Sheehy, J. E., Mitchell, P. L., Beerling, D. J., Tsukaguchi, T. and Woodward, F. I. (1998). Temperature of rice spikelets:
thermal damage and the concept of a thermal burden. Agronomie 18:449–460.Srivalli, B., Sharma, G. and Khanna-Chopra, R. (2003). Antioxidative defense system in an upland rice cultivar
subjected to increasing intensity of water stress followed by recovery. Physiologia Plantarum 119:503–512.Tanida, M. (1996). Catalase activity of rice seed embryo and its relation to germination rate at a low temperature.
Breeding Science 46:23–27.Türkan, Bor M., Zdemir, F. and Koca, H. (2005). Differential responses of lipid peroxidation and antioxidants in
the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycolmediated water stress. Plant Sciences 168:223–231.
Wahid, A., Gelani, S., Ashraf, M. and Foolad, M. R. (2007). Heat tolerance in plants: an overview. Environmental and
Experimental Botany 61:199–223.Wassmann, R., Jagadish, S. V. K., Sumfleth, K., Pathak, H., Howell, G., Ismail, A., Serraj, R., Redona, E., Singh, R.
K. and Heuer, S. (2009). Regional vulnerability of climate change impacts on Asian rice production and scope foradaptation. Advances in Agronomy 102:91–133.
Yamakawa, H. and Hakata, M. (2010). Atlas of rice grain filling-related metabolism under high temperature: jointanalysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of aminoacid accumulation. Plant Cell Physiology 51:795–809.
Yoshida, S., Satake, T. and Mackill, D. J. (1981). High-Temperature Stress in Rice (Review), IRRI Research Paper Series,67. Manila, Philippines: International Rice Research Institute, 1–15.
Zhao, X., Nishimura,, Y., Fukumoto, Y. and Li, J. (2011). Effect of high temperature on active oxygen species,senescence and photosynthetic properties in cucumber leaves. Environmental and Experimental Botany 70:212–216.
Copyright © 2022 FDOKUMEN




















