
Pyruvate-sensitive AOX exists as a non-covalently associated dimer in the homeothermic spadix of the...
7
Pyruvate-sensitive AOX exists as a non-covalently associated dimer in the homeothermic spadix of the skunk cabbage, Symplocarpus renifolius Yoshihiko Onda a , Yoshiaki Kato b , Yukie Abe b , Takanori Ito b , Yasuko Ito-Inaba b , Miyuki Morohashi a,c , Yuka Ito b , Megumi Ichikawa b , Kazushige Matsukawa a , Minoru Otsuka b , Hiroyuki Koiwa c , Kikukatsu Ito b, * a United Graduate School of Agricultural Science, Iwate University, Ueda, Morioka, Iwate 020 8550, Japan b Cryobiosystem Research Center, Faculty of Agriculture, Iwate University, Ueda, Morioka, Iwate 020 8550, Japan c Research Institute of Bio-system Informatics, Tohoku Chemical Co., Ltd., Odori, Morioka, Iwate 020 0022, Japan Received 17 October 2007; revised 20 November 2007; accepted 20 November 2007 Available online 3 December 2007 Edited by Michael R. Sussman Abstract The cyanide-resistant alternative oxidase (AOX) is a homodimeric protein whose activity can be regulated by the oxi- dation/reduction state and by a-keto acids. To further clarify the role of AOX in the skunk cabbage, Symplocarpus renifolius, we have performed expression and functional analyses of the encod- ing gene. Among the various tissues in the skunk cabbage, SrAOX transcripts were found to be specifically expressed in the thermogenic spadix. Moreover, our data demonstrate that the SrAOX protein exists as a non-covalently associated dimer in the thermogenic spadix, and is more sensitive to pyruvate than to other carboxylic acids. Our results suggest that the pyruvate- mediated modification of SrAOX activity plays a significant role in thermoregulation in the skunk cabbage. Ó 2007 Federation of European Biochemical Societies. Pub- lished by Elsevier B.V. All rights reserved. Keywords: Thermogenic plants; Mitochondria; Respiration; Alternative oxidase; Skunk cabbage; Pyruvate 1. Introduction The skunk cabbage is a homeothermic plant that flowers in early spring and maintains its spadix temperature at around 20 °C for about a week even when the ambient temperature drops below freezing [1–4]. It has been shown that the mecha- nism underlying this temperature regulation resides in the spa- dix tissue alone, and not in the spathe, leaves or spadix stalk [3]. When the ambient temperature decreases, the spadix will normally lose heat and its temperature will thus begin to de- crease. However, through some undetermined biochemical mechanism, a small drop in the temperature of the skunk cab- bage spadices results in an increase in their respiration rate, rather than the expected decrease [1,2,4]. Heat production in thermogenic plants has been attributed to a large increase in their alternative oxidase (AOX) activity [5–11]. AOX acts as an alternative terminal oxidase in the elec- tron transfer chain, reducing molecular oxygen to water. How- ever, in contrast to the mitochondrial terminal oxidase, cytochrome c oxidase, AOX does not pump protons, and this allows the dramatic drop in free energy between ubiquinol and oxygen to be dissipated as heat. Plant AOX activity is highly regulated not only through transcriptional control [12], but also by two post-translational mechanisms. In the first of these, the reduction state of the AOX protein (i.e. whether the protein exists as an inactive oxidized dimer or reduced ac- tive dimer) regulates its activity [13,14]. In the second mecha- nism, some a-keto acids further stimulate the activity of the reduced AOX protein [15–17]. However, although a partial purification of the cyanide-resistant alternative oxidase from thermogenic skunk cabbage has been achieved [6], no informa- tion is yet available concerning the reduction state and effects of a-keto acids on the activities of AOX protein. It has been shown that the homeothermic skunk cabbage spe- cies that is indigenous to Japan is Symplocarpus renifolius [18], and we have therefore isolated a novel AOX gene from this spe- cies (SrAOX) and found that it is specifically expressed in the thermogenic spadix of this plant. Interestingly, among a variety of organic acids that we tested, including some a-keto acids, only pyruvate was found to stimulate SrAOX, which was also found to exist as a non-covalently associated dimer in vivo. 2. Materials and methods 2.1. Plant materials S. renifolius plants were collected from fields situated within the Iwate University campus (39°43 0 N, 141°08 0 E), the Shizukuishi factory of Taishi Food Inc. (39°45 0 N, 141°00 0 E), from Hakuba in the Nagano prefecture (36°39 0 N, 137°50 0 E) and also from Omori, Akita prefecture (39°19 0 N, 141°20 0 E), Japan, during the period between March and April from 2000 to 2007. 2.2. Isolation of the full-length SrAOX cDNA and sequence analysis For the isolation of SrAOX cDNA by RT-PCR, total RNA was first extracted from flash-frozen, pulverized thermogenic spadices accord- ing to our previously reported method [19,20]. Oligo (dT)-primed first-strand cDNAs were prepared from 1 lg samples of total RNA using the Advantage RT-for-PCR Kit (Clontech Laboratories Inc., Palo Alto, CA, USA). First-strand cDNA was then used to generate partial SrAOX cDNAs by PCR amplification. Primers were designed based on conserved cDNA sequences between the AOXs in thermogenic plants (SgAOX [21] and DvAOX [22]) that cover the di- iron binding motifs as follows: Sgaox_F0 (22 mer) 5 0 -AAA GAG Abbreviations: AOX, alternative oxidase; BSA, bovine serum albumin; PVPP, polyvinylpolypyrrolidone; DTT, dithiothreitol; DMSO, di- methyl sulfoxide; IO-SMP, inside-out submitochondrial particles * Corresponding author. Fax: +81 19 621 6143. E-mail address: [email protected] (K. Ito). 0014-5793/$32.00 Ó 2007 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2007.11.061 FEBS Letters 581 (2007) 5852–5858
Transcript of Pyruvate-sensitive AOX exists as a non-covalently associated dimer in the homeothermic spadix of the...

FEBS Letters 581 (2007) 5852–5858
Pyruvate-sensitive AOX exists as a non-covalently associated dimer inthe homeothermic spadix of the skunk cabbage, Symplocarpus renifolius
Yoshihiko Ondaa, Yoshiaki Katob, Yukie Abeb, Takanori Itob, Yasuko Ito-Inabab,Miyuki Morohashia,c, Yuka Itob, Megumi Ichikawab, Kazushige Matsukawaa, Minoru Otsukab,
Hiroyuki Koiwac, Kikukatsu Itob,*
a United Graduate School of Agricultural Science, Iwate University, Ueda, Morioka, Iwate 020 8550, Japanb Cryobiosystem Research Center, Faculty of Agriculture, Iwate University, Ueda, Morioka, Iwate 020 8550, Japanc Research Institute of Bio-system Informatics, Tohoku Chemical Co., Ltd., Odori, Morioka, Iwate 020 0022, Japan
Received 17 October 2007; revised 20 November 2007; accepted 20 November 2007
Available online 3 December 2007
Edited by Michael R. Sussman
Abstract The cyanide-resistant alternative oxidase (AOX) is ahomodimeric protein whose activity can be regulated by the oxi-dation/reduction state and by a-keto acids. To further clarify therole of AOX in the skunk cabbage, Symplocarpus renifolius, wehave performed expression and functional analyses of the encod-ing gene. Among the various tissues in the skunk cabbage,SrAOX transcripts were found to be specifically expressed inthe thermogenic spadix. Moreover, our data demonstrate thatthe SrAOX protein exists as a non-covalently associated dimerin the thermogenic spadix, and is more sensitive to pyruvate thanto other carboxylic acids. Our results suggest that the pyruvate-mediated modification of SrAOX activity plays a significant rolein thermoregulation in the skunk cabbage.� 2007 Federation of European Biochemical Societies. Pub-lished by Elsevier B.V. All rights reserved.
Keywords: Thermogenic plants; Mitochondria; Respiration;Alternative oxidase; Skunk cabbage; Pyruvate
1. Introduction
The skunk cabbage is a homeothermic plant that flowers in
early spring and maintains its spadix temperature at around
20 �C for about a week even when the ambient temperature
drops below freezing [1–4]. It has been shown that the mecha-
nism underlying this temperature regulation resides in the spa-
dix tissue alone, and not in the spathe, leaves or spadix stalk
[3]. When the ambient temperature decreases, the spadix will
normally lose heat and its temperature will thus begin to de-
crease. However, through some undetermined biochemical
mechanism, a small drop in the temperature of the skunk cab-
bage spadices results in an increase in their respiration rate,
rather than the expected decrease [1,2,4].
Heat production in thermogenic plants has been attributed
to a large increase in their alternative oxidase (AOX) activity
[5–11]. AOX acts as an alternative terminal oxidase in the elec-
Abbreviations: AOX, alternative oxidase; BSA, bovine serum albumin;PVPP, polyvinylpolypyrrolidone; DTT, dithiothreitol; DMSO, di-methyl sulfoxide; IO-SMP, inside-out submitochondrial particles
*Corresponding author. Fax: +81 19 621 6143.E-mail address: [email protected] (K. Ito).
0014-5793/$32.00 � 2007 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2007.11.061
tron transfer chain, reducing molecular oxygen to water. How-
ever, in contrast to the mitochondrial terminal oxidase,
cytochrome c oxidase, AOX does not pump protons, and this
allows the dramatic drop in free energy between ubiquinol and
oxygen to be dissipated as heat. Plant AOX activity is highly
regulated not only through transcriptional control [12], but
also by two post-translational mechanisms. In the first of
these, the reduction state of the AOX protein (i.e. whether
the protein exists as an inactive oxidized dimer or reduced ac-
tive dimer) regulates its activity [13,14]. In the second mecha-
nism, some a-keto acids further stimulate the activity of the
reduced AOX protein [15–17]. However, although a partial
purification of the cyanide-resistant alternative oxidase from
thermogenic skunk cabbage has been achieved [6], no informa-
tion is yet available concerning the reduction state and effects
of a-keto acids on the activities of AOX protein.
It has been shown that the homeothermic skunk cabbage spe-
cies that is indigenous to Japan is Symplocarpus renifolius [18],
and we have therefore isolated a novel AOX gene from this spe-
cies (SrAOX) and found that it is specifically expressed in the
thermogenic spadix of this plant. Interestingly, among a variety
of organic acids that we tested, including some a-keto acids,
only pyruvate was found to stimulate SrAOX, which was also
found to exist as a non-covalently associated dimer in vivo.
2. Materials and methods
2.1. Plant materialsS. renifolius plants were collected from fields situated within the
Iwate University campus (39�43 0N, 141�08 0E), the Shizukuishi factoryof Taishi Food Inc. (39�45 0N, 141�00 0E), from Hakuba in the Naganoprefecture (36�39 0N, 137�50 0E) and also from Omori, Akita prefecture(39�19 0N, 141�20 0E), Japan, during the period between March andApril from 2000 to 2007.
2.2. Isolation of the full-length SrAOX cDNA and sequence analysisFor the isolation of SrAOX cDNA by RT-PCR, total RNA was first
extracted from flash-frozen, pulverized thermogenic spadices accord-ing to our previously reported method [19,20]. Oligo (dT)-primedfirst-strand cDNAs were prepared from 1 lg samples of total RNAusing the Advantage RT-for-PCR Kit (Clontech Laboratories Inc.,Palo Alto, CA, USA). First-strand cDNA was then used to generatepartial SrAOX cDNAs by PCR amplification. Primers weredesigned based on conserved cDNA sequences between the AOXs inthermogenic plants (SgAOX [21] and DvAOX [22]) that cover the di-iron binding motifs as follows: Sgaox_F0 (22 mer) 5 0-AAA GAG
blished by Elsevier B.V. All rights reserved.

Table 1Respiration rates of mitochondria isolated from stigma stage ofthermogenic spadices of the skunk cabbage
Sequential additions O2 uptake
nmol O2 min�1 mg�1 protein
NADH 532.7 ± 34.5+ADP 650.1 ± 37.9+KCN 494.6 ± 25.8+n-Propyl gallate 8.6 ± 1.1
Rates of O2 uptake were determined using an O2 electrode cuvette after
Y. Onda et al. / FEBS Letters 581 (2007) 5852–5858 5853
GAC GGC TCC GAG TGG C-3 0, and Sgaox_R1 (22 mer) 5 0-ATCAGT GGT ACC CGA GCG GCG C-3 0.
The 5 0 and 3 0 ends of the SrAOX cDNA regions were generatedusing the SMART RACE cDNA Amplification Kit (Clontech Labora-tories Inc., Palo Alto, CA, USA) with the primers 5 0-ATA AGT GGTACC CGA GCG GCG C-3 0 (Sraox50gsp1 (22 mer)), 5 0-CGA GGCGAA GTG GTT GAC-3 0 (Sraox50gsp2 (18 mer)), and 5 0-CTC CTGGAG GAG GCC GAG CCC G-30(Sraox30gsp1 (22 mer)). To obtainthe full length SrAOX cDNA, additional PCR amplifications were per-formed using the primers Sraox_full_F0 (20 mer) 5 0-TTC TTG TGGATT TCT CCC GG-3 0, and Sraox_full_R1 (24 mer) 5 0-CGA TTGCTC TTT TAT CTC CTC CTC-3 0. PCR products of 1.2 kb were ob-tained and were then gel purified and subcloned into the pBluescript IIKS+ vector (Toyobo Inc., Tokyo, Japan). The resulting plasmid,pSraox1-1, carrying the full length SrAOX cDNA, was sequenced onboth strands with the BigDye Terminator Cycle Sequencing Kit (Ap-plied Biosystems Inc., Foster City, CA, USA) and an automatedDNA sequencer (ABI3700, Applied Biosystems, Foster City, CA,USA). These sequence data were then analyzed using GENETYX soft-ware (Software Development Inc., Tokyo, Japan).
To amplify SrAOX-related cDNAs using total RNAs prepared fromthermogenic spadices, 3 0-RACE was performed using the SMARTRACE cDNA amplification kit and with primers that were designedbased on the highly conserved sequences among plant AOXs. These in-cluded Sraox _F3.5 for the putative iron-binding motif located moreproximally to the N-terminus of the SrAOX protein, and Sraox_F1,Sraox F2, and Sraox F3 for the more upstream regions of Sraox_F3.5. The sequences are as follows: Sraox_F3.5 (25 mer) 5 0-GAACGA GAG GAT GCA CCT GAT GAC C-3 0, Sraox_F1 (22 mer)5 0-ATG TCT TCT TCC AGA GGC GGT A-3 0, Sraox F2 (23 mer)5 0-ACG TAC GAG GCG GAC CTG TCG AT-3 0, and Sraox F3(23 mer) 5 0-ATG GTG GGC GGG CTG CTC CTC CA-3 0.
2.3. Northern blot analysisTotal RNA was isolated from the leaves, spathes, spadices, and
roots of Symplocarpus plants as described above. Five micrograms ofeach RNA sample was then fractionated on a 1.0% agarose gel, trans-ferred to a Hybond N+ membrane (Amersham Biosciences Inc., Piscat-away, NJ, USA), and hybridized with a full-length SrAOX cDNAprobe. Bands were visualized using the AlkPhos Direct Labeling andDetection System (Amersham Biosciences Inc., Piscataway, NJ,USA) according to the manufacturer�s instructions.
2.4. Isolation of mitochondriaMitochondria were isolated from the thermogenic florets of skunk
cabbages as described previously [23] with minor modifications.Briefly, florets (3–4 g fresh weight) were diced and suspended in20 mL of ice-cold ‘‘grinding buffer’’ (0.4 M mannitol, 25 mM MOPS/KOH, 2 mM EDTA, 10 mM KH2PO4, 1% PVP-40, 20 mM ascorbicacid, 4 mM cysteine, 2 mM pyruvate, 1% (w/v) defatted bovine serumalbumin (BSA), 2% (w/v) polyvinylpolypyrrolidone (PVPP), pH 7.2).These thermogenic floret preparations were then homogenized and fil-tered through eight layers of Miracloth (Calbiochem Inc., Darmstadt,Germany). Mitochondria were enriched using a three-step centrifuga-tion process: once at 1000 · g for 10 min (organelles in supernatant)and twice at 10000 · g for 20 min (organelles in pellet). The mitochon-drial fraction was resuspended in 2 mL of ‘‘washing buffer’’ (0.4 Mmannitol, 10 mM MOPS/KOH, 2 mM pyruvate, and 0.1% [w/v]BSA, pH 7.2) and layered on the top of three-step Percoll gradients(two gradients of 36 mL each) containing 5 mL of 60% [v/v], 15 mLof 45% [v/v] and 15 mL of 27% [v/v] Percoll in 0.25 M sucrose,10 mM HEPES/KOH, 2 mM pyruvate 0.2% [w/v] BSA, pH 7.2). Aftercentrifugation for 60 min at 20000 · g the mitochondria were isolatedfrom the 27%/45% interface. To remove the Percoll, the purified mito-chondria were centrifuged twice in washing buffer for 10 min at10000 · g. The protein concentrations were measured with the dye-binding method of Bradford [24] using BSA as a standard. The yieldof a typical preparation was about 0.8 mg of mitochondrial proteinper 3 g of thermogenic florets.
sequential addition of the indicated compounds at the following finalconcentrations: 1 mM NADH, 0.5 mM ADP, 0.5 mM KCN, and100 lM n-propyl gallate. The data are the mean ± S.D. of three mea-surements using mitochondria from the same preparation. Typicalresults are shown, representing three independent mitochondrialpreparations.
2.5. Western blotting of SrAOXWestern blotting analysis of SrAOX was performed by equal
loading of the samples onto 12.5% acrylamide gels and transfer ontopolyvinylidene difluoride membranes. Prestained protein standards
(11–170 kDa; Fermentas, Burlington, Canada) were used for molecu-lar mass estimation. The filters were incubated for 1 h at room temper-ature in 5% (w/v) milk powder in Tris-buffered saline (137 mM NaCl,2.68 mM KCl and 25 mM Tris, pH adjusted to 7.4 with HCl) that con-tained 0.1% (v/v) Tween-20 (TBS-T) and then for a further 1 h at roomtemperature with 5% (w/v) milk powder in TBS-T containing antibod-ies. Monoclonal antibodies to Sauromatum guttatum AOX [25] werediluted to 1:500. Following a wash in TBS-T, the filters were incubatedfor an additional hour at room temperature in 5% (w/v) milk powderin TBS-T that contained anti-mouse secondary antibodies (1:2000 dilu-tion) conjugated to horseradish peroxidase. The filters were washedagain with TBS-T and cross-reacted antibodies were detected usingthe SuperSignal West Femto Maximum Sensitivity Substrate (PierceInc., Perbio Science, UK) with a digital image analyzer (Light-CaptureAE6955, Atto Inc., Tokyo, Japan).
For total extracts from the thermogenic florets, the tissue was pow-dered in liquid N2. This sample powder was suspended in a solutioncontaining 62.5 mM Tris–HCl, pH 6.8, 2% (w/v) SDS, 10% glycerolat 95 �C for 5 min, and centrifuged (16000 · g for 30 min) followingthe removal of polyphenolics, tannins and other interfering substancesusing the Plant Total Protein Extraction Kit (Sigma, MO, USA). Theresulting supernatants were then used for immunoblotting analysis.
2.6. Mitochondrial respiration measurementsOxygen uptake by mitochondria (�0.1–0.25 mg of protein in a final
volume of 1.2 mL) was measured using a Clark-type electrode cuvette(Oxy1, Hansatech Instrument Inc., Norfolk, UK) at 25 �C in a reac-tion medium containing 0.3 M sucrose, 5 mM KH2PO4, 10 mM KCl,1 mM MgCl2, 0.1% (w/v) defatted BSA, and 20 mM MOPS/KOH,pH 7.2. Other additions are described in Tables 1 and 2. The O2 con-centration in air-saturated H2O at 25 �C was estimated at 240 lM ineach experiment.
2.7. Treatment of mitochondria with DTT and diamidePercoll-purified mitochondria were treated either with dithiothreitol
(DTT) or diamide by the addition of these reagents to an aliquot ofmitochondria at a final concentration of 3 and 20 mM, respectively.Dimethyl sulfoxide (DMSO) was added to the purified mitochondriato give the final concentration as in the DTT- or diamide-treated ali-quot as a control. For incubations in which DTT treatment followeddiamide treatment, DTT (final 20 mM) was added directly to the dia-mide-treated mitochondria without washing, followed by incubationfor 1 h on ice. Fresh DTT and diamide stocks were made on the dayof the experiment in each case.
2.8. Production of submitochondrial particlesFrozen mitochondria were thawed and diluted with a high salt med-
ium containing 0.4 M sucrose, 5 mM MES, and 20 mM MgCl2 (pH6.0). The suspension was then sonicated five times for 5 s each (ampli-tude 38%) at 4 �C with a vibra cell VCX500 (Sonics & Materials, Inc.,CT, USA). The sonicated mitochondria were then diluted 10-fold inthe high salt medium, and both intact mitochondria and large mem-brane fragments were pelleted at 48400 · g for 10 min and discarded.The supernatant was then centrifuged again at 105000 · g for

Table 2Effects of mono-, di-, and tri-carboxylic organic acids on NADHoxidation via AOX in mitochondria from stigma stage of thermogenicspadices of skunk cabbage
Organic acids Relative AOX activity
Without organic acids 1.00
Mono-carboxylic acidsPyruvate 2.07 ± 0.05Pyruvate* 1.87 ± 0.14Lactate 1.02 ± 0.01PEP 1.19 ± 0.01Propionate 0.99 ± 0.02Acetate 0.97 ± 0.04Gly 1.05 ± 0.05Ala 1.03 ± 0.04Crotonate 0.91 ± 0.01Acrylate 0.96 ± 0.01Acetoacetate 0.71 ± 0.02
Di-carboxylic acidsa-Ketoglutarate 1.49 ± 0.05a-Ketoglutarate* 1.10 ± 0.00Succinate 1.22 ± 0.04Malonate 0.98 ± 0.01Fumarate 1.19 ± 0.00Oxalate 0.90 ± 0.01Tartrate 1.01 ± 0.01LL-Malate 1.04 ± 0.02DD-Malate 0.94 ± 0.03
Tri-carboxylic acidsCitrate 0.86 ± 0.00Isocitrate 0.89 ± 0.01
AOX activity was measured as myxothiazol-insensitive (5 lM) and n-propyl gallate-sensitive (50 lM) oxygen consumption in the presenceof NADH (1 mM) in a standard reaction medium (pH 7.2). Changes ofthe AOX activity was assayed by adding each organic acid (10 mM)after a steady rate in the presence of myxothiazol. Effects of pyruvateand a-ketoglutarate were also assayed at the final concentrations of1 mM, which are indicated by asterisks. Changes of the AOX activateswere expressed as a relative value to that of without organic acids. Thedata are the mean ± S.D. of three assays using mitochondria from thesame preparation. Typical results are shown, representing two to threeindependent mitochondrial preparations.
Lea
f
Spat
heSp
adix
Roo
tL
eaf
Spat
heSp
adix
Roo
t
SrAOX
Thermogenic Post-thermogenic
Fig. 1. Expression analysis of SrAOX transcripts from both thermo-genic and post-thermogenic skunk cabbage samples. Five microgramaliquots of total RNA extracted from the leaf, spathe, spadix and roottissues of this plant were resolved on formaldehyde gels, blotted onto anylon filter, and hybridized with a labeled cDNA probe for SrAOX.The rRNA band intensities in the ethidium bromide-stained gels areshown as a loading control in the lower panel. Typical results areshown, representing three independent Northern analyses with sepa-rate RNA preparations.
5854 Y. Onda et al. / FEBS Letters 581 (2007) 5852–5858
60 min, and the pellet containing the inside-out submitochondrial par-ticles (IO-SMPs) was resuspended in incubation medium for respira-tion measurements. The percentage of IO-SMPs was defined as beingequal to the percentage of latent cytochrome c oxidase using a Cyto-chrome c Oxidase Assay Kit (Sigma). The yield of IO-SMPs for respi-ration analysis was found to be 88.2%.
3. Results and discussion
3.1. Expression patterns of the SrAOX transcripts
RT-PCR based cloning of the AOX cDNA from skunk cab-
bage (SrAOX) was performed using total RNA extracts from
stigma stage thermogenic spadices. The full-length cDNA of
this gene was subsequently isolated and found to encode a ma-
ture protein with a predicted molecular mass of 32.5 kDa
(DDBJ Accession No. AB183695). The following amino acids,
which are highly conserved among plant AOX proteins, were
also found to be conserved in SrAOX: (1) a cysteine residue
that is proximal to the N-terminus (termed CysI; [26]) and is
predicted to be involved in the redox regulation of AOX activ-
ity [13] and in the stimulation response to a-keto acids [27];
and (2) two glutamate and histidine residue pairs that are puta-
tive iron-binding motifs [28]. Moreover, by Southern analysis
we observed that the skunk cabbage genome contains at least
two SrAOX-related genes (data not shown).
Since it has been shown that heat production in the skunk
cabbage occurs only in the stigma and early male stages of
the spadices [2], and not in either the spathe or leaf [3], it
was of interest to determine whether SrAOX is specifically ex-
pressed in the thermogenic spadix. Therefore, we examined the
expression of this gene at the organ level in leaf, spathe, spadix
and root tissues, during stigma and late male stages, which are
thermogenic and post-thermogenic, respectively. The expres-
sion of SrAOX transcripts was found to be entirely specific
to the thermogenic spadix (Fig. 1). These results suggest that
SrAOX is involved in both tissue- and stage-specific heat pro-
duction in the skunk cabbage.
3.2. Alternative respiration of mitochondrial isolates from
thermogenic florets
We previously identified the florets as the thermogenic tis-
sues in the spadix of the skunk cabbage (Onda et al., unpub-
lished data) and speculated that if SrAOX indeed directs
spadix-specific heat production, the mitochondrial isolates
from the thermogenic florets would show high SrAOX capac-
ity. Respiratory analyses were conducted with fresh mitochon-
drial isolates from thermogenic florets using a Clark-type
oxygen electrode. As indicated in Table 1, 1 mM NADH was
added as a substrate to induce respiration and the subsequent
addition of 0.5 mM ADP resulted in a state 2–state 3 transi-
tion. After a steady rate was obtained in these isolates,
0.5 mM KCN was added to inhibit the cytochrome respiratory
pathway. Cyanide only inhibited oxygen consumption within
the range of 14–24% among three different mitochondrial
preparations (Table 1), and the subsequent addition of
100 lM n-propyl gallate, an inhibitor of the AOX [29], almost
completely stopped oxygen consumption. In contrast, it has
been shown that the oxygen consumption of mitochondria iso-
lated from non-thermogenic soybean cotyledons and old roots
is severely inhibited by 81% and 96%, respectively, upon treat-
ment with cyanide [15]. These results suggest that there are
substantial differences in the AOX capacities of thermogenic
and non-thermogenic plants and that the SrAOX protein plays
Y. Onda et al. / FEBS Letters 581 (2007) 5852–5858 5855
a substantial role in thermoregulation in the florets of skunk
cabbage.
Previously, it has been shown that pyruvate is the most effec-
tive a-keto acid activator of the soybean AOX protein [17]. To
further investigate the involvement of a-keto acids in the acti-
vation of SrAOX, we examined the effects of several organic
acids upon SrAOX regulation in purified mitochondria from
thermogenic florets of the skunk cabbage by measuring their
levels of myxothiazol-insensitive NADH oxidation. The oxy-
gen consumption rate of the control (buffer only added) was
assigned a value of 1.00, and this was compared with the val-
ues obtained following the addition of various organic acids
(Table 2). It was apparent that the SrAOX activity was mainly
activated by pyruvate and a-ketoglutarate at 10 mM. How-
ever, only pyruvate could activate SrAOX at lower concentra-
tions (1 mM) within the range of 1.9–2.1.
3.3. Effects of DTT and diamide on the SrAOX protein in
purified mitochondria
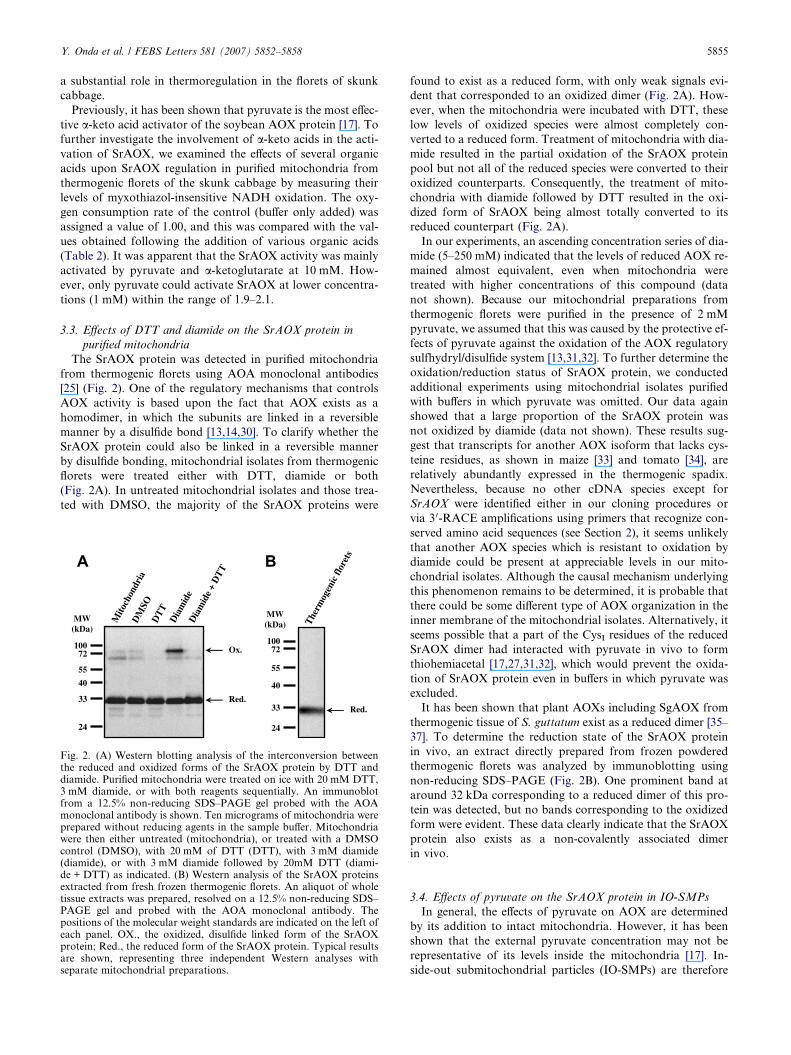
The SrAOX protein was detected in purified mitochondria
from thermogenic florets using AOA monoclonal antibodies
[25] (Fig. 2). One of the regulatory mechanisms that controls
AOX activity is based upon the fact that AOX exists as a
homodimer, in which the subunits are linked in a reversible
manner by a disulfide bond [13,14,30]. To clarify whether the
SrAOX protein could also be linked in a reversible manner
by disulfide bonding, mitochondrial isolates from thermogenic
florets were treated either with DTT, diamide or both
(Fig. 2A). In untreated mitochondrial isolates and those trea-
ted with DMSO, the majority of the SrAOX proteins were
Mito
chon
dria
DM
SOD
TT
Dia
mid
eD
iam
ide
+ D
TT
Red.
Ox.
33
24
40
55
72100
MW(kDa)
33
24
40
55
72100
MW(kDa)
Red.
A B
The
rmog
enic
flor
ets
Fig. 2. (A) Western blotting analysis of the interconversion betweenthe reduced and oxidized forms of the SrAOX protein by DTT anddiamide. Purified mitochondria were treated on ice with 20 mM DTT,3 mM diamide, or with both reagents sequentially. An immunoblotfrom a 12.5% non-reducing SDS–PAGE gel probed with the AOAmonoclonal antibody is shown. Ten micrograms of mitochondria wereprepared without reducing agents in the sample buffer. Mitochondriawere then either untreated (mitochondria), or treated with a DMSOcontrol (DMSO), with 20 mM of DTT (DTT), with 3 mM diamide(diamide), or with 3 mM diamide followed by 20mM DTT (diami-de + DTT) as indicated. (B) Western analysis of the SrAOX proteinsextracted from fresh frozen thermogenic florets. An aliquot of wholetissue extracts was prepared, resolved on a 12.5% non-reducing SDS–PAGE gel and probed with the AOA monoclonal antibody. Thepositions of the molecular weight standards are indicated on the left ofeach panel. OX., the oxidized, disulfide linked form of the SrAOXprotein; Red., the reduced form of the SrAOX protein. Typical resultsare shown, representing three independent Western analyses withseparate mitochondrial preparations.
found to exist as a reduced form, with only weak signals evi-
dent that corresponded to an oxidized dimer (Fig. 2A). How-
ever, when the mitochondria were incubated with DTT, these
low levels of oxidized species were almost completely con-
verted to a reduced form. Treatment of mitochondria with dia-
mide resulted in the partial oxidation of the SrAOX protein
pool but not all of the reduced species were converted to their
oxidized counterparts. Consequently, the treatment of mito-
chondria with diamide followed by DTT resulted in the oxi-
dized form of SrAOX being almost totally converted to its
reduced counterpart (Fig. 2A).
In our experiments, an ascending concentration series of dia-
mide (5–250 mM) indicated that the levels of reduced AOX re-
mained almost equivalent, even when mitochondria were
treated with higher concentrations of this compound (data
not shown). Because our mitochondrial preparations from
thermogenic florets were purified in the presence of 2 mM
pyruvate, we assumed that this was caused by the protective ef-
fects of pyruvate against the oxidation of the AOX regulatory
sulfhydryl/disulfide system [13,31,32]. To further determine the
oxidation/reduction status of SrAOX protein, we conducted
additional experiments using mitochondrial isolates purified
with buffers in which pyruvate was omitted. Our data again
showed that a large proportion of the SrAOX protein was
not oxidized by diamide (data not shown). These results sug-
gest that transcripts for another AOX isoform that lacks cys-
teine residues, as shown in maize [33] and tomato [34], are
relatively abundantly expressed in the thermogenic spadix.
Nevertheless, because no other cDNA species except for
SrAOX were identified either in our cloning procedures or
via 3 0-RACE amplifications using primers that recognize con-
served amino acid sequences (see Section 2), it seems unlikely
that another AOX species which is resistant to oxidation by
diamide could be present at appreciable levels in our mito-
chondrial isolates. Although the causal mechanism underlying
this phenomenon remains to be determined, it is probable that
there could be some different type of AOX organization in the
inner membrane of the mitochondrial isolates. Alternatively, it
seems possible that a part of the CysI residues of the reduced
SrAOX dimer had interacted with pyruvate in vivo to form
thiohemiacetal [17,27,31,32], which would prevent the oxida-
tion of SrAOX protein even in buffers in which pyruvate was
excluded.
It has been shown that plant AOXs including SgAOX from
thermogenic tissue of S. guttatum exist as a reduced dimer [35–
37]. To determine the reduction state of the SrAOX protein
in vivo, an extract directly prepared from frozen powdered
thermogenic florets was analyzed by immunoblotting using
non-reducing SDS–PAGE (Fig. 2B). One prominent band at
around 32 kDa corresponding to a reduced dimer of this pro-
tein was detected, but no bands corresponding to the oxidized
form were evident. These data clearly indicate that the SrAOX
protein also exists as a non-covalently associated dimer
in vivo.
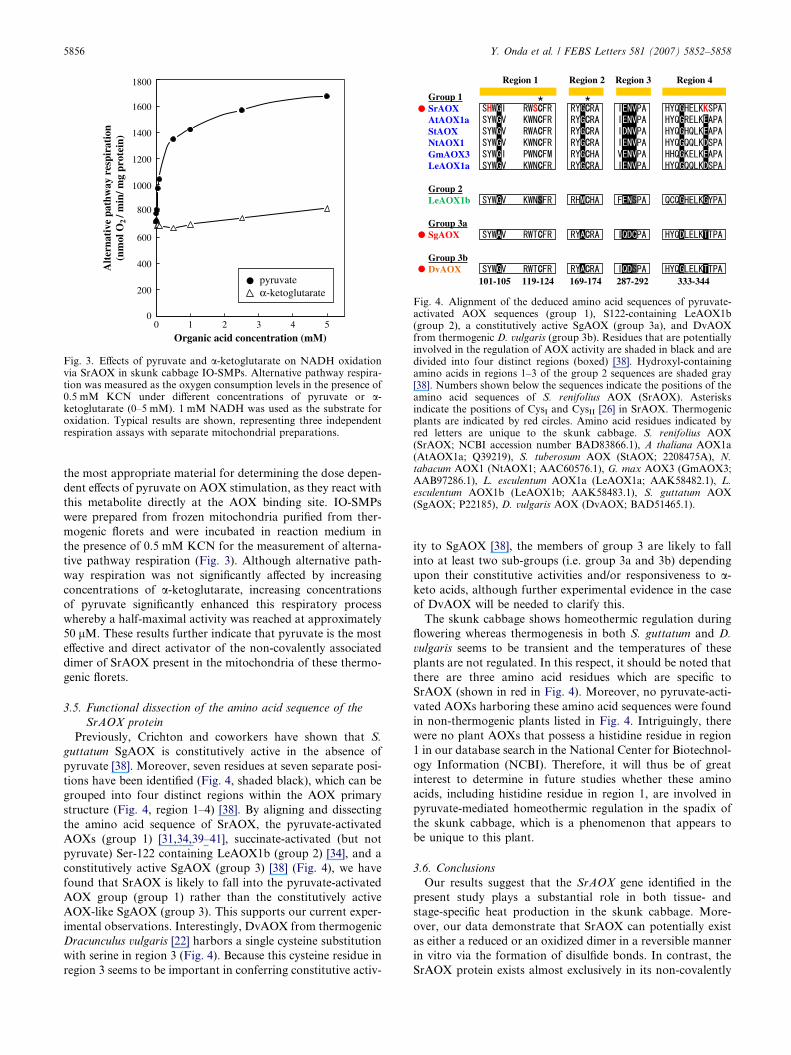
3.4. Effects of pyruvate on the SrAOX protein in IO-SMPs
In general, the effects of pyruvate on AOX are determined
by its addition to intact mitochondria. However, it has been
shown that the external pyruvate concentration may not be
representative of its levels inside the mitochondria [17]. In-
side-out submitochondrial particles (IO-SMPs) are therefore
1 2 3 4 500
200
400
600
800
1000
1200
1400
1600
1800
Alt
erna
tive
pat
hway
res
pira
tion
(nm
ol O
2 / m
in/ m
g pr
otei
n)
Organic acid concentration (mM)
pyruvateα-ketoglutarate
Fig. 3. Effects of pyruvate and a-ketoglutarate on NADH oxidationvia SrAOX in skunk cabbage IO-SMPs. Alternative pathway respira-tion was measured as the oxygen consumption levels in the presence of0.5 mM KCN under different concentrations of pyruvate or a-ketoglutarate (0–5 mM). 1 mM NADH was used as the substrate foroxidation. Typical results are shown, representing three independentrespiration assays with separate mitochondrial preparations.
Fig. 4. Alignment of the deduced amino acid sequences of pyruvate-activated AOX sequences (group 1), S122-containing LeAOX1b(group 2), a constitutively active SgAOX (group 3a), and DvAOXfrom thermogenic D. vulgaris (group 3b). Residues that are potentiallyinvolved in the regulation of AOX activity are shaded in black and aredivided into four distinct regions (boxed) [38]. Hydroxyl-containingamino acids in regions 1–3 of the group 2 sequences are shaded gray[38]. Numbers shown below the sequences indicate the positions of theamino acid sequences of S. renifolius AOX (SrAOX). Asterisksindicate the positions of CysI and CysII [26] in SrAOX. Thermogenicplants are indicated by red circles. Amino acid residues indicated byred letters are unique to the skunk cabbage. S. renifolius AOX(SrAOX; NCBI accession number BAD83866.1), A thaliana AOX1a(AtAOX1a; Q39219), S. tuberosum AOX (StAOX; 2208475A), N.tabacum AOX1 (NtAOX1; AAC60576.1), G. max AOX3 (GmAOX3;AAB97286.1), L. esculentum AOX1a (LeAOX1a; AAK58482.1), L.esculentum AOX1b (LeAOX1b; AAK58483.1), S. guttatum AOX(SgAOX; P22185), D. vulgaris AOX (DvAOX; BAD51465.1).
5856 Y. Onda et al. / FEBS Letters 581 (2007) 5852–5858
the most appropriate material for determining the dose depen-
dent effects of pyruvate on AOX stimulation, as they react with
this metabolite directly at the AOX binding site. IO-SMPs
were prepared from frozen mitochondria purified from ther-
mogenic florets and were incubated in reaction medium in
the presence of 0.5 mM KCN for the measurement of alterna-
tive pathway respiration (Fig. 3). Although alternative path-
way respiration was not significantly affected by increasing
concentrations of a-ketoglutarate, increasing concentrations
of pyruvate significantly enhanced this respiratory process
whereby a half-maximal activity was reached at approximately
50 lM. These results further indicate that pyruvate is the most
effective and direct activator of the non-covalently associated
dimer of SrAOX present in the mitochondria of these thermo-
genic florets.
3.5. Functional dissection of the amino acid sequence of the
SrAOX protein
Previously, Crichton and coworkers have shown that S.
guttatum SgAOX is constitutively active in the absence of
pyruvate [38]. Moreover, seven residues at seven separate posi-
tions have been identified (Fig. 4, shaded black), which can be
grouped into four distinct regions within the AOX primary
structure (Fig. 4, region 1–4) [38]. By aligning and dissecting
the amino acid sequence of SrAOX, the pyruvate-activated
AOXs (group 1) [31,34,39–41], succinate-activated (but not
pyruvate) Ser-122 containing LeAOX1b (group 2) [34], and a
constitutively active SgAOX (group 3) [38] (Fig. 4), we have
found that SrAOX is likely to fall into the pyruvate-activated
AOX group (group 1) rather than the constitutively active
AOX-like SgAOX (group 3). This supports our current exper-
imental observations. Interestingly, DvAOX from thermogenic
Dracunculus vulgaris [22] harbors a single cysteine substitution
with serine in region 3 (Fig. 4). Because this cysteine residue in
region 3 seems to be important in conferring constitutive activ-
ity to SgAOX [38], the members of group 3 are likely to fall
into at least two sub-groups (i.e. group 3a and 3b) depending
upon their constitutive activities and/or responsiveness to a-
keto acids, although further experimental evidence in the case
of DvAOX will be needed to clarify this.
The skunk cabbage shows homeothermic regulation during
flowering whereas thermogenesis in both S. guttatum and D.
vulgaris seems to be transient and the temperatures of these
plants are not regulated. In this respect, it should be noted that
there are three amino acid residues which are specific to
SrAOX (shown in red in Fig. 4). Moreover, no pyruvate-acti-
vated AOXs harboring these amino acid sequences were found
in non-thermogenic plants listed in Fig. 4. Intriguingly, there
were no plant AOXs that possess a histidine residue in region
1 in our database search in the National Center for Biotechnol-
ogy Information (NCBI). Therefore, it will thus be of great
interest to determine in future studies whether these amino
acids, including histidine residue in region 1, are involved in
pyruvate-mediated homeothermic regulation in the spadix of
the skunk cabbage, which is a phenomenon that appears to
be unique to this plant.
3.6. Conclusions
Our results suggest that the SrAOX gene identified in the
present study plays a substantial role in both tissue- and
stage-specific heat production in the skunk cabbage. More-
over, our data demonstrate that SrAOX can potentially exist
as either a reduced or an oxidized dimer in a reversible manner
in vitro via the formation of disulfide bonds. In contrast, the
SrAOX protein exists almost exclusively in its non-covalently

Y. Onda et al. / FEBS Letters 581 (2007) 5852–5858 5857
associated dimer in the thermogenic tissues of skunk cabbage
in vivo. Among the various organic acids that we tested in
our present experiments, including some a-keto acids, only
pyruvate was found to stimulate SrAOX activity at lower con-
centrations. Accordingly, we conclude that the SrAOX protein
exists as a non-covalently associated dimer and is primarily
activated by pyruvate during thermoregulation in the skunk
cabbage.
Acknowledgements: This work was supported by a Grant-in-Aid forScientific Research (B) (Grant No. 18380196) from the Japan Societyfor the Promotion of Science (JSPS), the Program for the Promotionof Basic Research Activities for Innovative Biosciences, and by the21st Century COE program from JSPS. Y.O. is supported by the JSPSResearch Fellowships for Young Scientists.
References
[1] Knutson, R.M. (1974) Heat production and temperature regula-tion in eastern skunk cabbage. Science 186, 746–747.
[2] Seymour, R.S. and Blaylock, A.J. (1999) Switching off the heater:influence of ambient temperature on thermoregulation by easternskunk cabbage Symplocarpus foetidus. J. Exp. Bot. 50, 1525–1532.
[3] Ito, K., Onda, Y., Sato, T., Abe, Y. and Uemura, M. (2003)Structural requirements for the perception of ambient tempera-ture signals in homeothermic heat production of skunk cabbage(Symplocarpus foetidus). Plant Cell Environ. 26, 783–788.
[4] Seymour, R.S. (2004) Dynamics and precision of thermoregula-tory responses of eastern skunk cabbage Symplocarpus foetidus.Plant Cell Environ. 27, 1014–1022.
[5] Guy, R.D., Berry, J.A., Fogel, M.L. and Hoering, T.C. (1989)Differential fractionation of oxygen isotopes by cyanide-resistantand cyanide-sensitive respiration in plants. Planta 177, 483–491.
[6] Berthold, D.A. and Siedow, J.N. (1993) Partial purification of thecyanide-resistant alternative oxidase of skunk cabbage (Symplo-carpus foetidus) mitochondria. Plant Physiol. 101, 113–119.
[7] McIntosh, L. (1994) Molecular biology of the alternative oxidase.Plant Physiol. 105, 781–786.
[8] Day, D.A. and Wiskich, J.T. (1995) Regulation of alternativeoxidase activity in higher plants. J. Bioenerg. Biomembr. 27, 379–385.
[9] Wagner, A.M. and Krab, K. (1995) The alternative respirationpathway in plants: role and regulation. Physiol. Plant. 95, 318–325.
[10] Moore, A.L., Albury, M.S., Crichton, P.G. and Affourtit, C.(2002) Function of the alternative oxidase: is it still a scavenger?Trends Plant Sci. 7, 478–481.
[11] Watling, J.R., Robinson, S.A. and Seymour, R.S. (2006) Contri-bution of the alternative pathway to respiration during thermo-genesis in flowers of the sacred lotus. Plant Physiol. 140, 1367–1373.
[12] Mackenzie, S. and McIntosh, L. (1999) Higher plant mitochon-dria. Plant Cell 11, 571–585.
[13] Umbach, A.L. and Siedow, J.N. (1993) Covalent and noncovalentdimers of the cyanide-resistant alternative oxidase protein inhigher plant mitochondria and their relationship to enzymeactivity. Plant Physiol. 103, 845–854.
[14] Umbach, A.L., Wiskich, J.T. and Siedow, J.N. (1994) Regulationof alternative oxidase kinetics by pyruvate and intermoleculardisulfide bond redox status in soybean seedling mitochondria.FEBS Lett. 348, 181–184.
[15] Millar, A.H., Wiskich, J.T., Whelan, J. and Day, D.A. (1993)Organic acid activation of the alternative oxidase of plantmitochondria. FEBS Lett. 329, 259–262.
[16] Day, D.A., Millar, A.H., Wiskich, J.T. and Whelan, J. (1994)Regulation of alternative oxidase activity by pyruvate in soybeanmitochondria. Plant Physiol. 106, 1421–1427.
[17] Millar, A.H., Hoefnagel, M.H.N., Day, D.A. and Wiskich, J.T.(1996) Specificity of the organic acid activation of alternativeoxidase in plant mitochondria. Plant Physiol. 111, 613–618.
[18] Nie, Z-L., Sun, H., Li, H. and Wen, J. (2006) Intercontinentalbiogeography of subfamily Orontioideae (Symplocarpus, Lysichi-
ton, and Orontium) of Araceae in eastern Asia and NorthAmerica. Mol. Phylogenet. Evol. 40, 155–165.
[19] Ito, K., Kusano, T. and Tsutsumi, K. (1999) A cold-induciblebZIP protein gene in radish root regulated by calcium- andcycloheximide-mediated signals. Plant Sci. 142, 57–65.
[20] Ito, K. (1999) Isolation of two distinct cold-inducible cDNAsencoding plant uncoupling proteins from the spadix of skunkcabbage (Symplocarpus foetidus). Plant Sci. 149, 167–173.
[21] Rhoads, D.M. and McIntosh, L. (1991) Isolation and character-ization of a cDNA clone encoding an alternative oxidase proteinof Sauromatum guttatum (Schott). Proc. Natl. Acad. Sci. USA 88,2122–2126.
[22] Ito, K. and Seymour, R.S. (2005) Expression of uncouplingprotein and alternative oxidase depends on lipid or carbohydratesubstrates in thermogenic plants. Biol. Lett. 1, 427–430.
[23] Douce, R. (1985) American Society of Plant Physiologist Mono-graph Series, Academic Press, pp. 1–44.
[24] Bradford, M.M. (1976) A rapid and sensitive method for thequantification of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72, 248–254.
[25] Elthon, T.E., Nickels, R.L. and McIntosh, L. (1989) Monoclonalantibodies to the alternative oxidase of higher plant mitochon-dria. Plant Physiol. 89, 1311–1317.
[26] Berthold, D.A., Andersson, M.E. and Nordlund, P. (2000) Newinsight into the structure and function of the alternative oxidase.Biochem. Biophys. Acta 1460, 241–254.
[27] Umbach, A.L. and Siedow, J.N. (1996) The reaction of thesoybean cotyledon mitochondrial cyanide-resistant oxidase withsulfhydryl reagents suggests that a-keto acid activation involvesthe formation of a thiohemiacetal. J. Biol. Chem. 271, 25019–25026.
[28] Siedow, J.N., Umbach, A.L. and Moore, A.L. (1995) The activesite of the cyanide-resistant oxidase from plant mitochondriacontains a binuclear iron center. FEBS Lett. 362, 10–14.
[29] Siedow, J.N. and Girvin, M.E. (1980) Alternative RespiratoryPathway: its role in seed respiration and its inhibition by propylgallate. Plant Physiol. 65, 669–674.
[30] Vanlerberghe, G.C., McIntosh, L. and Yip, J.Y.H. (1998)Molecular localization of a redox-modulated process regulatingplant mitochondrial electrontransport. Plant Cell 10, 1551–1560.
[31] Rhoads, D.M., Umbach, A.L., Sweet, C.R., Lennon, A.M.,Rauch, G.S. and Siedow, J.N. (1998) Regulation of the cyanide-resistant alternative oxidase of plant mitochondria: Identificationof the cysteine residue involved in alpha-keto acid stimulation andintersubunit disulfide bond formation. J. Biol. Chem. 273, 30750–30756.
[32] Vanlerberghe, G.C., Yip, J.Y.H. and Parsons, H.L. (1999) Inorganello and in vivo evidence of the importance of the regulatorysulfhydryl/disulfide system and pyruvate for alternative oxidaseactivity in tobacco. Plant Physiol. 121, 793–803.
[33] Karpova, O.V., Kuzmin, E.V., Elthon, T.E. and Newton, J.(2002) Differential expression of alternative oxidase genes in maizemitochondrial mutants. Plant Cell 14, 3271–3284.
[34] Holtzapffel, R.C., Castelli, J., Finnegan, P.M., Millar, A.H.,Whelan, J. and Day, D.A. (2003) A tomato alternative oxidaseprotein with altered regulatory properties. Biochim. Biophys.Acta 1606, 153–162.
[35] Umbach, A.L. and Siedow, J.N. (1997) Changes in the redox stateof the alternative oxidase regulatory sulfhydryl/disulfide systemduring mitochondrial isolation: implications for inferences ofactivity in vivo. Plant Sci. 123, 19–28.
[36] Millar, A.H., Atkin, O.K., Menz, R.I., Henry, B., Farquhar, G.and Day, D.A. (1998) Analysis of respiratory chain regulation inroots of soybean seedlings. Plant Physiol. 117, 1083–1093.
[37] Millenaar, F.F., Benschop, J.J., Wagner, A.M. and Lambers, H.(1998) The role of the alternative oxidase in stabilizing the in vivoreduction state of the ubiquinone pool and the activation state ofthe alternative oxidase. Plant Physiol. 118, 599–607.
[38] Crichton, P.G., Affourtit, C., Albury, M.S., Carre, J.E. andMoore, A.L. (2005) Constitutive activity of Sauromatum guttatumalternative oxidase in Schizosaccharomyces pombe implicatesresidues in addition to conserved cysteines in a-keto acidactivation. FEBS Lett. 579, 331–336.
[39] Djajanegara, I., Holtzapffel, R., Finnegan, P.M., Hoefnagel,M.H.N., Berthold, D.A., Wiskich, J.T. and Day, D.A. (1999) A

5858 Y. Onda et al. / FEBS Letters 581 (2007) 5852–5858
single amino acid change in the plant alternative oxidase alters thespecificity of organic acid activation. FEBS Lett. 454, 220–224.
[40] Hiser, C., Kapranov, P. and McIntosh, L. (1996) Geneticmodification of respiratory capacity in potato. Plant Physiol.110, 277–286.
[41] Vanlerberghe, G.C., Day, D., Wiskich, J.T., Vanlerberghe, A.E.and McIntosh, L. (1995) Alternative oxidase activity in tobaccoleaf mitochondria: dependence on tricarboxylic acid cycle-medi-ated redox regulation and pyruvate activation. Plant Physiol. 109,353–361.












![Drug Delivery System Based on Covalently Bonded Poly[N-Isopropylacrylamide-co-2-Hydroxyethylacrylate]-Based Nanoparticle Networks](https://static.fdokumen.com/doc/165x107/6340d5f6e0dac3b265042228/drug-delivery-system-based-on-covalently-bonded-polyn-isopropylacrylamide-co-2-hydroxyethylacrylate-based.jpg)




![Fluorescent cellulose aerogels containing covalently immobilized (ZnS) ₓ (CuInS ₂) ₁₋ ₓ/ZnS (core/shell) quantum dots [2013]](https://static.fdokumen.com/doc/165x107/63372dc94554fe9f0c05b209/fluorescent-cellulose-aerogels-containing-covalently-immobilized-zns-cuins.jpg)


