
Patterns of growth in coccoid, aggregate forming cyanobacteria
Upload
independentCategory
view
1download
0
Ecotoxicology and Environmental Safety 74 (2011) 1981–1993
Contents lists available at ScienceDirect
Ecotoxicology and Environmental Safety
0147-65
doi:10.1
n Corr
E-m
prabhat
profsmp
journal homepage: www.elsevier.com/locate/ecoenv
Differential physiological and biochemical responses of two cyanobacteriaNostoc muscorum and Phormidium foveolarum against oxyfluorfenand UV-B radiation
Sheeba, Vijay Pratap Singh, Prabhat Kumar Srivastava, Sheo Mohan Prasad n
Ranjan Plant Physiology and Biochemistry Laboratory, Department of Botany, University of Allahabad (A Central University of India), Allahabad (UP) 211002, India
a r t i c l e i n f o
Article history:
Received 16 December 2010
Received in revised form
6 July 2011
Accepted 11 July 2011Available online 28 July 2011
Keywords:
Cyanobacteria
Differential sensitivity
Oxidative stress
Oxyfluorfen
Photosynthesis
UV-B radiation
13/$ - see front matter & 2011 Elsevier Inc. A
016/j.ecoenv.2011.07.006
esponding author. Mobile: þ919451373143.
ail addresses: [email protected] (V.P.
[email protected] (P.K. Srivastava),
[email protected] (S.M. Prasad).
a b s t r a c t
In the present study, degree of tolerance and tolerance strategies of two paddy field cyanobacteria viz.
Nostoc muscorum and Phormidium foveolarum against oxyfluorfen (10 and 20 mg ml�1) and UV-B
(7.2 kJ m�2 d�1) stress were investigated. Oxyfluorfen and UV-B decreased growth, photosynthesis,
nutrient uptake, nitrate reductase, acid and alkaline phosphatase activities, which accompanied with
the increase in the level of oxidative stress. However, growth was more affected in N. muscorum than
P. foveolarum. Antioxidants exhibited differential responses against oxyfluorfen and UV-B stress.
Ascorbate and proline levels were higher in P. foveolarum. A protein of 66 kDa was expressed in
N. muscorum, however, it was absent in P. foveolarum than those of N. muscorum. Besides this, a protein
of 29 kDa appeared in P. foveolarum under all the treatments, but it was present only in control cells of
N. muscorum cells. Overall results indicated resistant nature of P. foveolarum against oxyfluorfen and
UV-B stress in comparison to N. muscorum.
& 2011 Elsevier Inc. All rights reserved.
1. Introduction
Soil and water pollution due to pesticides has become acommon concern among environmentalists. Use of pesticidesbecame indispensible and an integral part of modern agricultureand their use under Integrated Pest Management Programme tosave the crop losses becomes quite decisive in countries like Indiain the wake of second green revolution likely to be experienced innext few years. We cannot rule out the use of pesticides in ouragricultural fields because of steady but continuous rise inpopulation and lesser availability of agricultural fields. But exces-sive and imprudent use of pesticides will definitely lead toenvironmental degradation in several ways. Pesticides that areused in agricultural practices are transported to water bodiesthrough run-off, drift and leaching, and increase the risk ofexposure to non-target organisms (Chen et al., 2007). Oxyfluor-fen(2-chloro-1-(3-ethoxy-4-nitrophenyl)-4-trifluoromethyl)ben-zene) is an ortho-substituted diphenylether, which is a selectiveherbicide and used to control pre- and post-emergent broad leafand grassy weeds in agricultural fields (Ahrens, 1979). Studieshave demonstrated that diphenyl ethers inhibit photosynthesis
ll rights reserved.
Singh),
by affecting electron transport activities in chloroplasts of higherplants (Moreland et al., 1970; Bugg et al., 1980; Pritchard et al.,1980). Bugg et al. (1980) suggested that the site of inhibition inelectron transport by diphenyl ethers is in the region of plasto-quinone and cytochrome f. However, Moreland et al. (1970)suggested that interference with ATP generation could be one ofthe mechanisms of diphenyl ether phytotoxicity. Besides this,oxyfluorfen has the ability to decrease growth indirectly bygenerating reactive oxygen species (ROS) such as singlet oxygen(1O2), superoxide radical (O2
d�), hydrogen peroxide (H2O2) andhydroxyl radicals (dOH) (Geoffroy et al., 2002). Oxyfluorfen haslong residence time (half life 72–160 days) in natural conditions;it persists in fertile layer of soil and harms non-target organismsof soil including cyanobacteria leading to considerable losses incrop yield (Baruah and Mishra, 1986; Yen et al., 2003). Severalstudies demonstrated impact of oxyfluorfen on various metabolicactivities of higher plants; however, impact of oxyfluorfen onmetabolic activities of cyanobacteria is less explored despite theirecological as well as economic importance.
Besides this, incoming solar UV-B radiation may also adverselyaffect cyanobacteria. Solar UV-B radiation comes on the earthsurface due to depletion of stratospheric ozone layer. It isreported that chlorofluorocarbons, which can deplete the ozonelayer, can remain in the upper atmosphere for 40–150 years,hence, the global UV-B radiation will not recover to the levels ofthe pre-industrialization era by 2050, even if all the nations

Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–19931982
implement the Montreal Protocol (World MeteorologicalOrganization, 2007; Mohammed and Tarpley, 2011). Ultraviolet-B radiation affects plants including cyanobacteria from themolecular level to the ecosystem level (He and Hader, 2002). Ithas been reported that UV-B induces decrease in growth andphotosynthesis by enhancing damage to lipids, proteins andnucleic acids (He and Hader, 2002).
Cyanobacteria, in general, are exposed to harsh conditionssuch as extremes of high and low light, UV radiations, heavymetals, salinity, drought, temperature, xenobiotics, etc. andrespond by generating ROS (Gadkari, 1988; Geoffroy et al.,2002; He and Hader, 2002; Prasad and Zeeshan, 2005; Wanget al., 2008). ROS are the inevitable product of oxygen metabo-lism. They may originate in various electron transport chains—
photosynthetic or respiratory (Boger and Sandmann, 1998). Theirleast amount is necessary in cell signaling but when they arepresent in greater amount than needed, they harm variouscomponents of cells—a situation termed as oxidative stress. Boththe stresses—UV-B and oxyfluorfen may also generate ROS inplants often more than required (Geoffroy et al., 2002; He andHader, 2002). These ROS may readily peroxidise membrane lipidsand proteins (He and Hader, 2002; Prasad and Zeeshan, 2005). Sotheir concentration ought to be controlled. In order to controltheir amount, a full-fledged antioxidant system has evolved incellular system.
It is known that cyanobacteria occupy the lowest trophic levelwithin food webs. Therefore, changes in their community mayhave indirect but significant effects on the rest of the fresh watercommunities, nitrogen status and organic matter accumulation(Li et al., 2010). It is also known that organisms differ in theirsensitivity to different stress factors. In modern agriculturaltechnological programs cyanobacteria are utilized to increasethe availability of nitrogen and organic matter in soil for betterproduction of rice in rice producing countries. Nostoc muscorum, aheterocystous cyanobacterium, fixes molecular nitrogen underaerobic condition while Phormidium foveolarum, a non-heterocys-tous strain, fixes nitrogen in darkness. Herbicide and UV-B havebeen reported to inhibit nitrogenase activity (Gadkari, 1988;Sinha et al., 1996). Therefore, for better exploitation of cyano-bacteria as biofertilizer we must know about their degree oftolerance and tolerance strategies to various stresses. Therefore,in the present study, an attempt has been made to investigatethe degree of tolerance of two paddy field cyanobacteria viz.
N. muscorum and P. foveolarum against oxyfluorfen and UV-B byanalyzing different physiological and biochemical parameters.
2. Materials and methods
2.1. Reagents
Oxyfluorfen (trade name ‘‘Oxyflour’’ (EC 23.5%), active ingredient oxyfluorfen
(2-chloro-1-(3-ethoxy-4-nitrophenyl)-4-trifluoromethyl)benzene) used in the
present study was purchased from Divine Tree Limited, Gujarat, India. All
chemicals used in present work were of analytical grade and purchased from
different companies. Bovine serum albumin, 3-(3, 4-dichloro diphenyl)-1,
1-dimethyl urea, methyl viologen, p-benzoquinone and diphenyl carbazide were
purchased from Sigma Aldrich, USA. Methanol, acid ninhydrin, sucrose, hydro-
xylamine, sulphuric acid, toluene, trichloroacetic acid, thiobarbituric acid and
hydrogen peroxide were purchased from Merck, India. Sodium nitrate, K2HPO4,
MgCl2, sodium azide, manganese chloride, potassium nitrate, succinate, Tris–HCl
buffer, potassium nitrite, sulfanilamide, naphthylethylene diamine dihydrochlor-
ide, sodium nitrite, methionine, sulfosalicylic, sodium molybdate, sodium dodecyl
sulphate and polyacrylamide were purchased from Himedia, India. HEPES, NaOH
and sodium carbonate were purchased from Qualigens Fine Chemicals, India. 2,6-
dichlorophenol indophenols and nitroblue tetrazolium chloride were obtained
from LOBA Chemie, Mumbai, India. p-nitrophenyl phosphate was obtained from
CDH, India, riboflavin from Hoffmann-La Roche, Inc., Nutley, New Jersey, USA, and
pyrogallol from S.D. fine-CHEM Limited, India.
2.2. Organisms and culture conditions
The cyanobacteria N. muscorum (filamentous and heterocystous) and
P. foveolarum (filamentous non-heterocystous) were isolated in pure forms from
rice fields of Allahabad city using standard microbiological techniques. Their pure
stocks were maintained in our laboratory. Homogenous cultures of N. muscorum
and P. foveolarum were grown in BG 11 medium without and with nitrogen
(NaNO3 as nitrogen source), respectively, at 2572 1C in culture room under
the photosynthetically active radiation (PAR) of 70 mM photons m�2 s�1 with a
14:10 h light:dark period. All experiments were performed with 7 days old
exponential phase of culture.
2.3. Oxyfluorfen and UV-B treatments
After series of screening experiments with various doses of oxyfluorfen,
effective doses of 10 and 20 mg ml�1of oxyfluorfen were selected for the present
study. These two concentrations were outcome of number of screening experi-
ments ranging from 1 to 100 mg ml�1, toxic to the test cyanobacteria at different
levels. The exponentially grown cyanobacterial cells were harvested by centrifu-
gation at 4000g for 5 min and washed twice with distilled water and then the
pellets were resuspended in BG 11 medium containing 0, 10 and 20 mg ml�1 of
oxyfluorfen. Oxyfluorfen treated and untreated cultures were kept in Petri dishes
under 70 mM photons m�2 s�1 of PAR. Petri dishes were kept on a magnetic
shaker to reduce the cell aggregation and sedimentation and then cultures were
irradiated with artificial UV-B radiation equivalent to 7.2 kJ m�2 d�1, which is
biologically effective. Ultraviolet-B radiation was provided by UV-lamp (TL 40 W/12
Philips, Holland) along with 70 mM photons m�2 s�1 of PAR. The irradiance was
measured with the help of power meter (Spectra physics, USA Model 407, A–2). The
artificial UV-B radiation was equivalent to 15% stratospheric ozone depletion
at Varanasi, an adjoining city of Allahabad, and having fluence rate of 0.4 W m�2.
To filter out UV-C radiation (o280 nm), the lamp was wrapped with 0.127 mm
cellulose acetate film (Johnston Industrial Plastics, Toronto, Canada). After 72 h of
experiment, different parameters were analyzed by harvesting the cyanobacterial
cells. Cyanobacterial cells, which were not treated with oxyfluorfen and UV-B, were
regarded as ‘control’.
2.4. Measurement of growth and extraction of photosynthetic pigments
Growth was measured by estimating dry mass after 72 h of experiment.
Treated and untreated cells (80 ml) were harvested by centrifugation at 4000g for
10 min. The cells were washed thrice with 1 mM of ethylene diaminetetra-acetic
acid (EDTA) to remove surface bound metals. Cells of each sample were oven dried
at 80 1C for 48 h then weighted. For chlorophyll a (Chl a) and carotenoids
estimations, 40 ml of culture [equivalent to 66 mg dry mass (ml culture)�1 and
33 mg dry mass (ml culture)�1, respectively, in unstressed N. muscorum and
P. foveolarum cells] from each sample was centrifuged at 4000g for 10 min, and
pigments from cells were extracted in 100% methanol. The concentrations of Chl a
and carotenoids were quantified by the method of Porra et al. (1989) and Goodwin
(1954), respectively. For phycocyanin determination, cells of each sample were
treated with toluene and extracted with 2.5 mM potassium phosphate buffer
(pH 7.0) after repeated freezing and thawing, and the absorbance of the super-
natant was recorded at 620 nm. The amount of phycocyanin was determined by
the method of Blumwald and Tel-Or (1982).
2.5. Measurements of whole cell photosynthetic O2-evolution and respiration
Photosynthetic activity in terms of whole cell O2-evolution of treated and
untreated cyanobacterial cells was estimated by measuring O2-evolution for 5 min
with a Clark type O2-electrode (Rank Brothers, UK) in a temperature controlled
airtight reaction vessel at 25 1C. The cyanobacterial cells were illuminated by a
projector lamp providing 360 mM photons m�2 s�1 PAR at the surface of vessel.
Respiration rate of treated and untreated cyanobacterial cells was estimated by
measuring the O2-consumption in darkness with the Clark type oxygen electrode
at 25 1C for 5 min.
2.6. Measurements of photosynthetic electron transport activities
Spheroplasts of N. muscorum and P. foveolarum were prepared according to the
method of Spiller (1980) and resuspended in the HEPES-NaOH buffer (pH 6.9)
containing 0.5 M sucrose, 10 mM HEPES-NaOH, 5 mM K2HPO4, 10 mM MgCl2 and
2% (w:v) bovine serum albumin (BSA). The rates of photosystem I (PS I) and whole
photosynthetic chain reaction were measured in terms of O2-consumption.
The 6 ml reaction mixture consisted of buffer (pH 6.9), 10 mM [3-(3,4-dichloro
diphenyl)-1,1-dimethyl urea)] (DCMU), 1 mM sodium ascorbate, 50 mM 2,
6-dichlorophenol indophenol (DCPIP), 50 mM sodium azide (NaN3), 100 mM
methyl viologen (MV) and spheroplasts, which were used for measuring PS I
activity, while for whole photosynthetic chain reaction activity the mixture

Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–1993 1983
consisted of buffer (pH 6.9), 50 mM NaN3, 100 mM MV and spheroplasts. Photo-
system II (PS II) electron transport activity was measured in terms of O2-evolution.
The 6 ml reaction mixture contained buffer (pH 6.9), p-benzoquinone (p-BQ;
1 mM) and spheroplasts. Spectrophotometric assay of PS II activity in terms of
DCPIP photoreduction was also monitored by measuring changes in absorption at
600 nm in the presence of various exogenous electron donors, i.e. hydroxylamine
(NH2OH), diphenyl carbazide (DPC) and manganese chloride (MnCl2).
2.7. Nutrient uptake
The uptake of nitrate (NO3�) was determined by measuring its depletion from
the external medium following the method of Nicholas and Nason (1957). Cells
from each sample (40 ml) were harvested by centrifugation at 4000g for 10 min,
washed twice with double distilled water and resuspended in growth medium
(20 ml) containing 1 mM KNO3. NO3� uptake experiments were carried out under
75 mM photons m�2 s�1 PAR at 25 1C. Cell suspension (10 ml) from each sample
was withdrawn after 4 h of incubation and centrifuged to obtain cell free super-
natant (5 ml). To this, 2 ml of brucine reagent was added and then samples were
kept in ice bath and 5 ml of H2SO4 was added slowly drop by drop. Later on, these
were placed in a water bath at 90 1C for 10 min until the chloroform evaporates.
The color developed was read spectrophotometrically at 410 nm and NO3� present
in the sample was calculated using the standard curve.
The uptake of PO43� was determined by measuring the depletion of PO4
3� from
the external medium. For PO43� uptake, cells (40 ml) from each sample were
harvested by centrifugation at 4000g for 10 min, and resuspended in fresh BG 11
medium containing 100 mM phosphate and incubated at 25 1C under a PAR
of 75 mM photons m�2 s�1. For PO43� uptake experiments, glassware were rinsed
3–4 times with double distilled water to avoid any possibility of presence of PO43� .
Samples were withdrawn after 4 h of incubation, immediately centrifuged and the
cell-free supernatants were analyzed for residual PO43� spectrophotometrically at
660 nm following the method of Ames and Dubin (1960).
2.8. Measurements of acid and alkaline phosphatase and nitrate reductase activities
Acid phosphatase activity (ACP, EC 3.1.3.2) was determined by measuring
formation of p-nitrophenol from substrate p-nitrophenyl phosphate (p-NPP)
according to the method of Torriani (1960). A definite volume (40 ml) of each
sample was harvested, washed and resuspended in 4 ml of succinate buffer
(pH 6.0) and then treated with toluene (1%, v/v) for 30 min to permeabilize the
cells. The reaction was started by the addition of 0.2 ml of p-NPP (final
concentration of 5.2 mM) at 37 1C and terminated by the addition of 1 ml of 1 N
NaOH after 30 min of incubation. Samples were centrifuged and absorbance of
supernatant was recorded at 410 nm and the amount of p-nitrophenol in each
sample was calculated with the help of standard curve.
Alkaline phosphatase activity (ALP, EC 3.1.3.1) was assayed by the determina-
tion of p-nitrophenol formed from the substrate p-nitrophenyl phosphate (p-NPP)
following the method of Ihlenfeldt and Gibson (1975). A known volume (40 ml) of
treated and untreated cells was harvested, washed and resuspended in 4 ml of
0.2 M Tris–HCl buffer (pH 8.5) and then treated with toluene (1%, v/v) for 30 min.
The reaction was initiated by the addition of 0.2 ml of p-NPP (final concentration
of 5.2 mM) at 37 1C and terminated after 30 min of incubation by the addition of
0.2 ml of 1 M K2HPO4. Samples were centrifuged and absorbance of supernatant
was read at 420 nm. The amount of p-nitrophenol formed was calculated with the
help of standard curve.
Nitrate reductase (NR, EC 1.6.6.1) activity was measured by the method of
Camm and Stein (1974). Measurement of NR activity was based on total nitrite
(NO2�) formed. Cyanobacterial suspension (40 ml) was centrifuged, washed thrice
with double distilled water and incubated in growth medium containing 5 mM
KNO3. Samples were withdrawn at desired time and formation of NO2� was
measured by diazo-coupling method of Lowe and Evans (1964). The absorbance of
pink color was read at 540 nm.
2.9. Measurements of superoxide radicals (SOR; O2d�) and hydrogen peroxide (H2O2)
O2d� in each sample was measured by the method of Elstner and Heupel
(1976). This assay is based on the formation of NO2� from hydroxylamine in the
presence of O2d� . Cells obtained from 40 ml culture of each sample were
homogenized in 65 mM potassium phosphate buffer (pH 7.8) and centrifuged at
10,000g for 10 min at 4 1C. The reaction mixture consisted of 65 mM potassium
phosphate buffer (pH 7.8), 10 mM hydroxylamine hydrochloride and cell extract
was incubated for 20 min at 25 1C. After this, 17 mM sulfanilamide and 7 mM
naphthylethylene diamine dihydrochloride (NEDD) were mixed to the incubated
reaction mixture. After 15 min of incubation, diethyl ether was mixed to the same
reaction mixture gently and centrifuged at 2000 g for 5 min. The absorbance of the
colored aqueous phase was recorded at 530 nm. Production of O2d� was calculated
with the help of standard curve prepared with NaNO2� . For H2O2 measurement,
each sample (40 ml) was centrifuged at 4000g for 10 min and the cells were
homogenized in 3.5 ml of 5% (w/v) trichloroacetic acid (TCA) and centrifuged at
10,000g for 15 min. The supernatant obtained was used to analyze total peroxide
by the method of Sagisaka (1976).
2.10. Measurements of lipid peroxidation (as malondialdehyde; MDA content) and
electrolyte leakage
Lipid peroxidation was determined in terms of MDA (malondialdehyde)
content, a product of unsaturated fatty acid peroxidation known as 2-thiobarbi-
turic acid (TBA) reactive metabolite. MDA concentration was estimated by the
method of Heath and Packer (1968). Culture suspension (40 ml) from each sample
was harvested by centrifugation and washed twice in 50 mM phosphate buffer
(pH 7.0). Cells were homogenized in 5% (w/v) TCA. The resulting homogenate was
centrifuged at 10,000g for 10 min. To 0.5 ml of the aliquot of the supernatant, 2 ml
of 20% TCA containing 0.5% (w/v) TBA was added. The mixture was heated at 90 1C
for 20 min and then quickly cooled in ice bath followed by centrifugation.
Absorbance of the supernatant was read at 532 and 600 nm. The value for non-
specific absorption of each sample at 600 nm was subtracted from absorption
recorded at 532 nm. The MDA concentration was calculated using the extinction
coefficient 155 mM�1 cm�1.
The intactness of plasma membrane in cells of each sample was estimated in
terms of electrolyte leakage by the method of Gong et al. (1998). Each sample
(40 ml) was centrifuged at 4000g for 10 min and the cells were washed thoroughly
with double distilled water and placed in test tubes containing 30 ml double
distilled water. The tubes were incubated in a water bath at 30 1C for 2 h and
the initial electrical conductivity (EC1) of supernatant was measured by Con-
ductivity Meter. One of the samples from control set was boiled at 100 1C for
15 min to release all electrolytes, cooled and the final electrical conductivity (EC2)
was measured. The percentage of electrolyte leakage was calculated using the
formula:
EC ¼EC1
EC2� 100ð%Þ:
2.11. Assay of antioxidant enzymes
In vivo catalase (CAT, EC 1.11.1.6) activity was determined polarographically
by the method of Egashira et al. (1989). Cyanobacterial cells (40 ml) were
harvested from different test samples by centrifugation, washed twice with
double distilled water and finally suspended in 50 mM of phosphate buffer
(pH 7.0). In each sample, CAT activity was determined by recording O2-evolution
for 1 min in darkness after the addition of 5 ml of 50 mM phosphate buffer
(pH 7.0) containing 50 mM H2O2. To this, 1 ml of cell suspension was added
in darkness and O2-evolution was monitored by Clark type O2-electrode
(Rank Brothers, UK). Temperature around the vessel was maintained at 25 1C.
One unit of CAT is the amount of the enzyme producing 1 nM O2 min�1.
Superoxide dismutase (SOD, EC 1.15.1.1) activity was assayed by the method
of Giannopolitis and Ries (1977). Cells collected from each sample (40 ml) by
centrifugation were washed twice with 100 mM EDTA–phosphate buffer (pH 7.8).
Cells were homogenized in an ice cold 100 mM EDTA–phosphate buffer (pH 7.8).
The homogenate was centrifuged for 20 min at 10,000g. The supernatant fraction
was used as the enzyme source. The 3 ml reaction mixture contained 1.3 mM
riboflavin, 13 mM L-methionine, 0.05 M Na2CO3, (pH 10.2), 63 mM p-nitroblue
tetrazolium chloride (NBT) and 0.1 ml of enzyme extract. Reaction was carried
out for 15 min in similar test tubes at 25 1C under an illumination of
75 mM photons m�2 s�1. The initial rate of reaction as measured by the difference
in the increase in absorbance at 560 nm in the presence and absence of extract
was proportional to the amount of enzyme. The unit of SOD activity is defined as
the amount of enzyme, which caused a 50% inhibition of the reaction observed in
the absence of enzyme. For the blank, the reaction was run in darkness.
Peroxidase (POD, EC 1.11.1.7) activity was determined by the method of
Gahagen et al. (1968). Cells obtained from 40 ml culture suspension of each
sample were homogenized in 2 ml 100 mM phosphate buffer (pH 7.0) at 5 1C. The
homogenate was centrifuged at 10,000g for 30 min. The supernatant was con-
sidered as the enzyme extract. The 3 ml reaction mixture consisted of 1 ml 25 mM
H2O2, 1 ml 100 mM pyrogallol and 1 ml enzyme extract. After thorough mixing
of the reaction mixture, change in optical density was monitored at 430 nm for
2–3 min.
2.12. Measurement of ascorbate and proline
Ascorbate content in cells of each sample was estimated by the method of
Oser (1979). Cells (40 ml) were homogenized in 2 ml of 5% (w/v) sulfosalicylic
acid, and then the homogenate was centrifuged at 10,000g for 10 min. The
reaction mixture consisted of 2 ml 2% (w/v) sodium molybdate, 2 ml 0.15 N
H2SO4, 1 ml 1.5 mM K2HPO4 and 1 ml crude extract. The reaction mixture was
incubated at 60 1C in water bath for 40 min, cooled, centrifuged at 8000g for
10 min and the absorbance was recorded at 660 nm. The amount of ascorbate was
calculated by comparing absorbance with standard curve.

Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–19931984
Proline content was estimated according to the method of Bates et al. (1973).
Cells obtained from 40 ml culture suspension of each sample were crushed in 3%
(w/v) sulfosalicylic acid, centrifuged at 10000g and 1 ml of this extract was mixed
with 3% (v/v) glacial acetic acid and acid ninhydrin solution. Samples were heated
for 1 h in a water bath maintained at 95 1C, cooled and extracted with 4 ml toluene
by vortexing for 5 min with a test tube mixer. The toluene layer was taken for
recording the absorbance at 520 nm using toluene as blank. The proline content in
each sample was calculated with the help of standard curve.
2.13. SDS-PAGE analysis of proteins
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) of
both the cyanobacterial proteins was done on 12% polyacrylamide resolving
gel and the separation of individual protein was carried out in a vertical gel
electrophoresis system (Perfit, India) with gels of 160�120 mm�2 and 1 mm
thick, according to the method of Laemmli (1970).
2.14. Statistical analysis
All the experiments were conducted in triplicate and repeated at least thrice to
confirm the reproducibility of the results. A one-way ANOVA test was applied to
confirm the significance of data. Comparison between control and treatment’s
means was carried out using Duncan’s multiple range test (DMRT) at Po0.05
significance level and different parameters were correlated by regression analysis
in which growth (dry mass) was regarded as independent variable. SPSS-10
software was used for DMRT and regression analysis.
3. Results and discussion
3.1. Growth, pigments, photosynthesis and respiration
Oxyfluorfen showed (differential) inhibitory effects on both thecyanobacteria as indicated by decreased biomass accumulation,
Table 1Effect of oxyfluorfen and UV-B alone and together on photosynthetic pigments in N. m
Treatments Photosynthetic pigments (mg (ml culture)�1)
N. muscorum
Chlorophyll a Carotenoids Phycocyanin
Oxyfluorfen (mg/ml)
0 1.1470.051a 0.39670.020a 7.3070.33a
10 0.71470.032 (�37)c 0.27670.014 (�30)c 3.8070.17 (�48
20 0.59570.030(�48)d 0.24070.011 (�39)d 3.2770.15 (�55
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 0.91870.037 (�19)b 0.34270.016 (�14)b 5.2370.24 (�28
10 0.32370.013 (�72)e 0.17470.009 (�56)e 1.2070.064 (�8
20 0.22170.011 (�81)f 0.13870.006 (�65)f 0.87270.040 (�8
Data are means7SE of three independent experiments. Values with different letter
Duncan’s multiple range test. Values in parentheses indicate percent decrease (�) ove
Table 2Effect of oxyfluorfen and UV�B alone and together on photosynthetic activity and res
Treatments N. muscorum
Photosynthetic activity (mM O2
evolved (mg protein)�1 h�1)
Respiration (mM O2
consumed (mg protein)�1 h�1
Oxyfluorfen (mg/ml)
0 18.570.85a 2.770.12ab
10 9.970.51 (�46)c 3.070.14 (þ11)ab
20 8.370.40 (�55)d 3.170.13 (þ15)ab
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 13.970.71 (�25)b 3.070.16 (þ11)ab
10 6.670.30 (�64)e 3.270.17 (þ19)a
20 5.570.29 (�70)f 3.570.17 (þ30)a
Data are means7SE of three independent experiments. Values with different letters w
multiple range test. Values in parentheses indicate percent decrease (�) or increase (
photosynthetic pigments and photosynthetic activities (Tables 1–5;Fig. 1). Exposure of 10 and 20 mg ml�1 oxyfluorfen caused reductionin dry masses by 41% and 50% in N. muscorum and only by 6% and15% in P. foveolarum, respectively, in comparison to their respectivecontrols (Fig. 1). These reductions were site manifestation of variouspigments (Chl a, carotenoids, phycocyanin) and physiologicalchanges (Tables 1–8, Figs. 1–3 and 6). Chlorophyll biosynthesis iscatalyzed by several enzymes in multiple steps. Out of these, in thelast step protoporphyrinogen IX is converted to protoporphyrin IX,which is catalyzed by protoporphyrinogen IX oxidase (EC 1.3.3.4)(Jacobs and Jacobs, 1987; Matringe et al., 1992; Boger andSandmann, 1998; Geoffroy et al., 2003). Proper activity of thisprotoporphyrinogen IX oxidase is necessary for chlorophyll biosynth-esis (Jung, 2011). Oxyfluorfen (chemically diphenyl ether) is aperoxidizing herbicide, which has been reported to alter the activityof enzyme protoporphyrinogen oxidase (Duke et al., 1991). Theinhibition in pigment synthesis due to alteration in protoporphyr-inogen oxidase activity may disturb the function of light harvestingcomplex and energy transfer within photosystems (Karapetyan et al.,1983), which ultimately results in alterations in photosyntheticelectron transport reactions as indicated by inhibition in PS II, PS Iand whole photosynthetic electron transport chain reactions(Table 3). Further, the alteration in protoporphyrinogen IX oxidaseactivity will cause an accumulation of protoporphyrinogen IX, whichis oxidized to protoporphyrin IX by non-specific peroxidases (Aizawaand Brown, 1999). Protoporphyrin IX has strong photosensitizingproperty and is responsible for generation of ROS and causingoxidative stress (Boger and Sandmann, 1998; Aizawa and Brown,1999). Besides protoporphyrin IX-mediated generation of ROS, over-reduction of photosynthetic electron transport chain due to herbicidemay also generate ROS by spilling out electrons and giving it to
uscorum and P. foveolarum.
P. foveolarum
Chlorophyll a Carotenoids Phycocyanin
0.53670.027a 0.25670.012a 4.3570.20a
)c 0.40870.019 (�24)c 0.20870.009 (�19)c 2.8670.14 (�34)c
)d 0.36870.018 (�31)d 0.20070.010 (�22)cd 2.5870.13 (�41)d
)b 0.51270.026 (�4)ab 0.22470.012 (�13)b 4.0170.18 (�8)b
4)e 0.40070.020 (�25)c 0.20870.010 (�19)c 2.7270.14 (�37)c
8)ef 0.34470.018 (�36)e 0.17270.009 (�33)e 2.4570.13 (�44)de
s within the same column are significantly different (Po0.05) according to the
r control value.
piration in N. muscorum and P. foveolarum.
P. foveolarum
)
Photosynthetic activity
(mM O2 evolved (mg protein)�1 h�1)
Respiration (mM O2
consumed (mg protein)�1 h�1)
14.170.68a 1.670.072a
10.070.52 (�29)c 1.870.091 (þ13)a
9.170.41 (�35)cd 1.970.087 (þ19)a
11.670.63 (�18)b 1.870.094 (þ13)a
8.570.43 (�40)d 1.870.077 (þ13)a
7.970.41 (�44)de 2.070.098 (þ25)a
ithin same column are significantly different (Po0.05) according to the Duncan’s
þ) over control value.

Table 4Restoration of photosystem II activity (PS II; mmol DCPIP reduced (mg Chl a)�1 h�1) in spheroplasts prepared from N. muscorum exposed to oxyfluorfen and UV-B
radiation.
Treatments PS II activity (mM DCPIP reduced (mg Chl a)�1 h�1) with donors
Without donor DPC NH2OH MnCl2
Oxyfluorfen (mg/ml)
0 90.073.8a 110.075.2a 104.075.0a 95.074.4a
10 77.474.0 (�14)b 106.775.0 (�3)ab 102.074.9 (�2)a 90.274.7 (�5)b
20 71.073.7 (�21)c 103.475.0 (�6)b 99.875.2 (�4)ab 85.574.1 (�10)c
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 78.373.6 (�13)b 101.274.7 (�8)b 97.875.1 (�6)b 91.274.2 (�4)b
10 65.773.0 (�27)d 94.674.8 (�14)c 94.674.7 (�9)bc 80.773.9 (�15)d
20 62.173.2 (�31)de 91.374.7 (�17)c 92.674.3 (�11)c 76.974.0 (�19)de
All the values are means7SE of three independent experiments. Values with different letters within same column are significantly different (Po0.05) according to the
Duncan’s multiple range test. Values in parentheses indicate percent decrease (�) over control value.
Table 5Restoration of photosystem II activity (PS II; mmol DCPIP reduced (mg Chl a)�1 h�1) in spheroplasts prepared from P. foveolarum exposed to oxyfluorfen and UV-B
radiation.
Treatments PS II activity (mM DCPIP reduced (mg Chl a)�1 h�1) with donors
Without donor DPC NH2OH MnCl2
Oxyfluorfen (mg/ml)
0 80.073.4a 95.074.4a 89.074.1a 84.074.7a
10 72.873.5 (�9)b 93.174.3 (�2)ab 87.273.9 (�2)ab 80.674.2 (�4)ab
20 68.073.3 (�15)c 87.474.5 (�8)c 83.774.0 (�6)c 75.673.7 (�10)b
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 73.673.5 (�8)b 93.174.8 (�2)ab 87.274.2 (�2)ab 80.673.2 (�4)ab
10 66.473.1 (�17)c 82.674.0 (�13)d 80.174.0 (�10)cd 70.573.4 (�16)c
20 59.272.9 (�26)d 79.674.3 (�16)de 78.373.8 (�12)cd 68.973.4 (�18)cd
All the values are means7SE of three independent experiments. Values with different letters within same column are significantly different (Po0.05) according to the
Duncan’s multiple range test. Values in parentheses indicate percent decrease (�) over control value.
Table 3Effect of oxyfluorfen and UV�B radiation alone and together on photosynthetic electron transport activities in N. muscorum and P. foveolarum.
Treatments Photosynthetic electron transport activities
N. muscorum P. foveolarum
PS I PS II Whole chain PS I PS II Whole chain
Oxyfluorfen (mg/ml)
0 620727.9a 360716.2a 205710.7a 540723.2a 320713.8a 19078.7a
10 607725.5 (�2)ab 306715.6 (�15)b 16677.6 (�19)b 529725.4 (�2)ab 288715.6 (�10)b 16177.4 (�15)b
20 601727.0 (�3)ab 288712.4 (�20)c 15577.4 (�24)bc 523727.7 (�3)ab 268711.3 (�16)c 15277.0 (�20)bc
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 607728.5 (�2)ab 306714.4 (�15)b 16478.4 (� 20)b 529724.3 (�2)ab 291713.1 (�9)b 15277.3 (�20)bc
10 595730.9 (�4)ab 270712.4 (�25)cd 14377.4 (�30)c 518724.9 (�4)ab 256713.3 (�20)cd 14276.5 (�25)cd
20 582729.7 (�6)b 255713.3 (�29)d 13375.6 (�35)d 507723.3 (�6)b 243712.6 (�24)d 13375.6 (�30)d
All the values are means7SE of three independent experiments. Values with different letters within the same column are significantly different (Po0.05) according to the
Duncan’s multiple range test. Values in parentheses indicate percent decrease (�) over control value. Photosystem I activity (PS I; mM O2 consumed (mg Chl a)�1 h�1),
photosystem II activity (PS II; mM O2 evolved (mg Chl a)�1 h�1) and whole chain electron transport activity [mM O2 consumed (mg Chl a)�1 h�1).
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–1993 1985
molecular oxygen (O2) instead of NADPþ (Boger and Sandmann,1998). The formation of ROS may further trigger lipid peroxidation,which then alters the structure of thylakoid membrane and itsfluidity, which may again reduce photosystem-mediated electrontransport efficiency as we report in our study. Alteration in mem-brane fluidity due to oxyfluorfen treatment is supported by the dataof electrolyte leakage (Table 9). Pritchard et al. (1980) have alsoreported oxyfluorfen-mediated electrolyte leakage and was corre-lated with one of the sites of oxyfluorfen action. Our results alsoshowed that oxyfluorfen significantly increased O2
d� and H2O2
production, which was accompanied by a significant increase in lipidperoxidation and electrolyte leakage (Tables 8 and 9). Thus, results
showed that oxyfluorfen directly affects growth of both the cyano-bacteria by inhibiting chlorophyll synthesis and electron transportactivities, and indirectly by generating ROS and oxidative damage.Other herbicide 2,4-dichlorophynoxyacetic acid (2,4-D) has also beenreported to decrease growth of cyanobacterium Anabaena fertilissima
by decreasing the level of photosynthetic pigments and activities ofnitrate reductase (Kumar et al., 2010). Furthermore, Li et al. (2010)have reported that insecticide acetamiprid reduces photosyntheticperformance of the cyanobacterium Synechocystis sp. by inhibitingelectron transport and inactivating reaction centers. Lesser negativeimpact of oxyfluorfen on P. foveolarum than N. muscorum may beattributed to lesser damage on pigments and photosynthetic activity

Gro
wth
[µg
dry
mas
s (m
l cul
ture
)-1]
0
20
40
60
80
0
9
18
27
36
a
b
c
d
e
f
ab b
ccd
d
- UV-B
0 µg/ml 10 µg/ml 20 µg/ml
+ UV-B
Fig. 1. Effects of oxyfluorfen and UV-B radiation alone and together on growth of
N. muscorum (A) and P. foveolarum (B). Data are means7SE of three independent
experiments. Mean values followed by different letters on bars are significantly
different (Po0.05) according to the Duncan’s multiple range test.
Table 6Effect of oxyfluorfen and UV-B alone and together on NO3
� uptake [mM NO3 (g dry m
P. foveolarum.
Treatments N. muscorum
NO3� PO4
3�
Oxyfluorfen (mg/ml)
0 223.0710.7a 158.077.4a
10 103.075.2 (�54)c 135.076.5 (�1
20 92.074.2 (�59)cd 117.075.5 (�2
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 188.079.8 (�16)b 129.076.2 (�1
10 83.074.4 (�63)e 84.074.0 (�4
20 67.072.7 (�70)f 49.072.5 (�6
Data are means7SE of three independent experiments. Values with different letters w
multiple range test. Values in parentheses indicate percent decrease (�) over control
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–19931986
due to the lesser accumulation of ROS, hence lesser oxidative damage(MDA and electrolyte leakage) as shown by their respective regres-sion curves (Figs. 4 and 5).
UV-B also caused drastic decrease in biomass accumulation,pigments and photosynthesis (Fig. 1; Tables 1–3). Exposure ofUV-B caused a reduction in dry mass by 15% in N. muscorum andby 6% in P. foveolarum in comparison to control (Fig. 1). Reductionin dry mass has further increased when the two cyanobacteriawere given the combined treatments of oxyfluorfen and UV-B.Treatment of 20 mg ml�1 oxyfluorfenþUV-B led to a dry massreduction of 77% in N. muscorum while it was able to cause only24% reduction in P. foveolarum in comparison to control, provingP. foveolarum more tolerant against these two stresses in compar-ison to N. muscorum (Fig. 1). It is well known that increased UV-Bradiation can directly decrease growth and survival and causephoto-bleaching of photosynthetic pigments (He and Hader,2002; Gao et al., 2007). UV-B radiation is reported to decreasephotosynthetic parameters such as oxygen evolution, photosyn-thetic activity and 14CO2 fixation (Sinha and Hader, 2008). Thedecrease in these processes has been linked with UV-B-mediatedalterations in light harvesting complex and damage to oxygenevolving complex, and D1 and D2 polypeptides of PS II reactioncenter (Greenberg et al., 1989). UV-B can also indirectly affectvarious physiological and biochemical processes by producingexcess ROS resulting into oxidative stress (He and Hader, 2002;Wang et al., 2008). UV-B can induce lipid peroxidation andprotein oxidation of biological membranes, DNA damage andhormone inactivation (He and Hader, 2002; Wang et al., 2008).Prasad and Zeeshan (2004) have also reported that UV-B inducesreduction in biomass to which they have associated the reductionin chlorophyll. Mitchell and Karentz (1993) have showed thatUV-B radiation caused damage in growth, which was due toirreversible damage to DNA. Thus, UV-B-mediated reduction ingrowth of both the cyanobactria may be correlated withdecreased photosynthetic pigments and thereby decreased photo-synthetic activities (Fig. 1; Tables 1–5).
Results related to photosynthetic pigments in N. muscorum
and P. foveolarum are shown in Table 1. Oxyfluorfen and UV-Binduced the reduction in pigments in both the cyanobacteria,however, pigments were severely affected in N. muscroum thanP. foveolarum. Comparing the effects of oxyfluorfen and UV-B onphotosynthetic pigments, phycocyanin was the most affected andcarotenoids were the least affected (Table 1). It is known thatphotosynthetic pigments such as Chl a and carotenoids areintegrated in the membrane, while phycocyanin is found attachedto the outer surface of the thylakoid membrane. Because of thisreason, phycocyanin appeared to be highly sensitive to oxyfluor-fen and UV-B. Phycocyanins are major light harvesting pigments
ass)�1 h�1) and PO4�3 uptake (mM PO4
3 (g dry mass)�1 h�1) in N. muscorum and
P. foveolarum
NO3� PO4
3�
184.078.3a 148.077.5a
5)b 160.078.3 (�13)c 107.075.1 (�28)c
6)c 113.075.9 (�39)e 95.074.6 (�36)d
8)b 199.079.2 (�8)b 139.077.2 (�6)b
7)d 137.076.3 (�26)d 94.074.9 (�36)d
9)e 97.075.0 (�47)f 92.074.3 (�38)d
ithin same column are significantly different (Po0.05) according to the Duncan’s
value.

Table 7Effect of oxyfluorfen and UV-B alone and together on activities of acid phosphatase (ACP; nM p-NP formed (mg protein)�1 min�1), alkaline phosphatase (ALP; nM p-NP
formed (mg protein)�1 min�1] and nitrate reductase (NR; mM NO2� produced (g protein)�1 h�1) in N. muscorum and P. foveolarum.
Treatments Enzyme activities
N. muscorum P. foveolarum
ACP ALP NR ACP ALP NR
Oxyfluorfen (mg/ml)
0 4.6470.22a 8.8070.35a 35.071.6b 3.1070.16a 11.270.58a 59.773.2c
10 3.0470.15 (�34)c 5.7270.27 (�35)c 30.871.6 (�12)c 2.2170.11 (�29)c 9.4470.46 (�16)c 70.473.6 (þ18)a
20 2.8070.14 (�40)d 5.1070.27 (�42)cd 29.571.4 (�16)c 2.1270.10 (�32)cd 8.6470.39 (�23)d 71.573.2 (þ20)a
Oxyfluorfen (mg/ml) þ 7.2 kJ m�2 d�1 (UV-B)
0 3.6870.19 (�21)b 7.7070.35 (�13)b 44.072.3 (þ26)a 2.4470.13 (�21)b 10.170.53 (�10)b 69.373.3 (þ16)a
10 2.1670.11 (�53)e 2.9170.14 (�67)e 27.571.3 (�21)cd 2.0270.084 (�35)d 8.3270.42 (�26)de 65.173.5 (þ9)b
20 1.7670.089 (�62)f 2.5370.14 (�71)ef 14.070.64 (�60)e 1.7470.083 (�44)e 7.3670.33 (�34)e 40.572.2 (�32)d
All the values are means7SE of three independent experiments. Values with different letters within same column are significantly different (Po0.05) according to the
Duncan’s multiple range test. Values in parentheses indicate percent decrease (�) or increase (þ) over control value.
Table 8Effect of oxyfluorfen and UV-B alone and together on level of superoxide radicals (SOR; nM (mg dry mass)�1) and hydrogen peroxide (H2O2; mM (g dry mss)�1) in
N. muscorum and P. foveolarum.
Treatments N. muscorum P. foveolarum
SOR H2O2 SOR H2O2
Oxyfluorfen (mg/ml)
0 11.970.57f 28.071.3d 12.470.56cd 39.272.1e
10 14.370.74 (þ20)d 36.471.7 (þ30)c 16.770.77 (þ35)b 43.972.1 (þ12)d
20 15.370.64 (þ29)c 40.072.1 (þ43)b 17.670.84 (þ42)ab 48.572.0 (þ24)c
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 12.770.58 (þ7)e 36.871.9 (þ31)c 13.070.67 (þ5)c 44.872.2 (þ14)d
10 17.370.90 (þ45)b 40.072.2 (þ43)b 17.370.83 (þ40)b 51.372.7 (þ31)b
20 18.170.85 (þ52)a 46.471.9 (þ66)a 18.170.85 (þ46)a 58.373.1 (þ49)a
All the values are means7SE of three independent experiments. Values with different letters within same column are significantly different (Po0.05) according to the
Duncan’s multiple range test. Values in parentheses indicate percent increase (þ) over control value.
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–1993 1987
and reserves of nitrogen in cyanobacteria (Cohen-Bazire andBryant, 1982). Thus, the steeper decline in phycocyanin inN. muscorum suggests severe destruction in growth in comparisonto P. foveolarum as justified by data of dry mass (Fig. 1; Table 1).
The essential function of carotenoids in protecting the photo-synthetic system from photo-oxidative damage is well documen-ted and the carotenoids of the xanthophyll cycle play a major rolein photoprotection (Carletti et al., 2003). Carotenoids wereobserved to be least affected pigment in comparison to Chl a
and phycocyanin for the similar treatments (except in UV-B aloneexposed P. foveolarum cells). Lesser reduction in carotenoids(in comparison to Chl a, phycocyanin and dry mass) is suggestingtheir protective role against oxyfluorfen and UV-B stress. Carotenoidsplays major role in directly quenching singlet oxygen (1O2), since noantioxidant has been reported to directly quench 1O2 (Siefermann-Harms, 1987).
Results pertaining to photosynthetic activities are shown inTables 2–5. We report that exposure of both the cyanobacteriato oxyfluorfen and UV-B led to a reduction in whole cellO2-evolution and photosynthetic electron transport activities (PSII, PS I and whole chain) in comparison to control (Tables 2 and 3).Again, these processes were more affected in N. muscorum incomparison to P. foveolarum. Under oxyfluorfen and UV-B treat-ments decrease in whole cell O2-evolution can directly becorrelated with decreased photosynthetic electron transportactivities (Tables 2 and 3). Among photosynthetic electron trans-port activities, whole chain electron transport activity was com-paratively more affected followed by PS II and PS I underoxyfluorfen and UV-B stress (Table 3). Oxyfluorfen and UV-Bmediated decrease in photosynthetic electron transport activities
can indirectly be correlated with increased level of ROS, lipidperoxidation and electrolyte leakage (Tables 8 and 9). As shown inearlier studies that ROS cause damaging effects on photosyntheticpigments associated with PS II, I and also on water oxidation sideand reaction center complexes (He and Hader, 2002; Prasad andZeeshan, 2004). It is known that oxyfluorfen inhibits the photo-synthetic electron transport system by interacting with electroncarriers of photosynthetic chain reaction (Bugg et al., 1980).Recently, it has been reported that insecticide acetamiprid causedtoxicity in the cyanobacterium Synechocystis sp. by arrestingelectron transfer at acceptor side of PS II (Li et al., 2010).Ultraviolet-B affects photosynthetic electron transport activitiesby damaging the integrity of membrane system, thereby inter-rupting the electron transport across the thylakoid membrane(Renger et al., 1989). Under oxyfluorfen and UV-B stress, com-paratively more decrease in whole chain electron transportactivity suggests that these two stresses also damaged othercomponents of photosynthetic electron transport chain in addi-tion to PS II.
In order to find out possible site of damage in photosyntheticelectron transport chain caused by oxyfluorfen and UV-B, theDCPIP photoreduction was measured in the presence of variousexogenous electron donors—DPC, NH2OH, MnCl2 and comparedwith DCPIP photoreduction in PS II without donor (Tables 4 and 5).Results indicated that all the three exogenous electron donorssuccessfully restored PS II activity but DPC was comparativelymore effective. This shows that the site of damage within PS II liesin between oxygen evolving complex (OEC) and PS II.
Respiration rate showed increasing trend under oxyfluorfenand UV-B treatments (Table 2). Similar results were reported in

- UV-B
POD
act
ivity
[un
its (
mg
prot
ein)
-1 m
in-1
]
0.0
0.1
0.2
0.3
aab
ccd
d
e
POD
act
ivity
[ u
nits
(m
g pr
otei
n)-1
min
-1]
0.0
0.1
0.2
0.3
a aabb b
c
CA
T a
ctiv
ity [
units
(m
g pr
otei
n)-1
min
-1]
0.0
2.5
5.0
7.5
10.0
a
bb
cc
CA
T a
ctiv
ity [
units
(m
g pr
otei
n)-1
min
-1]
0.0
2.5
5.0
7.5
10.0
aab bbb
c
SOD
act
ivity
[un
its (
mg
prot
ein)
-1 m
in-1
]
0
4
8
12
16
20
b
aa
c c
d
0
4
8
12
16
20
dc
a
bcd d
SOD
act
ivity
-1 m
in-1
] [u
nits
(m
g pr
otei
n) 0 µg/ml 10 µg/ml 20 µg/ml
+ UV-B
Fig. 2. Effects of oxyfluorfen and UV-B radiation alone and together on enzymatic antioxidants in N. muscorum (A–C) and P. foveolarum (D–F). Data are means7SE of three
independent experiments. Mean values followed by different letters on bars are significantly different (Po0.05) according to the Duncan’s multiple range test.
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–19931988
the cyanobacterium Plectonema boryanum by Prasad and Zeeshan(2005). Our results indicated that exposure of both the cyano-bacteria to oxyfluorfen and UV-B inhibited photosynthetic elec-tron transport activities (Table 3), thereby reducing the supply ofATP. Thus, under oxyfluorfen and UV-B stress, increased respira-tory rate is to meet out the demand of ATP for carrying out thebasic life processes in the cell.
3.2. NO3� and PO4
3� uptake
NO3� and PO4
3� uptake, both decreased after oxyfluorfen andUV-B treatments, however, these declines were more withN. muscorum (Table 6). Inhibition in nutrient uptake by oxyfluor-fen and UV-B stress may be due to alterations in the membranepermeability as well as cellular homeostasis as evidenced byincreased ROS, lipid peroxidation and electrolyte leakage(Tables 8 and 9). It has been reported that pesticides and UV-Bdecreased nutrient uptake and probable cause was decreasedATP pool as a consequence of damaged photosynthetic electrontransport chain, which led to reduced photosynthesis (Tables 2and 3), thus limiting the availability of assimilatory powers
(NADPH and ATP) (Prasad et al., 2008). It is well known thatavailability of phosphate and nitrate to cyanobacteria is criticalfor their growth. Our results also prove this interlink (Fig. 1;Table 6).
3.3. ACP, ALP and NR activities
Results pertaining to ACP and ALP activities have been pre-sented in Table 7. Exposure of both the cyanobacteria to oxy-fluorfen and UV-B resulted into decreased activities in ACP andALP. However, decrease in these enzyme activities were lower inP. foveolarum in comparison to N. muscorum. Between the twoenzymes, ACP was more affected in P. foveolarum while ALP wasmore affected in N. muscorum (Table 7). Reduced activities ofACP and ALP activities were reported by Prasad et al. (2008) inP. boryanum under endosulfan toxicity. Acid and alkaline phos-phatase activities are used to detect pesticides in the environment(Dyk and Pletschke, 2010). Besides this, ACP and ALP are regardedas the principal enzymes responsible for hydrolysis of polypho-sphates and are shown to be localized around the polyphosphatebodies (Dyk and Pletschke, 2010) and their inhibition may starve
Prol
ine
[nM
(m
g dr
y m
ass)
-1]
Asc
orba
te [
nM (
mg
dry
mas
s)-1
]
0
200
400
600
800
a
b
c ccd cd
0 µg/ml 10 µg/ml 20 µg/ml
Asc
orba
te [
nM (
mg
dry
mas
s)-1
]
0
200
400
600
800b b
c
a
dd
0
3
6
9
12
cb
a
de
- UV-B
b
Prol
ine
[nM
(m
g dr
y m
ass)
-1]
0
3
6
9
12
bc
a
cd
d
b
- UV-B + UV-B+ UV-B
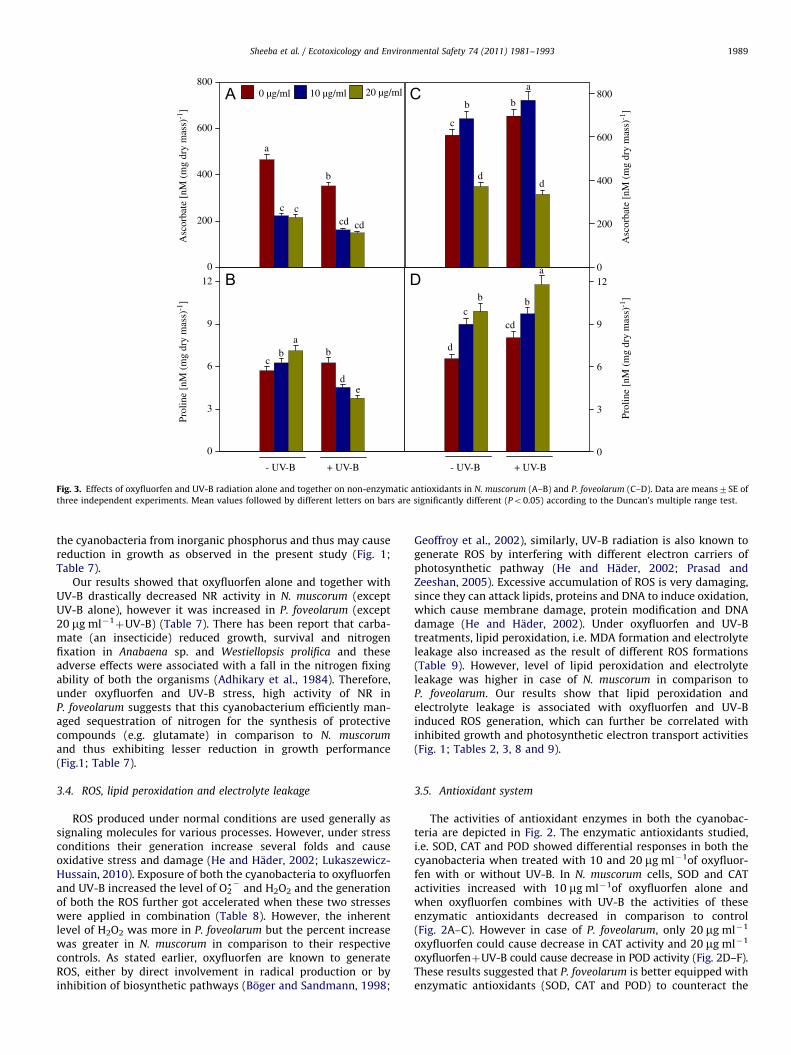
Fig. 3. Effects of oxyfluorfen and UV-B radiation alone and together on non-enzymatic antioxidants in N. muscorum (A–B) and P. foveolarum (C–D). Data are means7SE of
three independent experiments. Mean values followed by different letters on bars are significantly different (Po0.05) according to the Duncan’s multiple range test.
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–1993 1989
the cyanobacteria from inorganic phosphorus and thus may causereduction in growth as observed in the present study (Fig. 1;Table 7).
Our results showed that oxyfluorfen alone and together withUV-B drastically decreased NR activity in N. muscorum (exceptUV-B alone), however it was increased in P. foveolarum (except20 mg ml�1
þUV-B) (Table 7). There has been report that carba-mate (an insecticide) reduced growth, survival and nitrogenfixation in Anabaena sp. and Westiellopsis prolifica and theseadverse effects were associated with a fall in the nitrogen fixingability of both the organisms (Adhikary et al., 1984). Therefore,under oxyfluorfen and UV-B stress, high activity of NR inP. foveolarum suggests that this cyanobacterium efficiently man-aged sequestration of nitrogen for the synthesis of protectivecompounds (e.g. glutamate) in comparison to N. muscorum
and thus exhibiting lesser reduction in growth performance(Fig.1; Table 7).
3.4. ROS, lipid peroxidation and electrolyte leakage
ROS produced under normal conditions are used generally assignaling molecules for various processes. However, under stressconditions their generation increase several folds and causeoxidative stress and damage (He and Hader, 2002; Lukaszewicz-Hussain, 2010). Exposure of both the cyanobacteria to oxyfluorfenand UV-B increased the level of O2
d� and H2O2 and the generationof both the ROS further got accelerated when these two stresseswere applied in combination (Table 8). However, the inherentlevel of H2O2 was more in P. foveolarum but the percent increasewas greater in N. muscorum in comparison to their respectivecontrols. As stated earlier, oxyfluorfen are known to generateROS, either by direct involvement in radical production or byinhibition of biosynthetic pathways (Boger and Sandmann, 1998;
Geoffroy et al., 2002), similarly, UV-B radiation is also known togenerate ROS by interfering with different electron carriers ofphotosynthetic pathway (He and Hader, 2002; Prasad andZeeshan, 2005). Excessive accumulation of ROS is very damaging,since they can attack lipids, proteins and DNA to induce oxidation,which cause membrane damage, protein modification and DNAdamage (He and Hader, 2002). Under oxyfluorfen and UV-Btreatments, lipid peroxidation, i.e. MDA formation and electrolyteleakage also increased as the result of different ROS formations(Table 9). However, level of lipid peroxidation and electrolyteleakage was higher in case of N. muscorum in comparison toP. foveolarum. Our results show that lipid peroxidation andelectrolyte leakage is associated with oxyfluorfen and UV-Binduced ROS generation, which can further be correlated withinhibited growth and photosynthetic electron transport activities(Fig. 1; Tables 2, 3, 8 and 9).
3.5. Antioxidant system
The activities of antioxidant enzymes in both the cyanobac-teria are depicted in Fig. 2. The enzymatic antioxidants studied,i.e. SOD, CAT and POD showed differential responses in both thecyanobacteria when treated with 10 and 20 mg ml�1of oxyfluor-fen with or without UV-B. In N. muscorum cells, SOD and CATactivities increased with 10 mg ml�1of oxyfluorfen alone andwhen oxyfluorfen combines with UV-B the activities of theseenzymatic antioxidants decreased in comparison to control(Fig. 2A–C). However in case of P. foveolarum, only 20 mg ml�1
oxyfluorfen could cause decrease in CAT activity and 20 mg ml�1
oxyfluorfenþUV-B could cause decrease in POD activity (Fig. 2D–F).These results suggested that P. foveolarum is better equipped withenzymatic antioxidants (SOD, CAT and POD) to counteract the

Chlorophyll a
70605040302010
1.2
1.0
0.8
0.6
0.4
0.2
r = 0.993
Carotenoids
70605040302010
0.5
0.4
0.3
0.2
0.1
r = 0.997
Phycocyanin
70605040302010
876543210
r = 0.988
Photosynthetic oxygen yield
70605040302010
201816141210
864
r = 0.982
Respiration
70605040302010
3.6
3.4
3.2
3.0
2.8
2.6
r = -0.923
Photosystem I
70605040302010
630
620
610
600
590
580
r= 0.935
Photosystem II
70605040302010
380
360
340
320
300
280
260
240
r= 0.942
Whole Chain
70605040302010
220
200
180
160
140
120
r= 0.922
Nitrate uptake
70605040302010
300
200
100
0
r= 0.966
Phosphate uptake
70605040302010
180
160
140
120
100
80
6040
r= 0.907
Acid phosphatase
70605040302010
5.0
4.5
4.0
3.5
3.0
2.5
2.0
1.5
r = 0.990
Alkaline phosphatase
70605040302010
1098765432
r = 0.994
Nitrate reductase
70605040302010
50
40
30
20
10
r = 0.834
Superoxide radical
70605040302010
191817161514131211
r = -0.988
Hydrogen peroxide
70605040302010
50
40
30
20
r = -0.910
Lipid peroxidation
GROWTH
70605040302010
2.0
1.8
1.6
1.4
1.2
1.0
0.8
r = -0.946
Electrolyte leakage
70605040302010
40
30
20
10
0
r = -0.969
Superoxide dismutase
70605040302010
16
14
12
10
8
6
r = 0.702
Catalase
70605040302010
13
12
11
10
9
8
r = 0.247
Peroxidase
70605040302010
11
10
9
8
7
6
5
r = 0.590
Ascorbate
70605040302010
500
400
300
200
100
r= 0.971
Proline
70605040302010
7.57.06.56.05.55.04.54.03.5
r= 0.554
Fig. 4. Regression curves for N. muscorum. X-axis represents dry mass, which is an independent variable in all curves, while Y-axis represents different parameters, which
are dependent variables in N. muscorum.
Table 9Effect of oxyfluorfen and UV-B alone and together on level of lipid peroxidation (LPO; nM MDA (mg dry mass)�1) and electrolyte leakage (EL; %) in N. muscorum and
P. foveolarum.
Treatments N. muscorum P. foveolarum
LPO EL LPO EL
Oxyfluorfen (mg/ml)
0 0.8770.042d 6.070.27f 0.7470.038cd 4.070.21e
10 1.1370.054 (þ30)c 17.570.81 (þ192)d 0.8270.043 (þ11)c 10.370.47 (þ158)c
20 1.6770.081 (þ92)ab 25.571.3 (þ325)bc 0.8770.047 (þ18)b 14.070.71 (þ250)b
Oxyfluorfen (mg/ml)þ7.2 kJ m�2 d�1 (UV-B)
0 1.1070.053 (þ26)c 11.070.57 (þ83)e 0.8970.045 (þ20)b 7.370.33 (þ83)d
10 1.7770.092 (þ103)ab 24.071.2 (þ300)b 1.0070.052 (þ35)a 15.070.78 (þ275)b
20 1.9770.083 (þ126)a 33.071.7 (þ450)a 1.1670.049 (þ57)a 18.771.0 (þ368)a
All the values are means7SE of three independent experiments. Values with different letters within same column are significantly different (Po0.05) according to the
Duncan’s multiple range test. Values in parentheses indicate percent increase (þ) over control value.
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–19931990
Chlorophyll a
343230282624
r = 0.856
Carotenoids
343230282624
0.26
0.24
0.22
0.20
0.18
0.16
r = 0.853
Phycocyanin
343230282624
4.5
4.0
3.5
3.0
2.5
2.0
r = 0.838
Photosynthetic oxygen yield
343230282624
r = 0.916
Respiration
343230282624
2.1
2.0
1.9
1.8
1.7
1.6
1.5
r= -0.810
Photosystem I
343230282624r = 0.967
Photosystem II
343230282624
340
320
300
280
260
240
r = 0.981
Whole chain
343230282624
200
190
180
170
160
150
140130
r = 0.889
Nitrate uptake
343230282624r = 0.888
Phosphate uptake
343230282624
150
140
130
120
110
100
9080
r = 0.877
Acid phosphatase
343230282624
r = 0.887
Alkaline phosphatase
343230282624
12
11
10
9
8
7
r = 0.964
Nitrate reductase
343230282624
80
70
60
50
40
r = 0.586
Superoxide radical
343230282624
r = -0.844
Hydrogen peroxide
343230282624
60
50
40
30
r = -0.969
Lipid peroxidation
343230282624r = -0.953
Electrolyte leakage
343230282624
2018161412108642
r= -0.969
Superoxide dismutase
343230282624
6.5
6.0
5.5
5.0
4.5
4.0
r = 0.567
Catalase
343230282624
151413121110987
200
180
160
140
120
100
80
19
18
17
16
15
14
1312
0.4
0.3
0.2
0.1
r = 0.911
Peroxidase
343230282624
0.21
0.20
0.19
0.18
0.17
0.16
0.15
r = 0.277 Ascorbate
343230282624
0.6
0.5
0.4
0.3
550
540
530
520
510
500
3.23.02.82.62.42.22.01.81.6
1.2
1.1
1.0
0.9
0.8
0.7
800
700
600
500
400
300
r = 0.395
Proline
343230282624
12
11
10
9
8
7
6
r = -0.920
Fig. 5. Regression curves for P. foveolarum. X-axis represents dry mass, which is an independent variable in all curves, while Y-axis represents different parameters, which
are dependent variables in P. foveolarum.
Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–1993 1991
adverse effects of oxyfluorfen and UV-B induced ROS and thus hasgreater tolerance in comparison to N. muscorum.
The level of ascorbate drastically decreased in N. muscorum
when cells exposed to single and combined doses of oxyfluorfenand UV-B (Fig. 3A). However, in the case of P. foveolarum, theascorbate level is less affected under oxyfluorfen and UV-Btreatments in comparison to N. muscorum (Fig. 3C). Ascorbate isone of the most studied and powerful non-enzymatic antioxi-dants. It can directly scavenge O2
d� , dOH and reduce H2O2 to watervia ascorbate peroxidase reaction (Noctor and Foyer, 1998).Ascorbate regenerates tocopherol from tocopheroxyl radical pro-viding membrane protection (Noctor and Foyer, 1998). Higheramount of ascorbate in cells is expected to provide greaterprotection to sulphydryl groups—a functional integrity of proteinmolecules. These results indicated that ascorbate level is higherand less affected in P. foveolarum than N. muscorum and can becorrelated with greater tolerance of P. foveolarum. Regressioncurves for ascorbate also show that decrease in growth ispositively correlated in both the cyanobacteria but ‘r’ value is
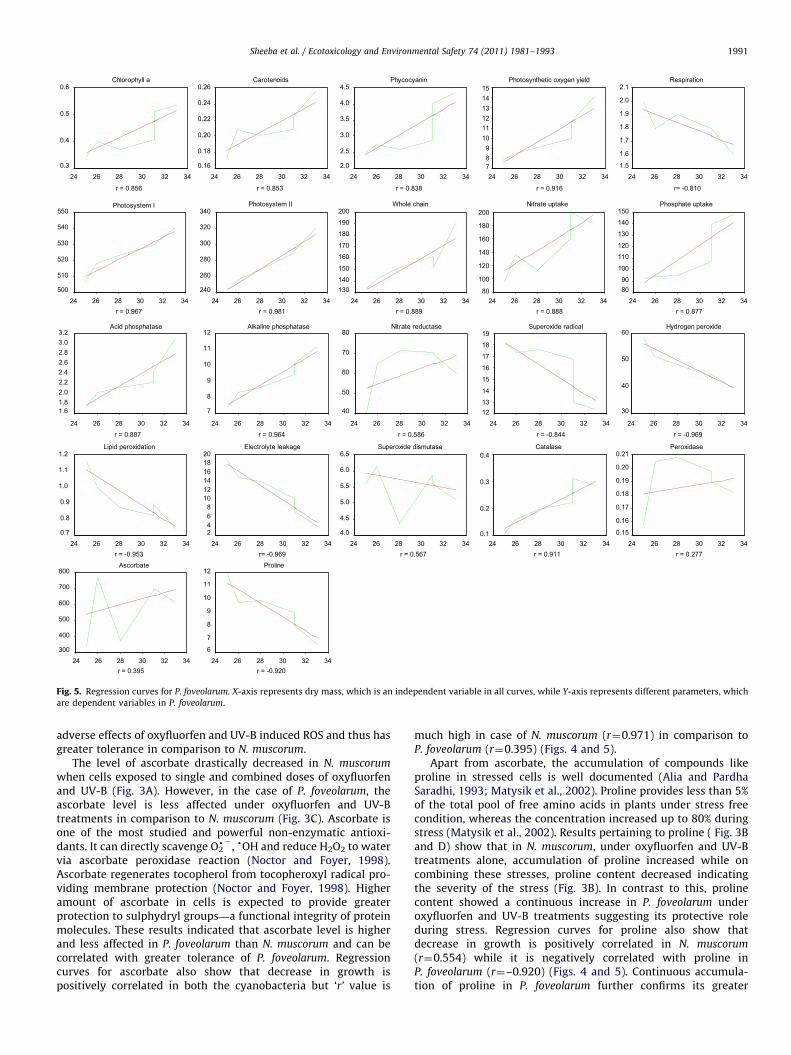
much high in case of N. muscorum (r¼0.971) in comparison toP. foveolarum (r¼0.395) (Figs. 4 and 5).
Apart from ascorbate, the accumulation of compounds likeproline in stressed cells is well documented (Alia and PardhaSaradhi, 1993; Matysik et al., 2002). Proline provides less than 5%of the total pool of free amino acids in plants under stress freecondition, whereas the concentration increased up to 80% duringstress (Matysik et al., 2002). Results pertaining to proline ( Fig. 3Band D) show that in N. muscorum, under oxyfluorfen and UV-Btreatments alone, accumulation of proline increased while oncombining these stresses, proline content decreased indicatingthe severity of the stress (Fig. 3B). In contrast to this, prolinecontent showed a continuous increase in P. foveolarum underoxyfluorfen and UV-B treatments suggesting its protective roleduring stress. Regression curves for proline also show thatdecrease in growth is positively correlated in N. muscorum
(r¼0.554) while it is negatively correlated with proline inP. foveolarum (r¼–0.920) (Figs. 4 and 5). Continuous accumula-tion of proline in P. foveolarum further confirms its greater

Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–19931992
resistance against oxyfluorfen and UV-B stress in comparison toN. muscorum.
3.6. SDS-PAGE analysis
Sodium dodecyl sulphate polyacrylamide gel electrophoresis(SDS-PAGE) analysis showed differential expression of proteins inboth the cyanobacteria under oxyfluorfen and UV-B treatments(Fig. 6). A protein of 29 kDa was detected in P. foveolarum underall treatments of oxyfluorfen and UV-B, which is not present at allin N. muscorum except control cells, suggesting its important rolein normal functioning of physiological processes. Brown et al.(1998) have shown that in plants and animals glutathione-S-transferases (GSTs) have molecular weights in the range of24–29 kDa. Therefore, in the present study, this protein of 29 kDa
Fig. 6. Effects of oxyfluorfen and UV-B radiation alone and together on SDS PAGE
protein profile of N. muscorum (A) and P. foveolarum (B). Lane 1: marker; lane 2:
control; lane 3: 7.2 kJ m�2 d�1 UV-B alone; lane 4: 10 mg ml�1 oxyfluorfen; lane
5: 20 mg ml�1 oxyfluorfen; lane 6: 10 mg ml�1 oxyfluorfenþ7.2 kJ m�2 d�1 UV-B
and lane 7: 20 mg ml�1 oxyfluorfenþ7.2 kJ m�2 d�1 UV-B.
may be ascribed as GST. Glutathione-S-transferases are enzymesthat remove xenobiotics including pesticides and decrease thelevel of oxidative stress in plants as well as in animals (Brownet al., 1998). On the other hand, a protein of 66 kDa appeared in N.
muscorum cells treated with oxyfluorfen and UV-B, which wascompletely absent in P. foveolarum under oxyfluorfen and UV-Btreatments. Ferreira et al. (1996) reported that 66 kDa protein is aphotomodified form of the large subunit of ribulose-1,5-bispho-sphate carboxylase/oxygenase (RUBISCO) enzyme. Thus underoxyfluorfen and UV-B treatments, enormous accumulation of66 kDa protein suggests the modification of RUBISCO enzymethat might have led to a weakened photosynthesis in N. muscorum
in comparison to P. foveolarum (Fig. 6)
4. Conclusion
Our study shows the inhibitory effect of pesticide oxyfluorfenand UV-B on both the cyanobacteria and effect of one stress isaggravated by other stress and vice-versa. Oxyfluorfen decreasedgrowth of both the cyanobacteria by directly affecting chlorophyllbiosynthesis, other pigments and photosynthetic electron trans-port activities and indirectly by increasing ROS and oxidativedamage (MDA and electrolyte leakage). UV-B stress also followedsimilar mechanisms. Further, greater tolerance of P. foveolarum
against oxyfluorfen and UV-B stress in comparison to theN. muscorum may be attributed to strong antioxidant defensesystem (particularly ascorbate and proline), low level of oxidativestress and absence of 66 kDa protein in comparison to N. muscorum.Besides this, presence of 29 kDa protein in P. foveolarum furthermakes it more tolerant against oxyfluorfen and UV-B stressin comparison to N. muscorum. Our study indicates that use ofP. foveolarum may be a better option over N. muscorum to sustainfertility and productivity of soils under such stressed conditions.
Acknowledgments
The authors are thankful to The Head, Department of Botany,University of Allahabad, Allahabad, for providing necessarylaboratory facilities. The authors are also thankful to UGC, NewDelhi, for providing financial support to carry out this work.
References
Adhikary, S.P., Das, P., Pattnaik, H., 1984. Effect of carbamate insecticide Sevin onAnabaena sp. and Westiellopsis prolifica. J. Gen. Appl. Microbiol. 35, 335–338.
Ahrens, J.F., 1979. Repeated applications of granular herbicides on container–grown ornamentals. Abstr. Weed Sci. Soc. Am., 53.
Aizawa, H., Brown, H.M., 1999. Metabolism and degradation of porphyrinbiosynthesis herbicides. In: Boger, P., Wakabayashi, K. (Eds.), PeroxidizingHerbicides. Springer-Verlag, Berlin, Heidelberg, pp. 348–381.
Alia, Pardha Saradhi, P., 1993. Suppression in mitochondrial electron transport isthe prime cause behind stress induced proline accumulation. Biochem.Biophys. Res. Commun 193, 54–58.
Ames, B.N., Dubin, D.T., 1960. The role of polyamines in the neutralization ofbacteriophage deoxyribonucleic acid. J. Biol. Chem. 235, 769–775.
Baruah, M., Mishra, R.R., 1986. Effect of herbicides, butachlor, 2,4-D and oxyfluor-fen on enzyme activities and CO2 evolution in submerged paddy field soil.Plant Soil 96, 287–291.
Bates, L.S., Waldren, R.P., Teare, I.D., 1973. Rapid determination of free proline forwater stress studies. Plant Soil 39, 205–207.
Blumwald, E., Tel-Or, E., 1982. Structural aspects of the adaptation of Nostocmuscorum to salt. Arch. Microbiol. 132, 163–167.
Boger, P., Sandmann, G., 1998. Action of modern herbicides. In: Raghavendra, A.S.(Ed.), Photosynthesis: A Comprehensive Treatise. Cambridge University Press,London, pp. 337–351.
Brown, J.C., Prasad, P., Wu, M.J., Irzyk, G.P., Jones, A.M., 1998. Purification of a65-kiloDolton nuclear protein with structural homology to glutathione-S-transferase. Plant Sci. 136, 227–236.

Sheeba et al. / Ecotoxicology and Environmental Safety 74 (2011) 1981–1993 1993
Bugg, M.W., Whitmarsh, W.J., Rieck, C.E., Cohen, W.S., 1980. Inhibition of photo-synthetic electron transport by diphenyl ether herbicides. Plant Physiol.65, 47–50.
Camm, E.L., Stein, J.R., 1974. Some aspects of the nitrogen metabolism of Nodulariaspumigena (Cyanophyceae). Can. J. Bot. 52, 719–726.
Carletti, P., Masi, A., Wonisch, A., Grill, D., Tausz, M., Ferretti, M., 2003. Changes inantioxidants and pigment pool dimensions in UV-B irradiated maize seedlings.Environ. Exp. Bot. 50, 149–157.
Chen, Z., Juneau, P., Qiu, B., 2007. Effects of three pesticides on the growth,photosynthesis and photoinhibition of the edible cyanobacterium Ge-Xian-Mi(Nostoc). Aquat. Toxicol 81, 256–265.
Cohen-Bazire, G., Bryant, D.A., 1982. Phycobilisomes: composition and structure.In: Carr, N.G., Whitton, S.C. (Eds.), The Biology of Cyanobacteria. BlackwellScientific Publications, Oxford.
Duke, S.O., Lydon, J., Becerill, J.M., Sherman, T.D., Lehnen Jr., L.P., Matsumoto, H.,1991. Protoporphyrinogen oxidase inhibiting herbicides. Weed Sci. 39,465–473.
Dyk, J.S.V., Pletschke, B., 2010. Review on the use of enzymes for the detection oforganochlorine, organophosphate and carbamate pesticides in the environ-ment. Chemosphere 82, 291–307.
Egashira, T., Takahama, U., Nakamura, K., 1989. A reduced activity of catalase as abasis for light dependent methionine sensitivity of a Chlamydomonas reinhardtiimutant. Plant Cell Physiol. 30, 1171–1175.
Elstner, E.F., Heupel, A., 1976. Inhibition of nitrite formation from hydroxylammonium chloride: a simple assay for superoxide dismutase. Anal. Biochem.70, 616–620.
Ferreira, R.M.B., Franco, E., Teixeira, A.R.N., 1996. Covalent dimerization of ribulosebisphosphate carboxylase subunits by UV-radiation. Biochem. J. 318, 227–234.
Gadkari, D., 1988. Effect of some photosynthesis-inhibiting herbicides on growthand nitrogenase activity of a new isolate of cyanobacteria, Nostoc G3. J. BasicMicrobiol. 28, 419–426.
Gahagen, H.E., Halm, R.E., Abeles, F.B., 1968. Effect of ethylene on peroxidaseactivity. Physiol. Plant. 21, 1270–1279.
Gao, K., Yu, H., Brown, M.T., 2007. Solar PAR and UV radiation affects thephysiology and morphology of the cyanobacterium. J. Photochem. Photobiol.B: Biol. 89, 117–124.
Geoffroy, L., Dewez, D., Vernet, G., Popovic, R., 2003. Oxyfluorfen toxic effect onS. obliquus evaluated by different photosynthetic and enzymatic biomarkers.Arch. Environ. Contam. Toxicol. 45, 445–452.
Geoffroy, L., Teisseire, H., Couderchet, M., Vernet, G., 2002. Effect of oxyfluorfenand diuran alone and in mixture on antioxidative enzymes of Scenedesmusobliquus. Pestic. Biochem. Physiol. 72, 178–185.
Giannopolitis, C.N., Ries, S.K., 1977. Superoxide dismutase I: occurrence in higherplants. Plant Physiol. 59, 309–314.
Gong, M., Li, Y.J., Chen, S.Z., 1998. Abscisic acid-induced thermo-tolerance in maizeseedlings is mediated by calcium and associated with antioxidant system.J. Plant Physiol. 153, 488–496.
Goodwin, T.W., 1954. Carotenoids. In: Paech, K., Tracey, M.V. (Eds.), Handbook ofPlant Analysis, Vol. 3. Springer-Varlag, Berlin, pp. 272–311.
Greenberg, B.M., Gaba, V., Mattoo, A.K., Edelman, M., 1989. Degradation of 32 kDaphotosystem II reaction centre protein in UV-B, Visible and Far red light occursthrough a common 23.5 kDa intermediate. Z. NatuforSch 44, 450–452.
He, Y.Y., Hader, D.P., 2002. UV-B-induced formation of reactive oxygen species andoxidative damage of the cyanobacterium Anabaena sp.: protective effectsof ascorbic acid and N-acetyl-L-cysteine. J. Photochem. Photobiol. B: Biol66, 115–124.
Heath, R.L., Packer, L., 1968. Photoperoxidation in isolated chloroplast. I. kineticsand stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125,189–198.
Ihlenfeldt, M.J.A., Gibson, J., 1975. Phosphate utilization and alkaline phosphataseactivity in Anacystis nidulans (Synechococcus). Arch. Microbiol. 102, 23–28.
Jacobs, J.M., Jacobs, J.N., 1987. Oxidation of protoporphyrinogen to protoporphyrin,a step in chlorophyll and haem biosynthesis. Purification and partial char-acterization of the enzyme from barley organelles. Biochem. J 244, 219–224.
Jung, S., 2011. Level of protoporphyrinogen oxidase activity tightly correlates withphotodynamic and defense responses in oxyfluorfen-treated transgenic rice.J. Pestic. Sci. 36, 16–21.
Karapetyan, N.V., Strasser, R., Boger, P., 1983. Variable fluorescence spectra ofalgae after herbicide-induced pigment bleaching. Z. Natuforsch 38C, 556–562.
Kumar, N., Amb, M.K., Bora, A., 2010. Chronic response of Anabaena fertilissima Rao,C.B. on growth, metabolites and enzymatic activities by chlorophenoxyherbicide. Pestic. Biochem. Physiol. 98, 168–174.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227, 680–685.
Li, L., Chen, X., Zhang, D., Pan, X., 2010. Effects of insecticide acetamiprid onphotosystem II (PS II) activity of Synechocystis sp. (FACHB-898). Pestic.Biochem. Physiol. 98, 300–304.
Lowe, R.H., Evans, H.J., 1964. Preparation and some properties of a soluble nitratereductase from Rhizobium joponicum. Biochim. Biophys. Acta 85, 377–389.
Lukaszewicz-Hussain, A., 2010. Role of oxidative stress in organophosphateinsecticide toxicity-short review. Pestic. Biochem. Physiol. 98, 145–150.
Matringe, M., Camadro, J.M., Block, M.A., Joyard, J., Scalla, R., Labbe, P., Douce, R.,1992. Localization within chloroplasts of protoporphyrinogen oxidase, thetarget enzyme for diphenylether-like herbicides. J. Biol. Chem. 267,4646–4651.
Matysik, J., Alia, Bhalu, B., Mohanty, P., 2002. Molecular mechanisms of quenchingof reactive oxygen species by proline under stress in plants. Curr. Sci.82, 525–532.
Mitchell, D.L., Karentz, D., 1993. The induction and repair of DNA photodamage inthe environmemt. In: Young, A.R.L., Bjorn, J., Nultsch, W. (Eds.), EnvironmentalUV Photobiology. Plenum Press, London, pp. 345–377.
Mohammed, A.R., Tarpley, L., 2011. Morphological and physiological responses ofnine southern U.S. rice cultivars differing in their tolerance to enhancedultraviolet-B radiation. Environ. Exp. Bot. 70, 174–184.
Moreland, D.E., Blackman, W.J., Todd, J.G., Farmer, F.S., 1970. Effects of dipheny-lether herbicides on reactions of mitochondria and chloroplasts. Weed Sci.18, 636–642.
Nicholas, D.J.D., Nason, A., 1957. Determination of nitrate and nitrite. In: Colowick,S.P., Kaplan, N.O. (Eds.), Methods in Enzymology, Vol. III. Academic Press, NewYork, pp. 981–984.
Noctor, G., Foyer, C.H., 1998. Ascorbate and glutathione: keeping active oxygenunder control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 249–279.
Oser, B.L., 1979. Hawks Physiological Chemistry. McGraw Hill, pp. 702-705.Porra, R.J., Thompson, W.A., Kriedemann, P.E., 1989. Determination of accurate
extinction coefficients and simultaneous equations for assaying chlorophylls a
and b extracted with four different solvents; verification of the concentrationof chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys.Acta 975, 384–394.
Prasad, S.M., Srivastava, P.K., Singh, V.P., Kumar, D., 2008. Endosulfan inducedeffect on growth, photosynthesis and assimilation of nitrogen and phosphorusin non-heterocystous cyanobacterium Plectonema boryanum. Biochem. Cell.Arch. 8, 109–118.
Prasad, S.M., Zeeshan, M., 2004. Effect of UV-B and monocrotophos, singly and incombination, on photosynthetic activity and growth of non-heterocystouscyanobacterium Plectonema boryanum. Environ. Exp. Bot. 52, 175–184.
Prasad, S.M., Zeeshan, M., 2005. UV-B radiation and cadmium induced changesin growth, photosynthesis, and antioxidant enzymes of cyanobacteriumPlectonema boryanum. Biol. Plant. 49, 229–236.
Pritchard, M.K., Warren, G.F., Dilley, R.A., 1980. Site of action of oxyfluorfen. WeedSci. 28, 640–645.
Renger, G., Volker, M., Eckert, H.J., Frome, R., Hohm-Veit, S., Graber, P., 1989. Onthe mechanism of photosystem II deterioration by UV-B irradiation. Photo-chem. Photobiol. 49, 97–105.
Sagisaka, S., 1976. The occurrence of peroxide in a perennial plant Populas gelrica.Plant Physiol. 57, 308–309.
Siefermann-Harms, D., 1987. The light harvesting protective functions of carote-noids in photosynthetic membranes. Physiol. Plant. 69, 561–568.
Sinha, R.P., Hader, D.P., 2008. UV-B protectant in cyanobacteria. Plant Sci.174, 278–289.
Sinha, R.P., Singh, N., Kumar, A., Kumar, H.D., Hader, M., Hader, D.P., 1996. Effectsof UV irradiation on certain physiological and biochemical processes incyanobacteria. J. Photochem. Photobiol. B Biol. 32, 107–113.
Spiller, H., 1980. Photophosphorylation capacity of stable spheroplasts prepara-tions of Anabaena. Plant Physiol. 66, 446–450.
Torriani, A., 1960. Influence of inorganic phosphate in the formation of phospha-tases by Escherichia coli. Biochim. Biophys. Acta 38, 460–469.
Wang, G., Chen, K., Chen, L., Hu, C., Zhang, D., Liu, Y., 2008. The involvement of theantioxidant system in protection of desert cyanobacterium Nostoc sp. againstUV-B radiation and the effects of exogenous antioxidants. Ecotoxicol. Environ.Saf. 69, 150–157.
World Meteorological Organization, 2007. Scientific assessment of ozone deple-tion: 2006. World Meteorological Organization, Global Ozone Research andMonitoring Project No. 50 Geneva, Switzerland, p. 572.
Yen, J.H., Sheu, W.S., Wang, Y.S., 2003. Dissipation of herbicide oxyfluorfen insubtropical soils and its potential to contaminate groundwater. Ecotoxicol.Environ. Saf. 54, 151–156.
Copyright © 2022 FDOKUMEN




















