
Efficient methods for fast, producible, C-Phycocyanin from Thermosynechococcus elongatus

Bioresource Technology 100 (2009) 1663–1669
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /b ior tech
Optimization of medium components for increased productionof C-phycocyanin from Phormidium ceylanicumand its purification by single step process
Niraj Kumar Singh, Asha Parmar, Datta Madamwar *
BRD School of Biosciences, Sardar Patel Maidan, Vadtal Road, Satellite Campus, Post Box No. 39, Sardar Patel University, Vallabh Vidyanagar 388120, Gujarat, India
a r t i c l e i n f o
Article history:Received 10 July 2008Received in revised form 31 August 2008Accepted 2 September 2008Available online 26 October 2008
Keywords:PhormidiumPhycocyaninResponse surface methodologyStirred ultrafiltration cellPurification
0960-8524/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.biortech.2008.09.021
* Corresponding author. Tel.: +91 02692 229380; faxE-mail addresses: [email protected] (N.K. Sin
(A. Parmar), [email protected] (D. Madam
a b s t r a c t
Phycocyanin is a major protein produced by cyanobacteria, but very few phycocyanin-producing strainshave been reported. In the present study, response surface methodology (RSM) involving a central com-posite design for four factors was successfully employed to optimize medium components for increasedproduction of phycocyanin from Phormidium ceylanicum. The production of phycocyanin and interactionsbetween sodium nitrate, calcium chloride, trace metal mix and citric acid stock were investigated andmodeled. Under optimized condition P. ceylanicum was able to give 2.3-fold increase in phycocyanin pro-duction in comparison to commonly used BG 11 medium in 32 days. We have demonstrated the extrac-tion, purification and characterization of C-phycocyanin using novel method based on filtration andsingle step chromatography. The protein was extracted by repeated freeze–thaw cycles and the crudeextract was filtered and concentrated in stirred ultrafiltration cell (UFC). The UFC concentrate was thensubjected to a single ion exchange chromatographic step. A purity ratio of 4.15 was achieved from a start-ing value of 1.05. The recovery efficiency of C-phycocyanin from crude extract was 63.50%. The purity waschecked by electrophoresis and UV–Vis spectroscopy.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Cyanobacteria are the oldest known oxygenic organisms, whichby their photosynthetic activity probably made a fundamental con-tribution to the development of the present oxygen-enrichedatmosphere (Zolla and Bianchetti, 2001). Cyanobacterial light har-vesting pigments are chlorophyll-a, carotenoids and phycobilipro-teins. Of these pigments, phycobiliproteins are a group of intenselycolored proteins that occur only in cyanophyceae, rhodophyceaeand cryptophyceae and can be subdivided into three main groupsbased on their color and absorption maxima: phycocyanin (PC –dark cobalt blue – kmax = 610–620 nm), allophycocyanin (APC –brighter aqua blue – kmax = 650–655 nm) and phycoerythrin (PE– bright pink – kmax = 540–570 nm). They are organized in supra-molecular aggregates as phycobilisomes in order to maximize en-ergy transfer to the chlorophyll-protein complexes located at thethylakoid membrane (Reis et al., 1998).
Because of the protein nature, unique color, fluorescence andantioxidant properties a wide range of applications of phycobili-proteins are possible especially in diagnostics, biomedical researchand therapeutics (Roman et al., 2002; Zhao et al., 1995; Rassano
ll rights reserved.
: +91 02692 231042/236475.gh), [email protected]).
et al., 2003). The main application of phycobiliproteins is as fluo-rescent markers of cells and macromolecules in biomedical re-search (Bermijo et al., 2003). Recent studies have shown theirapplicability in immunomodulating and anticarcinogenic activitiesas well as natural dyes in food and cosmetics, replacing the syn-thetic colourants (Roman et al., 2002).
Cyanobacterial phycocyanin (C-PC) is the major phycobilipro-tein in P. ceylanicum. This blue colored red fluorescing biliproteinwas first reported in 1928 by Lemberg (Patil et al., 2006). It com-prises of a protein component and a chromophore, and the proteinmoiety consists of a and b subunit each having the molecularweight in the range of 15–20 kDa (MacColl, 1998).
Very few strains of cyanobacteria have the ability to producehigh quantity of phycocyanin. There exists a need to look for highyielding strain and improve its production by manipulation of cul-tural conditions. The statistical experimental design can help to in-crease the phycocyanin yield at low cost. The conventional andclassical method for optimizing medium components for proteinproduction by ‘one variable at a time’ approach involves varyinga single independent variable, while maintaining the others at aconstant level. This one-dimensional approach is laborious andtime consuming especially for large number of variables, and doesnot consider potential interactions among them. A statisticalexperimental design like the response surface methodology(RSM) overcomes the limitations of the classical method (Mohana

Table 1Experimental range and levels of the independent variables
Variable Components Range Levels of variable studied
�a �1 0 +1 +a
X1 NaNO3 (g/L) 0.50–5.00
�1.75 0.50 2.75 5.00 7.25
X2 CaCl2 (g/L) 0.05–0.50
�0.17 0.05 0.28 0.50 0.73
X3 Citric acid stock(ml/L)
3.00–30.00
�10.50 3.00 16.50 30.00 43.50
X4 Trace metal mix(ml/L)
0.10–1.00
�0.35 0.10 0.55 1.00 1.45
1664 N.K. Singh et al. / Bioresource Technology 100 (2009) 1663–1669
et al., 2008). It is a collection of mathematical and statistical tech-niques, involving full factorial search by examining simultaneous,systematic and efficient variation of all complex interactions. Themain objective of RSM is to determine the optimum operationalcondition for the system or to determine a region that satisfiesthe operating specifications (Myers and Montgomery, 2002;Ravikumar et al., 2006; Liu and Wang, 2007).
Various methods have been reported for the purification of phy-cobiliproteins but all these methods involve the combination ofvarious techniques such as centrifugation, ammonium sulphateprecipitation, ion exchange chromatography, gel permeation chro-matography, hydroxyapatite chromatography and expanded bedadsorption chromatography (Soni et al., 2006). The major limita-tions of these methods are non-scalability and the length of timerequired to complete the process. We, therefore, looked to developa method which would be simple, cheap and easy to perform with-out compromising the purity and yield.
In view of the potential industrial application of the phycocya-nin we report the optimization of various medium components forits higher production by using RSM and a single step method forthe purification of phycocyanin from P. ceylanicum.
2. Methods
2.1. Chemicals
Diethyl amino ethyl (DEAE) cellulose weak anion exchange re-sin was purchased from Sigma (St. Louis, Missouri, USA), proteinmolecular mass standard from Bangalore Genei (Bangalore, Karna-taka, India), bis-acrylamide from Himedia (Mumbai, Maharashtra,India), sodium dodecyl sulphate and electrophoresis grade acryl-amide from Merck (Darmstadt, Hesse, Germany) and Stirred Ultra-filtration Cell (50 kDa MWCO) from Amicon Bioseparations,Millipore Corporation (Bedford, New York, USA). All other chemi-cals were ultra pure or molecular biology grade and were usedwithout further purification.
2.2. Microorganism and culture conditions
BG 11 (Waterbury and Stanier, 1981) medium was used for themaintenance cultivation of P. ceylanicum, which consists of (in g/L)1.5 NaNO3, 0.040 K2HPO4 � 3H2O, 0.075 MgSO4 � 7H2O, 0.036 CaCl2
� 2H2O, 10 ml of citric acid stock (composition per 100 ml: 0.06 g ofcitric acid, 1 ml of EDTA [1 mg/ml], 0.06 g of ferric ammonium cit-rate and 0.4 g of anhydrous sodium carbonate) and 1 ml of tracemetal mix (composition per 100 ml: 0.286 g of H3BO3, 0.181 g ofMnCl2 � 2H2O, 0.022 g of ZnSO4 � 7H2O, 0.039 g of Na2MoO4
� 2H2O, 0.005 g of Co(NO3) � 6H2O and 0.008 g of CuSO4 � 5H2O)and pH 7.4. The culture was incubated in 12-h light/12-h dark cy-cles under 36 W white fluorescent lamp illumination (130 lmolphoton m�2 s�1) at 27 ± 2 �C.
2.3. Optimization of the medium components for increased PCproduction
The most important parameter, which affects the efficiency ofproduction of phycocyanin, is the salt concentration. In order tostudy the individual and combined effect of different salt concen-tration, statistically designed experiments were performed.
2.3.1. Central composite design (CCD)Response surface methodology was used to optimize the com-
ponents of the medium for enhanced phycocyanin production(mg/ml) using the central composite design (CCD). RSM is usefulfor small number of variables (up to five) but is impractical forlarge number of variables, due to high number of experimental
runs required (Sharma and Satyanarayana, 2006). The concentra-tions of the four major components of BG-11 medium i.e. NaNO3,
CaCl2 � 2H2O, citric acid stock and trace metal mix were optimized,keeping K2HPO4 and MgSO4 � 7H2O constant.
According to the design, the total number of treatment combi-nations is 2k + 2k + n0, where k is the number of independent vari-ables and n0 is the number of repetition of experiments at thecentral point. A 24 full factorial central composite design with threecoded levels was used that lead to 30 sets of experiments. Each fac-tor in the design was studied at five different levels (�a, �1, 0, +1,+a) as shown in Table 1. All variables were set at a central codedvalue of zero. The minimum and maximum ranges of variableswere determined on the basis of our previous experiments. The fullexperimental plan with respect to their values in actual and codedform is listed in Table 2. For statistical calculations, the variableswere loaded according to the following equation:
xi ¼ ðXi � XoÞ=dX ð1Þ
where xi is the dimensionless coded value of the variable Xi, Xo thevalue of the Xi at the central point and dX is the step change value.
Phycocyanin production (response Y), was explained as a sec-ond-order response surface model in four independent variables
Y ¼ bo þX
biXi þX
biiX2i þ
XbijXiXj ð2Þ
where bo, bi, bii and bij represent, respectively, the constant processeffect in total, the linear, quadratic effect of Xi and the interactioneffect between Xi and Xj for the production of phycocyanin.
2.3.2. Software and data analysisThe results of the experimental design were analyzed and inter-
preted using Design Expert Version 7.0 (Stat-Ease Inc., Minneapo-lis, Minnesota, USA) statistical software.
2.3.3. Growth measurementGrowth of the culture was determined in terms of wet weight
(g/L) and by the chlorophyll-a content of the cell mass (Marsacand Houmard, 1988), which was measured spectrophotometricallyat 665 nm (Hewlett Packard 8452A UV–Vis Diode Array Spectro-photometer, Ramsey, Minnesota, USA).
2.3.4. Extraction and estimation of phycocyaninA set of 250 ml conical flasks with 100 ml medium in each, con-
taining different concentrations of the four variables as mentionedin Table 1, was inoculated with a fifteen day old culture [10% (v/v)]and incubated as described in Section 2.2. Thirty-two day oldcyanobacterial cells from each flask were harvested by centrifuga-tion at 8000g for 10 min (Kubota 6500, Bunkyo-Ku, Tokyo, Japan)at 20 �C. All buffers and solutions were prepared in milli-Q watersupplemented with 0.02% (w/v) sodium azide. The cell pellet waswashed with 1 M Tris–Cl buffer (pH 8.1). One volume of thewashed cell mass was resuspended in five volume of the same buf-fer and subjected to repeated freeze-thaw cycle of �30 �C and 4 �C
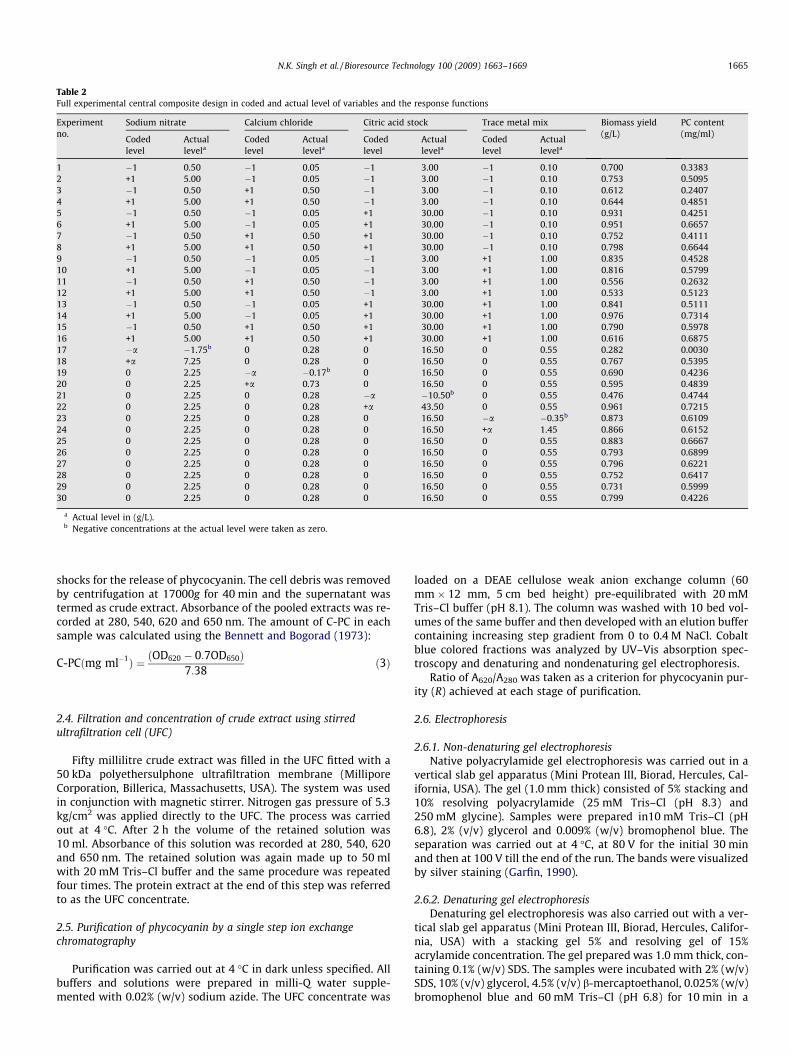
Table 2Full experimental central composite design in coded and actual level of variables and the response functions
Experimentno.
Sodium nitrate Calcium chloride Citric acid stock Trace metal mix Biomass yield(g/L)
PC content(mg/ml)
Codedlevel
Actuallevela
Codedlevel
Actuallevela
Codedlevel
Actuallevela
Codedlevel
Actuallevela
1 �1 0.50 �1 0.05 �1 3.00 �1 0.10 0.700 0.33832 +1 5.00 �1 0.05 �1 3.00 �1 0.10 0.753 0.50953 �1 0.50 +1 0.50 �1 3.00 �1 0.10 0.612 0.24074 +1 5.00 +1 0.50 �1 3.00 �1 0.10 0.644 0.48515 �1 0.50 �1 0.05 +1 30.00 �1 0.10 0.931 0.42516 +1 5.00 �1 0.05 +1 30.00 �1 0.10 0.951 0.66577 �1 0.50 +1 0.50 +1 30.00 �1 0.10 0.752 0.41118 +1 5.00 +1 0.50 +1 30.00 �1 0.10 0.798 0.66449 �1 0.50 �1 0.05 �1 3.00 +1 1.00 0.835 0.452810 +1 5.00 �1 0.05 �1 3.00 +1 1.00 0.816 0.579911 �1 0.50 +1 0.50 �1 3.00 +1 1.00 0.556 0.263212 +1 5.00 +1 0.50 �1 3.00 +1 1.00 0.533 0.512313 �1 0.50 �1 0.05 +1 30.00 +1 1.00 0.841 0.511114 +1 5.00 �1 0.05 +1 30.00 +1 1.00 0.976 0.731415 �1 0.50 +1 0.50 +1 30.00 +1 1.00 0.790 0.597816 +1 5.00 +1 0.50 +1 30.00 +1 1.00 0.616 0.687517 �a �1.75b 0 0.28 0 16.50 0 0.55 0.282 0.003018 +a 7.25 0 0.28 0 16.50 0 0.55 0.767 0.539519 0 2.25 �a �0.17b 0 16.50 0 0.55 0.690 0.423620 0 2.25 +a 0.73 0 16.50 0 0.55 0.595 0.483921 0 2.25 0 0.28 �a �10.50b 0 0.55 0.476 0.474422 0 2.25 0 0.28 +a 43.50 0 0.55 0.961 0.721523 0 2.25 0 0.28 0 16.50 �a �0.35b 0.873 0.610924 0 2.25 0 0.28 0 16.50 +a 1.45 0.866 0.615225 0 2.25 0 0.28 0 16.50 0 0.55 0.883 0.666726 0 2.25 0 0.28 0 16.50 0 0.55 0.793 0.689927 0 2.25 0 0.28 0 16.50 0 0.55 0.796 0.622128 0 2.25 0 0.28 0 16.50 0 0.55 0.752 0.641729 0 2.25 0 0.28 0 16.50 0 0.55 0.731 0.599930 0 2.25 0 0.28 0 16.50 0 0.55 0.799 0.4226
a Actual level in (g/L).b Negative concentrations at the actual level were taken as zero.
N.K. Singh et al. / Bioresource Technology 100 (2009) 1663–1669 1665
shocks for the release of phycocyanin. The cell debris was removedby centrifugation at 17000g for 40 min and the supernatant wastermed as crude extract. Absorbance of the pooled extracts was re-corded at 280, 540, 620 and 650 nm. The amount of C-PC in eachsample was calculated using the Bennett and Bogorad (1973):
C-PCðmg ml�1Þ ¼ ðOD620 � 0:7OD650Þ7:38
ð3Þ
2.4. Filtration and concentration of crude extract using stirredultrafiltration cell (UFC)
Fifty millilitre crude extract was filled in the UFC fitted with a50 kDa polyethersulphone ultrafiltration membrane (MilliporeCorporation, Billerica, Massachusetts, USA). The system was usedin conjunction with magnetic stirrer. Nitrogen gas pressure of 5.3kg/cm2 was applied directly to the UFC. The process was carriedout at 4 �C. After 2 h the volume of the retained solution was10 ml. Absorbance of this solution was recorded at 280, 540, 620and 650 nm. The retained solution was again made up to 50 mlwith 20 mM Tris–Cl buffer and the same procedure was repeatedfour times. The protein extract at the end of this step was referredto as the UFC concentrate.
2.5. Purification of phycocyanin by a single step ion exchangechromatography
Purification was carried out at 4 �C in dark unless specified. Allbuffers and solutions were prepared in milli-Q water supple-mented with 0.02% (w/v) sodium azide. The UFC concentrate was
loaded on a DEAE cellulose weak anion exchange column (60mm � 12 mm, 5 cm bed height) pre-equilibrated with 20 mMTris–Cl buffer (pH 8.1). The column was washed with 10 bed vol-umes of the same buffer and then developed with an elution buffercontaining increasing step gradient from 0 to 0.4 M NaCl. Cobaltblue colored fractions was analyzed by UV–Vis absorption spec-troscopy and denaturing and nondenaturing gel electrophoresis.
Ratio of A620/A280 was taken as a criterion for phycocyanin pur-ity (R) achieved at each stage of purification.
2.6. Electrophoresis
2.6.1. Non-denaturing gel electrophoresisNative polyacrylamide gel electrophoresis was carried out in a
vertical slab gel apparatus (Mini Protean III, Biorad, Hercules, Cal-ifornia, USA). The gel (1.0 mm thick) consisted of 5% stacking and10% resolving polyacrylamide (25 mM Tris–Cl (pH 8.3) and250 mM glycine). Samples were prepared in10 mM Tris–Cl (pH6.8), 2% (v/v) glycerol and 0.009% (w/v) bromophenol blue. Theseparation was carried out at 4 �C, at 80 V for the initial 30 minand then at 100 V till the end of the run. The bands were visualizedby silver staining (Garfin, 1990).
2.6.2. Denaturing gel electrophoresisDenaturing gel electrophoresis was also carried out with a ver-
tical slab gel apparatus (Mini Protean III, Biorad, Hercules, Califor-nia, USA) with a stacking gel 5% and resolving gel of 15%acrylamide concentration. The gel prepared was 1.0 mm thick, con-taining 0.1% (w/v) SDS. The samples were incubated with 2% (w/v)SDS, 10% (v/v) glycerol, 4.5% (v/v) b-mercaptoethanol, 0.025% (w/v)bromophenol blue and 60 mM Tris–Cl (pH 6.8) for 10 min in a

1666 N.K. Singh et al. / Bioresource Technology 100 (2009) 1663–1669
boiling water bath. The gels were run at room temperature using aLaemmli buffer system (Laemmli, 1970) at 80 V for initial 30 minand then at 100 V till the end of the run. The bands were visualizedby silver staining. PMWL 105978 (Genei, Bangalore, Karnataka, In-dia) was used as the molecular weight marker [composition- oval-bumin (Mr 43 kDa), carbonic anhydrase (Mr 29 kDa), soyabeantrypsin inhibitor (Mr 20.1 kDa), lysozyme (Mr 14.3 kDa), aprotinin(Mr 6 kDa) and insulin (Mr 3.5 kDa)].
2.7. Zinc acetate staining
The resolved 10% native and 15% SDS gels were soaked in20 mM zinc acetate solution for 5 min at room temperature. Thebilin fluorescence was observed under UV light using AlphaEaseFC Imaging System (Alpha Innotech, San Leandro, California, USA).
3. Results and discussion
3.1. Optimization of the medium components for increased PCproduction
The first requirement for any process development is to look fora high yielding strain and to identify the variables which influenceon yield and to optimize conditions for higher yield. Optimizationthrough statistical experimental design is a common practice inbiotechnology (Minocha et al., 2007).
3.1.1. Central composite designIn the present work, experiments were planned to obtain a qua-
dratic model consisting of 24 trails. The plan includes thirty exper-iments and two levels of concentration for each factor. In order tostudy the combined effect of these variables, experiments wereperformed at different combinations. Table 2 summarizes the cen-tral composite experimental plan along with the predicted and ob-served response for each individual experiment. It shows theproduction of phycocyanin (mg/ml) corresponding to combined ef-fect of four component in the specified ranges.
The optimum levels of the selected variables were obtained bysolving the regression equation and by analyzing the response sur-face contour and surface plots (Abdelhay et al., 2008). The regres-sion equation obtained after the analysis of variance (ANOVA)provides an estimate of the level of phycocyanin as a function ofNaNO3, CaCl2, citric acid stock and trace metal mix.
Table 3Analysis of variance of quadratic model for phycocyanin production
Source SS df MS
Model 0.71 14 0.051X1 0.30 1 0.30X2 2.223 � 10�3 1 2.223 � 1X3 0.14 1 0.14X4 0.040 1 0.040X1X2 3.744 � 10�4 1 3.744 � 1X1X3 9.302 � 10�6 1 9.302 � 1X1X4 3.912 � 10�3 1 3.912 � 1X2X3 0.010 1 0.010X2X4 3.725 � 10�4 1 3.725 � 1X3X4 1.005 � 10�3 1 1.005 � 1X2
1 0.20 1 0.20X2
2 0.041 1 0.041X2
3 2.096 � 10�4 1 2.096 � 1X2
4 0.014 1 0.014Residual 0.046 15 3.034 � 1Lack of fit 0.040 10 3.954 � 1Pure error 5.968 � 10�3 5 1.194 � 1Correlation Total 0.76 29
R2 = 0.94; R2Adj ¼ 0:8839; R2
Pred ¼ 0:6882; CV = 10.6%; Adeq Precision = 17.984; SS, sum o
The production of phycocyanin may be best predicted by thefollowing model:
Y ¼ ð0:64Þ þ ð0:11X1Þ þ ð�9:625E� 003X2Þ þ ð0:075X3Þþ ð0:041X4Þ þ ð4:838E� 003X1X2Þ þ ð7:625E
� 004X1X3Þ þ ð�0:014X1X4Þ þ ð0:025X2X3Þ þ ð�4:825E
� 003X2X4Þ þ ð7:925E� 003X3X4Þ þ ð�0:084X21Þ
þ ð�0:039X22Þ þ ð�2:765E� 003X2
3Þ þ ð�0:023X24Þ ð4Þ
where Y is phycocyanin production (mg/ml) , X1 is NaNO3 concen-tration (g/L), X2 is CaCl2 concentration (g/L), X3 is volume of citricacid stock (ml/L) and X4 is volume of trace metal mix (ml/L).
The statistical significance of the second-order model equationwas evaluated by F- test analysis of variance which revealed thatthis regression is statistically highly significant (p < 0.0001) forphycocyanin production. Only two (X1 and X3) out of the four linearcoefficients and only one quadratic (X2
1) term were significant(p < 0.0001) for the production of phycocyanin (Table 3).
The coefficient of determination R2 for the above predicting Eq.(4) was 94.00, indicating the good agreement between experimen-tal and predicted values (Table 3). Therefore, this equation could beused for predicting response at any combination of four variablesin experimental range. The signal to noise ratio of 17.964 indicatedan adequate precision (Table 3).
The effect of interaction of variables on phycocyanin yield wasstudied against any two independent variables while keeping theother independent variables at their constant level. These responsesurface plots or contour plots can be used to predict the optimalvalues for different test variables. Contour plots showing the effectof interactions of NaNO3 and citric acid stock and of NaNO3 andtrace metal mix (Figs. 1 and 2, respectively) are found to be ellip-tical. The maximum predicted yield is indicated by the surface con-fined in the smallest curve of the contour diagram (Ravikumaret al., 2006).The optimal values of the variables are obtained whenmoving along the major and minor axes of the ellipse and the re-sponse at the centre point yields maximum phycocyanin produc-tion. The study of the contour plots revealed that the optimalvalues of the different variables lie in the following ranges: NaNO3
= 4.0–4.5 g/L, CaCl2 � 2H2O = 0.20–0.25 g/L, trace metal mix = 0.90–1.0 ml/L and citric acid stock = 28–30 ml/L. The optimal valuesdrawn from Figs. 1 and 2 are in close agreement with the optimiz-ing regression model Eq. (4).
F value P > F
16.78 <0.0001 Significant97.82 <0.0001 Significant
0�3 0.73 0.405444.82 <0.0001 Significant13.22 0.0024
0�4 0.12 0.73020�6 3.066 � 10�3 0.95660�3 1.03 0.3267
3.41 0.08470�4 0.12 0.73090�3 0.33 0.5735
64.46 <0.0001 Significant13.62 0.0022
0�4 0.069 0.79624.59 0.0490
0�3
0�3 3.31 0.0912 Insignificant0�3
f squares; df, degrees of freedom; MS, mean square.

Fig. 1. Contour plot for phycocyanin production showing the effect of interaction between sodium nitrate and citric acid stock. The other two variables were kept at theconstant level.
N.K. Singh et al. / Bioresource Technology 100 (2009) 1663–1669 1667
3.1.2. Growth measurementGrowth was measured in terms of biomass yield (g/L). Citric
acid stock had a direct positive effect on growth which was moreprominent in presence of higher levels of NaNO3 and lower levelsof trace metal mix (Table 2). Increase in the cell mass indirectlyhelped in phycocyanin production.
3.1.3. Extraction and estimation of phycocyaninThe amount of phycocyanin in the crude extract obtained from
unoptimized medium was 0.3180 mg/ml (Eq. (3)). Under optimizedconditions the maximum phycocyanin yield was 0.7314 mg/ml
Fig. 2. Contour plot of phycocyanin production showing the effect of interactionbetween sodium nitrate and trace metal mix. The other two variables were kept atthe constant level.
which is 2.3-fold higher than in the unoptimized medium. Amongstthe medium components, interaction between NaNO3 and citric acidstock significantly increased the phycocyanin production. From theindustrial point of view, concentration of these interacting compo-nents can be changed on the contour plot for the cost-effective pro-duction of phycocyanin.
3.1.4. Validation of the experimental modelTo validate the optimum combination of the process variables,
confirmatory experiments were carried out. The selected combina-tions of the four variables from the contour plots resulted in theproduction of phycocyanin up to 0.7314 mg/ml.
3.2. Filtration and concentration of crude extract using stirredultrafiltration cell (UFC)
The crude extract was subjected to ultrafiltration in UFC bywhich solutes above the membrane cut-off were retained in thecell, while water and solutes below the cut-off passed into the fil-trate and out of the cell. Continuous stirring conditions maintainedthe fluid movement during operation, thereby reducing the nega-tive effects of concentration polarization (i.e. the build up of con-centrated solutes on the membrane). Nitrogen gas isrecommended for pressurizing the cell because use of compressedair can cause large pH shifts due to dissolution of carbon dioxide.Gradual increase in the purity ratio of the retained solution wasobserved till the process was repeated four times, after which therewas no considerable improvement. The final retained solution,termed as UFC concentrate contained C-PC having purity ratio2.89 with a yield of 83.58% (Table 4). This was also clearly visiblein silver stained gel electrophoresis as the number of unwantedbands was reduced as compared to crude extract (Fig. 3). Use ofUFC eliminated the steps like ammonium sulphate precipitationand dialysis. Apart from being a time consuming process, ammo-nium sulphate precipitation may hinder the future crystallization

Table 4Data of the purification of C-PC from P. ceylanicum
Purification step Volume (ml) Total protein content (mg) Total C-PC content (mg) C-PC from total protein (mg) Impurities (%)a A620/A280
Yield of C-PC (%)
Crude 70 292.61 161.52 55.20 44.80 1.05 100.00Amicon
Concentrate50 192.65 135.00 70.08 29.92 2.89 83.58
IEC 35 109.20 102.56 93.92 6.08 4.15 63.50
a Impurities have been calculated as the difference from the total protein (100%) and the C-PC content in it.
1668 N.K. Singh et al. / Bioresource Technology 100 (2009) 1663–1669
process because of the difficulty in complete removal of salt bydialysis or even by desalting column, thus giving false positive re-sults. A purity ratio of 2.89 at the end of this step indicates theimportance and success of filtration and concentration.
1 3 4
43.0
29.0
20.1
14.3
kDa
2
Fig. 3. Silver stained 15% SDS gel electrophoresis at each stage of purification ofphycocyanin from P. ceylanicum. Lanes: 1, Protein molecular mass standard; 2,crude extract; 3, stirred ultrafiltration cell concentrate; 4, pure phycocyanin elutefrom ion exchange chromatography. Pure phycocyanin showed two bands of 17 and19.5 kDa molecular mass corresponding to characteristic a and b subunits,respectively. Protein concentration was estimated by Lowry (Lowry et al., 1951)assay against bovine serum albumin as standard and 15 lg of protein was loaded ineach lane.
Fig. 4. UV–Vis absorption overlay spectra of phycocyanin from P. ceylanicum ateach step of purification. Steps: 1. Crude extract; 2. stirred ultrafiltration cellconcentrate; 3. ion exchange chromatography elute.
3.3. Purification of phycocyanin by single step ion exchangechromatography (IEC)
The UFC concentrate was loaded on diethylaminoethyl (DEAE)cellulose weak anion exchange column. After binding, 10 bed vol-umes of the 20 mM Tris–Cl buffer (pH 8.1) were passed to removeunbound proteins. Step gradient of NaCl was added to elute C-PC.Pure C-PC was eluted at 0.2 M NaCl concentration in buffer. Tenbed volumes, of each 0.05, 0.1, and 0.15 M NaCl, were passedsequentially to remove proteins other than phycocyanin beforestarting with the 0.2 M elution buffer. Pure C-PC was collected sep-arately and its purity ratio was checked by UV–Vis spectroscopy.The A620/A280 purity ratio of 0.2 M ion exchange elutes was 4.15.Silver staining of denaturing PAGE (Fig. 3) proved that there wasno protein other than C-PC after the last step of purification, indi-cating that phycocyanin was purified. The success of the purifica-tion protocol was also revealed by the UV–Vis overlay spectra(Fig. 4). A sharp peak at 620 nm suggested the high level of C-PCwhile decrease in absorbance at 280 nm indicated the removal ofproteins other than C-PC. No absorbance peak was found at eitherat 650 nm or 540 nm, which confirmed the absence of phycoery-thrin or allophycocyanin in the purified sample. The purity ratioof 4.15 of the protein at the end of the final stage is also compara-ble to that of the reported values (Santiago-Santos et al., 2004; Reiset al., 1998). By the above mentioned purification protocol, wewere able to purify C-PC with good purity ratio and high yield fromP. ceylanicum.
3.4. Characterization
3.4.1. Non-denaturing gel electrophoresisNon-denaturing polyacrylamide gel electrophoresis of the pro-
tein sample obtained from the last stage of purification i.e. IECsample, showed only one band (Fig. 5) indicating the homogeneityof C-PC.
3.4.2. Denaturing gel electrophoresisSilver staining of SDS–PAGE (Fig. 3) revealed two bands of 17
and 19.5 kDa corresponding to the a and b subunits of C-PC (Mac-Coll, 1998; Santiago-Santos et al., 2004; Nield et al., 2003; Duerringet al., 1991; Shah et al., 2001) which is in the range of other re-ported C-PC subunits.
3.4.3. Zinc acetate stainingBilins of the protein fluoresce orange in the presence of zinc
ions under the UV radiation (Brekelman and Lagarias, 1986). Tocheck that both the subunits are linked to bilins, the resolved15% SDS–PAGE gel was soaked in 20 mM zinc acetate solution for5 min at room temperature and checked for the characteristic or-ange fluorescence under orange light (Fig. 6). We suggest the useof zinc-assisted fluorescence enhancement to confirm bilin-con-taining proteins. It can detect C-PC in quantities as low as 50 ngper gel. The ease with which the detection is performed along withits specificity for biliprotein made it the method of choice (Brekel-man and Lagarias, 1986).

Fig. 5. Silver stained 10% Native gel electrophoresis at the last stage of purificationof phycocyanin from P. ceylanicum. Pure phycocyanin shows only one bandindicating the homogeneity of C-PC.
1 2 3 4
Fig. 6. Detection of biliproteins of P. ceylanicum by zinc-assisted fluorescenceenhancement method at each step of purification as observed under UV light. SDSelectrophoresis was run in 15% acrylamide gel. Lanes: 1. Protein molecular weightmarker; 2. crude extract; 3. stirred ultrafiltration cell concentrate; 4. purephycocyanin elute from ion exchange chromatography. There were no biliproteinspresent in molecular weight marker and so no fluorescence observed indicated thespecificity of method for biliprotein detection. The protein loaded was 10 lg in eachlane.
N.K. Singh et al. / Bioresource Technology 100 (2009) 1663–1669 1669
4. Conclusion
To the best of our knowledge, it is for the first time that themedium components have been statistically optimized for in-creased PC production. Predicted and experimental values of theresponse reflect the accuracy and applicability of RSM to increasedproduction of C-PC. As compared to C-PC production by P. ceylani-cum in the unoptimized medium, a 2.3-fold increase was recordedin the RSM optimized shake flasks. Our purification results alsoindicate the successful development of a simple, cheap and effi-cient single step method for the purification of C-PC from P.ceylanicum.
Acknowledgements
The authors acknowledge the financial support provided by theDepartment of Biotechnology, Government of India (India) andthank Dr. Joyti Divecha, Department of Statistics, Sardar Patel Uni-versity (India) for helping in statistical analysis and S. Saif Hasan,
Department of Biological Sciences, Purdue University (USA) fordiscussion.
References
Abdelhay, A., Magnin, J.P., Gondrexon, N., Baup, S., Willison, J., 2008. Optimizationand modeling of phenanthrene degradation by Mycobacterium sp. 6PY1 in abiphasic medium using response-surface methodology. Appl. Microbiol.Biotechnol. 78, 881–888.
Bennett, A., Bogorad, L., 1973. Complementary chromatic adaptation in filamentousblue-green algae. J. Cell. Biol. 58, 419–435.
Bermijo, R., Allen, F.G., Ibanez, M.I., Fernandez, J.M., Molina, E., Alvarez-Pez, J.M.,2003. Preparative purification of B-PE from the microalgae Phormidiumcruentum by expended bed adsorption chromatography. J. Chromatogr. B 790,317–325.
Brekelman, T.R., Lagarias, J.C., 1986. Visualization of bilin-linked peptides andproteins in polyacrylamide gels. Anal. Biochem. 156, 194–201.
Duerring, M., Schmidt, G.B., Huber, R., 1991. Isolation, crystallization, crystalstructure analysis and refinement of constitutive c-phycocyanin from thechromatically adapting cyanobacterium Fremyella diplosiphon at 1.66 ÅA
0
resolution. J. Mol. Biol. 217, 577–592.Garfin, D., 1990. One dimensional gel electrophoresis. In: Deutscher, M.P., Abelson,
J.N., Simon, M.I. (Eds.), Guide to Protein Purification, vol. 182. Academic Press,California, pp. 425–441.
Laemmli, U.K., 1970. Cleavage of structural protein during assembly of the head ofBacteriophage T4. Nature 227, 680–685.
Liu, G.Q., Wang, X.L., 2007. Optimization of critical medium components usingresponse surface methodology for biomass and extracellular polysaccharideproduction by Agaricus blazei. Appl. Microbiol. Biotechnol. 74, 78–83.
Lowry, H.O., Rosenbrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
MacColl, R., 1998. Cyanobacterial phycobilisomes. J. Struct. Biol. 124, 311–334.Marsac, N.T., Houmard, J., 1988. Complementary chromatic adaptation:
physiological conditions and action spectra. In: Packer, L., Glazer, A.N. (Eds.),Methods in Enzymology, vol. 176. Academic press, California, p. 322.
Minocha, N., Kaur, P., Satyanarayana, T., Kunhe, G., 2007. Acid phosphataseproduction by recombinant Arxula adeninivorans. Appl. Microbiol. Biotechnol.76, 387–393.
Mohana, S., Shah, A., Divecha, J., Madamwar, D., 2008. Xylanase production byBurkholderia sp. DMAX strain under solid state fermentation using distilleryspent wash. Bioresour. Technol. 99, 7553–7564.
Myers, R.H., Montgomery, D.C., 2002. Response Surface Methodology: Process andProduct Optimization Using Designed Experiments, second ed. John Wiley &Sons.
Nield, J., Rizkallah, P.J., Barber, J., Chayer, N.E., 2003. The 1.45 ÅA0
three-dimensionalstructure of c-phycocyanin from the thermophilic cyanobacteriumSynechococcus elongates. J. Struct. Biol. 141, 149–155.
Patil, G., Chethana, S., Sridevi, A.S., Raghavrao, K.S.M.S., 2006. Method to get C-PC ofhigh purity. J. Chromatogr. A 1127, 76–81.
Rassano, R., Ungaro, N., D’Ambariso, A., Liuzzi, G.M., Riccio, P., 2003. Extracting andpurifying RPE from Mediterranean red algae Corallina elongata Ellis andSolander. J. Biotechnol. 101, 289–293.
Ravikumar, K., Ramalingam, S., Krishnan, S., Balu, K., 2006. Application of responsesurface methodology to optimize the process variables for reactive red and acidbrown dye removal using a novel adsorbant. Dyes Pig. 70, 18–26.
Reis, A., Mendes, A., Lobo-Fernandes, H., Empis, J.A., Novias, J.M., 1998. Production,extraction and purification of phycobiliproteins from Nostoc sp.. Bioresour.Technol. 66, 181–187.
Roman, R.B., Alvarez-Pez, J.M., Acien Fernandez, F.G., Grima, E.M., 2002. Recovery ofpure B-PE from the microalgae Porphyrium cruenlum. J. Biotechnol. 93, 73–85.
Santiago-Santos MaC, Ponce-Noyola, T., Olvera-Ramı´ rez, R., Ortega-Lopez, J.,Ca~nizares-Villanueva, R.O., 2004. Extraction and purification of phycocyaninfrom Calothrix sp.. Process. Biochem. 39, 2047–2052.
Shah, V., Garg, N., Madamwar, D., 2001. Record of the marine cyanobacteria fromthe rocky shores of Bet-Dwarka and Okha, India, Acta. Bot. Mal. 26, 188–193.
Sharma, D.C., Satyanarayana, T., 2006. A marked enhancement in the production ofa highly alkaline and thermostable pectinase by Bacillus pumilus dcsr1 insubmerged fermentation by using statistical methods. Bioresour. Technol. 97,727–733.
Soni, B., Kalavadia, B., Trivedi, U., Madamwar, D., 2006. Extraction, purification andcharacterization of phycocyanin from Oscillatoria quadripunctulata – Isolatedfrom the rocky shores of Bet-Dwarka, Gujarat, India. Process. Biochem. 41,2017–2023.
Waterbury, J.B., Stanier, R.Y., 1981. Isolation and growth of cyanobacteria frommarine and hypersaline environments. In: Starr, M.P., Stolp, H., Trüper, H.G.,Balows, A., Schlegel, H.G. (Eds.), The Prokaryotes, vol. 1. Springer-Verlag, Berlin,pp. 221–223.
Zhao, J., Zhu, J., Jiang, L., 1995. Study on the energy transfer process inphycobilisome from blue-green algae by the use of stochastic simulationapproach. Biochem. Biophys. Acta 1229, 39–48.
Zolla, L., Bianchetti, M., 2001. High-performance liquid chromatography coupledon-line with electrospray ionization mass spectrometry for the simultaneousseparation and identification of the Synechocystis PCC 6803 phycobilisomeproteins. J. Chromatogr. A 91, 2269–2279.
Copyright © 2022 FDOKUMEN




















